173 CHAPTER 12 12.1 Signal Transduction Depends on Molecular Circuits 12.2 Receptor Proteins Transmit Information Into the Cell 12.3 Metabolism in Context: Insulin Signaling Regulates Metabolism 12.4 Calcium Ion Is a Ubiquitous Cytoplasmic Messenger 12.5 Defects in Signaling Pathways Can Lead to Diseases Signal transduction is an important facility in all life forms. It allows an organism to sense the environment and formulate the proper biochemical response. Just as the computer chip has “on–off” switches that allow the transmission of information, cells have molecular “on–off” switches that allow the transmission of information in the cell and between cells. [Courtesy of Intel.] Signal-Transduction Pathways T his chapter provides an overview of how cells receive, process, and respond to information from the environment. Signal-transduction cascades mediate the sensing and processing of these stimuli. These molecular circuits detect, amplify, and integrate diverse external signals to generate responses such as changes in enzyme activity, gene expression, or ion-channel activity. This chapter introduces some of the basic principles of signal transduction and important classes of mol- ecules that participate in common signal-transduction pathways.
Transcript
173
CHAPTER
12
12.1 Signal Transduction Depends onMolecular Circuits
12.2 Receptor Proteins TransmitInformation Into the Cell
12.3 Metabolism in Context: InsulinSignaling Regulates Metabolism
12.4 Calcium Ion Is a UbiquitousCytoplasmic Messenger
12.5 Defects in Signaling PathwaysCan Lead to Diseases
Signal transduction is an important facility in all life forms. It allows an organism to sensethe environment and formulate the proper biochemical response. Just as the computer chiphas “on–off” switches that allow the transmission of information, cells have molecular“on–off” switches that allow the transmission of information in the cell and between cells.[Courtesy of Intel.]
Signal-TransductionPathways
This chapter provides an overview of how cells receive, process, and respond toinformation from the environment. Signal-transduction cascades mediate the
sensing and processing of these stimuli. These molecular circuits detect, amplify,and integrate diverse external signals to generate responses such as changes inenzyme activity, gene expression, or ion-channel activity. This chapter introducessome of the basic principles of signal transduction and important classes of mol-ecules that participate in common signal-transduction pathways.
12.1 Signal Transduction Depends on Molecular CircuitsSignal-transduction pathways follow a broadly similar course that can be viewedas a molecular circuit (Figure 12.1). All such circuits contain certain key steps:
1. Release of the Primary Messenger. A stimulus such as a wound or digested mealtriggers the release of the signal molecule, also called the primary messenger.
2. Reception of the Primary Messenger. Most signal molecules are too large and toopolar to pass through the cell membrane or through transporters. Thus, theinformation presented by signal molecules must be transmitted across the cellmembrane without the molecules themselves entering the cell. Membranereceptors transfer information from the environment to a cell’s interior. Suchreceptors are intrinsic membrane proteins that have both extracellular and intra-cellular domains. A binding site on the extracellular domain specifically recog-nizes the signal molecule (often referred to as the ligand). The formation of thereceptor–ligand complex alters the tertiary or quaternary structure of the recep-tor, including the intracellular domain. However, structural changes in the fewreceptors that are bound to ligands are not sufficient to yield a response from thecell. The information conveyed by the receptor must be transduced into otherforms of information that can alter the biochemistry of the cell.
3. Relay of Information by the Second Messenger. Structural changes in receptorslead to changes in the concentration of small molecules, called second messengers,which are used to relay information from the receptor–ligand complex. Partic-ularly important second messengers include cyclic AMP (cAMP) and cyclicGMP (cGMP, or cyclic guanosine monophosphate), calcium ion, inositol1,4,5-trisphosphate, (IP3), and diacylglycerol (DAG; Figure 12.2).
The use of second messengers has several consequences. First, second mes-sengers are often free to diffuse to other compartments of the cell, such as thenucleus, where they can influence gene expression and other processes. Sec-ond, the signal may be amplified significantly in the generation of second mes-sengers. Each activated receptor–ligand complex can lead to the generation ofmany second messengers within the cell. Thus, a low concentration of signal inthe environment, even as little as a single molecule, can yield a large intracellularsignal and response.
4. Activation of Effectors That Directly Alter the Physiological Response. The ulti-mate effect of the signal pathway is to activate (or inhibit) the pumps, enzymes,and gene-transcription factors that directly control metabolic pathways, geneactivation, and processes such as nerve transmission.
17412 Signal-Transduction Pathways
Signal
Reception
Transduction
Response(s)
Amplification
Figure 12.1 Principles of signaltransduction. An environmental signal isfirst received by interaction with a cellularcomponent, most often a cell-surfacereceptor. The information that the signalhas arrived is then converted into otherchemical forms, or transduced. Thetransduction process often comprises manysteps. The signal is often amplified beforeevoking a response. Feedback pathwaysregulate the entire signaling process.
A or GO
OH
O
P OO
O
OH2
Ca
OH2
H2O
H2OOH2
OH2H2O
OH
OH2–O3PO
OPO32–
OPO32–
HO
O
O
O
Calcium ion
2+
Cyclic AMP (cAMP),Cyclic GMP (cGMP)
Inositol 1,4,5-trisphosphate (IP3)
Diacylglycerol (DAG)
–
OH
O
H
Figure 12.2 Common secondmessengers. Second messengers areintracellular molecules that change inconcentration in response toenvironmental signals. That changein concentration conveys informationinside the cell.
5. Termination of the Signal. After a signaling process has been initiated and theinformation has been transduced to affect other cellular processes, the signalingprocesses must be terminated. Without such termination, cells lose their respon-siveness to new signals. Moreover, signaling processes that fail to be terminatedproperly may lead to uncontrolled cell growth and the possibility of cancer.
Essentially every biochemical process presented in the remainder of this bookeither is a component of a signal-transduction pathway or can be affected by one.
12.2 Receptor Proteins Transmit Information Into the CellMost receptor proteins that mediate information transfer into the cell interior fallinto three classes: seven-transmembrane-helix receptors, dimeric receptors thatrecruit protein kinases, and dimeric receptors that are protein kinases. We beginby considering the largest and one of the most important classes of receptor, theseven-transmembrane-helix receptors.
Seven-Transmembrane-Helix Receptors Change Conformation in Response to Ligand Binding and Activate G ProteinsThe seven-transmembrane-helix (7TM) receptors are responsible for transmit-ting information initiated by signals as diverse as photons, odorants, tastants,hormones, and neurotransmitters (Table 12.1). Several thousand such receptorsare known, and the list continues to grow. Indeed, approximately 50% of thedrugs that we use alter receptors of this class. As the name indicates, these recep-tors contain seven helices that span the membrane bilayer (Figure 12.3).An example of a 7TM receptor that responds to chemical signals is the b-adrenergicreceptor. This protein binds epinephrine (also called adrenaline), a hormone respon-sible for the “fight or flight” response. We will address the biochemical roles of thishormone in more detail later (p. 375).
A variety of evidence reveals that the 7TM receptors, particularly their cyto-plasmic domains, change conformation in response to ligand binding. Thus,the binding of a ligand from outside the cell induces a conformational change in the7TM receptor that can be detected inside the cell. As we shall see, 7TM receptors alsohave in common the next step in their signal-transduction cascades.
17512.2 Receptor Proteins
Table 12.1 Biological functionsmediated by 7TM receptors
Hormone actionHormone secretionNeurotransmissionChemotaxisExocytosisControl of blood pressureEmbryogenesisCell growth and differentiationDevelopmentSmellTasteVisionViral infection
Source: After J. S. Gutkind, J. Biol. Chem.273:1839–1842, 1998.
C
N
(A) (B)
Cytoplasmic loops
N
C
Ligand-bindingsite
Figure 12.3 The structure of 7TMreceptors. (A) Schematic
representation of a 7TM receptor showinghow it passes through the membraneseven times. (B) Three-dimensionalstructure of rhodopsin, a 7TM receptortaking part in visual signal transduction.Notice the ligand-binding site near theextracellular surface. As the first 7TMreceptor for which the structure wasdetermined, rhodopsin provides aframework for understanding other 7TMreceptors. [Drawn from 1F88.pdb.]
HOH H
NCH3
HO
HOEpinephrine
Ligand Binding to 7TM Receptors Leads to the Activation of G ProteinsLet us focus on the �-adrenergic receptor as a model of the 7TM receptor class.What is the next step in the pathway after the binding of epinephrine by the �-adrenergic receptor? The conformational change in the cytoplasmic domain ofthe receptor activates a GTP-binding protein. This signal-coupling protein istermed a G protein (G for guanyl nucleotide). The activated G protein stimulatesthe activity of adenylate cyclase, an enzyme that increases the concentration of thesecond messenger cAMP by forming it from ATP (Figure 12.4).
How do these G proteins operate? In the unactivated state, the guanylnucleotide bound to the G protein is GDP. In this form, the G protein exists as a het-erotrimer consisting of �, �, and � subunits; the � subunit (referred to as Ga)binds the nucleotide (Figure 12.5). The � and � subunits are usually anchored tothe membrane by covalently attached fatty acids. The exchange of the bound GDPfor GTP is catalyzed by the hormone-bound receptor. The hormone–receptor com-plex interacts with the heterotrimeric G protein and opens the nucleotide-bindingsite so that GDP can depart and GTP from solution can bind. The � subunitsimultaneously dissociates from the �� dimer (Gbg), see Figure 12.4). The disso-ciation of the G-protein heterotrimer into Ga and Gbg units transmits the signal thatthe receptor has bound its ligand.
17612 Signal-Transduction Pathways
GTP
γ
β-Adrenergicreceptor
Epinephrine Adenylatecyclase
β
αGDP
Proteinkinase A
ATPCyclicAMP
Proteinkinase A
Figure 12.4 The activation of proteinkinase A by a G-protein pathway. Hormonebinding to a 7TM receptor initiates asignal-transduction pathway that actsthrough a G protein and cAMP to activateprotein kinase A.
α
γ
βGDP
GDP
(B)(A)
Figure 12.5 A heterotrimeric Gprotein. (A) A ribbon diagram shows
the relation between the three subunits. Inthis complex, the � subunit (gray andpurple) is bound to GDP. Notice that GDPis bound in a pocket close to the surface atwhich the � subunit interacts with the ��dimer. (B) A schematic representation ofthe heterotrimeric G protein. [Drawn from1GOT.pdb.]
A single hormone–receptor complex can stimulate nucleotide exchange in manyG-protein heterotrimers. Thus, hundreds of G� molecules are converted from theirGDP into their GTP forms for each bound molecule of hormone, giving an ampli-fied response. All 7TM receptors appear to be coupled to G proteins, and so the7TM receptors are sometimes referred to as G-protein-coupled receptors or GPCRs.
Activated G Proteins Transmit Signals by Binding to Other ProteinsAs described in the preceding subsection, the formation of the hormone–receptorcomplex activates a G protein. How does the G protein propagate the messagethat the hormone is present? It does so by a variety of means, depending on thespecific type of G protein. We will begin by examining one target of a G protein,the enzyme adenylate cyclase (Figure 12.6). The adenylate cyclase enzyme that isactivated by the �-adrenergic signaling pathway is a membrane protein thatcontains 12 presumed membrane-spanning helices. The G� protein binds toadenylate cyclase on the G� surface that had bound the �� dimer when the G�
protein was in its GDP form. G�s (where “s” stands for “stimulatory”) stimulatesadenylate cyclase activity, thus increasing cAMP production. The net result is thatthe binding of epinephrine to the receptor on the cell surface increases the rate ofcAMP production inside the cell.
Adenylate cyclase fragment
Gαs (GTP form)
Adenylate cyclase
(A) (B)
CN
Figure 12.6 Adenylate cyclase activation. (A) Adenylate cyclase is a membraneprotein with two large intracellular domains (red and orange) that contain the
catalytic apparatus. (B) The structure of the complex between G� in its GTP form bound to acatalytic fragment of adenylate cyclase. Notice that the surface of G� that had been boundto the �� dimer (see Figure 12.5) now binds adenylate cyclase. [Drawn from 1AZS.pdb.]
Cyclic AMP Stimulates the Phosphorylation of Many Target Proteins by Activating Protein Kinase AThe increased concentration of cAMP can affect a wide range of cellular processes,depending on the cell type. For example, it enhances the degradation of storagefuels, increases the secretion of acid by the gastric mucosa in the cells of the stom-ach and intestines, leads to the dispersion of melanin pigment granules in skincells, diminishes the aggregation of blood platelets, and induces the opening ofchloride channels in the pancreas. How does cAMP influence so many cellularprocesses? Is there a common denominator for its diverse effects? Indeed there is.Most effects of cyclic AMP in eukaryotic cells are mediated by the activation of a sin-gle protein kinase. This key enzyme is called protein kinase A (PKA). Kinases areenzymes that phosphorylate a substrate at the expense of a molecule of ATP. PKA
17712.2 Receptor Proteins
consists of two regulatory (R) chains and two catalytic (C) chains. In the absenceof cAMP, the R2C2 complex is catalytically inactive (Figure 12.7). The binding ofcAMP to the regulatory chains releases the catalytic chains, which are enzymati-cally active on their own. Activated PKA then phosphorylates specific serine andthreonine residues in many targets to alter their activity. The cAMP cascade isturned off by cAMP phosphodiesterase, an enzyme that converts cAMP into AMP,which does not activate PKA. The C and R subunits subsequently rejoin to formthe inactive enzyme.
G Proteins Spontaneously Reset Themselves Through GTP HydrolysisThe ability to shut down signal-transduction pathways is as critical as the abilityto turn them on. How is the signal initiated by activated 7TM receptors switchedoff? Ga subunits have intrinsic GTPase activity, hydrolyzing bound GTP to GDPand Pi (inorganic orthophosphate) and thereby deactivating itself. This hydrol-ysis reaction is slow, however, requiring from seconds to minutes and thus allow-ing the GTP form of G� to activate downstream components of thesignal-transduction pathway before GTP hydrolysis deactivates the subunit. Inessence, the bound GTP acts as a built-in clock that spontaneously resets the Ga sub-unit after a short time period. After GTP hydrolysis and the release of Pi, the GDP-bound form of G� then reassociates with G�� to reform the heterotrimericprotein (Figure 12.8).
CC C
C
cAMP-bindingdomains
cAMP
+ 4 cAMPR
RR
R++
Active Active
Figure 12.7 The regulation of protein kinase A. The binding of four molecules of cAMPactivates protein kinase A by dissociating the inhibited holoenzyme (R2C2) into a regulatorysubunit (R2) and two catalytically active subunits (C).
GDPGDP
Adenylatecyclase
GTPH2O Pi
Figure 12.8 Resetting G�. On hydrolysis of the bound GTP by the intrinsic GTPase activity ofG�, G� reassociates with the �� dimer to form the heterotrimeric G protein, therebyterminating the activation of adenylate cyclase.
178
The hormone-bound activated receptor must be reset as well to prevent thecontinuous activation of G proteins. A key step in the inactivation of the receptorrests on the fact that the receptor–ligand interaction is reversible (Figure 12.9).When the hormone dissociates, the receptor returns to its initial, unactivated state.The likelihood that the receptor remains in its unbound state depends on the con-centration of hormone in the environment.
Clinical Insight
Cholera and Whooping Cough Are Due to Altered G-Protein ActivityThe alteration of G-protein-dependent signal pathways can result in pathologicalconditions. Let us first consider the mechanism of action of the cholera toxin,secreted by the intestinal bacterium Vibrio cholera. Cholera is an acute diarrhealdisease that can be life threatening. It causes a voluminous secretion of electrolytesand fluids from the intestines of infected persons (Figure12.10). The cholera toxin, choleragen, is a protein composedof two functional units—a B subunit that binds to cells of theintestinal epithelium and a catalytic A subunit that enters thecell. The A subunit catalyzes the covalent modification of aG�s protein. This modification stabilizes the active GTP-bound form of G�s, trapping the molecule in the active con-formation. The active G protein, in turn, continuouslyactivates protein kinase A. PKA opens a chloride channel (aCFTR channel) and inhibits the Na�–H� exchanger byphosphorylation. The net result of the phosphorylation ofthese channels is an excessive loss of NaCl and the loss oflarge amounts of water into the intestine. Patients sufferingfrom cholera for 4 to 6 days may pass as much as twice theirbody weight in fluid. Treatment consists of rehydration witha glucose–electrolyte solution.
Whereas cholera is a result of a G protein trapped in theactive conformation, causing the signal-transduction path-way to be perpetually stimulated, pertussis, or whoopingcough, is a result of the opposite situation. The toxin alsomodifies a G� protein called G�i, which normally inhibitsadenylate cyclase, closes Ca2� channels, and opens K� chan-nels. The effect of this modification is to lower the G protein’saffinity for GTP, effectively trapping it in the “off” conforma-tion. The symptoms of whooping cough, such as prolongedcoughing that ends with a whoop as the patient gasps for air,have not yet been traced to the inhibition of any single targetof the G�i protein. Pertussis toxin is secreted by Bordetellapertussis, the bacterium responsible for whooping cough. ■
17912.2 Receptor Proteins
Dissociation
Figure 12.9 Signal termination. Signaltransduction by the 7TM receptor ishalted, in part, by dissociation of thesignal molecule (yellow) from the receptor.
QUICK QUIZ 1 List the means bywhich the �-adrenergic pathway is
terminated.
Figure 12.10 Death’s dispensary. An 1866 cartoon illustrating thatcontaminated water is a frequent source of cholera infection. [The Granger Collection.]
The Hydrolysis of Phosphatidyl Inositol Bisphosphate by Phospholipase C Generates Two Second MessengersCyclic AMP is not the only second messenger employed by 7TM receptors andthe G proteins. We turn now to another ubiquitous second-messenger cascadeused by many hormones to evoke a variety of responses. The phosphoinositidecascade, like the adenylate cyclase cascade, converts extracellular signals intointracellular ones. The intracellular messengers formed by activation of thispathway arise from the cleavage of phosphatidylinositol 4,5-bisphosphate (PIP2),a phospholipid present in cell membranes. The binding of a hormone such asvasopressin to a 7TM receptor leads to the activation of phospholipase C. TheG� protein that activates phospholipase C is called G�q. The activated enzymethen hydrolyzes the phosphodiester linkage joining the phosphorylated inosi-tol unit to the acylated glycerol moiety. The cleavage of PIP2 produces two mes-sengers: inositol 1,4,5-trisphosphate (IP3), a soluble molecule that can diffusefrom the membrane, and diacylglycerol (DAG), which stays in the membrane(Figure 12.11).
What are the biochemical effects of the second messenger IP3? UnlikecAMP, IP3 does not cause a cascade of phosphorylation to elicit a response fromthe cell. IP3 directly causes the rapid release of Ca2� from intracellular stores—theendoplasmic reticulum and, in smooth muscle cells, the sarcoplasmic reticulum. IP3associates with a membrane protein called the IP3-gated channel or IP3 receptorto allow the flow of Ca2� from the endoplasmic reticulum into the cell cyto-plasm. The elevated level of Ca2� in the cytoplasm then triggers a variety of bio-chemical processes such as smooth-muscle contraction, glycogen breakdown,and vesicle release.
The lifetime of IP3 in the cell is very short—less than a few seconds. It israpidly converted into derivatives that have no effect on the IP3-gated channel.
Figure 12.11 The phospholipase C reaction. Phospholipase C cleaves the membrane lipidphosphatidylinositol 4,5-bisphosphate into two second messengers: diacylglycerol, whichremains in the membrane, and inositol 1,4,5-trisphosphate, which diffuses away from themembrane.
Calciumion
CytoplasmIP3receptor
IP3
PIP2
Protein kinase C
Calciumion
ER membrane
Cell membraneDiacylglycerol (DAG)
Phospholipase Ccleavage
DAG
Figure 12.12 The phosphoinositide cascade. The cleavage of phosphatidylinositol 4,5-bisphosphate (PIP2) into diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3)results in the release of calcium ions (owing to the opening of the IP3 receptor ionchannels) and the activation of protein kinase C (owing to the binding of protein kinaseC to free DAG in the membrane). Calcium ions bind to protein kinase C and helpfacilitate its activation.
18112.2 Receptor Proteins
Lithium ion, widely used to treat bipolar affective disorder, may act by inhibit-ing the recycling of IP3, although the details of lithium action remain to bedetermined.
Diacylglycerol, the other molecule formed by the receptor-triggered hydrol-ysis of PIP2, also is a second messenger that, in conjunction with Ca2�, activatesprotein kinase C (PKC), a protein kinase that phosphorylates serine and threonineresidues in many target proteins (Figure 12.12).
Some Receptors Dimerize in Response to Ligand Binding and Recruit Tyrosine KinasesThe 7TM receptors initiate signal-transduction pathways through changes in ter-tiary structure that are induced by ligand binding. A fundamentally differentmechanism is utilized by a number of other classes of receptors. For these recep-tors, ligand binding leads to changes in quaternary structures—specifically, theformation of receptor dimers.
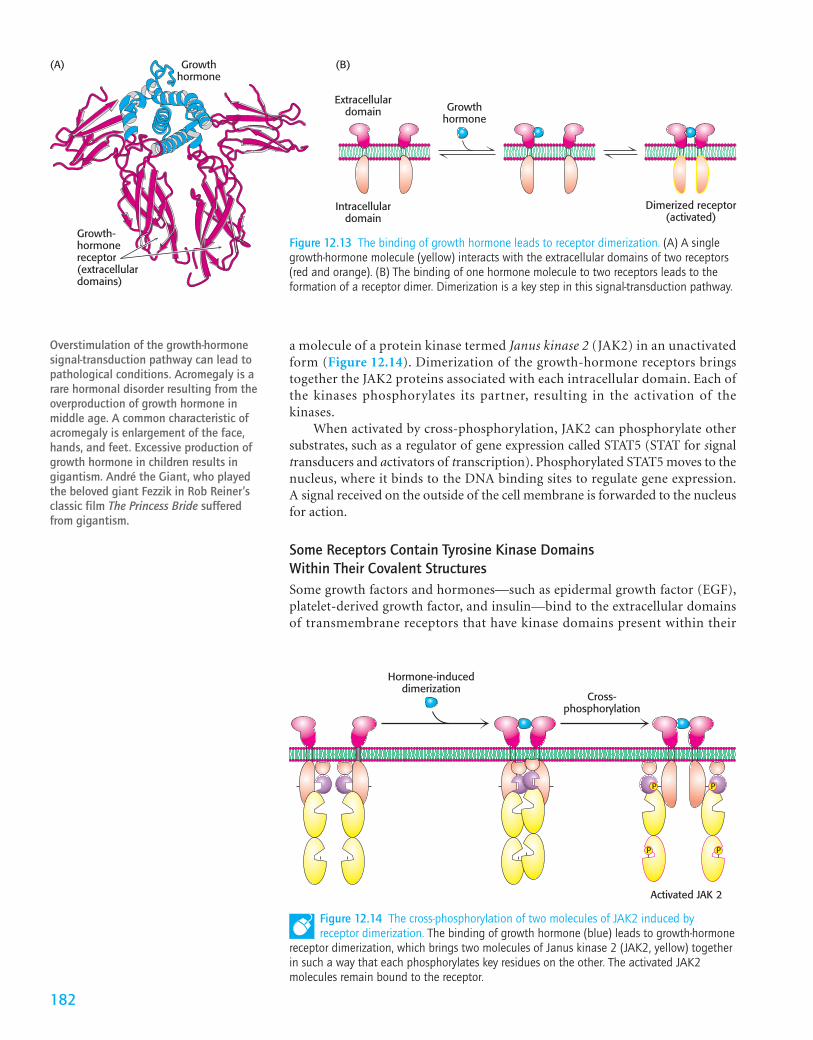
We consider human growth hormone and its receptor as an example. Growthhormone is a monomeric protein of 217 amino acids. The growth-hormonereceptor has an extracellular domain, a single membrane-spanning helix, and anintracellular domain. In the absence of bound hormone, the receptor is presentas a monomer. Each monomeric hormone binds to the extracellular domains oftwo receptor molecules, thus promoting the formation of a dimer of the recep-tor (Figure 12.13).
Dimerization of the extracellular domains of the receptor brings togetherthe intracellular domains as well. Associated with each intracellular domain is
a molecule of a protein kinase termed Janus kinase 2 (JAK2) in an unactivatedform (Figure 12.14). Dimerization of the growth-hormone receptors bringstogether the JAK2 proteins associated with each intracellular domain. Each ofthe kinases phosphorylates its partner, resulting in the activation of thekinases.
When activated by cross-phosphorylation, JAK2 can phosphorylate othersubstrates, such as a regulator of gene expression called STAT5 (STAT for signaltransducers and activators of transcription). Phosphorylated STAT5 moves to thenucleus, where it binds to the DNA binding sites to regulate gene expression.A signal received on the outside of the cell membrane is forwarded to the nucleusfor action.
Some Receptors Contain Tyrosine Kinase Domains Within Their Covalent StructuresSome growth factors and hormones—such as epidermal growth factor (EGF),platelet-derived growth factor, and insulin—bind to the extracellular domainsof transmembrane receptors that have kinase domains present within their
Overstimulation of the growth-hormonesignal-transduction pathway can lead topathological conditions. Acromegaly is arare hormonal disorder resulting from theoverproduction of growth hormone inmiddle age. A common characteristic ofacromegaly is enlargement of the face,hands, and feet. Excessive production ofgrowth hormone in children results ingigantism. André the Giant, who playedthe beloved giant Fezzik in Rob Reiner’sclassic film The Princess Bride sufferedfrom gigantism.
Intracellulardomain
Dimerized receptor(activated)
Extracellulardomain
Growth-hormonereceptor(extracellulardomains)
Growthhormone
Growthhormone
(A) (B)
Figure 12.13 The binding of growth hormone leads to receptor dimerization. (A) A singlegrowth-hormone molecule (yellow) interacts with the extracellular domains of two receptors(red and orange). (B) The binding of one hormone molecule to two receptors leads to theformation of a receptor dimer. Dimerization is a key step in this signal-transduction pathway.
Hormone-induceddimerization
Cross-phosphorylation
Activated JAK 2
P P
P P
Figure 12.14 The cross-phosphorylation of two molecules of JAK2 induced byreceptor dimerization. The binding of growth hormone (blue) leads to growth-hormone
receptor dimerization, which brings two molecules of Janus kinase 2 (JAK2, yellow) togetherin such a way that each phosphorylates key residues on the other. The activated JAK2molecules remain bound to the receptor.
182
intracellular domains. Such receptors have a specific kind of kinase called tyro-sine kinase, which phosphorylates proteins on the hydroxyl group of tyrosineresidues. These receptor tyrosine kinases (RTKs) signal by mechanisms quite sim-ilar to those described for the pathway initiated by the growth-hormone recep-tor discussed in the preceding subsection. Of the more than 500 kinase genes inthe human genome, fewer than 100 encode tyrosine kinases, and all of thesetyrosine kinases appear to regulate the control of growth.
Consider, for example, epidermal growth factor, a 6-kd polypeptide that stimu-lates the growth of epidermal and epithelial cells by binding to the epidermal growthfactor receptor, a single polypeptide chain consisting of 1186 residues (Figure 12.15).The receptor tyrosine kinase is monomeric and enzymatically inactive in theabsence of the growth factor. The binding of EGF to its extracellular domain causesthe receptor to dimerize and undergo cross-phosphorylation and activation.
18312.2 Receptor Proteins
Transmembranehelix
Kinase domain C-terminal tail(tyrosine-rich)
EGF-binding domain
Figure 12.15 The modular structure of the EGF receptor. This schematic view of the aminoacid sequence of the EGF receptor shows the EGF-binding domain that lies outside the cell, asingle transmembrane helix-forming region, the intracellular tyrosine kinase domain, and thetyrosine-rich domain at the carboxyl terminus.
P P GDP GTPGTPRas GDP
Grb-2 ActivatedRasSos
EGFreceptor
EGF
Extracellulardomain
Intracellulardomain P P
Figure 12.16 The EGF signaling pathway. The binding of epidermal growth factor (EGF) toits receptor leads to cross-phosphorylation of the receptor. The phosphorylated receptorbinds Grb-2, which, in turn, binds Sos. Sos stimulates the exchange of GTP for GDP in Ras.Activated Ras binds to protein kinases and stimulates them (not shown).
How is the signal transferred beyond the receptor tyrosine kinase? A keyadaptor protein, called Grb-2, links the phosphorylation of the EGF receptor tothe stimulation of cell growth through a chain of protein phosphorylations(Figure 12.16). On phosphorylation of the receptor, Grb-2 binds to the phos-photyrosine residues of the receptor tyrosine kinase. Grb-2 then recruits a pro-tein called Sos. Sos, in turn, binds to Ras and activates it. Ras is a veryprominent signal-transduction component that we will consider shortly.Finally, Ras, in its activated form, binds to other components of the molecularcircuitry, leading to the activation of the specific protein kinases that phosphory-late specific targets that promote cell growth. We see here another example of howa signal-transduction pathway is constructed. Specific protein–protein interac-tions link the original ligand-binding event to the final result—the stimulationof cell growth.
Ras Belongs to Another Class of Signaling G ProteinThe signal-transduction protein Ras is member of an important family of signalproteins—the small G proteins, or small GTPases. This large superfamily ofproteins—grouped into subfamilies called Ras, Rho, Arf, Rab, and Ran—plays amajor role in a host of cell functions including growth, differentiation, cell motil-ity, cytokinesis, and the transport of materials throughout the cell (Table 12.2).Like their relatives the heterotrimeric G proteins (p. 176), the small G proteinscycle between an active GTP-bound form and an inactive GDP-bound form. Theydiffer from the heterotrimeric G proteins in being smaller (20–25 kd comparedwith 30–35 kd) and monomeric.
In their activated GTP-bound form, small G proteins such as Ras stimulatecell growth and differentiation. Recall that Sos is the immediate upstream link toRas in the circuit conveying the EGF signal. How does Sos activate Ras? Sos bindsto Ras, reaches into the nucleotide-binding pocket, and opens it up, allowing GDPto escape and GTP to enter in its place. Sos is referred to as a guanine-nucleotideexchange factor (GEF).
Like the G� protein, Ras possesses an intrinsic GTPase activity, which servesto terminate the signal and return the system to the inactive state. This activity isslow but is augmented by helper proteins termed GTPase-activating proteins(GAPs). The guanine-nucleotide exchange factors and the GTPase-activating pro-teins allow the G-protein cycle to proceed with rates appropriate for a balancedlevel of downstream signaling.
12.3 Metabolism in Context: Insulin Signaling Regulates Metabolism
Insulin is among the principal hormones that regulate metabolism, and we willexamine the effects of this hormone on many metabolic pathways later. This sec-tion presents an overview of its signal-transduction pathway. Insulin is the hor-mone released after eating a full meal and is the biochemical signal designatingthe fed state. In all of its detail, this multibranched pathway is quite complex; sowe will focus solely on the major branch. This branch leads to the mobilization ofglucose transporters to the cell surface. These transporters allow the cell to takeup the glucose that is plentiful in the bloodstream after a meal.
The Insulin Receptor Is a Dimer That Closes Around a Bound Insulin MoleculeInsulin is a peptide hormone that consists of two chains, linked by two disulfidebonds (Figure 12.17). The insulin receptor is a member of the receptor tyrosinekinase class of membrane proteins. The identical subunits form dimers on
18412 Signal-Transduction Pathways
Table 12.2 Ras superfamily of GTPases
Subfamily Function
Ras Regulates cell growth through serine or threonine protein kinasesRho Reorganizes cytoskeleton through serine or threonine protein kinasesArf Activates the ADP-ribosyltransferase of the cholera toxin A subunit;
regulates vesicular trafficking pathways; activates phospholipase DRab Plays a key role in secretory and endocytotic pathwaysRan Functions in the transport of RNA and protein into and out
of the nucleus
Figure 12.17 Insulin structure.Notice that insulin consists of two
chains (shown in blue and yellow) linkedby two interchain disulfide bonds. The �chain (blue) also has an intrachaindisulfide bond. [Drawn from IB2F.pdb.]
insulin binding, and this change in quaternary structure results in cross-phos-phorylation by the two kinase domains, activating the kinase activity (Figure12.18). Each subunit consists of one � chain and one � chain linked to oneanother by a single disulfide bond. Each � subunit lies completely outside thecell, whereas each � subunit lies primarily inside the cell, spanning the membrane with a single transmembrane segment. The two � subunitsmove together to form a binding site for a single insulin molecule.
The Activated Insulin Receptor Kinase Initiates a Kinase CascadeOn phosphorylation, the insulin receptor tyrosine kinase is activated. Because thetwo units of the receptor are held in close proximity to each other, additional siteswithin the receptor also are phosphorylated. These phosphorylated sites act asdocking sites for other substrates, including a class of molecules referred to asinsulin-receptor substrates (IRSs). The IRS proteins are subsequently phosphory-lated by the tyrosine kinase activity of the insulin receptor. The signal is conveyedto the cell interior by the IRS protein through a series of membrane-anchoredmolecules to a protein kinase that finally leaves the membrane (Figure 12.19).
185
Insulin-binding site
α subunit
β subunit
Figure 12.18 The insulin receptor. Thereceptor consists of two units, each ofwhich consists of an � subunit and a �subunit linked by a disulfide bond. The �subunit lies outside the cell and two �subunits come together to form a bindingsite for insulin. Each � subunit liesprimarily inside the cell and includes aprotein kinase domain.
P P
Insulinreceptor
Insulin
P
P
P
P
P
P
PP
P
ATPAkt
PDK1(PIP3-dependentprotein kinase)
IRS-1
Activated AktADP
Phosphoinositide3-kinase
PIP2
PIP3
Figure 12.19 Insulin signaling. Thebinding of insulin results in the cross-phosphorylation and activation of theinsulin receptor. Phosphorylated sites onthe receptor act as binding sites forinsulin-receptor substrates such as IRS-1.The lipid kinase phosphoinositide 3-kinasebinds to phosphorylated sites on IRS-1through its regulatory domain and thenconverts PIP2 into PIP3. Binding to PIP3activates PIP3-dependent protein kinase(PDK1), which phosphorylates andactivates kinases such as Akt. ActivatedAkt can then diffuse throughout the cell tocontinue the signal-transduction pathway.
Figure 12.20 The action of a lipid kinase in insulin signaling. Phosphorylated IRS-1 and IRS-2activate the enzyme phosphatidylinositide 3-kinase, an enzyme that converts PIP2 into PIP3.
In their phosphorylated form, the IRS molecules act as adaptor proteins. Thephosphotyrosine residues in the IRS proteins are recognized by other proteins, themost important of which is a lipid kinase that phosphorylates phosphatidylinos-itol 4,5-bisphosphate to form phosphatidylinositol 3,4,5-trisphosphate (PIP3;Figure 12.20, see also Figure 12.19). This lipid product, in turn, activates a protein
kinase, PDK1. This activated protein kinase phosphorylates and activates Akt,another protein kinase. Akt is not membrane anchored and moves through thecell to phosphorylate enzymes that stimulate glycogen synthesis as well as com-ponents that control the trafficking of the glucose transporter GLUT4 to the cellsurface. At the cell surface, GLUT4, one of a family of five glucose transporters,allows the entry of glucose down its concentration gradient into the cell.
The cascade initiated by the binding of insulin to the insulin receptor is sum-marized in Figure 12.21. The signal is amplified at several stages along this path-way. Because the activated insulin receptor itself is a protein kinase, each activatedreceptor can phosphorylate multiple IRS molecules Activated enzymes furtheramplify the signal in at least two of the subsequent steps. Thus, a small increase inthe concentration of circulating insulin can produce a robust intracellularresponse. Note that, as complicated as the pathway described here is, it is substan-tially less elaborate than the full network of pathways initiated by insulin.
Insulin Signaling Is Terminated by the Action of PhosphatasesWe have seen that the activated G protein promotes its own inactivation by therelease of a phosphoryl group from GTP. In contrast, proteins phosphorylated onserine, threonine, or tyrosine residues, such as insulin-receptor substrates, do nothydrolyze spontaneously; they are extremely stable kinetically. Specific enzymes,called protein phosphatases, are required to hydrolyze these phosphorylated pro-teins and convert them back into the states that they were in before the initiationof signaling. Similarly, lipid phosphatases are required to remove phosphorylgroups from inositol lipids that had been phosphorylated as part of a signalingcascade. In insulin signaling, three classes of enzymes are of particular impor-tance: protein tyrosine phosphatases that remove phosphoryl groups from tyro-sine residues on the insulin receptor, lipid phosphatases that hydrolyzephosphatidylinositol 3,4,5-trisphosphate to phosphatidylinositol 3,4-bisphos-phate, and protein serine phosphatases that remove phosphoryl groups from acti-vated protein kinases such as Akt. Many of these phosphatases are activated orrecruited as part of the response to insulin. Thus, the binding of the initial signalsets the stage for the eventual termination of the response.
12.4 Calcium Ion Is a Ubiquitous Cytoplasmic MessengerWe have already seen that Ca2� is an important component of one signal-transduction circuit, the phosphoinositide cascade. Indeed, Ca2� is itself an intra-cellular messenger in many eukaryotic signal-transducing pathways.
Calmodulin (CaM), a 17-kd protein with four calcium-binding sites, servesas a calcium sensor in nearly alleukaryotic cells. Calmodulin is acti-vated by the binding of Ca2� when thecytoplasmic calcium level is raised.Calmodulin is a member of the EF-hand protein family. The EF hand is aCa2�-binding motif that consists of ahelix, a loop, and a second helix. Thismotif, originally discovered in the pro-tein parvalbumin, was named the EFhand because the helices designated Eand F in parvalbumin that form thecalcium-binding motif are positionedlike the forefinger and thumb of theright hand (Figure 12.22).
Insulin
Activatedreceptor
Phosphorylated IRS proteins
Insulin receptor+
Localized phosphoinositide 3-kinase
Phosphatidyl inositol-3,4,5-trisphosphate (PIP3)
Activated PIP3-dependentprotein kinase
Cross-phosphorylation
Enzymaticreaction
Amplification
Protein–proteininteraction
Enzymaticreaction
Enzymaticreaction
Amplification
Activated Aktprotein kinase
Increased glucose transporteron cell surface
Protein–lipidinteraction
Amplification
Figure 12.21 Insulin signaling pathway.Key steps in the signal-transductionpathway initiated by the binding of insulinto the insulin receptor.
EF hand
Ca2+Figure 12.22 An EF hand. Formedby a helix-loop-helix unit, an EF
hand is a binding site for Ca2+ in manycalcium-sensing proteins. Here, the E helixis yellow, the F helix is blue, and calciumis represented by the green sphere. Noticethat the calcium ion is bound in a loopconnecting two nearly perpendicularhelices. [Drawn from 1CLL.pdb.]
QUICK QUIZ 2 Why does it makegood physiological sense for insulin
to increase the number of glucosetransporters in the cell membrane?
186
The Ca2�–calmodulin complex stimulates a wide array of enzymes, pumps, andother target proteins. Two targets are especially noteworthy: one that propagatesthe Ca2� signal and another that abrogates it. The binding of Ca2�–calmodulinto a calmodulin-dependent protein kinase (CaM kinase) activates the kinase andenables it to phosphorylate a wide variety of target proteins. CaM kinases regu-late the metabolism of fuel, ionic permeability, neurotransmitter synthesis, andneurotransmitter release through the action of the Ca2�–calmodulin complex.The plasma membrane Ca2�–ATPase pump is another important target ofCa2�–calmodulin. Stimulation of the pump by Ca2�–calmodulin drives the cal-cium level down to restore a low-calcium basal state to the cell, thus helping toterminate the signal.
12.5 Defects in Signal-Transduction Pathways Can Lead to Disease
In light of their complexity, it comes as no surprise that signal-transduction pathwaysoccasionally fail, leading to pathological or disease states.Cancer,a set of diseases char-acterized by uncontrolled or inappropriate cell growth, is strongly associated withdefects in signal-transduction proteins. Indeed, the study of cancer has contributedgreatly to our understanding of signal-transduction proteins and pathways.
Genes that, when mutated, cause cancer often normally regulate cell growth.The unmutated, normally expressed versions of these genes are termed proto-oncogenes, and the proteins that they encode are signal-transduction proteins thatregulate cell growth. If a proto-oncogene suffers a mutation that leads to unre-strained growth by the cell, the gene is then referred to as an oncogene.
The gene encoding Ras, a component of the EGF-initiated pathway, is one ofthe genes most commonly mutated in human tumors. Mammalian cells containthree Ras proteins (H-, K-, and N-Ras), each of which cycles between inactive GDPand active GTP forms. The most-common mutations in tumors lead to a loss of theability to hydrolyze GTP. Thus, the Ras protein is trapped in the “on” position andcontinues to stimulate cell growth, even in the absence of a continuing signal.
Mutated, or overexpressed, receptor tyrosine kinases also are frequentlyobserved in tumors. For instance, the epidermal-growth-factor receptor (EGFR)is overexpressed in some human epithelial cancers, including breast, ovarian, andcolorectal cancer. Because some small amount of the receptor can dimerize andactivate the signaling pathway even without binding to EGF, the overexpressionof the receptor increases the likelihood that a “grow and divide” signal will beinappropriately sent to the cell.
Other genes can contribute to cancer development only when both copies of thegene normally present in a cell are deleted or otherwise damaged.Such genes are calledtumor-suppressor genes. These genes encode proteins that either inhibit cell growth byturning off growth-promoting pathways or trigger the death of tumor cells.For exam-ple, genes for some of the phosphatases that participate in the termination of EGF sig-naling are tumor suppressors. Without any functional phosphatase present, EGFsignaling persists after its initiation, stimulating inappropriate cell growth.
Clinical Insight
Protein Kinase Inhibitors May Be Effective Anticancer DrugsThe widespread occurrence of overactive protein kinases in cancer cells suggeststhat molecules that inhibit these enzymes might act as antitumor agents. Recentresults have dramatically supported this concept. More that 90% of patientswith chronic myelogenous leukemia (CML) have a specific chromosomal defectin affected cells (Figure 12.23). The translocation of genetic material between
18712.5 Signaling Pathway Defects
bcr-abl gene
Chromosome9
Chromosome22
c-abl gene
bcr gene
Translocation
Figure 12.23 The formation of the bcr-ablgene by translocation. In chronicmyelogenous leukemia, parts ofchromosomes 9 and 22 are reciprocallyexchanged, causing the bcr and abl genesto fuse. The protein kinase encoded by thebcr-abl gene is expressed at higher levelsin cells having this translocation than isthe c-abl gene in normal cells.
chromosomes 9 and 22 causes the c-abl (“c” for cellular) gene, which encodes atyrosine kinase, to be inserted into the bcr gene on chromosome 22. The resultis the production of a fusion protein called Bcr-Abl that consists primarily ofsequences for the c-Abl kinase. However, the bcr-abl gene is expressed at higherlevels than the gene encoding the normal c-Abl kinase, leading to an excess of sig-nals for cell growth. In addition, the Bcr-Abl protein may have regulatory proper-ties that are subtly different from those of the c-Abl kinase itself. Thus, leukemiacells express a unique target for drugs. Recent clinical trials of a specific inhibitorof the Bcr-Abl kinase, imatinib mesylate (called Gleevec commercially), haveshown dramatic results; more than 90% of patients responded well to the treatment. Thus, our understanding of signal-transduction pathways is leading toconceptually new disease treatments. ■
SUMMARY
12.1 Signal Transduction Depends on Molecular CircuitsMost signal-transduction pathways are constructed with a similar set ofcomponents. A primary messenger, such as a hormone, binds to the extra-cellular part of a membrane-bound receptor. The messenger–receptor com-plex generates a second messenger inside the cell, which activates proteinsthat alter the biochemical environment inside the cell. Finally, means existto terminate the signal-transduction pathway.
12.2 Receptor Proteins Transmit Information Into the CellSeven-transmembrane-helix receptors operate in conjunction with het-erotrimeric G proteins. The binding of a hormone to a 7TM receptor trig-gers the exchange of GTP for GDP bound to the � subunit of the G protein.G� proteins can transmit information in a number of ways. G�s-GTP acti-vates adenylate cyclase, an integral membrane protein that catalyzes thesynthesis of cAMP. Cyclic AMP then activates protein kinase A by bindingto its regulatory subunit, thus unleashing its catalytic chains. 7TM recep-tors activate G�q proteins and the phosphoinositide pathway. The receptor-triggered activation of phospholipase C generates two intracellularmessengers by hydrolysis of phosphatidylinositol 4,5-bisphosphate. Inosi-tol trisphosphate opens calcium channels in the endoplasmic and sar-coplasmic reticulum membranes. Diacylglycerol activates protein kinase C.Some ligands induce dimerization of the receptors to which they bind. Sucha receptor contains an extracellular domain that binds the ligand, a trans-membrane region, and a cytoplasmic domain that either binds a proteinkinase or is a protein kinase. The growth-hormone receptor participates inan example of this type of signal-transduction pathway. Dimerization of thereceptor activates Janus kinase 2, a protein kinase associated with the intra-cellular part of the receptor.
Intrinsic tyrosine kinases are covalently incorporated in the intracellu-lar domains of some receptors, such as the epidermal-growth-factor recep-tor and the insulin receptor. When such receptor tyrosine kinases dimerizeand are activated, cross-phosphorylation takes place. The phosphorylatedtyrosines in activated receptor tyrosine kinases serve as docking sites for sig-naling proteins and permit further propagation of the signal. A prominentcomponent of such pathways is the small GTPase Ras. The Ras protein, likethe G� subunit, cycles between an inactive form bound to GDP and anactive form bound to GTP.
12.3 Metabolism in Context: Insulin Signaling Regulates MetabolismThe hormone insulin is secreted when blood levels of glucose are high.Insulin binds to the insulin receptor, which is a receptor tyrosine kinase.
18812 Signal-Transduction Pathways
The activated tyrosine kinase then phosphorylates insulin-receptor sub-strate. The signaling pathway continues, with the key result being anincrease in glucose transporters in the cell membrane.
12.4 Calcium Ion Is a Ubiquitous Cytoplasmic MessengerCalcium ion acts by binding to calmodulin and other calcium sensors.Calmodulin contains four calcium-binding modules called EF hands thatrecur in other proteins. Ca2�–calmodulin activates target proteins by bind-ing to positively charged amphipathic helices.
12.5 Defects in Signaling Pathways Can Lead to DiseaseIf the genes encoding components of the signal-transduction pathways arealtered by mutation, pathological conditions, most notably cancer, mayresult. In their mutated form, these genes are called oncogenes. The normalcounterparts are called proto-oncogenes and function in pathways thatcontrol cell growth and replication. Mutated versions of ras are frequentlyfound in human cancers.
adenylate cyclase (p. 177)protein kinase A (PKA) (p. 177)phosphoinositide cascade (p. 180)phospholipase C (p. 180)protein kinase C (PKC) (p. 181)receptor tyrosine kinase (RTK)
(p. 183)Ras (p. 183)calmodulin (CaM) (p. 186)EF hand (p. 186)
calmodulin-dependent protein(CaM) kinase (p. 187)
oncogene (p. 187)proto-oncogene (p. 187)
Answers to QUICK QUIZZES
1. Dissociation of epinephrine from the receptor. Conver-sion of cAMP into AMP by phosphodiesterase and the sub-sequent inhibition of PKA. Conversion GTP into GDP byG� and the subsequent reformation of the inactive het-erotrimeric G protein.
2. Insulin signifies the fed state. Its presence leads to the removal of glucose from the blood for storage ormetabolism. Increasing the number of glucose trans-porters available makes these biochemical processes moreefficient.
1. A reappearance. Ligand-gated channels can be thoughtof as receptors. Explain.
2. Magnification. Explain how a small number of hormonesbinding to the extracellular surface of a cell can have a largebiochemical effect inside the cell.
3. On–off. Why is the GTPase activity of G proteins cru-cial to the proper functioning of a cell?
4. Specificity. Hormones affect the biochemistry of a dis-tinct set of tissues. What accounts for the specificity of hor-mone action?
5. Making connections. Suppose that you were investigatinga newly discovered growth-factor signal-transduction path-way. You found that, if you added a GTP analog in which theterminal phosphate was replaced by sulfate, the duration ofthe hormonal response was increased. What can youconclude?
6. Viva la différence. Why is the fact that a monomeric hor-mone binds to two identical receptor molecules, thus pro-moting the formation of a dimer of the receptor, consideredremarkable?
Problems
7. Chimeras. In an elegant experiment on the nature ofreceptor tyrosine kinase signaling, a gene was synthesized thatencoded a chimeric receptor—the extracellular part camefrom the insulin receptor, and the membrane-spanning andcytoplasmic parts came from the EGF receptor. The strikingresult was that the binding of insulin induced tyrosinekinase activity, as evidenced by rapid autophosphorylation.What does this result tell you about the signaling mecha-nisms of the EGF and insulin receptors?
8. Active mutants. Some protein kinases are inactive unlessthey are phosphorylated on key serine or threonineresidues. In some cases, active enzymes can be generated bymutating these serine or threonine residues to glutamate.Propose an explanation.
9. Antibodies mimicking hormones. An antibody has twoidentical antigen-binding sites. Remarkably, antibodies tothe extracellular parts of growth-factor receptors often leadto the same cellular effects as does exposure to growth fac-tors. Explain this observation.
10. Facile exchange. A mutated form of the � subunit of theheterotrimeric G protein has been identified; this formreadily exchanges nucleotides even in the absence of an acti-vated receptor. What would be the effect on a signalingpathway containing the mutated � subunit?
11. Diffusion rates. Normally, rates of diffusion varyinversely with molecular weights; so smaller molecules dif-fuse faster than do larger ones. In cells, however, calcium iondiffuses more slowly than does cAMP. Propose a possibleexplanation.
12. Awash with glucose. Glucose is mobilized for ATP gen-eration in muscle in response to epinephrine, which acti-vates G�s. Cyclic AMP phosphodiesterase is an enzyme thatconverts cAMP into AMP. How would inhibitors of cAMPphosphodiesterase affect glucose mobilization in muscle?
13. Many defects. Considerable effort has been directedtoward determining the genes in which sequence variationcontributes to the development of type 2 diabetes, a diseasethat results from a loss of sensitivity of cells to insulin.Approximately 800 genes have been implicated. Propose anexplanation for this observation.
14. Growth-factor signaling. Human growth hormone bindsto a cell-surface membrane protein that is not a receptortyrosine kinase. The intracellular domain of the receptor canbind other proteins inside the cell. Furthermore, studiesindicate that the receptor is monomeric in the absence ofhormone but dimerizes on hormone binding. Propose apossible mechanism for growth-hormone signaling.
15. Total amplification. Suppose that each �-adrenergicreceptor bound to epinephrine converts 100 molecules ofG�s into their GTP forms and that each molecule of acti-
vated adenylate cyclase produces 1000 molecules of cAMPper second. With the assumption of a full response, howmany molecules of cAMP will be produced in 1 s after theformation of a single complex between epinephrine and the�-adrenergic receptor?
Data Interpretation Problems
16. Establishing specificity. You wish to determine the hormone-binding specificity of a newly identified membranereceptor. Three different hormones, X, Y, and Z, were mixedwith the receptor in separate experiments, and the percentageof binding capacity of the receptor was determined as a func-tion of hormone concentration, as shown in graph A.
Bin
ding
to r
ecep
tor
as a
perc
enta
ge o
f the
max
imum
Hormone concentration (M)10−210−6 10−410−8
Y
Z
X
(A)
100
80
60
40
20
(a) What concentrations of each hormone yield 50%maximal binding?(b) Which hormone shows the highest binding affinity forthe receptor?
You next wish to determine whether the hormone–receptor complex stimulates the adenylate cyclase cascade. Todo so, you measure adenylate cyclase activity as a function ofhormone concentration, as shown in graph B.
Stim
ulat
ion
of a
deny
late
cyc
lase
as a
per
cent
age
of m
axim
um
Hormone concentration (M)10−210−6 10−410−8
Y
Z
X(B)
100
80
60
40
20
(c) What is the relation between the binding affinity ofthe hormone–receptor complex and the ability of the hor-mone to enhance adenylate cyclase activity? What can youconclude about the mechanism of action of thehormone–receptor complex?
190 12 Signal-Transduction Pathways
(d) Suggest experiments that would determine whether aG�s protein is a component of the signal-transductionpathway.
17. Binding matters. A scientist wishes to determine thenumber of receptors specific for a ligand X, which he has inboth radioactive and nonradioactive form. In one experi-ment, he adds increasing amounts of the radioactive X and
measures how much of it is bound to the cells. The result isshown as total activity in the adjoining graph. Next, heperforms the same experiment, except that he includes aseveral hundredfold excess of nonradioactive X. This resultis shown as nonspecific binding. The difference between thetwo curves is the specific binding.
(a) Why is the total binding not an accurate representationof the number of receptors on the cell surface?(b) What is the purpose of performing the experiment inthe presence of excess nonradioactive ligand?(c) What is the significance of the fact that specific bindingattains a plateau?
18. Counting receptors. With the use of experiments such asthose described in problems 16 and 17, it is possible to cal-culate the number of receptors in the cell membrane. Sup-pose that the specific activity of the ligand is 1012 cpm permillimole and that the maximal specific binding is 104 cpmper milligram of membrane protein. There are 1010 cells permilligram of membrane protein. Assume that one ligandbinds per receptor. Calculate the number of receptor mole-cules present per cell.
Total binding
Specific binding
Nonspecific binding[Lig
and
boun
d]
[Ligand]
Problems 191
Selected readings for this chapter can be found online at www.whfreeman.com/Tymoczko






























![[VII]. Regulation of Gene Expression Via Signal Transduction Reading List VII: Signal transduction Signal transduction in biological systems.](https://static.documents.pub/doc/80x56/56649e385503460f94b28319/vii-regulation-of-gene-expression-via-signal-transduction-reading-list-vii.jpg)




