Page 1
CARRYING CAPACITY: A CRITIQUE OF THE CONCEPT A.L~
ITS USEFULNESS
T. Jon Davies
Supervisor: Prof P. A. R. Hockey
Fitzpatrick Institute
University ofCapeTown
Rondebosch 7700
There is considerable confusion surrounding the term carrying capacity in the literature. 1 suggest that
carrying capacity is the upper limit to abundance set by the resources in the environment. Due to temporal
variation in resource availability, carrying capacity should be considered as a dynamic value which
natural populations tend towards unless influenced otherwise by external factors such as predation. It can
be measured in terms of individual numbers, biomass or energy. By adopting units of mass or energy it
may be possible to define composite-species carrying capacities. However, due to. the differing resource
requirements of many species such an approach can be misleading, as the carrying capacity for one
species may not be indicative ofthe capacityfor another.
The concept of carrying capacity has several valuable applications to wild populations. It is particularly
relevant to the present day system of parks and reserves in which adaptive movement of animals is
restricted by fragmentation of the habitat and the construction offences. Estimates of carrying capacity
can be used to ensure that a population does not exceed the ability ofthe habitat to sustain it. The relative
value ofhabitats based on the potential population size they can support may be obtained, aiding reserve
selection processes. Lastly, understanding the mechanisms by which the carrying capacity is set may
enable identification of those populations at or near carrying capacity and those held below carrying
capacity. Such information could be used to aid management and potentially direct artificial manipulation
of the population to increase its density, by either removing a source of mortality or increasing the
resource base.
In partial fulfilment for the degree ofMaster of Science (Conservation Biology), March 1997
1
Page 2
The copyright of this thesis rests with the University of Cape Town. No
quotation from it or information derived from it is to be published
without full acknowledgement of the source. The thesis is to be used
for private study or non-commercial research purposes only.
Univers
ity of
Cap
e Tow
n
Page 3
CONTENTS
Page
1. INTRODUCTION ;................................................................ 3
2. TO WHAT DENSITY DOES CARRYING CAPACITY REFER? 52.1. Subsistence Density ·............................................................................................... 52.2. OptimUIn Density....... 72.3. Tolerance Density ,...................................................................................................... 92.4. Security Density 11
2.4.1. Does a predator-regulated prey equilibrium exist in nature? 11
J. TEMPORAL VARIATION AND CARRYING CAPACITy........................................ 143.1. Seasonal Variatton................................................................................................................... 173.2. AnnualVariation 18
3.2.1. Population cycles....................................................................................................... 183.2.2. Random fluctuations 20
4. WHAT SETS THE CARRYING CAPACITY?........................................................ 224.1. Scramble'Competition. 234.2 Contest Competition 254.3. The Special Case of Space 26
5. ESTIMATING CARRYING CAPACITy...................................................................... 285.1. Estimating Carrying Capacity in Terms of Supportable Animal Units................................... 29
5.1.1. The population density approach.............................................................................. 295.1.2. The biomass approach............................................................................................... 31
5.2. Estimating Carrying Capacity in Terms of Resources............................................................. 325.2.1. Energetics.................................................................................................................. 325.2.2. Nutrition :......................... . 38
5.3. Problems of Open Systems ·.................................................. 395.4. The Construction of Indices Relating Carrying Capacity to Measurable
Environmental Parameters....................................................................................................... 40
6. APPLICATIONS OF THE CONCEPT OF CARRYING CAPACITY TOCONSERVATION RELATED ISSUES......................................................................... 41
6.1. Avoidance of the Problems Associated with Overpopulation.................................................. 426.2. HabItat Evaluation.................................. 446.3. Assessing Population Status..................................................................................................... 466.4. Raising Population Densities 47
7. CONCLUSION ;......... 48
REFERENCES ;.............. 50
APPENDIX A...................................................................................................................... 63
APPENDIX B 67
2
Page 4
1. INTRODUCTION
Giles (1978) called carrying capacity a term "any 'barbershop biologist can use in confident
ignorance". The term has been used in a wide variety of fields including biology, ecology,
anthropology, geography, business management (Budd, 1992) and tourism (Briassoulis, 1992). The
various classifications of carrying capacity in the literature are enough to cause confusion on their
own (e.g. Budd, 1992~ Caughley and Sinclair, 1994~ Dhondt, 1988; Igbozurike, 1981~ Lien, 1993;
Odum, 1983; Zorn and Seelbach, 1995) many of them applied in a contradictory manner, e.g. the
use of "optimum carrying capacity" by Caughley (1979) and Norland et al. (1985). Where
appropriate I have attempted, in the following sections, to group those that apply to similar
biological concepts.
Budd (1992) traces the term to Verhulst in 1838, although he does not specify in what'context it
was applied. The term was first used in an ecological context in 1922 by Hadwen and Palmer(cited
in Dhondt, 1988).who defined it as "the number of stock which a range will support for a definite
period ofgrazing without injury". Thisdefinition indicates that the concept ofcarrying capacity was
first applied to domesticated animals. The term was subsequently used for wildlife and it is this that
causedthe majority of the early confusion surrounding the concept.
Giles (1978) lists a selection of terms that have been equated with carrying capacity in the
biological literature: maximum potential biomass of species i per unit area, maximum potential
biomass production of species i per unit area, current biomass of species i per unit area, maximum
animals or biomass per unit area of species i to achieve a particular objective and ecological
homeostasis (kcal/mvunit time). Further examples include: maximum population (Igbozurike,
1981), absorptive capacity (Igbozurike, 1981), environmental' limit (Igbozurike, 1981), supportive
ability (Igbozurike, 1981), productivity (Igbozurike, 1981), equilibrium population density (Barlow,
1996), maximum sustainable size (Buckland et aI., 1996) and K (Whittaker, 1972). This list is
indicative of the wide use the term carrying capacity has been put to and hence the problems
surrounding its definition (appendix A contains a selection of the various definitions of carrying
3
Page 5
capacity cited in the literature), casting some doubt on the credibility of Budd's (1992) statement
that "(a)pplication of the concept of carrying capacity has a fairly well defined meaning for
biologists and wildlife managers".
The defining ofcarrying capacity must not be approached in a superficial or over-simplistic manner.
Weeden (1974) states the following: "In its traditional biological context, carrying capacity is a
simple abstraction: the ability of an area or ecosystem to absorb stress without disruption of its
basic properties". Budd (1992) also gives a general definition as "the number of individuals that can
be supported in a given area". However, these simple definitions are open to a variety of
interpretations, and are often determined more by the phenomenon being investigated rather than
by any stringent definition of the concept (e.g. Allen and Cormier, 1996). Appendix B gives some
indication ofthe emphasis different authors have placed on the issues surrounding the term.
Dhondt (1988) proposes that the only way around the difficultyin defining the term is either not to
use it at all or to restrict its use to a vague concept. In theoretical ecology, where models refer to an
equilibrium density, the term "upper asymptote" should be used. If applied to wildlife biology, the
resource the population is being managed for must be defined. Moen (1973) also suggests that
carrying capacity is "more of a concept than a straightforward, definable biological relationship".
Begon et al.'s (1990) description of carrying capacity as an "idealised concept not to be taken
literally" echoes these sentiments. Despite these views, the use of the term has spread widely in the
scientific and semi-popular literature. In 1989 it was voted one of the 25 most popular ecological
concepts by the British Ecological Society (Peters, 1993).
This paper is an attempt to clarifywhat should be meant by the term carrying capacity when applied
to ecological issues. An examination of the various definitions <;U1d uses that the term has been put
to will be undertaken. Concentrating on animal populations, I will show that many applications of
this concept, either by design or misunderstanding, do not fall into what I understand the term to
mean. The second part ofthe paper addresses the problems associated with applying the concept of
a carrying capacity to real populations. The final sections will discuss attempts to measure carrying
capacity empirically, including the appropriate units that should be used and where it may be
applicable with particular reference to conservation and predicting impacts ofhabitat loss.
4
Page 6
2. TO WHAT DENSITY DOES CARRYING CAPACITY REFER?
Several measures' ofpopulation density have been equated to canying capacity in the literature (e.g.
Caughleyand Sinclair, 1994; Caughley and Walker, 1983; Giles, 1978; Mott, 1960, cited in Bartels
et aI., 1993). Dasmann (1981) proposes that these can be placed into four groups: "subsistence
density" to describe population densities set by the resource base; "optimum density" referring to
those populations that are managed with a goal in mind; "tolerance density" to describe population
densities limited by social constraints; and "security density" describing population densities limited
by disease, predation or parasitism . The above classifications are discussed briefly in the following
sections, highlighting the differences between them. Equivalent biological terms have been placed in
parenthesis following each subheading.
2.1. Subsistence Density (absolute carrying capacity, maximum canying capacity, K carrying
capacity, potential carrying capacity, environmental carrying capacity, ecological carrying
capacity): -
This subsistence density is so called because it is the potential density at which the resources of the
environment can just sustain the population without any tendency towards either an increase or
decrease in population size. It can be viewed as an equilibrium 1 between birth and death rates (Fig.
1) set by intraspecific competition (Begon et al., 1990). Caughley (1979) simply described carrying
capacity as the "equilibrium between animals and vegetation", although it may be better equated
I There is no implication that this is a stable equilibrium.
5
Page 7
with the equilibrium between a consumer and its resource base. Odum (1983) sees this density as
an energetic balance between the inputs and outputs of a system. At the subsistence density, energy
maintenance costs match the available energy (Odum, 1983), i.e. there is no energy available for an
increase in the population. I have taken this density to be representative of the canying capacity.
This definition assumes that density is resource limited and that the population does not degrade its
own resource base (Odum, 1971), i.e. the subsistence density is a sustainable population size and it
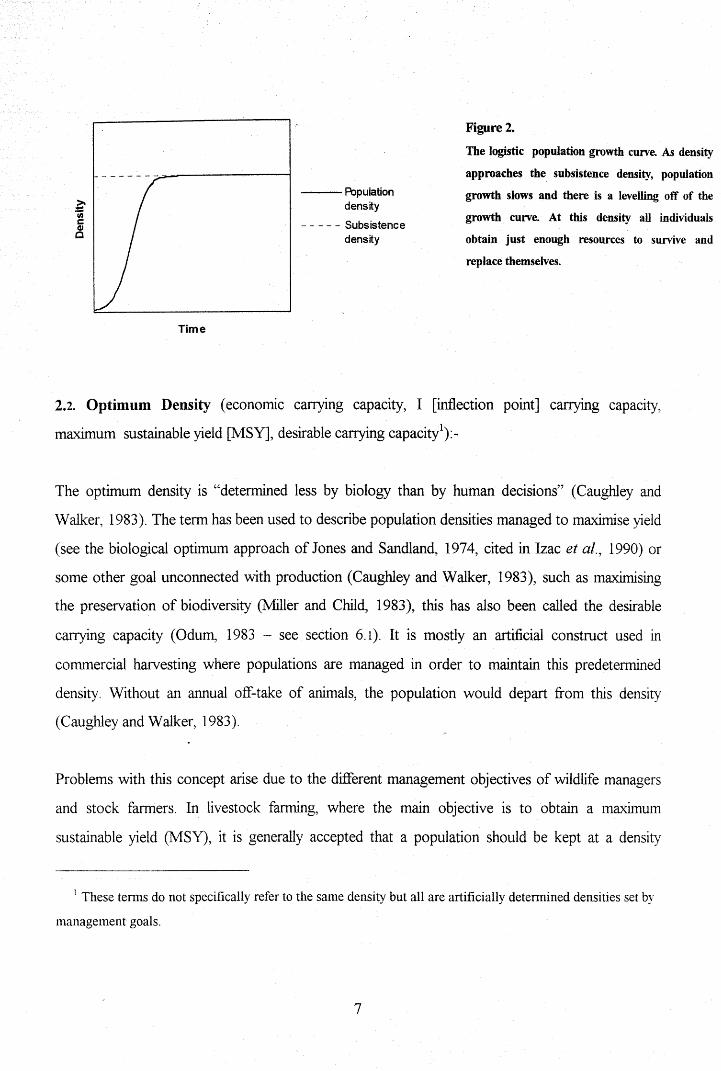
can be represented graphicallyby the logistic growth curve (Fig. 2).
Equilibrium population
density
Density
. - •. - •• rvbrtal~y
--Nataility
Figure 1.
Changes in the frequency of mortality
and natality with density. The
intersection of the two lines (indicated
by the arrow) represents the
equilibrium population density
(Murray, 1982). When this point is set
by resource limitation, it is equivalent
to the subsistence density.
An interesting consideration is whether one trophic category or another is more likely to be at the
subsistence density. The Green World Hypothesis, originally proposed by Hairston et al. (1960,
cited in Polis, 1994) suggests that the top predator trophic level will depress the level directly
beneath it. Consequently, the top level will always exhibit a biomass close to carrying capacity and
so will every alternate level down the trophic cascade. Although general rules such as these are
interesting, the, reality is probably much more complex (see Polis, 1994) and it is probably
dangerous to generalise.
6
Page 8
--------..-..---------1
Time
---Populationdensity
- - - - - Subsistencedensity
Figure2.
The logistic population growth curve. As density
approaches the subsistence density, population
growth slows and there is a levelling off of the
growth curve. At this density all individuals
obtain just enough resources to survive and
replace themselves.
2.2. Optimum Density (economic canying capacity, I [inflection point] carrying capacity,
maximum sustainable yield [MSY], desirable carrying capacityl):-
The optimum density is "determined less by biology than by human decisions" (Caughley and
Walker, 1983). The term has been used to describe population densities managed to maximise yield
(see the biological optimum approach of Jones and Sandland, 1974, cited in Izac et al., 1990) or
some other goal unconnected with production (Caughley and Walker, 1983), such as maximising
the preservation of biodiversity (Miller and Child, 1983), this has also been called the desirable
canying capacity (Odum, 1983 - see section 6.1). It is mostly an artificial construct used in
commercial harvesting where populations are managed in order to maintain this predetermined
density. Without an annual off-take of animals, the population would depart from this density
(Caughley and Walker, 1983).
Problems with this concept arise due to the different management objectives of wildlife managers
and stock farmers. In livestock farming, where the main objective is to obtain a maximum
sustainable yield (MSY), it is generally accepted that a population should be kept at a density
I These tenus do not specifically refer to the same density but all are artificially determined densities set by
management goals.
7
Page 9
between a half to a third of that potentially supportedby the available food supply i.e. the canying
capacity (Macnab, 1985 - Fig. 3)1. Management to maximise performance (yield) should be
regarded as striving for an optimal stocking rate, not the canying capacity (Mentis and Duke,
1976).
The relationship between canying capacity and productivity has led to statements such as that by
Raillard and Menesguen (1994) that "canying capacity... (in shellfish culture) corresponds to the
ability of the system to support shellfish production". Although this statement is not necessarily
wrong in itself it could easily be misinterpreted, leading to confusion over what density is being !
referred to. This is exemplified by Weeden's (1974) comment that a population at canying capacity
can "sustain a definable maximum harvest"2.
Z''u;ccuo
Time
_ .. - Population
density
- ~ - - - Subsistencedensity
- - -. -. Optimumdensity
Figure 3.
The optimum density intercepts the
population growth curve at its steepest point if
the management goal is to maximise
production.
1 This relationship may only be true for resources that are competed for by scramble competition and not
those competed for by contest competition (see sections 4.1 and 4.2 ).
2 Further confusion stems from inappropriate use of the term. Coe (1980), in comparing pastoral systems in
northern Kenya, used "optimum carrying capacity" when referring to the biomass of herbivores derived from
the regression of wild herbivore biomass against rainfall. This use of the term differs from that by others (e.g.
Dasmann, 1981; Macnab, 1985), i.e. there is no reference to management goals, only that large wild
herbivores occur at this density.
8
Page 10
avoid confusion I will refer to any density maintained by active management to obtain some
stated goal as the"cptimumdensity''iI supportMentis andDuke (1976) instating that this density
should not be referred to as the carrying capacity.
2.3. ToleranceDensity:-
Tolerance density, also referred to as "saturation density" by Leopold (1933, cited in Dhondt,
1988), is the density of individuals above whichintraspecific competition inhibits further increase in
numbers. I have adopted the term tolerance density in place of saturation density, which is
suggestive of a crowded population at subsistence level, whereas tolerance density is a density
above which intraspecific social tolerance allows no further population growth (Dhondt, 1988).
This also avoids confusion over other uses of the term saturation density (e.g. Odum, 1953, cited in
Dhondt, 1988.
Giles (1978) differentiates between potential carrymg capacity (equivalent to the subsistence
density) and tolerance density. He refers to the latter as the "theoretical maximum number of social
units" (for example, coveys of gamebirds) of a species that can occur in a prescribed area. Leopold
(1933, cited in Dhondt, 1988) also separated tolerance density and carrying capacity (subsistence
density) in a similar manner, noting that a carrying capacity exists for all species, but for some (e.g.
non-territorial species), no tolerance density existed. Giles (1978) suggests that unlike the
subsistence density, management cannot raise the tolerance density this is due" to the fact that the
tolerance density is a property of the species, whereas carrying capacity (subsistence density) is a
property of the habitat (Dasmann, 1981).
Dasmann (1981), in explaining the existence of populations at tolerance density, closely approaches
a group selectionist argument, suggesting that species with "a tolerance density equivalent to an
optimum density, so that self regulation of numbers provides the best conditions for the breeding
population" would have an "evolutionary advantage". Current evolutionary theory does not hold
9
Page 11
with the evolution of behavioural traits that would reduce the relative fitness of individuals that
adopted them for the benefit ofthe population' (Kaufinann, 1983).
I propose that a population density set by intraspecific interactions which is below the carrying
capacity ofthe environment is unlikely to exist in nature (see, however, the special case of space in
section 4.3). A decrease in territory size following food enrichment (e.g. Watson et al., 1984;
Watson and O'Hare, 1979) suggests that the primary reason for territorial behaviour is to gain
access to food resources (Sinclair, 1989). Consequently, if it exists, the tolerance density is either
equal to the carrying capacity or exists at some level above the carrying capacity where it is never
expressed (Fig. 4)2.
- - - -- - """---------1.2:' ._-_ •.. __ ._ .. _ •• _ .• - ••.u;c::GJo
Time
--Populationdensity
- - - - - Subsistencedensity
- - - - Tolerancedensity A
- - • - - Tolerancedensity B
Figure 4.
Tolerance density A is a purely theoretical
one, occurring above the subsistence density,
consequently it will never be seen in natural
populations. Tolerance density B would hold
the population below the level where
resources are limiting.
1 Any individ~al that passed down genes for a lower density intolerance (i.e. could thrive in more crowded
conditions) would produce offspring that would thrive when the population density was at a level that inhibited
recruitment of individuals with a high density intolerance. These genes would then spread through the
population until a density is reached in which availability of resources (other than space) become a limiting
factor.
2 It is possible that a population in which individuals defend territories throughout the year may be resource
limited only at certain times of the year (Kaufmann, 1983). However, this density should still be considered as
one set by resources other than space.
10
Page 12
2.4. Security Density:-
This is the level at which 'a population is maintained by predation (possibly also disease or
parasitism - Igbozurike, 1991) at a density below that of the subsistence density (this may be
visualised as equivalent to tolerance density B in Fig. 4). The security density is analogous to
Errington's (1945, cited in Dasmann, 1981) "threshold of security" in which populations exceeding
this density become unstable due to increased vulnerability to predation (which reduces their
number to that of the secure density). In effect, there is density-dependent mortality in the form of
predation (see Hill and Robertson, 1988). The strength of this factor on the prey population can be
seen by the population irruptions, that occur once a predator has been removed, for example,
population explosion of the mussel, Perumytilus purpuratus, following the removal of its main
predator the starfish, Heliaster helianthus (Paine et al., 1985).
Barlow (1996) used the term "threshold of security" to classify that density of potentially
susceptible hosts below which a disease is eliminated from a population due to the low rates of
transmission. Changes in the environment that favour population growth of a particular species,
raising its density above the threshold of security, will result in increased contact rates between
individuals and an increased transmission rate of the disease. This is only applicable for diseases
which are spread through physical contact and where contact rate depends upon population size
(Allen and Cormier, 1996). Sinclair (1989) proposed that parasitic infections may act in a similar
manner, although the strength of such population regulation is debatable (McCallum and Dobson,
1995).
2.4.1. Does a predator-regulated prey equilibrium exist in nature?
Predation can be either depensatory, having a destabilising effect on the prey population density or
compensatory, regulating the prey population density (Sinclair and Pech, 1996). An example of the
fornier may be if a predator takes a set number of individuals independent of prey density (Fig. 5).
This could be the case if the prey represented only one of many prey species available to the
predator. Therefore, as prey density increased, predation would remove an ever decreasing
11
Page 13
_~_ ........I"'I,nn of the prey (Caughley and Sinclair, 1994). This is a type II functional response by the
.......,.,rlgTC'lr (see Fig. 5), possibly exhibited b},' the caribou tRangifer tarandus) - wolf (Canis /upis)
L ...."'._~l'1nn in British,Colombia, with the moose (A/ces alces) being the main prey of the wolf
lSlJliClatf and Pech, 1996).
Prey Density
- - - - - Density-dependentpredation
---. Densityindependentpredation
Figure 5.
Change in predation pressure as prey
density increases. Density-dependent
predation gives a type ill functional
response and density-independent
predation gives a type II functional
response.
For predation to have a regulatory role on prey density it, would require density-dependent
feedback mechanisms' (Fig. 5). These can take one of two forms: firstly, a numerical response of
the predators to prey increase in numbers, or, secondly, a functional response such as prey
switching. This can occur almost instantaneously with a change in the relative prey densities and
therefore can reduce the time lag often associated with a numerical response due to the difference
in generation times between predator and prey. As a consequence, a numerical response may be
more likely to result in population cycles whilst the functional response may regulate prey density,
1 Colinvaux (1973) suggests that population regulation by predation is rare for large mammals and many of
the examples are from island or isolated populations. It is possible that small predators are more effective at
controlling their prey, due to their superior killing power and the predators' safety from injury (Colinvaux,
1973).
12
Page 14
called a type ill functional response! (Dale et al., 1994; Holling et aI., 1959, cited in Begon et al.,
1990), possibly exhibited by the moose-wolf interactions on the Isle ROYale (Dasmann, 1,981).
Messier (1994) found that in North America both wolf (Canis lupus) density and per capita killings
were strongly related to moose (Alces alces) density (exhibiting a type ill response) up to densities
of 0.65 moose/km" However, above this threshold density wolf predation was inversely density
dependent, no longer regulating population numbers (Messier, 1994)2. An empirical model using
these figures suggested that two possible equilibria existed in the presence of wolves. One at a low
density (0.2 - 0.4 moose/km-), where predation is the main regulatory factor on moose population
size. The other equilibrium occurred at a higher equilibrium density' of around 1.3 moose/km-. The
existence of these two equilibria shows the problems associated with generalities describing trophic
interactions such as the Green World Hypothesis (see Polis, 1994),
The existence of two possible equilibrium densities, as described above, means that those
definitions describing carrying capacity as an equilibrium population density are misleading.
Furthermore, contrary to being in equilibrium, it is often those populations near the carrying
capacity that show the greatest variability, being particularly vulnerable to malnutrition and disease
(Eberhardt and Siniff 1977; Mattson and Reid, 1991). Dasmann (1981) found that the original
moose population on the Isle Royale, Lake Superior, which was free from predation (therefore
presumably much closer to the subsistence density), was subject to severe die-offs. Following the
introduction of wolves the population settled to a lower, more stable density (Dasmann, 1981). The
issue oftemporal variabilityand carrying capacity is discussed in the follOWing section.
1 A type III functional response may be the result of learning, bioenergetic optimisation of search effort,
prey refugia, risk or vulnerability (Dale et al., 1994).
2 This may be due to food satiation or an upper limit set by handling time.
3 At this higher density food shortage is the main regulatory factor. although a limited amount of predation
keeps the population below the carrying capacity. Therefore at high moose densities wolves may influence, but
not regulate, moose numbers (Messier, 1994).
13
Page 15
3. TEMPORAL VARIATION AND CARRYING CAPACITY
carrying capacity (from here on referring exclusivelyto the subsistence 'density) is often depicted as
a stable state. However, it will almost invariably vary in both space and time (deLeeuw and Tothill,
1993) as summarised in the following quote:
"Carrying capacity is not fixed. It fluctuates naturally with weather and climate, and the operations
of other natural factors such as fire, floods. earthquakes and vulcanism. It is being modified
continually by human action" (Dasmann et al., 1973).
The concept of carrying capacity is most easily applied to populations at a stable equilibrium1.
However, the view that there is a "balance of nature" with populations at an equilibrium
density (Colinvaux, 1973) is now largely thought of as the exception and not the rule"
Therefore the idea of a constant carrying capacity is unrealistic (Potvin and Huot, 1983).
Pimm (1991) states that population dynamics can be placed into one of four categories: stable
equilibrium; essentially random climatic processes driving the population; cyclic; and widely
fluctuating (only encountering density dependence at extreme population densities).
In Figs 2-5, population growth has been represented as a sigmoidal curve stabilising at carrying
capacity. This is typical of population growth models derived from the logistic equation, although
1 I follow Begon et al. (1990) in defining the term stable equilibrium to be a population that returns to some
level after it has been displaced from this level by some disturbance.
2 What DeAngelis and Waterhouse (1987) call the "classical view" of ecological systems were based on the
existence of an equilibrium, although not necessarily a stable one. However, this view is becoming less
satisfactory in explaining the variation seen in nature, and the very existence of equilibrium states is being
called into question more and more often (DeAngelis and Waterhouse. 1987; Schaffer and Kot, 1986). S. L.
Pimm (pers. comm.) suggests that no population can be said to be stable: the longer any population is
observed. the greater the variation that is to be found in the population dynamics.
14
Page 16
other equations produce similar curves (Odum, 1983). odum (1953, cited in Dhondt, 1988) was
the first to introduce the term carrying capacity to the logistic equation as the upper asymptote (K).
However, Dhondt (1988) states that 'due to the assumptions implicit in the equation and its
deterministicnature, its application to carrying capacity is "deeply misleading".
The basic assumptions made by the logistic equationare:
1. a linear relationship between increasing density and negative feedback on population growth.
Emlen (1973) stated this as being one ofthe most unrealistic assumptions ofthe equation';
2. continuous birth and death (i.e. the population does not have discrete generations, which is not
true for most birds and mammals); and
3. every individual in the population is the same
( after Dasmann, 1981).
One further point not stressed by Dasmann (1981)· is that the logistic equation assumes that K is
stable. As Begon et al. (1990) state, "all environments are variable, the position of any balance
point is continually changing", consequently "no single carrying capacity can characterise a natural
population" (Begon and Mortimer, 1981). This continual flux in the carrying capacity can be
represented by adding stochastic variation to K (Fig. 6). I propose that the (current) carrying
capacity should be equated to K, although Bartels et al (1993) state that "carrying capacity is an
average number that populations fluctuate around" (Fig. 6). Other authors suggest that the troughs
in density should be considered as the carrying capacity (e.g. Edwards and Fowle, 1955, cited in
Dhondt, 1988 - Fig. 7) andCaughley and Walker (1983) represent carrying capacity as a fuzzy
boundary encompassing the densities around which the population fluctuates (Fig. 7).
1 It has been recognised that mechanisms of density dependence may be different at extremes of population
densities to those found at central (average) densities, i.e. when the population is close to its equilibrium
density (Sinclair. 1989). It is possible that "density vague phenomena" (sensu Strong, 1986) are a simple
consequence of "curvilinear density dependence" (Sinclair, 1989~ Sinclair and Pech, 1996) where, at some
densities, regulation is strong and at other densities regulation is weak, such as in the collared flycatcher
(Ficedula albicollis) in central Europe (Torok and Teth, 1988).
15
Page 17
Figure 6.
Logistic growth curve with K varying
stochastically. Carrying capacity
--Population density sensu Bartels et aL (1993),
represented by the dotted line, is an
average density around which the
population fluctuates.
Time
Figure 7.
Time
-- Population density
Carrying capacity, sensu Edwards and
Fowle (1955, cited in Dhondt, 1988),)
denoted by the lower dotted line,
representing the trough in population
fluctuations. Carrying capacity sensu
Caughley. and Walker (1983), depicted
as the area between the two dotted
lines, encompassing upper and lower
boundaries within which population
density fluctuates.
The types of fluctuations that populations are likely to undergo can be separated into different
classes:
1. seasonal (or intra-annual) changes, resulting from a combination of life history strategies and
seasonal changes in the environment; and
16
Page 18
2. annual (or supra-annual) fluctuations, due to either changes between years in the physical
environment or to population dynamics (after Odum, 1971).
3.1. Seasonal Variation
Where there is marked annual climatic variation, there are usually seasonal fluctuations in plant
growth (pietersen et al., 1993). Consequently, Liebig's "Law of the Minimum" (see section 5)
would be operating at only a certain time of year (Edwards and Fowle, 1955, cited in Dhondt,
1988). Therefore, a population may be held below the carrying capacity, when resources are
abundant, because of seasonal mortality when resources are limiting (Dhondt, 1988). This results in
a stable population in a fluctuating environment'.
However, many animals have adapted to these seasonal fluctuations and go through a cycle of
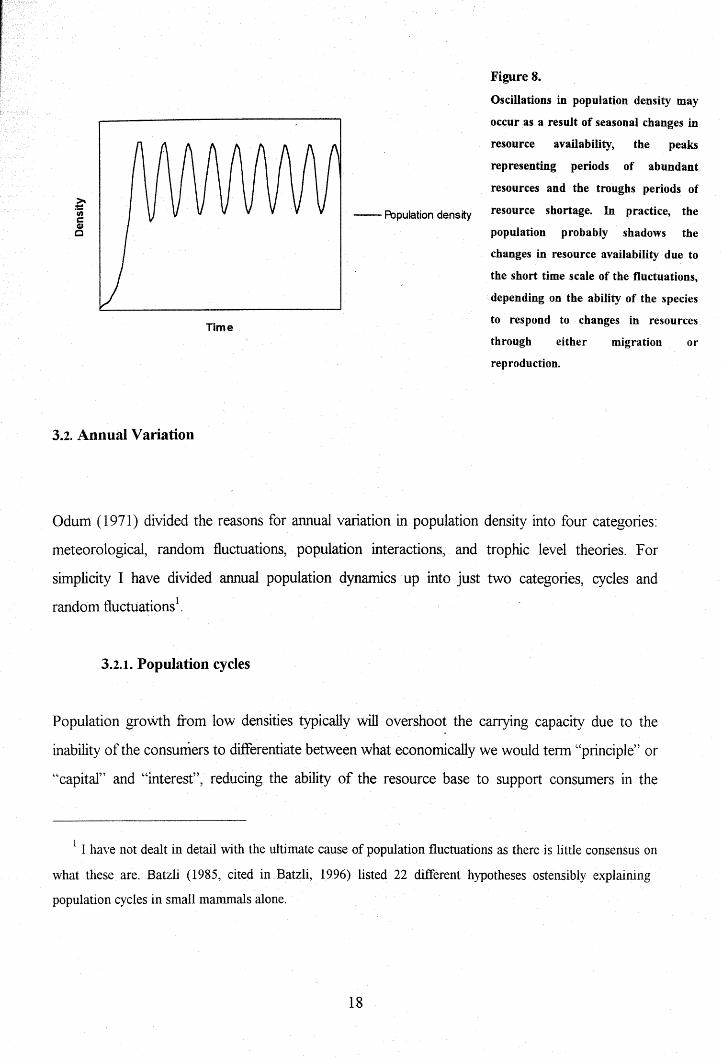
increase and decrease in phase with these changes (Fig. 8). Numbers increase dramatically in the
breeding season, frequently to be followed by high mortality. The population, therefore, oscillates
widely from season to season. However, the trough in population density may remain relatively
constant from year to year (Dasmann, 1981), such as that found in winter wader numbers in
northern latitudes (Goss-Custard, 1985). Edwards and Fowle (1955, cited in Dhondt, 1988)
propose that this trough should be the carrying capacity of the environment, differentiating it from
the seasonal fluctuations in resources, which set the "current carrying capacity".
I This would only be the case for populations with a generation time in excess of a year.
17
Page 19
Time
3.2. Annual Variation
Figure 8.
Oscillations in population density may
occur as a result of seasonal changes in
resource availability, the peaks
representing periods of abundant
resources and the troughs periods of
__ Population density resource shortage. In practice, the
population probably shadows the
changes in resource availability due to
the short time scale of the fluctuations,
depending on the ability of the species
to respond to changes in resources
through either migration or
reproduction.
Odum (1971) divided the reasons for annual variation in population density into four categories:
meteorological, random fluctuations, population interactions, and trophic level theories. For
simplicity I have divided annual population dynamicsup into just two categories, cycles and
random fluctuations1.
3.2.1. Population cycles
Population growth from low densities typically will overshoot the canying capacity due to the
inability of the consumers to differentiatebetween what economically we would term "principle" or
"'capital" and "interest", reducing the ability of the resource base to support consumers in the
1 I have not dealt in detail with the ultimate cause of population fluctuations as there is little consensus on
what .these. are. Batzli (1985, cited in Batzli, 1996) listed 22 different hypotheses ostensibly explaining
population cycles in small mammals alone.
18
Page 20
future. Depending on the length ofthe time lag and the magnitude ofr (the intrinsic rate ofgrowth),
the population may fluctuate around K with the oscillations dampening down to the point at which
K is reached (unless a further disturbance occurs to the population). This has been termed a
"relaxation oscillation" pattern (Odum, 1971 - Fig. 9). A greater time lag may result in population
cycles which do not dampen down over time (Caughley, 1979)1. Under this scenario a potential
equilibrium exists between the consumer and the resource, but it is never reached. Instead there is a
constant fluctuation in both carrying capacity and population density (Fig. 10).
Time
--_.- PopulationDensity
- - - - - CarryingCapacity
Figure 9.
A relaxation oscillation pattern in
which the population fluctuates
around and eventually stabilises at
carrying capacity.
I May (1976, cited in Hornfeldt, 1994) originally proposed thata ninemonthtime lag in feedback response could
explainthe 3-4 year population cycles of thevoles tMicrotus agrestis, Clethrionomys glareolus and C. rufocanus).
19
Page 21
Z.'iiicQ)
o
Time
3.2.2. Random fluctuations
--'- Populationdensity
- - ~ - - Carrying capacity
Figure 10.
Population cycles in which population
density initially overshoots carrying
capacity, causing a degradation of the
habitat, followed by a population
crash. As the habitat recovers and
carrying capacity increases once
more, the pattern repeats itself.
Long time lags or a very high intrinsic growth rate may result in high peaks followed by crashes in
population density (Caughley, 1979 - Fig. 11). This has been coined "deterministic chaos" and may
explain apparently random fluctuations in natural populations (Berryman and Millstein, 1989~
Massot et aI., 1992). However, truly random population fluctuations are due to density
independent environmental fluctuations either not affecting the resource base or temporarily
lowering the carrying capacity, causing the population to crash.
.A A .A A
"~ 1\ .~ 1\
Time
20
---- Populationdensity
....... Carrying
capacity
Figure 11.
Deterministic chaos: apparently
random oscillations in population
density as a result of overshooting the
carrying capacity due to a high
intrinsic rate of growth.
Page 22
populations in relatively stable environments will tend to be self-regulating, having been subject to
selection pressures favouring K-selected traits (e.g. large size, delayed reproduction, small
reproductive allocation, high levels of parental care and the production of few but large offspring),
which maximise competitive ability in high density populations (pianka, 1970; Elgar and Catterall,
1989). In variable environments, r-selected traits (e.g. small size, precocious reproduction,
semelparity, a large reproductive allocation and the production of many small offspring) which
maximise productivity, will be favoured (pianka, 1990). Consequently in low-diversity, physically
stressed environments, populations tend to be regulated by density-independent factors (Odum,
1971), resulting in the expression of non-equilibrium population dynamics (Scoones, 1993).
Depending on the frequency of environmental perturbations and the relative recovery time of the
population, it is possible that a harsh, variable climate may keep the population below the resource
defined carrying capacity (subsistence density) for the majority ofthe time.
It may therefore be inappropriate to apply the concept of carrying capacity to populations
undergoing random fluctuations below the canying capacity (typically species exhibiting r-selected
traits)1. Odum (1971) suggests that species' population densities fluctuate most strongly in
relatively simple, species-poor environments' such as the arid and semi-arid areas of sub-Saharan
Africa, in which Caughley and Walker (1983), Hoffinann et al. (1995) and Bartels et al. (1993) all
suggest that using the concept of carrying capacity in explaining population dynamics is
inappropriate.
1 Even in many relatively constant environments, small species often exhibit r-selected traits. This may be
due to the scale at which humans judge the variability of an environment. What may appear as a constant
environment at one scale may appear to be highly variable at another.
2 The conventional theory supported Odum's view that simple communities ere inherently more unstable
than more complex ones (Elton, 1958). However, more recent work has cast doubt over these assumptions and
the exact relationship between stability and complexity probably varies with trophic leveL top down and bottom
up systems, and the predictability of the environment (Nilsson and Grelsson, 1995).
21
Page 23
I suggest that questions of whether carrying' capacity is a meaningful concept when applied to
fluctuating populations, such as wildebeest (Connochaetes taurinus) where numbers were found to
vary between 250.000 and 1 000000 in the Serengeti.~ Mara region ofEast Africa (Talbot, 1977),
should depend on whether the population dynamics are driven by changes in the resource basel,
(typically k-selected species), or by other environmental factors (typically r-selected species). If the
former is the case, carrying capacity is central to the understanding ofpopulation dynamics, and the
factors setting the carrying capacity and the means in which they act should be investigated.
However, ifthe latter is the case, the concept ofcarrying capacity may be largely irrelevant.
4. WHAT SETS THE CARRYING CAPACITY?
The question of what determines carrying capacity is not an easy one to answer. Many obvious
factors can easily be identified, such as the presence and quantity of the right kind of food, presence
of water, appropriate soil and topography as well as the interaction between all these factors. All
relate to the provision of suitable habitats for the life history and behavioural requirements of a
particular species, such as nesting, resting, mating and predator avoidance, delineating the
fundamental niche of the species. With relatively little imagination this list could be extended almost
indefinitely.
It may be impossible to identify all the attributes of a given habitat required to support a given
number of a given species. Dasmann (1981) puts this in context by stating that even for the most
well known species, man, we do not know all the various factors. However, in some circumstances,
it may not be necessary to do this, as the determinant of population density of a given area of
habitat is set by only one or a handful offactors.
I The population density is some function of resource availability.
22
Page 24
The "law ofthe minimum" was a concept devised by Justus Liebig (Dasmann, 1981) when studying
plant nutrition. The growth ofa plant is dependent on the amount ofthe resource that is available to
it in "minimum quantity relative to the plant's needs" (Dasmann, 1981). This resource is the limiting
factor, and an increase in the availability of other resources will not enhance growth. This concept
can be applied to those factors which limit the total density of a species in a particular area, in effect
setting the carrying capacity.
Many of these factors are equivalent to those that regulate populations 1, such as energetic and
nutritional resources. In 1954, Lack (cited in Dhondt, 1988) suggested that most vertebrate
populations were limited by food. This is supported by the fact that 53% of studies of density
dependence cited food as the regulating factor (Sinclair, 1989), and the response of many terrestrial
vertebrate populations to food supplementation is to double in density (Duquette and Millar, 1995).
The food limitation hypothesis of population regulation states "that a stable equilibrium will exist
between a population and its food resource due to a density-dependent decrease in fecundity and!or
increase in mortality" (Seetheret a/., 1996). This is a typical response to scramble competition for
resources. Other factors merely limit the maximum size of the population, such as cover and nest
sites, being competed for by contest competition, although Hassell (1975) and Murray (1982)
suggest that these may also lead to apparently density-dependent population dynamics.
4.1. Scramble Competition
Scramble competition is the "most extreme form of overcompensating density dependence" where,
due to the effect of intraspecific competition, "all competing individuals are so adversely affected
that none ofthem survive" (Begon et a/., 1990). This definition takes the concept to extreme levels:
1 Regulation "is the tendency of the population to return, through density-dependent factors. to the
equilibrium level when disturbed" whereas limitation is the process which sets the potential equilibrium level.
Consequently all causes of mortality and reproductive loss are limiting factors (Sinclair and Pech, 1996).
23
Page 25
it may be easier to consider a limiting factor, such as water, of which there is only a set amount
available each day. Each individual in a population requires a litre of water a day to remain in
perfect health, however, they can survive on half a litre of water a day but are in a relatively poor
condition. The former scenario can be thought of as a population at the optimum density for
maximising productivity and the latterof oneat the carrying capacity where each individual obtains
just enough resources to survive and reproduce successfully. Between these two densities the
individuals can be classed in the range from healthy to unhealthy. If the population density exceeds
the number where each individual can get halfa litre of water a day they all suffer from lack of
water anddie1.
Under this scenario the population would undergo exponential growth until the population size
reaches the level where each individual could only get a litre of water. Once this point is exceeded.
density-dependent population regulation would be expected to reduce the population growth rate
until an equilibrium is reached at the subsistence density, giving an "S shaped" population growth
curve parallel to that obtained from the logistic equation 2. 1£ for some reason, the population
overshoots or the resource decreases after the population has reached this equilibrium, a population
crash would be expected.
I This is based on the assumption of symmetrical inter-individual competition and tolerance of resource
shortage. which is unlikely to be the case (see section 5.1.1) (HasselL 1975).
2 Many authors (e.g. Sinclair, 1989; Hassell, 1986: Scoones. 1993; Dennis and Taper. 1994; Cooch et al..
1989) have stressed the difficulty in identifying whether or not population regulation is occurring. However.
apparent examples of density dependence are common in the literature: for example. laboratory populations of
the fruit fly (Drosophila melanogasteri (Rodriguez. 1989). the lizard (Lacerta viviparai (Massot et al., 1992).
the snow goose (Chencaerulescens caerulescensy (Cooch et al., 1989). and the wood mouse iApodemus
sylvaticusi (Montgomery. 1989).
24
Page 26
However, Murray (1982) states that density-dependent factors are not required for this
relationship1. A limiting factor that is density-independent, such as minimum breeding territory size,
may set the maximum breeding popufation size (Murray, 1982); This would be an example of
contest competition.
4.2. Contest Competition
Contest competition is "intraspecific competition in which mortality compensates exactly for
increases in density, so that there are a constant (or approximately constant) number of survivors
irrespective of initial density" (Begon et al., 1990). Using the same analogy as above, contest
competition can be seen as competition for a set number of litres ofwater available every day.
Individuals either compete for water successfully, in which case they are healthy, or unsuccessfully,
and perish: there is no density-dependent population regulation (growth can be exponential) until
all available resources are used, in which case the population is at the subsistence density. Such
examples may be rare in nature. However a similareffect may be seen in populations where there is
considerable difference in competitive ability for a limiting resource. Superior competitors will
secure sufficient resources to maintain good health whilst poor competitors may perish or emigrate
from the area' (see section 5.1.1). This scenario would give a typical "J- shaped" population growth
1 The existence of density-dependence was often debated because of its perceived importance in buffering
populations from extinction (e.g. Ginzburg et aI., 1990). However, as Murray (1982) shows, true density
dependence is not required, only some form of population limitation at the extremes of population density.
2 If a limiting factor. such as nest sites, held the population at a level below that which could be sustained
by the depletable resources (such as food), it is possible that the ecological carrying capacity could be close to
or at the biologically optimal density (maximising production) (Macnab, 1985). However, as the optimum
density. is an artificial construct, any correspondence between the actual population density and the optimal
density would be purely coincidental.
25
Page 27
curve levelling off abruptly at the population density determined by the available resources (0dum,
1983 - Fig. 12).
Time
5.3. The Special Case of Space
---Populationdensity
....... ".. Carrying
Capacity
Figure 12.
The "J- shaped" population growth curve,
with exponential growth until the population
reaches the carrying capacity (determined by
density-independent factors) and growth is
halted.
Many studies suggest that breeding population size is determined by the number of available
territories, which remains relatively constant from year to year, regardless of fluctuations in the
numbers ofnon-territorial individuals (e.g. gamebirds - Watson and Jenkins, 1968). Examples such
as these have led people to believe in the existence of some sort of social space requirement, similar
to a tolerance density. However, examples of populations at the tolerance density cited in the
literature are often inconclusive. As stated in section 2.3, I believe that the existence of a tolerance
density is highly unlikely. Predation (e.g. Duquette and Millar, 1995; Errington, 1934, cited in
Dhondt 1988), unusually depressed population densities (e.g. Dasmann, 1981; van Zyl, 1992), mis
identification of the limiting resource' or population regulation during a different life history phase
are frequently the real reasons for what appears to be over-dispersion.
1 The Seychelles magpie robin (Copsychus sechellarum) on Fregate island was thought to be limited by
nesting sites and access to foraging habitat. However, supplementary feeding and the introduction of nest boxes
could not increase the number of territories beyond 11 (Komdeur, 1996). This is probably due to the limiting
effect of another (unidentified) resource as a result of the lack of foraging habitat.
26
Page 28
House mice (Mus musculus domesticus) in corn stocks, where resources are known not to be
limiting, show adrenal weight increases (indicating an alteration in the neural-endocrine balance),
thymus weight decreases (indicating increased adrenocortical activity), and decreases in the weight
of reproductive glands as density increases (Konnondy, 1976). It seems hard to argue against self
imposed regulation. However, I suggest that, in natural populations, in an enviromnent parallel to
one where mice evolved (in which superabundant resources in the form of com stocks did not
exist), such high population numbers may have been indicative that the population would soon
suffer from resource shortages. Therefore, individuals would be selected to alter their resource
allocation away from maximising the number of offspring, to survival or maximising the fitness ofa
single offspring1.
There is, however, one situation where a tolerance density may set the maximum population
density, or at least the number of breeding pairs. This scenario could arise if there is mate choice
based on the size of territory (Trombulak, 1990). For example, females select those males with the
largest territories'. resulting in male-male competition for space'. The fact that possession of a
territory can be essential for breeding in some bird species, such as the red grouse tLagopus
lagopus) (Watson and Jenkins, 1968) suggests that there is mate choice for territory holders. As
most territorial behaviour is linked to resource acquisition (Kaufinann, 1~83), territory size will
reflect resource availability. However, mate choice based on territory size, as in the pied flycatcher
(Ficedula hypoleuca) (Dale and Slagsvold, 1990), may result in apparent over-dispersal, with space
1 A similar explanation is offered by Dawkins (1976) to explain what appears to be group selection.
Apparent self-regulation. unconnected with resources in one environment (the laboratory). can be easily
explained in terms of resource limitations in another environment (the evolutionarily natural environment).
:; Random settlement by females would also mean that it would be advantageous for males to secure large
territories (Dale and Slagsvold, 1990)
3 Even in this example it could be thought that the real resource being competed for is a mate, not space in
itself.
27
Page 29
acting as the limiting factor. I have entitled this section "the special case of space" as the resource
being competed for may set the canying capacity although it does not enhance the physical fitness
of the successful competitor'. In fact defending such a large territory may be costly in time and
energy.
Once those factors that determine the carrying capacity have been identified, it may be possible to
measure the carrying capacity empirically.
5. ESTIMATING CARRYING CAPACITY
The following section examines the pros and cons of different approaches to the estimation of
canying capacity with regard to the aspects of interseasonal variation, spatial variation, plant
species composition, interactions between herbivores (consumers) and plants (producers), and
interactions among herbivores (consumers), which were identified by Walker (1982) as being
problematic to the estimation of carrying capacity. The particular problems associated with
composite-species carrying capacities will be discussed. Attempts to estimate carrying capacity fall
into two main categories, differentiated by the units in which they are measured: firstly, those that
are a measure of the supportable animal units, and secondly, those that are a measure of resource
availability.
I This applies to any arbitrary resource which is sexually selected for.
28
Page 30
5.1.Estimating Carrying Capacity in Terms of Supportable Animal Units
5.1.1. The population density approach
This approach is based on actual population densities and is susceptible to many of the problems
listed by Walker (1982), in that population density may vary over space and time. Crete (1989)
attempted to circumvent some of these problems associated with the interactions between
herbivores and plants, by using a stable population of herbivores which had had time to equilibrate
with the vegetation where hunting was absent.and predation pressure relatively low. However,
estimations of carrying capacity would still be no more than an educated guess as to the effects of
pathogens and migration on population density.
An interesting variation to this approach is adopted by Suter (1995) in estimating the carrying
capacity for Swiss winter populations of the great cormorants (Phalacrocorax carbo). The
wintering population densities were found to closely fit the sigmoidal growth curve with the
population close to the upper asymptote, indicating that this number was the carrying capacity.
However, as in the previous example, it is impossible to tell from these data if the population can be
classed as approaching the subsistence density or some other equilibrium, such as one set by
predation.
An alternative approach was used in an attempt to establish the carrying capacity of the
environment for coypu (Myocaster coypus) based on high population densities where density
dependent self-regulatory factors were starting to become apparent' (Reeves and Usher, 1989).
Alone, this method suffers from the some of the drawbacks as using demographic characteristics to
1 It wouldbe expectedthat a relatively low birth rate and high death rate would characterise a population near
carrying capacity. This is exemplified by the Seychelles warbler (Acrocephalus sechellensis), in which individuals
translocated from a saturatedhabitat (with the populationnear carryingcapacity), Cousin island, to one in which the
warbler was absent, Aride island, had increasedclutch size, adultsurvival, fledgling success and survival of young to
oneyearof age (Komdeur, 1994).
Page 31
measure population status (e.g. time lags - section 6.3). A combination of this method and the one
above may get around the problems associated with each on its own. A time series of population
sizes should establish if the population' is increasing, decreasing or is at equilibrium. By using
demographic criteria, SOIne indication may be given, ifthe population is at equilibrium, whether or
not this is set by the subsistence density (for example, self-regulation in the form of a decrease in
fecundity or an increase in emigration would suggest that the population was near the carrying
capacity).
It is therefore possible to estimate the carrying capacity for a certain habitat using population
densities (measured in number of individuals) by incorporating demographic information. However,
this may require long term studies of the population, and is applicable. only to the species
concerned. Such an approach to calculating carrying capacity is based on the assumption that
habitat use/availability reflects the carrying capacity of the habitat (Hobbs and Hanley, 1990). This
in turn is dependent on two other assumptions:
1. a species will selectan area in whichit willbe ableto best satisfy its needs; and
2. as a consequence of the above, higher quality habitats will be used to a greater extent.
This isbasedon the theory of the "idealfree distribution" (IFD)originally proposedby Fretwell and
Lucas (1969, cited in Kacelnik et al., 1992) in which each individuals settles in the patch where it
can obtain the highest intake rate of food, i.e. individuals distribute themselves evenly in terms'of
competition for resources.
Many of the assumptions' on which the theory of the IFD is grounded are questionable and
Kacelnik et al. (1992) state that most field studies of natural populations do not confirm the
existence of the IFD.. Selection of patches may be based on predator avoidance as well as food
1 The IFD assumes that..a) resources are distributed in patches, b) competitors are equal in all respects, c) there is
no exclusion of some individuals by others, they are free to settle anywhere, d) travel time between patches is
negligible, or settling is sequential with the best patches at the current time (those with the highest potential intake
rate) being occupied first, e) the potential intake rate in every patch is known at all times, f) resource density remains
constant i.e. there is no depletion, and g) intake rate increases with resource density (after Kacelnik et ai., 1992).
30
Page 32
resources, such as in McNamara and Houston's (1990, cited in Kacelnik et al., 1992) dynamic
"state-dependent" IFD model. Where risk of predation varies inversely with resource value of a
patch, competitors with a low body reserve (close to starvation) will select patches with high
resource value arid high risk of predation, as competitors with a high body reserve will select
patches of a relatively low resource with low rates of predation (Kacelnik, et al., 1992).
Consequently those habitats with greatest resources will not necessarily contain the greatest density
of consumers. Studies of the smooth newt (Triturus vulgaris) found that predation determined
whether or not the species occurred in a certain pond (Halley et al., 1996).
Where there is asymmetric intraspecific competition for resources, a sub-dominant individual may
be better off foraging in a lower quality habitat where interference competition is low (Goss
Custard et al., 1994). Dominant individuals may be able to maintain their food intake rate in the
presence of intraspecific competitors at a level equal to or above that achieved in the absence of
competition (Sutherland and Parker, 1985). This scenario is similar to "ideal despotic distribution"
(IDD) of Fretwell and Lucas (1969, cited in Kacelnik et al., 1992) in which superior competitors
monopolise the patches of high resource value, consequently these patches are exploited by fewer
individuals. This leads to a distribution opposite to that predicted by the IFD model, with a lower
ratio of competitors to resources in high quality patches (Kacelnik et al., 1992) complicating the
estimation ofcarrying capacity.
Populations may only tend towards the predicted IFD when numbers are at or over the carrying
capacity (e.g. Bautistaet al., 1995; Focardi et al., 1995). Furthermore, the estimation of carrying
capacity based on densities overlooks the large variation in mass between individuals.
5.1.2. The biomass approach
Within a sexually dimorphic species one sex is often larger than the other, requiring more resources
for growth and maintenance. The proportion of gestating females may also alter the resource
requirements of the total population. Consequently the resource requirements of a population are
31
Page 33
not only dependent upon the number of individuals but also the make-up ofthat population in terms
of sex ratio, size distribution and reproductive. status. It is generally accepted that plants do not
react to density of individuals as such but to the "effect of their neighbours". This is a function of
the number, size and proximity of neighbouring.plants (Kataoka et al, 1989). Therefore, a small
number of large individuals may have the same competitive effect' as a large number of small
individuals. Consequently, the numerical size of a population is regulated to a much lesser extent
than is its biomass.
In 1940 Mohr (cited in Peters and Raelson, 1984) proposed that biomass of mammalian
populations remained constant per unit area, i.e. was independent of size; as individual mass
increased, population density decreased. This is exhibited by the limpet, Patella cochlear, where
total supportable biomass is similar for populations of different densities (Branch, 1975), i.e.
independent of the size of the individuals'. It has been argued that community biomass is the best
means of making comparisons between areas (e.g. White, 1994). However, a biomass approach to
estimating composite-species carrying capacities should incorporate the relationship between mass
and metabolism.
5.2. Estimating Carrying Capacity in Terms of Resources
5.2.1. Energetics
Mentis (1977) shows that, although there are no major differences in the relative proportional
contributions of individual species in their resource requirements, estimates of carrying capacity
1 Competitive effect should be interpreted here as reflecting resource requirements rather than active
competition per se.
2 This. would require that the limpets follow a -1 thinning rule, which may be unlikely in animal
populations (see following section).
32
Page 34
based on biomass consistently underestimate the real situation. Most methods for estimating energy
flow through a particular population have concentrated on energy for metabolism which generally
accounts for the largest energy expenditure (especially for endothermic animals - Nagy, 1989).
Many attempts have been made to construct some sort of thinning rule (such a the -3/2 self
thinning rule in plants1- Westoby, 1984) to predict the carrying capacity ofan environment, having
knowledge ofthe habitat's energy resources.
Energetic allometries suggest a -4/3 thinning-rule' (Begon, 1986; Elliot, 1993) based on the
metabolic - weight allometry (power of 0.75 - Pietersen et aI., 1993)3. This relationship has
become known as the "Energetic Equivalence Rule" (Cotgreave, 1993). The theory is best
explained by using the mathematical relationships: as population density scales to W-O.75 and BMR
(basal metabolic rate) scales to »r:". the rate ofenergy use in any given area is equal to:
W-O.75 * W+O·75 =W = 1.
Therefore, the amount of energy flowing through each species in a community is independent of
body size.
1 The -3/2 slope is derived from plotting log biomass (B) against log density (N) (Westoby, 1984). When a stand
reaches the asymptote (the maximum attainable biomass or yield per unit area), it moves from a stage where it is
limited by space to one where it is limited by the carrying capacity of the site (Zhang et al., 1993). Once the carrying
capacity is reached, the slope becomes -1 (Begon, 1986;Hughes and Griffiths, 1986) obeying the Law of Constant
Yield (see Begon et al., 1990). This means that metabolism is intraspecifically mass-independent, therefore a
doubling in size results in a doubling of resource requirements. The carrying capacity referred to above is the
maximum biomass that can be supported, which is set by food limitations. The spatial limitation is not
considered to be setting the carrying capacity, as growth (increase in biomass) is still possible.
2 The parallel between these two thinning rules is spurious. The -3/2 rule is set by space and the -4/3 rule is
set by food resources. The latter may be thought of as closer to the -1 thinning rule in plants at carrying
capacity.
3 Sessile organisms at high densities which are space limited are likely to follow the same -3/2 thinning rule
as plants (Begon, 1986). Possibly without exception, all sessile animals are filter feeders and are, consequently.
unlikely to be food limited, therefore they are unlikely to reach a carrying capacity set by food requirements.
33
Page 35
Mean metabolic rate should be proportional to the mean amount of food consumed (this ignores
interspecific variation in assimilation efficiency). Theoretically, if food is evenly distributed
throughout the environment, the spatial requirements of a species would scale to its metabolism,
hence the -4/3 thinning rule'. Therefore, knowing the mass of individuals of certain classes and the
relative distribution of these classes in a population', it should be possible to calculate the carrying
capacity, in terms of total biomass, from the amount of available resources in the environment. It
should be remembered that when estimating the energetic value of resources, their net value (e.g.
total energy supplied by the resource - energy expended in foraging for the resources) should be
taken into account, not their absolute value'. What may appear as sufficient resources to supply the
energetic requirements ofa population could be insufficient ifthe cost offoraging is high.
For each trophic category the availableenergy can be calculated, and this sets the energetic carrying
capacity (a notable omission from the list ofvarious classifications ofcarrying capacity found in the
1 For these self-thinning rules to be valid it requires that the resource remains constant. i.e. growth of the
population does not alter the availability of the resource. This is one of the properties of sunlight, but it may be
less appropriate for resources required for animal populations. The self-thinning rule for mobile animals
should, therefore, take into account both the animal's energetic allometries and the interactions between the
animal and its resources (Begon, 1986).
2 Due to the fact that the demographic structure of a population may vary with density relative to carrying
capacity. the distribution of classes should be taken from a population at the carrying capacity.
3 The importance of using the net resource value can be seen in the following assessment of the carrying
capacity for white-tailed deer (Odocoileus virginianus) wintering in Quebec. It was found that all vegetation
cover types could provide a positive energy balance with up to 50 em of snow. However as snow depth
increased. the vegetation types that could still provide a positive energy balance decreased due to the energetic
cost of walking in deep snow (potvin and Huot, 1983). It should also be born in mind that all available
resources will never be consumed. due to the law of diminishing returns - as the resource becomes rarer, the
search effort increases until eventually it costs more energy to search and process the resource than is gained
from its consumption.
34
Page 36
literature). The actual density of individuals would vary depending on the mix of species and their
relative masses. Carrying capacities could therefore be expressed as an "index of total community
consumption" (Edgar, 1993). Collinsonand Goodman (1982) and Nanni (1979) suggest that
carrying capacity should be estimated for discrete classes of foragers; as the canying capacity for
one particular class of consumers may be greater than that for another of equivalent energetic
requirements (see Mentis, 1977). Estimates of the total potential canying capacity should be
divided depending on the proportion of different food types in the areal. Depending on the
management objectives of the area, each foraging class can be divided at the level of the species to
obtain the desirable community makeup (Collinson and Goodman, 1982).
However, the -4/3 self-thinning is based on several dubious assumptions; that similar amounts of
energy are available to species of all sizes (Cotgreave, 1993), that energy use is independent of
body size", and that the relationship between body mass and energy requirements is independent of
hi 3trop c category .
1 The interaction between consumers and producers such as facilitation, the opening up of the herb layer for small
herbivores by large roughage eaters, makes the estimation ofcomposite-species carrying capacities more complex
(Mentis and Duke, 1976; Sinclair and Norton-Griffiths, 1982; Grobler and Jones, 1980):
2 Silva and Downing (1995) found that, in fact, only mammals with a mass between 0.1-100 kg obeyed the
W·0 75 scaling rule with an apparent peak in energy use per unit mass in species with a body mass around 100 g
(Marquet et al., 1995). Of 45 mammals examined, the relationship between body mass and density varied from
positive to negative to no relationship at all (Silva and Downing, 1995) ..Silva and Downing (1995) proposed
that an upper limit to small mammal densities may be imposed by food, due to the energetic cost of living near
the ultimate body size constraint for homeothermy. The minimum population density for large mammals may
be set by the constraints of finding a mate. Furthermore, the exact relationship between body size and
metabolism may vary between biogeographic regions (Du Toit and Owen-Smith, 1989).
3 This was found not to betrue (peters and Raelson, 1984; Reiss, 1988), with the relationship changing from a
positive one (e.g. omnivores) to a negative one (e.g. carnivores and insectivores) (Marquet et al., 1995) .
35
Page 37
Further reasons why the -4/3 thinning rule may be inaccurate are given by Reiss (1988):
1. average daily metabolic rate may scale to mass more or less steeply than basal metabolic rate' ~
2. productivity of the .environment may scale negatively with size, i.e. large home ranges may
contain a greater proportion of"useless space";
3. larger species share home ranges with more conspecifics; and
4. stress is placed on daily energetic requirements instead of on some biologically relevant time
period, such as the breeding season, when energy requirements are highest (adapted from Reiss,
1988).
These problems are compounded ifestimates ofprimary production are used to predict the carrying
capacity ofhigher trophic levels.
The attraction of using energetic allometric scaling to calculate carrying capacity is that composite
species canying capacities can be estimated. Incidental evidence for the existence of a composite
species canying capacity comes from attempts to increase cheetah (Acinonyx jubatus) numbers in
the Kruger National Park. Translocated cheetahs either dispersed out ofthe park or died. Caughley
and Walker (1983) suggest that the resident cheetahs were at their carrying capacity, imposed by
the competitive influence of lions (Panthera leoi. Mentis (1978) and Mentis and Duke (1976)
give further support to the existence of composite-species canying capacities with their finding that
1 Nagy (1987; 1994) showed that although estimates offield metabolic rate (FMR) scaled with mass. the
relationship varied both from that of B:MR and between eutherian mammals, marsupials and birds (a log-log
plot ofFMR against mass gave slopes of 0.81, 0.58 and 0.64 respectively - Nagy, 1987).
2 The superior competitive ability of lions has restricted the cheetah to a particular (realised) niche, which it
occupies to its capacity. Cheetahs and lions could then be said to have separate capacities. However, in the
absence of lions, cheetahs would probably increase in numbers (filling its fundamental niche). This niche
expansion is.a result of competitive release enabling the one species to increase its range in the absence of a
competitor that would otherwise have excluded it (Diamond, 1975).
36
Page 38
wild herbivores did not have greater canying capacities on natural Afiican veld types than domestic
cattle, although the debate continues (Fritz et al., 1996)1.
The estimation of composite-species canying capacities is dependent on the fact that mammals
have, within a narrow range, similar nutritional requirements when. these are expressed as a
proportion of their energetic requirements (Mentis and Duke, 1976 and Mentis, 1980). However,
subtle differences in resource requirements and assimilation efficiency between species (Du Toit
and Owen-Smith, 1989) may mean that the application of simple composite-species canying
capacities could result in dangerous conclusions being drawn about the ability of a habitat to sustain
a community'. Tables equating indigenous herbivores to stocking units, presenting figures such as
0.4 giraffe (Giraffe camelopardalis) or 4.9 bushpigs(Potamochoerus porcus) equivalent to a single
steer (e.g. Mentis, 1978; Mentis and Duke, 1976) expose the extremely simplistic nature of this
approach, although it is widely applied in livestock and game farming (e.g. Le Houerou, 1981).
1 Early thoughts on the theory behind carrying capacity for large herbivores in natural African vegetation
types suggested that the carrying capacity should be higher than that for domestic cattle, as the wild herbivores
consumed a greater variety of plant species (Mentis, 1980). Mentis and Duke (1976) found a large dietary
overlap among indigenous herbivores but identified dietary differentiation at the level of the plant part
consumed. Mentis (1980) concede that a system consisting of both browsers and grazers may achieve a higher
realisable biomass than simple systems made up of pure browsers or pure grazer species.
2 Evans et ai. (1979) calculated the energetic value of benthic intertidal invertebrate prey species at Seal
Sands. Great Britain and the energetic requirements of shorebirds (estimated as a multiple of BMR)
incorporating assimilation efficiency. It was calculated that if proposed development plans went ahead the.
area would only just be able to meet the energetic requirements of the species present. Furthermore, if species
specific prey requirements are taken into account, the welfare of the different species differed remarkably.
37
Page 39
5.2.2. Nutrition
The previous section highlights the danger of using energy consumption at the expense of other
factors to determine carrying capacity. Where food is shown to be a limiting factor, it is often the
supply of particular nutrients that is limiting, not energy. It was found that the Asian elephant
(Elaphas maximus) required a minimum of 5% protein in its diet (Tudge, 1994) and the snowshoe
hare (Lepus americanus) 11% (Sinclair et al., 1982). A more specific example is that of the feral
donkey (Equus asinusi in northern Australia. Freed from its natural enemies the population is
limited by nutrients criticalfor lactation, in particular, calcium (Freeland and Choquenot, 1990).
To calculate the carrying capacity for a particular species in a certain habitat accurately, it is
necessary to measure the dietary requirements of the species in question and the ability of the
environment to match them. The importance of considering the quality of diet ingested, not just the
quantity ingested, is stressed due to the often inverse relationshipbetween quality ofherbage and its
abundance (Hobbs and Swift, 1985). Furthermore, the concentration of nutrients in the diet is
generally the main factor influencing the condition of the herbivore! (e.g. see Cook et aI., 1996).
Without taking into account both nutritional constraints, the carrying capacity would be
overestimated (Focardi et al., 1995).
Two general approaches based on the measurement of nutritional resources have been used in.
estimating carrying capacity. The first uses the minimum necessary concentration of the limiting
nutrient, denoted as C* (Focardi et al., 1995). To calculate C* the mean requirement per day is
divided by the maximal intake of forage (Kg of dry matter). The second. approach takes into
account the multiple needs of the species (Hobbs and Swift, 1985) where nutrient concentration is
determined by quality of available food items and the mixture of these items in the diet.
Carrying capacity may therefore be calculated by assessing how much food is present in the
1 Quality of diet should also be considered when estimating predator populations. Often prey are thought of
as uniform when in fact they may differ in age and size (Sutherland, 1982).
38
Page 40
environment that will, on average, provide some specified nutritional level in the diet'.
Carrying capacity can then be estimated. by dividing this biomass of forage by the average
individual intake'. Both.approaches suffer from the problems associated with measuring specific
nutritional requirements (see Moen, 1973; Pietersen et al., 1993).
5.3.Problems of Open Systems
Due to the variation in the mobility of the various species comprising.a community, it is unlikely
that a completely closed system exists. Obviously the greater the number of linkages between
species, the harder it will be to define a closed system. However, it is possible to distinguish
between systems in which the majority of primary productivity is allochothonous (derived from
outside the community) and those in which the majority of primary productivity is autochthonous
(generated within the community). In the present context I refer to the former as an open system.
With the increase in ocean ranching, e.g. of salmon (Cooney, 1993), the question of carrying
capacity is becoming increasingly relevant to marine/coastal systems. In such systems much of the
primary production is born in the water column, consequently a large but unknown proportion of
food may not be ingested but drifts out of the system, confounding estimates of carrying capacity
for assemblages of suspension feeders (Edgar, 1993).
Attempts to estimate the carrying capacity of Norwegian fjords for fish, where the availability of
zooplankton was the crucial factor for juvenile cod and other gadid competitors, found no
1 This can be done by determining the biomass and nutrient content of the available forage species and
summing them, starting with the highest nutrient categories, until the threshold nutrient concentration is
reached (Hobbs and Swift, 1985).
2 This approach is based on two assumptions: firstly, animals will select high quality forage in preference to
low quality forage (however, see section 7). Secondly, the forage categories used match the distinctions made
by the animals themselves (Hobbs and Swift, 1985).
39
Page 41
correlationbetween simulated fisheries' yield and local primary production (Salvanes et al., 1995).
It appears that the advective supply of zooplankton swamps the effect of local production.
Consequently, variation in this transport process (e.g. by alterations in large scale circulation
patterns)will greatlyaffect the fish stock.
Such open systems can have a surprising impact on the carrying capacity of a particular habitat.
Some islands in the Gulfof California have unusually high spider densities, and, counter-intuitively,
small islands have higher densities than large islands. This is because allochthonous marine input
provides the primary source of prey for the consumers swamping in situ terrestrial productivity
(Polis and Hurd, 1995). A small island with a relatively large perimeter results in a greater
proportion of marine detritus washing up on the island, explaining the inverse correlation between
island size and spiderdensity.
5.4. The Construction of Indices Relating Carrying Capacity to Measurable Environmental
Parameters
If carrying capacity can be estimated, it may be possible to create indices relating it to
environmental parameters, enabling the construction of maps delineating. the spatial variation in
carrying capacity. The majority of such work has been done in connection with ranching (Field,
1978). Schmidt et al. (1995) list three attempts to construct an index relating grazing capacity to
measurable environmental parameters:
1. the rainfall method ofCoe, Cumming and Phillipson' (1976, cited in Schmidt et al., 1995);
2. the combined veld condition and rainfall method ofDanckwerts (1989, cited in Schmidt et al.,
1995); and
1 This relationship between rainfall and biomass was especially clear for regions receiving less than 700
mm of rainfall per year (Coe, 1980). Ironically, it is in these semi-arid environments where the whole concept
ofany one carrying capacity (no matter how defined) may be hard to apply due to the seasonal and annual
changes in rainfall resulting in a wildly fluctuating environment (see section 3.2.2).
40
Page 42
3. the herbaceous phytomass method ofMoore and Odendaal, 1987·(cited in Schmidt et al., 1993).
All three methods give different results when applied to the same area (Schmidt et al., 1995). It
therefore appears that even for the most well studied relationship, the identification of any general
rules remains elusive. The use of simple equations that relate shorebird number to some measurable
environmental parameter also suffers from several shortcomings; the parameter measured may not
be a limiting factor, the knock on effect of re-distribution of individuals are ignored (Goss-Custard
et al., 1994) and the problems associated with measuring resource availability (less problematic for
herbivores). These problems are compounded when attempts to estimate composite-species
carrying capacities are made.
It would appear that simple indices may be misleading when removed from the context in which
they were constructed. Stoddart (1952, cited in Bartels et al., 1993) goes as far as saying that
"there is no method whereby any technician can go.into a new country and measure anything which
will automatically give him the grazing capacity". I suggest that this also applies to measuring the
carrying capacity. Consequently, no single regression lineis effective at predicting the abundance of
individual populations (Peters and Raelson, 1984). However, this shortcoming does not detract
from the potential usefulness of the concept ofcarrying capacity to conservation issues, just that the
difficulties in quantifying carrying capacity should not be underestimated.
6. APPLICATIONS OF THE CONCEPT OF CARRYING CAPACITY TO
CONSERVATION RELATED ISSUES
This section deals with the issues of overpopulation, habitat evaluation, assessment of population
status and increasing population densities, identifying the relevance of carrying capacity to
conservation.
41
Page 43
6.1. Avoidance of the Problems Associated with Overpopulation
The importance in managing for overabundance in wild populationsis probably greater now than it
has ever been before. It has been suggested that, at least for domestic cattle, high population
densities may be maintained by the abilityof the animals to move adaptively between key resource
patches (Scoones, 1993). The construction of fences may inhibit this ability and has been partly
blamed for over-grazing in the northern Hluhluwe Game Reserve (Macdonald, 1980) and may
result in excessive population densities in times of drought in the Etosha National Park (Berry,
1981~ Owen-Smith, 1983)1.
Small reserves may be particularly vulnerable to damage caused by mammal overabundance as
often they are subject to a single climatic regime and therefore do not maintain the heterogeneity of
larger areas which buffer fluctuations in animal populations. For example, if climatic conditions
favour a particular species, it is likely that the species will increase throughout the whole reserve
(Macdonald and Brooks, 1983).
Recognition of whether an area is overpopulated depends on the stated goals, which may range
from conserving biodiversity to maximising the recreational value of a habitat (Miller and Child,
1983). As a consequence ofthese varied goals the definition ofoverabundance' varies considerably
1 It is thought that high densities of wild herbivores used to occur but for only short periods at any given
site.
2 The definition of overabundance implies that a habitat supporting a population below this density would
be free from degradation. However it can be argued that natural succession will occur no matter what the
density of herbivores (Dahlberg, 1993). Therefore, setting population limits should be based on what is deemed
to be acceptable limits to degradation of the land. An exception to this is when some threshold level exists
below which degradation is accelerated. Dahlberg (1993) suggests that soil loss may be an example of this with
an exponential increase in soil loss once vegetation cover is reduced by 35-400/0.
42
Page 44
and often is referenced to set goals! (Macdonald and Brooks, 1982; 1983; Macnab, 1985). It is
interesting to note that often these definitions do not imply that the population has exceeded its
carrying capacity (Macnab, 1985).
The problems associated with overpopulation stem from two facts: firstly, it is possible for a
population to exceed its carrying capacity; secondly, the environment (resource base). cannot
sustain a population above its carrying capacity indefinitely (Budd, 1992). If a system has several
stable states, separated by "hills and watersheds" a large perturbation, such as the heavy over
consumption of resources, may result in the transition from one stable state to another', It has long
been recognised that heavy grazing can alter the characteristics (both biotic and abiotic) of the
habitat (Collinson and Goodman, 1982; Milton, 1991; Dean et al., 1995). As plants differ in their
palatability, grazing is often selective, altering species composition and increasing plant diversity'.
It is becoming accepted that a large number of natural systems exhibit these types of dynamics
(May, 1977). It is therefore possible that continuous variation in a variable can produce
discontinuous effects. Often one state is more stable than the other (May, 1977) requiring more
1 Often the determined stocking rate is a compromise due to the conflicting nature of many of the
management goals such as those set for bison (Bison bison) in the Theodore Roosevelt National Park, where
two of the goals were to maximise viewing opportunities for visitors whilst minimising the damage to the
vegetation. (Norland et al., 1985).
2 Sinclair (1989) finds indirect evidence for the existence of multiple stable states (care should be taken not
to confuse this concept with that of stable equilibrium populations) in the literature e.g. the spruce budworm
(Choristoneurafumiferana) among other forest insects, salmonids, marine fish stocks, and wildebeest. Many of
these examples suggest that predation maintains the population below the carrying capacity (see section 3.4.1).
Escape from this low density to the higher density equilibrium may be through rapid immigration or climatic
effects temporarily favouring the species in question, i.e. by increasing the food supply (Sinclair, 1989).
3· This may act in a parallel manner to the intermediate disturbance effect, where a moderate level of
predation maximises the diversity of a system (e.g. see Schneider, 1978).
43
Page 45
effort to reverse'. As a result Odum (1983) suggests that in terms of the long term safety and
stability of the population, the ideal density is somewhere between carrying capacity and .the
optimum density (maximising productivity): he calls this the "desirable carrying capacity". This
density allows a margin of safety which buffers the population against environmental.fluctuations'.
LeRoux (1979) and Pietersen et al. (1993) suggests that herbivore populations should be managed
to ensure that they do not exceed the level where they can be sustained in the season of low dry
matter availability in the preferred area.
Adopting such a management policy would require "pre-emptive culling" to inhibit population
irruptions (Caughley, 1982). Owen-Smith (1983) suggests the creation of dispersal sinks, adjacent
to core conservation areas, may simulate the regulating effect of dispersal by creating "vacuum
zones" in which animals entering the zone are culled, mimicking theeffect of dispersal out of the
area. Although the. problems associated with culling appear to be numerous (Caughley, 1982;
Caughley and Walker, 1983), the alternative of letting nature "take its course" is meaningless
because the present day system of parks and reserves are not "natural" (Bell, 1982). Furthermore,
the course ofnature and our own aims may differ significantly.
6.2. Habitat Evaluation
By estimating the carrying capacity ofan area for a particular species, it is possible to assess the size
of a population that an area could support and thus whether it can support a minimum viable
I Many grazing-induced changes cannot be reversed by resting (e.g. changes in vegetation due to
overgrazing in the Karoo - Dean et al., 1995) as both structure and species composition may have altered and
a new dominant species may inhibit the re-establishment of the original species. Alternatively, changes of the
physical conditions may have occurred (Milton and Hoffman, 1994).
2 This density would be almost impossible to define empirically and, although it is appealing, may be of
little practical use.
44
Page 46
population (MVP) ofthe species'. This approach was adopted for harbour seals (Phoca vitulina) in
the Dutch Delta (Reijnders, 1994) and the black footed ferret (Mustela nigripes) (Biggins et al..
1993)2. By comparing estimates of carrying capacity for different habitats, it is possible to rank the
habitats on their ability to support populations of a particular species, enabling the identification of
those habitats that are ofgreater value, i.e. may have a higher conservation potential, and to predict
the effect of habitat loss on existing populations (without the need for long term demographic
studies).
If a population in an area is not already at the carrying capacity, then as habitat is lost, density will
increase in the remaining areas. Increased densities are likely to both increase interference. and
exploitative competition. The relative importance of these changes depends on the behaviour and
ecology of the species in question (Goss-Custard et al., 1995)3. Whether or not this will affect the
status of the population depends on whether density-dependent feedback mechanisms come into
effect", i.e. whether carrying capacity is reached.
1 This has often concentrated on absolute numbers independent of the environment, such as the genetically
based 50 and 500 rules (e.g. Reed et al., 1986), although the actual population size would have to be
considerable greater (Shaffer, 1987). However, defining a population as viable is relative and there is probably
no "magic number" (Gilpin and Soule, 1986). Consequently an approach based upon mathematical population
models may be more appropriate (Wissel et al., 1994).
2 Reijnders (1994) suggests that the estimatedcarrying capacity is not necessarily a figure that should beaimed
foe but that it may act as abench mark.
3 For example, food depletion is more likely to be a limiting factor in flock foraging birds than is
interference (Goss-Custard, 1985; Goss-Custard and Charman, 1976, cited in Alonso et al., 1994).
4 Goss-Custard et al. (1994) suggest that there may be a threshold density (cW) above which density
dependent mortality and emigration occurs. Habitat loss would only have a serious deleterious effect on the
population if it pushed population densities over this threshold (Goss-Custard et al., 1994; 1995).
45
Page 47
6.3.Assessing population status
It may be possible to establish whether apopulation is close to the carrying capacity by examining a
set of demographic or behavioural criteria'. Eberhardt and Siniff (1977) suggest that these criteria
may be used as benchmarks indicating the position of the population relative to carrying capacity'.
This is based on the assumption that .density-dependent population regulation alters life history
parameters depending on how close the population is to carrying capacity. It should therefore be
possible to use indices oflife history parameters to evaluate population status, i.e. a population near
carrying capacity would have relatively low birth rates compared to a population well below
carrying capacity.
The use of such attributes as indicators have been adopted by several authors. Examples include the
use of reproductive characteristics and population demography. for the Yellowstone grizzly bear
(Ursus aretos horribilis) (Mattson and Reid, 1991) and behavioural attributes of the giant panda
(Ailuropoda melanoleuea) (Johnson et al., 1988). The best indices appear to be those comprised of
a set of characteristics rather than just one or two (Chivers and DeMaster, 1994; Munkittrick and
Dixon, 1989). If a population is shown to be below canying capacity, investigations can be made
into why this should be so and appropriate action taken. Following a crash in the wildebeest
(Connoehaetes taurinus) population in Etosha National Park, the carrying capacity of the park
was estimated in terms of its ability to fulfil nutritional requirements of the wildebeest
1 Characteristics that reflect population status include: behavioural attributes - antagonistic/displacement
behaviour. time spent searching for food or in tending young, and shifts in dietary components; individual
responses - physical condition, incidence of disease and parasitism; reproductive characteristics - age at first
reproduction, annual reproductive rates of mature females; and population aspects - age structure, survival
rate, occupancy of marginal range, rate of change of population size, effects on habitat or food base (after
Eberhardt and Siniff, 1977).
2 As Potvin and Huot (1983) recognise, it is difficult if not impossible to identify a population below
carrying capacity in a poor quality habitat or a population above carrying capacity in a damaged habitat
without referring to some benchmark.
46
Page 48
population. It was found that the park could support the previous high densities of wildebeest.
Therefore, alternative reasons for decline where investigated. It was thought that increased
predation and mortality from anthrax following the creation of artificial watering holes was the
main cause of decline (Berry, 1981). If a population is found to be at carrying capacity, it
indicates that there is little chance of increasing the population under present habitat conditions
(Mattson and Reid, 1991)1.
6.4. Raising Population Densities
Once the limiting factors regulating population densities have been identified, it may be possible to
alleviate their effects, enabling the population to increase in size. However, many attempts to do
this, such as supplemental feeding of the white-tailed deer (Odocoileus virginianusi (Mcbryde,
1995) have ended in failure. To raise population densities successfully it may require the
manipulation ofa variety of factors simulating the characteristics of a highly productive habitat,
such as accomplished in commercial fish ponds (Biro, 1995).
A danger identifiedwith increasing population densities by the manipulation of a single resource is
that in years where food is scarce, the artificially high population densities may cause greater
damage to the habitat than the population densities that would naturally have existed in that area.
This degradation in habitat is a form of density-dependent feedback which involves a time lag,
hence irreversible damage to the vegetation and substratum may occur before the population
declines. This n:ay be particularly so if regulation occurs through changes in the natality, not
1 Although this method sounds attractive, some contradictory findings such as the decrease in the
proportion of pregnant female spinner dolphins (Stenella longirostris) over time in spite of increased mortality
due to the tuna fishery suggests that there may be a time lag in the response of the population (Chivers and
DeMaster, 1994). Furthermore, it is essential that monitoring important resource or habitat requirements is
undertaken to differentiate between changes in carrying capacity and in the demographics of the population
(population status) in which carrying capacity is constant (Chivers and DeMaster, 1994).
47
Page 49
mortality, ofthe species: the longer lived the species, the more of a problem this may be. Therefore,
an increase in the .density of a particular species is not only difficult to achieve, but, even if
successful, hidden knock-on ecosystem effects may occur'.
7. CONCLUSION
Carrying capacity is the maximum biomass that can be supported by the available resources in an
environment without the degradation of those resources through over-consumption.
Fundamentally, it is a measure of the energetic value of the consumable resources within an
environment. Due to temporal fluctuations in resource availability, carrying capacity should be
considered as a dynamic value which populations track to a greater or lesser extent. It can be
measured in units of available energy, biomass or population density. If either biomass or energy is
used to determine the carrying capacity, not only can the maximum supportable population density
be calculated, but a range of densities, each varying in the relative fitness of the individuals
contained within it, can be obtained due to the relationship between density and productivity
(White, 1993).
Theoretically, carrying capacities based on energy may be extended to encompass a range of
species (composite-species carrying capacities). However, due to interspecific differences in
resource requirements, such composite-species capacities should be considered as only theoretical
concepts, unless the species in question exhibit an unusually high degree ofniche overlap.
1 This may be particularly relevant to areas with inherently low carrying capacities such as the main arid
vegetation types in Australia (spinifex, Triodia spp.; mitchell grass, Astrebla spp.; saltbush, Atriplex; mulga,
Acacia aneura, Roberts, 1987) which appear to be especially susceptible to degradation.
48
Page 50
'The present system of isolated reserves has raised the issue of carrying capacity to one of high
priority. It is no longer possible for species to move opportunistically between high quality patches,
they are now often restricted within limited ranges. By assessing the carrying capacity ofa habitat it
is possible to both evaluate its importance in terms ofthe numbers ofa particular species that it can
sustain, and hence its conservation value, and also, avoid problems associated with over-utilisation
ofresources by ensuring the population does not exceed this capacity.
In this triage situation, where it is important to identify those habitats that have the greatest
potential value for conservation, the use of carrying capacity offers the chance to plan for the
future. Instead of evaluating an area by the number and diversity of species that occur there in the
present day, it is possible to estimate its potential future ability to support a species. By using
estimates of carrying capacity the problem of the unequal spatial distribution of resources
associated with using a minimum area requirement (MAR) (Shaffer, 1987) approach can be
avoided. It has been proposed that many reserves are undergoing a period of relaxation following
fragmentation and isolation. Consequently, the diversity of species and their relative abundance
represented today may be lost from the area in the near future. Although estimates of carrying
capacity cannot predict the risk of extinction directly it may be derived from knowledge of
minimum population requirements and the ability of the habitat to sustain them. It is perhaps in this
field that the use ofcarrying capacity has the greatest potential.
Page 51
REFERENCES
Allaby, M. (1992). The concise Oxforddictiornuy ofzoology. Oxford University Press, Oxford.
Allen, L.J. S. and Cormier, P. J. (1996). Environmentally drivenepizootics.Mathematical
Biosciences 131: 51-80.
Alonso,1. C., Alonso.J. A andBautista, L. M. (1994). Carrying capacityofstaging areasand
facultative migration extension in common cranes. Journal ofApplied Ecology31: 212
222.
Barlow, N. D. (1996). The ecology of wildlife disease control: simple models revisited. Journal of
Applied Ecology33: 303-314.
Bartels, G. B., Norton, B. E. andPerrier, G. K. (1993). An examination of the carrying capacity
concept, In (R. H. Behnke Jr, I. Scoones and C. Kerven eds) Range ecology at
disequilibrium - newmodels of natural variability and pastoral adaptation in African
Savannas, pp. 89-103. Overseas Development Institute, London.
Batzli, G. O. (1996). Population cycles revisited. Trends in Ecology and Evolution 11: 488-489.
Bautista L. M., Alonso, J. C. and Alonso, J. A. (1995). A field test of ideal free distribution in
flock-feeding common cranes. Journal of Animal Ecology 64: 747-757.
Begon, M. (1986). Is there a self-thinning rule for animal populations? Oikos 46: 122-124.
Begon, M., Harper, J. L. andTownsend, C. R. (1990). Ecology: individuals, populations and
communities. Blackwell Scientific Publications, London.
Begon, M. and Mortimer, M. (1981). Population ecology: a unified study of animals and plants.
Blackwell Scientific Publications, London.
Bell, R. H. V. (1982). Decision making in wildlife management with reference/to problems of
overpopulation. Proceedings and abstracts of the international conference on the
management of large mammals in African conservation areas. Large Mammal Symposium
Paper 9. Council for Scientific and Industrial Research, Pretoria.
Berry, H. H (1981). The wildebeest problem at Etosha . a review of the findings and
recommendations for management. Symposium on Wildlife Ecology. SouthWestAfrica
Namibia government. Nature Conservation Branch.
50
Page 52
Berryman, A. A and Millstein, 1. A. (1989). Are ecological systemschaotic - and if not why not?
Trends in Ecology andEvolution4: 26-28.
Biggins, D. E., Miller, B. 1.,Hanebury, L. R., Oakleaf, B., Farmer, A. H., Crete, R. and Dood, A.
(1993). A techniquefor evaluating black-footedferret habitat. Biological Report 13: 73-88.
Biro, P. (1995). Managementofpond ecosystems and trophic webs. Aquaculture 129: 373-386.
Branch, G. M. (1975). Intraspecific competition inPatellacochlear Born. Journal of Animal
Ecology 44: 263-281.
Briassoulis, H. (1992). Environmental impacts oftourism: a frameworkfor analysis and evaluation.
In (Briassoulis, H. and van der Streaten, 1. eds) Tourism and the environment. Kluwer
Academic Publishers, Dordrecht.
Buckland, S. T., Ahmadi, S., Staines, B. W., Gordon, I. J. and Youngson, R. W. (1996).
Estimatingthe minimum population size that allowsa given annualnumber of mature red
deer stags to be culled sustainably. JournalofApplied Ecology 33: 118-130.
Budd, W. W. (1992). What capacity the land? Journalofsoil and water conservation 47: 28-31.
abstracts ofthe international conference on the management oflarge mammals in African
conservation areas. Large Mammal Symposium Paper 8. Councilfor Scientific and
Industrial Research, Pretoria.
Caughley, G. (1979). What is this thingcalled carrying capacity? in (Boyce, M. S. and Hayden
Wing, L. D. eds) North American elk: ecology, behaviourand management, pp. 2-8.
University of Wyoming.
Caughley, G. and Sinclair, A. R. E. (1994). Wildlife ecologyand tnanagement. Blackwell Scientific
Publications, London.
Caughley, G. and Walker, B. (1983). Working with ecological ideas. In (A. A. Ferrar ed.)
Guidelines for the management of large. mammals in African conservation areas, .pp. 13-33.
South African National Scientific Programmes: Report 69.
Chivers, S. 1. and DeMaster, D. P. (1994). Evaluation of biological indices for three easternPacific
dolphin species. Journal of Wildlife Management 58: 470-478.
Coe, M. (1980). African wildlife resources. In (Soule, M. E. ed.) Conservation biology: an
evolutionary-ecological perspective. Sinauer Associates, Inc. Sunderland.
51
Page 53
Colinvaux, P. A. (1973). Introduction to ecology. JohnWiley and Sons inc. New York.
Collinson, R F. H. and Goodman, P. S. (1982). An assessment ofrange condition and large
herbivore carrying capacity of the Pilansberg GameReserve, with guidelines and
recommendations for management. INKWE NO.1. Environmental Researchin
Bophuthatswana.
Cooch,E. G., Lank, D. B., Rockwell, R. F. and Cooke, F. (1989). Long-term decline in fecundity
in a snow goose population: evidence for density dependence? JournalofAnimal Ecology
58: 711-726.
Cook, J. G., Quinlan, L. J., Irwin, L. L., Bryant, L. D., Riggs, R. A. and Thomas, J. W. (1996).
Nutrition-growthrelations of elk calves during late summer and fall. Journal ofWildlife
Management 60: 528-54l.
Cooney, R. T. (1993). A theoretical evaluation of the carrying capacity of PrinceWilliam Sound,
Alaska, for juvenile Pacific salmon. Fisheries Research 18: 77-87.
Cotgreave, P. (1993). The relationship betweenbody sizeand population abundance in animals.
Trends in Ecology andEvolution 8: 244-248.
Crete, M. (1989). Approximation ofK carrying capacity for moose in eastern Quebec. Canadian
Journal ofZoology 67: 373-380.
Curtis, H. (1983). Biology4th edition. Worth Publishers inc. New York.
Dahlberg, A. (1993). The degradation debate: is clarification possible? In (N. Abel ed.) Carrying
capacity, rangeland degradation and livestock development policy for the communal
rangelands of Botswana; pp. 1-9. Pastoral Development Network 35c.
Dale, B. W., Adams, L. G. andBowyer, R. T. (1994). Functional response of wolvespreying on
barren-ground caribou in a multiple-prey ecosystem. Journal of Animal Ecology 63: 644
652.
Dale, S. and Slagsvold, T. (1990). Random settlement offemale piedflycatchers, Ficedula
hypoleuca: significance of male territory size. Animal Behaviour 39: 231-243.
Dasmann, R. F. (1981). Wildlife biology 2nd edition. JohnWiley and Sons, New York.
Dasmann, R. F., Milton, 1. P. andFreeman, P. H. (1973). Ecological principles for economic
development. John Wiley and SonsLtd, London.
Dawkins, R. (1976). The selfish gene. Oxford University Press, Oxford.
52
Page 54
Dean, W. R 1., Hoffman, M. T., Kerley, G. I. H. andMilton, S. 1. (1995). Desertification in
developed countries: in search of th~ silver bullet. SouthAfrican Journal ofScience 91:
2132-215.
DeAngelis, D. L. and Waterhouse,1. C. (1987). Equilibrium and nonequilibrium concepts in
ecological models. Ecological Monographs57: 1-21.
DeLeeuw, P. N. and Tothill, 1. C. (1993). The conceptof rangeland carrying capacity in sub
SaharanAfrica- mythor reality. In (R. H. Behnke Jr, I. Scoones and C. Kerveneels)
Rangeecology at giseQuilibrium - new models of natural variability and pastoral
adaptation in African savannas, pp. 77-88. Overseas Development Institute, London.
Dennis, B. and Taper, M. L. (1994). Densitydependence intimeseries observations ofnatural
populations: Estimation and testing. Ecological Monographs 64: 205-224.
Dhondt, A. A. (1988). Carrying capacity: a confusing concept. ActaOecologia 9: 337-346.
Diamond, 1. M. (1975). Assembly rules of species and communities. In (M. L. Codyand 1. M.
Diamond, eds) Ecology and evolution of communities, pp. 342-444. Belknap Press,
Cambridge.
Dias, P. C. (1996). Sources and sinks in population biology. Trends in Ecology and Evolution 11:
326-330.
Du Toit,1. T. and Owen-Smith, N. (1989). Body size. population metabolism, and habitat
specialisation among large African herbivores. American Naturalist 133: 736-740.
Duqette, L. S. and Millar, 1. S. (1995). The effect of supplemental food on life-history traits and
demography ofa tropical mousePeromyscus mexicanus. Journal of Animal Ecology 64:
348-360.
Eberhardt, L. L. and Siniff D. B. (1977). Population dynamics and marine mammal management
policies. Journal ofthe Fisheries Research Board Canada 34: 183-190.
Edgar, G. 1. (1993). Measurement of the carrying capacity of benthic habitats using a metabolic
rate based index. Oecologia 95: 115-121.
Elgar, M. A. and Catterall, C. P. (1989). Density-dependent natural selection. Trends in Ecology
andEvolution4: 95-96.
Elliot, 1. M. (1993). The self-thinning rule applied to juvenile sea-trout Salmo trutta. Journal of
Animal Ecology 62: 371-379.
53
Page 55
Elton, C. S. (1958). The ecology ofinvasion by animals and plants. Methuen, London.
Emlen, 1.M. (1973). Ecology: an evolutionary approach. Addison-Wesley, ReadingMass.
Evans, P. R., Herdson, D. M., Knights, Pl. 1. and Pienkowski, M. W. (1979). Short-termeffects of
reclamation of part of Seal Sands, Teesmouth, on wintering waders and shelduck.
Oecologia 41: 183-206.
Field, D. I. (1978). Potential carrying capacityof rangeland inBotswana. Ministry of Agriculture.
Focardi, S.,Poli,B. M. and Tinelli, A. (1995). The nutritional carrying capacity offour
Mediterranean habitats for fallow deer (Dama dama). Revue d'Ecologie 50: 97-107.
Freeland, W. 1. and Choquenot, D. (1990). Determinants of herbivore carrying capacity: plants,
nutrients, andEquus asinus in northern Australia. Ecology71: 589-597.
Fritz, H., Garine-Wichatitsky, M. De. and Letessier, G. (1996). Habitatuse bysympatric wildand
domestic herbivores in an African savanna woodland: the influence of cattle spatial
behaviour. Journal of Applied Ecology 33: 589-598.
Giles, Jr, R. H. (1978). Wildlife management. W. H. Freeman and Company, SanFrancisco.
Gilpin, M. E. and Soule, M. E. (1986). Minimum viable populations: processes of species
extinction. In (M. E. Souleed.) Conservation biology: the science of scarcity and diversity.
Sinauer Associates, Inc. Publishers, Massachusetts.
Ginzburg, L. R., Ferson, S. and Akcakaya, H. R. (1990). Reconsrtructibility of density dependence
and the conservative assessment of extinction risks. Conservation Biology 4: 63-70.
Goss-Custard, J. D. (1985). Foragingbehaviour of wading birdsandthe carrying capacity of
estuaries. In (R. M. Sibly andR. H. Smith eds)Behavioural ecology: ecological
consequences of adaptive behaviour, pp. 169-188. Blackwell Scientific Publications,
London.
Goss-Custard, 1.D., Caldow, R. W. G., Clarke, R. T., Durell, S. E. A. LeV., Urfi, 1. and West, A.
D. (1994). Consequences of habitat loss and change to populations of wintering migratory
birds: predicting the local andglobal effects from studies of individuals. Ibis 137: S56-S66.
Goss-Custard, 1.D., Caldow, R. W. G., Clarke, R. T. and West, A. D. (1995). Deriving population
parameters from individual variations in foraging behaviour. II. Modeltests and population
parameters. Journal of Animal Ecology64: 277-289.
, 54
Page 56
Grobler, 1. H. and Jones, M. A. (1980). Populationstatistics and carrying capacityoflarge
ungulates in the WhoviWildArea, Rhodes Matapos National Park, ZimbabweRhodesia.
South AfricanJournalofWildlife' Research 10: 38-42.
Hassell, M. P. (1975). Density-dependence in single-species populations. Journal ofAnimal
Ecology 44: 283-295.
Hassell, M. P. (1986). Detecting density dependence. Trends in Ecology and Evolution 1: 90-93.
Hickman, Jr. C. P., Roberts, L. S. and Larson, A. (1993). Integrated principles ofzoology 9th
edition. Mosby, Missouri.
Hill, D. and Robertson, P. (1988). The pheasantecology~ managementand conservation. BSP
ProfessionalBooks, Oxford.
Hobbs, N. T. and Hanley, T. A. (1990). Habitat evaluation: do use/availability data reflectcanying
capacity? Journal ofWildlife Management 54: 515-522.
Hobbs, N. T. and Swift, D. M. (1985). Estimatesofhabitatcarryingcapacity incorporating explicit
nutritionalconstraints. Journalof Wildlife Management49: 814-822.
Hoffinan, M. T., Sonnenberg, D., Hurford, 1. L. and Jagger, B. W. (1995). The ecologyand
management ofRiemvasmaaks' s natural resources. National Botanical Institute of South
Africa.
Hornfeldt, B. (1994). Delayeddensity dependence as a determinant ofvole cycles. Ecology 75:
791-806.
Hughes, R. N. and Griffiths, C. L. (1986). Application of the self-thinning rule to animal
populations. Unpublished M.S.
Igbozurike, U. M. (1981). The concept of carrying capacity. Journal of Geography 80: 141-146.
Izac, A. M. N., Anaman, K. A. and Jones, R. (1990). Biological and economic optima in a tropical
grazing ecosys~em in Australia. Agriculture, Ecosystemsand Environment 30: 265-279.
Johnson, K. G., Schaller, G. B. and Jinchu, H. (1988). Responses ofgiant pandas to a bamboodie
off National Geographic Research 4: 161-177.
Kacelnik, A., Krebs, 1. R. and Bernstein, C. (1992). The ideal free distribution and predator-prey
populations. Trends in Ecology and Evolution 7: 50-55.
Kataoka, M., Ibaraki, K. and Tokunga, H. (1989). Populationregulationof rhodegrass cultivars in
sward conditionsI. Genet density. Journal of Applied Ecology 26: 273-283.
55
Page 57
Kaufinann, 1.H. (1983). On the definitions and functions ofdominance and territoriality. Biological
Review 58: 1~20.
Komdeur, 1.(1994). Conservingthe Seychelles warbler Acrocephalus sechellensis by translocation
from Cousin Island, to the islandsofAride and Cousine. Biological Conservation 67: 143~
152.
Komdeur,1. (1996). Breeding ofthe Seychelles Magpie Robin Copsychus sechellarum and
implications for its conservation. Ibis 138: 485-498.
Kormondy, E. 1. (1976). Concepts ofecology. Prentice-Hall, Englewood Cliffs, N. 1.
Le Houerou, H. N. (1981). How to calculate livestockconversion factors in the Libyan rangelands,
FAO Project Lib/O18 TechnicalNote NO.1: 1-15.
Lekoux, C. 1. G. (1979). The grazing capacity ofthe plains in the Etosha NationalPark.
Proceedings of the Grassland Society of Southern Africa 14: 89-93.
Macdonald, I. A. W. (1980). The problem oflocalised overgrazing following fire in the Northern
HluhluweGame Reserve - The example ofQololenja Hill, 1978to 1980. Unpublished
M.S.
Macdonald, I. A. W. and Brooks, P. M. (1982). Monitoringfor the detection ofoverabundancein
mammals - the information deficit. Proceedingsand abstractsofthe.international conference
on the management of large mammals in African conservation areas. Large Mammal
Symposium Paper 11. Council for Scientific and Industrial Research, Pretoria.
Macdonald, I. A. W. and Brooks, P. M. (1983). Monitoring for the detection ofmammal
overabundancein small conservationareas. In Management of large mammals in African
conservation areas, pp. 187-200. Haum EducationalPublishers, Pretoria.
Macnab, 1. (1985). Carryingcapacity and related slippery shibboleths. Wildlife SocietyBulletin 13:
403-410.
Marquet, P. A., Navarrete, S. A. and Castilla, 1. C. (1995): Body size, populationdensity, and the
Energetic EquivalenceRule. Journal of Animal Ecology64: 325-332.
Massot, M., Clobert, 1., Pilorge, T., Lecomte, 1. and Barbault, R. (1992), Density dependence in
the common lizard: demographicconsequences of a density manipulation. Ecology 73:
1742-1756.
56
Page 58
Mattson, D. J. and Reid, M. M. (1991). Conservation ofthe Yellowstone grizzly bear.
Conservation Biology5: 364-372.
May, R. M. (1977). Thresholds and breakpoints in ecosystems with a multiplicity of stablestates.
Nature 269: 471-477.
McBryde, G. L. (1995). Economics ofsupplemental feeding andfood plots for white-tailed deer.
Wildlife Society Bulletin 23: 497-501.
McCallu~ H. andDobson, A. (1995). Detectingdisease and parasite threats to endangers species
and ecosystems. Trends in Ecology and Evolution 10: 190-194.
Mentis, M. T. (1977). Stockingrates and carrying capacities for ungulates on African rangelands.
South African Journal of Wildlife Research7: 89-98.
Mentis, M. T. (1978). Economically optimal species - mixes and stocking rates for ungulates in
SouthAfrica. In (Hydered.) Proceedings of the first international rangeland congress, pp.
146-149. DenverCo.
Mentis, M. T. (1980). The effect of animal sizeand adaptation on defoliation, selective defoliation,
annual production and veld condition. Proceedings of the Grassland Society of Southern
Africa 15: 147-151.
Mentis, M. T. andDuke, R. R. (1976). Carrying capacities of natural veld inNatal for large wild
herbivores. South African Journal of Wildlife Research 6: 65-74.
Messier, F. (1994). Ungulate population models with predation: a case studywith the North
American moose. Ecology75: 478-488.
Miller, K. and Child, G. (1983). Goalsfor protected areas. In (A. A. Ferrar ed.) Guidelines for the
management of largemammals in African conselVation areas, pp. 1-11. South African
Scientific Programmes; Report 69.
Milton, S. J. (1991). Plant spinescence in arid southern Africa: does moisture mediate selection by
animals? Oecologia 87: 279-287.
Milton, S. J. andHoffinan, M. T. (1994). The application of state-and-transition models to
rangeland research and management in arid succulent and semi-arid grassy Karoo, South
Africa. African Journal of Range andForestryScience 11: 18-26.
Moen,A. N. (1973). Wildlife ecology: and analytical approach. W. H. Freeman and company, San
Francisco.
57
Page 59
Montgomery, W. I. (1989). Population regulation in the wood mouse,Apodemussylvaticus. I.
Density dependence inthe annual cycleofabundance. Journalof Animal Ecology 58: 465-
475.
Munkittrick, K. R. andDixon, D. G. (1989). A holistic approachto ecosystem health assessment
usingfish population characteristics. Hydrobiologia 188/189: 123-135.
Murray, Jr. B. G. (1982). On the meaning ofdensity dependence. Oecologia 53: 370-373.
Nagy,K. A. (1987). Field metabolic rates and food requirement scaling in mammals and birds.
Ecological Monographs 57: 111-128.
Nagy, K. A. (1989). Fieldbioenergetics: accuracy ofmodels and methods. Physiological Zoology
62: 237-252.
Nagy, K. A. (1994). Field bioenergetics of mammals: what determines field metabolic rates?
Australian Journal of Zoology42: 43-53.
Nanni, U. W. (1979). Veldmanagement in the Umgeni Valley Nature Reserve. Proceedings of the
Grassland Society of Southern Africa 14:95-99.
Nilsson, C. and Grelsson, G. (1995). The fragility of ecosystems: a review. Journal of Applied
Ecology32: 677-692.
Norland, 1. E., Irby, L. R. andMarlow, C. B. (1985). Determination ofoptimum bisonstocking
rate inTheodoreRoosevelt National Park, North Dakota. Journal of Environmental
Management 21: 225-239.
Odum, E. P. (1983). Basicecology. CBS College Publishing, New York.
Odum, E. P. (1971). Fundamentals of ecology 3fd edition. Saunders College Publishing,
Philadelphia.
Owen-Smith, R.. N. (1983). Dispersal and the dynamics of largeherbivores in enclosed areas: I
implications for management. In (R. N. Owen-Smith ed.) Management of large mammals in
African conservation areas, pp. 127-143. Haum, Pretoria.
Paine, R. T., Castillq, 1. C. and Cancino, 1. (1985). Perturbations and recovery patterns of starfish
dominated intertidal assemblages in Chile, New Zealand, and Washington state. American
Naturalist 125: 679-691.
Peters, R. H. (1993). A critique for ecology. Cambridge University Press, Cambridge. p. 75.
58
Page 60
Peters, R. H. and Raelson, lV. (1984). Relation between individual size and mammalian
population density. The American Naturalist 124: 498-517.
Pianka, E. R (1970). On r- andk- seleclion. American Naturalist 104: 592-597.
Pietersen, L. M.,Meissner, H. H. andPietersen, E. W.(1993). Food selectionand intakeby male
impalasin the Timbavati area. SouthAfrican Journal of Wildlife Research23: 6-11.
Pimm, S. L. (1991). The balance of nature? Ecological issues in the conservation of species and
communities. University ofChicago Press, Chicago.
Polis, G. A. (1994). Food webs, trophiccascades and community structure. Australian Journal of
Ecology 19: 121-136.
Polis, G. A. and Hurd, S. D. (1995). Extraordinary high spiderdensities on islands: Flow of energy
from the marineto terrestrial food websandthe absence of predation. Proceedings of the
National Academy of Sciences, USA 92: 4382-4386.
Potvin, F. and Huot, 1. (1983). Estimating carrying capacity ofa white-tailed deer wintering area in
Quebec. Journal of Wildlife Management 47: 463-475.
Raillard, O. and Menesguen, A. (1994). An ecosystem box model for estimating the carrying
capacityofa macrotidal shellfish system. Marine Ecology Progress Series 115: 117-130.
Reed,M. J., Doerr, P. D. and Walters, J. R. (1986). Determining minimum population sizes for
birds and mammals. Wildlife Society Bulletin 14: 255-261.
Reeves, S. A and Usher, M. B. (1989). Application ofa diffusion modelto the spreadofan
invasive species: the coypuin GreatBritain. Ecological Modelling 47: 217-232.
Reijnders, P. 1. H. (1994). Historical population size of the harbourseal, Phoca vitulina, in the
Delta area, SW Netherlands. Hydrobiologia 282/283: 557-560.
Reiss, M. (1988). Scaling of homerange size: Body size, metabolic needs and ecology. Trendsin
Ecology and Evolution 3: 85-86.
Roberts, B. (1987). Australian land degradation andits control. Ambio 16: 272-276.
Rodriguez, D. 1. (1989). A model of population dynamics for the fruit fly Drosophilamelanogaster
with densitydependence in morethan one life stageand delayed density effects. Journal of
Animal Ecology 58: 349-365.
Srether, B., Anderson, R., Hjelord, O. andHeim, M. (1996). Ecological correlates of regional
variation in life history of the moose Alcesalces. Ecology 77: 1493-1500.
59
Page 61
Salvanes, A G. V., Aksnes, D., Fossa,1. H. and Giske, 1. (1995). Simulated carrying capacities of
fish in Norwegianfjords. Fisheries Oceanography 4: 17-32.
Schaffer, W. M. and Kot, M. (1986). Chaos in ecological systems: the coals that Newcastle
forgot. Trends in EcologyandEvolution 1: 58-63.
Schmidt, A G., Theron, G. K. andHoven, W. Van. (1995). A comparison ofsome methodsused
to estimate the grazing capacity ofa game ranch in Northern Province, South Africa.
Koedoe 38: 123-128.
Schneider, D. (1978). Equalisation ofpreynumbers by migratory shorebirds. Nature 271: 353-354.
Scoones, I. (1993). Why are there so many animals? Cattle population dynamics in the communal
areas ofZimbabwe. In (R H. Behnke, Jr, I. Scoonesand C. Kerven eds).Rangeecology at
disequilibrium - new models ofnatural variability and pastoral adaptation inAfiican
savannas, pp. 62-76. Overseas Development Institute, London.
Shaffer, M. (1987). Minimum viable populations: copingwith uncertainty. In (M. E. Soule ed.)
Viable populations for conservation. Cambridge University Press, Cambridge.
Silva, M. and Downing, 1. A (1995). The allometric scaling ofdensity and body mass: A non-linear
relationship for terrestrial mammals. The American Naturalist 145: 704-727.
Sinclair, A. R. E. (1989). Population regulation in animals. in (Cherret, 1. M. ed.) Ecological
Concepts. pp. 197-241. Blackwell Scientific Publications, Oxford.
Sinclair, A. R.E., Krebs, C. 1. and Smith, 1. N. M. (1982). Diet quality and food limitation in
herbivores: the case of the snowshoe hare. Canadian Journal ofZoology 60: 889-8?7.
Sinclair, A. R. E. and Norton-Griffiths, M. (1982). Does competition or facilitation regulate
migrant ungulate populations in the Serengeti? A test of hypotheses. Oecologia 53: 364
369.
Sinclair, A. R. E. and Pech, R. P. (1996). Density dependence, stochasticity, compensation and
predator regulation. Oikos75: 164-173.
Smith, R. L. (1966). Ecology andfield biology. Harper and Row Publishers, New York.
Solomon, E. P., Bery, L. R., Martin, D. W. and Villee, C. (I993). Biology 3rd edition. Saunders
College Publishers, London.
Strong, D. R. (1986). Density-vague population change. Trends in Ecologyand Evolution I: 39
42.
60
Page 62
Suter, W. (1995). Are connorantsPhalaeroeorax earbo wintering in Switzerland approaching
canyingcapacity? An analysis of increase patterns and habitat choice. Ardea83: 255-266.
Sutherland, W. 1. (1982). Spatial variation in the predationofcocklesby oystercatchers at Traeth
Melynog, Anglesey. I. The cocklepopulation. Journal ofAnimal Ecology5l: 481-489.
Sutherland, W. 1. and Parker, G. A. (1985). Distribution ofunequal competitors. In (R.M. Sibly
and R. H. Smith eds)Behavioural ecology: Ecological consequences.of adaptive behaviour.
pp. 255-273. Blackwell Scientific Publications, Oxford.
Talbot, L. M. (1977). Wildlife quotas sometimes ignoredthe realworld. Smithsonian year 8: 116
12l.
Torok, 1. and T6th, L. (1988). Density dependence in reproductionofthe collared flycatcher
(Fieedula albicollisi at high population levels. Journal ofAnimal Ecology57: 251-258.
Trollope, W. S. W., Trollope, L. A. andBosch, O. 1.H. (1990). Veld and pasture management
terminology in southern Africa. Journal of the Grassland Societyof Southern Africa 7: 52
6l.
Trombulak, S. C. (1990). Assessment of territoryvalueby a tropicalhummingbird (Amazilia
saueerottei). Biotropica 22: 9-15.
Tudge, C. (1994). Asia's elephants no placeto hide. New Scientist 141: 34-37.
van Zyl, A. 1. (1992). Do breeding pairs of martial eagleshold "super-normal" territories? Gabar
7: 9-10.
Walker, B. H. (1982)Estimation of stocking rates for multi species communities. M.S.
Unpublished, University of Witwatersrand.
Watson. A. and Jenkins, D. (1968). Experiments on population controlby territorial behaviour in
red grouse. Journal of Animal Ecology36: 595-614.
Watson, A., Moss, R..andParr,R. (1984). Effects offood enrichment on numbers and spacing
behaviour of red grouse. Journal of Animal Ecology 53: 663-678.
Watson, A. and O'Hare, P. 1. (1979). Red grouse populations on experimentally treated and
untreatedIrishbog. Journal of Applied Ecology 16: 433-452.
Weeden, R. (1974). Technology andwildlife. Technology Review76: 38-45.
Weisz, P. B. andKeogh, R. N. (1982). The science ofbiology 5th edition. McGraw-Hill Book
Company, New York.
61
Page 63
Westoby, M. (1984). The self-thinning rule. Advances in EcologicalResearch 14: 167-225.
White, L. 1. T. (1994). Biomass ofrain forest mammals in the Lope Reserve, Gabon. Journalof
Animal Ecology 63: 499~512.
White,R (1993). Comments. In (N. Abeled.) Carrying capacity, rangelanddegradationand
livestockdevelopmentpolicyfor the communal rangelands ofBotswana, pp. 15-18.
Pastoral DevelopmentNetwork 35c.
Whittaker,R. H. (1972). Communities and ecosystems. The Macmillan company, London.
Wissel, C., Stephan, T and Zaschke, S. H. (1994). Modelling extinctionand survival ofsmall
populations. In (H. Remmert ed.) Minimum animal populations. Ecological Studies 106:
67-103. Springer-Verlag, Berlin.
Zhang, L., Moore, 1. A. and Newbeny, 1. D. (1993). Estimatingasymptoticattributes offorest
stands based on bio-mathematical rationales. Ecological Applications 3: 743-748.
Zorn, T. G. and Seelbach, P. W. (1995). The relation between habitat availability and the short
term canying capacityof a streamreachfor smallmouth bass. North American Journalof
Fisheries Management 15: 773-783.
62
Page 64
APPENDIX A
Definitions ofcarrying capacity citedin the literature:
"...the number of stock which a range will support for a definite period ofgrazing without injury to
the range." (Hadwen and Palmer, 1922, cited inBartels et al., 1993).
"...the number of stock of one or more species which an area will support in good condition during
the time that the forage is palatable and accessible, without decreasing the forage production in
subsequent seasons." (Sampson, 1923, cited inBartels et al., 1993).
"...the maximum number of grazing animals of a given class that can be maintained in good flesh
year after year on a grazing unit without injury to the range forage stock or to the basic resource."
(Dasmann, 1945, cited in Bartels et al., 1993).
"...the maximum standing crop that can maintain itself indefinitely in an area" (Clarke, 1954, cited in
Dhondt, 1988).
"...the maximum number of animals of a given species and quality that can in a given ecosystem
survive through the least favourable environmental conditions occurring within a stated time
interval. For practical purposes this time interval is usually one year." (Edwards and Fowle, 1955,
cited in Bartels et al.,. 1993).
"The stocking rate at the optimum grazing pressure" (Mott, 1960, cited in Bartels et aI., 1993).
"In its true sense, the maximum number of individual animals that can survive the greatest period of
stress each year on a given land area. It does not refer to sustained production. In range
management, the term has become erroneously synonymous with grazing capacity." (Society for
Range Management, 1964, cited in Bartels et al., 1993).
63
Page 65
"...the total number of individuals of a species that will live in an ecosystem (or habitat) under
certain conditions" (Knight, 1965).
"...the maximum number that can be supported in a given habitat." (Smith, 1966)
"...the number of animals of a specified type that can subsist on a unit area and produce at a
required rate over a specified period, usually a season, a year, or longer." .(Cowlishaw, 1969, cited
in Bartels et al., 1993).
"...the population of animals of a particular size that, over a long period of time, can be supported
by the vegetation growing in the region without adversely affecting vegetation production" (Talbot
et al., 1970, cited in Igbozurike, 1981).
"...the maximum population that can be sustained in a habitat without the degradation of the life
support systems" (Odum, 1971, cited in Budd, 1992).
The "number of individuals which an environment can support". (Emlen, 1973).
cc..• the ability of an area or ecosystem to absorb stress without disruption of its basic properties"
(Weeden, 1974).
"The maximum stocking rate possible which is consistent with maintaining or improving vegetation
or related resources. It may vary from year to year on the same area due to fluctuating forage
production." (Society.for Range Management, 1974, cited in Bartels et al.,.1993).
"The population density that the environment can support on a continuing, steady-state basis"
(Whittaker, 1975, cited in Igbozurike, 1981).
64
Page 66
~' ...the user-specified quality biomass of a particular species under the influence of social or
behavioural constraints, for which a particular area, having user-specified objectives, will supply all
energetic and physiological requirements over a long (but specified) period" (Giles, 1978).
" ...the point where the rate of production of forage equals the rate at which. that forage is
consumed." (Caughley, 1979)
" ... the equilibrium between animals and vegetation and the position of that equilibrium as,indexed
by its characteristic density ofanimals" (Collinson and Goodman, 1982).
" ...the maximum sustainable density" (Weisz and Keogh, 1982).
The "density a population assumes over a long period of time if it is not harvested" (Caughley,
1983).
" ...the density at which a population will stabilise if it is left alone." (Caughley and Walker, 1983)
" ...the number at which the population stabilises" (Curtis, 1983).
" ...the size (biomass) of a population that can be supported on a long-term sustainable basis without
detriment to the non-renewable resources ofa given area or its productivity" (Evans, 1984).
"The maximum number of organisms that can be supported in a given area or habitat: usually
denoted by K; the uPI:er asymptote of the logistic equation" (Lincoln et al., 1985).
"The population size for which the population growth rate is zero" (Hobbs and Hanley, 1990).
"...the total amount of [consumerable forage] produced and available for grazing from...a specified
land area, [including] harvested forages and even other feedstuffs used in conjunction with grazing,
65
Page 67
thereby providing a means of summarising total ranch oranimal enterprise capacity" (Vallentine,
1990, cited in Bartels et al., 1993).
"The maximum population size that can be supported 'indefinitely by a given environment, at which
intraspecific competition has reduced that per capita net rate of increase to zero. An idealised
concept not to be taken literally in practice" (Begon et al., 1990).
"Potential of an area to support livestock through grazing and/or browsing and/or fodder
production over an extended number of years without deterioration to the overall ecosystem".
(Trollope et aI., 1990).
"...the optimum stocking level to achieve specific objectives given specified management options."
(Scamecchia, 1990, cited in Bartels et aI., 1993).
"The maximum population of a given organism that a particular environment can sustain" (Allaby,
1992).
" ...the number of individuals that can be supported in a given area" (Budd, 1992).
"...the population that can be sustained by the resources which the animals use within a particular
area". (Bartels et al., 1993).
"...the maximum number oforganisms that a habitat can support" (Solomon et al., 1993).
The "maximumdensity the environment can support" (Hickman et al., 1993).
The "natural limit of a population set by resources in a particular environment". (Caughley and
Sinclair. 1994).
The "equilibrium population density in the absence ofdisease" (Barlow, 1996).
":.the population size reached at equilibrium in a given area" (Dias, 1996).
66
Page 68
APPENDIXB
Table indicating the emphasis vanous authors have placed on the definition of carrying
capacity. The column headings state. the different issues that have been included in definitions
of carrying capacity. An X indicates that the author has included that issue in his or her
definition.
What factors set the carrying capacity:
Reference Temporal Temporal Maximum
IAnthropogenic Unspecified Carrying
environmental variation in number goal limiting capacityisvariation only carrying supported determined factors a
capacity by the density sustainableresources population
density
Hadwen and Palmer, 1922 1 X X
Sampson, 1923 1 X X X
Dasmann, 1945 1 X X
Clarke. 19542 X X
Edwards and Fowle. 1955 1 X X
Mott, 1960 1 X
Society for range X Xmanagement 1964 1
Knight. 1965 X
Smith. 1966 X
Cowlishaw, 1969 1 X
Talbot et al., 19703 X X
Odum. 1971-1 X" X
Emlen. 1973 X
Weeden. 1974 X
Society for range X X Xmanagement. 19741
Whittaker, 1975 3 X X
Giles. 1978 X
Caughley, 1979 X X
67
Page 69
Collinson and Goodman, X X
1982
Weisz and Keogh, 1982 X X
Caughley, 1983 X
Caughley and Walker, 1983 X
Curtis, 1983 X
Evans, 1984 X X
Lincoln et al., 1985 X
Hobbs and Hanley, 1990 X
Vallentine, 19901 X
Begon et aI., 1990 X X
Trollope et aI., 1990 X X
Scarnecchia, 19901 X
Allaby, 1992 X
Budd. 1992 X
Bartels et al., 1993 X X
Solomon et al., 1993 X
Hickman etal., 1993 X
Caughley and Sinclair, X1994
Barlow, 1996 X
1 Cited in Bartels et al.(1993).2 Cited in Dhondt (1988).3 Cited in Igbozurike (1981).4 Cited in Budd (1992).
NB. A blank indicates that the authors have not included this factor into their definition.
As apparent from the above table the majority of authors have placed stress on the carrying capacity as a
function of sustainability, I.e. no degradation of the habitat occurs from animals stocked at this density. Most
definitions also see carrying capacity as the maximum population density that the environment can support (I
have taken this to he an indication that this density is set by resources). Little attention has been given to either
the variability of the environment or the variability of carrying capacity tracking the environmental
fluctuations.
68