A Chimeric Dengue Virus Vaccine using Japanese Encephalitis VirusVaccine Strain SA14-14-2 as Backbone Is Immunogenic and Protectiveagainst Either Parental Virus in Mice and Nonhuman Primates
Department of Virology, Beijing Institute of Microbiology and Epidemiology, Beijing, Chinaa; State Key Laboratory of Pathogen and Biosecurity, Beijing, Chinab; ChengduInstitute of Biological Products Co. Ltd., Chengdu, Chinac; National Institute for Food and Drug Control, Beijing, Chinad; Wadsworth Center, Albany, New York, USAe;Graduate School, Anhui Medical University, Hefei, Chinaf
The development of a safe and efficient dengue vaccine represents a global challenge in public health. Chimeric dengue viruses(DENV) based on an attenuated flavivirus have been well developed as vaccine candidates by using reverse genetics. In this study,based on the full-length infectious cDNA clone of the well-known Japanese encephalitis virus live vaccine strain SA14-14-2 as abackbone, a novel chimeric dengue virus (named ChinDENV) was rationally designed and constructed by replacement with thepremembrane and envelope genes of dengue 2 virus. The recovered chimeric virus showed growth and plaque properties similarto those of the parental DENV in mammalian and mosquito cells. ChinDENV was highly attenuated in mice, and no viremia wasinduced in rhesus monkeys upon subcutaneous inoculation. ChinDENV retained its genetic stability and attenuation phenotypeafter serial 15 passages in cultured cells. A single immunization with various doses of ChinDENV elicited strong neutralizingantibodies in a dose-dependent manner. When vaccinated monkeys were challenged with wild-type DENV, all animals exceptone that received the lower dose were protected against the development of viremia. Furthermore, immunization with Chin-DENV conferred efficient cross protection against lethal JEV challenge in mice in association with robust cellular immunity in-duced by the replicating nonstructural proteins. Taken together, the results of this preclinical study well demonstrate the greatpotential of ChinDENV for further development as a dengue vaccine candidate, and this kind of chimeric flavivirus based on JEvaccine virus represents a powerful tool to deliver foreign antigens.
Dengue has emerged as the most important mosquito-borneviral disease worldwide during the past decades. The spec-
trum of illness ranges from asymptomatic infections and denguefever (DF) to severe and potentially life-threatening dengue hem-orrhagic fever (DHF) and dengue shock syndrome (DSS). Dengueis transmitted to humans by infected Aedes mosquitoes and main-tained by a human-mosquito-human cycle of transmission in na-ture. The World Health Organization (WHO) estimates that over50 million infections occur annually, with 25,000 to 50,000 casesof DHF, and there are over 2.5 billion persons at risk of infectionin more than 100 countries throughout the tropical and subtrop-ical regions (1). In fact, the global health burden of dengue couldbe much higher than previously thought, and a recent modelingwork revealed that 390 million people around the world wereinfected with this mosquito-borne virus in 2010 (2). Currently,there is neither a licensed vaccine nor an effective and specificantiviral therapy for dengue. The four serotypes of dengue virus(DENV-1 to DENV-4) belong to the Flavivirus genus in the familyFlaviviridae together with Japanese encephalitis virus (JEV), Yel-low fever virus (YFV), West Nile virus (WNV), and tick-borneencephalitis virus (TBEV). Flaviviruses have an �11-kb single-stranded positive-sense RNA genome that contains a single openreading frame (ORF) flanked by two untranslated regions (5= and3=UTRs). The ORF encodes a single polyprotein, which is proteo-lytically processed into three structural proteins (C, prM, and E)and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A,NS4B, and NS5). The E glycoprotein is the principal antigen in-ducing protective immunity against virus infection, and it re-quires the coexpression of prM to acquire its native conformation.
All the nonstructural proteins are actively involved in viral ge-nome replication, translation, and regulation. Due to the highdegree of similarity among flaviviruses in genome organization,replication, and translation strategy, viable chimeric viruses havebeen rationally designed and generated by interchanging variousgenes among different flaviviruses (3–6) by using reverse genetics.This kind of chimeric approach has been widely utilized in denguevaccine development (6–11). The most advanced product of thiskind is the ChimeriVax-Dengue (CYD) vaccine developed bySanofi Pasteur, which was created based on the yellow fever vac-cine strain 17D (YF-17D) (reviewed in reference 12). The protec-tive efficacy of this tetravalent dengue vaccine in a phase 2b trial inThailand was recently reported (13), and large-scale phase 3 trialsare ongoing (14). Another chimeric dengue vaccine under clinicaltrial is developed based on the known attenuated DENV-2 strainPDK53 (6, 10, 15, 16). Additionally, recombinant DENV-4 with adeletion of 30 nucleotides in the 3= UTR (DEN4�30) has beensuccessfully utilized as a backbone for chimeric dengue vaccinedevelopment (11, 17–20). Generally, the well-characterized atten-
uated flaviviruses represent good candidates as a genetic backbonefor chimeric dengue vaccine development. A licensed Japaneseencephalitis virus live vaccine (strain SA14-14-2) has been widelyused in China and other Asian countries, including India, SouthKorea, Sri Lanka, and Nepal (21). Large-scale vaccination in morethan 300 million children has shown an excellent safety profile,genetic stability, and remarkable effectiveness and efficacy forSA14-14-2 (22–26). These attractive properties make JEV strainSA14-14-2 an ideal candidate as a genetic backbone for chimericdengue vaccine development. In this work, we report the earlydevelopment of a chimeric dengue vaccine using JEV vaccinestrain SA14-14-2 as the genetic backbone. The resulting chimera ishighly attenuated, immunogenic, and protective against eitherJEV or DENV challenge in mice or nonhuman primates, demon-strating great potential for further development. The use of theJEV SA14-14-2 genetic background to deliver protective antigenscould be an efficient method for the development of vaccinesagainst other flaviviruses.
MATERIALS AND METHODSCells and viruses. BHK-21 (baby hamster kidney) and Vero (Africangreen monkey kidney) cells were maintained in Dulbecco’s minimal es-sential medium (DMEM; Invitrogen) supplemented with 10% fetal bo-vine serum (FBS) and penicillin (100 U/ml)-streptomycin (100 �g/ml).Aedes albopictus C6/36 cells were cultured in RPMI 1640 supplementedwith nonessential amino acids (Invitrogen) and 10% FBS. Primary ham-ster kidney (PHK) cells were produced at the Chengdu Institute of Bio-logical Products under the good manufacturing practice (GMP) work-shop and cultured in MEM alpha supplemented with 10% FBS. All celllines were grown at 37°C in 5% CO2, except for C6/36 cells, which weremaintained at 28°C. DENV-2 strain D2-43 (GenBank accession no.AF204178) was isolated from the acute-phase serum of a DF patient inGuangxi Province, China, in 1987 (27). DENV-2 strain TSV01 (GenBankaccession no. AY037116) was recovered from the infectious clone andprepared from culture supernatants of infected C6/36 cells. JEV strainSX-06 was isolated from a mosquito in Shanxi Province, China, in 2006,and the 50% lethal dose (LD50) was measured in groups of 4-week-oldBALB/c mice accordingly. JEV vaccine strain SA14-14-2 (GenBank no.D90195) was provided by the Chengdu Institute of Biological Products.Virus titers were determined by standard plaque assay on BHK-21 cells,and virus stocks were stored in aliquots at �80°C until used.
Genetic construction of the JEV/DENV chimera. All plasmids wereconstructed using standard molecular biology protocols and confirmedby enzyme digestion and DNA sequencing. The DNA construct of thechimera is shown in Fig. 1A.
Genetic construction of the full-length infectious clone of JEV hasbeen described by Ye et al. (28). Briefly, plasmid pACYC-Linker was firstcreated from plasmid pACYC177 (Fermentas) and plasmid pSP64 (Pro-mega). Further details about the construction of pACYC-Linker andprimers used in this study are available from the authors on request. Theentire genome of JEV strain SA14-14-2 was cloned in plasmid pACYC-Linker. Cloning sites were engineered to permit replacement of the entirepre-M and E coding sequences of JEV for the corresponding sequences ofDENV-2 strain D2-43. Sites for posttranslational cleavage of the capsidand pre-M proteins and of the E and NS1 proteins were preserved (Fig.1A). The resulting plasmid contained the full-length cDNA of the JEV/DENV-2 chimera, named pChinDENV.
Transcription and transfection. Plasmid pChinDENV was linearizedwith XhoI and used as a template for SP6 RNA polymerase transcriptionin the presence of an m7GpppA cap analog. In vitro transcription was doneusing the RiboMAX Large Scale Production System (Promega) accordingto the manufacturer’s protocols. The yield and integrity of transcriptswere analyzed by gel electrophoresis under nondenaturing conditions.RNA transcripts (5 �g) were transfected with Lipofectamine 2000 (Invit-
rogen) into BHK-21 cells grown in 60-mm-diameter culture dishes. Su-pernatants were then harvested at about day 5 posttransfection, whentypical cytopathic effects (CPE) were observed, and clarified by low-speedcentrifugation. Working virus stocks (named ChinDENV) were preparedby amplification of the transfection harvest for one passage on Vero orPHK cells and stored in aliquots at �80°C until use. Virus titers weredetermined by standard plaque assay on BHK-21 cells.
Nucleotide sequencing. Viral RNA was isolated using an RNeasyminikit (Qiagen), and the genome cDNA was obtained by reverse tran-scription (RT) using SuperScript III (Invitrogen). For determination ofconsensus sequence, PCR products were directly sequenced in both direc-tions using virus-specific primers. The 5= and 3= termini were sequencedusing 5= and 3= rapid amplification of cDNA ends (RACE). Sequencefragments were assembled into a consensus sequence with DNA STARsoftware, version 7.0.
Growth curves. Growth curves were done by infecting confluentBHK-21, C6/36, Vero, or PHK cells in a 12-well plate at a multiplicity ofinfection (MOI) of 0.01. The inoculum was removed after 1 h of adsorp-tion, and culture medium was added. Cell supernatants were collected atsuccessive 24-h intervals postinfection. Yields of virus in each sample werethen quantitated by plaque assay on BHK-21 cells. Briefly, BHK-21 cells in12-well plates were infected with a 10-fold serial dilution of viruses. Theplates were incubated at 37°C for 1 h with gentle rocking every 15 min. Thesupernatant was removed, and cells were overlaid with 1% low-melting-point agarose (Promega) in DMEM containing 2% FBS. After furtherincubation at 37°C for 4 days, the cells were fixed with 4% formaldehydeand stained with 0.2% crystal violet to visualize the plaques.
IFA. Indirect immunofluorescence assays (IFA) were performed asfollows. Briefly, confluent BHK-21 cells grown in 60-mm-diameter cul-ture dishes containing a 1-cm2 coverslip inside were infected with virusesat an MOI of 0.01. At 48 h postinfection, the coverslips containing infectedcells were removed and directly used for IFA analysis. The cells seeded onthe coverslip were fixed with ice-cold acetone and incubated with primaryantibodies (monoclonal antibody [MAb] 4AD5F5D5D6for JEV E protein,MAb 2B8 for DENV E protein [27], or JN1 for JEV NS1 protein [Abcam])or serum of mice at 37°C for 1 h. Cells were washed three times withphosphate-buffered saline (PBS) and then incubated with secondary an-tibodies conjugated to Alexa Fluor 488 (Invitrogen) in PBS for 30 min at37°C. Fluorescent cells were examined using a fluorescence microscope(Olympus).
Genetic-stability assay. The chimeric virus ChinDENV was seriallypassaged in PHK cells at an MOI of 0.01. The complete genome sequenceand plaque phenotypes of ChinDENV virus of passages 3, 9, and 15 wereassessed as described above.
Neutralization assay. Neutralizing antibody titers were determinedby a constant virus-serum dilution 50% plaque reduction neutralizationtest (PRNT50). Briefly, a 1:10 dilution of serum was prepared in DMEMcontaining 2% FBS and then heat inactivated for 30 min at 56°C. Serial2-fold dilutions of inactivated sera were mixed with equal volumes ofDENV-2 in DMEM supplemented with 2% FBS to yield a mixture con-taining approximately 250 PFU of virus/ml. After incubation at 37°C for 1h, 200-�l volumes of the virus-antibody mixtures were added to wells of12-well plates containing confluent monolayers of BHK-21 cells. The con-centration of infectious virus was determined using the plaque assay de-scribed above. The endpoint neutralization titer was calculated by themethod of Reed and Muench (29).
Mouse experiments. Research involving animals was approved by andcarried out in strict accordance with the guidelines of the ExperimentalAnimal Welfare and Ethics Committee of Beijing Institute of Microbiol-ogy and Epidemiology. For neurovirulence tests, groups of 2-day-old and4-week-old BALB/c mice were inoculated by the intracerebral (i.c.) routewith 10-fold dilutions of ChinDENV and its parent viruses, respectively.Animals were monitored for 21 days after inoculation. Any mice found ina moribund condition were euthanatized and scored as dead. Virus dosesinducing a 50% mortality rate were calculated using the method of Reed
A Chimeric JEV/Dengue Vaccine
December 2013 Volume 87 Number 24 jvi.asm.org 13695
and Muench. Average survival times (AST) were calculated for animalsthat succumbed to infection.
Immunogenicity was assessed by subcutaneous (s.c.) inoculation ofvarious doses of ChinDENV virus diluted in DMEM plus 2% FBS into4-week-old female BALB/c mice. The mice were bled by tail vein puncture1 day prior to and periodically after immunizations at 2, 4, and 6 weekspostimmunization. The sera were stored at �20°C for determination ofIgG antibodies by IFA and neutralizing antibodies by PRNT50.
Additionally, groups of 4-week-old female BALB/c mice (n � 10) werefirst immunized with 6.85 log10 PFU of JEV live vaccine SA14-14-2 or PBSby the s.c. route, and then 14 days later, all the animals were immunizedwith the same dose of ChinDENV. At 28 and 42 days after primary im-munization, titers of neutralization antibody against DENV-2 in serafrom each animal were measured by using standard PRNT as describedabove.
For the protection assay, heat-inactivated sera from mice immunizedwith ChinDENV were mixed with equal volumes of DENV-2 (D2-43strain) diluted in DMEM supplemented to yield a mixture containingapproximately 500 PFU (�100 LD50s) of virus/ml. The virus-antibodymixtures were incubated at 37°C for 1 h and then injected by the i.c. routeinto groups of 2-day-old BALB/c suckling mice (n � 9). Animals werethen monitored for 15 days after inoculation.
JEV challenge experiments. For the cross protection study, groups of4-week-old BALB/c mice were inoculated by the s.c. route with 105 PFU ofChinDENV. Equal doses of JEV (SA14-14-2) and PBS were set as controls.Four weeks after the initial vaccination, all the mice were intraperitoneally(i.p.) challenged with 105 PFU (10 LD50s) of JEV SX-06 as described in therelevant figure legends. Signs of illness and death were monitored for atleast 15 days. Groups of three virus-immunized or PBS-immunized micewere anesthetized, and blood was collected by retro-orbital bleeding andsubjected to neutralizing antibody titration by plaque assay. Spleens weretaken after cervical dislocation for evaluation of cellular immune response(described below).
Cellular immune responses. Cellular immune responses were as-sessed in splenocytes by the gamma interferon (IFN-�) or interleukin 2(IL-2) enzyme-linked immunosorbent spot (ELISPOT) mouse set (BDBiosciences) according to the manufacturer’s protocol. Briefly, spleensexcised from anesthetized animals were thawed and washed with Hanksbalanced salt solution (HBSS). The cells were then centrifuged at roomtemperature at 2,500 rpm for 15 min without braking, followed by twowashes with HBSS. Then, the cells were resuspended in RPMI 1640 con-taining 10% FBS and diluted to a working concentration of 100 millioncells per ml. For ELISPOT analysis, 10 million cells per well were seededwith the appropriate antigen stimulation in 96-well tissue culture dishes
FIG 1 Design and construction of a genomic-length chimeric JEV/DENV-2 (ChinDENV) cDNA clone. (A) Strategy for the construction of the infectious cDNAclone of ChinDENV. The JEV prM signal peptide (blank box) is underlined. The C-terminal 3 amino acids of JEV E protein (E3, gray box) and the prM-E ofDENV-2 lacking the C-terminal 3 amino acids of the E protein (dark gray box) are indicated. (B) Alignment of flavivirus amino acid sequences around the C-prMand E-NS1 junctions. The prM signal peptides are boxed. The last and antepenultimate C-terminal amino acids of flavivirus E proteins are highlighted in gray.Sites of cleavage by the viral NS2B-3 protease and signal peptidase are indicated by the open arrow and solid arrows, respectively.
coated with 5 mg/ml of IFN-� or IL-2 capture monoclonal antibody.Nonstimulated and concanavalin A (5 �g/ml; Sigma-Aldrich)-stimulatedcells were used as negative and positive controls, respectively. The cellswere then cultured for 36 h at 37°C and 5% CO2. Plates were washed, andbiotinylated anti-mouse IFN-� or IL-2 antibody was added to each welland incubated for 2 h at room temperature. Thereafter, the plates werewashed and incubated for 1 h at room temperature with streptavidin-horseradish peroxidase (streptavidin-HRP). Finally, AEC substrate solu-tion (BD Biosciences) was added and spots were counted with an Immu-noSpot analyzer (Cellular Technology Ltd.). Assay results are expressed asthe value obtained by the following: (number of spots in experimentalwell � number of spots in medium control)/107 cells.
Monkey experiments. All experimental procedures involving rhesusmonkeys were performed under sodium pentobarbital anesthesia bytrained technicians, and all efforts were made to ameliorate the welfareand to minimize animal suffering in accordance with the recommenda-tions in The Use of Non-Human Primates in Research (30).
(i) Immunization and challenge of rhesus monkeys. Fourteen mon-keys, weighing 3.4 to 5.0 kg, were prescreened for negativity for antibodiesagainst DENV by IFA. Animals were randomly divided into four groupsand immunized s.c. in the deltoid region with 0.5 ml of ChinDENV con-taining 3.0, 4.0, or 5.0 log10 PFU or were sham inoculated with PBS. Bloodwas collected daily for 10 days to detect viremia. Blood samples for neu-tralizing-antibody tests were taken before immunization (day �1) andthen on days 15, 20, 30, 50, and 63 postimmunization. On day 64 postim-munization, all the immunized monkeys plus unimmunized controlswere challenged by s.c. inoculation of 0.5 ml containing 5 log10 PFU ofDENV-2 (TSV-01 strain). For the following 9 days, blood was collectedfor determination of viremia. Neutralizing-antibody levels in serum weremeasured by neutralization assay described above on days 15 and 30 post-challenge.
(ii) Viremia determination. Viremia in monkeys was measured byquantitative real-time reverse transcriptase PCR (qRT-PCR). Virus titerwas expressed in log10 PFU/ml. The number of PFU in serum detected wascalculated by generating a standard curve from 10-fold dilutions of RNAisolated from a serum sample containing a known number of PFU, thetiter of which was determined by plaque assay on BHK-21 cells as de-scribed above. Briefly, viral genomic RNA was extracted from 200 �l ofserum or an equal volume of sample serum by using the PureLinkTMRNA minikit (Invitrogen, USA) according to the manufacturer’s instruc-tions. RNA was eluted in 50 �l of RNase-free water, aliquoted, and storedat �80°C until use. The primers (5=-CAGGCTATGGCACTGTCACGAT-3= and 5=-CCATTTGCAGCAACACCATCTC-3=) and probe (5=-CTCTCCGAGAACGGGCCTCGACTTCAA-3=) were designed using the pro-gram Primer ExpressTM (PE Applied Biosystems, USA). The targetregion for the assays was based on the E protein region of the genome ofDENV-2. A TaqMan assay probe was 5= labeled with the reporter dye6-carboxyfluorescein (FAM) and labeled at the 3= end with the quencherdye 6-carboxytetramethylrhodamine (TAMRA). qRT-PCR was per-formed by using a One Step PrimeScript RT-PCR kit (TaKaRa, Japan).The 25-�l reaction mixtures were set up with a 0.5 �M concentration ofeach primer (qDVEF and qDVER, respectively), 0.25 �M probe, and 5 �lof RNA. Thermocycling programs consisted of 42°C for 5 min, 95°C for 10s, and 40 cycles of 95°C for 5 s and 60°C for 30 s.
Statistic analysis. For survival analysis, Kaplan-Meier survival curveswere analyzed by a log rank test using standard GraphPad Prism software,version 5.0.
RESULTSEngineering and characterization of the chimeric ChinDENV.To assess the potential of the infectious cDNA clone derived fromJEV SA14-14-2 to serve as a vector for vaccine development, weengineered a chimeric DENV-2 (termed ChinDENV) cDNAclone containing the structural genes of DENV-2 strain D2-43, arepresentative strain of the Asia 1 genotype, using a sub-
genomic replicon of JEV SA14-14-2 (31) as the genetic back-bone. ChinDENV retained the prM signal peptide of JEV as well asthe last three amino acids at the terminus of the E protein toensure efficient cleavage at the C-prM junction and E-NS1 junc-tion, respectively (Fig. 1).
Total RNA transcripts encoding ChinDENV were transfectedin BHK-21 cells, and immunostaining assays with specific mono-clonal antibodies characterized the recovered ChinDENV ex-pressing the E protein of DENV-2 and NS1 protein of JEV (Fig.2A). The recovered ChinDENV caused typical cytopathic effectsin BHK-21 cells almost the same as parental DENV, producing ahomogeneous population of plaques which were �0.5 mm in di-ameter, approximately half the size of those of parental JEV (Fig.2B). RT-PCR analysis and full-genome sequencing confirmed thatthe prM-E gene of DENV-2 was successfully engineered into thegenome of JEV SA14-14-2 as expected.
Full-genome sequencing showed that there were seven nucle-otide substitutions between ChinDENV and parental DENV-2 inthe prM-E genes, and four of them resulted in amino acid changes,including prM-28 Gly to Gln, prM-55 Leu to Phe, prM135 Met toIle, and E402 Phe to Ile. Within the UTRs and capsid protein aswell as nonstructural protein genes, there were four nucleotidedifferences (nucleotide positions 473, 3200, 3465, and 10843) be-tween ChinDENV and JEV SA14-14-2, all of which were silent anddid not cause amino acid substitutions.
Subsequently, the growth efficiencies of ChinDENV and itsparental viruses were examined in mammalian Vero and mos-quito C6/36 cells (Fig. 2D). The results showed that ChinDENVreplicated efficiently in Vero and C6/36 cells and peaked at 72 and48 h postinfection, with titers of 105.8 PFU/ml and 106.9 PFU/ml,respectively. ChinDENV replicated slightly more efficiently thanparental DENV-2, whereas both of them replicated less efficientlythan JEV in both cells. The growth kinetics of ChinDENV wasthen determined in primary hamster kidney (PHK) cells, the cer-tified cell bank intended for manufacturing ChinDENV. As shownin Fig. 2D, ChinDENV and DENV-2 exhibited very similar kinet-ics of virus production, with the same peak titer, approximately106.3 PFU/ml. Taken together, these results demonstrate that therescued ChinDENV chimera possesses the designed genomicstructure, expresses DENV-specific antigens, shows a small-plaque phenotype, and replicates efficiently in different cell lines.
Genetic and plaque phenotypic stability of ChinDENV. Toassess the phenotype and genetic stability of ChinDENV duringsequential passage in vitro, ChinDENV recovered from trans-fected cells (passage 0) was passaged up to 15 times in PHK cells.Viruses harvested at passages 3, 9, and 15 were subjected to aplaque forming assay and complete-genome sequencing in com-parison with that of passage 0. The small-plaque phenotype ofChinDENV in BHK-21 cells remained unchanged at each passage(Fig. 2C), indicating that there were no plaque variants overgrow-ing the original parent population. Only a single nucleotide mu-tation at position 2173 in the E gene occurred at passage 15, lead-ing to an amino acid change from Gln to Arg. Another silentnucleotide change at position 8666 in the NS5 gene was ob-served at passage 9. These data indicate that the chimeric virusChinDENV exhibits a genetic and plaque phenotypic stabilityduring in vitro passage.
Virulence of the chimeric ChinDENV in mice. The JEV vac-cine strain SA14-14-2 retains a degree of neurotropism in sucklingmice (21). To make sure ChinDENV does not exceed its parental
A Chimeric JEV/Dengue Vaccine
December 2013 Volume 87 Number 24 jvi.asm.org 13697
virus in neurovirulence, groups of 2-day-old BALB/c mice wereinoculated with serial 10-fold dilutions of ChinDENV and its pa-rental viruses, JEV SA14-14-2 and DENV-2 D2-43, by the i.c.route. All the infected animals were then observed for at least 21
days for signs of illness and death. As shown in Table 1, the LD50 ofChinDENV was 245.47 PFU, approximately 115-fold and 50-foldhigher than those of JEV SA14-14-2 and DENV D2-43. Addition-ally, we also found that neither the ChinDENV chimera nor itsparental viruses were lethal for 3- to 4-week-old BLAB/c miceinoculated at a dose of 105.0 PFU by the i.c. or i.p. route (data notshown). These results indicate that ChinDENV exhibits greatlyreduced neurovirulence in suckling mice compared with those ofits parental viruses and retains avirulence to immunocompetentadult mice.
Immunogenicity and protection in mice. To assess the im-munogenicity of ChinDENV in mice, groups of 4-week-oldBALB/c mice were immunized with various doses (104.8 and105.8 PFU per mouse) of ChinDENV by the s.c. route. SerumIgG and neutralizing antibodies against DENV-2 were mea-sured by IFA and PRNT at 2, 4, and 6 weeks after immunizationas previously described (32). All mice had seroconversion at 2weeks postimmunization, and IgG antibody levels increasedrapidly (Fig. 3A). High levels of IgG antibodies remained stableuntil day 42 in mice receiving either a low or high dose (104.8 or105.8 PFU) of ChinDENV. Importantly, DENV-specific neu-tralizing antibodies were induced in all mice immunized withChinDENV in a dose-dependent manner. The geometric meantiters (GMTs) of neutralizing antibody induced by low and
FIG 2 In vitro characterization of ChinDENV. (A) Immunostaining of JEV-, ChinDENV-, DENV-2-, or mock-infected BHK cells with specific anti-JEV E,DENV-2 E, and JEV NS1 antibodies. Cells were infected with viruses at an MOI of 0.01. At 48 h postinfection, JEV or DENV-2 antigen was detected withmonoclonal antibody 4AD5F5D5D6 (for JEV E protein), 2B8 (for DENV-2 E protein), and JN1 (for JEV NS1 protein). (B) Plaque morphology of JEV,ChinDENV, and DENV-2 on BHK-21 cells grown in 6-well plates were infected with a 10-fold serial dilution of viruses. The plates were incubated at 37°C for 1h. Supernatant was removed and cells were overlaid with 1% low-melting-point agarose in DMEM containing 2% FBS. After further incubation at 37°C for 4days, the cells were fixed with 4% formaldehyde and stained with 0.2% crystal violet to visualize the plaques. (C) Plaque morphology of ChinDENV passaged inPHK cells. The chimeric virus ChinDENV was passaged in PHK cells up to 15 times. The plaque phenotypes of passage 3, 9, and 15 viruses were examined onBHK-21 cells 4 days after infection by plaque assay. (D) Growth curves of the chimera ChinDENV and parental viruses JEV and DENV-2 in cell culture.Monolayers of Vero, C6/36, and PHK cells were infected with the indicated viruses at an MOI of 0.01. At each time point, the media were removed and virus titersin cell culture media were determined on BHK-21 cells by plaque assay.
TABLE 1 Neurovirulence of ChinDENV and its parental viruses insuckling mice
high doses of ChinDENV were 47.1 and 106.3 on day 42postimmunization, respectively (Fig. 3B).
To clarify whether preimmunization of JEV interferes with theimmunogenicity of ChinDENV, mice immunized with JEV SA14-14-2 were subjected to a second immunization with the same doseof ChinDENV. As shown in Table 2, in mice immunized with JEVSA14-14-2, high neutralization antibodies against DENV-2 weresuccessfully induced by the second ChinDENV immunization.However, the peak titer (55.00 8.85) at 42 days after primaryimmunization in the JEV-preimmunized mice was slightly lowerthan that in naive mice (84.00 17.33). And no neutralizationantibody was detected in the PBS control group, as expected.
Since no ideal immunocompetent-mouse model was currentlyavailable for DENV challenge, we initially evaluated the protec-tion efficacy of ChinDENV antisera in the established suckling-mouse model as previously described (27). Complete protectionwas observed for the group of animals inoculated with antiserafrom the ChinDENV-immunized mice, whereas all control micedied within 5 to 9 days postinoculation (data not shown). Takentogether, these results demonstrate that immunization with a sin-gle dose of ChinDENV could induce a neutralizing antibody re-sponse against DENV-2 that conferred full protection against le-thal DENV-2 challenge in suckling mice.
Viremia, immunogenicity, and protection in nonhuman pri-mates. Attenuation of ChinDENV was further evaluated in rhesusmonkeys, the most suitable model for preclinical evaluation ofsafety of DENV infections for humans (33, 34). Three groups ofadult rhesus monkeys were inoculated s.c. with ChinDENV atdoses of 1 103, 1 104, and 1 105 PFU per animal. Viremia inmonkeys was then determined by real-time RT-PCR for 10days postinjection. All the animals injected with wild-type (wt)
DENV-2 exhibited continual viremia (mean duration, 3.5 days),with a peak titer of 102.75 PFU/ml following DENV challenge.However, ChinDENV failed to produce any detectable viremia inany inoculated monkeys (Table 3). No signs of disease or clinicalmanifestation were observed in any of the ChinDENV-injectedanimals throughout the experiments. These results strongly dem-onstrate the excellent safety profile of ChinDENV in nonhumanprimates.
Next, we determined the neutralizing antibody response in-duced by the single dose of ChinDENV in monkeys. As shown inTable 4, all the ChinDENV-immune monkeys developed neutral-izing antibodies to DENV-2 from day 15 until day 50 postimmu-nization in a dose-dependent manner. Monkeys receiving highand middle doses of ChinDENV attained peak GMTs of 184 and204 on day 20 after inoculation, respectively. Animals immunizedwith the low dose of ChinDENV also developed strong neutraliz-ing antibodies on day 30 postimmunization, with a GMT of 82.No neutralizing antibody was detected for the control animalsimmunized with PBS, as expected.
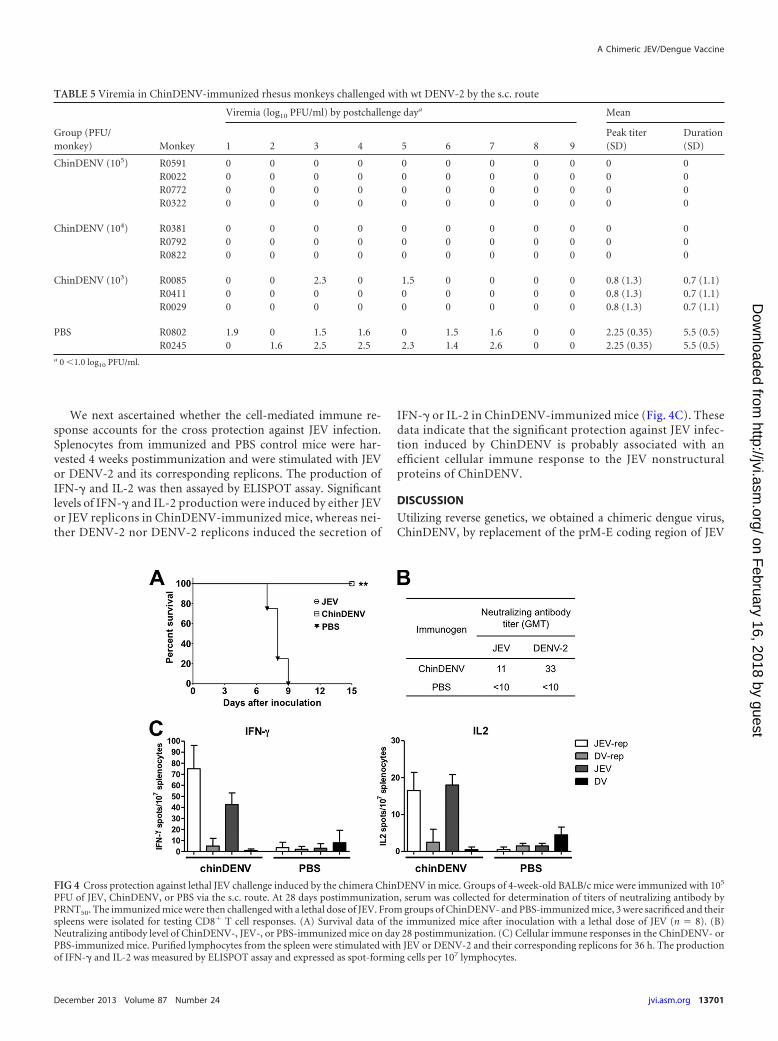
To assess the in vivo protective efficacy, all the vaccinated ani-mals were challenged with wt DENV-2 on day 63 postimmuniza-tion, and viremia was monitored for 10 days thereafter. As shownin Table 5, control animals immunized with PBS exhibited con-tinual viremia (mean duration, 5.5 days), with a peak titer of 102.25
PFU/ml following wt DENV-2 challenge. All the animals receivinghigh and middle doses (105 and 104 PFU) of ChinDENV devel-oped no viremia following wt DENV-2 challenge; only one animal(R0085) immunized with a low dose (103 PFU) of ChinDENVdeveloped low-level, transient viremia (Table 5). These observa-tions showed that the single dose of ChinDENV immunizationsignificantly protected against wt DENV-2-induced viremia innonhuman primates. Additionally, the anamnestic antibody re-sponse after virus challenge was analyzed in the sera collected atdays 15 and 30. It is noteworthy that all ChinDENV-immunemonkeys developed a significant (�4- to 6-fold) increase in serumneutralizing antibody titer after DENV-2 challenge, indicatingthat DENV-2 challenge had stimulated a strong anamnestic im-mune response (Table 4). Together, these results strongly demon-strate that a single immunization with ChinDENV successfullyinduced a robust protective immune response in nonhuman pri-mates.
Protection against lethal JEV challenge in mice. Clinical trials
FIG 3 Immunogenicity of ChinDENV in mice. Groups of five 4-week-old BALB/c mice were immunized with doses of 60,000 or 600,000 PFU of thechimera by the s.c. route. Serum was collected from animals at 14, 28, and 42 days postvaccination for determination of titers of IgG antibody (A) andneutralizing antibody (B) against DENV-2 by using IFA and PRNT50, respectively. The dotted line represents the limit of detection of the assay for eachserum.
TABLE 2 Immunogenicity assay of ChinDENV in mice preimmunizedwith JEV vaccine SA14-14-2
1stimmunization
2ndimmunization
PRNT50 titer (mean SD) onindicated day after 1stimmunization
have demonstrated that YFV-specific cellular immune responsecould be induced by the YFV-based CYD vaccination (35). In thisstudy, to make sure that the potential protective immunity againstJEV was conferred by ChinDENV immunization, groups of4-week-old BALB/c mice immunized with ChinDENV were chal-lenged with lethal JEV challenge. The same dose of SA14-14-2 wasalso immunized as controls together with PBS. All the mice werethen subjected to a 15-day period of observation for signs of death.All the mice immunized with SA14-14-2 survived, and all PBScontrols died within 9 days postinfection, as expected (Fig. 4A).Significantly, none of the ChinDENV-vaccinated animals diedfrom wt JEV challenge. This result demonstrates that a single im-
munization with ChinDENV could induce complete protectionagainst lethal JEV challenge.
To further investigate the putative roles of the humoraland cellular immune responses in the protection against JEVchallenge, PRNT against JEV and DENV-2 were assayed, re-spectively. As shown in Fig. 4B, ChinDENV immunization in-duced significant neutralizing antibodies (GMT, 1:33) againstDENV-2, whereas the titer of neutralizing antibody against JEVwas relatively low (GMT, 1:11). This result indicates that theChinDNEV-induced protection may not be attributable to thehumoral immune response to the chimeric DENV structuralproteins.
TABLE 3 Viremia in rhesus monkeys immunized with graded doses of ChinDENV
VirusDose(log10 PFU) Monkey Gendera
Viremia (log10 PFU/ml) by indicated day postimmunizationb Mean
R0411 M �10 49 73 78 56 411 538R0029 M �10 27 100 84 57 254 466
GMT lt]10 27 65 82 62 293 471
PBS R0802 F �10 �10 �10 NT NT 157 170R0245 M �10 �10 �10 NT NT 158 109
GMT 157 136a M, male; F, female.b 50% plaque reduction neutralization measured against DENV-2 strain TSV01. Monkeys were immunized with the indicated doses of ChinDENV and challenged by the s.c. routewith 105 PFU of DENV-2 strain TSV01 on day 63. Antibody levels were determined on �1, 15, 20, 30, and 50 days postimmunization and 15 and 30 days postchallenge. NT, nottested.
We next ascertained whether the cell-mediated immune re-sponse accounts for the cross protection against JEV infection.Splenocytes from immunized and PBS control mice were har-vested 4 weeks postimmunization and were stimulated with JEVor DENV-2 and its corresponding replicons. The production ofIFN-� and IL-2 was then assayed by ELISPOT assay. Significantlevels of IFN-� and IL-2 production were induced by either JEVor JEV replicons in ChinDENV-immunized mice, whereas nei-ther DENV-2 nor DENV-2 replicons induced the secretion of
IFN-� or IL-2 in ChinDENV-immunized mice (Fig. 4C). Thesedata indicate that the significant protection against JEV infec-tion induced by ChinDENV is probably associated with anefficient cellular immune response to the JEV nonstructuralproteins of ChinDENV.
DISCUSSION
Utilizing reverse genetics, we obtained a chimeric dengue virus,ChinDENV, by replacement of the prM-E coding region of JEV
TABLE 5 Viremia in ChinDENV-immunized rhesus monkeys challenged with wt DENV-2 by the s.c. route
FIG 4 Cross protection against lethal JEV challenge induced by the chimera ChinDENV in mice. Groups of 4-week-old BALB/c mice were immunized with 105
PFU of JEV, ChinDENV, or PBS via the s.c. route. At 28 days postimmunization, serum was collected for determination of titers of neutralizing antibody byPRNT50. The immunized mice were then challenged with a lethal dose of JEV. From groups of ChinDENV- and PBS-immunized mice, 3 were sacrificed and theirspleens were isolated for testing CD8� T cell responses. (A) Survival data of the immunized mice after inoculation with a lethal dose of JEV (n � 8). (B)Neutralizing antibody level of ChinDENV-, JEV-, or PBS-immunized mice on day 28 postimmunization. (C) Cellular immune responses in the ChinDENV- orPBS-immunized mice. Purified lymphocytes from the spleen were stimulated with JEV or DENV-2 and their corresponding replicons for 36 h. The productionof IFN-� and IL-2 was measured by ELISPOT assay and expressed as spot-forming cells per 107 lymphocytes.
A Chimeric JEV/Dengue Vaccine
December 2013 Volume 87 Number 24 jvi.asm.org 13701
vaccine strain SA14-14-2 with the corresponding segment ofDENV-2. Cleavage of the N-terminal signal sequence of prM by ahost cellular peptidase requires prior removal of the C proteinfrom this signal sequence by the viral NS2B/3 protease (36–38).These coordinated cleavage events are believed to play an impor-tant role in the process of the assembly of flavivirus virions. The Ctermini of the E proteins of all members of the Flaviviridae familyhave similar organizations (39) and consist of two antiparalleltransmembrane helices (TM1 and TM2). The TM2 helix serves asan internal signal sequence for the synthesis of the first nonstruc-tural protein, NS1 (40). As shown in Fig. 1B, the last and antepen-ultimate C-terminal amino acids of flavivirus E proteins are highlyconserved, indicating that these amino acids are likely to play animportant role in efficient cleavage of the E-NS1 junction. Thus,to ensure proper processing of prM and E proteins of DENV-2,the signal sequence of JEV between C and prM as well as the lastthree C-terminal amino acids of the JEV E protein were retained inthe ChinDENV chimera (Fig. 1B). As expected, the resulting via-ble ChinDENV was characterized by the chimeric structure of thegenome, and the proteins of ChinDENV appeared to be properlyprocessed, with the E protein retaining the antigenic properties ofDENV-2 (Fig. 2A). It was also observed that ChinDENV displayeda small-plaque phenotype in BHK-21 cells, similar to that ofprM-E donor virus, but its plaque size was significantly reducedcompared to that of JEV. We further found that ChinDENV grewequally as well as the parent virus DENV-2 in various cell lines,with decreased peak replication titers compared with JEV. Theseresults are consistent with the previous observations that chimerasexhibited growth characteristics similar to those of the virusescontributing structural protein genes (5, 41), indicating that thereduction in plaque size and replication efficiency for ChinDENVcompared to JEV are likely to be determined partly by the donorprM-E genes. In addition, the chimerization itself has been provedto contribute to the attenuation of chimeras (10, 42–50). Notably,the chimeric virus ChinDENV replicated efficiently in Vero andPHK cells, both of which are certified for vaccine production.Additionally, ChinDENV reached a peak titer of 107.7 PFU/ml inPHK cells after optimization. Serial passaging of ChinDENV inVero cells resulted in a single amino acid change (Q400R) in the Eprotein of ChinDENV at passage 15 and a single silent nucleotidemutation (C8666T) within NS5 gene at passage 9. Neither muta-tion affected the phenotypic trait of plaque size of ChinDENV(Fig. 2C). These results suggest that the ChinDENV chimera canbe convenient for scale-up and industrialization.
Loss of or low mouse virulence is an important attenuationmarker for flaviviruses. In our study, the chimera ChinDENV ex-hibited a significant attenuation phenotype, with an i.c. LD50
about 115-fold and 50-fold higher than those of its parental vi-ruses JEV SA14-14-2 and DENV-2 D2-43, respectively. In addi-tion, lack of neuroinvasiveness of ChinDENV as well as its paren-tal viruses in the adult mouse demonstrates that ChinDENV doesnot exceed its parental viruses in peripheral virulence. Attenua-tion of ChinDENV was further assessed by determining itsviremia profile in rhesus monkeys, which is correlated closely tothe phenotype of attenuation in humans (51, 52). No viremia wasdetected in any ChinDENV-inoculated monkey, even at a highdose (105 PFU) (Table 3), while monkeys developed high levels ofviremia following inoculation with the same dose of wt DENV-2(Table 3). These data indicate a favorable safety profile forChinDENV.
A single dose of ChinDENV stimulated both IgG and neutral-izing antibodies against DENV-2 in mice, and passive serum ofmice immunized with ChinDENV conferred complete protectionagainst DENV-2 challenge in suckling mice. In rhesus monkeys,the chimeric ChinDENV rapidly elicited a neutralizing antibodyresponse to wt DENV-2 in a dose-dependent manner (Table 4). Itwas observed that the monkeys immunized with high and middledoses of ChinDENV showed no signs of viremia postchallenge,and the mean peak titer and mean duration of viremia in low-dosegroups were significantly reduced compared with those of thePBS-immunized group after challenge with wt DENV-2 (Table 5).Especially, we found that one monkey with viremia postchallengein the low-dose group had a neutralizing antibody titer of 1:74.Sanofi’s recent phase 2b trial also showed a lack of protective effi-cacy against DENV-2 infection despite the induction of reason-able levels of neutralizing antibodies against all four serotypes onvaccination (13). The explanation for this phenomenon remains apuzzle.
Recall immunity after challenge plays a critical role in protec-tion against flavivirus (53). The role of anamnestic responses forreducing viremia after challenge has been supported by previousstudies (54–60). We also found that monkeys immunized withChinDENV and challenged s.c. with wt DENV-2 had dramaticincreases in serum neutralizing antibodies, suggesting a goodmemory component in ChinDENV immunity. However, thepresence of an anamnestic immune response, even in animals thatdeveloped no detectable viremia, demonstrating limited or local-ized viral replication, has been shown in those animals. Neverthe-less, the boost in antibody responses postchallenge would guaranteethat upon natural exposure, vaccinated individuals would respondwith a rapid and robust immune response. Taken together, theseresults demonstrate that the chimeric virus ChinDENV exhibits asatisfactory balance between attenuation and immunogenicityagainst DENV-2 in mice and monkeys.
JE vaccination has been proven to be the only effective measurefor disease control (61), and the live vaccine SA14-14-2 is widelyused in most areas where JE is endemic. Interestingly, a singleimmunization with ChinDENV provided full protection againstlethal JEV challenge in BALB/c mice (Fig. 4). The existence ofcross protection between heterologous but closely related mem-bers of the Flaviviridae family has been reported (62–66). Neutral-ization assay showed that the titer of neutralization antibodyagainst JEV was relatively low and may not account for the in vivoprotection. The contribution of the NS1-induced immune re-sponse should not be ruled out, since flavivirus NS1 protein hasbeen shown to induce protective immune responses in animals(67–73). Notably, immunization with ChinDENV elicited signif-icantly increased JEV antigen-specific T cell responses (IFN-� andIL-2), suggesting a possible role for cellular immunity in the de-fense against JEV challenge in ChinDENV-immunized mice. Tlymphocytes have been found to be necessary for protection fromWNV (74) and DENV (75, 76) infections in animal models. Mostidentified flavivirus T cell epitopes reside in the nonstructural pro-teins (77–80), the majority of which concentrated within the NS3protein (reviewed in reference 81). Although the contribution ofcellular immune responses to recovery from JEV infection re-mains unclear, a requirement for both CD4� and CD8� T cells tocontrol infections in the central nervous system has also been shownfor JEV (82). Collectively, these data suggest that ChinDENV canafford a cross protection against lethal JEV challenge in mice in
association with both the humoral and cellular immune re-sponses.
The DENV vaccine candidate ChinDENV described here rep-resents a significant advancement for dengue vaccine develop-ment. First, ChinDENV utilizes the JE vaccine SA14-14-2, whichhas demonstrated an ideal safety profile in large-scale clinical us-age, as the genetic backbone. Meanwhile, for YFV-17D, the back-bone for CYD in clinical trials, vaccination-associated adverseevents have been reported (83–87). Second, the Sanofi Pasteur’sCYD2 vaccine carries the prM-E genes of Asian genotype I ofDENV-2 (PUO-218 strain). The antigenic mismatch between theCYD2 vaccine virus and the DENV-2 circulating in Thailand cur-rently has been partially attributed to the failure of protection inthe clinical trials in Thailand (13). ChinDENV carries the protec-tive antigens of DENV-2 strain D2-43 (Asian genotype II). In par-ticular, there are five amino acid substitutions in the E protein(residues 71, 141, 164, 402, and 484) between ChinDENV andCYD2. Several of them are solvent exposed and involve potentialneutralizing antibody epitope recognition (88). Third, JEV andDENV are currently cocirculating in many Asian countries, wherethe socioeconomic burden of both diseases is significant. A vac-cine candidate that could protect against both DENV and JEV willbe highly attractive.
In summary, the chimeric virus ChinDENV is characterized bya highly attenuated profile in mice, a lack of viremia in rhesusmonkeys, satisfactory immunogenicity, and protective efficacy forDENV-2 infection in mice and monkeys. In particular, the chi-mera is capable of providing cross protection from JEV infectionin mice. This novel live attenuated chimeric dengue vaccine basedon JEV vaccine shows great potential for further development.
ACKNOWLEDGMENTS
We thank Guan-Mu Dong and Li-Li Jia (National Institute for Food andDrug Control, Beijing) for helpful discussion.
This work was supported by the National Basic Research Project ofChina (no. 2012CB518904), the National Natural Science Foundation ofChina (no. 31300151, 81101243, U1132002, and 31270974), and the Na-tional Major Special Program of Science and Technology of China(2008ZX10004-015 and 2013ZX10004001). C.-F.Q. was supported by theBeijing Nova Program of Science and Technology (no. 2010B041).
REFERENCES1. WHO. 2012. Dengue and severe dengue. Fact sheet no. 117. WHO, Ge-
Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB,Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. 2013.The global distribution and burden of dengue. Nature 496:504 –507.
3. Bray M, Lai CJ. 1991. Construction of intertypic chimeric dengue virusesby substitution of structural protein genes. Proc. Natl. Acad. Sci. U. S. A.88:10342–10346.
4. Pletnev AG, Bray M, Huggins J, Lai CJ. 1992. Construction and char-acterization of chimeric tick-borne encephalitis/dengue type 4 viruses.Proc. Natl. Acad. Sci. U. S. A. 89:10532–10536.
5. Chambers TJ, Nestorowicz A, Mason PW, Rice CM. 1999. Yellowfever/Japanese encephalitis chimeric viruses: construction and biologicalproperties. J. Virol. 73:3095–3101.
6. Huang CY, Butrapet S, Pierro DJ, Chang GJ, Hunt AR, BhamarapravatiN, Gubler DJ, Kinney RM. 2000. Chimeric dengue type 2 (vaccine strainPDK-53)/dengue type 1 virus as a potential candidate dengue type 1 virusvaccine. J. Virol. 74:3020 –3028.
7. Bray M, Men R, Lai CJ. 1996. Monkeys immunized with intertypicchimeric dengue viruses are protected against wild-type virus challenge. J.Virol. 70:4162– 4166.
8. Guirakhoo F, Weltzin R, Chambers TJ, Zhang ZX, Soike K, RatterreeM, Arroyo J, Georgakopoulos K, Catalan J, Monath TP. 2000. Recom-binant chimeric yellow fever-dengue type 2 virus is immunogenic andprotective in nonhuman primates. J. Virol. 74:5477–5485.
9. Guirakhoo F, Arroyo J, Pugachev KV, Miller C, Zhang ZX, Weltzin R,Georgakopoulos K, Catalan J, Ocran S, Soike K, Ratterree M, MonathTP. 2001. Construction, safety, and immunogenicity in nonhuman pri-mates of a chimeric yellow fever-dengue virus tetravalent vaccine. J. Virol.75:7290 –7304.
10. Huang CY, Butrapet S, Tsuchiya KR, Bhamarapravati N, Gubler DJ,Kinney RM. 2003. Dengue 2 PDK-53 virus as a chimeric carrier for tet-ravalent dengue vaccine development. J. Virol. 77:11436 –11447.
11. Whitehead SS, Hanley KA, Blaney JE, Jr, Gilmore LE, Elkins WR,Murphy BR. 2003. Substitution of the structural genes of dengue virustype 4 with those of type 2 results in chimeric vaccine candidates which areattenuated for mosquitoes, mice, and rhesus monkeys. Vaccine 21:4307–4316.
12. Guy B, Barrere B, Malinowski C, Saville M, Teyssou R, Lang J. 2011.From research to phase III: preclinical, industrial and clinical develop-ment of the Sanofi Pasteur tetravalent dengue vaccine. Vaccine 29:7229 –7241.
13. Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chantha-vanich P, Suvannadabba S, Jiwariyavej V, Dulyachai W, Pengsaa K,Wartel TA, Moureau A, Saville M, Bouckenooghe A, Viviani S, Torn-ieporth NG, Lang J. 2012. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a ran-domised, controlled phase 2b trial. Lancet 380:1559 –1567.
14. Guy B, Almond J, Lang J. 2011. Dengue vaccine prospects: a step forward.Lancet 377:381–382.
15. Butrapet S, Huang CY, Pierro DJ, Bhamarapravati N, Gubler DJ,Kinney RM. 2000. Attenuation markers of a candidate dengue type 2vaccine virus, strain 16681 (PDK-53), are defined by mutations in the 5=noncoding region and nonstructural proteins 1 and 3. J. Virol. 74:3011–3019.
16. Osorio JE, Huang CY, Kinney RM, Stinchcomb DT. 2011. Developmentof DENVax: a chimeric dengue-2 PDK-53-based tetravalent vaccine forprotection against dengue fever. Vaccine 29:7251–7260.
17. Blaney JE, Jr, Matro JM, Murphy BR, Whitehead SS. 2005. Recombi-nant, live-attenuated tetravalent dengue virus vaccine formulations in-duce a balanced, broad, and protective neutralizing antibody responseagainst each of the four serotypes in rhesus monkeys. J. Virol. 79:5516 –5528.
18. Durbin AP, McArthur JH, Marron JA, Blaney JE, Thumar B, WanionekK, Murphy BR, Whitehead SS. 2006. rDEN2/4Delta30(ME), a live atten-uated chimeric dengue serotype 2 vaccine is safe and highly immunogenicin healthy dengue-naive adults. Hum. Vaccin. 2:255–260.
19. Pletnev AG, Swayne DE, Speicher J, Rumyantsev AA, Murphy BR.2006. Chimeric West Nile/dengue virus vaccine candidate: preclinicalevaluation in mice, geese and monkeys for safety and immunogenicity.Vaccine 24:6392– 6404.
20. Durbin AP, Whitehead SS, Shaffer D, Elwood D, Wanionek K, ThumarB, Blaney JE, Murphy BR, Schmidt AC. 2011. A single dose of theDENV-1 candidate vaccine rDEN1Delta30 is strongly immunogenic andinduces resistance to a second dose in a randomized trial. PLoS Negl. Trop.Dis. 5:e1267. doi:10.1371/journal.pntd.0001267.
21. Yu Y. 2010. Phenotypic and genotypic characteristics of Japanese enceph-alitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine28:3635–3641.
22. Bista MB, Banerjee MK, Shin SH, Tandan JB, Kim MH, Sohn YM, OhrrHC, Tang JL, Halstead SB. 2001. Efficacy of single-dose SA 14-14-2vaccine against Japanese encephalitis: a case control study. Lancet 358:791–795.
23. Gatchalian S, Yao Y, Zhou B, Zhang L, Yoksan S, Kelly K, Neuzil KM,Yaich M, Jacobson J. 2008. Comparison of the immunogenicity andsafety of measles vaccine administered alone or with live, attenuated Jap-anese encephalitis SA 14-14-2 vaccine in Philippine infants. Vaccine 26:2234 –2241.
24. Hennessy S, Liu Z, Tsai TF, Strom BL, Wan CM, Liu HL, Wu TX, YuHJ, Liu QM, Karabatsos N, Bilker WB, Halstead SB. 1996. Effectivenessof live-attenuated Japanese encephalitis vaccine (SA14-14-2): a case-control study. Lancet 347:1583–1586.
25. Kumar R, Tripathi P, Rizvi A. 2009. Effectiveness of one dose of SA
A Chimeric JEV/Dengue Vaccine
December 2013 Volume 87 Number 24 jvi.asm.org 13703
14-14-2 vaccine against Japanese encephalitis. N. Engl. J. Med. 360:1465–1466.
26. Tandan JB, Ohrr H, Sohn YM, Yoksan S, Ji M, Nam CM, Halstead SB.2007. Single dose of SA 14-14-2 vaccine provides long-term protectionagainst Japanese encephalitis: a case-control study in Nepalese children 5years after immunization. Vaccine 25:5041–5045.
27. Deng YQ, Dai JX, Ji GH, Jiang T, Wang HJ, Yang HO, Tan WL, Liu R,Yu M, Ge BX, Zhu QY, Qin ED, Guo YJ, Qin CF. 2011. A broadlyflavivirus cross-neutralizing monoclonal antibody that recognizes a novelepitope within the fusion loop of E protein. PLoS One 6:e16059. doi:10.1371/journal.pone.0016059.
28. Ye Q, Li XF, Zhao H, Li SH, Deng YQ, Cao RY, Song KY, Wang HJ,Hua RH, Yu YX, Zhou X, Qin ED, Qin CF. 2012. A single nucleotidemutation in NS2A of Japanese encephalitis-live vaccine virus (SA14-14-2)ablates NS1= formation and contributes to attenuation. J. Gen. Virol. 93:1959 –1964.
29. Reed LR, Muench H. 1938. A simple method of estimating fifty percentendpoints. Am. J. Hyg. 27:493– 497.
30. Weatherall D 2006. The use of non-human primates in research. TheRoyal Society, London, United Kingdom.
31. Li SH, Li XF, Zhao H, Deng YQ, Yu XD, Zhu SY, Jiang T, Ye Q, QinED, Qin CF. 2013. Development and characterization of the repliconsystem of Japanese encephalitis live vaccine virus SA14-14-2. Virol. J. 10:64. doi:10.1186/1743-422X-10-64.
32. Cao F, Li XF, Yu XD, Deng YQ, Jiang T, Zhu QY, Qin ED, Qin CF.2011. A DNA-based West Nile virus replicon elicits humoral and cellularimmune responses in mice. J. Virol. Methods 178:87–93.
33. Rosen L. 1999. Comments on the epidemiology, pathogenesis and controlof dengue. Med. Trop. 59:495– 498.
34. Whitehead RH, Chaicumpa V, Olson LC, Russell PK. 1970. Sequentialdengue virus infections in the white-handed gibbon (Hylobates lar). Am.J. Trop. Med. Hyg. 19:94 –102.
35. Guy B, Nougarede N, Begue S, Sanchez V, Souag N, Carre M, Cham-bonneau L, Morrisson DN, Shaw D, Qiao M, Dumas R, Lang J, ForratR. 2008. Cell-mediated immunity induced by chimeric tetravalent denguevaccine in naive or flavivirus-primed subjects. Vaccine 26:5712–5721.
36. Amberg SM, Rice CM. 1999. Mutagenesis of the NS2B-NS3-mediatedcleavage site in the flavivirus capsid protein demonstrates a requirementfor coordinated processing. J. Virol. 73:8083– 8094.
37. Lobigs M, Lee E. 2004. Inefficient signalase cleavage promotes efficientnucleocapsid incorporation into budding flavivirus membranes. J. Virol.78:178 –186.
38. Yamshchikov VF, Compans RW. 1995. Formation of the flavivirus en-velope: role of the viral NS2B-NS3 protease. J. Virol. 69:1995–2003.
39. Cocquerel L, Wychowski C, Minner F, Penin F, Dubuisson J. 2000.Charged residues in the transmembrane domains of hepatitis C virus gly-coproteins play a major role in the processing, subcellular localization,and assembly of these envelope proteins. J. Virol. 74:3623–3633.
40. Lindenbach BD, Thiel H-J, Rice CM. 2007. Flaviviruses: the viruses andtheir replication, p 1101–1152. In Knipe DM, Howley PM, Griffin DE,Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed.Lippincott Williams and Wilkins, Philadelphia, PA.
41. Bhatt TR, Crabtree MB, Guirakhoo F, Monath TP, Miller BR. 2000.Growth characteristics of the chimeric Japanese encephalitis virus vaccinecandidate, ChimeriVax-JE (YF/JE SA14 –14 –2), in Culex tritaenio-rhynchus, Aedes albopictus, and Aedes aegypti mosquitoes. Am. J. Trop.Med. Hyg. 62:480 – 484.
42. Chambers TJ, Liang Y, Droll DA, Schlesinger JJ, Davidson AD, WrightPJ, Jiang X. 2003. Yellow fever virus/dengue-2 virus and yellow fevervirus/dengue-4 virus chimeras: biological characterization, immunoge-nicity, and protection against dengue encephalitis in the mouse model. J.Virol. 77:3655–3668.
43. Johnson BW, Chambers TV, Crabtree MB, Arroyo J, Monath TP,Miller BR. 2003. Growth characteristics of the veterinary vaccine candi-date ChimeriVax-West Nile (WN) virus in Aedes and Culex mosquitoes.Med. Vet. Entomol. 17:235–243.
44. Johnson BW, Chambers TV, Crabtree MB, Bhatt TR, Guirakhoo F,Monath TP, Miller BR. 2002. Growth characteristics of ChimeriVax-DEN2 vaccine virus in Aedes aegypti and Aedes albopictus mosquitoes.Am. J. Trop. Med. Hyg. 67:260 –265.
45. Johnson BW, Chambers TV, Crabtree MB, Guirakhoo F, Monath TP,Miller BR. 2004. Analysis of the replication kinetics of the ChimeriVax-DEN 1, 2, 3, 4 tetravalent virus mixture in Aedes aegypti by real-time
reverse transcriptase-polymerase chain reaction. Am. J. Trop. Med. Hyg.70:89 –97.
46. McGee CE, Lewis MG, Claire MS, Wagner W, Lang J, Guy B, TsetsarkinK, Higgs S, Decelle T. 2008. Recombinant chimeric virus with wild-typedengue 4 virus premembrane and envelope and virulent yellow fever virusAsibi backbone sequences is dramatically attenuated in nonhuman pri-mates. J. Infect. Dis. 197:693– 697.
47. McGee CE, Tsetsarkin K, Vanlandingham DL, McElroy KL, Lang J, GuyB, Decelle T, Higgs S. 2008. Substitution of wild-type yellow fever Asibisequences for 17D vaccine sequences in ChimeriVax-dengue 4 does notenhance infection of Aedes aegypti mosquitoes. J. Infect. Dis. 197:686 –692.
48. Pletnev AG, Men R. 1998. Attenuation of the Langat tick-borne flavivirusby chimerization with mosquito-borne flavivirus dengue type 4. Proc.Natl. Acad. Sci. U. S. A. 95:1746 –1751.
49. Pugachev KV, Schwaiger J, Brown N, Zhang ZX, Catalan J, Mitchell FS,Ocran SW, Rumyantsev AA, Khromykh AA, Monath TP, Guirakhoo F.2007. Construction and biological characterization of artificial recombi-nants between a wild type flavivirus (Kunjin) and a live chimeric flavivirusvaccine (ChimeriVax-JE). Vaccine 25:6661– 6671.
50. Taucher C, Berger A, Mandl CW. 2010. A trans-complementing recom-bination trap demonstrates a low propensity of flaviviruses for intermo-lecular recombination. J. Virol. 84:599 – 611.
51. Durbin AP, Karron RA, Sun W, Vaughn DW, Reynolds MJ, PerreaultJR, Thumar B, Men R, Lai CJ, Elkins WR, Chanock RM, Murphy BR,Whitehead SS. 2001. Attenuation and immunogenicity in humans of alive dengue virus type-4 vaccine candidate with a 30 nucleotide deletion inits 3=-untranslated region. Am. J. Trop. Med. Hyg. 65:405– 413.
52. Innis BL, Eckels KH, Kraiselburd E, Dubois DR, Meadors GF, GublerDJ, Burke DS, Bancroft WH. 1988. Virulence of a live dengue virusvaccine candidate: a possible new marker of dengue virus attenuation. J.Infect. Dis. 158:876 – 880.
53. Konishi E, Yamaoka M, Khin Sane W, Kurane I, Takada K, Mason PW.1999. The anamnestic neutralizing antibody response is critical for pro-tection of mice from challenge following vaccination with a plasmid en-coding the Japanese encephalitis virus premembrane and envelope genes.J. Virol. 73:5527–5534.
54. Bernardo L, Hermida L, Martin J, Alvarez M, Prado I, Lopez C,Martinez R, Rodriguez-Roche R, Zulueta A, Lazo L, Rosario D, GuillenG, Guzman MG. 2008. Anamnestic antibody response after viral chal-lenge in monkeys immunized with dengue 2 recombinant fusion proteins.Arch. Virol. 153:849 – 854.
55. Bernardo L, Izquierdo A, Alvarez M, Rosario D, Prado I, Lopez C,Martinez R, Castro J, Santana E, Hermida L, Guillen G, Guzman MG.2008. Immunogenicity and protective efficacy of a recombinant fusionprotein containing the domain III of the dengue 1 envelope protein innon-human primates. Antiviral Res. 80:194 –199.
56. Blair PJ, Kochel TJ, Raviprakash K, Guevara C, Salazar M, Wu SJ,Olson JG, Porter KR. 2006. Evaluation of immunity and protective effi-cacy of a dengue-3 pre-membrane and envelope DNA vaccine in Aotusnancymae monkeys. Vaccine 24:1427–1432.
57. Guzmán MG, Rodriguez R, Hermida L, Alvarez M, Lazo L, Mune M,Rosario D, Valdes K, Vazquez S, Martinez R, Serrano T, Paez J,Espinosa R, Pumariega T, Guillen G. 2003. Induction of neutralizingantibodies and partial protection from viral challenge in Macaca fascicu-laris immunized with recombinant dengue 4 virus envelope glycoproteinexpressed in Pichia pastoris. Am. J. Trop. Med. Hyg. 69:129 –134.
58. Markoff L, Pang X, Houng HS, Falgout B, Olsen R, Jones E, Polo S.2002. Derivation and characterization of a dengue type 1 host range-restricted mutant virus that is attenuated and highly immunogenic inmonkeys. J. Virol. 76:3318 –3328.
59. Raviprakash K, Porter KR, Kochel TJ, Ewing D, Simmons M, Phillips I,Murphy GS, Weiss WR, Hayes CG. 2000. Dengue virus type 1 DNAvaccine induces protective immune responses in rhesus macaques. J. Gen.Virol. 81:1659 –1667.
60. Robert Putnak J, Coller BA, Voss G, Vaughn DW, Clements D, PetersI, Bignami G, Houng HS, Chen RC, Barvir DA, Seriwatana J, CayphasS, Garcon N, Gheysen D, Kanesa-Thasan N, McDonell M, HumphreysT, Eckels KH, Prieels JP, Innis BL. 2005. An evaluation of dengue type-2inactivated, recombinant subunit, and live-attenuated vaccine candidatesin the rhesus macaque model. Vaccine 23:4442– 4452.
61. Halstead SB, Thomas SJ. 2011. New Japanese encephalitis vaccines: al-
ternatives to production in mouse brain. Expert Rev. Vaccines 10:355–364.
62. Yamshchikov G, Borisevich V, Kwok CW, Nistler R, Kohlmeier J,Seregin A, Chaporgina E, Benedict S, Yamshchikov V. 2005. The suit-ability of yellow fever and Japanese encephalitis vaccines for immuniza-tion against West Nile virus. Vaccine 23:4785– 4792.
63. Williams DT, Daniels PW, Lunt RA, Wang LF, Newberry KM, Mack-enzie JS. 2001. Experimental infections of pigs with Japanese encephalitisvirus and closely related Australian flaviviruses. Am. J. Trop. Med. Hyg.65:379 –387.
64. Tesh RB, Travassos da Rosa AP, Guzman H, Araujo TP, Xiao SY. 2002.Immunization with heterologous flaviviruses protective against fatal WestNile encephalitis. Emerg. Infect. Dis. 8:245–251.
65. Takasaki T, Yabe S, Nerome R, Ito M, Yamada K, Kurane I. 2003.Partial protective effect of inactivated Japanese encephalitis vaccine onlethal West Nile virus infection in mice. Vaccine 21:4514 – 4518.
66. Kanesa-Thasan N, Putnak JR, Mangiafico JA, Saluzzo JE, Ludwig GV.2002. Short report: absence of protective neutralizng antibodies to WestNile virus in subjects following vaccination with Japanese encephalitis ordengue vaccines. Am. J. Trop. Med. Hyg. 66:115–116.
67. Schlesinger JJ, Brandriss MW, Cropp CB, Monath TP. 1986. Protectionagainst yellow fever in monkeys by immunization with yellow fever virusnonstructural protein NS1. J. Virol. 60:1153–1155.
68. Falgout B, Bray M, Schlesinger JJ, Lai CJ. 1990. Immunization of micewith recombinant vaccinia virus expressing authentic dengue virus non-structural protein NS1 protects against lethal dengue virus encephalitis. J.Virol. 64:4356 – 4363.
69. Amorim JH, Diniz MO, Cariri FA, Rodrigues JF, Bizerra RS, GoncalvesAJ, de Barcelos Alves AM, de Souza Ferreira LC. 2012. Protectiveimmunity to DENV2 after immunization with a recombinant NS1 proteinusing a genetically detoxified heat-labile toxin as an adjuvant. Vaccine30:837– 845.
70. Li Y, Counor D, Lu P, Duong V, Yu Y, Deubel V. 2012. Protectiveimmunity to Japanese encephalitis virus associated with anti-NS1 anti-bodies in a mouse model. Virol. J. 9:135.
71. Chung KM, Nybakken GE, Thompson BS, Engle MJ, Marri A, FremontDH, Diamond MS. 2006. Antibodies against West Nile virus nonstruc-tural protein NS1 prevent lethal infection through Fc gamma receptor-dependent and -independent mechanisms. J. Virol. 80:1340 –1351.
72. Lee TH, Song BH, Yun SI, Woo HR, Lee YM, Diamond MS, ChungKM. 2012. A cross-protective mAb recognizes a novel epitope within theflavivirus NS1 protein. J. Gen. Virol. 93:20 –26.
73. Lin YL, Chen LK, Liao CL, Yeh CT, Ma SH, Chen JL, Huang YL, ChenSS, Chiang HY. 1998. DNA immunization with Japanese encephalitisvirus nonstructural protein NS1 elicits protective immunity in mice. J.Virol. 72:191–200.
74. Shresta S, Kyle JL, Snider HM, Basavapatna M, Beatty PR, Harris E.2004. Interferon-dependent immunity is essential for resistance to pri-mary dengue virus infection in mice, whereas T- and B-cell-dependentimmunity are less critical. J. Virol. 78:2701–2710.
75. Yauch LE, Zellweger RM, Kotturi MF, Qutubuddin A, Sidney J, PetersB, Prestwood TR, Sette A, Shresta S. 2009. A protective role for denguevirus-specific CD8� T cells. J. Immunol. 182:4865– 4873.
76. Yauch LE, Prestwood TR, May MM, Morar MM, Zellweger RM, PetersB, Sette A, Shresta S. 2010. CD4� T cells are not required for the induc-tion of dengue virus-specific CD8� T cell or antibody responses but con-tribute to protection after vaccination. J. Immunol. 185:5405–5416.
77. Weiskopf D, Yauch LE, Angelo MA, John DV, Greenbaum JA, SidneyJ, Kolla RV, De Silva AD, de Silva AM, Grey H, Peters B, Shresta S, SetteA. 2011. Insights into HLA-restricted T cell responses in a novel mousemodel of dengue virus infection point toward new implications for vac-cine design. J. Immunol. 187:4268 – 4279.
78. Duangchinda T, Dejnirattisai W, Vasanawathana S, Limpitikul W,Tangthawornchaikul N, Malasit P, Mongkolsapaya J, Screaton G. 2010.Immunodominant T-cell responses to dengue virus NS3 are associatedwith DHF. Proc. Natl. Acad. Sci. U. S. A. 107:16922–16927.
79. Bashyam HS, Green S, Rothman AL. 2006. Dengue virus-reactive CD8�T cells display quantitative and qualitative differences in their response tovariant epitopes of heterologous viral serotypes. J. Immunol. 176:2817–2824.
80. Akondy RS, Monson ND, Miller JD, Edupuganti S, Teuwen D, Wu H,Quyyumi F, Garg S, Altman JD, Del Rio C, Keyserling HL, Ploss A, RiceCM, Orenstein WA, Mulligan MJ, Ahmed R. 2009. The yellow fevervirus vaccine induces a broad and polyfunctional human memory CD8�T cell response. J. Immunol. 183:7919 –7930.
81. Vaughan K, Greenbaum J, Blythe M, Peters B, Sette A. 2010. Meta-analysis of all immune epitope data in the Flavivirus genus: inventory ofcurrent immune epitope data status in the context of virus immunity andimmunopathology. Viral Immunol. 23:259 –284.
82. Shrestha B, Pinto AK, Green S, Bosch I, Diamond MS. 2012. CD8� Tcells use TRAIL to restrict West Nile virus pathogenesis by controllinginfection in neurons. J. Virol. 86:8937– 8948.
83. Khromava AY, Eidex RB, Weld LH, Kohl KS, Bradshaw RD, Chen RT,Cetron MS. 2005. Yellow fever vaccine: an updated assessment of ad-vanced age as a risk factor for serious adverse events. Vaccine 23:3256 –3263.
84. Lindsey NP, Schroeder BA, Miller ER, Braun MM, Hinckley AF,Marano N, Slade BA, Barnett ED, Brunette GW, Horan K, Staples JE,Kozarsky PE, Hayes EB. 2008. Adverse event reports following yellowfever vaccination. Vaccine 26:6077– 6082.
85. Pulendran B, Miller J, Querec TD, Akondy R, Moseley N, Laur O,Glidewell J, Monson N, Zhu T, Zhu H, Staprans S, Lee D, Brinton MA,Perelygin AA, Vellozzi C, Brachman P, Jr, Lalor S, Teuwen D, Eidex RB,Cetron M, Priddy F, del Rio C, Altman J, Ahmed R. 2008. Case of yellowfever vaccine-associated viscerotropic disease with prolonged viremia, ro-bust adaptive immune responses, and polymorphisms in CCR5 andRANTES genes. J. Infect. Dis. 198:500 –507.
86. Silva ML, Espirito-Santo LR, Martins MA, Silveira-Lemos D, Peruhype-Magalhaes V, Caminha RC, de Andrade Maranhao-Filho P, Auxili-adora-Martins M, de Menezes Martins R, Galler R, da Silva Freire M,Marcovistz R, Homma A, Teuwen DE, Eloi-Santos SM, Andrade MC,Teixeira-Carvalho A, Martins-Filho OA. 2010. Clinical and immunolog-ical insights on severe, adverse neurotropic and viscerotropic disease fol-lowing 17D yellow fever vaccination. Clin. Vaccine Immunol. 17:118 –126.
88. Sukupolvi-Petty S, Austin SK, Engle M, Brien JD, Dowd KA, WilliamsKL, Johnson S, Rico-Hesse R, Harris E, Pierson TC, Fremont DH,Diamond MS. 2010. Structure and function analysis of therapeuticmonoclonal antibodies against dengue virus type 2. J. Virol. 84:9227–9239.
A Chimeric JEV/Dengue Vaccine
December 2013 Volume 87 Number 24 jvi.asm.org 13705