Alginate degradation in situ accompanying thermal drying of the marine algae Nereocystis luetkeana and integrifolia by J. N. C. Whyte and J. R. Englar andM . Kung LIBRI\R T ,. .. ..-;:" D Efl (I .": l' . . . I. . FISHERIES AND MARINE SERVICE SERVICE DES PECHES ET DES SCIENCES DE LA MER TECHNICAL REPORT No. RAPPORT TECHNIQUE N° 1976 643 1+ Environment Canada Fisheries and Marine Service Environ nement Canada Service des peches et des sciences de la mer
Transcript
Alginate degradation in situ accompanying thermal drying of the marine algae Nereocystis luetkeana and M~crocystis integrifolia
by
J . N. C. Whyte and J. R. Englar
andM. Kung
LIBRI\R T ,. .. ..-;:"
DEfl (I .": l' . . . I. .
FISHERIES AND MARINE SERVICE
SERVICE DES PECHES ET DES SCIENCES DE LA MER
TECHNICAL REPORT No. RAPPORT TECHNIQUE N°
1976
643
1+ Environment Canada
Fisheries and Marine Service
Environ nement Canada
Service des peches et des sciences de la mer
Technical Reports
Technical Reports are research documents that are of sufficient importance to be preserved, but which for some reason are not appropriate for primary scientific publication. Inquiries concerning any particular Report should be directed to the issuing establishment.
Rapports Techniques
Les rapports techniques sont des documents de recherche qui revetent une assez grande importance pour etre conserves mais qui, pour une raison ou pour une autre, ne conviennent pas a une publication scientifique prioritaire. Pour toute demande de renseignements concernant un rapport particulier, il faut s'adresser au service responsable.
Department of the Environment
Fisheries and Marine Service
Research and Development Directorate
TECHNICAL REPORT NO. 643
(Numbers 1-456 in this series were issued
as Technical Reports of the Fisheries
Research Board of Canada. The series
name was changed with report number 457)
Minist~re de l'Environnement
Services des Peches et des Sciences de
la mer Direction de la Recherche et
D~veloppement
RAPPORT TECHNIQUE NO. 643
(Les num~ros 1-456 dans cette serie
furent utilises comme Rapports
Techniques de l'office des recherches
sur les pecheries du Canada. Le nom
de la serie fut change avec Ie
rapport numero 457)
Alginate degradation in situ accompanying
thermal drying of the marine algae
Nereocystis luetkeana and Macrocystis integrifolia
by
J.N.C. Whyte, J.R. Englar
and
This is the thirty~third Technical
Report from the Research and
Development Directorate
Vancouver Laboratory
Vancouver, B.C.
M. Kung
Ceci est Ie trente troisieme Rapport
de la Direction de la Recherche et
Developpement, Laboratoire de
Vancouver
Vancouver, (C.-B.)
1976
i i
TABLE OF CONTENTS
Page No.
ABSTRACT
INTRODUCTION
EXPERIMENTAL:
iv
1
5
(a) Collection and preparation of seaweed samples 5
(b) Procedures and methods of analysis 5
(i) Drying of algal material on aluminum weighing dishes
(ii) Drying of algal material on a fine-meshscreen
5
5
(iii) Extraction of sodium alginate 6
(iv) Effect of heat on pH of the minced algae 7
(v) Dry weight of algal material 7
(vi) Sodium alginate determination 7
(vi i) Viscosity measurements 7
RESULTS AND DISCUSSION 8
ACKNOWLEDGMENT
REFERENCES
Scheme 1.
Table l.
Table 2.
Table 3.
Table 4.
Table 5.
Fundamental steps in making sodium alginate
Drying of Nereocystis luetkeana in aluminum . wei ghi ng di shes
Drying of Macrocystis integrifolia in aluminum weighing dishes
Intrinsic sodium alginate in freeze-dried kelp
16
17
10
20
21
samples 22
pH of algal tissue maintained at different temperatures for 1 hour 23
Correlation between drying and extracted sodium alginate from Nereocystis 1uetkeana 24
iii
Page No.
Table 6. Correlation between drying and extracted sodium alginate from Macrocystis integrifolia 25
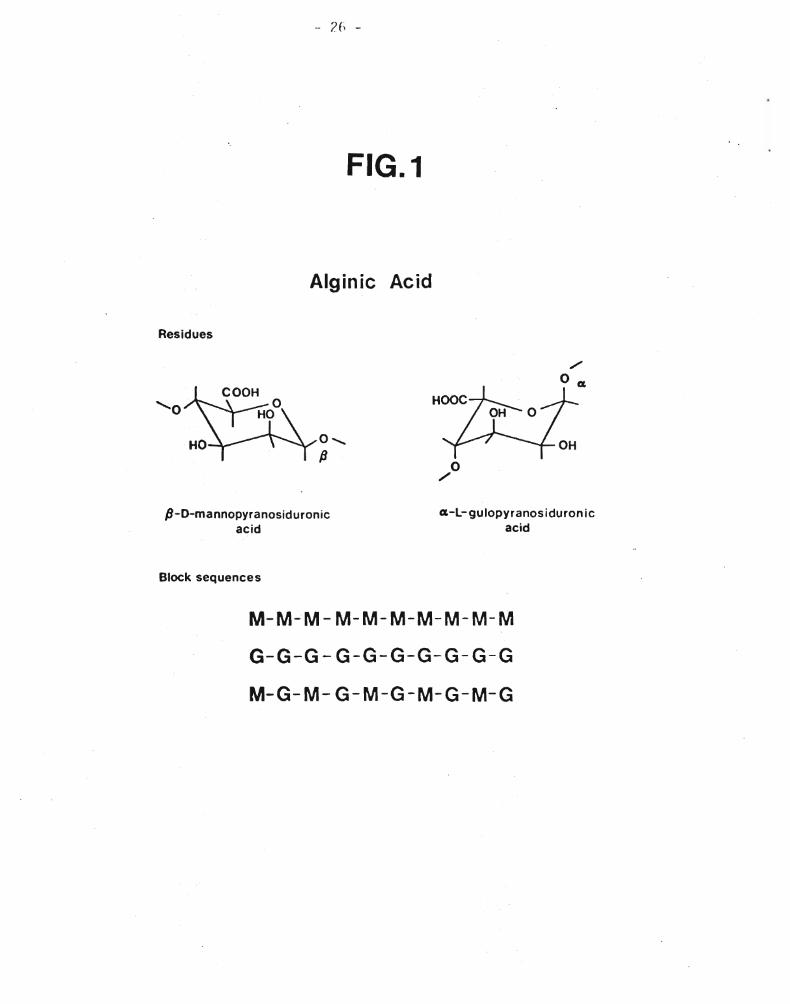
Figure l. Alginic acid 26
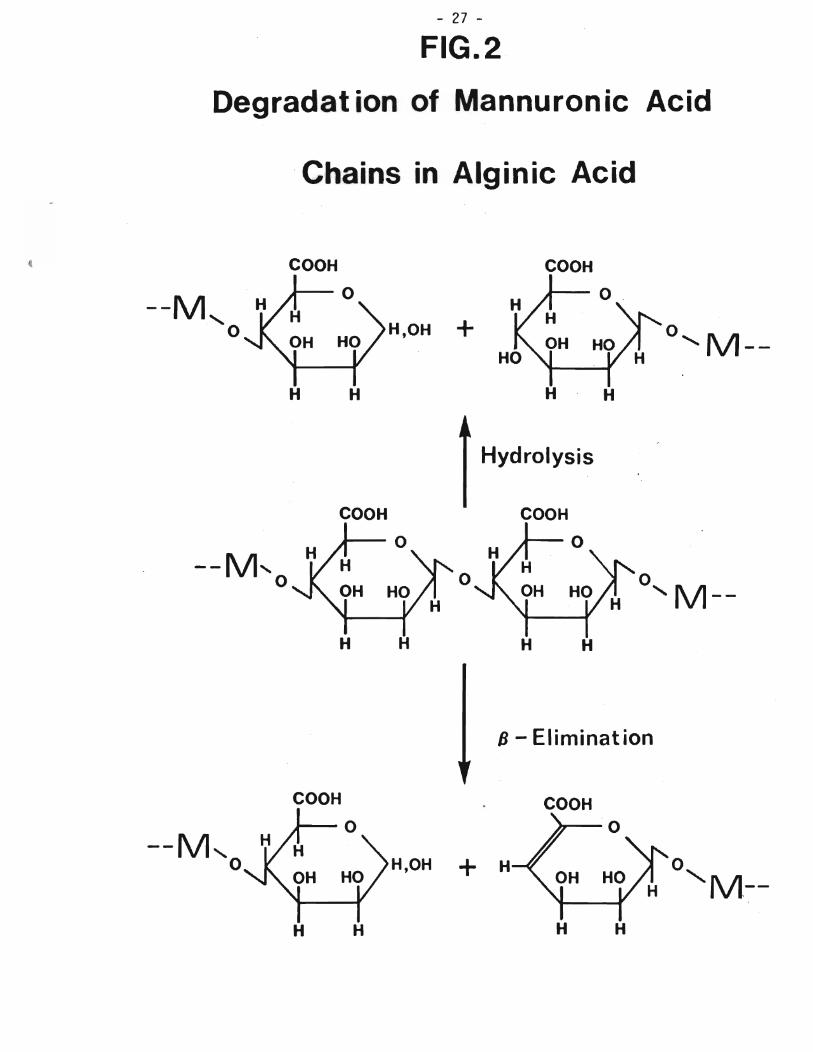
Figure 2. Degradation of mannuronic acid chains in alginic acid 27
Figure 3. Effect of temperature on the drying rate of Nereocystis 28
Fi gure 4. Effect of temperature on the drying rate of Macrocystis 29
Figure 5. Effect on viscosity of alginate with drying temperature of Nereocystis 30
Fi gure 6. Effect on viscosity of alginate with drying temperature of Macrocystis 31
,
iv
ABSTRACT
In the general introduction to this report previous work associated with thermal drying of marine algae is presented together with a comparison of the moisture contents of Nereocystis luetkeana, Macrocystis integrifolia and other commonly utilized brown algae.
The drying of static-beds of minced Nereocystis and Macrocystis on aluminum dishes and fine-mesh screens indicated that the former alga
generally required about 1.3 times the drying period of the latter with
the resistance to dehydration below the 30% moisture level in Nereocystis accounting for the additional drying time.
The extraction processes for sodium alginate and the types of chemical degradation associated with this polyuronide are presented and discussed. The degradation of the alginate accompanying drying of Nereocystis and
Macrocystis was detected by viscosity measurements of the isolated polysaccharide. On drying Nereocystis severe depolymerization of the component alginate resulted
from drying conditions in excess of 80oC, whereas the polymer in Macrocystis
tended to resist thermal degradation to the extent that a low to medium grade of alginate could be recovered from the plant tissue dried even at 300oC.
0 '
Although these laboratory experiments cannot be fully equated with a commercial scale thermal drying unit the results would suggest that Macrocystis integrifolia should be dried in preference to Nereocystis luetkeana when consideration is afforded the economics of drying and the subsequent use of the dried product as a source of sodium alginate.
v
, ,/
RESUME
Dansl'introduction generale du present rapport on donne Ie compte
rendu de travaux anterieurs qui avaient porte sur la deshydratation a la
chaleur des algues marines, en meme temps que la teneur en humidite de
Nereocystis luetkeana, de Macrocystis integrifolia et d'autres algues brunes
communement utilisees.
Le sechage de couches statiques de Nereocystis et de Macrocystis
emincees sur des recipients plats d'aluminium ou des tamis fins a permis
de constater que la duree de la deshydratation etait habituellement 1, 3
fois plus longue pour la premiere et que ceci etait attribuable au fait
qu'a partir de 30 pour cent d'humidite l'operation devenait plus difficile.
On traite des pro cedes d'extraction de l'alginate de sodium ainsi
que des types de degradation chimique associes a ce polyuronide. La
degradation de l'alginate qui accompagne la deshydratation de Nereocystis
et de Macrocystis a ete mise en evidence par viscosimetrie du polysaccharide
isole. Pendant la deshydratation de Nereocystis, on a observe une importante
depolymerisation de l'alginate, au-dessus de 800 e, tandis que Ie polymere
chez Macrocystis, semble resister a la temperature au point qu'il serait
possible de recuperer de l'alginate de basse ou de moyenne qualite, meme
apres deshydratation a 300oe.
Les conditions d'experience ne sont pas tout a fait comparables a
celles qu'on retrouve dans les installations industrielles, mais, d'apres
les resultats, il semblerait preferable de traiter Macerocystis integrifolia
plut6t que Nereocystis luetkeana, compte tenu des facteurs economiques de
l'operation et de l'emploi subsequent du produit seche comme source d'alginate
de sodium.
- 1 -
I NTROOUCTI ON
The principal reasons for drying brown seaweeds, which contain from
70% to 92% moisture in living tissue, are to allow the material to be stored
without subsequent fungal or bacterial decomposition, to reduce the cost of
transportation, to provide material suitable for compounding into animal feed or fertilizer products and to provide a ready source of raw material
for extraction of the commercially important polysaccharide, alginic acid.
In Britain during the 1950 l s most seaweeds were dried, often after
air drying, in through-circulation dryers and considerable efforts were made
by the Institute of Seaweed Research to develop cheap and efficient procedures for drying seaweed (Anon., 1968). Through-circulation drying, which consisted
essentially of blowing he~ted air through static chopped seaweed beds was extensively studied and the drying of Laminaria c1oustoni stipes (Gardner et a1., 1953 (a)), L. c1oustoni fronds (Gardner et a1. 1953 (b)), L. digitata fronds and stipes and L. saccharina fronds (Gardner et a1. 1953 (c)) illustrated
that the drying characteristics of the stipes and fronds of the different species
were similar but that the physical differences between these two segments of
the plants exerted a greater influence on the drying time than any differences
between these sublittoral species. In general however, it was noted that
L. digitata plants dried faster than, and L. saccharina slower than L. c1oustoni .
From these laboratory tests a graphical design for continuous multi-stage
through-circulation drying of seaweed was developed (Gardner et a1. 1954). Further work on this type of drying on members of the Fucaceae, Ascophy11um nodosum, Fucus serratus and F. vesicu10sus (Mitchell et a1., 1955) demonstrated that the mucilaginous nature of the chopped seaweed was primarily responsible
for the problems and inefficiency associated with through-circulation drying, since the glutinous mass often formed from chopped algae presented an impermeable layer to the flow of air in the dryer.
Drying of plant tissue is governed by the rate of internal migration of the moisture to the surface of the tissue. The rate of drying is therefore
dependent on the particle surface size together with the surface temperature
- 2 -
of the particle. The larger the particle the longer the moisture will
take to migrate to the surface and hence the longer the drying time.
The inefficiency of the through-circulation dryers resulted from the relatively large particle sizes used in the process. However, even
when the particle size was reduced the matted product produced by many
algae provided an equally inefficient system as a result of the impervious nature of the chopped seaweed bed to the flow of drying air. To overcome these problems use was made of revolving drum dryers which by agitation of the seaweed bed continually provided new surfaces of the chopped tissue to the drying forces, which were generated by pressurized steam or oil-fired
burners. The drying of several members of the Laminariaceae and Fucaceae
from the coast of Britain with a steam-jacketed drum dryer illustrated that
seaweeds could be dried successfully with the drum operating at 1620 C,
conditions under which it is considered that the alga would be in contact
with the drum for about 15 seconds and the dry seaweed would leave the drum
at 400 C. Nevertheless, even under these relatively mild operating conditions the alginate component of these algae was demonstrated to have undergone
partial depo1ymerization (Booth, 1956).
In the late 1960's in British Columbia a kelp drying-plant was constructed at Masset, Queen Charlotte Islands, to dry the giant kelp Macrocystis integrifo1ia
and Nereocystis 1uetkeana with a rotary, diesel-oil fired, concurrent flow triple-pass kiln type dryer which had a capability of evaporating 18,000 1bs. of water per hour. In a trial operation of the facility in 1973 (Vlhyte et a1., 1974(a)) the efficiency of the drying system was readily exhibited by the drying of 1,200 tons of chopped Nereocystis. Although the pre-drying transportation facilities proved to be totally inefficient the final dried.
ke1pmea1 was obtained in a condition suitable for formulation into animal feed and fertilizer products. However subsequent chemical analysis of the
ke1pmea1 demonstrated that the alginate component of Nereocystis had undergone
severe depo1ymerization during the drying process and was therefore of limited
commercial utility. To have decreased the air temperature within the dryer in an effort to minimize the thermal degradation of the alginate would probably
- 3 -
have made impractical the drying of Nereocystis since, unlike most other
brown algae, it contains in excess of 90% moisture (Whyte et a1. 1974(b)).
Seaweeds must be dried to less than 20% moisture content to be suitable for storage without fungal or bacterial degradation. Therefore the economics of drying brown algae must be based to a large extent on the percentage of moisture existing in the living tissue. As a comparative guide to the drying requirements of certain kelps the following list of moisture contents of
The giant floating kelps on the coast of British Columbia, Nereocystis luetkeana and Macrocystis integrifo1ia, are noted to contain the higher levels of moisture in the fresh state and would certainly be more costly to dry than Ascophyllum nodosum which is the principal component of Norwegian kelpmeal (Jensen, 1971).
The Canada Kelp Co., which first owned the drying plant at Masset, originally intended a future expansion of the facilities to include an alginate extraction plant with the intention of using the dried seaweed
meal as a source of sodium alginate in the off-season. We expressed scepticism
- 4 -
about these plans because the proposal would depend on the nature of the alginate component remaining in the Nereocystis and Macrocystis following the drying procedure. In fact, the trial operation of the system at Masset did demonstrate that the alginate component, (with a viscosity of 5000-7000 centipoise in fresh Nereocystis) was subsequently degraded on drying in the Heil dryer to afford a low molecular weight polymer in the kelpmeal which
exhibited a viscosity of 19-30 centipoise and was of little commercial value.
This present study was therefore instigated to determine the comparative drying times of the two giant kelps under identical laboratory conditions and to determine the subsequent effects of thermal drying on the alginate component in situ in Nereocystis luetkeana and Macrocystis integrifol ia.
,
- 5 -
EXPERIMENTAL
(a) Collection and Preparation of Seaweed Samples
Samples of Nereocystis luetkeana and Macrocystis integrifolia
were collected at Sooke, Vancouver Island in July 1974. Only healthy
attached plants were selected and cut approximately 5ft. from the apices of the stipes. The specimens were placed in plastic bags and transported
to the laboratory in insulated coolers containing ice layered over crushed dry ice placed on the bottom of the coolers. The plants were then freed from extraneous epiphytes and epifauna and minced through a Blakeslee butcher's mincer. The combined minced tissue was stirred to attain a
homogeneous consistency then packaged into zip-lock plastic bags, weighed
and stored at -3l oC.
(b) Procedures and methods of analysis
(i) Drying of algal material on aluminum weighing dishes
Samples of the thawed minced algal tissue (lOg) were spread evenly
on the bottom of aluminum weighing dishes (54mm diameter) and positioned in a forced draft airflow oven regulated at a specific temperature. The
uniform performance characteristics of the 3.75 cu.ft. oven, with a control
sensitivity of <± 0.250 C, a uniformity at 200°C of <± 1.50 C, a set point
accuracy of ± 2.00C and exceptionally rapid recovery time, allowed for
reproducible assessments of drying rates to be executed. The weight of the dishes with contents was recorded at selected time intervals and continued until constant weight was achieved. Triplicate samples were dried and the median values recorded.
(ii) Drying of algal material on a fine-mesh screen
A grill-frame (42.6 x 30 cm) was made from 4 mm diameter stainless steel rod with transverse support rods positioned every 2.5 cm along the
longitudinal sides of the frame. Two longitudinal skids made from the same rod elevated the grill - frame by 6 cm. The surface of the grill was covered
with fine-mesh screen (fly-screen) and between the two skids aluminum foil was positioned to form a catch-pan. The whole apparatus was weighed on a top-loading
- 6 -
balance. The frozen algal material was equilibrated to lOC in a cold
room then a sample (250 g) spread evenly over the surface of the screen.
The fluid which drained from the screen was caught in the fabricated
aluminum foil pan and the whole apparatus was again weighed. The weight
requirement to obtain a 10% moisture level in the final dried product
was calculated as follows:
Weight of algal material dried to X% moisture level
= weight wet alga x percentage absolute dry weight
100 - X
The apparatus with sample was inserted into the oven at a selected
temperature and as the drying process proceeded the grill-frame was removed and weighed. The drying was continued until the requisite amount of water
had been lost from the sample and the remaining dried material was removed
for subsequent extraction.
(iii) Extraction of Sodium Alginate (Whyte et al., 1975)
The dried algal material retrieved from the screen (20-35 g) was
extracted with 500 ml of 1% aqueous calcium chloride at 600 C for 20 minutes
and the resultant mixture centrifuged. The centrifugate was discarded and the residue further extracted twice with calcium chloride to remove all "fucoidan" material. The residue was then treated with three successive washings of 0.05 N aqueous hydrochloric acid at room temperature to remove
as much of the calcium component as possible prior to being extracted with 500 ml of 0.5% aqueous sodium carbonate at 450 C for 30 minutes. The resultant
mixture was centrifuged and the residue accorded two further treatments with
sodium carbonate solution. To the combined extracts, 6 volumes of ethanol
were added to precipitate the sodium alginate polymer. The polysaccharide
was then washed with 75% aqueous ethanol until free from alkali then dried
by solvent exchange, using acetone and diethyl ether, and finally dried under
high vacuum and weighed.
- 7-
(iv) Effect of heat on pH of the minced algae
Samples of the thawed minced algal tissue (approx. 200 g) were heated at various temperatures for 1 hour, cooled to room temperature and the pH of the resultant mixture, after stirring, was measured by an
Orion Research Iona1yzer Model 801 digital pH/mv meter coupled to an
Orion laboratory pH electrode Model 91-01-00 and reference electrode.
(v) Dry weight of algal material
The absolute dry weight of the minced algae was determined by
freeze drying samples to constant weight.
(vi) Sodium alginate determinations
The exact content of sodium alginate in the minced algal samples
was assessed by an acidic decarboxylation technique described previously
(Whyte et a1. 1974 (c)).
(vii) Viscosity measurements
Aqueous solutions of the isolated sodium alginate, in 1% concentrations,
were measured at 2SoC with a Brookfield viscometer using a #2 or #3 spindle operating at 60 rpm.
- 8 -
RESULTS AND DISCUSSION
Samples of Nereocystis luetkeana and Macrocystis integrifolia,
collected at Sooke in the month of July, vlere minced and thoroughly
stirred to provide a homogeneous mixture. The minced Nereocystis
was easy to handle and consisted of loosely held particles of vegetative
tissue admixed with cell fluid which could be readily separated into con
stituent parts. By comparison the minced Macrocystis produced a glutinous
mass, lacking expressed cell fluid, which entrapped considerable air and
defied separation into constituent parts.
To assess the drying times of these algal samples portions were
placed in aluminum weighing dishes and dried at selected temperatures
in an isothermal forced draft airflow oven. Graphs were drawn of time
versus percentage moisture remaining in the algal tissue and from these
graphs the duration of drying at a specific temperature to attain moisture
levels of 30, 20, 15, 10 and 0% were obtained and are presented in Tables
and 2 for Nereocystis and Macrocystis respectively. It was noted that
Macrocystis required a longer period of drying than Nereocystis when
heated at l050 C and 1500 C yet this situation was reversed when the samples
were heated above 2000 C. This might be explained by the entrapment of
moisture under a skin formed on drying the mucilaginous Macrocystis sample
at lower temperatures and the prevention or disruption of this surface layer
when rapid egression of moisture was encountered at the more elevated temper
atures. Apart from this anomaly, which would occur only when drying in a
static bed like a dish, Nereocystis would tend to take longer to dry than Macrocystis in accordance with the fact that the former alga contained 91.6%
moisture in comparison to 88.3% moisture in the latter. The samples when
heated above 1500 C invariably experienced peripheral charring as the moisture
content tended towards 0% which accounted for the lower solids content
reported for the algae when dried at the higher temperatures, Tables 1 and
2.
Of considerable importance in this study was the standardizing of the
t
- 9 -
sodium alginate extraction procedure on the dried algal matr.rial since
degradation of the polymer was possible during subsequent isolation with
chemical extractants. The fundamental steps for the chemical extraction
of sodium alginate, whether in the laboratory or industrial scale, involves variations of the general extraction procedure outlined in Scheme 1. The
Green's cold process which is used for the extraction of Macrocystis pyrifera
involves leaching the fresh kelp for several hours with 0.3% hydrochloric acid to remove salts and fucoidan prior to alkaline extraction of the alginate.
The corresponding G1oahec-Herter process, which is associated more with
the littoral and sublittoral species, involves leaching of the dried or
partially dried algae with dilute calcium chloride followed by dilute hydrochloric acid to remove salts, 1aminaran and fucoidan prior to digestion with
alkali at 600 C to extract the alginate component. Other alginate products are produced by neutralizing the alginic acid, Scheme 1, with appropriate bases to afford other salts or by reacting the acid with propylene oxide
to provide propylene glycol alginate. Although this information on the
commercial extraction of alginate is available in the literature (Guise1ey,
1968) details of current extraction processes are closely guarded and have
not been made known.
Following an assessment of a number of slightly modified alginate extraction procedures the most reproducible, as regards yield and viscosity
of isolated polysaccharide, consisted of treating the algal material with 1% aqueous calcium chloride at 60°C followed by washing with dilute hydro
chloric acid at room temperature to remove all salts and fucoidan material
then extracting the alginate with 0.5% aqueous sodium carbonate at 450 C for 30 minutes. The polysaccharide was precipitated with ethanol and washed free from all alkali with 75% aqueous ethanol and dried by solvent exchange using acetone and diethyl ether. From other preliminary extractions it was demonstrated that the polysaccharide was mor~ susceptible to degradation
when isolated with 1% aqueous sodium carbonate at 600 C. Similarly it was
demonstrated that an acid wash following the calcium chloride pretreatment
was necessary prior to extraction with 0.5% sodium carbonate since the
- 10-
SCHEME 1
Fundamental steps in making Sodium Alginate
I WET OR DRY SEAWEED
1% CaC1 2 or 0.3% HCl ---........ 1 LEACHING :I------+r
Soda ash and water ---+.1 DISSOLUTION OF ALGINATE I--L_--! ...
Dehydration (hot air _ 1 SODIUM ALGINATE PASTE .r---. or alcohol)
I MILLING AND SCREENING
I DRY SODIUM ALGINATE PQ\lJDER J
Washings
Insoluble residue
Waste and cellulose
Waste liquor
CaC1 2, water and
dissolved impurities
Waste alcohol
CO 2 and waste liquor
Water and waste alcohol
- 11 -
carry-over of trace quantities of calcium ions into the alginate fraction subsequently produced partial gel formations in the aqueous solutions resulting in extremely elevated, yet fallacious, viscosity measurements.
The sodium alginate content, based on weight of material isolated
by the extraction procedure, was 22.4% and 23.0% for Nereocystis and
Macrocystis respectively, Table 3. These values were 2% to 3% higher than the figures obtained by acidic decarboxylation estimations (Whyte et a1., 1974(c) which however were more accurate since they were less
prone to the vagaries of isolation. Nevertheless, for the purposes of this experiment a comparison of the content of isolated sodium alginate was more significant than the absolute values. Viscosity readings of 5625
centipoise and 5900 centipoise for 1% aqueous solutions of sodium alginate
from Nereocystis and Macrocystis respectively indicated a high degree of
polymerization for the polysaccharides in the fresh algae, Table 3. Yet, as a po1yuronide, alginic acid and corresponding salts are susceptible to
ready degradation in acidic, alkaline and even neutral solutions.
Alginic acid is a linear copolymer composed of block segments of
B-~-(1+4)-linked mannopyranosiduronic acid residues, a-~-(1+4)-linked gu10pyranosiduronic acid residues and alternating sequences of these
residues as illustrated in Figure 1 (Haug et a1., 1967 (a)). Degradation
of these block polymers can involve proton-catalyzed hydrolytic cleavage of the glycuronan chain leading to saturated oligosaccharide fragments (Smidsrod et al., 1969) or can entail B-elimination at the 4-Q-glycosidic linkage to afford unsaturated deoxyhexopyranuronate-oligosaccharide fragments (Haug et a1., 1967(b)) such as those depicted as the possible degradation products of the mannuronic acid chains in alginic acid, Figure 2 (Kiss, 1974). It has been demonstrated for alginic acid that the rate of S-eliminative
degradation increases above pH 10 and that hy~rolytic cleavage is more prominent below pH 5, yet between these pH values in slightly alkaline. acidic or neutral medium a significant portion of the degradation occurs as a B-eliminative process (Haug et al., 1963). Degradation of alginic acid
- 12 -
can also result from oxidative depolymerization caused by autoxidation
of reducing compounds in the algae. The reducing phenolic compound
IIfucosan ll present in varying amounts in brown algae, particularly members
of the Fucaceae, is considered, by autoxidation, t o liberate hydrogen
peroxide which subsequently forms free radicals that degrade the alginic
acid molecule by chain rupture. (Smidsrod et al., 1963).
The pH values of 6.29 and 6.19 for minced Nereocystis and Macrocystis
dec l ined only slightly when the tissue was heated at various temperatures
up to 1000e for 1 hour, Table 4. The relatively neutral condition of the
algal tissue suggested in accordance with the theories presented in the
previous paragraph that during the process of drying the alginate molecule
would be depolymerized by a combination of Belimination and oxidative
degradation. Thus when a 1% aqueous solution of a commercial sample of
sodium alginate, at pH 7.2, with a viscosity of 236 centipoise was heated
at 1000e for 1 hour the viscosity of the resultant solution, at pH 7.5,
was recorded as 74 centipoise.
To determine the effects on the alginate ~ situ in the algae at
various drying temperatures sufficient quantities of algal tissue had to
be dried to yield adequate material for subsequent extraction. Although
the drying of 10 gram quantities in aluminum dishes, alluded to previously, provided a general guide to the time requirements for drying at specific
temperatures, the non-uniformity associated with drying in these types of
containers, especially with larger quantities of vegetative tissue, prompted
the use of an elevated screen-covered grill frame as the drying vehicle.
The minced alga was spread evenly and thinly on the screen such that the
circulating air
uniform drying.
level the algal
in the oven dried the layer on either side to provide more
As commercial kelpmeal is dried to a 10-20% moisture
samples were dried to approximately 10% moisture levels
and the drying curves obtained for Nereocystis and Macrocystis are
illustrated in Figure 3 and 4 respectively. With drying temperatures
up to 2000 e, the times to attain approximately 10% moisture contents in
- 13 -
Nereocystis were about 1.3 times as long as that for Macrocystis. From the shape of the drying curves the resistance to dehydration below the 30%
moisture level in Nereocystis would appear to be responsible for the additional
drying time. Drying on the screen rather than in dishes considerably curtailed
the drying times of both algal tissues especially when the lower drying temper
ature of 105°C was employed, cf. Tables 1, 2~ 5 and 6. As it was not plausible
to dry fresh material to an exact moisture level the actual content of moisture remaining in the dried Nereocystis and Macrocystis samples are presented in
Tables 5 and 6 respectively.
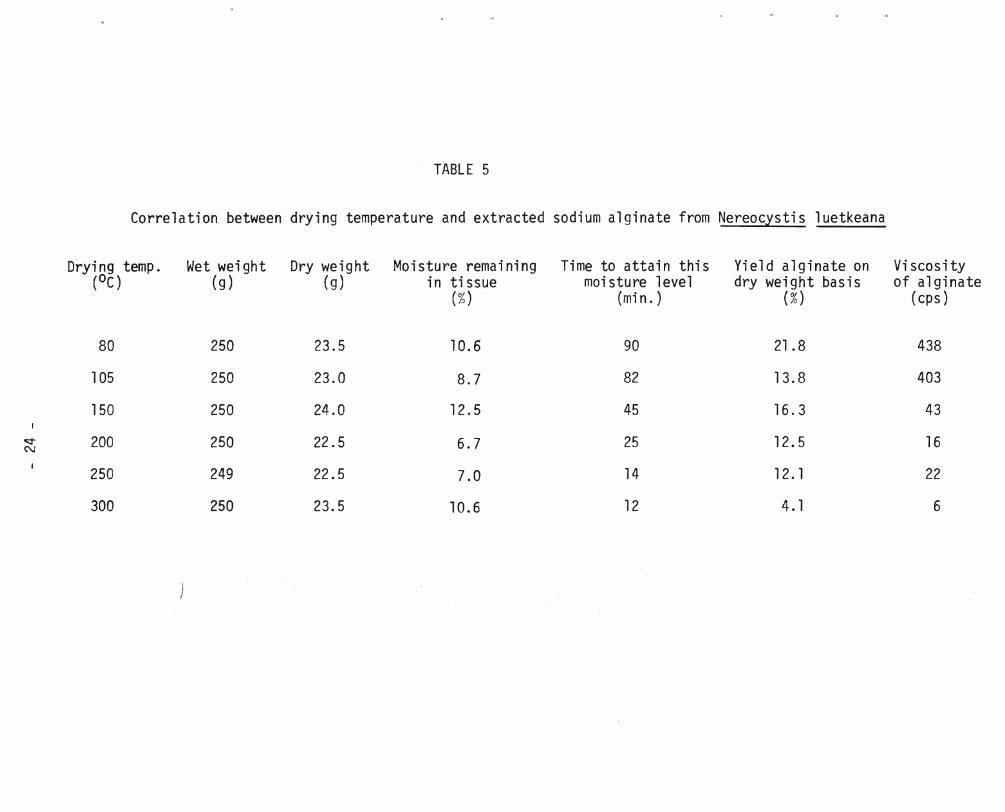
Extraction of the dried samples of Nereocystis provided sodium
alginate in yields which ranged from 21.B% to 4.1% corresponding to drying
temperatures of BOoe and 300°C respectively, and indicated severe degradation
at the latter temperature. When dried at 105°C to 250°C the yields of
alginate were similar at approximately 14±2% although a marked difference in the viscosities of these isolates was observed, Table 5. The correlation
between the viscosities of the isolated alginates and drying temperatures of
Nereocystis clearly illustrated the accelerated degradation of the polysaccharide
in the alga when dried above 105°C, Figure 5.
The extent of degradation in the alginate in Nereocystis accompanying
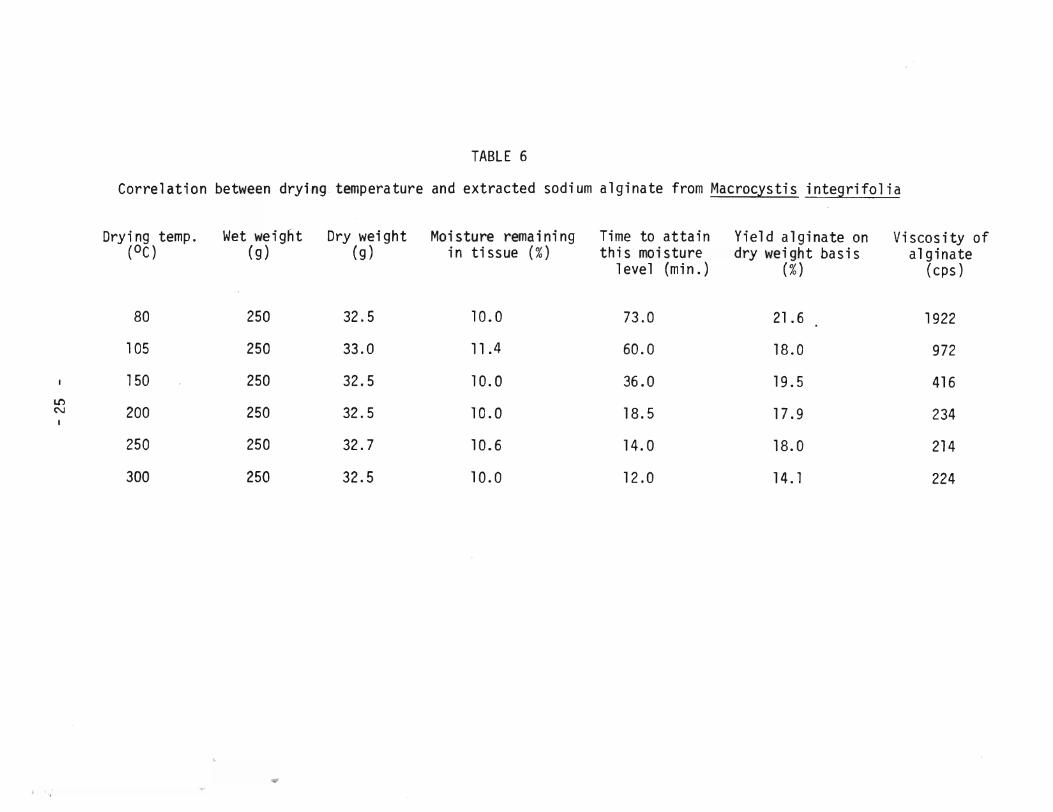
thermal drying was not as evident in the corresponding polymer isolated from the dried Macrocystis. The yield of alginate isolated from this alga
ranged from 21.6% to 14.1% when drying was performed at BOoe to 300°C respectively and at the intermediate temperatures of 105°C to 250°C the yield varied only slightly, lB.7±0.B%, Table 6. Relatively high viscosities of approximately 1000 - 2000 centipoise were registered for the alginate isolated from Macrocystis
dried below 105°C, hO\</ever when this temperature was exceeded a noticeable decline in the degree of polymerization of the polysaccharide ensued, Figure 6. Interestingly enough the alginate from Macrocystis appeared to attain a lower
limit of depolymerization, with viscosities recorded in excess of 200 centipoise even for the algal samples dried at the extreme temperatures of 200°C
to 300°C. This is in marked contrast to the alginate from Nereocystis which
_ 14 _
displayed viscosities under 43 centipoise when the associat~d drying temperature exceeded 1050 C.
Commercial sodium alginates provide 1% aqueous solutions with viscosities varying from 100 to 2000 centipoise for medium to high viscosity grades and 20 to 100 centipoise for extra low to low viscosity grades (Anon., 1961). The initial thermal degradation of the alginic acid
in Nereocystis and the further depolymerization accompanying subsequent
industrial isolation would render this dried alga unsuitable for the
production of medium to high grade algi nates. Nevertheless, it would appear from our results that dried Macrocystis could be extracted for
low to medium grade alginate depending on the temperature of drying, with the lowest temperatures according the highest viscosity alginate in the final dried alga.
The removal of water from these seaweeds by mechanical dewatering or
natural air drying would negate the adverse effects on the nature of the
alginate component which accompany thermal drying. Preliminary results from roller and screw pressing of chopped Nereocystis and Macrocystis have demonstrated that cellular fluid resisted expression by these devices due to the flexible nature of the plant cellular matrix (Whyte et al., 1974 (a)). A minor percentage of cellular fluid exuding from chopped Nereocystis could be separated by centrifugation, however, the glutinous mass resulting from chopped Macrocystis would prevent any such separation. Intact Macrocystis plants have been dried successfully by natural air drying, in contrast to whole Nereocystis plants which tended to decompose before drying as a result of the inseparable matting of the fronds hindering the flow of air to all
surfaces of the plant (Whyte, unpublished observations). The bulk storage of fresh Nereocystis and Macrocystis by anaerobic ensiling (Black, 1955) or the use of "curing tanks" in which chopped algae could be preserved by
formalin, sulphur dioxide or other preservatives have not been investigated.
The observations presented in this report would indicate that the
- 15 -
industrial isolation of alginates from the giant kelps Nereocystis and Macrocystis on the coast of B.C . should preferably be performed on fresh raw material. However. should the storage of dried kelp be essential for
. the economic operation of an alginate extraction plant then Macrocystis integrifolia should be dried in preference to Nereocystis luetkeana since the alginate component of the former alga tends to resist thermal degradation to the extent that a low to medium grade of alginate could be isolated ultimately from the resultant kelpmeal.
- 16 -
ACKNOWLEDGMENT
As a recipient of a summer grant provided by the Work-in-Government
1975 Program of the Government of British Columbia, Miss M. Kung was seconded from the Marine Resources (Commercial Fisheries) Branch to the
Vancouver Laboratory of the Fisheries and Marine Service to assist the research being conducted in the marine plants project. The authors wish to express their appreciation to the Marine Resources Branch and in particular to Mr. L.M. Coon, Head, Marine Plant Section, for providing the funds and arranging the transfer which proved to be mutually beneficial.
- 17 -
REFERENCES
Anon., 1961. Ke1co algin, the story of its control over the water in products we use in our daily lives. Ke1co Co., Calif., 13pp.
Anon., 1968. Institute of Seaweed Research, Inveresk, Midlothian.
Annual report for 1968, 26 pp.
BLACK, W.A.P., 1955. The preservation of seaweed by ensiling and
bactericides. J. Sci. Food Agric., ~, 14.
BOOTH, E., 1956. A method of drying seaweed using a steam-heated drum dryer. J. Sci. Food Agric., I, 705.
GARDNER, R.G. and MITCHELL, T.J., 1953 (a). Through-circulation drying of seaweed I. Laminaria c1oustoni stipe. J.Sci. Food Agric.,
1, 113.
GARDNER, R.G. and MITCHELL, T.J., 1953 (b). Through-circulation drying of seaweed II. Laminaria c1oustoni frond. J. Sci. Food Agric.,
i, 237.
GARDNER, R.G. and MITCHELL, T.J., 1953 (c). Through-circulation drying of
seaweed III. Laminaria digitata frond and stipe; Laminaria saccharina frond. J. Sci. Food Agric., 1, 364.
GARDNER, R.G. and MITCHELL, T.J., 1954. Through-circulation drying of seaweed IV. A graphical design method for continuous mUlti-stage driers. J. Sci. Food Agric., i, 481.
GUISELEY, K.B.,1968. Seaweed colloids. Encyclopedia of Chemical Technology, ed. R.E. Kirk and V.F. Othmer, Interscience Pub.
Inc., New York, 17, p. 763. ==-==
HAUG, A., LARSEN, B. and SMIDSROD, 0., 1963. The degradation of alginates at different pH values. Acta. Chem. Scand., 17, 1466.
- 18 -
HAUG, A., LARSEN, B. and SMIDSROD, 0., 1967(a). Studies on the sequence of uronic acid residues in alginic acid. Acta, Chern. Scand., n, 691.
HAUG, A., LARSEN, B. and SMIDSROD, 0., 1967(b). Alkaline degradation of alginate. Acta. Chern. Scand., n, 2859.
JENSEN, A., 1971. The nutritional value of seaweed meal for domestic animals. Proc. Intll. Seaweed Symp. I, 7.
KISS, J., 1974. B-Eliminative degradation of carbohydrates containing uronic acid residues. Advances in Carbohydrate Chemistry and Biochemistry, 29, 306.
MITCHELL, T.J. and POTTS, C.S., 1955. Through-circulation drying of seaweed v. Ascophy11um nodosum; Fucus serratus; Fucus vesiculosus. J. Sci. Food Agric., ~, 402.
SMIDSROD, 0., HAUG, A. and LARSEN, B., 1963. Degradation of alginate in the presence of reducing compounds. Acta. Chern. Scand.,
lZ' 2628.
SMIDSROD, 0., LARSEN, B., PAINTER, T. and HAUG, A., 1969. The role of intramolecular autocatalysis in the acid hydrolysis of polysaccharides containing 1,4-linked hexuronic acid. Acta. Chern. Scand., 23, 1573.
WHYTE, J.N.C. and ENGLAR, J.R., 1974(a). Commercial kelp drying operation at Masset, 1973. Fisheries Research Board of Canada, Technical Report No. 453, 30 pp.
WHYTE, J.N.C. and ENGLAR, J.R., 1974(b). Elemental composition of the marine alga Nereocystis luetkeana over the growing season. Fisheries and Marine Service Technical Report No. 509, 29 pp.
- 19 -
WHYTE, J.N,C. and ENGLAR, J.R. 1974(c). Determination of uronic acid content of saccharides by acidic decarboxylation: a simplified procedure. Analytical Biochemistry 59, 426.
WHYTE, J.N.C. and ENGLAR, J.R., 1975. Basic organic chemical parameters of the marine alga Nereocystis luetkeana over the growing season. Fisheries and Marine Service Technical Report No. 589, 42 pp.
Drying
Temperature °c
105
150
200
250
300
- 20 -
TABLE 1
Drying of Nereocystis luetkeana in
Aluminum Weighing Dishes
Time (min.) to Attain Percentage Moisture Dry solids
30% 20% 15% 10% 0% Obtained %
73 92 105 118 240 8.5
32 39 43 55 75 8.3
26 29 32 35 50* 8.1
18 22 24 27 40* 7.9
12 15 17 19 24* 7.3
* Peripheral charring observed at this time
- 21 -
TABLE 2
Drying of Macrocystisintegrifolia in
Aluminum Weighing Dishes
Drying Time (min.) to Attain Percentage Moisture Dry solids
Temperature °c 30% 20% 15% 10% 0% Obtained %
105 88 109 123 138 255 12.3
150 40 47 53 58 90 11. 2
200 16 19 21 23 35* 10.9
250 11 13 15 17 24* 10.8 300 9 10 11 12 16* 10.6
* Peripheral charring observed at this time
- 22 -
TABLE 3
Intrinsic sodium alginate in freeze-dried kelp samples
Figures in parenthesis obtained by acidic decarboxylation
- 23 -
TABLE 4
pH of Algal Tissue Maintained at
Different Temperatures for 1 hour
Temperature Nereocystis Macrocysti s
23°C 6.294 6.192
500C 6.294 6.188
75°C 6.271 6.153
100°C 6.219 6.047
TABLE 5
Correlation between drying temperature and extracted sodium alginate from Nereocystis 1uetkeana
Drying temp. Wet weight Dry weight Moisture remaining Time to attain this Yield alginate on Viscosity (OC) (g) (g) in tissue moi sture level dry weight basis of alginate
(%) (mi n. ) (%) (cps)
80 250 23.5 10.6 90 21.8 438
105 250 23.0 8.7 82 13.8 403
150 250 24.0 12.5 45 16.3 43
o:::t" 200 250 22.5 6.7 25 12.5 16 N
250 249 22.5 7.0 14 12. 1 22
300 250 23.5 10.6 12 4. 1 6
TABLE 6
Correlation between drying temperature and extracted sodium alginate from Macrocysti~ integrifolia
Dry i ng temp. Wet weight Dry weight Moisture remaining Time to attain Yield alginate on Vi seos ity of (OC) (g) (g) in tissue (%) this moisture dry weight basis alginate