35
9. Abbau der Glukose zum Pyruvat 236

9.
Abbau der Glukose zum Pyruvat
236

Presence of O2 (aerobic)
Absence of O2 (anaerobic)
Pentosephosphate
Pathway (PPP)Glycolysis
9.1. Übersicht: Abbau von Glucose
Oxidative
Phosphorylation Alcoholic
Fermentation
NADP+
NADPH
Homolactic
Fermentation
Glucose
237
Citric Acid
CycleCitric Acid
Cycle
PDH
Complex

9.2. Glycolyse
Hauptabbauweg der Glucose in (fast) allen Organismen
Zweck: Gewinnung von ATP und NADH
Gesamtreaktion:
2 NADH
ADP +
Pi
(inorganic phosphate) 2 ATP
2 22 NAD+
CC
CCC
C
Glucose
C C CPyruvat
238
-I
±0
±0 ±0
+1
±0
S Oxidationstufen der C-Atome:
-I + 4(±0) + -I = ±0
S Oxidationstufen der C-Atome:
2(-III + +II + +III ) = +IV
-III
+II
+III
Beim Abbau der Glucose in der Glycolyse werden den C-Atomen durch NAD+
4 Elektronen entzogen

9.2.1. Einzelreaktionen der Glycolyse
239

240
Die Glycolyse ist ein Beispiel für Substratkettenphosphorylierung:
der schrittweise Abbau/Umbau eines organischen Moleküls wird gekoppelt mit
der Bildung eines phosphorylierten Zwischenprodukts, das in der Lage ist, ADP
zu ATP zu phosphorylieren
Ablauf und Zweck der Reaktionsschritte in der Glycolyse
▪ Schritt 1: Hexokinase Reaktion
Festhalten” der Glucose im Cytosol
▪ Schritt 2: Phosphoglucomutase (PGM) Reaktion
Umgestaltung des C6-Molekülgerüsts, um in Schritt 4 die Spaltung in zwei
C3-Moleküle zu ermöglichen
▪ Schritt 3: Phosphofruktokinase (PFK) Reaktion
Einführung einer zweiten negativ geladenen Gruppe, um die Spaltung des
Molekülgerüsts in Schritt 4 zu begünstigen
▪ Schritt 4 & 5: Aldolase Reaktion und Triosephosphatisomerase (TIM) Reaktion:
Erzeugung von 2 identischen Aldehyden, die in Schritt 6 oxidiert werden
können
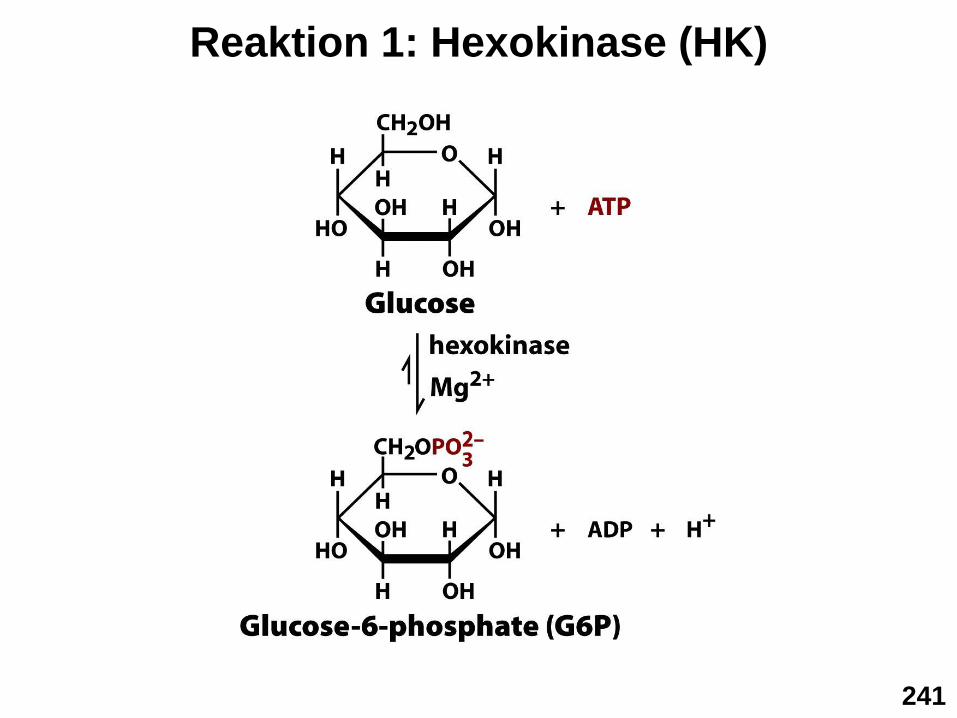
Reaktion 1: Hexokinase (HK)
241

Kinase Substrat: ATP-Mg2+-Komplex
242

Reaktion 2: Phosphoglucoisomerase (PGI)
243

PGI Reaktion: Säure-Base Katalyse
B+ = ε-NH3+-Gruppe
von K
B’ = Imidazol-Gruppe
von H
244

Reaktion 3: Phosphofructokinase (PFK)
245

Reaktion 4: Aldolase
246

Mechanismus der Aldolase Reaktion
Säure-Base Katalyse durch b-Carboxylatgruppe
von D-Rest im aktiven Zentrum
-COO- + H+ -COOH
247

Reaktion 5: Triosephosphatisomerase (TIM)
248

249
▪ Zwischenbilanz der Glykolyse nach Ablauf der Schritte 1-5:
Energie: -2 ATP
Elektronen: ±0 NADH C-Atome in GAP auf gleicher Oxid.stufe wie Glucose
▪ Schritt 6: Glycerinaldehydphosphat Dehydrogenase (GAPDH) Reaktion
Oxidation der Aldehydgruppe mit gleichzeitiger Phosphorylierung der
entstandenen Carbonsäure → Bildung von NADH
→ Bildung eines energiereichen
Phosphoanhydrids (ohne ATP Verbrauch!)
Warum ist es in der GAPDH Reaktion thermodynamisch möglich, dass
eine energiereiche Phosphorylverbindung entsteht?
Oxidation eines Aldehyds zur Carbonsäure mittels NAD+ liefert
DG ≈ -50 kJ/mol
Die Bildung eines Phosphoanhydrids aus Carbonsäure und Phosphat
benötigt DG ≈ +50 kJ/mol
→ die Oxidation liefert gerade genug Energie für die Phosphorylierung
Wie wird es mechanistisch ermöglicht Oxidation und Phosphorylierung
zu koppeln? → siehe Folie 252

Reaktion 6: Glycerinaldehyd-3-phosphate
Dehydrogenase (GAPDH)
250

GAPDH: Katalysemechanismus
251

252
▪ Schritt 7: Phosphoglyceratkinase (PGK) Reaktion
Nutzung der energiereichen Phosphoanhydrid-Bindung zur ATP Synthese
▪ Zwischenbilanz der Glycolyse nach Ablauf der Schritte 1-7:
Energie: ± 0 ATP
Elektronen: + 2 NADH (pro Molekül Glucose)
C-Atome im 3PG sind auf der gleichen Oxid.stufe wie
Pyruvat → die verbleibenden Schritte der Glykolyse
enthalten keine weitere Oxidation des C-Gerüsts
Die Phosphorylgruppe im 3PG ist nicht energiereich genug, um ADP zu
phosphorylieren (Hydrolyse DG ≈ -10 kJ/mol).
→ Warum ist es trotzdem möglich, dass 2 ATP in der Glycolyse
gewonnen werden?
▪ Schritt 8: Phosphoglycerat Mutase Reaktion
Verlagerung der Phosphorylgruppe auf das mittlere C-Atom, um Bildung des
Phosphoenolpyruvats (enthält sehr energiereiche Phosphorylgruppe!)
vorzubereiten

Reaktion 7: Phosphoglyceratkinase (PGK)
253

Reaktion 8: Phosphoglyceratmutase (PGM)
254

PGM: Katalysemechanismus
255

256
▪ Schritt 9: Enolase Reaktion
Bildung des energiearmen H2O Moleküls und Entropiegewinn durch
Molekülspaltung wird zur Ausbildung einer energiereichen C=C
Doppelbindung genutzt
▪ Schritt 10: Pyruvatkinase (PK) Reaktion
Die extrem energiereiche Phosphorylgruppe des Phosphoenolpyruvats
(PEP) wird zur ATP Bildung genutzt
Warum ist die Phosphorylgruppe im PEP so extrem energiereich?
Durch Keto-Enol Tautomerie wird die Enol-Gruppe in die
wesentlich energieärmere Ketogruppe umgewandelt (C=C
Bindungen sind energiereicher als C=O Bindungen)
C C O H
H C C O
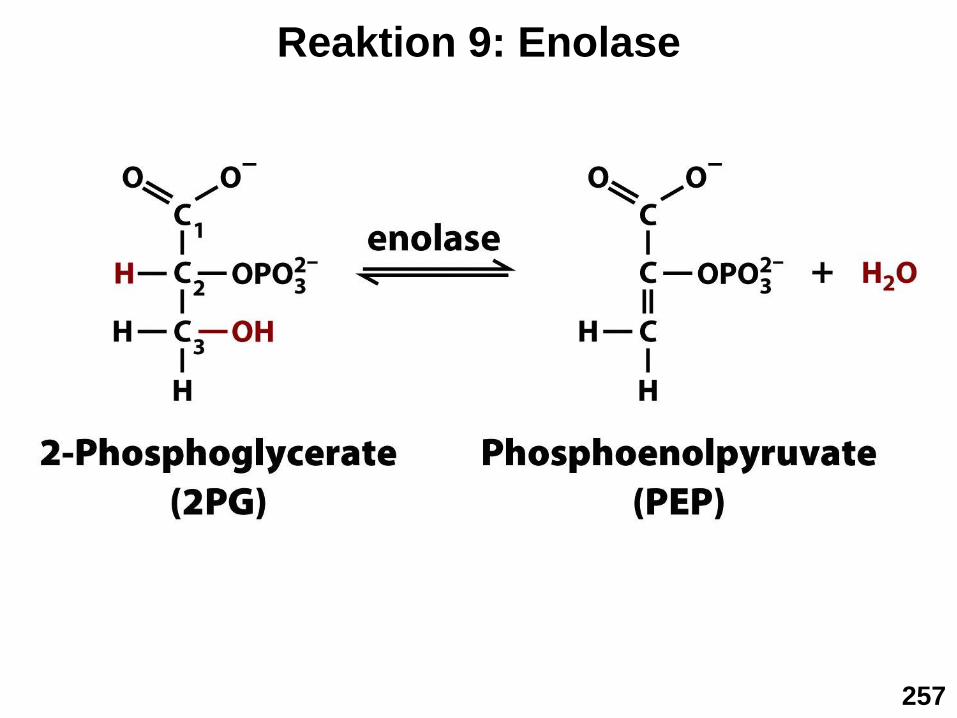
Reaktion 9: Enolase
257

Reaktion 10: Pyruvatkinase (PK)
258
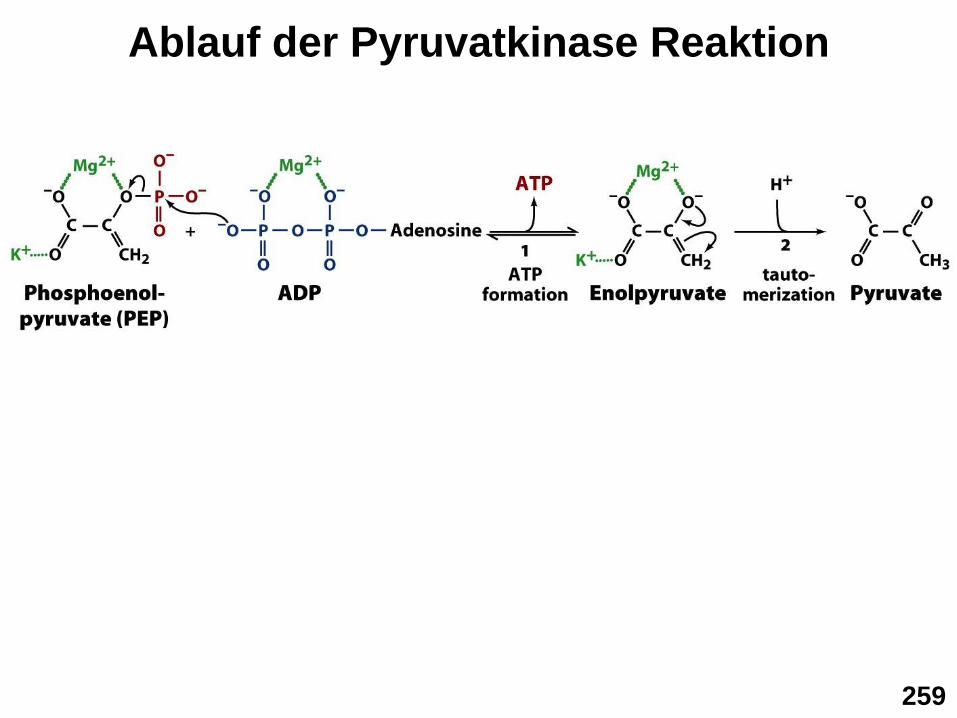
Ablauf der Pyruvatkinase Reaktion
259

Energiebilanz der Pyruvatkinase Reaktion
260

261
Zweck: ▪ Gewinnung von NADPH (für anabole Redoxreaktionen)
▪ Bereitstellung von Ribose-5-Phosphat für Biosynthese von Nukleotiden
▪ Bereitstellung von Erythrose-4-Phosphat für Biosynthese aromatischer
Aminosäuren
Warum wird NAD+/NADH für katabole und NADP+/NADPH für anabiole
Redoxreaktionen verwendet?
▪ Zelluläre Konzentrationsverhältnisse der Redox-Cofaktoren:
9.3. Pentosephosphat Weg (Pentosephosphat Pathway = PPP)
[NADP+]
[NADPH]
[NADH]
[NAD+]≈ 1000
≈ 0.01
→ das NAD+/NADH System ist relativ elektronenarm
und fungiert daher vorwiegend als Oxidationsmittel
→ das NADP+/NADPH System ist relativ elektronenreich
und fungiertt daher vorwiegend als Reduktionsmittel

262
9.3. Pentosephosphat Weg (Pentosephosphat Pathway = PPP)
Gesamtreaktion des PPP:
6 NADPH
6 NADP+
S Oxidationstufen
der C-Atome = ±0
S Oxidationstufen
der C-Atome:
+I
+IV±0
-IFructose-6-
phosphatGlycerinaldehyd-
3-phosphat
3(+IV) ±02(±0) + + = +XII

Verknüpfung von PPP und Glycolyse
263

Oxidative
Phase
Nicht-oxidative
Phase
9.3.1. Einzelreraktionen des PPP
264

265
1. Hälfte des PPP: die oxidative Phase
▪ Die PPP Endprodukte NADPH und CO2 werden gebildet
▪ Ribulose-5-phosphat (C5-Körper) dient als Ausgangssubstrat für 2. Hälfte des
▪ Gesamtreaktion der oxidativen Phase:
3 Glucose-6-phosphat
+
3 H2O
6 NADPH6 NADP+
3 Ribulose-5-phosphat
+
3 CO2 + 6 H+
▪ NADPH-Bildung ist mit CO2-Abspaltung gekoppelt = oxidative Decarboxylierung
Mechanismus: nach Oxidation durch NADP+ entsteht ein Zwischenprodukt, das
instabil ist, da CO2 Abspaltung aus b-Ketocarbonsäuren nur eine geringe
Aktivierungsenergie aufweist (Mesomeriestabilisierung des Übergangszustands)
Mesomerie EnolatCarbanion
Übergangszustand
Keto-Enol
Tautomerie

6-Phosphogluconatdehydrogenase
Reaktion
instabil
(β-Ketocarbonsäure!)
266

267
2. Hälfte des PPP: die nicht-oxidative Phase
▪ Die PPP Endprodukte Fructose-6-phosphat und Glycerinaldehyd-3-phosphat
werden gebildet
▪ Umbau des C5 Körpers Ribulose-5-phosphat (Endproduct der oxidativen Phase)
in C6 Körper (Fructose-6-phosphate) und C3 Körper (GAP), die in Glycolyse
unter Gewinnung von ATP und NAD+ abgebaut werden können
▪ Umbau der Molekülgerüste wird durch Transaldolasen und Transketolasen
katalysiert (gekoppelte Aldol-/Ketolspaltung und Aldol-/Ketoladdition)
▪ C-Körper der nicht-oxidativen Phase können als Grundgerüste für andere
Biosynthesen dienen (z. B. Ribose-5-phosphat, Erythrose-4-phosphat)
▪ Gesamtreaktion der nicht-oxidativen Phase:
3 Ribulose-5-phosphat
(3x C5 = 15 C-Atome)
2 Fructose-6-phosphat
(2x C6 = 12 C-Atome)
+
Glycerinaldehyd-3-phosphat
(1x C3 = 3 C-Atome)

268
Regulation des Metabolismus kann auf Ebene der Gene oder der durch sie
codierten Enzyme erfolgen
▪ schnelle Regulation durch Veränderung der Enzymaktivität mittels
allosterischer Effektoren (Aktivatoren, Inhibitoren; siehe Kapitel 4.5.)
▪ langsame Regulation durch Veränderung der Genexpression (siehe Genetik VL)
9.4. Regulation von Stoffwechselwegen
Welche Enzyme eines Stoffwechselwegs werden reguliert?
Das Enzym muß
▪ spezifisch für den Stoffwechselweg sein (Selektivität)
▪ eine Reaktion katalysieren, die möglichst nahe am Beginn des
Stoffwechselwegs stattfindet (Effizienz)
▪ eine “irreversible” Reaktion (DG <<0) katalysieren (Richtung)
→ Phosphofructokinase (PFK) ist der Hauptregulationspunkt der Glycolyse

Energiebilanz der Glycolyse-Reaktionen
Standardbedingungen:
Konzentration der
Reaktanden ist 1 M
berücksichtigt
zelluläre Konzentrationen
der Reaktanden (≠1 M)
269DG = DG0’ + RTln K
Verhältnis der Konzentrationen der
Reaktanden im zellulären Gleichgewicht

Energiebilanz der Glycolyse-Reaktionen (II)
DG
“irreversibel”
(stark negatives DG)
vollständig reversibel
(DG ≈ 0)
270






![MTB RACE FASTER Ernährungs-Guide Offiziller ... · * [Anmerkung 1] Traditionelle Sportgetränkformeln beziehen sich auf Getränke aus Glukose, Dextrose, Maltodextrin oder anderen](https://static.documents.pub/doc/80x56/5d58a88588c993a1608b490c/mtb-race-faster-ernaehrungs-guide-offiziller-anmerkung-1-traditionelle.jpg)










