31
CHAPTER -2 AIMS & OBJECTIVES

CHAPTER -2
AIMS & OBJECTIVES

AIM OF THE STUDY
The aim of the study was to evaluate the antifungal activity of leaf extracts of nine tree
species selected based on their use in respiratory and other disorders in traditional systems of
medicine and then to isolate and characterize the bioactive compound from the most active
plant species.
OBJECTIVES
The present study was undertaken with the following objectives:
Screening of various extracts of plants to evaluate their in vitro antifungal activity.
Isolation and purification of antifungal molecule from plants having in vitro activity
against pathogenic fungi.
Evaluation of antifungal potential of isolated molecule.
Studies on toxicity of purified molecule.
Biochemical characterization of purified molecule.

CHAPTER -3
REVIEW OF LITERATURE......

There have been continuous efforts for the improvement of antifungal therapies over
the last 30 years, the phenomenon of antifungal resistance is still of major concern in clinical
practice. With increasing numbers of immune-compromised patients with malignancy,
hematologic disease, and HIV, as well as those receiving immunosuppressive drug regimens
for the management of organ transplantation or autoimmune inflammatory conditions, the
incidence of fungal infections has dramatically increased over recent years. Although
invasive fungal diseases are now more frequent than during the first half of the century, they
are still difficult to diagnose clinically. During the latter half of the century, particularly
during the past two decades, a number of different classes of antifungal agents have been
discovered. Although, since the discovery of amphotericin B, there has been much progress
in this field, there is still a critical need for new antifungal agents to treat life threatening
invasive mycoses. The differences between the human host cells and the fungal pathogen
cells, both of which are eukaryotic, and have same molecular processes; therefore, there is
always the risk that what is toxic to the fungal cells will be toxic to the host cells (Wilson et
al., 2002). That's why; there is a great demand for novel antifungals belonging to a wide
range of structural classes, selectively acting on new targets with fewer side effects.
Therefore, one approach might be the testing of traditionally medicinal plants for their
antifungal activities as potential sources for drug development.
The present review is a brief account of the research works of various scientific
groups about the conventional and new strategies currently undertaken to discover alternative
therapy targets and antifungals.
3.1. Fungal Infection: A Renewed Threat
There are about 300 fungal species which have been reported to be pathogenic to
humans (Hay, 2006). Disseminated fungal infections in the hospital settings are not rare any
more as approximately 200,000 cases are reported every year from developed countries
which are considered to be associated with a high mortality rate of upto more than 40 %. The
fatal infections are mainly due to species of Candida and Aspergillus. While, the
retrospective analysis of US mortality trends due to invasive mycotic diseases conducted by
McNeil et al (2001) showed a marked and steady increase in mortality due to mycoses other
than Candida and Aspergillus during the period from 1980 to 1997. Overall mortality due to
other-mycoses has been found to be increased by 329 % from 0.14 to 0.60 deaths per 100,000
people during the study period. Hence, fungi such as Fusarium, Pnemocystis, Zygomycetes
or Penicillium which were normally considered to be non pathogenic, are emerging as a
threat to public health.

Mycotic infections are classified broadly into three groups namely superficial,
subcutaneous and systemic infections. A brief description of different groups of these
infections has been given below:
3.1.1. Superficial Infections
Superficial fungal infections are among the most common skin diseases affecting
millions of people throughout the world. These infections, which occur in both healthy and
immunocompromised persons, are caused by dermatophytes, yeasts and nondermatophyte
molds. The common dermatophytes that cause superficial skin infections in humans include
Trichophyton, Epidermophyton and Microsporum species. The disorders caused by
dermatophytes are commonly known as tinea corporis, tinea pedis, tinea cruris, tinea
unguum and tinea capitis, etc (Tosanger and Crutchfield, 2004).
In infected individuals, the yeast usually occurs in moist, occluded sites e.g. under
breasts, axillae or between buttocks. The skin folds between fingers of hands and toes of feet
are other very common sites of infections (Mandell et al., 1994). The Candida albicans is
common yeast that has been found to cause superficial infections. Candidal skin infections
often present with erythema, cracking or maceration (Hay, 1996). Malassezia is yeast which
can cause superficial infections. Malassezia furfur (Pityrosporum orbiculare and
Pityrosporum ovale) are reported to cause pityrosporum folliculitis and pityriasis (tinea)
versicolor which is a chronic, asymptomatic condition with well marked depigmented scaling
patches found mainly on the trunk (Inamadar and Palit, 2003).
Although the skin and nail infections due to molds are less common, they often cause
indolent infections in healthy or immunocompromised individuals, especially the elderly
people. The soil contaminated with infective organisms such as Scopulariopsis brevicaulis,
Fusarium, Aspergillus, Alternaria, Acremonium, Scytalidinum dimidiatum (Hendersonula
toruloides), Scytalidinum hyalinum has been considered to be the important source of
infections (Escobar and Carmona-Fonseca, 2003). Molds may cause ophthalmologic diseases
such as keratitis. The keratitis caused due to infections with Fusarium, Aspergillus and
Alternaria species have been common problems in India and other parts of the world (Guarro
et al., 2003).
3.1.2. Subcutaneous Fungal Infections
There have been three general types of subcutaneous fungal infections:
chromoblastomycosis, mycetoma and sporotrichosis. All these infections appear to be caused
by traumatic inoculation of the etiological fungi in to the subcutaneous tissue.
Chromoblastomycosis has been subcutaneous mycoses characterized by verrucoid lesions of

the skin, usually of the lower extremities. Histological examination of lesions revealed
muriform cells with perpendicular septations or so-called "copper pennies" that are
characteristic of this infection (Sayal et al., 2002). The most common causes of
chromoblastomycosis are Fonsecaea pedrosoi, F. compacta, Cladosporium carionii and
Phialophora verrucosa. These infections generally remain limited to the subcutaneous tissue
with no involvement of bones, tendons or muscle cells. Many of the fungi causing mycetoma
are pigmented brown to black. The causes of mycetoma have been quite diverse but could be
classified as eumycotic and actinomycotic mycetomas. The most common causative agents of
eumycotic mycetoma and actinomycotic mycetoma in United States have been reported to be
Pseudallescheria boydii and Nocardia brasiliensis respectively (Mariat et al., 1977). These
organisms are known as dematiaceous (melanized) fungi. The melanin pigment is deposited
in the cell walls of these organisms. These fungi may produce a range of infections from
superficial to subcutaneous to deep (visceral) infection characterized by the presence of
dematiaceous hyphal and/or yeast-like cells in tissues (Rippon, 1988). Sporotrichosis has
been the third general class of subcutaneous mycoses. The main causative agent of
sporotrichosis has been the Sporothrix schenckii which necrotizes the subcutaneous tissues at
the site of traumatic inoculation and finally causes nodular lymphangitis (Tomimori-
Yamashita et al., 1998). The infection usually spreads along cutaneous lymphatic channels of
the extremities involved.
3.1.3. Systemic Fungal Infections
The systemic fungal infections can either be worldwide in distribution or restricted to
some geographic region of globe. Histoplasma capsulatum, Cryptococcus neoformans and
Aspergillus sp. are worldwide in distribution. However, some of the pathogenic fungi such as
Coccidioides and Paracoccidioides have been found in the American continent only.
Penicilliosis marneffei is so far known only from Southeast Asia. Blastomyces dermatitis has
been found in India, America, Africa and other Asian countries. It has been reported that
endemic zone of blastomycosis has extended during the last 15 years from American and
Africa to Asia.
Pathogenic fungal organisms might gain entry through lungs, gastrointestinal tract or
intravenous route to cause invasive infections. Immunocompromised patients generally
remain at risk of systemic diseases; however, healthy individuals also may develop invasive
infections. Moreover, invasive fungal pathogens can be divided into two general classes,
primary pathogens and opportunistic pathogens (Pisseri et al., 2009). Fungi in the former
class usually have an environmental reservoir and infect individuals who have either been

exposed to a large dose or who are immunologically naive to the fungus. Opportunistic
pathogens take advantage of debilitated or immunocompromised hosts to cause infection.
They may have an environmental reservoir e.g., Cryptococcus neoformans, Aspergillus
fumigatus or exist as commensals in healthy organisms e.g., Candida species (Morgan et al.,
2005).
The opportunistic mycoses constitute a large category of fungal diseases. There are
etiological agents, which are ordinarily incapable of causing diseases in healthy individuals
but turn potential pathogens in immunocompromised or debilitated patients. Species of
Candida and Aspergillus are classical examples of opportunistic fungi, which are reported
from world over being very common in India. The number of opportunistic fungal diseases is
very large and ever increasing. Opportunistic human fungal pathogens have become
increasingly important over the past 20 years, paradoxically because the success of modern
medical practice has led to the survival of debilitated and immunosuppressed patients (Hay,
2006). Such patients are highly susceptible to infections by opportunistic pathogens such as
Candida species, C. neoformans, A. fumigatus and other Aspergillus species, and the
zygomycetes.
3.2. Fungal Pathogens: An Emerging Problem Agent
Fungi have emerged as the most common pathogens in causing infections particularly
in patients with a reduced defence such as those with haematological malignancies, transplant
recipients, cancer, diabetes, HIV and other immunodeficiencies (Pisseri et al., 2009). Since
opportunistic mycoses pose a serious threat to such patients, it is anticipated that these
infections may break out in epidemic proportions. While aspergillosis, candidiasis,
zygomycosis and infections with Fusarium species may be encountered with greater
frequency in neutropenics or in organ/bone marrow transplant recipients, cryptococcosis and
histoplasmosis are likely to be found often in patients with AIDS (Rhiannon, 2002). Besides,
the infections due to pathogenic fungi under immunocompromised conditions often become
invasive which are generally untreatable and fatal. Invasive fungal infections, particularly
in immunosuppressed patients, have continued to increase in incidence during the
past 30 years and invasive aspergillosis causes approximately 30% of invasive fungal
infections in patients. The overall mortality due to invasive aspergillosis has been around 85
% which may fall to 50 % if aggressively treated (Denning, 1996). The new drugs in trial
may further reduce the mortality but only moderately and still a very large portion of patients
remain at risk of fatal consequences (Ullmann, 2003). Moreover, invasive aspergillosis has
overtaken candidiasis as the most frequent fungal pathogen detected post mortem in tertiary

care hospitals in Europe (Vogeser et al., 1999). It was observed on autopsy that 4 % of all
patients died, had invasive aspergillosis as compared to about 2 % with invasive candidiasis
(Groll et al., 1996). Despite a better understanding of the epidemiology of Aspergillus
infections, diagnostic limitations persist. The Aspergillus has more than 260 species (Samson
and Varga, 2009), however, only 4 of them, namely A. fumigatus, A. flavus, A. niger and A.
terreus have been considered mainly to be pathogenic to humans. A. nidulans has been
isolated as the less common opportunistic mould among pathogenic species of Aspergillus.
Of these species, the A. fumigatus has been reported to be responsible for more than 90% of
Aspergillus induced infections and the marked predominance of A. fumigatus on clinical
samples reflects its environmental preponderance over other pathogenic species of
Aspergillus (Latge, 2001).
3.2.1. Aspergillus fumigatus: Ecological Significance
A. fumigatus has been a saprophytic fungus that plays an essential role in recycling
environmental carbon and nitrogen (Gugnani, 2003). The soil has been considered as the
natural ecological niche of A. fumigatus, where it survives and grows on organic debris. The
A. fumigatus has been very frequently found in the human habitats (Hirsch et al., 2000)
including pillows (Woodcock et al., 2006) and in the epicentre of vegetable matter compost
(Ryckeboer et al., 2003). The hyphae of A. fumigatus develop conidiophores and each of
which may produce large number of tiny (2.0 to 3.0 μm) green brown conidia (Levdansky et
al., 2007). The conidia released into the atmosphere become airborne as micro particles both
indoors and outdoors. The A. fumigatus does not have an elaborated mechanism for releasing
its conidia into the air and their dispersal depends simply on disturbances in the environment
and the air currents. Environmental surveys have indicated that all humans will inhale at least
several hundred A. fumigatus conidia every day and they counter with the immune system of
the host. In susceptible individuals, the germination of A. fumigatus conidia in the airways
causes a wide spectrum of diseases.
3.2.2. A. fumigatus as a Human Pathogen
Among the human pathogenic species of Aspergillus, the A. fumigatus has been found
to be primary causative agent of human infections, followed by A. flavus, A. terreus, A. niger
and A. nidulans (Morgan et al., 2005). Though A. fumigatus can establish infection in any
organ of the body, the involvement of lung has been the most common due to uptake of
conidia by inhalation (O'Gorman et al., 2009) (Figure 3.1). Repeated exposure to conidia of
A. fumigatus may cause allergic conditions, including asthma, allergic sinusitis, and
alveolitis. The A. fumigatus may invade the lung tissues and disseminate to the deeper body

parts to cause invasive aspergillosis. The invasive aspergillosis has been considered to be the
most devastating form of aspergillosis which very frequently occurs in severely
immunocompromised patients (Post et al., 2007). The mortality rate in invasive aspergillosis
has been reported up to 90% in high-risk populations and to be dependent on factors such as
host immune status, the site of infection, and the treatment regimen applied (Nicolle et al.,
2011).
Figure 3.1- Life cycle of pathogenic A. fumigatus showing infection of human host by
conidia.
3.3. Biochemical Targets for Antifungal Chemotherapy
Fungal cells are complex organisms that share many biochemical targets with other
eukaryotic cells. Thus fungal and human cells are similar at the molecular level. This makes
it more difficult to find or design drugs that target fungi without affecting human cells. As a
consequence, many antifungal drugs cause side-effects (Neely and Ghannoum, 2000).
Therefore, agents that interact with fungal targets (Figure 3.2) not found toxic in eukaryotic
cells are needed. The fungal cell wall is a unique organelle that fulfills the criteria for
selective toxicity. The fungal cell wall differs greatly from the bacterial cell wall and is not
affected by antibacterial cell wall inhibitors such as the β-lactams or vancomycin (Sable et
al., 2008). Arrangement of the biomolecular components of the cell wall accounts for the
individual identity of the organism. Although, each organism has a different biochemical

composition, their gross cell wall structure is similar. There are three general mechanisms of
action for the antifungal agents: cell membrane disruption, inhibition of cell division and
inhibition of cell wall formation.
Figure 3.2- Biochemical targets for various antifungal agents used in current chemotherapy.
Interference with fungal cell wall biosynthesis has not been as successful and
effective as penicilins and cephalosporins against bacteria. Many chemicals have been
discovered that interfere with various steps in fungal cell wall synthesis with excellent
antifungal activity in vitro (Neely and Ghannoum, 2000). Unfortunately, development of
these agents into useful drugs has proven very difficult. Many of these agents are developed
to target β-glucan synthesis. Moreover, antifungal agents that disrupt the cell membrane do so
by targeting ergosterol, either by binding to the sterol, forming pores and causing the
membrane to become leaky (as with polyene antifungals), or inhibiting ergosterol
biosynthesis (as seen with azole antifungal agents) (Milhaud et al., 2002). Ergosterol is
similar to mammalian cholesterol, thus agents binding ergosterol may have a cytotoxic effect
in the host tissue. Ergosterol has two conjugated double bonds that are lacking in mammalian
sterols. Furthermore, Nucleoside antifungal agents affect cell division by targeting the
microtubule effects in forming the mitotic spindle.
3.4. Conventional Antifungal Drugs and their Clinical Limitations
Until the 1940s, relatively few agents were available for the treatment of systemic
fungal infections (Wingard and Leather, 2003). The development of the polyene antifungals,
nystatin in the late 1940s, and the broader spectrum, more effective amphotericin B by the

late 1950s represented a major advance in the treatment of fungal infections (Sutton et al.,
2004). Until 1990 there was only one drug useful for treatment of invasive Aspergillus
disease, amphotericin B, which has to be given intravenously (Chamilos et al., 2008).
However, its clinical use is associated with numerous adverse effects related to both the drug
(nephrotoxicity) and its administration (fever, rigors, and hypotension). Additionally,
mortality associated with systemic fungal infections remains unacceptably high despite the
use of amphotericin B. The search for newer systemic antifungals led to the discovery of the
azoles several decades later, with the release of ketoconazole in the early 1980s followed by
fluconazole and itraconazole in the early 1990s. In 1990 itraconazole capsules became
available that included Aspergillus species in the spectrum although the drug was mainly used
in the prophylactic setting due to poor bioavailability (Naschimento et al., 2003). Ten years
later an intravenous formulation of itraconazole became available allowing the drug to be
used for the empiric or pre-emptive treatment of high risk patients. With the
registration of voriconazole the arsenal of drugs available has further increased (Pastor and
Guarro, 2007). Voriconazole is the newest azole antifungal agent for the treatment of
systemic mycosis. Voriconazole was developed as part of a program designed to enhance the
potency and spectrum of activity of fluconazole (Alexander et al., 2005). These agents were
available in oral formulation and demonstrated a relatively improved safety profile compared
with that of amphotericin B. Due to the azoles fungistatic nature, the gold standard for serious
systemic infections continues to be amphotericin B; however, a clear role for the azoles has
been established in the treatment of mild to moderate and refractory infections (Kretzer et al.,
2006). However, despite antifungal therapy, mortality in patients with invasive
aspergillosis remains very high and clearly new therapeutic approaches are needed. The
various agents available to treat fungal infections include
(a) Polyenes: Amphotericin B, Nystatin.
(b) Azoles: Clotrimazole, Miconazole, Ketoconazole, Fluconazole, Itraconazole.
(c) Echinocandins: Micafungin, Caspofungin, Pneumocandins
(d) Nucleoside analogues: Flucytosine
(e) Other antifungal Agents: Cationic peptides, Sordarins, Allylamines and thiocarbamates,
Pradimicins and benanomicins, Nikkomycins.
3.4.1. Polyenes
The polyene antifungal agents form complexes with ergosterol and disrupt the
fungal plasma membrane, resulting in increased membrane permeability, leakage of
the cytoplasmic contents and death of the fungal cell (Zotchev, 2003). Thus, the

polyenes are fungicidal and have the broadest spectrum antifungal activity. They are also
somewhat less toxic to mammalian cells because the affinity of the polyenes for ergosterol
in fungal cells is higher than the affinity for cholesterol in mammalian cells (Bruheim
et al., 2004). The minimum inhibitory concentration of amphotericin B ranges from 0.03 to
30.0 µg/ml against various fungal species (Sabatelli et al., 2008). Amphotericin B was
isolated from a strain of Streptomyces nodosus recovered from a soil sample obtained at
Tembladora on the Orinoco River in Venezuela by Gold and colleagues. Brown & Hazen
discovered nystatin in 1949 in a soil sample, obtained from a farm in Virginia, USA,
containing a strain of Streptomyces noursei (Zotchev, 2003). The minimum inhibitory
concentration of Nystatin ranges from 1.56 to 6.25 ug/ml (Fjaervik and Zotchev, 2005).
3.4.2. Azoles
The initial azole compounds were the imidazoles (clotrimazole, miconazole and
ketoconazole), which were then followed by the triazoles, fluconazole and itraconazole
(Diekema et al., 2003). The azoles inhibit fungal cytochrome P450 3A-dependent C14-a-
demethylase which is responsible for the conversion of lanosterol to ergosterol
(Rodriguez–Tudela et al., 2008). This leads to the depletion of ergosterol in the fungal cell
membrane. Clotrimazole, discovered in 1969,
cannot be given parenterally, has poor
oral absorption and is used for the treatment of oral and vaginal candidosis. The
minimum inhibitory concentration of Clotrimazole ranges from 1.0 to 10.0 µg/ml.
Miconazole was also discovered in 1969 and also has poor oral bioavailability.
Miconazole has been found to be active against a wide range of fungi and its MIC against
different organisms varies from 0.25 to 8.0 µg/ml (Panackal et al., 2006). Ketoconazole was
discovered in 1978, and has good oral absorption, a broad spectrum of activity and low
toxicity, although it may be hepatotoxic and does produce endocrine abnormalities by
suppression of testosterone and ACTH-stimulated cortisol synthesis (Kretzer et al., 2006).

The minimum inhibitory concentration of Ketoconazole ranges from 0.02 to 128.0 µg/ml
against various fungal species.
Fluconazole was formulated in 1981. It is a novel bistriazole, is metabolically stable
and water soluble and has low lipophilicity and plasma protein binding (Garey et al.,
2006). It is well tolerated and has a very low incidence of side effects and a broad spectrum
of antifungal activity, except against Aspergillus spp. Itraconazole, discovered in 1986, is
another triazole antifungal agent with broad-spectrum antifungal activity, including
activity against Aspergillus spp (Wheat et al., 2006). It is very insoluble, is only available in
oral form and may be given once daily. The minimum inhibitory concentrations of both the
drugs (Itraconazole and Fluconazole) have been found to ranges from 0.12 to 2.0 µg/ml
(Sabatelli et al., 2008).

3.4.3. Echinocandins
The echinocandins are cyclic lipopeptide fungicidal agents. They act by preventing
cell wall synthesis by non-competitive inhibition of 1,3-beta-D-glucan synthase, an enzyme
which is absent in mammalian cells (Balani et al., 2000). This inhibition is highly specific
and brief exposure to these drugs leads to cell death. Micafungin is a natural antifungal
product derived from other fungi as a defense mechanism for competition of nutrients, etc.
More specifically, micafungin is produced by Coleophoma empetri. Compared to
amphotericin B, caspofungin seems to have a relatively low incidence of side effects (Balani
et al., 2000). Both are administered intravenously.
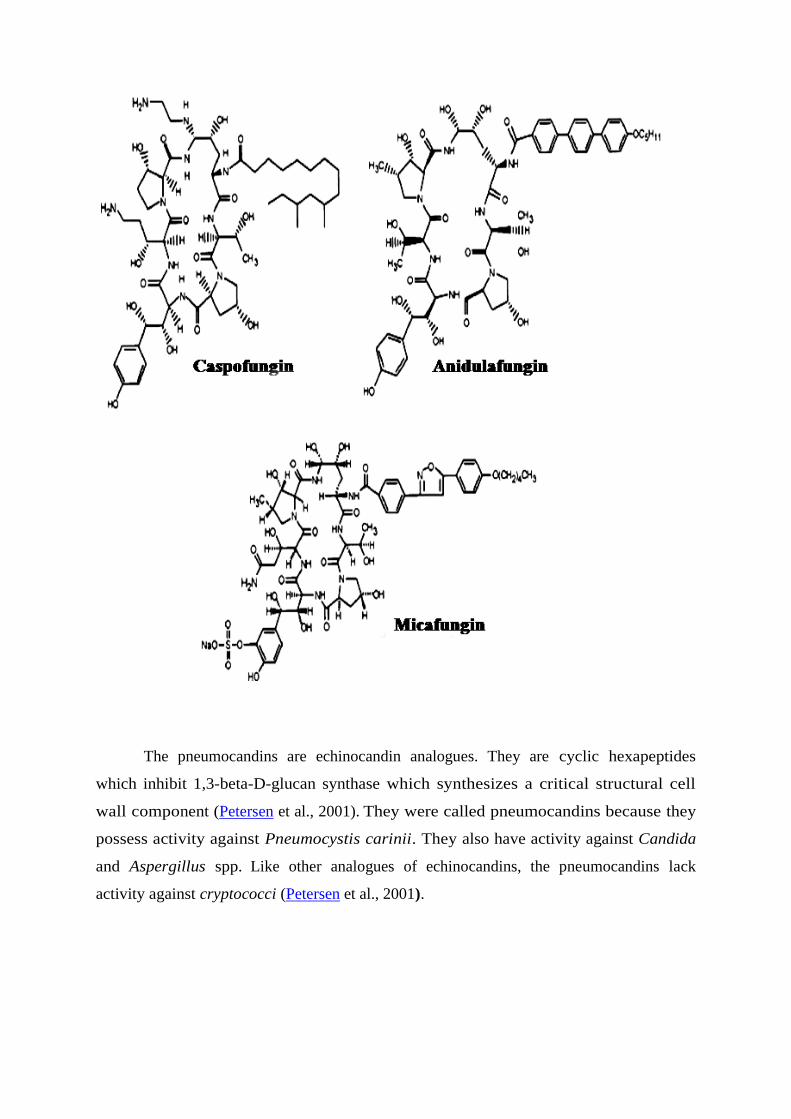
The pneumocandins are echinocandin analogues. They are cyclic hexapeptides
which inhibit 1,3-beta-D-glucan synthase which synthesizes a critical structural cell
wall component (Petersen et al., 2001). They were called pneumocandins because they
possess activity against Pneumocystis carinii. They also have activity against Candida
and Aspergillus spp.
Like other analogues of echinocandins, the pneumocandins lack
activity against cryptococci (Petersen et al., 2001).

3.4.4. Nucleoside analogues
5-Fluorocytosine (flucytosine, 5-FC) is a fluorinated pyrimidine that was
synthesized in 1957 as a cytosine analogue for the treatment of leukaemia; however, it
was ineffective because it had no cytotoxic activity. The antifungal activity of 5-
fluorocytosine was discovered later and was reported in 1963 in a murine model of
candidosis (Thomas and Katiyar, 2010). 5-Fluorocytosine had low activity against
Aspergillus species and the MIC was recorded greater than 100.0 µg/ml. It inhibits
pyrimidine metabolism by interfering with RNA and protein synthesis in the fungal cell.
Flucytosine enters fungal cells via cytosine permease and is deaminated to 5-
fluorouracil, which is incorporated into RNA. Uridine 5-monophosphate
pyrophosphorylase then converts 5-fluorouracil into fluorodeoxyuridine
monophosphate, which inhibits thymidylate synthetase and interferes with DNA
synthesis. This drug is selectively toxic to fungi because mammalian cells lack
cytosine permease and do not convert large amounts of flucytosine to 5-fluorouracil.
3.4.5. Other Antifungal Agents
The search for new antifungal agents has been expanded as progress in molecular
biology has led to a better understanding of important and essential pathways in fungal
cell growth and multiplication (Borowski, 2000). One group includes cationic peptides,
both naturally occurring and synthetic derivatives, bind to ergosterol and cholesterol in
fungal cell membranes, ultimately leading to cell lysis (Kretzer et al., 2006). These
peptides have antifungal activity against Aspergillus spp., Candida spp., C. neoformans
and Fusarium spp. Naturally occurring cationic peptides include cecropins, dermaseptins,
indolicin, histatins, bactericidal permeability-increasing factor (BPI), lactoferrin and
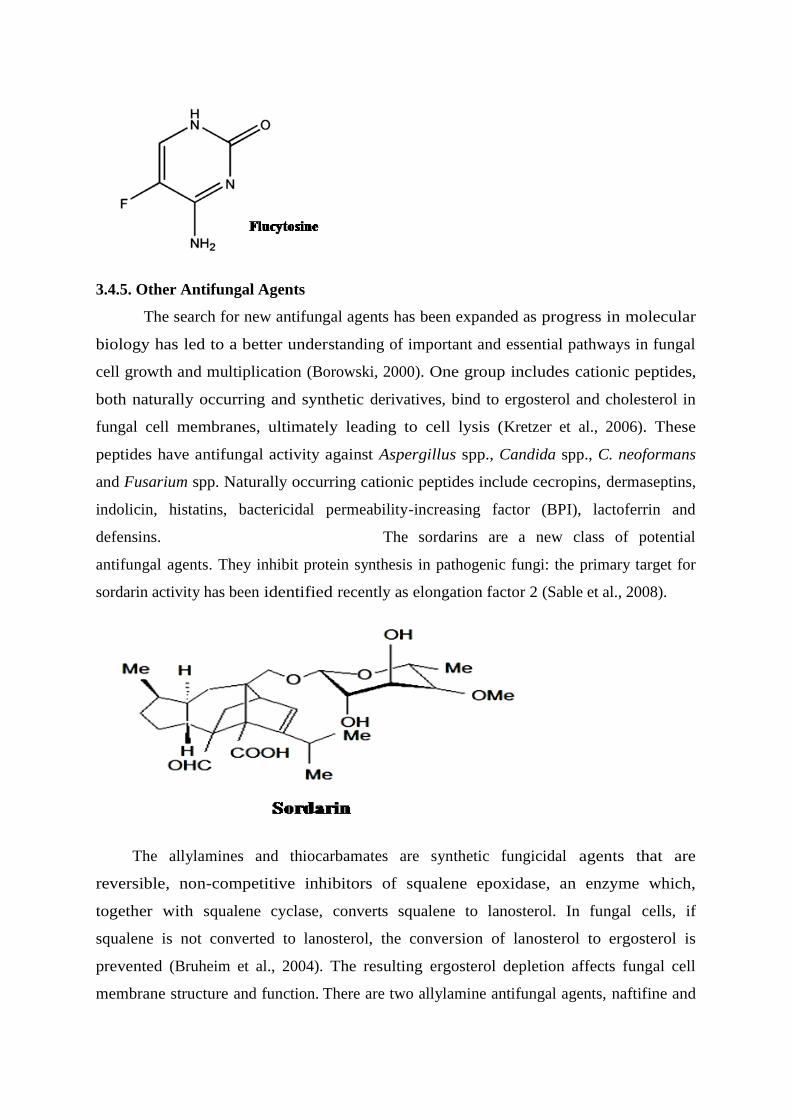
defensins. The sordarins are a new class of potential
antifungal agents. They inhibit protein synthesis in pathogenic fungi: the primary target for
sordarin activity has been identified recently as elongation factor 2 (Sable et al., 2008).
The allylamines and thiocarbamates are synthetic fungicidal agents that are
reversible, non-competitive inhibitors of squalene epoxidase, an enzyme which,
together with squalene cyclase, converts squalene to lanosterol. In fungal cells, if
squalene is not converted to lanosterol, the conversion of lanosterol to ergosterol is
prevented (Bruheim et al., 2004). The resulting ergosterol depletion affects fungal cell
membrane structure and function. There are two allylamine antifungal agents, naftifine and

terbinafine, and one thiocarbamate, talnaftate. Naftifine is a topical preparation whereas
terbinafine is an oral systemic agent (Borowski, 2000). The allylamine, naftifine, is
considered an effective topical agent for treatment of dermatophytes infections of the skin.
Similarly, the pradimicins and benanomicins are fungicidal compounds. They
appear to bind, in a calcium-dependent manner, to cell wall mannoproteins and this causes
osmotic lysis and leakage of intracellular contents, particularly potassium, ultimately leading to
cell death.
The nikkomycins are competitive inhibitors of fungal chitin synthase enzymes
which are necessary for fungal cell wall synthesis (Sable et al., 2008). Chitin is a
linear polymer of â-(1,4)-linked N-acetylglucosamine residues and is synthesized on
the cytoplasmic surface of the plasma membrane. Chitin synthase catalyses the
polymerization of N-acetylglucosamine in the formation of chitin.

Also, we need to learn more about combination antifungal therapy, e.g. about
the effects of sequential blockade at two or more sites, and about the combination of
antifungal agents with cytokines in an attempt to augment the inflammatory and immune
responses of patients (Baddley and Pappas, 2005).
3.4.6. Limitations of Conventional Approaches
The molecules available for treatment of aspergillosis are not only fewer in number,
they have limited efficacy. The amphotericin B which was developed in early 1960s still
remains a preferred drug inspite of the fact that it has been highly toxic to the host (Bruheim
et al., 2004). A new antifungal agent called caspofungin was launched in 2002 and included
in the existing therapeutic options for candidiasis and invasive aspergillosis in patients
refractory to, or intolerant of, treatment with other drugs. Caspofungin showed good activity
against Aspergillus and Candida, however, its intravenous route of administration remained
an unsolved problem. Other anti-Aspergillus drugs also have been found to cause toxic
effects (Howard and Arendrup, 2011). Most drugs used for treating aspergillosis have been
shown to be immunosuppressive (Steinbach et al., 2004). The development of drug resistance
in A. fumigatus has become an increasingly serious problem (Camps et al., 2012) which has
made treatment of aspergillosis almost impossible with limited available drugs. The drug
resistance can be primary (fungal pathogen inherently resistant to antifungal drug) or
secondary (development of resistance during treatment). The resistance in Aspergillus to
polyenes has been shown to be primary rather than secondary resistance. The primary
resistance to polyenes in A. fumigatus has been less frequent (Masia et al., 2002) than in
A. terreus. Most A. terreus isolates were found to be resistant to amphotericin B in vitro but
were susceptible to itraconazole or voriconazole (Sutton et al., 2004). There are reports which
have demonstrated resistance in Aspergillus species against voriconazole among stem cell
transplant recipients (Siwek et al., 2004). The development of resistance in A. fumigatus

against most of antifungal drugs makes chemotherapy of aspergillosis less efficient;
therefore, immunotherapy by using vaccine may be important for the better management of
aspergillosis.
3.5. Phytopharmaceutical Approaches and their Significance
3.5.1. Historical Perspectives
The medicinal plants have been one of the important sources of
medicines since the dawn of human civilization. According to medicinal records, in India,
the earliest mention about the use of medicinal plants is to be found in the Rig-Veda which was
written between 4500-1600 B.C (Padma, 2005). The Materia Medica of Hippocrates, who
is now referred to as the father of medicine consisted essentially of herbal recipes, some 400
simple remedies having been compiled and described by him. Theophrastus of Athens
(370-287 B.C.) was another famous biologist-botanist who produced a number of
manuscripts including the famous Historia Plantarium. About 500 plants, mostly cultivated,
were described in this manuscript. Pliny, the elder (23-79 A.D.), a Roman naturalist and
philosopher, described 1000 plants with their medicinal properties, anatomy and
horticultural practices in his book, Historia Naturalis. Dioscorides (60 A.D) wrote “De
Materia Medica” describing 600 plant species of medicinal value from Mediterranean
region. In the middle Ages, the writing of Galen (131 A.D.) becomes popular. Galen is
considered today to be the most distinguished physician of antiquity after Hippocrates. He treated
diseases essentially by the use of herbs. Allopathic as
well as homeopathic systems of medicine today are based on the doctrine explained by Galen.
In the early stages, the science of medicine developed around those plants which
had curative properties. A continued search for medicinal plants during the last several
centuries has given rise to a long list of plants which are of great use in the treatment of
diseases, and for promoting health. It can be stated, more or less truthfully, that every disease
has a cure in a plant growing in nature. In ancient times, medicinal plants were chosen for
their colour or the shape of their leaves (Park, 2008). For example, heart-shaped leaves were
used for heart problems, while plants with red flowers were used to treat bleeding disorders. The
formal study of herbs, called herbology, dates back to the ancient cultures revered the power of
nature and developed herbal remedies based on plants found in their in home
environments (Kamboj, 2000). Herbal therapy is also a major component of India’s
Ayurvedic medicine, traditional Chinese medicine, Native American medicine, homeopathy,
and naturopathy (Patwardhan et al., 2003).

3.5.2. Current Status of Herbal Medicine
A total of 122 biologically active compounds have been identified derived only from
94 species of plants. A conservative estimate of the number of flowering plants occurring on
the planet is 250,000. Of these, only about 6% have been screened for biological activity and
a reported 15% have been evaluated phytochemically (Turker and Usta, 2008). This means
that about 3.5 to 4 billion people in the world rely on plants as a source of drugs. According to
World Health Organisation (WHO), about 80% of the world’s population, primarily those of
developing countries rely on plant-derived medicines for their healthcare (Gurib-Fakim,
2006). The world Health Organization is now actively encouraging developing countries to
use herbal medicine which they have been traditionally used for centuries. They have
identified 3000 plants from the forests of India and other tropical countries which can be used as
medicine. The active ingredients from these plants are worth nearly Rs. 2000 crores of rupees for
the US market alone and nearly 8 times that for the world market.
India has a rich diversity of medicinal plants and only a relatively small number of
plants species have been scientifically validated, some of indigenous plant species may
become extinct before their potential as sources of pharmaceutical drugs is investigated and
applied (Kong et al., 2003). Some of the medicinal plants that had been used traditionally are
becoming endangered, rare, or threatened due to unsustainable harvesting methods (Ming et
al., 2003). Other factors such as exposure to modern culture and urbanization also play a
role in the loss of the traditional use of plants. If plant leaf extracts are as effective as the
bark, bulbs or roots, these endangered species can be utilized in a sustainable way. Plants
have the major advantage of still being the cheapest and most effective alternative source of
drugs. Natural products, either as pure compounds or as standardized plant extracts, provide
unlimited opportunities for new drug leads because of the unmatched availability of chemical
diversity (Cos et al., 2006). Since plants produce a variety of compounds with antifungal
properties, screening plant leaf extracts and isolation of antifungal compounds may become
the base for the development of a medicine, a natural blueprint for the development of new
drugs, or a phytomedicine to be used for the treatment of fungal infections (Kala and Sajwan,
2007).
3.5.3. Market Potential of Herbal Drugs
Herbal medicines are readily available in the market from health food stores without
prescriptions and are widely used all over the world. The utilization of herbal drugs is on the
flow and the market is growing step by step. The annual turnover of the Indian herbal
medicinal industry is about Rs. 2,300 crores as against the pharmaceutical industry’s turnover

of Rs. 14,500 crores with a growth rate of 15 percent (Sharma et al., 2008). The major
pharmaceuticals exported from India in the recent years are Isabgol, Opium alkaloids, Senna
derivatives, Vinca extract, Cinchona alkaloids, Ipecac root alkaloids, solasodine, menthol,
Gudmar herb, papian, Rauwolfia guar gum, Jasmine oil, Agar wood oil, Sandal wood oil, etc
(Kokate et al., 2005). The global market for herbal medicines currently stands at over $60
billion annually. The sale of herbal medicines is expected to get higher at 6.4% an average
annual growth rate (Inamdar et al., 2008). Herbal drugs are marketed in various forms. They
are available in both classical forms (tablets, powder, decoction, medicated oil, medicated
ghee, fermented products) and modern drug presentation forms like capsules, lotions, syrups,
ointments, creams, granules etc. There are more than 8500 manufacturers of herbal drugs in
India (Jain, 2001). Thus, plant-based therapeutic agents continue to have scientific, social,
and commercial significance and appear to be gathering a momentum in health relevant areas.
A study of the process by which the traditional or more recent plant-based molecular drugs or
the new breed of herbal drugs came to be used in present-day medicine reveals that, in over
70% of the cases, the starting point has been some reference to the use of that plant as an
indigenous cure in a folklore or traditional system of medicine of one culture or other.
3.5.4. Traditional Herbal Medicine in Medical Health Practices
During the last decade, the use of TM (traditional medicine) has expanded globally
and has gained popularity (Fabricant and Farnsworth, 2001). It has not only continued to be
used for primary health care of the poor in developing countries, but has also been used in
countries where conventional medicine is predominant in the National health care system.
The practice of traditional medicine is widespread throughout Asia including India, China,
Japan, Pakistan, Srilanka, and Thailand (Venkatasubramanian, 2007). In Japan, herbal
medicinal preparations are more in demand than mainstream pharmaceutical products. 60
to 70% of allopathic doctors in Japan prescribe TM for their patients. In Malaysia, traditional
forms of Malay, Chinese and Indian medicine are used extensively. China is the leading country
for incorporating traditional herbal medicine into a modern health care system. In this
country, TM accounts for around 40% of all health care delivered and are used to treat
roughly 200 million patients annually. According to a recent survey, almost 7,300 plants have
been used in traditional Chinese medicine (Venkatasubramanian, 2007). The traditional
system of medicine is so engrained in Indian culture that, even now 75% of the Indian
population depend on this indigenous system for relief (Ravishankar and Shukla, 2007). With
such a huge section of an ever-increasing population relying on herbal remedies, it is imperative
that the plant products which have been in use for such a long time be scientifically supported

for their efficacy.
The
potential toxicity of the traditional medicines is an important consideration when studying
their biological activities (McGaw et al., 2007). Plant extracts might be very toxic as they
contain many different compounds; therefore it is very important to investigate cytotoxicity
of both crude extracts and isolated compounds. The use of medicinal plants in the form of
crude extracts presents several difficulties. The amount of the bioactive compound(s) from
plants may vary with both the locality and the season in which they are collected. Also,
bioactive molecules of many plants are powerful poisons when taken in excess, and if the
plant extract contains a lower content of bioactive compound(s) than usual, suboptimal
dosage may not be effective. Medicinal properties of many plants are also rapidly lost on
storage, for example, Foxglove leaf’s bioactive molecules decompose on long storage, unless
dried quickly after collection. Furthermore, crude extracts from many medicinal plants may
contain, in addition to the bioactive molecules, other constituents which have harmful effects.
It is therefore important to isolate and identify the bioactive molecules from plant extracts.
The advantage of using pure drugs instead of crude plant extracts includes, amongst others,
accurately prescribed dosage. Structural modification of isolated and identified bioactive
compounds from plant extracts may allow an improvement in the efficacy and moderation of
side effects. Pure bioactive molecule can frequently be synthesized economically, thus
preventing dependence on plants as sources.
3.6. Re-emergence of Antifungal Herbs as the Treatment of Choice
Plants are the oldest source of pharmacologically active compounds, and have
provided humankind with many medically useful compounds for centuries. The primary
benefits of using plant derived medicines are that they are relatively safer than synthetic
alternatives, offering profound therapeutic benefits and more affordable treatment (Singh,
2007). It is estimated that more than two thirds of the world’s population relies on plant
derived drugs; some 7000 medicinal compounds used in the Western pharmacopoeia are
derived from plants. Several methods have been used to acquire compounds for drug
discovery including isolation from plants and other natural sources, synthetic chemistry,
combinatorial chemistry, and molecular modeling (Lombardino and Lowe, 2004). However,
natural products, and particularly medicinal plants, remain an important source of new drugs,
new drugs leads, and new chemical entities (Newman et al., 2003). The compounds have
provided the basic scaffold for medicinal chemistry modifications to expand the spectrum
and/or potency of improved analogues in subsequent years (Walsh, 2003).

3.7. Antifungal Compounds from Plants
Plants are storehouses of a wide variety of secondary metabolites, such as tannins,
terpenoids, alkaloids and flavonoids which have demonstrated their antimicrobial properties
in vitro (Rawat et al., 2008). Moreover, the plant derived fungicides are easily biodegradable
and selective in their toxicity. These antifungal metabolites can be performed in the plant, the
so-called 'constitutive antifungal substances' or they are produced by plants in response to an
infecting organism i.e., phytoalexins. In other words phytoalexins are chemical compounds
formed in plants via a metabolic sequence induced either biotically or in response to chemical
or environmental factors (Rawat et al., 2008). In many plants, a significant proportion of the
assimilated carbon and energy is diverted to the synthesis of organic molecules that may have
no obvious role in growth and development. These molecules are known as secondary
metabolites. The important secondary metabolite includes terpenoids, phenolic compounds,
saponins, cardiac glycosides, cyanogenic glycosides, glucosinolates and alkaloids.
3.7.1. Terpenoids having Antifungal Activity
A large number of studies have been done in recent years on the antifungal activity of
terpenoids of natural origin. These reports concern mainly sesquiterpenes and sesquiterpene
lactones. The fragrance of plants is carried in essential oil fraction. These oils are secondary
metabolites that are highly enriched in compounds based on an isoprene structure. They are
called terpenes, their general chemical structure is C10H16, and they occur as diterpenes,
triterpenes, and tetraterpenes (C20, C30, and C40), as well as hemiterpenes (C5) and
sesquiterpenes (C15). The mechanism of action of terpenes is not fully understood but is
speculated to involve membrane disruption by the lipophilic nature (Cheng et al., 2007). The
terpene family includes hormones, the carotenoid pigments, sterols and sterol derivatives,
latex and many of the essential oils that give plants their distinctive odours and flavours
(Adam et al., 2002). Monoterpenoid from essential oils are well known for their antifungal
activities. Iridoids are a group of monoterpenoid lactones, which usually occur as glycosides
since their aglycones tend to be highly unstable. However, antifungal activity of iridoids
appears to be associated with the few stable unglycosylated structures known. Sesquiterpene
lactones, showing a wide range of biological activities are found to be more active than
sesquiterpenes (Cheng et al., 2007) as well as many diterpenoids also show antifungal
activity. Another important source of antifungal triterpenoids is the saponins (Adam et al.,
2002). Saponins are terpene glycosides. They may be steroid glycosides, steroid alkaloid
glycosides or triterpene glycosides. The antifungal activity of saponins is usually correlated
with the sugar moiety glycosylated to the 3- hydroxy group of the triterpenoids, whereas most

other antifungal constituents tend to be strongly lipophilic and inactive in glycosidic form
(Chiang and Kuo, 2000). They are stored in plant cells as inactive precursors but are readily
converted into biological active compounds by enzymes in response to pathogen attack.
3.7.2. Nitrogenous Compounds having Antifungal Activity
Nitrogen first appears in organic form as glutamic acid, the key reaction being the
transfer of ammonia to a ketoglutarate, catalysed by glutamic dehydrogenase. The other
amino acids are subsequently synthesized from glutamic acid through the catalytic action of
nonspecific transaminases. Amino acids are involved in the biosynthesis of all other
nitrogenous plant compounds, from the proteins to alkaloids, amines, cyanogenic glycosides,
porphyrins, purines, pyrimidines and cytokiins (Bandaranayake, 2002). Alkaloids and amines
are nitrogen-containing compounds, which include representatives showing anti-fungal
activities. Heterocyclic nitrogen compounds are called alkaloids. The first medically useful
example of an alkaloid was morphine; isolated in 1805 from the Papaver somniferum,
codeine and heroin are both derivatives of morphine. The polyamines, spermidine and
spermine, which have a universal occurrence in plants inhibit spore germination of
Penicillium species. Both glucosinolates and cyanogenic glucosides are further groups of
nitrogen and sulphur containing plant compounds which occur in plants in an inactive form
called post-inhibitins. After plant damage these are transformed into the active
isothiocyanates and HCN, which are toxic to insects and microorganisms (Dewick and Paul,
2002).
3.7.3. Aromatic Compounds having Antifungal Activity
Many of the aromatic compounds are emerging as a major group of antifungal plant
compounds. They include simple and alkylated phenols, phenolic acids, phenyl propanoids,
coumarins, flavonoids, isoflavonoids, stiibenoids, quinones and xanthones (Begley and
Tadhg, 2009). Some of the simplest bioactive phytochemicals consist of a single substituted
phenolic ring. The site(s) and number of hydroxyl groups on the phenol group are thought to
be related to their relative toxicity to microorganisms, with evidence that increased
hydroxylation results in increased toxicity (Ross, 2005). In addition, it was also reported that
more highly oxidized phenols are inhibitorier. The mechanisms thought to be responsible for
phenolic toxicity to microorganisms include enzyme inhibition by the oxidized compounds,
possibly through reaction with sulfhydryl groups or through more nonspecific interactions
with the proteins. Many phenolic acids like benzoic, protocatechuic and gentisic acids have
been reported as constitutive antifungal compounds (Bandaranayake, 2002). Antifungal
phenyl propanoid or hydroxy cinnamic acids include p-coumaric, ferulic, caffeic, sinapic and

chlorogenic acids. All these compounds have a widespread distribution in plants. The
structure - activity relationship studies have proved that the aromatic group rather than the
carboxyl group seems to be necessary for the fungal inhibition.
Xanthones are a restricted group of plant polyphenols, biosynthetically related to the
flavonoids. These are planar-six carbon molecules in a conjugated ring system consisting of a
backbone molecule and various chemical groups attached to it (Dewick and Paul, 2002).
Xanthone backbone consists of two benzene rings attached through a carbonyl group and
oxygen not allowing free rotation about the carbon - carbon bonds. The unique backbone
along with type and position of the attached chemical groups defines specific properties of
xanthones. Xanthones possess numerous bioactive capability including antifungal properties.
Coumarins are phenolic substances made of fused benzene and α-pyrone rings which have a
phenyl propanoid nucleus, are another group of aromatic substances rich in antifungal
representations (Dewick and Paul, 2002). Coumarins have been reported to stimulate
macrophages which could have an indirect negative effect on infections. Examples are
coumarin, esculetin, hermiarin, scopoletin and umbelliferone all of which have a wide
distribution in higher plants. There are about twelve recognized
groups of flavanoids that differ from one another only by the oxidation state of this
heterocyclic ring. Three major groups of flavonoids are flavones, falvonols and
anthocyanidins (Begley and Tadhg, 2009). Flavones are phenolic structures containing one
carbonyl group and the addition of a 3 hydroxyl group yields a flavonol. Flavonoids are
hydroxylated phenolic substances synthesized by plants in response to microbial infection.
They have been found to be effective antimicrobial substances against a wide array of
microorganisms. Their activity is probably due to their ability to complex with extracellular
and soluble proteins and to complex with fungal cell walls. More lipophilic nature of
flavonoids may also disrupt fungal membranes (Ross, 2005).
One group of flavonoids, the isoflavonoids have become known for their
antimicrobial activities. lsoflavonoids are one of the several classes of chemicals of diverse
structures formed as phytoalexins. In addition to isoflavonoids; flavonoids, flavanones,
flavans, flavones, flavonols, certain biflavones, chalcones and dihydro chalcones are known
to be associated with antifungal activity (Dewick and Paul, 2002). Quinones are aromatic
rings with two ketone substitutions and characteristically highly reactive. They can switch
between diphenol (hydroquinone) and diketone (quinone) easily through oxidation and
reduction reactions. These compounds, being colored, are responsible for the browning
reaction in cut or injured fruits and vegetables (Ross, 2005). In addition to providing a source

of stable free radicals, quinones are known to complex irreversibly with nucleophilic amino
acids in proteins. Therefore the quinones inactivate the protein and impair their function.
Quinones bind with surface exposed adhesins, cell wall polypeptides, membrane-bound
enzymes and form complex which inactivate the enzymes.
3.7.4. Aliphatic Compounds having Antifungal Activity
These compounds form the simplest form of lipids; they contain only carbon and
hydrogen. These molecules are found mainly in petroleum but living organisms, eukaryotic
or prokaryotic, contain frequently hydrocarbons which are directly derived from fatty
acids. Several hydrocarbons (octane, nonane, dodecane, hexadecane.) belong to aroma
compounds which are found in environmental or food systems. Such as aliphatic C(17)-
polyacetylenes of the falcarinol type, which occur in common food plants of the Apiaceae
family such as Carrot, Celeriac, Parsnip and Parsley, have demonstrated interesting
bioactivities including antibacterial, antimycobacterial, and antifungal activity as well as anti-
inflammatory, anti-platelet-aggregatory, neuritogenic and serotonergic effects (Christensen,
2011). In addition, the cytotoxicity of falcarinol type polyacetylenes towards human cancer
cells, bioavailability, and their potential anticancer effect in vivo indicates that these
compounds may contribute to the health effects of certain vegetables and hence could be
important nutraceuticals (Christensen, 2011). Isolation
and characterization of pharmacologically active compounds from medicinal plants continue
even today. It has only been in the past ten years or so that interest in higher plant antifungal
agents has been reawakened worldwide, and the literature in this area is becoming
substantial. The use of medicinal plants for the treatment of fungal infections predates written
records. Thus, phytochemical screening of plants species, especially of ethnopharmaceutical
use, will provide valuable baseline information in the search for new pharmaceuticals.
Therefore, the presence of antifungal activities demonstrated in various extracts and
compounds of different plant species which used in traditional medicine practices have been
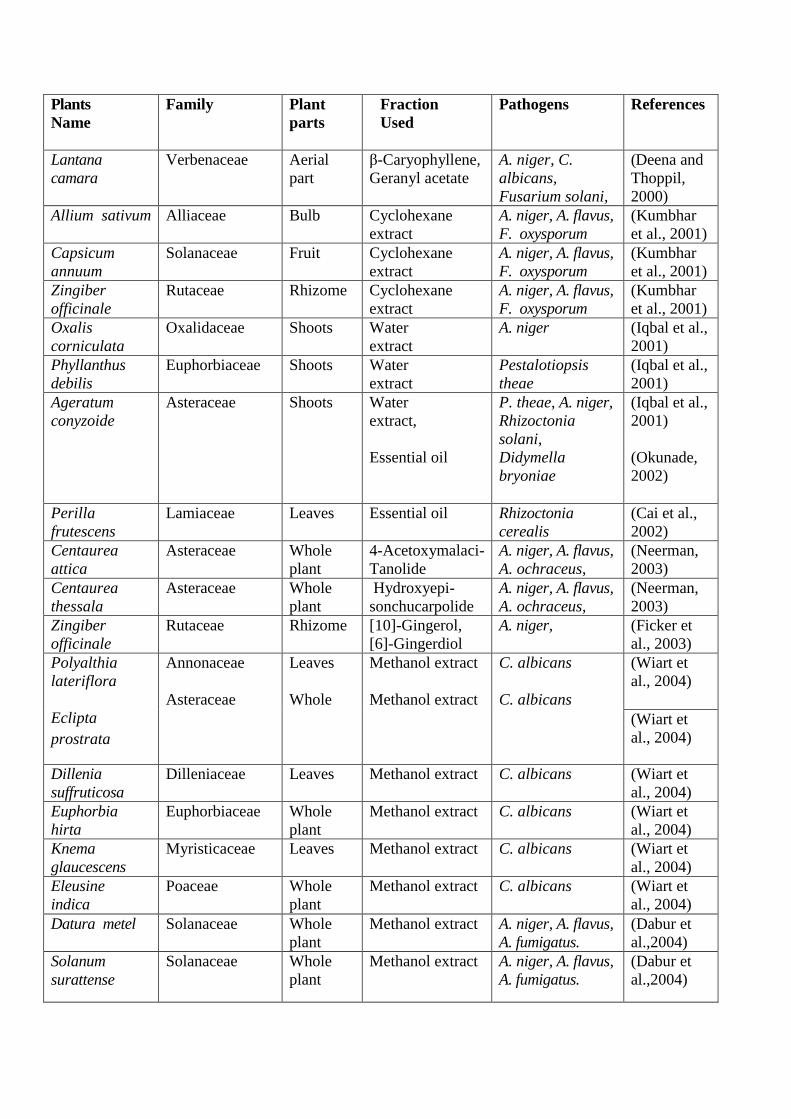
listed in Table 3.1.
Table 3.1- List of various selected plants with reported antifungal activity.
Plants
Name
Family
Plant
parts
Fraction
Used
Pathogens
References
Lantana
camara
Verbenaceae Aerial
part
β-Caryophyllene,
Geranyl acetate
A. niger, C.
albicans,
Fusarium solani,
(Deena and
Thoppil,
2000)
Allium sativum Alliaceae Bulb Cyclohexane
extract
A. niger, A. flavus,
F. oxysporum
(Kumbhar
et al., 2001)
Capsicum
annuum
Solanaceae Fruit Cyclohexane
extract
A. niger, A. flavus,
F. oxysporum
(Kumbhar
et al., 2001)
Zingiber
officinale
Rutaceae Rhizome Cyclohexane
extract
A. niger, A. flavus,
F. oxysporum
(Kumbhar
et al., 2001)
Oxalis
corniculata
Oxalidaceae Shoots Water
extract
A. niger (Iqbal et al.,
2001)
Phyllanthus
debilis
Euphorbiaceae Shoots Water
extract
Pestalotiopsis
theae
(Iqbal et al.,
2001)
Ageratum
conyzoide
Asteraceae Shoots Water
extract,
Essential oil
P. theae, A. niger,
Rhizoctonia
solani,
Didymella
bryoniae
(Iqbal et al.,
2001)
(Okunade,
2002)
Perilla
frutescens
Lamiaceae Leaves Essential oil Rhizoctonia
cerealis
(Cai et al.,
2002)
Centaurea
attica
Asteraceae Whole
plant
4-Acetoxymalaci-
Tanolide
A. niger, A. flavus,
A. ochraceus,
(Neerman,
2003)
Centaurea
thessala
Asteraceae Whole
plant
Hydroxyepi-
sonchucarpolide
A. niger, A. flavus,
A. ochraceus,
(Neerman,
2003)
Zingiber
officinale
Rutaceae Rhizome [10]-Gingerol,
[6]-Gingerdiol
A. niger, (Ficker et
al., 2003)
Polyalthia
lateriflora
Eclipta
prostrata
Annonaceae
Asteraceae
Leaves
Whole
Methanol extract
Methanol extract
C. albicans
C. albicans
(Wiart et
al., 2004)
(Wiart et
al., 2004)
Dillenia
suffruticosa
Dilleniaceae Leaves Methanol extract C. albicans (Wiart et
al., 2004)
Euphorbia
hirta
Euphorbiaceae Whole
plant
Methanol extract C. albicans (Wiart et
al., 2004)
Knema
glaucescens
Myristicaceae Leaves Methanol extract C. albicans (Wiart et
al., 2004)
Eleusine
indica
Poaceae Whole
plant
Methanol extract C. albicans (Wiart et
al., 2004)
Datura metel Solanaceae Whole
plant
Methanol extract
A. niger, A. flavus,
A. fumigatus.
(Dabur et
al.,2004)
Solanum
surattense
Solanaceae Whole
plant
Methanol extract
A. niger, A. flavus,
A. fumigatus.
(Dabur et
al.,2004)

Chenopodium
botrys
Chenopodiaceae Aerial
part
Essential oil A. niger,
C. albicans
(Maksimovi
c et
al.,2005)
Cynara
scolymus
Asteraceae Leaves,
Flower
Chloroform,
ethanol extract
A. niger,
C. albicans
(Zhu et al.,
2005)
Helichrysum
italicum
Asteraceae Whole
plant
Terpenoid,
Terpene oil.
C. albicans (Mastelic et
al., 2005)
Ailanthus
excelsa
Simaroubaceae Stem,
Bark.
Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Albizia lebbeck Mimosaceae Pod Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Ceratonia
siliqua
Mimosaceae Pod Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Gloriosa
superba
Liliaceae Root Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Ruta
graveolens
Rutaceae Leaves Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Sesbania
sesban
Fabaceae Leaves Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Solanum
indicum
Solanaceae Fruit Dichloromethane,
methanol extract.
C. albicans,
S. cerevisiae
(Kumar et
al., 2006)
Citrus
aurantifolia
Rutaceae Whole
plant
Essential oil A. niger (Verma et
al.,2007)
Citrus limon Rutaceae Whole Essential oil A. niger (Verma et
al.,2007)
Citrus
paradisi
Rutaceae Whole
plant
Essential oil A. niger (Verma et
al.,2007)
Fragaria
virginiana
Rosaceae Leaves Water extract Yeast isolates (Ducan et
al., 2008)
Acacia nilotica Fabaceae Bark,
Leaves
Methanol extract A. flavus (Mahesh
and Satish,
2008)
Ziziphus
mauritiana
Rhamnaceae Bark,
Leaves
Methanol extract Dreschlera
turcica
(Mahesh
and Satish,
2008)
Sida cordifolia Malvaceae Bark,
Leaves
Methanol extract Fusarium
verticillioides
(Mahesh
and Satish,
2008)

Thymus
vulgaris
Lamiaceae Aerial
part
Essential oil A. niger, A. flavus,
A. fumigatus
(Soković et
al., 2009)
Mentha spicata Labiatae Aerial
part
Essential oil A. niger, A. flavus,
A. fumigatus
(Soković et
al., 2009)
Piper
regnellii
Piperaceae Leaves Hexane extract Paracoccidioides
brasiliensis
(Johann et
al., 2010)
Baccharis
dracunculifolia
Asteraceae Leaves Hexane extract Paracoccidioides
brasiliensis
(Johann et
al., 2010)
Grindelia
camporum
Asteraceae Leaves Methanol extract F. oxysporum,
A. flavus,
A.fumigatus
(Zabka et
al., 2011)
Cajanus cajan Fabaceae Leaves,
Root
Ethanolic extract C. albicans,
C. tropicalis,
C. krusei,
(Brito et al.,
2012)
Arctotis
arctotoides
Asteraceae Leaves, Hexane,
Acetone extract
C. glabrata,
C.krusei,
(Otang et
al., 2012)
Gasteria
bicolor
Asphodelaceae Leaves, Hexane,
Acetone extract
C. glabrata,
C.krusei,
(Otang et
al., 2012)
Psoralea
corylifolia
Fabaceae Seeds petroleum ether
extract
F.oxysporum,
F. moniliforme,
F.graminearum,
(Srinivasan
and Sarada,
2012)

3.8. Selection of Plants Species for Screening
To available estimates, the total number of higher plants species is approximately
250,000 species. Of them, only 6% have been reportedly screened for biological activity and
about 15% have been screened for phytochemical activity (Fabricant and Farnsworth, 2001).
Initial listing of the aspirants’ species for screening of biological activity is a major task of
specific importance in itself. The selection of suitable plants could be done, including
traditional use, chemical content, toxicity, randomized selection or a combination of several
criteria. Here, the reasonable plants selection will be on the bases that have been exploited for
human use as traditional medicines in some place. The method involves a careful observation
of the use of natural resources in folk medicine in different cultures.
As mentioned above, Achyranthes aspera, Aegle marmelos, Argemona mexicana,
Callistemon lanceolatus, Capparis aphylla, Catharanthus roseus, Commelina bengalensis,
Justicia adhatoda and Syzygium cumini have been selected for present study on the basis of
traditional medicine systems, which have been used for thousands of years in India.
Moreover, the use of these plants in the traditional medicine systems of many other cultures
has been extensively documented. Therefore, a search for antifungal drugs has been
examined where the investigator determined the well-defined pharmacological activity and
performs a randomized exploration.

















