Br. J. Cancer (1976) 33, 27 STUDIES ON THE STIMULATION AND SUPPRESSION OF DEOXYRIBONUCLEIC ACID (DNA) SYNTHESIS IN LYMPH NODE CELLS OF MICE BEARING PROGRESSIVELY GROWING TUMOURS K. D. CHANDRADASA AND J. BRADLEY From the Sub-Department of Immunology, The Univer8ity of Liverpool, Liverpool L69 3BX Received 24 July 1975 Accepted 19 September 1975 Summary.-Host responsiveness to a progressively growing methylcholanthrene (MC) induced tumour (MC6/2) was studied at varying intervals following subcuta- neous (s.c.) tumour implantation by monitoring the in vitro incorporation of tritiated thymidine (3H-TdR) into lymph node cells (LNC) undergoing stimulation in vivo and concurrently determining the total numbers of the lymphoid cells present in these organs at each of the time intervals. It was found that an initial period of rapidly increasing stimulation of DNA synthesis in lymph nodes was soon followed by the onset of a stage of decrease of this activity. Within limits, the larger the tumour inoculum the stronger the initial response. The suppression of stimulation of DNA synthesis that ensued appeared to be directly related to the tumour mass and to the dose of tumour cells implanted. The total numbers of the cells accumulating in nodes also increased initially but remained elevated during the subsequent period of tumour growth. Continued presence of the tumour was essential for the increased DNA synthesis in lymph nodes since tumour removal leads to a rapid decrease to levels found in tumour-free animals. These findings demonstrate that the failure to eradicate an antigenic tumour by its host may not be solely due to " desensitizing" and " blocking " factors but that other important mechanisms are also involved. We suggest that the inability to reject the tumour in this situation is dependent in considerable measure on the development of a state of hyporeactivity in the host due to the partial inhibition of the DNA synthetic response, possibly in T cells of the tumour host, due to " suppressor factor(s) " interacting with the immunocompetent cells. EVIDENCE available at present indi- cates that " blocking mechanisms " (Hellstrom and Hellstrom, 1969; Hellstrom et al., 1969; Sjogren et al., 1971; Baldwin, Price and Robins, 1972) play a considerable role in the depression of immunity developing in the tumour bear- ing individual at the effector or at the target cell level. On the other hand, inappropriate or inadequate stimulation of the tumour host is implicated in the works of Evans et al. (1962) Haddow and Alexander (1964) and Haddow (1965). A number of investigators have reported that the large pyroninophilic cell response in the paracortex of draining lymph nodes of tumour implanted animals is rapidly exhausted during tumour growth (Rosenau and Moon, 1966; Alexander et al., 1969; Edwards et alt., 1971; Chandradasa, 1973). In certain tumour systems enhancing or blocking activity could not be detected in the serum either in vivo (Vaage, 1972) or in vitro (Deckers et al., 1973). Evidence was presented in a previous communica- tion (Chandradasa, 1973) that concomitant immunity developed in the host is sub- jected to a rapid and specific suppression during the progressive growth of tumours. In transfer studies it was also observed that the specifically cytotoxic lymphoid cells developing in the lymphoid organs of tumour implanted mice decreased either in proportion or in effectiveness during
Transcript
Br. J. Cancer (1976) 33, 27
STUDIES ON THE STIMULATION AND SUPPRESSION OFDEOXYRIBONUCLEIC ACID (DNA) SYNTHESIS IN LYMPH NODECELLS OF MICE BEARING PROGRESSIVELY GROWING TUMOURS
K. D. CHANDRADASA AND J. BRADLEY
From the Sub-Department of Immunology, The Univer8ity of Liverpool, Liverpool L69 3BX
Received 24 July 1975 Accepted 19 September 1975
Summary.-Host responsiveness to a progressively growing methylcholanthrene(MC) induced tumour (MC6/2) was studied at varying intervals following subcuta-neous (s.c.) tumour implantation by monitoring the in vitro incorporation of tritiatedthymidine (3H-TdR) into lymph node cells (LNC) undergoing stimulation in vivo andconcurrently determining the total numbers of the lymphoid cells present in theseorgans at each of the time intervals. It was found that an initial period of rapidlyincreasing stimulation of DNA synthesis in lymph nodes was soon followed by theonset of a stage of decrease of this activity. Within limits, the larger the tumourinoculum the stronger the initial response. The suppression of stimulation of DNAsynthesis that ensued appeared to be directly related to the tumour mass and to thedose of tumour cells implanted. The total numbers of the cells accumulating innodes also increased initially but remained elevated during the subsequent periodof tumour growth. Continued presence of the tumour was essential for the increasedDNA synthesis in lymph nodes since tumour removal leads to a rapid decrease tolevels found in tumour-free animals. These findings demonstrate that the failure toeradicate an antigenic tumour by its host may not be solely due to " desensitizing"and " blocking " factors but that other important mechanisms are also involved.
We suggest that the inability to reject the tumour in this situation is dependent inconsiderable measure on the development of a state of hyporeactivity in the host dueto the partial inhibition of the DNA synthetic response, possibly in T cells of thetumour host, due to " suppressor factor(s) " interacting with the immunocompetentcells.
EVIDENCE available at present indi-cates that " blocking mechanisms "(Hellstrom and Hellstrom, 1969;Hellstrom et al., 1969; Sjogren et al., 1971;Baldwin, Price and Robins, 1972) play aconsiderable role in the depression ofimmunity developing in the tumour bear-ing individual at the effector or at thetarget cell level. On the other hand,inappropriate or inadequate stimulationof the tumour host is implicated in theworks of Evans et al. (1962) Haddow andAlexander (1964) and Haddow (1965). Anumber ofinvestigators have reported thatthe large pyroninophilic cell response inthe paracortex of draining lymph nodes oftumour implanted animals is rapidly
exhausted during tumour growth (Rosenauand Moon, 1966; Alexander et al., 1969;Edwards et alt., 1971; Chandradasa, 1973).In certain tumour systems enhancing orblocking activity could not be detected inthe serum either in vivo (Vaage, 1972) orin vitro (Deckers et al., 1973). Evidencewas presented in a previous communica-tion (Chandradasa, 1973) that concomitantimmunity developed in the host is sub-jected to a rapid and specific suppressionduring the progressive growth of tumours.In transfer studies it was also observedthat the specifically cytotoxic lymphoidcells developing in the lymphoid organs oftumour implanted mice decreased eitherin proportion or in effectiveness during
K. D. CHANDRADASA AND J. BRADLEY
tumour bearing. These findings havebeen supported by the observations of anumber of other workers (Vaage, 1973;Deckers et al., 1973; Whitney, Levy andSmith, 1974). Both Deckers et al. (1973)as well as Whitney et al. (1974) haveprovided evidence from in vitro studiesthat progressive growth oftumours beyonda certain early stage results in a loss ordecline of the cell mediated immunity ofthe tumour host. These experimentswere conducted under conditions where"blocking factors " were considered un-likely to intervene and hence add supportfor an afferent or a central block of theimmune response.
We present evidence in this paper forthe development of a state of hypo-reactivity to the tumour within the lymphnode cell population of the host duringtumour bearing.
MATERIALS AND METHODS
Mice.-Inbred mice of the Balb/c strain,as previously described (Chandradasa, 1973)were used in all experiments performed inthis work.
Tumour.-Tumour MC6/2 used in thepresent studies is a tumour line derived fromthe MC6 described previously (Chandradasa,1973) by the conversion of that tumour to theascitic form after the 15th transplantgeneration. Thereafter it was propagatedintraperitoneally under aseptic conditions.The MC6/2, like its parental form, induced ahigh level of concomitant immunity durings.c. growth enabling the rejection of achallenge tumour dose of 2 x 106 cells by16/17 mice, while the minimum overthresholddose of 2 x 104 cells gave rise to progressivelygrowing tumours in 100% of the normal ani-mals. The tumour utilized in these studies wastaken only from the 10th-36th transplantgenerations passaged in the ascitic form.
In vivo techniques of tumour transplanta-tion and tumour challenge.-The tumour cellsobtained in the ascitic form were washedtwice in Eagle's Minimum Essential Medium(MEM) containing 100 jig streptomycin and100 i.u. benzylpenicillin per ml, resuspendedin the same medium and the required numberof viable cells were inoculated subcutaneously
onto the middle area of the flank, underaseptic conditions.
The cell inocula used in the followingexperiments, viz. 2 x 104, 105, 5 x 105and 107 all gave rise to progressively growingtumours. The survival of mice bearingsuch tumours was restricted to a period ofbetween 30 and 55 days, the life expectancybeing longer with a smaller size of tumourinoculum.
Surgical excision of tumours -Tumourexcision under aseptic conditions was per-formed under ether anaesthesia. The edgesof the skin were sutured using surgical silkand the wound covered with collodion.Wounds healed completely within 2 weeks.
Determination of the weights of tumours.-Tumours were carefully freed from tissues ofhost origin. They were blotted with severallayers of " Kleenex " tissue paper 3 times beforebeing weighed to the nearest mg.
Preparation of lymph node cell suspensions.-Ipsilateral and contralateral axillary andinguinal lymph nodes were harvested underaseptic conditions into RPMI 1640 mediumand were washed twice in the same medium.They were gently teased apart using finelypointed forceps in RPMI 1640 and the cellswere passed through a 160 mesh stainlesssteel gauze. Cell suspensions were washedonce at 250 g for 7 min and the sedimentedcells were resuspended in 2 ml RPMI 1640containing 10% heat inactivated foetal calfserum (FCS) plus 100 ,ug streptomycin and100 i.u. benzylpenicillin per ml (RPMI/FCS).The cell viability and total viable cells/mlwere determined as for tumour cells. TheLNC suspensions thus prepared were used forthe incorporation of tritiated thymidine.
To determine the total number of mono-nuclear cells present in the lymph nodes,whole nodes were first teased apart inphosphate buffered saline (PBS) using finelypointed forceps. The liberated cells werefirst harvested and the remaining tissuepieces were gently ground in Griffiths tubes(Baird and Tatlock Ltd, Manchester, Eng-land) using 5 ml of PBS until all the cells hadbeen liberated. The cells were washed in 5 mlPBS, spinning at 250 g for 10 min. The cellsediment was resuspended in a suitablevolume of PBS and the cell counts wereperformed in leucocyte diluting fluid in aNeubauer haemacytometer. The total cellsobtained from lymph nodes or similarlytreated spleens were then calculated.
2'S
STIMULATION AND SUPPRESSION OF DNA IN MICE
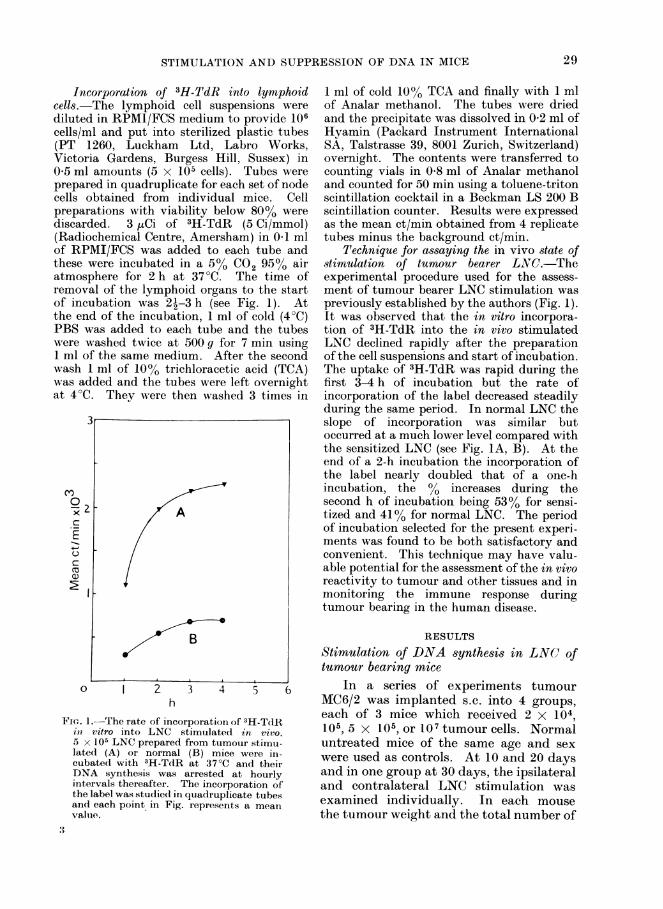
Incorporation of 3H-TdR into lyrnphoidcells.-The lymphoid cell suspensions werediluted in RPMI/FCS medium to provide 106cells/ml and put into sterilized plastic tubes(PT 1260, Luckham Ltd, Labro Works,Victoria Gardens, Burgess Hill, Sussex) in0 5 ml amounts (5 x 105 cells). Tubes wereprepared in quadruplicate for each set of nodecells obtained from individual mice. Cellpreparations with viability below 80% werediscarded. 3 ,uCi of 3H-TdR (5 Ci/mmol)(Radiochemical Centre, Amersham) in 0-1 mlof RPMI/FCS was added to each tube andthese were incubated in a 50 CO2 950 airatmosphere for 2 h at 37°C. The time ofremoval of the lymphoid organs to the startof incubation was 21-3 h (see Fig. 1). Atthe end of the incubation, 1 ml of cold (4°C)PBS was added to each tube and the tubeswere washed twice at 500 g for 7 min using1 ml of the same medium. After the secondwash 1 ml of 10% trichloracetic acid (TCA)was added and the tubes were left overnightat 4°C. They wvere then washed 3 times in
Ox '
.c
Ea,
Fic
II
t]a
v
1 ml of cold 10% TCA and finally with 1 mlof Analar methanol. The tubes were driedand the precipitate was dissolved in 0-2 ml ofHyamin (Packard Instrument InternationalSA, Talstrasse 39, 8001 Zurich, Switzerland)overnight. The contents were transferred tocounting vials in 0-8 ml of Analar methanoland counted for 50 min using a toluene-tritonscintillation cocktail in a Beckman LS 200 Bscintillation counter. Results were expressedas the mean ct/min obtained from 4 replicatetubes minus the background ct/min.
Technique for assaying the in vivo state ofstimulation of tumour bearer LNC.-Theexperimental procedure used for the assess-ment of tumour bearer LNC stimulation waspreviously established by the authors (Fig. 1).It was observed that the in vitro incorpora-tion of 3H-TdR into the in vivo stimulatedLNC declined rapidly after the preparationof the cell suspensions and start of incubation.The uptake of 3H-TdR was rapid during thefirst 3-4 h of incubation but the rate ofincorporation of the label decreased steadilyduring the same period. In normal LNC theslope of incorporation was similar butoccurred at a much lower level compared withthe sensitized LNC (see Fig. lA, B). At theend of a 2-h incubation the incorporation ofthe label nearly doubled that of a one-hincubation, the 00 increases during thesecond h of incubation being 53% for sensi-tized and 410% for normal LNC. The periodof incubation selected for the present experi-ments was found to be both satisfactory andconvenient. This technique may have valu-able potential for the assessment of the in vivoreactivity to tumour and other tissues and inmonitoring the immune response duringtumour bearing in the human disease.
RESULTS
Stimulation of DNA synthesis in LNC oftumour bearing mice
o 1 2 3 4 5) 6 Ln a series ot experiments tumourh MC6/2 was implanted s.c. into 4 groups,
each of 3 mice which received 2 x 1041. The rate of incorporation of 3H-TdRo vitro into LNC stimulated in vivo. 105, 5 X 105, or 107 tumour cells. Normal; x 105 LNC prepared from tumour stimu- untreated mice of the same age and sexated (A) or normal (B) mice were in- were used as controls. At 10 and 20 days-ubated with 3H-TdR at 37°C and their)NA synthesis was arrested at hourly and in one group at 30 days, the ipsilateralntervals thereafter. The incorporation of and contralateral LNC stimulation was'he label was studied in quadruplicate tubes examined individually. In each mousend each point in Fig. represents a meani'alue. the tumour weight and the total number of
29
T- - -- -- _r J- A.--
K. D. CHANDRADASA AND J. BRADLEY
cells present in the spleen and in theipsilateral and contralateral nodes werecarefully determined.
The results of these experiments areshown in Table I and in Fig. 2A, B andFig. 3. It is obvious that the ct/min/5 x 105 cells increase, but this incrementdoes not reflect the total activity of thelymph nodes because the total cells innodes also increase in number. Thereforethe ct/min/LNC have been multiplied bythe total cell content to give the totalct/min/set of nodes.
The incorporation of 3H-TdR has beenexpressed as: (1) the ct/min obtained byincubating 5 x 105 LNC (proportional
l0Days
L0 0 lo LO
post tumour implantation
incorporation): (2) as a total ct/minobtained by calculating the ct/min for thetotal number of cells in the ipsi or contra-lateral nodes, providing a measure of thetotal stimulation: (3) as an index ofrelative stimulation obtained by dividingthe ct/min/5 x 105 LNC in the test nodesby the equivalent of a normal controlgroup; and (4) as an index of totalstimulation which is the value obtained bydividing the total ct/min in test nodes bythat of the normal controls.
The results show that in the differentgroups of mice implanted with tumours,the incorporation of 3H-TdR into 5 x 105ipsilateral LNC during the first 10 days
c0.Qa)
- E_ 4--
0
a)
L-
o
, oQ
-oa)C:
-c
4)
-C
.oo
I
a
Days post tumour implantation
FIG. 2A FIG. 2B
FIG. 2A, B.-Stimulation of DNA synthesis in lymph nodes of Balb/c mice bearing MC6/2 tumour ata s.c. site. 2 x 104, 105, 5 x 105 or 107 tumour cells were implanted s.c. at Day 0 and at Days 10and 20 incorporation of 3H-TdR by 5 x 105 ipsilateral or contralateral LNC were studied in vitroin replicates of 4 tubes. Incorporation of the label into (5 x 105) LNC from normal mice was
studied simultaneously as controls. Fig. 2A indicates the uptake of 3H-TdR by 5 x 105 LNC atthe two-time intervals. Fig. 2B shows the index of relative stimulation which is the mean incor-poration in 5 x 105 test LNC/corresponding value in normal LNC. Each column represents a
mean value obtained from a group of 3 mice and the lines on top of columns show the s.e. mean.DNA synthesis (2A) and the index of relative stimulation (2B) in Fig. are indicated as follows: clearcolumns, ipsilateral LNC: dotted columns, contralateral LNC. Hatched columns show the DNAsynthesis in normal LNC. The number of tumour cells implanted at Day 0, are indicated by simpleletters, a = 107, b = 5 x 105, C = 105 and d = 2 x 104, and the changes in mean tumour weight* 0) are shown. S.e., Standard error.
m0
x
C-
7-6 a
5
2
30
0
31STIMULATION AND SUPPRESSION OF DNA IN MICE
t- 8 Yo G m co to co co 00
4-4 -H VH -H H -H -H -f -H -H
lGo11--r 6Z '3 0 00Ct 00eD to _I tCO00-4nd
.>d H -H-- -H r--HNC°<<X XON-f, H,c3dL o lco:locOO:
c3 t-1LgO 00O eD Cq 0X ° 0 0 CZ
d 00 N to tz >> °oLO 04 C* gl "d<tb 0
*-IfIH -HC3 _-HN~O O C N£ C-H -HoHO4s++{+-c
o Stt Cd D00 M co 00 o Lo 0 0 00 00 co,- °* ) -q m8 l qmcc=cq czo oul~uc co0 aZ *
.65 -4
-H E-°V *4 c.5 o: Oc o cs~omc"-4 = m c 8olde | 4 t; r4 ¢r- c, 0 0 m N coU- -I -, cq Ml t 40 3;
W to V t-M)t+ 0 04"°to "~C to -4 a 3co
:3| -$e c)oO eX><>r.. dX.9 4 Qo N -Hm -Hc4 H HH `-nH0-H H -°L ; + +++++>>> =d
4-l c3d,-Hj Hc3L4°>OQ
d.o".) dce o qc e
X zz_ 9~~~~~~~~~~s0=0
cec o-Ia qt Lod c t- toONeC oc Loco ° °>x oc- q0-°r-.a3*tcesti oC 5t oo -I N CO v 0 CS 4..
+-lL -H -H H -H aq -H m -Hc -H " -H " -H mt-HX. agJd'tn
Le~~~~~~~~~~~~~~~~;- 4)c38
A~~~~~~~~~~~~~~~co ;Z<Cs
'O X.5O.H md H
o~~~~~~- 0x Cx Lo ",- 0 =-tn~~~~~~~~~~~~~~~~ 8.cq COe
aO
co
* Qs
4':.5
4c',
otQ
C.a)X
K. D. CHANDRADASA AND J. BRADLEY
4
3
21
0
E
Z3
xE42
U) lo 2o 0 lo 20Days post tumour implantation
I
4--,c*cc
L-
°Ec3
Fie. 3. Changes in iindex of total stimulationof the LNC in tumour bearing mice. Theindex of total stimulation = total ct/min intest lymph nodes/total ct/min in controllymph nodes. (These values are shown inTable I.) Clear columns, ipsilateral nodes:(lotted columns, contralateral nodes. Eachcolumn represents a mean value obtaine(dfrom a group of 3 mice. The s.e. meain isindicated by the lines on top of the columnsand the changes in mean tumour weight(0* 0) and the number of tumouicells implanted at Day 0, a = 101, b = 5
X 105, C= 105 and d = 2 x 104 are also
shown. S.e. mean-- Standard error of themean.
increased 4-6 fold whereas the total ct/minfor the nodes increased 10-36 fold over thecorresponding values in normal controlmice. Stimulation of DNA synthesis inLNC reached peak levels at 10 days, withmice implanted with the larger doses of5 x 105 or 107 tumour cells incorporatingmarkedly more 3H-TdR in LNC than thegroups implanted with the smaller doses-this effect being more marked in the totalct/min and in the indices of total ct/minthan in the ct/min/5 x 105 LNC values.There is, however, a limit to furtherstimulation by increasing the initialtumour inoculum since the group im-
planted with 107 tumour cells incorporatedless 3H-TdR by the ipsilateral LNC com-
pared with the mice implanted witha x 105 tumour cells. The difference in thect/min/5 x 105 LNC in the 2 groups at10 days was found to be significant at the500 level (0.02 < P < 0.05) and the differ-ences in the corresponding indices ofrelative stimulation (0.001 < P < 0.01)or total stimulation (P < 0-001) were
highly significant.The total numbers of the LNC present
in the ipsilateral and contralateral nodeswere found to be increased in the differentgroups of mice at 10 days, in directrelationship to the number of tumour cellsimplanted up to 5 x 1_05. A similarrelationship was observed with the in-crease of the cells present in spleens, up tothe largest tumour cell dose of 107employed.
At 20 days, mice implanted withx 105 or 107 tumour cells showed a
rapid decline of their DNA syntheticresponse to tumour in the LNC. Thedecrease in the incorporation of 3H-TdRinto 5 x 105 LNC or into the total cellpopulation of the ipsilateral nodes of thefirst category of mice bore a high levelof significance (P < 0.001). The factthat the normal control LNC at 20days also had decreased DNA synthesiscould leave the possibility that in testLNC the fall of DNA synthesis may
have been exaggerated, but the testLNC activity, when related to the per-
formance in normal LNC (see relativeindex of stimulation, Table 1) still showedsignificance (0.01 < P < 0.02). Theircontralateral LNC also showed a signi-ficant decrease of 3H-TdR incorporationboth proportionally (P < 0.01) as well as
totally (0.01 < P < 0.02). Similarchanges were observed in the ipsilateralLNC of mice implanted with 107 tumourcells with significant decreases in propor-
tional (0.001 < P < 0.002) as well as intotal incorporation of 3H-TdR (0.01 < P< 0.02), although no significant changewas noted in the contralateral node cells.These observations were made during, a
-40 ~~~~~~~3
-2
.0
lo- ~ ~ ~
0 d~~~~~~~~~~~~1
32
"'.
I1
STIMULATION AND SUPPRESSION OF DNA IN MICE
period when the tumour weight increasedmore than three-fold in the group im-planted with 5 x 105 tumour cells ornearly twice as in the group implantedwith 107 cells.
Mice which received smaller doses of2 x 104 or 105 tumour cells provided adifferent picture. In the first category ofmice no significant change in either thect/min/5 x 105 cells or in the indices ofstimulation of LNC was noted between 10and 20 days, while in terms of totalstimulation the 20-day values of both ipsi-and contralateral LNC were apparentlyelevated, although this failed to reachsignificance at the 50% level (0.05 < P< 0.1). There was a significant (P< 0.001) increase in the accumulatedLNC in both ipsi- and contralaterallymph nodes at 20 days, which no doubtcontributed to the increased total radio-activity. In mice implanted with 105tumour cells, no significant increase ofeither the proportional stimulation or theindices of relative or total stimulation wasnoted but the total stimulation in bothipsi- and contralateral nodes was raisedabove the values at 10 days. This wasfound to be significant (0.01 < P < 0.05).
The tumours in these 2 groups of miceremained below 1 g during this period ofinvestigation, but had more than doubledin weight between 10 and 20 days. It wasalso noted that their spleens had accumu-lated numbers of cells comparable with
that present in other groups at the samestage.
In the group of mice tested at 30 daysin which the tumours were initiated withan inoculum of 2 x 104 tumour cells, thect/min/5 x 105 LNC or the relative indexof stimulation in both ipsi- and contra-lateral nodes did not show a significantdecrease below the corresponding levels at20 days. However, the decreases in theindices of total stimulation were signi-ficant in both the ipsilateral (0.02 < P< 0.05) as well as in the contralateralnodes (0.002 < P < 0.01) as was the totalct/min in the contralateral nodes (0.002 <P < 0.01).
Effect of tumour excision on the uptake of3H-TdR by LNC1
Tumour MC6/2 was implanted s.c. intothe right flank of 14 mice. A third groupof 7 mice was set aside as normal controls.Ten days later the tumours were com-pletely excised surgically from Group I(6 mice) while Group II (8 mice) andGroup III (7 mice) received sham opera-tions. Seven days following these treat-ments the axillary and inguinal lymphnodes were carefully removed from thetumour excised or tumour bearing flanksof the Group I and Group II mice andfrom the sham operated flank of the GroupIII mice. The LNC preparations from the3 groups of mice were then tested for theirability to incorporate 3H-TdR in vitro.
TABLE IT.-Effect of Excision of Tumour on the Tumour Stimulated DNASynthesis in LNC*
Treatmernt,
Group IITumour MC 6/2 s.c.
Group I for 10 days Group IIITuimouir MC 6/2 s.c. sham excisioni Normal controls
for 10 days of tumour sham operationexcision of tumour + 7 days following + 7 days following
7 days free of tumour sham excision sham operationNo. of mice in group 6 8 7ct/min/5 x 105LNC -4- s.e. meant 673-27 X 232-9 1917-37 ± 234-8 866-27 + 43-5
I06MC 6/2 cells were implanted subcutaneously at Day 0 into groups I and II and the tumours wereexcised from group I on Day 10, while the groups II and III were sham operated at the same time. OnDay 17 the ipsilateral LNC of groups I and II and the corresponding LNC from the sham operated flank ofgrotup III wvere tested for their stimulated DNA synthesis by incubating 5 x 105 cells with 3H-TdR.
* LNC = LLymph node cells. t s.e. mean = Standard error of the mean.
33
K. D. CHANDRADASA AND J. BRADLEY
The results of this experiment areshown in Table II. The mice in Group IIbearing tumours incorporated 3H-TdRsignificantly more than those in Group I(P < 0.001) or those in Group III (P< 0-001) while no significant differencewas noted between 3H-TdR incorporationin Groups I and III.
DISCUSSIONThese results (see Table I, Fig. 2A, B
and Fig. 3) indicate the existence of 2different phases of stimulation of DNAsynthesis within the LNC of the tumourbearing mice studied here. During thefirst phase, which lasted about 10 days,the stimulation of DNA synthesis bore,within limits, a direct relation to thetumour mass present. Thus, it can beseen from Table I and Fig. 2A, B that peaklevels of incorporation of 3H-TdR into5 x 105 LNC resulted in all groups ofmice at 10 days. During the secondphase, a marked retardation in the rateof stimulation of LNC was observed. Inthe groups that were implanted with thelarger doses of 5 x 105 or 107 tumour cellsthis led to a significant decrease in thestimulation of LNC at 20 days, whereas inthose that were given the smaller doses of2 x 104 or 105 cells it resulted in acomplete arrest of any further increase inthe rate of stimulation of DNA synthesis.The increase in the total stimulation inthese mice at 20 days resulted from anaccumulation of LNC at this stage.
The specific nature of the response inthe MC6/2 implanted mice is indicated bythe findings on the effect of tumourexcision (Table II). These clearly showthe requirement of the continued presenceof tumour for the maintenance of the DNAsynthetic response in lymph nodes.
These observations may indicate thepresence of a mechanism of suppression orregulation of the immune response to thetumours within the tumour host. Theinitial period of about 10 days duringwhich the rate ofLNC stimulation steadilyincreased may also represent a phase duringwhich the factors responsible for the
controlling influence begin to make theirappearance in the tumour host.
At present, much emphasis has beenfocussed on the blocking effect of anti-body, antigen or their complexes either atthe effector or at the target cell level.Although this is now well established, ithas not always been adequate to explainthe experimental results observed (Deckerset al., 1973; Vanky et al., 1973; Whitney etal., 1974). The possibility that the tumourhost is not responding fully is oftenmasked by the demonstration of its abilityto destroy a limited number of tumourcells when reinoculated and also by thedetection in vitro of specifically cytotoxiclymphoid cells present within its system.
In a previous report on the backgroundDNA synthesis in the spleen cells oftumour bearing mice, Konda, Nakao andSmith (1973) observed that this activityinitially increased but subsequently de-creased with continuing growth of thetumour. However, the absolute valuesfor the whole spleen remained elevatedduring the course of tumour bearing.This study was carried out using the grosspopulation of the spleen cells and thereforecannot be taken as a measure of thesplenic lymphocyte DNA synthesis. Theseworkers also reported a marked increase ofcells in the spleen responding to a varietyof non-tumour antigens and also that thesplenic haematopoietic cells capable ofcolony formation on adoptive transfer wereobserved to be greatly increased duringtumour bearing. Whilst these observa-tions are not contrary to those reportedhere, our findings may gain strong supportfrom the observations of Vanky et at.(1973) who showed that in mixed lympho-cyte-target cell interaction tests, LNCdraining large tumours of long duration inhuman cancer, failed to respond to theautochthonous tumour cells but retainedtheir ability to react to allogeneic lympho-cytes or to phytohaemmaglutinin, indi-cating the specificity of non-responsive-ness.
We have shown that the host's LNCresponse to the tumour becomes depressed
34
STIMULATION AND SUPPRESSION OF DNA IN MICE 35
soon after the early period of tumourgrowth. This may find an interestingcorrelation in the control of the delayedhypersensitivity reaction in mice paintedwith certain skin sensitizing agents (Asher-son and Barnes, 1973; Zembala et al.,1975). It has been shown that pretreat-ment of mice with the picrylating agentpicryl sulphonic acid depresses or abolishesthe DNA synthetic response to the sameagent as well as to picryl chloride in thelymph nodes, but not in the spleen.Delayed hypersensitivity, but not theantibody response, is markedly affected,reflecting on the composition of these 2types of lymphoid organs-the lymphnodes consisting predominantly of " T "cells while the spleen containing nearlyequal proportions of both " T " and " B "cells (Raff, 1971). Although the DNAsynthetic response in the LNC of thetumour bearing mice studied here maynot undergo such dramatic changes, it isnevertheless possible that a similar basismay underlie both these phenomena. Itwould therefore be interesting to see if such" suppressor factor(s) " could be demon-strable in the tumour hosts and if so toidentify and study their immunobiochemi-cal nature.
We wish to thank Miss D. Murray andMr R. Clay for their valuable technicalassistance.
The continued support of the NorthWest Cancer Research Fund is gratefullyacknowledged.
REFERENCESALEXANDER, P., BENSTEAD, J., DELORME, E. J. &HODGETT, J. (1969) The Cellular Immune Responseto Primary Sarcomata in Rats. II. AbnormalResponse of Nodes Draining the Tumour. Proc.R. Soc. B, 174, 237.
AsHERSON, G. L. & BARNES, R. M. R. (1973) ContactSensitivity in the Mouse: XII. The Use of DNASynthesis in vivo to Determine the AnatomicalLocation of Immunological Unresponsiveness toPicryl Chloride. Immunology, 25, 495.
BALDWIN, R. W., PRICE, M. R. & RoBsINs, R. A.(1972) Blocking of Lymphocyte Mediated Cyto-toxicity for Rat Hepatoma Cells by Tumour.specific Antigen-antibody Complexes. Nature,New Biol., 238, 185.
CHANDRADASA, K. D. (1973) Development and
Specific Suppression of Concomitant Immunity intwo Syngeneic Tumour-host Systems. Int. J.Cancer, 11, 648.
DECKERS, P. J., DAVIS, R. C., PARKER, G. A. &MANNICK, J. A. (1973) The Effect of Tumor Sizeon Concomitant Tumor Immunity. Cancer Re8.,33, 33.
EDWARDS, A. J., SUMNER, G. F., ROWLAND, G. F. &HUND, C. M. (1971) Changes in LymphoreticularTissues during Growth of Murine Adenocarcinoma.I. Histology and Weights of Lymph Nodes,SpleenandThymus. J. natn. CancerInst.,47,301.
EVANS, C. A., GORMAN, L. R., ITO, Y. & WEISER,R. S. (1962) Antitumour Immunity in ShopePapilloma Carcinoma Complex of Rabbits. I.Papilloma Regression Induced by Homologous andAutologous Tissue Vaccines. J. natn. CancerInst., 29, 277.
HADDOW, A. (1965) Immunology of the Cancer Cell.Tumour Specific Antigens. Br. med. Bull., 21, 133.
HADDOW, A. & ALEXANDER, P. (1964) An Immuno-logical Method of Increasing the Sensitivity ofPrimary Sarcomas to Local Irradiation withX-rays. Lancet, i, 452.
HELLSTROM, I. & HELLSTROM, K. E. (1969) Studieson the Cellular Immunity and its Serum MediatedInhibition by Moloney-virus Induced MouseSarcoma. Int. J. Cancer, 4, 587.
HELLSTROM, I., HELLSTR6M, K. E., EVANS, C. A.,HEPPNER, G. H., PIERCE, G. E. & YOUNGE,J. P. S. (1969) Serum Mediated Protection ofNeoplastic Cells from Inhibition by LymphocytesImmune to the Tumour Specific Antigens.Proc. natn. Acad. Sci. U.S.A., 62, 362.
KONDA, S., NAKAO, I. & SMITH, R. T. (1973) TheStimulatory Effect of Tumor Bearing upon the Tand B Cell Subpopulations of the Mouse Spleen.Cancer Res., 33, 2247.
RAFF, M. C. (1971) Surface Antigen Markers forDistinguishing T and B Lymphocytes in Mice.Transplantn Rev., 6, 52.
ROSENAU, W. & MOON, H. D. (1966) Cellular Re-action to Methylcholanthrene-induced SarcomasTransplanted to Isogeneic Mice. Lab. Invest., 15,1212.
SJ6GREN, H. O., HELLSTROM, I., BANSAL, S. C. &HELLSTR6M, K. E. (1971) Suggestive Evidence thatthe " Blocking Antibodies " of Tumour BearingIndividuals may be Antigen Antibody Complexes.Proc. natn. Acad. Sci. U.S.A., 68, 1372.
VAAGE, J. (1972) Specific Densensitization ofResistance against a Syngeneic Methylcholan-threne-induced Sarcoma in C3 Hf Mice. CancerRes., 32, 193.
VAAGEE, J. (1973) Influence of Tumor Antigen onMaintenance versus Depression of Tumour-specific Immunity. Cancer Res., 33, 493.
VANKY, F., STJERNSWARD, J., NILSONNE, U. &SUNDBLAD, R. (1973) Differences in the Tumor-associated Reactivity of Blood Lymphocytes andTumor-draining Lymph Node Cells in SarcomaPatients. J. natn. Cancer Inst., 51, 17.
WHITNEY, R. B., LEVY, J. G. & SMITH, A. G.(1974) Influence of Tumor Size and SurgicalResection on Cell-mediated Immunity in Mice.J. natn. Cancer Inst., 53, 111.
ZEMBALA, M., ASHERSON, G. L., MAYHEW, B. &KREJCI, J. (1975) In vitro Absorption andMolecular Weight of Specific T-cell SuppressorFactor. Nature, Lond., 253, 72.