1
PHOTOPHYSIOLOGICAL PROPERTIES OF THE MARINE PICOEUKARYOTE
PICOCHLORUM RCC 237 (TREBOUXIOPHYCEAE, CHLOROPHYTA)1
Céline Dimier, Federico Corato, Giovanni Saviello and Christophe Brunet2
Stazione Zoologica "Anton Dohrn", Villa Comunale, 80121 Naples, Italy
Running title: Photophysiology of Picochlorum
1Received Accepted
2Author for correspondence: email [email protected]
2
Key index words: Fluorescence quantum yield; Non-Photochemical Quenching of fluorescence;
Photoacclimation; Picoplankton; Xanthophyll Cycle; Zeaxanthin.
Abbreviations: Ax, antheraxanthin; chl a, chlorophyll a; DPS, de-epoxidation state
[=(Ax+Zx)/(Vx+Ax+Zx)]; DTT, Dithiotreitol; ETR, electron transport rate; FALS, Forward
Angle Light Scatter; Fq'/Fm', operating efficiency of PSII; Fv/Fm, maximum photochemical
efficiency of PSII; HL, High Light; LHC, Light-Harvesting Complex; LL, Low Light; ML,
Moderate Light; NF, Norflurazon; NPQ, Non-photochemical quenching; PAM, Pulse Amplitude
Modulation; PFD, Photon flux density; PSII, Photosystem II; RALS, Right Angle Light Scatter;
RLC, Rapid light curve; Vx, violaxanthin; Zx, zeaxanthin.
3
The photophysiological properties of strain RCC 237 belonging to the marine
picoplanktonic Picochlorum (Henley et al.) genus were investigated under different photon flux
densities (PFD, ranging from 40 to 400 µmol photons·m-2·s-1), mainly focusing on the
development of the xanthophyll cycle and its relationship with the non-photochemical quenching
of fluorescence (NPQ). The xanthophyll cycle functioning and its photoprotective role was
investigated by applying a progressive increase of PFD and using Dithiotreitol and Norflurazon
to block specific enzymatic reactions in order to deeply study the relationship between
xanthophyll cycle and NPQ. These two processes were significantly related only during the
gradually increasing light periods and not during stable light periods, where NPQ and zeaxanthin
were decoupled. This result reveals that NPQ is a photoprotective process developed by algae
only when cells are experiencing increasing PFD or in response to stressful light variations, for
instance after a sudden light shift. Results showed that the photobiological properties of
Picochlorum strain RCC 237 seem to be well related to the surface water characteristics, since it
is able to maintain its photosynthetic characteristics under different PFDs and to quickly activate
the xanthophyll cycle under high light.
4
INTRODUCTION
In natural environments, phytoplankton cells continuously experience light variations due
to the diel cycle and the vertical displacement of cells within the mixed layer due to
hydrodynamics. Since light is crucial for survival and success of algae in aquatic ecosystems,
they have to continuously photoacclimate, through biochemical, physiological and/or
morphological variations. Photophysiological properties exhibited by algae seemed to be related
to the environment where they grow (Stomp et al. 2004, Strzepek and Harrison 2004),
converging toward the hypothesis that light is a relevant factor driving competition, exclusion
and/or diversity in the algal community (Litchman and Klausmeir 2001, Floder and Burns 2005).
An important trait of phytoplankton functional diversity is cell size that is in part responsible for
the variety of biological and/or ecological behaviors with respect to the environment, e.g. sinking
rate, nutrient consumption, light utilization and packaging effect (Fogg 1991, Raven 1998).
Picoplankton (size < 3 µm) are known to be important contributors to autotroph communities in
many coastal and oceanic ecosystems (e.g. Raven 1998, Not et al. 2005). The high level of
taxonomic biodiversity in the picoeukaryotic fraction (Moon-van der Staay et al. 2001, Diez et
al. 2004) reinforces the key role of this group in marine ecosystem structure and functioning.
Nevertheless, very few data exist on their photophysiology (e.g. Wilhem et al. 1989;
Timmermans et al. 2005), in contrast to prokaryotes (e.g. Bibby et al. 2003 and references
therein, Stomp et al. 2004) and to the larger cell size eukaryotic community (e.g. Casper- Lindley
and Bjorkman 1998, Lavaud et al. 2004).
An important photoprotective mechanism active at short temporal scales is the dissipation of
excess energy, through the non-photochemical quenching of chl a fluorescence that is catalyzed
5
by the xanthophyll cycle (Finazzi et al. 2006). This process involves enzymatic conversion from
violaxanthin to antheraxanthin and zeaxanthin in plants and green algae (Gilmore et al. 1994,
Masojidek et al. 1999) and from diadinoxanthin to diatoxanthin in chromophyte alge (Lavaud et
al. 2002). Accumulation of zeaxanthin or diatoxanthin is triggered by the formation of a pH
gradient across the thylakoid membrane (Demmig-Adams and Adams 2000). Much attention has
been paid to the photoprotection mechanism in chromophyte algae (e.g Lavaud et al. 2004 and
references therein, Harris et al. 2005), while few studies have dealt with marine green micro-
algae (e.g. Gilmore and Yamamoto 2001, Garcia-Mendoza et al. 2002). The presumed minor
contribution of green algae into the phytoplankton community, as well as the similarity of their
xanthophyll cycle to the well-documented one found in terrestrial plants, (e.g. Havaux and
Niyogi 1999, Demmig-Adams and Adams 2000) make the study of photoregulation in this group
of low ecological interest. Recently, it has been shown that these algae are among the main
picoeukaryote contributors in many ecosystems (Diez et al. 2004, Not et al. 2005), highlighting
their capacity to adapt to different ecosystem characteristics and their probably relevant role in
ecosystem structure and functioning.
This study investigates the photophysiological properties of the strain RCC 237
belonging to the picoeukaryotic genus Picochlorum (Chlorophyta, Trebouxiophyceae). The
short-term photoacclimative process is dissected in order to test the presence of an efficient
acclimation to high photon flux density, through rapid xanthophyll cycle activation. Our
assumption is based on the biological peculiarities of picoeukaryotes, e.g. low sinking rate and
packaging effect (Raven 1998), that may lead this group to be more sensitive to light than large
cells. This could be the case of the strain RCC 237 that was isolated from Mediterranean surface
waters. Pigments, quenching of fluorescence and PSII efficiency were used as photoacclimative
6
indices during different light treatments and in presence or absence of enzymatic inhibitors
linked to xanthophyll cycle activity.
MATERIALS AND METHODS
Algal model and culture conditions. The strain RCC 237 of Picochlorum (Chlorophyta,
Trebouxiophyceae), isolated by Dr. D. Vaulot from the surface layer (20 m depth) of the
Mediterranean Sea, was provided by the Roscoff Culture Collection (France; Vaulot et al. 2004).
This small non-flagellated coccoid strain (diameter 1.5 µm) was cultivated non-axenically at 20°
C under 40 µmol photons·m-2·s-1 (measured by a 4 π QSL-2100, Biospherical instruments INC,
San Diego, USA) provided by one lamp (OSRAM, Decostar 51, 50 W; Munich, Germany) with
a 12:12 light:dark photoperiod. Algae were grown in Keller medium (Keller et al. 1987) in 3-L
glass cylinders and the cultures were continuously aerated and maintained in exponential phase
by daily dilution of half of the culture with fresh medium during more than 10 days before the
experiments. The pH and temperature were estimated daily with a HI- 9214-Stick pHmeter
(Hanna Instruments, Woonsocket, USA) while cell concentration was controlled by flow
cytometry (see below).
Experimental design and sampling. The four experiments were conducted in triplicate
aerated cultures at 20° C. Each culture flask was illuminated by one lamp (OSRAM, Decostar
51, 50 W; Munich, Germany), and the three lamps were mounted on a prototype device, called
“PLIS” (“Progressive Light Increase System”) allowing gradual changes of PFD on the culture
flasks, controlled by a BASIC program. A 3-L glass culture flask was maintained under the
7
initial conditions as an experimental control. During the four experiments, 30 mL of culture was
sampled (see below) for HPLC-analyzed pigments, absorption spectrum on a filter by a
spectrophotometer, cellular parameters by flow cytometry, quantum yield of fluorescence and
rapid light curves (RLCs) by Phyto-Pam.
Low light to high light experiment. The “PLIS” was setup as follows: after 30 minutes at
40 µmol photons·m-2·s-1, light was gradually increased to 200 µmol photons·m-2·s-1, over 60 min.
For the following three hours, light remained constant at 200 µmol photons·m-2·s-1 and then
increased to 400 µmol photons·m-2·s-1 over 30 min, remaining constant at this high light value for
2 hours. Frequency of sampling was high, especially during the two increasing light periods,
with a total of 23 sampling points (Fig. 1a)
High light to low light experiment. This experiment was conducted on cells acclimated at
400 µmol photons·m-2·s-1 during 6 hours (i.e. the duration of the previous experiment) in order to
study the relaxation of the photosynthetic apparatus under low light (40 µmol photons·m-2·s-1)
during 3 hours.
DTT addition experiment. After one hour of illumination at 40 µmol photons·m-2·s-1, the
DTT was injected in aqueous solution into the culture to a final concentration of 500 µmol·L-1
(Lohr and Wilhelm 2001). Cells were incubated for 10 minutes with the inhibitor before the shift
to high light (400 µmol photons·m-2·s-1). The control culture with no-addition of DTT followed
the same light change as the three other flasks.
8
NF addition experiment. A methanolic solution of NF was injected into the culture flask
to a final concentration of 10 µmol·L-1 methanol (2.5% v/v methanol; Garcia-Plazaola et al.
2002). Cells were incubated for 10 minutes with the inhibitor before being shifted to 400 µmol
photons·m-2·s-1. It was previously verified that methanol (without NF) at this concentration has
no effect on the algal photophysiology (data not shown).
Pigment analysis. Samples of 10 mL were filtered onto GF/F glass-fiber filters
(Whatman, Maidstone, UK) and immediately stored in liquid nitrogen until analysis. Using the
method outlined in Casotti et al. (2005), the pigment solution extract was injected in a Hewlett
Packard series 1100 HPLC (Hewlett-Packard, Kennett Square, PA, USA) with a C8 BDS 3 µm
Hypersil, IP column (Thermo Electron corporation, USA). The mobile phase was composed of
two solvents: A, methanol, aqueous ammonium acetate (70:30) and B, methanol. Pigments were
detected spectrophotometrically at 440 nm using a Hewlett Packard photodiode array detector
model DAD series 1100 while fluorescent pigments were detected using a Hewlett Packard
standard FLD cell series 1100 with excitation and emission wavelengths setup at 407 nm and
665 nm, respectively. Determination and quantification of pigments were realized using pigment
standards from the D.H.I. Water & Environment (Hørsholm, Denmark).
Absorption spectrum. Absorption was measured on 10 ml culture samples filtered onto
GF/F filters (Whatman, 25 mm), following the procedure of Tassan and Ferrari (1995), between
400 and 800 nm with 1-nm increment on a spectrophotometer (Hewlett Packard HP-8453E)
equipped with an integrating sphere (Labsphere RSA-HP-53).
9
Flow cytometry analysis. Duplicate 1 ml samples were fixed in 10% paraformaldehyde
and immediately stored in liquid nitrogen. Analysis was done with a FACScalibur (Becton
Dickinson, USA) using 6 µm beads (Flow set, Coulter) as internal standard. The method and
apparatus was the same as in Casotti et al. (2005).
Fluorescence measurement. Photochemical efficiency of PSII was estimated with a
Phyto-PAM fluorometer (Heinz Walz GmbH, Effeltrich, Germany). Maximum photochemical
efficiency (Fv/Fm, with Fv = Fm-Fo) was measured on 15 min dark-adapted sample while the
operating photochemical efficiency (Fq’/Fm’, with Fq’ = Fm’- F’) was measured on actinic light-
exposed sample (Harris et al. 2005). The effective non-photochemical quenching of fluorescence
was quantified by the Stern-Volmer expression:
effNPQ = (Fm/Fm’) - 1 (1)
where, Fm and Fm’ are the maximum fluorescence values from dark- and actinic light-exposed
samples, respectively. Fm and Fm’ were measured after a saturation pulse of bright red light
(intensity of 2400 µmol photons·m-2·s-1) applied during 450 ms, causing a complete reduction of
the PSII acceptor pool.
The RLCs (rapid light curves) were determined at 5 sampling points (0, 70, 210, 310 and
390 min) on 15 min dark-adapted samples by applying ten increasing actinic irradiances (from 8
to 1500 µmol photons·m-2·s-1, two minutes at each PFD level). The absolute electron transport
rate was calculated as:
10
absETR = (Fq’/Fm’) x PFD x (a*ph/2) (2)
where, PFD was the incident irradiance (µmol photons·m-2·s-1) and (a*ph/2) the mean absorption
value of phytoplankton normalized by chl a (m2·mg chl a-1) and divided by 2 under the
assumption that half of the absorbed light is distributed to PSII. The RLCs were fitted with the
equation of Eilers and Peeters (1988) to estimate the photosynthetic parameters αetr, Ek and
absETRmax.
In the RLC, operating photochemical efficiency decreases as NPQ develops (Villareal
2004). This NPQ capacity was calculated after each actinic irradiance level according to equation
(1), where Fm was the maximum fluorescence measured after the 15 min dark-adaptation and
Fm’ was the maximum fluorescence measured after each actinic light level. The maximum NPQ
(maxNPQ) was determined as the highest value reached on the NPQ coefficient vs PFD curve.
RESULTS
The low light acclimated cells were characterized by a dominance of violaxanthin (Vx,
72%) among the three-xanthophyll cycle pigments (violaxanthin-antheraxanthin-zeaxanthin, Vx-
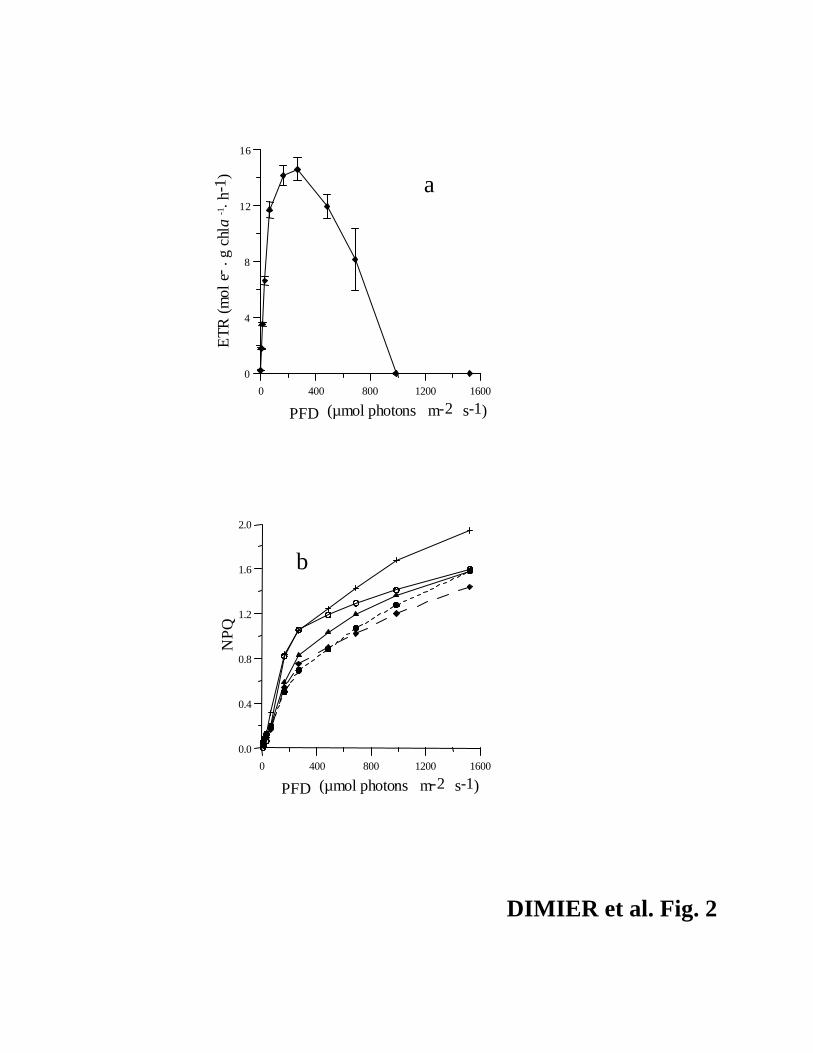
Ax-Zx, Fig. 1b) and a high value of maximum photochemical efficiency (Fv/Fm = 0.69). These
values as well as the sustained growth rate (0.39 d-1) and the high values of absETRmax and
photosynthetic efficiency (0.85 mol e-·g chl a-1·h-1 and 0.013 mol e-·g chl a-1·h-1·(µmol photons·m-
2·s-1)-1, respectively) revealed a good acclimative state to low light. The strong decrease of
Fq’/Fm’ for PFDs > 250 µmol photons·m-2·s-1 along the RLC (Fig. 2a) showed a high capacity
for energy dissipation, consistent with the NPQ capacity evolution and its maximum value (1.94,
Fig. 2b).
11
Gradual increasing light experiment. The de-epoxidation of Vx into Ax and Zx mainly
occurred during the two increasing light periods (Fig. 1, b and c) and was negatively correlated
with the maximum photochemical efficiency (p<0.001), which slightly decreased with time (-
8.5%, data not shown). Activation of the Vx de-epoxidase already occurred at low light since the
synthesis of Ax was initiated after 12 min (i.e. 72 µmol photons·m-2·s-1, Fig. 1b). Significant
linear relationships were found between Zx/chl a and effNPQ only during the two increasing light
periods:
effNPQ = 6.0 (Zx/chl a) – 0.31 (r2 = 0.75, n=9) (3)
for the 40 to 200 µmol photons·m-2·s-1 light shift, and
effNPQ = 0.67 (Zx/chl a) – 0.07 (r2 = 0.85, n=9) (4)
for the 200 to 400 µmol photons·m-2·s-1 light shift.
The relationships (3) and (4) revealed that not all the intracellular zeaxanthin was linked
to energy dissipation. Indeed, effNPQ reached approximately the same value at the end of the two
increasing light periods (around 0.10, Fig. 1c) despite a Zx/chl a two fold higher under 400 with
respect to 200 µmol photons·m-2·s-1 (0.025 vs 0.012 mol Zx·mol chl a-1, respectively; Fig. 1b).
During the two stable light periods, the effNPQ decreased while Ax/chl a and Zx/chl a continued
to increase and Vx/chl a reached a plateau, showing a decoupling with the xanthophyll cycle
12
(Fig. 1, b and c). The effNPQ decreased faster at high light than under moderate light (< 30 min
and 2 hours after light stability, respectively). On the contrary, no difference was revealed in the
NPQ capacity evolution along the RLC for these two sampling points (Fig. 2b). The higher
zeaxanthin content under moderate light with respect to low light allows a faster photoprotective
response when cells experience increasing values of PFD, with little need to dissipate excess
energy through NPQ. This increase in zeaxanthin in relation to the difference in PFD (200 or 400
µmol photons·m-2·s-1) was probably related to the different conformation of the light harvesting
complexes (Pascal et al. 2005). The amount of other accessory pigments, such as chl b/chl a and
ß-carotene/chl a, remained constant during the time course of the experiment, whereas lutein/chl
a increased by 20% (data not shown). Photoregulation process developed by cells during this
experiment seemed to be efficient since the photosynthetic characteristics (e.g. Fig. 2a) did not
vary with increasing PFD. Also, the effNPQ remained low and the capacity to develop NPQ did
not significantly change with time (Fig. 2b). Algal growth was enhanced under high light, as
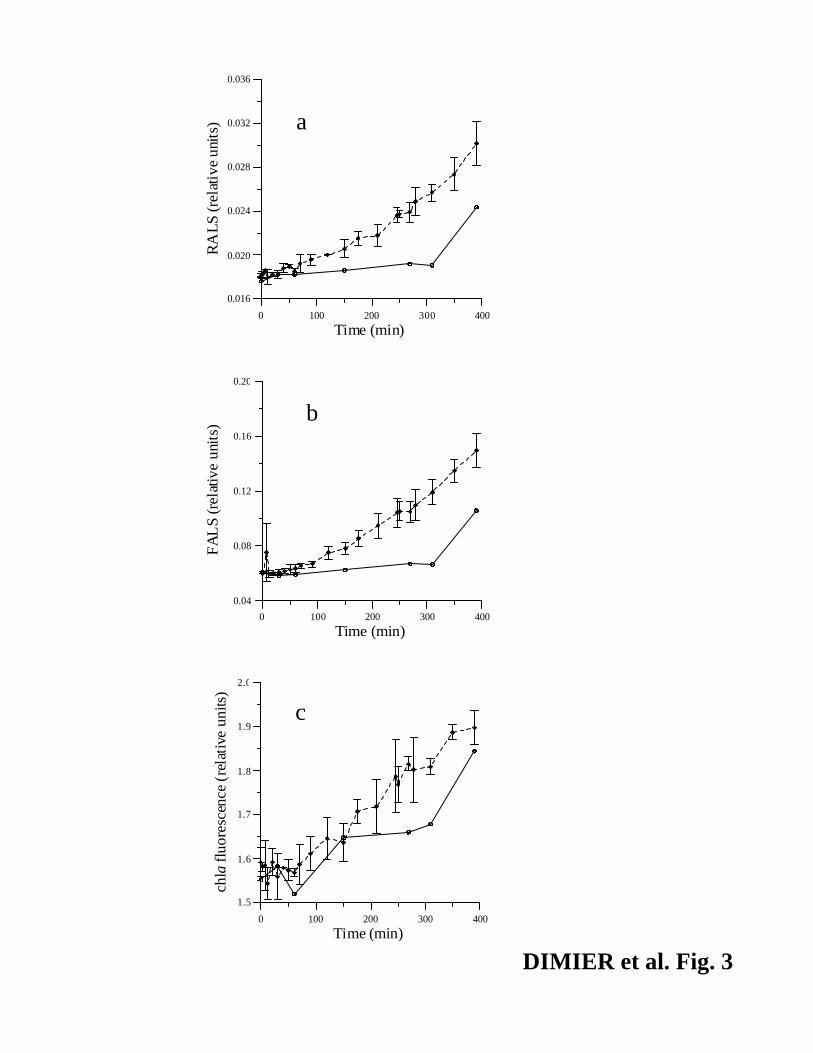
revealed by the stronger increase of FALS and RALS (Fig. 3, a and b; +150.4 % and +67%,
respectively) with respect to low light (+74% and +38.8%, respectively). Increase of red chl a
fluorescence and similar mean of chl a·cell-1 in both the light conditions (Fig. 3c, p > 0.05, mean
± SD = 0.052 ± 0.005 fmole chl a·cell-1) revealed that cells were not negatively affected by high
light.
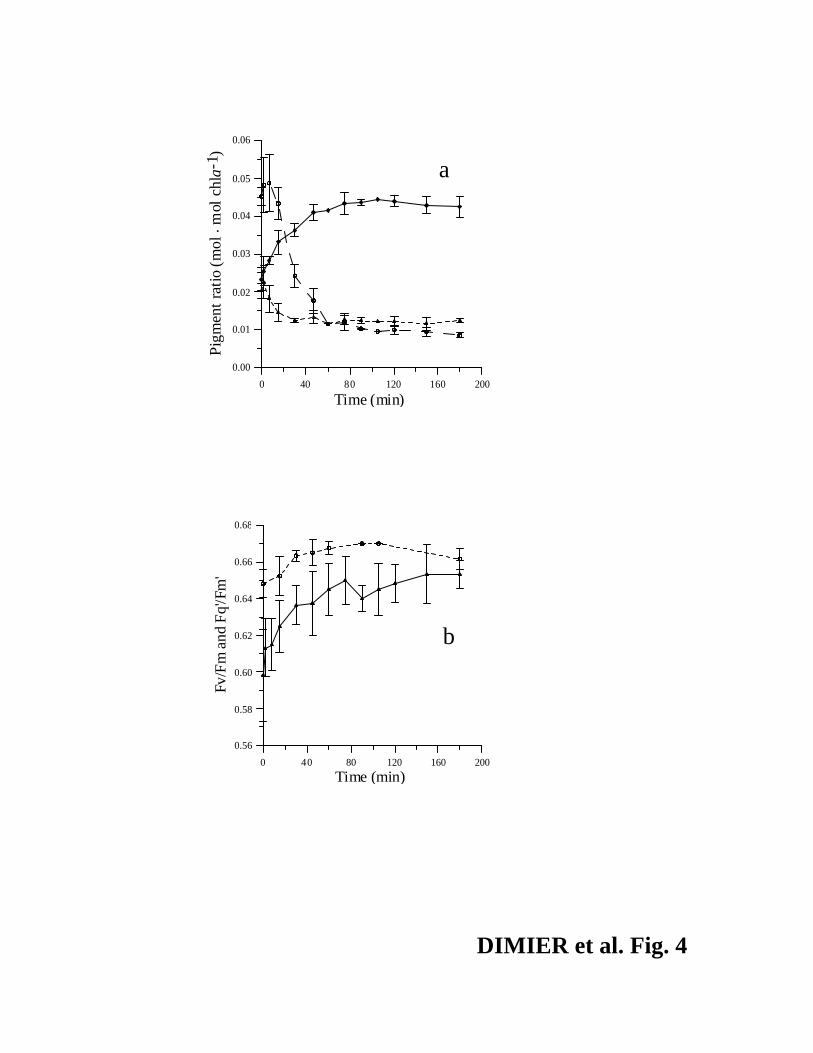
High light to low light experiment. The xanthophyll pool of the high-light-acclimated
cells (6 hours at 400 µmol photons·m-2·s-1) was dominated by Ax (51% vs 26.1% (Vx) and
22.7% (Zx), initial values in Fig. 4a). As at the end of the previous experiment (PFD = 400 µmol
photons·m-2·s-1), the effNPQ was very low (0.02) and the maximum photochemical efficiency was
13
high (Fv/Fm = 0.65). Under LL, the Zx/chl a started to decrease after 2 min and Vx/chl a
strongly increased (Fig. 4a). In less than two hours, the xanthophyll pigments reached a plateau
(Fig. 4a), with Vx, Ax and Zx contributing for 64%, 18% and 18%, respectively, which was
similar to the values found for the low light acclimated cells. The PSII reaction centers quickly
opened, as suggested by the rapid increase of the Fq'/Fm', which almost reached the value of
Fv/Fm (Fig. 4b). The maxNPQ decreased during this experiment (from 2.36 to 1, data not
shown) in agreement with the lowering of the photoprotective pigment pool.
Use of dithiotreitol. Death or stress were not evident by flow cytometry (see Casotti et al.
2005; data not shown) during the experiment. The initial rise of effNPQ in the control and +DTT
cultures (Fig. 5a) was not related to Zx synthesis (p>0.05), revealing a rapid formation of a Zx-
independent NPQ due to the sudden light shift from 40 to 400 µmol photons·m-2·s-1. Thus the
effNPQ decreased only in +DTT, probably because Zx pool was not high enough to maintain the
thermal dissipation rate of excess of energy in the cells (Fig. 5, b and c). Indeed, the de-
epoxidation was inhibited and zeaxanthin decreased in +DTT after 30 minutes (Fig. 5, b and c),
in agreement with results from Gilmore et al. (1994) who showed that epoxidation can be
activated during light-induced lumen acidification after inhibition of the de-epoxidase with DTT.
This hypothesis would also explain the decrease of Ax as already described by Gilmore et al.
(1998) in higher plants.
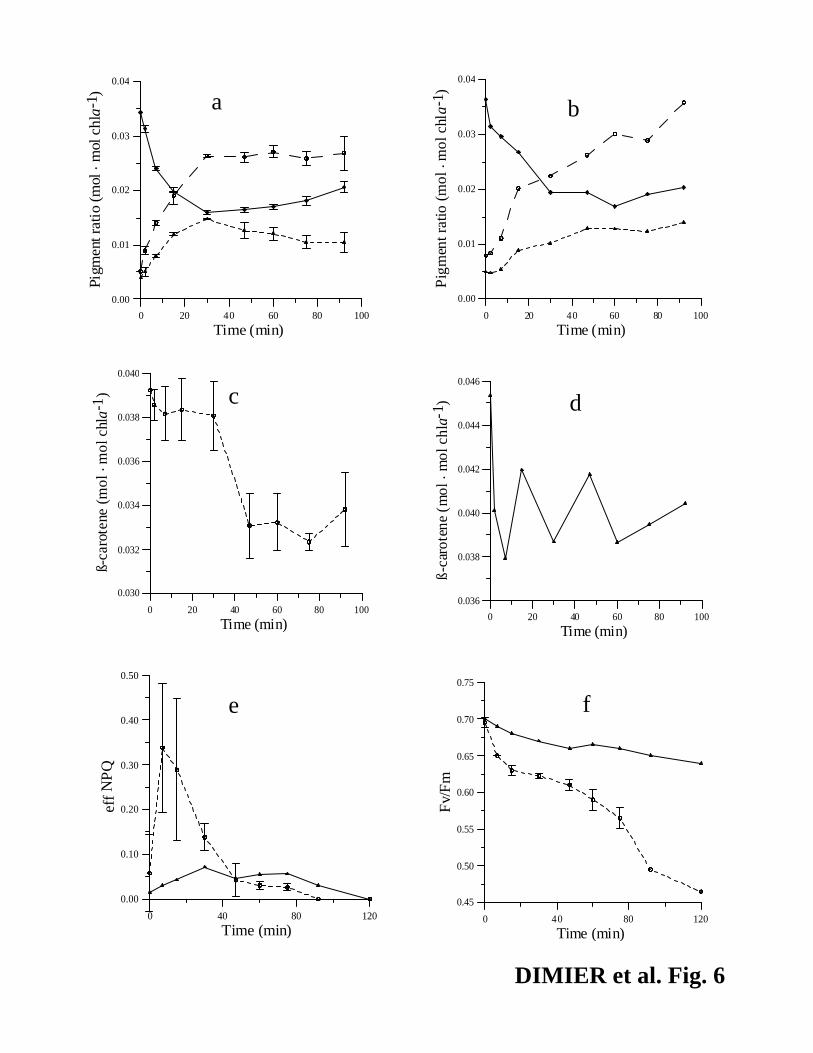
Use of norflurazon. The addition of NF did not obviously affect the amount of Vx, Ax
and Zx, at least in the first 60 min of the experiment, under high light compared to the control
(Fig. 6, a and b). Since NF inhibited the phytoene desaturase, cells apparently used the pre-
14
existing pool of ß-carotene to produce Vx, Ax and Zx. Indeed, the decrease of ß-carotene (-
0.007 mol·mol chl a-1) almost corresponded to the increase in VxAxZx pool (+0.010 mol·mol chl
a-1, Fig. 6, a and c). The maxNPQ estimated from the RLC decreased from 1.35 to 0.35 after 1
hour in +NF, whereas it remained constant in the control (data not shown), revealing a decrease
of the cell’s capacity for thermal energy dissipation. This suggests photoinhibition, consistent
with a ~30% reduction of Fv/Fm in the NF-treated culture with respect to the control (Fig. 6f), in
part due to an increase in Fo in +NF (+ 36%, data not shown). We have no explanation for the
peak of effNPQ in +NF cells 10 min after the shift to high light (Fig. 6e).
DISCUSSION
During this study the photoprotective processes developed at short temporal scale by
strain RCC 237 of Picochlorum was investigated. Little physiological information is available on
the picoeukaryotes (e.g. Sobrino et al. 2005, Timmermans et al. 2005, Veldhuis et al. 2005,
Finazzi et al. 2006), whereas their diversity in marine ecosystems is usually high (Diez et al.
2004, Not et al. 2005). Indeed, the suggested differentiation in ecotypes of some picoeukaryotes
(Rodriguez et al. 2005) makes it difficult to make any generalizations about the physiological
results obtained from one strain. Meanwhile, their biological peculiarities (e.g. low sinking rate,
small package effect, efficient resource acquisition and use for growth; Raven, 1998) make them
a relevant model for comparison with larger cells, such as diatoms. Whereas this strain cannot be
considered as a model for chl b-containing picoeukaryotes nor for the genus Picochlorum, results
obtained in this study provide information about the photoacclimation properties of a green
picoeukaryote growing in oceanic surface waters. Photo-physiological and photoprotective
15
properties differ among phytoplankton according to their phylogeny and ecology (Casper-
Lindley and Bjorkman 1998, Strzepek and Harrison 2004). The strain RCC 237 grows better
under 400 µmol photons·m-2·s-1 than under 40 µmol photons·m-2·s-1 (µ = 0.80 d-1 and 0.40 d-1,
respectively) and its photophysiological properties seem to be adapted to the high and variable
irradiance characteristics of oceanic surface water. This strain is able to photoacclimate to a
broad range of PFD by maintaining its photosynthetic characteristics, such as absETRmax (0.85
mol e-·g chl a-1·h-1), photosynthetic efficiency (αetr = 0.013 mol e-·g chl a-1·h-1·(µmol photons·m-
2·s-1)-1), and light saturation (Ek = 65 µmol photons·m-2·s-1). Assuming a transfer of four
electrons for 1 mol O2, a rate of 0.21 mol O2·g chl a-1·h-1 has been estimated from the absETRmax
value, indicating similar photosynthetic performance as for instance Emiliania huxleyi grown
under 50 µmol photons·m-2·s-1 (Harris et al. 2005). The fast recovery of PS II properties (< 1
hour, e.g. pigments or maximum photochemical efficiency) at low PFD reveals the high
plasticity of PSII for optimizing photochemistry, in agreement with the ability to sustain growth
under variable PFD.
Comparing the de-epoxidation state (DPS) values reached under high PFD, the strain RCC 237
seems able to activate the xanthophyll cycle more rapidly with respect to other green algae, such
as Dunaliella tertiolecta (Casper-Lindley and Bjorkman 1998). More generally, the DPS value
reached under different PFD is higher in phytoplankton (> 0.25 for a range of light from 8 to 40
µmol photons·m-2·s-1, Schubert et al. 1994; Havelkova-Dousova et al. 2004; this study) than in
terrestrial plants (<0.12 for a range of light from 50 to 220 µmol photons·m-2·s-1, Havaux et al.
2000, Garcia-Plazaola et al. 2002). This feature could be linked to the more variable PFD
experienced by micro-algae that would probably require faster and/or stronger photoprotection
responses as compared to terrestrial plants. This hypothesis agrees with the results of Lavaud et
16
al. (2002) and Garcia-Mendoza et al. (2002), who showed that the xanthophyll content increased
under intermittent high light in diatoms and chlorophytes. The progressive increase of PFD
seems to enhance the synthesis of photoprotective pigment (reaching 0.025 mol Zx·mol chl a-1,
Fig. 1b) with respect to a sudden light shift (reaching 0.015 mol Zx·mol chl a-1, Fig. 6b). The
increase and maintenance of the Zx pool under moderate or high PFD allows a quicker effNPQ
formation when cells experience a further increase of PFD. During stable light periods, the
decrease of effNPQ with a constant DPS agrees with the fact that only ∆pH and not de-
epoxidation state is relaxed after light increase (Ruban and Horton 1999). The quicker decrease
of effNPQ under constant 400 µmol photons·m-2·s-1 than under 200 µmol photons·m-2·s-1 may
relate to the higher content of Zx even though light is two fold higher. The ecological advantage
may be a faster photoregulation under changing PFD, with little necessity to dissipate excess
light. The overall lack of relationship between NPQ and zeaxanthin is due to the fact that not all
the zeaxantin participates to the quenching of fluorescence and to the fast development of NPQ
independently from zeaxanthin. NPQ is directly related to the synthesis of zeaxanthin only
during gradual increasing light periods. The effNPQ per mole of Zx is much higher during the
change from 40 to 200 µmol photons·m-2·s-1 than from 200 to 400 µmol photons·m-2·s-1 (see
relationships 4 and 5) and the zeaxanthin synthesis was higher during the second light shift than
during the first one (+ 115% and 40 %, respectively). Tardy and Havaux (1996) showed that a
specific subset of Zx molecules could participate to NPQ formation. Since the effNPQ reaches the
same value at the end of the two increasing light periods, the value of 0.012 mol Zx·mol chl a-1
(maximal value obtained after the first increasing light period) must correspond to the upper limit
of Zx contributing to NPQ in the strain RCC 237 of Picochlorum. The same quantity of Zx per
chl a must be devoted to another function under high PFD, such as a slow down of D1 protein
17
degradation rate (Jahns et al. 2000), or lipid peroxidation prevention (Havaux and Niyogi 1999).
The synthesis of zeaxanthin begins at the photosynthetic light saturation (Ek), i.e. around 65
µmol photons·m-2·s-1. On the contrary, a rapid quenching of fluorescence occurred without any
increase in zeaxanthin when cells experienced either sudden or gradual increase of PFD meaning
that is caused by an increasing light rather than excessive light per se. Indeed, the development
of effNPQ occurs at subsaturating PFD (significant increase at 50 µmol photons·m-2·s-1),
revealing that energy dissipation begins before photosynthetic light saturation.
Zeaxanthin-independent NPQ can develop rapidly in parallel with the protonation of the PsbS
protein in plants (Dall’Osto et al. 2005, Pascal et al. 2005). After formation, this NPQ is
maintained by binding of Zx on PsbS (Niyogi et al. 2004). This two-step functioning is
conserved in chl b-containing photoautotrophs from terrestrial plants (Niyogi et al. 2004) to
phytoplankton (e.g. Casper-Lindley and Bjorkman 1998, Masojidek et al. 1999, this study).
Nevertheless, the NPQ seems to be mainly zeaxanthin-dependent when this pigment is already
present in relatively high amount in the cell as revealed during the second light increase (from
200 to 400 µmol photons·m-2·s-1, Fig. 1, b and c behavior could present ecological advantage,
since the protein PsbS enables fast and reversible NPQ, depending on zeaxanthin removal from
the binding sites and not on its epoxidation (Horton and Ruban 2005). The presence of this
mechanism in green algae and terrestrial plants could explain the lower pH needed for the
activation of the de-epoxidase enzyme (= 6.5, Emanuelson et al. 2003) with respect to
chromophyte algae, such as diatoms (= 7.2, Jakob et al. 2001), in which the PsbS protein has not
yet been found (Horton and Ruban 2005). State transition, an additional photoprotective
mechanism responsible for the uncoupling between zeaxanthin and NPQ in some green algae
18
(Masojidek 2004), has still to be investigated in green marine picoeukaryotes such as the
Picochlorum genus.
Acknowledgements
The authors thank the Roscoff Culture Collection (Roscoff, France) for sending us the strain of
Picochlorum sp. and R. Casotti for her flow cytometry experience. We also thank S. McDonald
for the English revision of the manuscript. Two anonymous referees and D. Vaulot are gratefully
acknowledged for the numerous and helpful comments on the manuscript. C.D. is supported by a
Ph.D grant from SZN. This publication represents a contribution to the aims of the MarBef
Network of Excellence “Marine Biodiversity and Ecosystem Functioning” which is funded in the
Community’s Sixth Framework Programme (contract no. GOCE-CT-2003-505446). This
publication is contribution number MPS07002 of MarBef.
19
List of references
Bibby, T.S., Mary, I., Nield, J., Partensky, F. & Barber, J. 2003. Low-light adapted
Prochloroccus species possess specific antennae for each photosystem. Nature 424: 1051-1054.
Casotti, R., Mazza, S., Brunet, C., Vantrepotte, V., Ianora, A. & Miralto, A. 2005. Growth
inhibition and toxicity of the diatom aldehyde2-trans, 4-trans-decadienal on Thalassiosira
weissflogii (Bacillariophyceae). J. Phycol. 41: 7-20.
Casper-Lindley, C. & Bjorkman, O. 1998. Fluorescence quenching in four unicellular algae with
different light-harvesting and xanthophyll-cycle pigments. Photosynth. Res. 56: 277-289.
Dall'Osto, L., Caffari, S. & Bassi, R. 2005. A mechanism of non-photochemical energy
dissipation, independent from PsbS, revealed by a conformational change in the antenna protein
CP26. Plant Cell 17: 1-16.
Demming-Adams, B. & Adams III, W.W. 2000. Harvesting sunlight safely. Nature 403: 371-
374.
Díez, B., Massana, R., Estrada, M. & Pedrós-Alió, C. 2004. Distribution of eukaryotic
picoplankton assemblages across hydrographic fronts in the Southern ocean, studied by
denaturating gradient gel electrophoresis. Limnol. Oceanog. 49: 1022-1034.
Eilers, P.H.C. & Peeters, J.C.H. 1988. A model for the relationship between light intensity and
the rate of photosynthesis in phytoplankton. Ecol. Model. 42: 199-215.
Emanuelson, A., Eskling, M. & Akerlund, H.E. 2003. Chemical and mutational modifications of
histidines in violaxanthin de-epoxidase from Spinacia oleracea. Physiol. Plant. 119: 97-104.
20
Finazzi, G., Johnson, G.N., Dall’osto, L., Zito, F., Bonente, G., Bassi, R. & Wollman, F.A. 2006.
Nonphotochemical quenching of chlorophyll fluorescence in Chlamydomonas reinhardtii.
Biochemistry, 45: 1490-1498.
Fisher, T., Berner, T., Iluz, D. & Dubinsky, Z. 1998. The kinetics of the photoacclimation
response of Nannochloropsis sp. (Eustigmatophyceae): A study of changes in ultrastructure and
PSU density. J. Phycol. 34: 818-824.
Floder, S. & Burns, C.W. 2005. The influence of fluctuating light on diversity and species
number of nutrient-limited phytoplankton. J. Phycol. 41: 950-956.
Fogg, G.E. 1991. The phytoplanktonic ways of life. New phytol. 118 : 191-232.
Garcia-Mendoza, E., Matthijs, H.C.P., Schubert H. & Mur, L.R. 2002. Non-photochemical
quenching of chlorophyll fluorescence in Chlorella fusca acclimated to constant and dynamic
light conditions. Photosynth. Res. 74: 303-315.
Garcia-Plazaola, J.I., Hernandez, A., Artetxe, U. & Becerril, J.M. 2002. Regulation of the
Xanthophyll cycle pool size in duckweed (Lemna minor) plants. Physiol. Plant. 116: 121-126.
Gilmore, A.M., Mohanty, N. & Yamamoto, H.Y. 1994. Epoxidation of zeaxanthin and
antheraxanthin reverses non-photochemical quenching of photosystem II chlorophyll a
fluorescence in the presence of trans-thylakoid delta-pH. FEBS Lett. 350: 271-274.
Gilmore, A. M., Shinkarev, V.P., Hazlett, T. L. & Govindjee 1998. Quantitative analysis of the
effects of intrathylakoid pH and xanthophyll pigments on chlorophyll a fluorescence lifetime
distributions and intensity in thylakoids. Biochemistry 37: 13582-13593.
Gilmore, A. M. & Yamamoto, H.Y. 2001. Time-resolution of the antheraxanthin- and delta pH-
dependent chlorophyll a fluorescence components associated with photosystem II energy
dissipation in Mantoniella squamata. Photochem. Photobiol. 74: 291-302.
21
Harris, G.N., Scanlan, D.J. & Geider, R.J. 2005. Acclimation of Emiliania huxleyi
(Prymnesiophyceae) to photon flux density. J. Phycol. 41: 851-862.
Havaux, M. & Niyogi, K.K. 1999. The violaxanthin cycle protects plants from photooxidative
damage by more than one mechanism. Proc. Natl. Acad. Sci. U.S.A. 96: 8762-8767.
Havaux, M., Bonfils, J.P., Lutz, C. & Niyogi, K.K. 2000. Photodamage of the photosynthetic
apparatus and its dependence on the leaf developmental stage in the npq1 Arabidopsis mutant
deficient in the xanthophyll cycle enzyme violaxanthin de-epoxidase. Plant Physiol. 124: 273-
284.
Havelkova-Dousova, H., Prasil, O. & Behrenfeld, M.J. 2004. Photoacclimation of Dunaliella
tertiolecta (Chlorophyceae) under fluctuating irradiance. Photosynthetica 42: 273-281.
Henley, W.J., Hironaka, J.L. Guillou, L., Buchheim, M.A., Buchheim, J.A., Fawley, M.W. &
Fawley, K.P. 2004. Phylogenetic analysis of the “Nannochloris-like” algae and diagnoses of
Picochlorum oklahomensis gen. et sp. nov. (Trebouxiophyceae, Chlorophyta). Phycologia 43:
641-652.
Horton, P. & Ruban, A.V. 2005. Molecular design of the photosystem II light-harvesting
antenna: photosynthesis and photoprotection. J. Exp. Bot. 56: 365-373.
Jahns, P., Depka, B. & Trebst, A. 2000. Xanthophyll cycle mutants from Chlamydomonas
reinhardtii indicate a role for zeaxanthin in the D1 protein turnover. Plant Physiol. Biochem. 38:
371-376.
Jakob, T., Goss, R. & Wilhelm, C. 2001. Unusual pH-dependence of diadinoxanthin de-
epoxidase activation causes chlororespiratory induced accumulation of diatoxanthin in the
diatom Phaeodactylum tricornutum. J. Plant Physiol. 158: 383-390.
22
Keller, M.D., Selvin, R.C., Claus, W. & Guillard, R.R.L. 1987. Media for the culture of oceanic
ultraphytoplantkon. J. Phycol. 23: 633-638.
Lavaud, J., Rousseau, B., van Gorkom, H. J. & Etienne, A.L. 2002. Influence of the
diadinoxanthin pool size on photoprotection in the marine planktonic diatom Phaeodactylum
tricornutum. Plant Physiol. 129: 1398-1406.
Lavaud, J., Rousseau, B. & Etienne, A.L. 2004. General features of photoprotection by energy
dissipation in planktonic diatoms (Bacillariophyceae). J. Phycol. 40: 130-137.
Litchman, E. & Klausmeier, C.A. 2001. Competition of phytoplankton under fluctuating light.
Am. Nat. 157: 170-187.
Lohr, M. & Wilhelm, C. 2001. Xanthophyll synthesis in diatoms: quantification of putative
intermediates and comparison of pigments conversion kinetics with rates constants derived from
a model. Planta 212: 382-391.
Masojidek, J., Kopecky, J., Koblizek, M. & Torzillo, G. 2004. The xanthophyll cycle in green
algae (Chlorophyta): Its role in the photosynthetic apparatus. Plant Biology 6: 342-349.
Masojidek, J., Torzillo, G., Koblizek, J., Bernardini, P., Sacchi, A. & Komenda, J. 1999.
Photoadaptation of two members of the Chlorophyta (Scenedesmus and Chlorella) in laboratory
and outdoor cultures: changes in chlorophyll fluorescence quenching and the xanthophyll cycle.
Planta 209: 126-135.
Moon-van der staay, S. Y., De Watcher, R. & Vaulot, D. 2001. Oceanic 18S rDNA sequences
from picoplankton reveal unsuspected eukaryotic diversity. Nature 409: 607-610.
Niyogi, K. K., Li, X.P., Rosenberg, V., Jung, H.S. 2004. Is PsbS the site of non-photochemical
quenching in photosynthesis? J. Exp. Bot. 56: 375-382.
23
Not, F., Massana, R., Latasa, M., Marie, D., Colson, C., Eikrem, W., Pedrós-Alió, C., Vaulot, D.
& Simon, N. 2005. Late summer community composition and abundance of photosynthetic
picoeukaryotes in Norwegian and Barents Sea. Limnol. Oceanogr. 50: 1677-1686.
Pascal, A. A., Liu, Z., Broess, K., van Oort, B., van Amerongen, H., Wang, C., Horton, P.,
Robert, B., Chang, W. & Ruban, A. 2005. Molecular basis of photoprotection and control of
photosynthetic light-harvesting. Nature 436: 134-138.
Raven, J.A. 1998. The twelfth Tansley lecture. Small is beautiful: the picophytoplankton. Funct.
Ecol. 12: 503.
Ruban, A. & Horton, P. 1999. The xanthophyll cycle modulates the kinetics of non-
photochemical energy dissipation in isolated light-harvesting complexes, intact chloroplasts and
leaves. Plant Physiol. 119: 531-542.
Schubert, H., Kroon, B.M.A. & Matthijs, H.C.P. 1994. In vivo Manipulation of the xanthophyll
cycle and the role of zeaxanthin in the protection against photodamage in the green alga
Chlorella pyrenoidosa. J. Biol. Chem. 269: 7267-7272.
Sobrino, C., Neale, P.J. & Lubian, L.M. 2005. Interaction of UV radiation and inorganic carbon
supply in the inhibition of photosynthesis: Spectral and temporal responses of two marine
picoplankton. Photochem. Photobiol. 81: 384-393.
Stomp, M., Huisman, J., de Jongh, F., Veraart, A.J., Gerla, D., Riijkeboer, M., Ibelings, B.W.,
Wollenzien, U.I.A. & Stal, L.J. 2004. Adaptive divergence in pigment composition promotes
phytoplankton biodiversity. Nature 432: 104-107.
Strzepek, R.F. & Harrison, P.J. 2004. Photosynthetic architecture differs in coastal and oceanic
diatoms. Nature 431: 689-692.
24
Tardy, F. & Havaux, M. 1996. Photosynthesis, chlorophyll fluorescence, light-harvesting system
and photoinhibition resistance of zeaxanthin-accumulating mutant of Arabidopsis thaliana.
Photochem. Photobiol. 34: 87-94.
Tassan, S. & Ferrari, G.M. 1995. An alternative approach to absorption measurements of aquatic
particles retained on filters. Limnol. Oceanogr. 40: 1358-1368.
Timmermans, K.R., van der Wagt, B., Veldhuis, M.J.W., Maatman, A. & de Baar, H.J.W. 2005.
Physiological responses of three species of marine pico-phytoplankton to ammonium, phosphate,
iron and light limitation. J. Sea Res. 53: 109-120.
Vaulot, D., Le Gall, F., Marie, D., Guillou, L. & Partensky, F. 2004. The Roscoff Culture
Collection (RCC): a collection dedicated to marine picoplankton. Nova Hedwigia 79: 49-70.
Veldhuis, M.J.W., Timmermans, K.R., Croot, P. & van der Wagt, B. 2005. Picophytoplantkon: a
comparative study of their biochemical composition and photosynthetic properties. J. Sea Res.
53: 7-24. Villareal, T.A. 2004. Single-cell pulse amplitude modulation fluorescence
measurements of the giant diatom Ethmodiscus (Bacillariophyceae). J. Phycol. 40: 1052-1061.
Wilhelm, C., Kramer, P.P. & Lenartz-Weiler, I. 1989. The energy distribution between the
photosystems and light-induced changes in the stoichiometry of system I and system II reaction
centers in the chlorophyll b-containing alga Mantoniella squamata (Prasinophyceae).
Photosynth. Res. 20: 221-233.
25
LEGENDS
Fig.1: Progressive light-increase experiment. Temporal variation of (a): light intensity (µmol
photons·m-2·s-1), (b): Vx/chl a (diamond), Ax/chl a (circle) and Zx/chl a (triangle) in mol·mol chl
a-1 and (c): DPS (triangle) and effNPQ (circle). Values are means (± SD), n = 3 from independent
experiments.
Fig. 2: Progressive light-increase experiment. Mean curves of ETR (a) and NPQ (b) vs PFD
obtained from RLC measurements.
Fig. 3: Progressive light-increase experiment. Temporal variation of cellular RALS (a), FALS
(b) and red fluorescence (c) in the changing light culture (diamond) and in the control (circle).
Values are means (± SD) expressed in units relative to the beads, n = 3 from independent
experiments.
Fig. 4: High to low PFD experiment. Temporal variation of: (a): Vx/chl a (diamond), Ax/chl a
(circle) and Zx/chl a (triangle) in mol·mol chl a-1 and (b): Fq'/Fm' (operating efficiency of PSII,
triangle) and Fv/Fm (maximum photochemical efficiency, circle). Values are means (± SD), n =
3 from independent experiments.
Fig. 5: DTT-added experiment. Temporal variation of (a): effNPQ (DTT-treated culture: (circle;
control: triangle), and Vx/chl a (diamond), Ax/chl a (circle) and Zx/chl a (triangle) in mol·mol
26
chl a-1 in the DTT-treated culture (b) and in the control (c). Values are means (± SD) with n = 3
from independent experiments, except the control (n = 1).
Fig. 6: NF-added experiment. Temporal variation of: Vx/chl a (diamond), Ax/chl a (circle) and
Zx/chl a (triangle) in the NF-treated culture (a) and in the control (b); ß-carotene/chl a in the NF-
treated culture (c) and in the control (d); effNPQ (e, NF-treated culture: circle; control: triangle)
and Fv/Fm (f, NF-treated culture: circle; control: triangle). All the pigment ratios are in mol·mol
chl a-1 and the values are means (± SD) with n = 3 from independent experiments, except the
control (n =1).
0 100 200 300 400
Time (min)
0
100
200
300
400
Lig
ht in
tens
ity (
µmol
pho
tons
. m
-2 .
s-1 )
0 100 200 300 400Time (min)
0.00
0.01
0.02
0.03
0.04
0.05
Pigm
ent r
atio
(mol
. m
ol c
hla-
1 )
0 100 200 300 400
Time (min)
0.0
0.2
0.4
0.6
0.8
1.0
DPS
0.0
0.1
0.2
0.3
0.4
0.5
eff NPQ
a
b
c
DIMIER et al. Fig. 1
DIMIER et al. Fig. 2
a
0 400 800 1200 1600
0
4
8
12
16
ETR
(mol
e-. g
chla
-1. h
-1)
(µmol photons m-2 s-1)PFD
0 400 800 1200 1600. .
0.0
0.4
0.8
1.2
1.6
2.0
NPQ
b
(µmol photons m-2 s-1)PFD
DIMIER et al. Fig. 3
0 100 200 300 400
Time (min)
0.016
0.020
0.024
0.028
0.032
0.036
RA
LS (r
elat
ive
units
) a
0 100 200 300 400
Time (min)
1.5
1.6
1.7
1.8
1.9
2.0
chla
fluo
resc
ence
(rel
ativ
e un
its)
c
0 100 200 300 400
Time (min)
0.04
0.08
0.12
0.16
0.20
FAL
S (r
elat
ive
units
) b
DIMIER et al. Fig. 4
0 40 80 120 160 200
Time (min)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
Pigm
ent r
atio
(mol
. m
ol c
hla-
1 ) a
0 40 80 120 160 200
Time (min)
0.56
0.58
0.60
0.62
0.64
0.66
0.68
Fv/F
m a
nd F
q'/F
m'
b
0 40 80 120
Time (min)
0.00
0.04
0.08
0.12
eff N
PQ
0 40 80 120Time (min)
0.00
0.01
0.02
0.03
0.04
Pigm
ent r
atio
(mol
. m
ol c
hla-
1 )
0 40 80 120
Time (min)
0.00
0.01
0.02
0.03
0.04
Pigm
ent r
atio
(mol
. m
ol c
hla-
1 )
b
c
a
DIMIER et al. Fig. 5
a b
0 20 40 60 80 100
Time (min)
0.00
0.01
0.02
0.03
0.04
Pigm
ent r
atio
(mol
. m
ol c
hla-
1 )
c d
0 20 40 60 80 100
Time (min)
0.036
0.038
0.040
0.042
0.044
0.046
ß-ca
rote
ne (m
ol .
mol
chl
a-1 )
0 20 40 60 80 100
Time (min)
0.030
0.032
0.034
0.036
0.038
0.040
ß-ca
rote
ne (m
ol .
mol
chl
a-1 )
e f
0 20 40 60 80 100
Time (min)
0.00
0.01
0.02
0.03
0.04Pi
gmen
t rat
io (m
ol .
mol
chl
a-1 )
DIMIER et al. Fig. 6
0 40 80 120
Time (min)
0.45
0.50
0.55
0.60
0.65
0.70
0.75
Fv/F
m
0 40 80 120Time (min)
0.00
0.10
0.20
0.30
0.40
0.50
eff N
PQ