Introduction 1 1. INTRODUCTION High salt concentration in the soil or salinity is one of the major abiotic stresses affecting plant growth and productivity (Giri et al. 2003; Al Karaki 2006). Soils are considered saline when the electrical conductivity of the soil is 4 dS m -1 or more (Richards 1954), which is equivalent to approximately 40 mM NaCl and generates an osmotic pressure of approximately 0.2 MPa (Munns and Tester 2008). Salinity arises due to deposition of salts (Fig. 1.1) via two natural processes – weathering of rocks containing soluble salts of various types, mainly chlorides of sodium, calcium and magnesium, and to a lesser extent, sulphates and carbonates; and deposition of oceanic salt (mainly NaCl) carried by inland wind and rain (Munns and Tester 2008). Thus, the major ions contributing to soil salinity include cations (Na + , Ca 2+ , Mg 2+ and K + ) and anions (Cl - , SO 4 2- , HCO 3 - , CO 3 2- and NO 3 - ). Other constituents contributing to salinity in hyper saline soils and water include B, Sr 2+ , SiO 2 , Mo, Ba 2+ and Al 3+ (Hu and Schmidhalter 2002). Besides, irrigation water and fertilizers used in agriculture; low precipitation and over-exploitation of available water resources also contribute significantly to soil salinity (Cantrell and Lindermann 2001; Al Karaki 2006). Of all the salts, sodium chloride is the most soluble and abundant salt released (Munns and Tester 2008). At present, cultivated land affected by salt amounts to 77 million hectares and constitutes 5% of 1.5 billion hectares cultivated land around the world. It is expected that increased salinization will result in 50% of arable land loss by the middle of the 21 st century (Wang et al. 2003). The alarming rate of increase in soil salinity in agricultural land is a setback to agriculture as most of the economically important crop species are very sensitive to soil salinity (Mahajan and Tuteja 2005) and have Fig. 1.1 Salt deposition seen as a white cover on the soil.
Transcript
Introduction
1
1 INTRODUCTION High salt concentration in the soil or salinity is one of the major abiotic stresses affecting plant growth and productivity (Giri et al 2003 Al Karaki 2006) Soils are considered saline when the electrical conductivity of the soil is 4 dS m-1 or more (Richards 1954) which is equivalent to approximately 40 mM NaCl and generates an osmotic pressure of approximately 02 MPa (Munns and Tester 2008) Salinity arises due to deposition of salts (Fig 11) via two natural processes ndash weathering of rocks containing soluble salts of various types mainly chlorides of sodium calcium and magnesium and to a lesser extent sulphates and carbonates and deposition of oceanic salt (mainly NaCl) carried by inland wind and rain (Munns and Tester 2008) Thus the major ions contributing to soil salinity include cations (Na+ Ca2+ Mg2+ and K+) and anions (Cl- SO4
2- HCO3- CO3
2- and NO3-) Other constituents contributing to salinity in hyper
saline soils and water include B Sr2+ SiO2 Mo Ba2+ and Al3+ (Hu and Schmidhalter 2002) Besides irrigation water and fertilizers used in agriculture low precipitation and over-exploitation of available water resources also contribute significantly to soil salinity (Cantrell and Lindermann 2001 Al Karaki 2006) Of all the salts sodium chloride is the most soluble and abundant salt released (Munns and Tester 2008)
At present cultivated land affected by salt amounts to 77 million hectares and constitutes 5 of 15 billion hectares cultivated land around the world It is expected that increased salinization will result in 50 of arable land loss by the middle of the 21st century (Wang et al 2003) The alarming rate of increase in soil salinity in agricultural land is a setback to agriculture as most of the economically important crop species are very sensitive to soil salinity (Mahajan and Tuteja 2005) and have
Fig 11 Salt deposition seen as a white cover on the soil
Introduction
2
resulted in decreased crop production of more than 20 irrigated land worldwide (Porcel et al 2012) 11 Effect of salinity on plants In agriculture the impact of soil salinity on plant is alarming as it affects the establishment growth and development of plants leading to enormous decline in productivity (Tester and Davenport 2003 Giri et al 2003) Excessive salts in soil in particular Na+ ions alter the basic organization of the soil (Mahajan and Tuteja 2005) The occurrence of Na+ ions in the cation exchange complex makes the soil compact and subsequently reduces soil porosity and hampers soil aeration (Manchanda and Garg 2008) Low soil aeration due to high salt concentration has a direct relation with all major living processes such as reduction in growth photosynthesis protein and lipid metabolism (due to salt-induced osmotic imbalance) nutritional disorder and ion toxicity in plants (Porcel et al 2012) (Fig 12) Osmotic inequilibrium in a salt-stressed plant is often translated as retardation in growth of the plants with the leaves and stems appearing stunted (Singh and Charath 2001) This effect of salt is primarily due to ndash (i) decrease in the plantrsquos ability to take up water and nutrients as a result of osmotic or water-deficit (physiological drought) effect of salt and (ii) uptake of salt by plants from the soil through transpiration stream injure cells in the transpiring leaves thereby inhibiting cell division and enlargement in plantrsquos growing point (Manchanda and Garg 2008) Stunted growth of the plants grown in saline soils may also be a consequence of sidetracking of energy to counterbalance the accumulation of salts in the cells Continued uptake of salt by plants and subsequent significant increase in the concentration of salts decreases the size of the leaves (Singh and Charath 2001) affect the structure of chlorophyll molecules resulting in disruption of photosynthesis and leaf senescence Specific effects of salt stress on leaf senescence have been related to accumulation of toxic ions (Na+ and Cl-) or to K+ and Ca2+ depletion (Yeo et al 1991) Salt induced ionic inequilibrium and toxicity is precursor for disruption in the plant mineral relations This may be elucidated by the effects of salinity on nutrient availability competitive uptake transport or partitioning within the plant or may be caused by physiological inactivation of a given nutrient resulting in an increase in the
Introduction
3
plants internal requirement for that essential element thereby causing ionic imbalance in the cell (Grattan and Grieve 1999) At the whole plant level salinity frequently induces an increase in Na+ and Cl- ions as well as a decrease in K+ Ca2+ NO3
- and Pi
concentrations (Shokri and Maadi 2009) Therefore high concentrations of Na+ and Cl- ions in the soil solution may depress nutrient-ion activities and produce extreme ratios of Na+ K+ Na+ Ca2+ Ca2+ Mg2+ and Cl- NO3
- (Grattan and Grieve 1999)
Fig 12 Effect of salt stress on plants Salt stress causes physiological drought to plants resulting in hyperosmotic stress On the other hand excessive uptake of Na+ and Cl- ions lead to hyperionic stress These stresses generate a secondary stress ldquooxidative stressrdquo The combined effect of these stresses ultimately affect plant growth and reduce the yield 12 Strategies to minimize crop loss due to salinity Improving plant tolerance against salinity and maintaining crop productivity in saline soils is a major challenge for sustainable agriculture Huge decline in crop productivity due to salt stress have encouraged the researchers around the world to develop strategies to minimize crop loss Searching for new salt tolerant crop plants and development of salt tolerant crops through breeding are major strategies used so far (Gallagher 1985 Glenn and OrsquoLeary 1985 Cuartero and Fernandez-Munoz 1999)
Introduction
4
Other methods include leaching of excessive salts or desalinizing seawater for use in irrigation (Muralev et al 1997) Though successful these approaches are costly and beyond the economic means of developing nations (Cantrell and Linderman 2001)
Under natural conditions plants can establish symbiosis with microorganisms such as bacteria and fungi These associations have been shown to improve plant performance under stress environments and consequently improve their yield (Brown 1974 Levy et al 1983 Creus et al 1998) Of these microorganisms arbuscular mycorrhizal fungi (AMF) have proved to be an efficient bio-ameliorator of salinity effects in plants The wide host range of AMF ndash it can establish symbiosis with roots of 70 terrestrial plant species including halophytes hydrophytes and xerophytes is an added advantage (Smith and Read 2008 Hejiden et al 1998) AM symbioses have been shown to increase tolerance to soil salinity in a variety of host plants such as Capsicum annum Astragalus sinicus Zea mays (Turkmen et al 2008 Peng et al 2011 Sheng et al 2011) It is clear that AMF ameliorate growth inhibition caused by salinity however the mechanism involved remains unresolved (Porcel et al 2012) 13 Arbuscular mycorrhizal fungi
Arbuscular mycorrhizal fungi are aseptate coenocytic organisms belonging to the phylum Glomeromycota (Hibbett et al 2007) They have been in existence for more than 400 million years and all this while the morphology of AMF remained unaltered therefore AMF are regarded as living fossils (Parniske 2008) They are considered by many to be ancient asexuals a characteristic that defies the predictions of evolutionary theory (Parniske 2008) Although there is no confirmed report of a sexual stage in the life cycle of AMF it is possible that genetic material is exchanged and recombined anastomosis between hyphae allows the exchange of nuclei but has so far
Fig 13 Arbuscular mycorrhizal spores as seen under a stereo zoom microscope
Introduction
5
only been observed between hyphae of closely related fungal strains (Giovanetti et al 2004 de la Providencia et al 2005) Although spores of AMF (Fig 13) can germinate in the absence of host plants they are obligate biotrophs and therefore depend on a living photoautotrophic partner to complete their life cycle and produce the next generation of spores (Parniske 2008)
Plant root symbioses with fungi occur in several different forms and are referred to as mycorrhiza (from the Greek lsquomycosrsquo meaning fungus and lsquorhizarsquo meaning root) (Parniske 2008) Symbiotic development results in the formation of dichotomously branched intra-cellular structures within plant cells These structures which are known as arbuscules (from the Latin lsquoarbusculumrsquo meaning bush or little tree) (Fig 14) are considered to be the primary site of nutrient exchange between the fungal and plant symbiotic partners (Smith and Read 2008 Parniske 2008)
Fig 14 Light micrograph of an arbuscule in root cortical cell (A) and its diagrammatic representation (B Parniske 2008) The extra radical mycelium (ERM) surrounding the root is profusely branched absorbing hyphae which form a network extending into the soil ERM increases the total absorptive surface area of root and help in acquiring nutrients even beyond the depletion zone which develops around plant roots (George et al 1992) Thus AM intimately connects plants to the hyphal network of the fungi which can be in excess of 100 meters of hyphae per cubic centimeter of soil (Miller et al 1995) This hyphal network is specialized for nutrient (predominantly phosphate) and water uptake (Finlay 2008) In return for supplying plants with nutrients and water AMF obtain
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
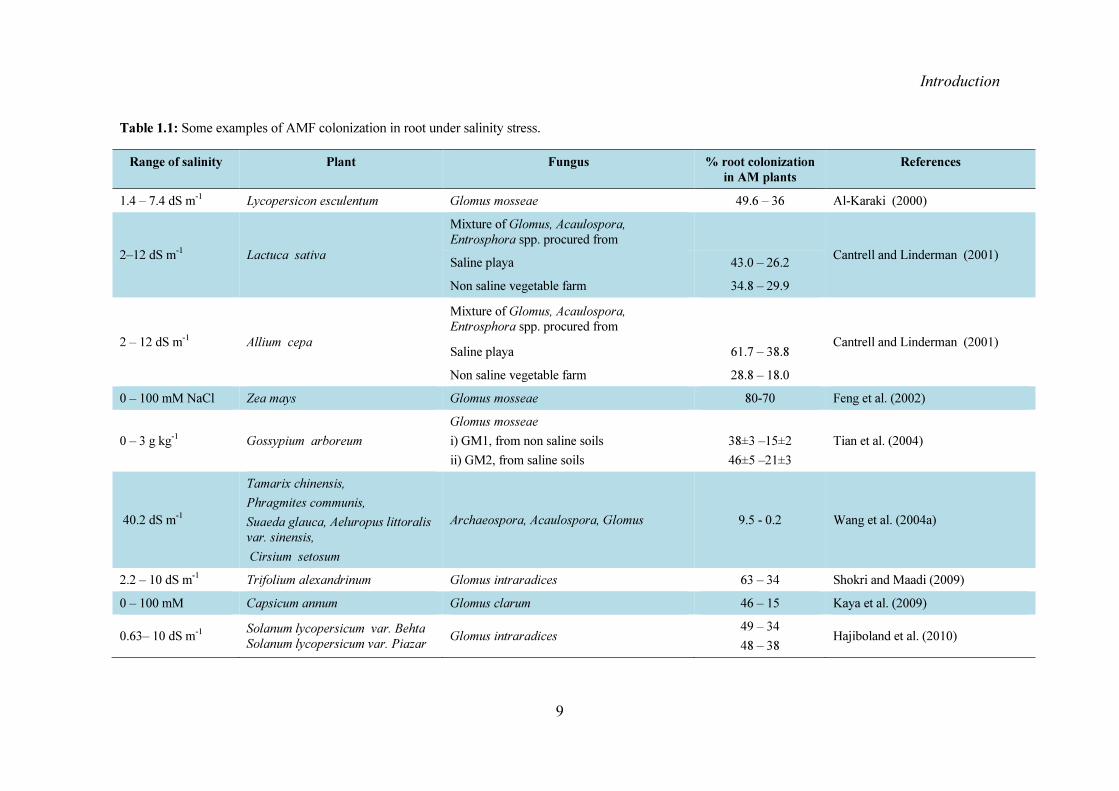
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
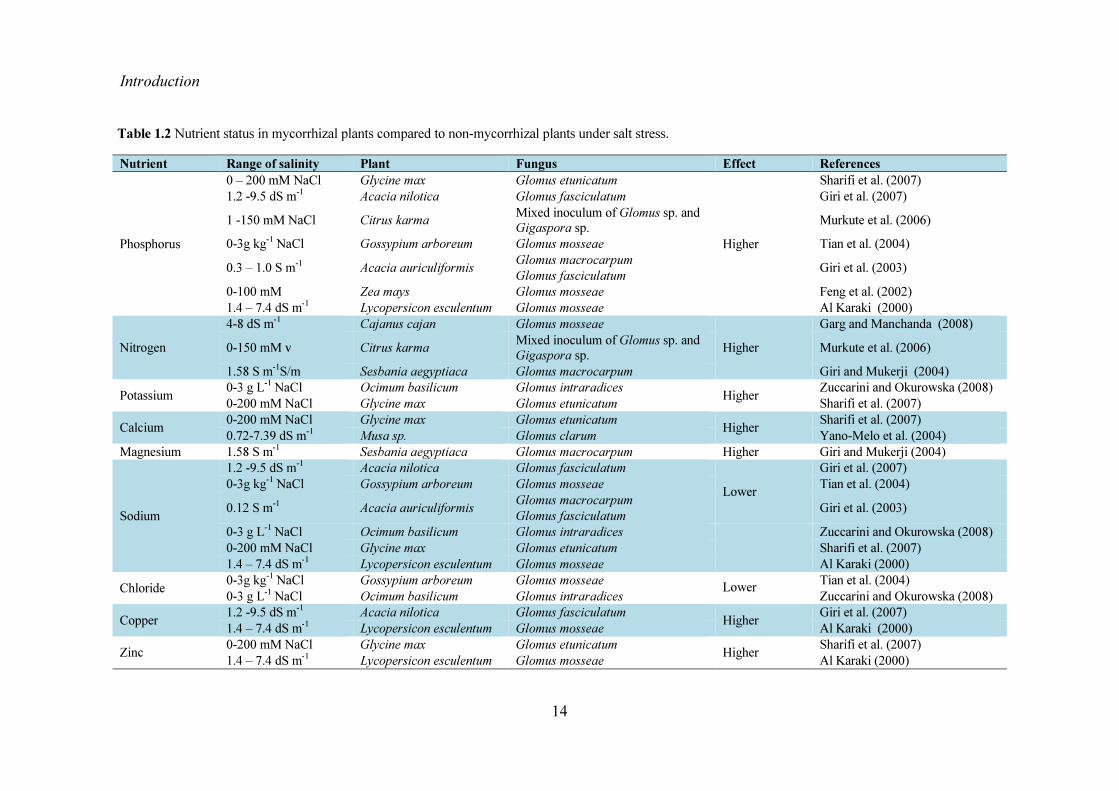
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
2
resulted in decreased crop production of more than 20 irrigated land worldwide (Porcel et al 2012) 11 Effect of salinity on plants In agriculture the impact of soil salinity on plant is alarming as it affects the establishment growth and development of plants leading to enormous decline in productivity (Tester and Davenport 2003 Giri et al 2003) Excessive salts in soil in particular Na+ ions alter the basic organization of the soil (Mahajan and Tuteja 2005) The occurrence of Na+ ions in the cation exchange complex makes the soil compact and subsequently reduces soil porosity and hampers soil aeration (Manchanda and Garg 2008) Low soil aeration due to high salt concentration has a direct relation with all major living processes such as reduction in growth photosynthesis protein and lipid metabolism (due to salt-induced osmotic imbalance) nutritional disorder and ion toxicity in plants (Porcel et al 2012) (Fig 12) Osmotic inequilibrium in a salt-stressed plant is often translated as retardation in growth of the plants with the leaves and stems appearing stunted (Singh and Charath 2001) This effect of salt is primarily due to ndash (i) decrease in the plantrsquos ability to take up water and nutrients as a result of osmotic or water-deficit (physiological drought) effect of salt and (ii) uptake of salt by plants from the soil through transpiration stream injure cells in the transpiring leaves thereby inhibiting cell division and enlargement in plantrsquos growing point (Manchanda and Garg 2008) Stunted growth of the plants grown in saline soils may also be a consequence of sidetracking of energy to counterbalance the accumulation of salts in the cells Continued uptake of salt by plants and subsequent significant increase in the concentration of salts decreases the size of the leaves (Singh and Charath 2001) affect the structure of chlorophyll molecules resulting in disruption of photosynthesis and leaf senescence Specific effects of salt stress on leaf senescence have been related to accumulation of toxic ions (Na+ and Cl-) or to K+ and Ca2+ depletion (Yeo et al 1991) Salt induced ionic inequilibrium and toxicity is precursor for disruption in the plant mineral relations This may be elucidated by the effects of salinity on nutrient availability competitive uptake transport or partitioning within the plant or may be caused by physiological inactivation of a given nutrient resulting in an increase in the
Introduction
3
plants internal requirement for that essential element thereby causing ionic imbalance in the cell (Grattan and Grieve 1999) At the whole plant level salinity frequently induces an increase in Na+ and Cl- ions as well as a decrease in K+ Ca2+ NO3
- and Pi
concentrations (Shokri and Maadi 2009) Therefore high concentrations of Na+ and Cl- ions in the soil solution may depress nutrient-ion activities and produce extreme ratios of Na+ K+ Na+ Ca2+ Ca2+ Mg2+ and Cl- NO3
- (Grattan and Grieve 1999)
Fig 12 Effect of salt stress on plants Salt stress causes physiological drought to plants resulting in hyperosmotic stress On the other hand excessive uptake of Na+ and Cl- ions lead to hyperionic stress These stresses generate a secondary stress ldquooxidative stressrdquo The combined effect of these stresses ultimately affect plant growth and reduce the yield 12 Strategies to minimize crop loss due to salinity Improving plant tolerance against salinity and maintaining crop productivity in saline soils is a major challenge for sustainable agriculture Huge decline in crop productivity due to salt stress have encouraged the researchers around the world to develop strategies to minimize crop loss Searching for new salt tolerant crop plants and development of salt tolerant crops through breeding are major strategies used so far (Gallagher 1985 Glenn and OrsquoLeary 1985 Cuartero and Fernandez-Munoz 1999)
Introduction
4
Other methods include leaching of excessive salts or desalinizing seawater for use in irrigation (Muralev et al 1997) Though successful these approaches are costly and beyond the economic means of developing nations (Cantrell and Linderman 2001)
Under natural conditions plants can establish symbiosis with microorganisms such as bacteria and fungi These associations have been shown to improve plant performance under stress environments and consequently improve their yield (Brown 1974 Levy et al 1983 Creus et al 1998) Of these microorganisms arbuscular mycorrhizal fungi (AMF) have proved to be an efficient bio-ameliorator of salinity effects in plants The wide host range of AMF ndash it can establish symbiosis with roots of 70 terrestrial plant species including halophytes hydrophytes and xerophytes is an added advantage (Smith and Read 2008 Hejiden et al 1998) AM symbioses have been shown to increase tolerance to soil salinity in a variety of host plants such as Capsicum annum Astragalus sinicus Zea mays (Turkmen et al 2008 Peng et al 2011 Sheng et al 2011) It is clear that AMF ameliorate growth inhibition caused by salinity however the mechanism involved remains unresolved (Porcel et al 2012) 13 Arbuscular mycorrhizal fungi
Arbuscular mycorrhizal fungi are aseptate coenocytic organisms belonging to the phylum Glomeromycota (Hibbett et al 2007) They have been in existence for more than 400 million years and all this while the morphology of AMF remained unaltered therefore AMF are regarded as living fossils (Parniske 2008) They are considered by many to be ancient asexuals a characteristic that defies the predictions of evolutionary theory (Parniske 2008) Although there is no confirmed report of a sexual stage in the life cycle of AMF it is possible that genetic material is exchanged and recombined anastomosis between hyphae allows the exchange of nuclei but has so far
Fig 13 Arbuscular mycorrhizal spores as seen under a stereo zoom microscope
Introduction
5
only been observed between hyphae of closely related fungal strains (Giovanetti et al 2004 de la Providencia et al 2005) Although spores of AMF (Fig 13) can germinate in the absence of host plants they are obligate biotrophs and therefore depend on a living photoautotrophic partner to complete their life cycle and produce the next generation of spores (Parniske 2008)
Plant root symbioses with fungi occur in several different forms and are referred to as mycorrhiza (from the Greek lsquomycosrsquo meaning fungus and lsquorhizarsquo meaning root) (Parniske 2008) Symbiotic development results in the formation of dichotomously branched intra-cellular structures within plant cells These structures which are known as arbuscules (from the Latin lsquoarbusculumrsquo meaning bush or little tree) (Fig 14) are considered to be the primary site of nutrient exchange between the fungal and plant symbiotic partners (Smith and Read 2008 Parniske 2008)
Fig 14 Light micrograph of an arbuscule in root cortical cell (A) and its diagrammatic representation (B Parniske 2008) The extra radical mycelium (ERM) surrounding the root is profusely branched absorbing hyphae which form a network extending into the soil ERM increases the total absorptive surface area of root and help in acquiring nutrients even beyond the depletion zone which develops around plant roots (George et al 1992) Thus AM intimately connects plants to the hyphal network of the fungi which can be in excess of 100 meters of hyphae per cubic centimeter of soil (Miller et al 1995) This hyphal network is specialized for nutrient (predominantly phosphate) and water uptake (Finlay 2008) In return for supplying plants with nutrients and water AMF obtain
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
3
plants internal requirement for that essential element thereby causing ionic imbalance in the cell (Grattan and Grieve 1999) At the whole plant level salinity frequently induces an increase in Na+ and Cl- ions as well as a decrease in K+ Ca2+ NO3
- and Pi
concentrations (Shokri and Maadi 2009) Therefore high concentrations of Na+ and Cl- ions in the soil solution may depress nutrient-ion activities and produce extreme ratios of Na+ K+ Na+ Ca2+ Ca2+ Mg2+ and Cl- NO3
- (Grattan and Grieve 1999)
Fig 12 Effect of salt stress on plants Salt stress causes physiological drought to plants resulting in hyperosmotic stress On the other hand excessive uptake of Na+ and Cl- ions lead to hyperionic stress These stresses generate a secondary stress ldquooxidative stressrdquo The combined effect of these stresses ultimately affect plant growth and reduce the yield 12 Strategies to minimize crop loss due to salinity Improving plant tolerance against salinity and maintaining crop productivity in saline soils is a major challenge for sustainable agriculture Huge decline in crop productivity due to salt stress have encouraged the researchers around the world to develop strategies to minimize crop loss Searching for new salt tolerant crop plants and development of salt tolerant crops through breeding are major strategies used so far (Gallagher 1985 Glenn and OrsquoLeary 1985 Cuartero and Fernandez-Munoz 1999)
Introduction
4
Other methods include leaching of excessive salts or desalinizing seawater for use in irrigation (Muralev et al 1997) Though successful these approaches are costly and beyond the economic means of developing nations (Cantrell and Linderman 2001)
Under natural conditions plants can establish symbiosis with microorganisms such as bacteria and fungi These associations have been shown to improve plant performance under stress environments and consequently improve their yield (Brown 1974 Levy et al 1983 Creus et al 1998) Of these microorganisms arbuscular mycorrhizal fungi (AMF) have proved to be an efficient bio-ameliorator of salinity effects in plants The wide host range of AMF ndash it can establish symbiosis with roots of 70 terrestrial plant species including halophytes hydrophytes and xerophytes is an added advantage (Smith and Read 2008 Hejiden et al 1998) AM symbioses have been shown to increase tolerance to soil salinity in a variety of host plants such as Capsicum annum Astragalus sinicus Zea mays (Turkmen et al 2008 Peng et al 2011 Sheng et al 2011) It is clear that AMF ameliorate growth inhibition caused by salinity however the mechanism involved remains unresolved (Porcel et al 2012) 13 Arbuscular mycorrhizal fungi
Arbuscular mycorrhizal fungi are aseptate coenocytic organisms belonging to the phylum Glomeromycota (Hibbett et al 2007) They have been in existence for more than 400 million years and all this while the morphology of AMF remained unaltered therefore AMF are regarded as living fossils (Parniske 2008) They are considered by many to be ancient asexuals a characteristic that defies the predictions of evolutionary theory (Parniske 2008) Although there is no confirmed report of a sexual stage in the life cycle of AMF it is possible that genetic material is exchanged and recombined anastomosis between hyphae allows the exchange of nuclei but has so far
Fig 13 Arbuscular mycorrhizal spores as seen under a stereo zoom microscope
Introduction
5
only been observed between hyphae of closely related fungal strains (Giovanetti et al 2004 de la Providencia et al 2005) Although spores of AMF (Fig 13) can germinate in the absence of host plants they are obligate biotrophs and therefore depend on a living photoautotrophic partner to complete their life cycle and produce the next generation of spores (Parniske 2008)
Plant root symbioses with fungi occur in several different forms and are referred to as mycorrhiza (from the Greek lsquomycosrsquo meaning fungus and lsquorhizarsquo meaning root) (Parniske 2008) Symbiotic development results in the formation of dichotomously branched intra-cellular structures within plant cells These structures which are known as arbuscules (from the Latin lsquoarbusculumrsquo meaning bush or little tree) (Fig 14) are considered to be the primary site of nutrient exchange between the fungal and plant symbiotic partners (Smith and Read 2008 Parniske 2008)
Fig 14 Light micrograph of an arbuscule in root cortical cell (A) and its diagrammatic representation (B Parniske 2008) The extra radical mycelium (ERM) surrounding the root is profusely branched absorbing hyphae which form a network extending into the soil ERM increases the total absorptive surface area of root and help in acquiring nutrients even beyond the depletion zone which develops around plant roots (George et al 1992) Thus AM intimately connects plants to the hyphal network of the fungi which can be in excess of 100 meters of hyphae per cubic centimeter of soil (Miller et al 1995) This hyphal network is specialized for nutrient (predominantly phosphate) and water uptake (Finlay 2008) In return for supplying plants with nutrients and water AMF obtain
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
4
Other methods include leaching of excessive salts or desalinizing seawater for use in irrigation (Muralev et al 1997) Though successful these approaches are costly and beyond the economic means of developing nations (Cantrell and Linderman 2001)
Under natural conditions plants can establish symbiosis with microorganisms such as bacteria and fungi These associations have been shown to improve plant performance under stress environments and consequently improve their yield (Brown 1974 Levy et al 1983 Creus et al 1998) Of these microorganisms arbuscular mycorrhizal fungi (AMF) have proved to be an efficient bio-ameliorator of salinity effects in plants The wide host range of AMF ndash it can establish symbiosis with roots of 70 terrestrial plant species including halophytes hydrophytes and xerophytes is an added advantage (Smith and Read 2008 Hejiden et al 1998) AM symbioses have been shown to increase tolerance to soil salinity in a variety of host plants such as Capsicum annum Astragalus sinicus Zea mays (Turkmen et al 2008 Peng et al 2011 Sheng et al 2011) It is clear that AMF ameliorate growth inhibition caused by salinity however the mechanism involved remains unresolved (Porcel et al 2012) 13 Arbuscular mycorrhizal fungi
Arbuscular mycorrhizal fungi are aseptate coenocytic organisms belonging to the phylum Glomeromycota (Hibbett et al 2007) They have been in existence for more than 400 million years and all this while the morphology of AMF remained unaltered therefore AMF are regarded as living fossils (Parniske 2008) They are considered by many to be ancient asexuals a characteristic that defies the predictions of evolutionary theory (Parniske 2008) Although there is no confirmed report of a sexual stage in the life cycle of AMF it is possible that genetic material is exchanged and recombined anastomosis between hyphae allows the exchange of nuclei but has so far
Fig 13 Arbuscular mycorrhizal spores as seen under a stereo zoom microscope
Introduction
5
only been observed between hyphae of closely related fungal strains (Giovanetti et al 2004 de la Providencia et al 2005) Although spores of AMF (Fig 13) can germinate in the absence of host plants they are obligate biotrophs and therefore depend on a living photoautotrophic partner to complete their life cycle and produce the next generation of spores (Parniske 2008)
Plant root symbioses with fungi occur in several different forms and are referred to as mycorrhiza (from the Greek lsquomycosrsquo meaning fungus and lsquorhizarsquo meaning root) (Parniske 2008) Symbiotic development results in the formation of dichotomously branched intra-cellular structures within plant cells These structures which are known as arbuscules (from the Latin lsquoarbusculumrsquo meaning bush or little tree) (Fig 14) are considered to be the primary site of nutrient exchange between the fungal and plant symbiotic partners (Smith and Read 2008 Parniske 2008)
Fig 14 Light micrograph of an arbuscule in root cortical cell (A) and its diagrammatic representation (B Parniske 2008) The extra radical mycelium (ERM) surrounding the root is profusely branched absorbing hyphae which form a network extending into the soil ERM increases the total absorptive surface area of root and help in acquiring nutrients even beyond the depletion zone which develops around plant roots (George et al 1992) Thus AM intimately connects plants to the hyphal network of the fungi which can be in excess of 100 meters of hyphae per cubic centimeter of soil (Miller et al 1995) This hyphal network is specialized for nutrient (predominantly phosphate) and water uptake (Finlay 2008) In return for supplying plants with nutrients and water AMF obtain
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
5
only been observed between hyphae of closely related fungal strains (Giovanetti et al 2004 de la Providencia et al 2005) Although spores of AMF (Fig 13) can germinate in the absence of host plants they are obligate biotrophs and therefore depend on a living photoautotrophic partner to complete their life cycle and produce the next generation of spores (Parniske 2008)
Plant root symbioses with fungi occur in several different forms and are referred to as mycorrhiza (from the Greek lsquomycosrsquo meaning fungus and lsquorhizarsquo meaning root) (Parniske 2008) Symbiotic development results in the formation of dichotomously branched intra-cellular structures within plant cells These structures which are known as arbuscules (from the Latin lsquoarbusculumrsquo meaning bush or little tree) (Fig 14) are considered to be the primary site of nutrient exchange between the fungal and plant symbiotic partners (Smith and Read 2008 Parniske 2008)
Fig 14 Light micrograph of an arbuscule in root cortical cell (A) and its diagrammatic representation (B Parniske 2008) The extra radical mycelium (ERM) surrounding the root is profusely branched absorbing hyphae which form a network extending into the soil ERM increases the total absorptive surface area of root and help in acquiring nutrients even beyond the depletion zone which develops around plant roots (George et al 1992) Thus AM intimately connects plants to the hyphal network of the fungi which can be in excess of 100 meters of hyphae per cubic centimeter of soil (Miller et al 1995) This hyphal network is specialized for nutrient (predominantly phosphate) and water uptake (Finlay 2008) In return for supplying plants with nutrients and water AMF obtain
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
6
carbohydrates from plants Up to 20 of the photosynthesis products of terrestrial plants (approximately 5 billion tonnes of carbon per year) are estimated to be consumed by AMF (Solaiman and Saito 1997 Bago et al 2003) Therefore AM symbiosis is a significant contributor to global phosphate and carbon cycling influencing primary productivity in terrestrial ecosystems (Bago et al 2000) The beneficial effects of AM are most apparent under conditions of limited nutrient availability (Fitter 2005) Although the underlying regulatory mechanisms are not understood the amount of root colonization typically decreases when nutrients are in abundance (Parniske 2008) 14 AMF in saline soils Saline environment is a deterrent for organisms except the halophiles AMF despite its low affinity for halophytes (Brundrett 1991) have been found in saline soils (Khan 1974 Allen and Cunningham 1983 Pond et al 1984 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001 Harisnaut et al 2003 Yamato et al 2008) While some researchers report low density of AMF spores in saline soils (Barrow et al 1997 Carvalho et al 2001) some others found high spore density (Khan 1974 Bhaskaran and Selvaraj 1997 Aliagharzadeh et al 2001 Landwehr et al 2002) Aliasgharzadeh et al (2001) observed spores of Glomus intraradices G versiforme and G etunicatum in the severely saline soils of Tabriz plains with ~ 162 dS m-1 electrical conductivity The authors also found that the number of AMF spores did not significantly decrease with soil salinity and reported relatively a high spore number (mean of 100 per 10 g soil) The accumulation of spores may be explained by the inhibition of spore germination and hyphal growth of AMF by salt (McMillen et al 1998 Aliasgharzadeh et al 2001) Wang et al (2004a) while investigating the relationship between the distribution of AMF in the rhizosphere of different wild plants in Yellow River Delta (EC ~ 402 dS m-1) observed a total of 33 species representing three genera of AMF including two species of Archaeospora seven in Acaulospora and twenty four in Glomus They also found most spores at a depth of 0ndash40 cm Number of AMF spores decreased with increasing soil depth in the rhizosphere Ho (1987) had reported the same while studying AM of halophytic grasses in the Alvord Desert of Oregon
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
7
The occurrence of AMF in salt marsh plants has also been reported by many authors (Khan 1974 Rozema et al 1986 Sengupta and Chaudhuri 1990 Carvalho et al 2001 Hilderbrandt et al 2001) The occurrence of AMF spores in soils is independent of NaCl Na2CO3 Na2SO4 or CaSO4 salt types though the degree of colonization varied from one individual to the next (Landwehr et al 2002) AMF spores are also found in extremely alkaline soils of pH values up to 11 Evaluation of the distribution of AMF spores in two salt marshes ndash Terschelling (an almost natural site at the Atlantic Coast where the NaCl content in the sea is about 35 ECe ~ 62 ndash 190 mS m-1) The Netherlands and at Schreyahn (of anthropogenic origin due to potash mining ECe ~ 63 ndash 201 mS m-1) Germany revealed that the distribution of AMF spores unlikely followed the salt gradient at both sites (Wilde et al 2009) The finding may implicate that distribution and or abundance of AMF spores may not entirely depend on salt concentration but also different on edaphoclimatic factors AMF spore density share a positive correlation with soil pH sand per cent and organic carbon (Ho 1987 Mohammed et al 2003 Mathur et al 2007 Aliasgharzadeh et al 2001) and a negative correlation available soil Mg Ca Cl clay electrical conductivity SO4 sodium absorption ratio (Aliasgharzadeh et al 2001 Saint Etienne et al 2006)
In most of the studies above the AMF spores were identified based on morphological and molecular characteristics (Landwehr et al 2002 Regvar et al 2003 Wilde et al 2009) The most commonly found AMF in saline soils are Glomus spp (Allen and Cunningham 1983 Ho 1987 Wang et al 2004a) Molecular analyses revealed that on an average 80 of these spores belonged to one single species Glomus geosporum (Wilde et al 2009) Though molecular identification techniques can overcome some of the pitfalls in morphology based identification it is not advisable to solely rely on molecular identification techniques because gene content of the different nuclei within an AMF individual is to some extent variable (Lloyd-MacGilip et al 1996 Sanders et al 1996) The sequences of the ITS region within a single spore can be more variable than between spores of a single isolate (Pringle et al 2000) The divergence of the ITS region within a single spore can be between 24 and 57 (Antoniolli et al 2000) Therefore to define a species any characterization of a microbe by sequencing of ribosomal genes needs to be complemented by morphological andor physiological characterization (Wilde et al 2009)
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
8
15 Effect of salinity on colonization spore germination and hyphal growth Salinity not only affects the host plant but also its symbiotic partner the arbuscular mycorrhizal fungi It can upset the ability of AMF spore to germinate grow and colonize the roots of host plant (Hirrel 1981 Estaun 1989 Juniper and Abbott 1993 McMillen et al 1998) Table 11 shows some of the studies on per cent colonization of AMF under salt stress conditions Direct effect of NaCl on the fungus the plant or both the plant and the fungus suppresses colonization of plant roots by some AMF fungi under saline conditions (Hirrel and Gerdemann 1980 Ojala et al 1983 Menconi et al 1995 Poss et al 1985 Rozema et al 1986 Duke et al 1986 Giri et al 2007 Juniper and Abbott 2006 Sheng et al 2008 McMillen et al 1998 Juniper and Abbott 2006) Salt-induced inhibition of AMF colonization is evident more in the early than in the later stages of the symbiosis (McMillen et al 1998) In the presence of NaCl germination of spores is delayed rather than prevented (Hirrel 1981 Juniper and Abbott 2006) Studies showed that pre-inoculation of a transplant with AMF fungi bypasses the inhibitory effects that salt could have on spore germination (Al Karaki 2006 Cantrell and Lindermann 2001)The rate of germination and maximum germination of AMF spores may also depend on the salt type (Juniper and Abbott 1993) According to them different salts NaNO3 and Na2SO4 with similar osmotic potentials (-048 and -043 MPa respectively) impart differential effects on the rate and maximum germination of spores They attributed the difference to a higher concentration of Na+ in the latter They also reported that NaCl allows faster rate and maximum germination as compared to KCl with similar osmotic potentials
Contrary to the reports above a few studies showed no effect of NaCl on colonization (Levy et al 1983 Hartmond et al 1987) In fact increased AMF fungal sporulation and colonization were observed when the plants are treated with 200 mM NaCl (Aliasgharzadeh et al 2001 Yamato et al 2008) Here it may be recalled that the establishment of arbuscular mycorrhiza may also be influenced by soil properties host plants and species and isolates of AMF which differ in their tolerance to adverse physical and chemical conditions in soil (Hayman 1982 Juniper and Abbott 1993 Joshi and Singh 1995) For instance chenopod plants are not commonly mycorrhizal but may become mycorrhizal when exposed to saline conditions (Sengupta and Chaudhuri 1990)
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
9
Table 11 Some examples of AMF colonization in root under salinity stress
Range of salinity Plant Fungus root colonization in AM plants
2ndash12 dS m-1 Lactuca sativa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 430 ndash 262 Non saline vegetable farm 348 ndash 299
2 ndash 12 dS m-1 Allium cepa Mixture of Glomus Acaulospora Entrosphora spp procured from
Cantrell and Linderman (2001) Saline playa 617 ndash 388 Non saline vegetable farm 288 ndash 180
0 ndash 100 mM NaCl Zea mays Glomus mosseae 80-70 Feng et al (2002)
0 ndash 3 g kg-1 Gossypium arboreum Glomus mosseae i) GM1 from non saline soils ii) GM2 from saline soils
38plusmn3 ndash15plusmn2 46plusmn5 ndash21plusmn3
Tian et al (2004)
402 dS m-1
Tamarix chinensis Phragmites communis Suaeda glauca Aeluropus littoralis var sinensis Cirsium setosum
Archaeospora Acaulospora Glomus 95 - 02 Wang et al (2004a)
22 ndash 10 dS m-1 Trifolium alexandrinum Glomus intraradices 63 ndash 34 Shokri and Maadi (2009) 0 ndash 100 mM Capsicum annum Glomus clarum 46 ndash 15 Kaya et al (2009)
063ndash 10 dS m-1 Solanum lycopersicum var Behta Solanum lycopersicum var Piazar Glomus intraradices 49 ndash 34
48 ndash 38 Hajiboland et al (2010)
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
10
Range of salinity Plant Fungus root colonization in AM plants
References
0 ndash 100 mM Poncirus trifoliate Glomus versiforme G mosseae
653-595 775 ndash 678 Wu et al (2006a)
0 ndash 100 mM NaCl Citrus tangerine Glomus mosseae ca 70 ndash 40 Wu et al (2006b)
100 ndash 300 mM NaCl Pennisetum glaucum Glomus fasciculatum 60 days 90 days
33 ndash 20 50 ndash 33
Borde et al (2011)
0 ndash 100 mM NaCl Lycopersicon esculentum Glomus mosseae 55 ndash 27 Latef and Chaoxing (2011) 0 ndash 200 mM NaCl Prosopis alba Glomus intraradices 70 Scambato et al (2011) 08 ndash 36 dS m-1 Zea mays Glomus mosseae 99-79 Sheng et al (2011)
0 ndash 069 g kg-1 Astragalus sinicus Glomus mosseae Gintraradices G claroideum
85-30 90-40 95-85
Peng et al (2011)
015 ndash 938 mS cm-1 Triticum aestivum var Sakha 8 Triticum aestivum var Giza Mixture of Glomus spp ca 85 ndash 60
ca 78 ndash 50 Talaat and Shawky (2011)
1ndash 6 dS m-1 Dianthus caryophyllus Glomus intraradices 26 ndash 4 Navarro et al (2012)
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
11
Inhibition of hyphal growth of AMF in saline soil may be a result of ion toxicity or salt-induced osmotic stress (McMillen et al 1998 Cantrell and Lindermann 2001 Juniper and Abbott 2006) Hyphal growth is an energy-dependent process and requires the AMF spores to maintain turgor (Cooke and Whipps 1993) Under such circumstances osmotic stress caused by salinity imposes strain on hyphal extension thus limiting its growth Therefore the number of hyphae produced as well as the growth rate of hyphae of AMF growing from propagules (eg spores and hyphal fragments) may decrease in soil containing high concentrations of NaCl (McMillen et al 1998) These reports suggests that hyphal growth of the fungi can be taken as being more sensitive to NaCl than spore germination which is delayed but not necessarily reduced
The relative tolerance of different types of the same fungal genus can vary as is reported in the case of Glomus sp While propagules of Glomus sp within the colonized root pieces grow in 300 mM NaCl spores of the same fungi extracted from the soil did not This variation in tolerance may be attributed to difference in the energy status between them or differences in the amount of water and energy required to initiate germination (Juniper and Abbott 2006) Tommerup (1984) indicated that initial hyphal extensions have higher requirements for water than the intermediate stages for activation and germ tube production Therefore it may be possible that the germination rate of a prehydrated spore will have greater percentage than the non prehydrated spore in saline environment (Juniper and Abbott 2006) This contrasting result invites further studies in this aspect
In a first report on the effect of salinity on Glomus intraradices under in vitro conditions Jahromi et al (2008) found no significant difference in hyphal length and number of branched absorbing structures (BAS) between control (no salt) and 50 mM NaCl However at 100 mM NaCl the authors observed a significant decrease in hyphal length and the number of BAS Also salinity reduced the number of spores produced by Glomus intraradices Therefore decrease in AMF colonization of host roots under saline conditions may be related to the reduced ability of AMF to sporulate and the decreasing hyphal length BAS is believed to be associated with spore formation Bago et al (1998) had also shown that BAS can gradually form spores in their ramification So the reduction in the number of BAS can also further reduce sporulation
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
12
16 Contributions of arbuscular mycorrhizal fungi in amelioration of salt stress in plants
Studies have shown that under salt stress AMF colonization of host roots often results in better plant growth and biomass compared to non-mycorrhizal (NM) plants Better growth and biomass in mycorrhizal (M) plants have been correlated with AMF-mediated increased nutrient uptake accumulation of osmoregulators increase in photosynthetic rate and water use efficiency The strategies used by AM to protect plants from the detrimental effects of salinity are discussed below 161 Improving plant growth and biomass Plants grown in saline soils exhibit reduced fitness measured in terms of growth and biomass (Al Karaki 2000 Giri and Mukerji 2004 Porcel et al 2012) This may be due to inability of plants to acquire nutrients from the soil as nutrient ions either gets converted to non-available forms or competes with Na+ or Cl- ions for its uptake by plants Another factor that limits plant growth under salinity is the forced channelization of energy towards establishing tolerance to salinity Salinity induced retardation of growth and fitness of plants have been shown to be alleviated by AM Higher growth and biomass in M than the NM plants under salt stress were reported in many plants (Al Karaki 2000 Cantrell and Linderman 2001 Giri et al 2003 Giri et al 2007 Sannazzaro et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Latef and Chaoxing 2011) The increased dependency of host plants on AMF under salt stress is an affirmation that AMF enhance plant growth (Borde et al 2011 Kumar et al 2011) Better growth of M plants as compared to NM plants under salt stress is attributed to improved nutrient uptake especially phosphorus (P) and other nutrients mycorrhiza-mediated effects on water absorption and increased photosynthetic system (Giri et al 2003 Garg and Manchanda 2009 Abdel-Fattah and Asrar 2011 Kumar et al 2011) 162 Nutrient uptake Plants for their growth and survival require 14 essential nutrients of which the macro-nutrients nitrogen phosphorus potassium calcium magnesium and sulphur are present
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
13
in large amounts in tissues The micro-nutrients (copper iron molybdenum manganese zinc) though required in very small amount by the plants are indispensable for plant growth and survival The effects of salinity on availability uptake transport or physiological inactivation of a given nutrient render them unavailable for uptake by plant (Grattan and Grieve 1999) However many studies have shown that this negative effect of salinity on nutrient uptake can be alleviated by colonization of plants by AMF Table 12 lists some examples of AMF influence on nutrient uptake in plants under salt stress The effects of NaCl and AMF colonization on nutrients are described below 1621 Nitrogen Soil salinity affects total N uptake and soil N contribution leading to reduced plant growth Under salt stress the otherwise mobile forms of N are rendered immobile (Hodge and Fitter 2010 Miransari 2010) Salinity interferes with the uptake of both forms of available N nitrate and ammonium ions (Frechilla et al 2001) Uptake of nitrates by plants is challenged by salinity at two levels (i) by direct competition of chloride with nitrate and (ii) at the level of the membrane andor the membrane proteins by changing plasma lemma integrity (Cramer et al 1985 Frechilla et al 2001) However under salt stress plants colonized by AMF displayed higher N-concentration as compared to NM plants (Cantrell and Linderman 2001 Silveira et al 2001 Giri and Mukerji 2004) In M plants N is taken up by ERM as inorganic N from the soil in the form of nitrate and assimilated via nitrate reductase located in the arbuscule-containing cells (Karldorf et al 1998) and converted to arginine via the GS-GOGAT (glutamine synthetase- glutamine 2-oxoglutarate amidotransferase) cycle Arginine in the hyphae is broken down to urea and ultimately transferred to the plant as NH4
+ with the resulting C skeletons from arginine breakdown re-incorporated in to the fungal C pools (Bago et al 2002 Govindarajulu et al 2005) The identification of ammonium transporter gene in ERM of Glomus intraradices (Loacutepez-Pedrosa et al 2006) a mycorrhiza-specific plant ammonium transporter in Lotus japonicus that is expressed in arbusculated cells (Guether et al 2009) up-regulation of an ammonium transporter in Medicago truncatula (Gomez et al 2009) makes the mechanism clearer
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
14
Nutrient Range of salinity Plant Fungus Effect References
Phosphorus
0 ndash 200 mM NaCl Glycine max Glomus etunicatum
Higher
Sharifi et al (2007) 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Giri et al (2007) 1 -150 mM NaCl Citrus karma Mixed inoculum of Glomus sp and
Gigaspora sp Murkute et al (2006) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 03 ndash 10 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-100 mM Zea mays Glomus mosseae Feng et al (2002) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Nitrogen 4-8 dS m-1 Cajanus cajan Glomus mosseae
Higher Garg and Manchanda (2008)
0-150 mM v Citrus karma Mixed inoculum of Glomus sp and Gigaspora sp Murkute et al (2006)
158 S m-1Sm Sesbania aegyptiaca Glomus macrocarpum Giri and Mukerji (2004) Potassium 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Higher Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) Calcium 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007)
072-739 dS m-1 Musa sp Glomus clarum Yano-Melo et al (2004) Magnesium 158 S m-1 Sesbania aegyptiaca Glomus macrocarpum Higher Giri and Mukerji (2004)
Giri et al (2007) 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Tian et al (2004) 012 S m-1 Acacia auriculiformis Glomus macrocarpum
Glomus fasciculatum Giri et al (2003) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices
Zuccarini and Okurowska (2008)
0-200 mM NaCl Glycine max Glomus etunicatum Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Chloride 0-3g kg-1 NaCl Gossypium arboreum Glomus mosseae Lower Tian et al (2004) 0-3 g L-1 NaCl Ocimum basilicum Glomus intraradices Zuccarini and Okurowska (2008)
Copper 12 -95 dS m-1 Acacia nilotica Glomus fasciculatum Higher Giri et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Zinc 0-200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 14 ndash 74 dS m-1 Lycopersicon esculentum Glomus mosseae Al Karaki (2000)
Table 12 Nutrient status in mycorrhizal plants compared to non-mycorrhizal plants under salt stress
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
15
Cliquet and Stewart (1993) observed that increased N uptake in M plant is due to change in N metabolism brought about by changes in enzymes of N metabolism (Mathur and Vyas 1996) Studies have reported that improved N nutrition may help to decrease the toxic effects of Na+ ions by decreasing its uptake and this may indirectly help in maintaining the chlorophyll content of the plant
Higher N concentration in leguminous M plants has also been related to AMF-mediated promotion of nodulation and prevents pre-mature nodule senescence during salt stress (Garg and Manchanda 2009) Under salt stress nodule senescence prematurely due to acceleration of lytic activities formation of green pigments from leghaemoglobin (Sarath et al 1986) and loss of nitrogen fixation (Delgardo et al 1994) In M plants higher leghaemoglobin concentration delays the change of colour in nodule from pink to brownish pink due to synthesis of green pigments from leghaemoglobin Mycorrhizal plants also possess a higher nitrogenase activity All these parameters contribute to a higher nitrogen fixing ability of M plants The increase in nitrogenase activity and nitrogen fixation in M plants than NM plants has been attributed to relief from P-stress which is beneficial for the functioning of nitrogenase enzyme of the bacterial symbionts and possibly due to uptake of some essential micro-nutrients which results both in improved growth of plants (Founoune et al 2002) or vice versa (Rabie and Almadini 2005) Therefore it may be suggested that mycorrhizal and nodule symbioses often act synergistically on infection rate mineral nutrition and plant growth (Patreze and Cordeiro 2004) which supports the need for both N and P and increased tolerance of plants to salinity stress (Rabie and Almadini 2005) 1622 Phosphorus Phosphate is a mineral nutrient limiting plant growth at many natural stands due to poor solubility (Hause and Fester 2005) This problem is further deteriorated under saline conditions as phosphate ions precipitates with Ca2+ Mg2+ and Zn2+ ions thereby rendering them unavailable to plants (Azcon-Aguilar et al 1979) Therefore P-solubilization or fertilization is necessary to make them available for uptake by plants However this problem may be ameliorated by AMF colonization of host plants In fact studies have shown that M plants have higher P-concentration than their NM
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
16
counterparts under salt stress (Giri et al 2003 2007 Hajiboland et al 2010 Latef and Chaoxing 2011) Mycorrhizal inoculation can increase P concentration in plants by enhancing its uptake aided by the extensive hyphae of the fungus which allows them to explore more soil volume than the non mycorrhizal plants (Ruiz-Lozano and Azcon 2000) It is estimated that external hyphae deliver up to 80 of a plantrsquos P requirements (Matamoros et al 1999) High affinity phosphate transporters in ERM take up P and transport it within the fungus as polyphosphate and once in the intra-radical hyphae the long chains are hydrolyzed facilitating transfer to the host plant via a phosphate transporter (Harrison and van Burren 1995 Harrison 1999 Bago et al 2002 Ohtomo and Saito 2005) Adding up higher affinity for phosphate ions lower threshold concentration for P ability to store larger amounts of absorbed P than the plant roots also aids the continued movement of P by AM symbiosis into the hyphae (Bolan 1991) Improved P nutrition in AM inoculated plants may improve their growth rate increase antioxidant production and enhanced nodulation and nitrogen fixation in legumes (Feng et al 2002 Alguacil et al 2003 Garg and Manchanda 2008) Enhanced uptake of P on M plants may reduce the negative effects of Na+ and Cl- ions by maintaining vacuolar membrane integrity which facilitates compartmentalization within vacuoles and selective ion intake (Rinaldelli and Mancuso 1996) thereby preventing ions from interfering in metabolic pathways of growth (Cantrell and Linderman 2001) 1623 K+ Na+ ratio Similar physiological properties of K+ and Na+ impose an inevitable competition between these cations for uptake by plants Thus when plants are exposed to NaCl stress K+ Na+ ratio decreases However inoculation of a plant with AMF can minimize the effect of salinity on K+ and Na+ nutrition and maintain favourable K+ Na+ ratio Greater acquisition of K+ ions in M plants than NM plants under saline stress have been reported in many plants (Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008 Shokri and Maadi 2009 Porras-Soriano et al 2009) Subsequently K+ Na+ ratio was higher in M plants compared to NM plants Higher K+ Na+ ratio in M plants is accomplished by regulating the expression and activity of K+ and Na+
transporters and of H+ pumps that generate the driving force for transport of ions (Parida
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
17
and Das 2005) The Na+ H+ antiporter catalyze the transfer of Na+ out of the cytoplasm into either vacuole or apoplasm (Ouziad et al 2006) The higher K+ Na+ ratio helps in preventing the disruption of various K-mediated enzymatic processes and inhibition of protein synthesis High K+ Na+ ratio is also beneficial in influencing the ionic balance of the cytoplasm or Na+ efflux from plants (Allen and Cunningham 1983 Founoune et al 2002 Colla et al 2008) Lower Na+ in the M plants than NM plants may also be explained by the dilution effect due to growth enhancement (Al Karaki 2000 2006)
There are contrasting reports that AMF sometimes enhance Na+ uptake (Allen and Cunningham 1983) while others suggest that AM colonized plants have lower levels of Na+ (Dixon et al 1993 Giri et al 2007 Sharifi et al 2007 Zuccarini and Okurowska 2008) Giri et al (2007) reported increase in Na+ concentration in AM plants with increasing salinity levels up to a certain level and subsequently decreased at higher salinity This suggests that AMF induce buffering effect on the uptake of Na+
when the content of Na+ is within the permissible limit (Allen and Cunningham 1983) This also indicates the possibility of a regulatory mechanism operating in the plant to contain Na+ ions 1624 Calcium NaCl induced Ca2+ deficiency has been reported in different plants such as strawberry tomato and beans (Kaya et al 2002 Tuna et al 2007 Cabot et al 2009) Studies revealed that mycorrhizal colonization has a strong effect on Ca2+ concentration of plants Under salt stress mycorrhizal lettuce and banana were found to possess higher Ca2+ concentration than their NM equivalents (Cantrell and Lindermann 2001 Yano-Melo et al 2003) High Ca2+ facilitates higher K+ Na+ selectivity in plants leading to salt adaptation (Cramer et al 1985 Rabie and Almadini 2005) Also high Ca2+ was also found to enhance colonization and sporulation of AMF (Jarstfer et al 1998) However in contrast to the reports above Giri et al (2003) reported that Ca2+ concentration remains unchanged in shoot tissues of M and NM plants This suggests that AMF may not be so important for the nutrients moving to plant roots by mass flow as compared to nutrients moving by diffusion (Tinker 1975) Rhodes and Gerdemann (1978) indicated that Ca+2 is not translocated to roots through mycorrhizal hyphae as readily and
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
18
efficiently as P Moreover AMF inoculation depressed CaP ratio by increased production of oxalate in the mycorrhizosphere which is able to scavenge Ca+2 from the solution (Azcon and Barea 1992) 1625 Magnesium The macro-nutrient Mg constitutes an integral component of the chlorophyll molecule Biosynthesis of chlorophyll is impeded by salt stress This results in disruption in light harvesting and photosynthesis Mycorrhiza by improving Mg2+
can support higher chlorophyll concentration (Giri et al 2003) This suggests that interference of salt on chlorophyll synthesis is less in M than NM plants (Giri and Mukerji 2004) Though reports of decreased chlorophyll concentration and photosynthesis in response to salinity and its amelioration by application of AMF are widely reported (Sheng et al 2008 Latef and Chaoxing 2011) study on Mg2+ nutrition as affected by salinity and mycorrhization has drawn attention of a very few workers (Cantrell and Linderman 2001 Colla et al 2008 Shokri and Maadi 2009) 1626 Chloride ions High tissue Cl- concentrations can be toxic to crop plants and may restrict the agriculture of saline regions (Xu et al 2000) Root cells take up Cl- from the soil solution through H+Cl- symporters at low [Cl-]ext and also through anion channels under saline condition To reach the xylem and then the shoot Cl-traverses the root by a symplastic pathway and is released from cells within the stele through specific anion channels At high salinity Cl- accumulation in aerial parts increases greatly though it remains constant in roots (White and Broadley 2001) Toxicity of Cl- ions may be controlled by the inoculation of host plants with AMF (Zuccarini and Okurowska 2008) The Cl- ions can be compartmentalized in vacuolar membranes thereby preventing it from interfering with the metabolic pathways in the plant (Cantrell and Lindermann 2001) However there are reports of increased Cl- accumulation due to AMF colonization This may be explained by the carbon drain imposed by mycorrhizal hyphae on plants which enhances the translocation of highly mobile anions like Cl- from the soil (Graham and Syversten 1984)
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
19
A careful analysis of the above studies revealed that plant nutritionndashAM symbiosis studies under salt stress has been emphasized on enhanced K+ uptake and improved K+ Na+ ratio in plants and a far and a few have discussed Ca2+ and Mg2+ uptake under salinity stress Although salinity studies have analyzed plant tissue for micronutrients (Cu Fe Mn2+ and Zn2+) in the plants grown in saline soils they have directed little attention to micronutrients as affected by salinity and mycorrhizal colonization Therefore much more can be learned if interactions between salinity levels and mycorrhizal colonization are studied taking more nutrients in consideration 163 Biochemical changes As soil dries out and lowers the soil water potential the water potential in plant cell decreases leading to decrease in cell volume Under salt stress despite the availability of soil water plants face a similar situation known as ldquophysiological droughtrdquo At such conditions metabolic shifts occur and result in changes in the levels of a variety of cellular metabolites Such changes in response to abiotic stress appear to be related with the increased ability to tolerate such conditions (Chen and Murata 2011) One important group of such metabolites includes the so-called lsquocompatible solutesrsquo also known as osmolytes These water-soluble metabolites are non-toxic at high concentrations and do not interfere with normal biochemical reactions rather they replace water in biochemical reactions Frequently investigated osmolytes include proline glycine betaine sugars and polyols (Hasegawa et al 2000 Parida and Das 2005) Osmoregulation allows cells to maintain turgor and turgor-dependent processes including cellular expansion and growth stomatal opening and photosynthesis as well as keeping a gradient of water potential favorable to water entrance into the plant Table 13 lists some examples on the variable AMF effect on some biochemical compounds in plants under salt stress 1631 Proline Proline is one of the most frequently studied osmolytes in relation to water and salt stress in plants When exposed to saline conditions many plants accumulate proline as a
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
20
Table 13 Some studies showing variable AMF effect (higherlower compared to non-mycorrhizal plants) on the following parameters in plants under salt stress
Compound Range of salinity Plant Fungus Effect References
Proline
0 ndash 200 mM NaCl Glycine max Glomus etunicatum Higher Sharifi et al (2007) 4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 0 ndash 6 dS m-1 Vicia faba Glomus clarum Lower Rabie and Almadini (2005) 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Sannazzaro et al (2007)
Polyamines 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007)
Carbohydrates 0 ndash300 mM NaCl Phragmites australis Glomus fasciculatum Higher Al-Garni (2006) 0 ndash 200 mM NaCl Glycine max Glomus etunicatum Lower Sharifi et al (2007)
Amino acids 0 ndash 1 g kg-1 NaCl Zea mays Glomus mosseae Higher Sheng et al 2001
Antioxidants 0 ndash100 mM NaCl Glycine max Glomus etunicatum
Higher Ghorbanli et al (2004)
4 ndash 8 dS m-1 Cajanus cajan Glomus mosseae Garg and Manchanda (2009) 09 ndash 71 dS m-1 Lycopersicon esculentum Glomus mosseae ZhongQun et al (2007)
Abscissic acid 0 ndash 200 mM NaCl Lotus glaber Glomus intraradices Higher Sannazzaro et al (2007) 0 ndash 100 mM NaCl Lactuca sativa Glomus intraradices Lower Jahromi et al (2008)
Chlorophyll 158 S m-1 Sesbania aegyptiaca
Sesbania grandiflora Glomus macrocarpum Giri and Mukerji (2004)
0 ndash 2 g kg-1 Zea mays Glomus mosseae Higher Sheng et al (2008) 084 ndash58 dS m-1 Lactuca sativa Mixture of Glomus mosseae
G intraradices and G coronatum Zuccarini (2007)
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
21
non-toxic and protective osmolyte to maintain osmotic balance under low water potentials (Stewart and Lee 1974 Jain et al 2001 Parida et al 2002 Ashraf and Foolad 2007 Sannazzaro et al 2007) It also acts as a reservoir of energy and nitrogen for utilization during salt stress (Goas et al 1982) Proline accumulation in plants under salt stress has been found to increase when the plant is colonized by AMF Mycorrhizal mung bean (Vigna radiata) plants were reported to have higher proline content than NM plants at 125 and 25 mM NaCl at 40 and 60 days after sowing (Jindal et al 1993) Sharifi et al (2007) also reported a higher proline concentration in M soybean plants than the NM plants They also observed that in M plants a higher level of proline concentration is found in roots than shoots This may be due to the fact that the roots are the primary sites of water absorption and therefore must maintain osmotic balance between water absorbing root cells and the external media However in contrast to the reports above Rabie and Almadini (2005) reported that NM Vicia faba plants accumulated more proline than M plants at various salinity levels This may be explained by the fact that besides its role as an osmo-protectant proline is also an indicator of level of stress (Wang et al 2004b) The accumulation of proline may also be due to salinity and not necessarily by mycorrhizal colonization as reported by Sannazzaro et al (2006) 1632 Glycinebetaine Glycinebetaines are quaternary ammonium compounds which are N-methylated derivatives of amino acids These compounds accumulate in plant cells upon exposure to salt stress Once formed they are seldom metabolized (Duke et al 1986 Grattan and Grieve 1985) This can therefore be used as an effective indicator of salt stress (Duke et al 1986) The role of glycinebetaine is not limited to maintaining turgor but it can also enhance tolerance to salt stress by various ways (i) stabilize the structures and activities of enzymes and protein complexes (ii) maintain the integrity of membranes against the damaging effects of excessive salt (iii) protection of the photosynthetic machinery (iv) induction of specific genes whose products are involved in stress tolerance (v) reductions in levels of ROS under stress and (vi) regulation of the activity of ion-channel proteins either directly or via protection of the plasma membrane (Chen and Murata 2008)
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
22
Inoculation of plants with AMF has been shown to enhance accumulation of glycinebetaine under salt stress (Al Garni 2006 Manchanda and Garg 2008) Glycine betaine concentrations in mycorrhizal Phragmites australis exposed to saline conditions were two-fold greater than the NM equivalents respectively (Al Garni 2006) 1633 Carbohydrates Carbohydrates are a major contributor of osmoregulation in plant cells under saline stress (Thanna and Nawar 1994 Al Garni 2006) It is the plantrsquos inherent adaptive strategy to accumulate more sugars in cells upon exposure to salt stress in order to prevent turgor breakdown Sugar concentration showed significant increase when plants are exposed to increasing concentrations of NaCl (Al Garni 2006) The increase in total carbohydrates was found to be positively correlated with AMF colonization of host plant as reported by Thomson et al (1990) Under salt stress mycorrhizal Phragmites australis and Glycine max had higher sugar concentration compared to their NM counterparts (Al-Garni 2006 Porcel and Ruiz-Lozano 2004) The positive correlation between sugar content and AMF colonization is due to the sink effect of fungus demanding sugars from the shoot tissues (Aue 2000) Mycorrhizal colonization frequently leads to increased rates of photosynthesis and of carbon compound to the root systems of host plants (Finlay and Soumlderstroumlm 1992) The increased sugar accumulation may also be due to hydrolysis of starch to sugars in the mycorrhiza inoculated seedlings (Nemec 1981) Feng et al (2002) studied the prevalence of correlation between P concentration and sugar accumulation in host plants under saline conditions They reported that in spite of similar P concentrations the soluble sugar concentration in roots of mycorrhizal plants was higher than that of the NM maize plants This suggests that the higher soluble sugar concentration in mycorrhizal roots is due to AMF colonization and not due to improvement of the P status of the plants Conversely some authors reported negative correlations between AMF colonization and sugar accumulation in host plants Pearson and Schweiger (1993) reported decrease in carbohydrate concentration with an increase in the per cent root colonization Sharifi et al (2007) observed no role of soluble sugars in responses of Glomus etunicatum colonized soybean plants to salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
23
1634 Polyamines Polyamines as a function of AMF and salinity stress have drawn the least attention among the osmolytes Polyamines are small organic cations that are mandatory for eukaryotic cell growth There are three main polyamines in plants putrescine (Put) spermidine (Spd) and spermine (Spm) Putrescine is synthesized directly from ornithine via ornithine decarboxylase or indirectly following decarboxylation of arginine by arginine decarboxylase Spermidine and Spermine are synthesized from Put by successive addition of aminopropyl groups These cations are thought to play an important role in plant responses to a wide array of environmental stressors such as salinity (Delauney and Verma 1993 Krishnamurthy and Bhagwat 1989) high osmolarity (Besford et al 1993) and oxidative stress (Langebartels et al 1991 Kurepa et al 1998) Under saline conditions free polyamine pools are reduced however it remains unclear which polyamine is more responsible in imparting salt tolerance in plants In a sole study Sannazzaro et al (2007) proposed that AMF modulate polyamine pool to impart salinity tolerance in Lotus glaber The beneficial effect of polyamines under salt stress has been attributed to their ability to interact with negatively charged macromolecules such as DNA and RNA proteins and phospholipids (Galston and Kaur Sawhney 1990 Gill and Tuteja 2010a) 1635 Antioxidants Generation of reactive oxygen species (ROS) constitute salt-induced secondary stress ldquooxidative stressrdquo in plants ROS include singlet oxygen (1O2) superoxide anion (O2-) hydrogen peroxide (H2O2) and hydroxyl radical (OH) Major sources of ROS in cells are chloroplast and mitochondria which house the electron transport system (Scandalios 1993 Asada 1999 Moller 2001) All ROS can react with DNA proteins and lipids (Fridovich 1986) and in the absence of the protective mechanism they can damage cell structure and function (Gill and Tuteja 2010b) ROS is an inevitable by-product of plant metabolism produced essentially from photosynthesis photorespiration and respiration However salt stress encourages the production of ROS in excess While at low concentrations ROS are required for signaling growth and behavior high concentrations of ROS is a threat to cells as it causes membrane lipid peroxidation
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
24
protein oxidation enzyme inhibition and DNA and RNA damage (Miller et al 2010 Singh et al 2011) Enhanced generation of ROS under salt stress is accomplished in four ways first plants responds to salt stress by decreasing stomatal conductance to avoid excessive water loss This in turn decreases the internal CO2 concentrations (Ci) and slows the reduction of CO2 by Calvin cycle This response leads to depletion of oxidized NADP+ which acts as the final acceptor of electrons in PSI and alternatingly increases the leakage of electrons to O2 forming O-
2 (Hsu and Kao 2003) Furthermore Na+ or Cl- toxicity resulting from salt stress could disrupt the photosynthetic electron transport and provoke electron leakage to O2 (Borsani et al 2001 Slesak et al 2002) Second the decrease in Ci slows down the reactions of Calvin cycle and induces photorespiration particularly in C3 plants resulting in generation of more H2O2 in the peroxisome (Wingler et al 2000 Ghannoum 2009) Third the cell membrane-bound NADPH oxidase and the apoplastic diamine oxidase get activated during salt stress and therefore contribute to generation of ROS (Hernandez et al 2001 Mazel et al 2004 Tsai et al 2005) Fourth salt stress increases the rate of respiration resulting in higher respiratory electron leakage to O2 (Fry et al 1986 Moser et al 1991 Jeanjean et al 1993)
To protect from oxidative damage plants have antioxidant defense system comprising of non-enzymatic antioxidant molecules such as ascorbic acid glutathione carotenoids tocopherols and enzymes such as superoxide dismutase (SOD) peroxidase (POX) catalase (CAT) ascorbate peroxidase (APX) glutathione reductase (GR) (Jiang and Zhang 2002 Nuacutenez et al 2003 Yamane et al 2004) A correlation between antioxidant capacity and NaCl tolerance has been demonstrated in several plant species (Gossett et al 1994 Benavides et al 2000 Nuacutenez et al 2003) Plants with high concentrations of antioxidants have been reported to have greater resistance to these antioxidative damages (Spychalla and Desbough 1990 Dionisio-Sese and Tobita 1998 Jiang and Zhang 2002 Gill and Tuteja 2010b)
AM-mediated salt tolerance by host plants has also been attributed to its ability to detoxify reactive oxygen species (ROS) and maintain the delicate balance between ROS and antioxidants (Wu et al 2010a b Hajiboland et al 2010 Borde et al 2011 Singh et al 2011) Mycorrhizal plants had lower concentration of malondialdehyde while
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
25
maintaining higher activity of SOD CAT APX POX and GR compared to NM plants (Wu et al 2010 Latef and Chaoxing 2011) The increased SOD will help detoxify O2
- to H2O2 (Smirnoff 1993) This H2O2 generated are scavenged by CAT APX and POX (Benavides et al 2000 Lopez et al 1996) The elevated levels of GR activity may serve to ensure the availability of NADP+ to accept electrons derived from photosynthetic electron transport thereby directing electrons away from oxygen and minimizing the production of O2
- (Gamble and Burke 1984 Menconi et al 1995) This AM-mediated rapid removal of excess ROS helps in maintaining the optimum concentration of ROS to perform its physiological role at the same time preventing the shift towards destructive mode
A detailed analysis of the influence of AM symbiosis on antioxidant system of plants under salt stress revealed that this aspect of AMF is in nascent stage and sporadic with certain gaps ndash (i) despite root being the organ that harbors the mycorrhiza and in direct contact with saline soil antioxidant enzymes have rarely been studied in roots (ii) although antioxidant enzyme activities are known to vary with severity and duration of stress (Abodagallah 2010) most of the studies involve a single sampling time (iii) so far the influence of AM symbiosis on the accumulation of non-enzymatic antioxidants such as ascorbic acid α-tocopherols glutathione and carotenoids in host plants has been less studied (Ruiz-Lozano et al 2012) and (iv) antioxidant enzyme activities are rarely correlated with ROS measurements (Ruiz-Lozano et al 2012) Furthermore enzyme activity depended largely on the plant organ (Rios-Gonzalez et al 2002) Therefore in order to obtain a clear picture of the influence of AM symbiosis during salt stress on antioxidant system of plants studies should be directed towards evaluation of the influence of AM symbiosis on (i) activities of non-specific ROS scavenging enzymes (SOD CAT POX APX GR) and accumulation of non-enzymatic antioxidants (ascorbic acid glutathione carotenoids α-tocopherols) in both rots as well as leaves at different time intervals and (ii) to correlate changes in antioxidative capacity with levels of H2O2 and oxidative damage in the same tissue as ROS accumulation depends largely on ROS production and scavenging (Miller et al 2010)
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
26
164 Physiological changes Salt stress has a negative effect on the physiology of the plant It disturbs photosynthetic efficiency gas exchange membrane disruption and water status Evidences are accumulating that AM symbiosis can alleviate such effects by employing various mechanisms which are discussed below 1641 Photosynthetic efficiency In nature chlorophyll is the central molecule differentiating the heterotrophs and autotrophs the producer In the presence of salt in the soil solution there is disruption in the structure and concentration of chlorophyll molecule resulting in reduced photosynthetic efficiency in plants (Sheng et al 2008) The decrease in chlorophyll molecule can be attributed to ndash (i) suppression of specific enzymes that are responsible for the synthesis of photosynthetic pigments (Murkute et al 2006) (ii) decrease in the uptake of minerals (eg Mg) needed for the chlorophyll biosynthesis (El-Desouky and Atawia 1998) and (iii) destruction of PSII reaction center and disrupt electron transport in photosynthetic apparatus of the plants (Sheng et al 2008) On the positive side these salt-induced toxic effects on photosynthetic machinery of the plant may be restricted or prevented by inoculating plant with AMF In this regard many authors have demonstrated that M plants had higher chlorophyll concentration and non-photochemical quenching capacity as compared to their NM equivalents (Sheng et al 2008 Zuccarini and Okurowska 2008 Hajiboland et al 2010 Kumar et al 2011) AM-facilitated improved uptake of nutrients (higher K+ Ca2+ and Mg2+ ions) helps in avoiding the specific effects of salt stress on chlorophyll degradation and leaf senescence Furthermore the ability of M plants to regulate the energy bifurcation between photochemical and non-photochemical events also helps in maintaining photosynthesis (Sheng et al 2008) 1642 Membrane integrity Electrolyte leakage from the cytoplasm is a measure of membrane damage in plants In plants exposed to salinity electrolyte leakage is accelerated by the ROS generated ROS can attack the lipid molecules in the membranes thereby rendering the membranes
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
27
permeable for electrolytes to leach out It is now becoming clearer that the presence of AMF in roots of plants help in maintaining the integrity and stability of plasma membrane under saline stress Under NaCl stress lower lipid peroxidation and concurrent lower electrolyte leakage were reported in mycorrhizal maize pigeon pea and fenugreek as compared to their NM counterparts (Feng et al 2002 Garg and Manchanda 2009) It is suggested that AM-conferred resistance to peroxidation of membrane lipids and hence leakage of electrolytes from the cell mediated through improved nutrition especially P and maintenance of higher Ca2+Na+ are the key factors contributing to this beneficial effect of AM-colonization on membrane integrity Other mechanisms include maintenance of higher antioxidant capacity and containment of ROS concentration in M plants as against NM plants 1643 Abscissic acid (ABA) concentration Plant growth and response to a stress condition is largely under the control of hormones (Mahajan and Tuteja 2005) Under salt stress the increase in transpiration results in increase in pH of leaf and accumulation of ABA Abscissic acid in turn promotes the efflux of K+ ions from the guard cells resulting in loss of turgor leading to stomatal closure On the other hand the analogous findings that AMF colonization can alter the ABA levels in host plant sponsor for an AM-induced regulatory mechanism for ABA accumulation (Duan et al 1996 Ludwig-Muller 2000 Estrada-Luna and Davies 2003) AM-mediated higher ABA levels regulate free polyamine pools in the plant (Sannazzaro et al 2007) However in contrast to this report Jahromi et al (2008) reported lower ABA levels in Glomus intraradices colonized lettuce plants than the NM plants indicating that M plants were less strained than NM plants by salinity stress imposed hence they accumulated less ABA 1644 Water status Arbuscular mycorrhizal colonization and the effective development of external mycelium is an important means for uptake of water in plants grown in saline soils Under salt stress AMF colonization have shown to provide better water status by maintaining higher relative water content over the NM plants (Aroca et al 2006 Colla et al 2008 Jahromi et
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
28
al 2008 Sheng et al 2008) Improved water status due to AMF colonization may also be attributed to its role in ensuring liquid continuity high hydraulic conductivity of roots and hence water uptake (Smith et al 2010) Since hydraulic conductivity is dependent on P-concentration (Carvajal et al 1996) it is likely that water uptake would be more strongly expressed in P-sufficient M plant than in P-deficient NM plants Mycorrhizal plants are also shown to accumulate solutes and maintain the osmotic balance Lower water saturation deficit and higher turgor potential in M plants also improves the water status of the plant (Al-Garni 2006 Sheng et al 2008)
Many studies have shown better water status in M plants than their corresponding NM plants however it is still unclear of how water from the AMF is translocated to the plant system The possibility of direct water transfer to plants via fungal hyphae have been put forward however the idea remains controversial to be established (Smith et al 2010) This gap in our knowledge is surprising given the importance of AM-mediated uptake of water in plants under salt stress However a positive step towards bringing down this gap is the recent discovery of an aquaporin gene GintAQP1 from Glomus intraradices (Aroca et al 2009) the expression of GintAQP1 is a compensatory alternative to plant aquaporins (Aroca et al 2009) Aquaporins belong to the major intrinsic protein (MIP) family of transmembrane channels which permit the selective membrane passage of water (and other few compounds) but not of H+ and other ions (Chen et al 2001 Hill et al 2004) through the plasma lemma (by plasma lemma intrinsic proteins (PIPs)) and the tonoplast (by tonoplast intrinsic protein (TIPs)) Though a few studies have shown AMF influence on plant aquaporins (Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008) these findings are not persuasive enough to establish and characterize the role of aquaporins and point to the possibility that AMF differentially exert control on each PIP (plasma membrane intrinsic protein) gene and each PIP gene analyzed may have a different function and regulation in AM-mediated alleviation of water stress 165 Molecular changes The plant function is ultimately explained by operation of genes in cells and tissues to regulate its growth with a symbiont in coordination with the environmental stress Notable absence of knowledge regarding the molecular basis for AM-induced tolerance
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
29
to salt stress in plants indicates that study aimed at molecular levels is at nascent stage Only a handful of reports in molecular studies (Harrison and van Burren 1995 Gonzalez-Guerrero et al 2005 Ouziad et al 2006 Aroca et al 2007 Jahromi et al 2008 Aroca et al 2009 Guether et al 2009) are a testimony to the declaration Though molecular studies are racing at a fast pace in other plant biology microbiology studies the AM-plant molecular studies are often impeded due to the obligate and heterokaryotic nature of AMF So far the scientists have concerted their efforts in unraveling the mechanisms for nutrient and water uptake in plants In this regard expression analyses of plant aquaporins have been studied in tomato (Ouziad et al 2006) Phaseolus vulgaris (Aroca et al 2007) and lettuce (Jahromi et al 2008) (as described in the section on improved water status) Recently an aquaporins gene (GintAQP1) from Glomus intraradices was reported (Aroca et al 2009) This is a step nearer to elucidation of water uptake mechanism in AM symbioses however it deserves more research to reach a conclusion
Molecular studies on AM-plant mineral uptake have revealed a phosphate transporter (Harrison and van Burren 1995) an ammonium transporter (Lopez-Pedrosa et al 2006 Guether et al 2009) a putative zinc transporter (Gonzalez-Guerrero et al 2005) in AM fungus The role of AMF in nutrient uptake at the molecular level is only beginning to be understood
Expression studies have been conducted for Na+H+ antiporters ndash LeNHX1 and LeNHX2 in tomato (Ouziad et al 2006) The authors reported that salt and mycorrhizal colonization had no significant effects on these two antiporters
In another study Jahromi et al (2008) analyzed the expression pattern of genes encoding ∆1-pyrroline-5-carboxylate synthetase (LsP5CS) late embryogenesis abundant protein (LsLea) and abscissic acid (Lsnced) in mycorrhizal and non-mycorrhizal lettuce plants subjected to varied salt treatments (0 ndash 100 mM NaCl) The PC5S enzyme catalyzes the rate limiting step in the biosynthesis of proline (Kishor et al 1995) an osmoregulator in plants Late embryogenesis abundant protein acts as stress marker They also possess chaperone like activity thus having a protective role during osmotic stress Lsnced encodes for 9-cis-epoxycarotenoid dioxygenase a key enzyme for the biosynthesis of stress hormone abscissic acid (ABA) Abscissic acid
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
30
promotes stomatal closure to minimize transpirational water loss It also mitigates stress damage through the activation of many stress-responsive genes which collectively increase plant stress tolerance (Bray 2002) The authors reported a higher expression of genes LsP5CS and Lsnced in NM plants than M plants at 50 mM NaCl though at 100 mM the levels were similar LsLea gene was found to express under conditions of salt stress and the induction of this gene was found to be lower in M plants than NM plants The lower expression of this gene suggests that M plants suffer less stress than non-AM plants which may be likely due to primary salt avoidance mechanism 166 Ultrastructural changes Presence of salts in the soil has been reported to affect the ultrastructure of cells (Pareek et al 1997 Yamane et al 2004 Miyake et al 2006 Mahmoodzadeh 2008 Andrea and Tani 2009) At high soil salinity most of the structural components of plants are damaged leading to loss of function and subsequently death of the plants Salt stress caused an increase in membrane surface and quantity of vesicles in root cells of Sorghum (Koyro 1997) Salt stress caused thickened cell wall increased frequency of plasmodesmata and vacuolization of cytoplasm in shoot apical meristem of canola (Mahmoodzadeh 2008) Chloroplast showed disruptions in thylakoid and thylakoid membrane (Yamane et al 2004) As AM symbioses have been shown to impart plant tolerance against salt stress it is very tempting to speculate that it might prevent salt-induced ultrastructural damage in plants However there are no studies on the role of AMF in mitigating salt stress in host plant at the ultrastructure of the cell Therefore our attention in host-AMF relationship under salt stress needs be directed towards unveiling the mechanism of salt stress alleviation by AMF 17 Objectives
1 To estimate the degree of damage and extent of tolerance in mycorrhizal and non-mycorrhizal fenugreek plants subjected to varying degrees of NaCl stress
2 To investigate the effects of different levels of salinity on nutrient acquisition and evaluate the role of AMF in mitigation of NaCl-induced ionic imbalance in plants
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity
Introduction
31
3 To study the influence of varying levels of salt stress on accumulation of endogenous osmolytes and alterations in cell ultrastructure and further to evaluate the contributions of AMF in maintaining osmotic homeostasis and prevention of ultrastructural damage in plants
4 To evaluate the influence of AMF on antioxidant system in fenugreek plants subjected to varying levels of salinity