BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. A Method for Extracting High-Quality RNA from Diverse Plants for Next- Generation Sequencing and Gene Expression Analyses Author(s): Roxana Yockteng Ana M. R. Almeida , Stephen Yee , Thiago Andre , Colin Hill , and Chelsea D. Specht Source: Applications in Plant Sciences, 1(12) 2014. Published By: Botanical Society of America DOI: http://dx.doi.org/10.3732/apps.1300070 URL: http://www.bioone.org/doi/full/10.3732/apps.1300070 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
A Method for Extracting High-Quality RNA from Diverse Plants for Next-Generation Sequencing and Gene Expression AnalysesAuthor(s): Roxana Yockteng Ana M. R. Almeida , Stephen Yee , Thiago Andre , Colin Hill , and ChelseaD. SpechtSource: Applications in Plant Sciences, 1(12) 2014.Published By: Botanical Society of AmericaDOI: http://dx.doi.org/10.3732/apps.1300070URL: http://www.bioone.org/doi/full/10.3732/apps.1300070
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.
High-throughput, next-generation sequencing of genomes and transcriptomes requires high-quality, clean, and concentrated ge-netic material. While getting high-quality genomic extractions from plants is not trivial, it is even more challenging to consis-tently obtain suffi cient quantity of pure RNA for RNA-Seq (tran-scriptome) or chromatin immunoprecipitation followed by next-generation sequencing (ChIP-Seq) experiments. Due to its instability, RNA is more delicate to manipulate than genomic DNA. It is often single stranded and contains ribose sugars; the 2 ′ -hydroxyl group attached to the ribose makes RNA more prone to hydrolysis. For studies of plant biology, high-quality RNA
extraction is further complicated when plant samples contain large quantities of polysaccharides, high levels of RNases, vari-ous different kinds of phenolics including tannins, low concen-trations of nucleic acids (high water content), and/or fi brous tissues such as lignin (wood) that are diffi cult to break up and remove ( MacRae, 2007 ).
A large number of plant-specifi c protocols are available, rec-ommending a diversity of strategies for tissue storage, maceration, preparation, and extraction to obtain suffi cient RNA from plants depending on the species or lineage of plant, the tissue type be-ing extracted, and the presence of secondary metabolites and inhibitors for any given plant lineage or plant part (for reviews see MacRae, 2007 ; Accerbi et al., 2010 ). For example, Ghawana et al. (2011) propose a phenol-based method to isolate RNA in species with high presence of secondary metabolites such as rheum ( Rheum austral D . Don) and arnebia ( Arnebia euchroma (Royle) I. M. Johnst.). A method proposed by Gehrig et al. (2000) uses high-molecular-weight polyethylene glycol (PEG) to extract RNA from species with high presence of polyphenols and polysaccharides (e.g., Aloe L., Ananas Mill., Clusia L., Euphorbia L.). Other protocols are designed to extract RNA from a specifi c type of tissue, such as the trizol-based methods proposed to extract RNA from siliques and seeds of Arabidop-sis (DC.) Heynh. ( Meng and Feldman, 2010 ) or the seeds of Davidia involucrata Baill. ( Qi et al., 2009 ).
Other methods focus on rapid methods for RNA extraction, although these are typically proposed only for model organ-isms (e.g., Arabidopsis leaves) and are limited to PCR-based
1 Manuscript received 29 August 2013; revision accepted 18 November 2013.
The authors thank the Specht Laboratory for sharing ideas and data; T. Renner (carnivores), S. Salzman, I. Terry, and B. Dorsey (cycads), F. Zapata (Burseraceae), A. Jackson and Mi Yeon Lee ( Nicotiana and Hordeum ), and K. Morioka (monocots) for troubleshooting and testing; M. Bartlett for introducing the laboratory to Invitrogen’s RNA reagent; the Evolutionary Genetics Lab, Museum of Vertebrate Zoology, for supporting next-gen sequencing among the Berkeley Natural History Museums; and T. Gregory for providing cycad tissue and fi nancial support. Additional funding was provided by the National Science Foundation (IOS 0845641, DEB 1110461), the University of California Institute for Mexico (UC MEXUS)–Consejo Nacional de Ciencia y Tecnología (CONACYT), the Hellman Family Fund, and a Prytanean Junior Faculty Award.
A METHOD FOR EXTRACTING HIGH-QUALITY RNA FROM DIVERSE PLANTS FOR NEXT-GENERATION SEQUENCING AND GENE
EXPRESSION ANALYSES 1
ROXANA YOCKTENG 2,3 , ANA M. R. ALMEIDA 2 , STEPHEN YEE 2 , THIAGO ANDRE 4 , COLIN HILL 2 , AND CHELSEA D. SPECHT 2,5
2 Department of Plant and Microbial Biology, Department of Integrative Biology, and the University and Jepson Herbaria, University of California, Berkeley, California 94720 USA; 3 Origine, Structure et Evolution de la Diversité (UMR 7205 CNRS),
Muséum National d’Histoire Naturelle, CP39, 16 rue Buffon, 75231 Paris Cedex 05, France; and 4 Laboratório de Taxonomia Vegetal e Biologia Reprodutiva, Departamento de Botânica, Universidade Federal do Rio de Janeiro, 21941-902 Rio de Janeiro,
Rio de Janeiro, Brazil
• Premise of the study: To study gene expression in plants, high-quality RNA must be extracted in quantities suffi cient for sub-sequent cDNA library construction. Field-based collections are often limited in quantity and quality of tissue and are typically preserved in RNA later . Obtaining suffi cient and high-quality yield from variously preserved samples is essential to studies of comparative biology. We present a protocol for the extraction of high-quality RNA from even the most recalcitrant plant tissues.
• Methods and Results: Tissues from mosses, cycads, and angiosperm fl oral organs and leaves were preserved in RNA later or frozen fresh at −80 ° C. Extractions were performed and quality was measured for yield and purity.
• Conclusions: This protocol results in the extraction of high-quality RNA from a variety of plant tissues representing vascular and nonvascular plants. RNA was used for cDNA synthesis to generate libraries for next-generation sequencing and for expres-sion studies using quantitative PCR (qPCR) and semiquantitative reverse transcription PCR (RT-PCR).
Applications in Plant Sciences 2013 1 ( 12 ): 1300070 Yockteng et al.—RNA extraction from diverse plant tissues doi:10.3732/apps.1300070
http://www.bioone.org/loi/apps
manually ground sample can be added to the FastPrep tube with Lysing Matrix beads (see above) for additional pulverization.
RNA extraction— Once the tissue is suffi ciently homogenized: 1. Add 0.6 mL of cold (4 ° C) Plant RNA Reagent (Life Technol ogies) to
pulverized tissue. Mix by brief vortexing or fl icking the bottom of the tube until the sample is thoroughly resuspended. If tissue was ground with a FastPrep, homogenize with the cold buffer for an additional 40 s. If the tissue is not com-pletely ground, repeat 1–2 × until the tissue is pulverized. If the tissue was ground with a mortar and pestle and does not need additional grinding, continue with the next step.
2. Incubate 5 min at room temperature. Placing the samples on a rotator or nutator will help to maximize surface area of the tissue with the extraction buffer.
3. Clarify the solution by centrifuging for 2 min at 12,000 × g in a microcen-trifuge at room temperature. Transfer the supernatant to a tube with Phase Lock Gel (5 Prime, Gaithersburg, Maryland, USA). Although the Phase Lock Gel tubes are not required, they greatly facilitate separation of the organic and aque-ous phases and help ensure cleanliness of the sample.
4. Add 0.1 mL of 5 M NaCl to the sample, tap tube to mix, and add 0.3 mL of chloroform–isoamyl alcohol (24 : 1). Mix thoroughly by inversion and cen-trifuge the sample at 4 ° C for 10 min at 12,000 × g to separate the phases. Trans-fer the aqueous (top) phase to an RNase-free tube.
5. Add to the aqueous phase an equal volume of a mix of LiCl (4 M) (3/4 v) and isopropyl alcohol (1/4 v). Mix and let stand at −20 ° C for 30 min to over-night. If the tissue was stored in RNA later , mix and let stand at −20 ° C for a maximum of 3 h (not overnight). If the precipitation is longer, salts from the RNA later solution could also precipitate. (Note: We have also let the sample stand at −80 ° C for 3 h, and this works as well.)
6. Centrifuge the sample at 4 ° C for 20 min at 12,000 × g . 7. Decant or remove supernatant with a pipette, taking care not to lose the
pellet. Add 1 mL of 75% ethanol to the pellet. The pellet may be diffi cult to see. To help to see the pellet, you can add 1 μ L of GlycoBlue (Ambion) in step 5.
8. Centrifuge at 4 ° C for 5 min at 12,000 × g . Decant the liquid carefully, taking care not to lose the pellet. Briefl y centrifuge to collect the residual liquid, and remove it with a pipette.
9. Let dry on ice for 15 min at room temperature and elute pellet in 10–30 μ L of RNase-free water. Pipette the water up and down over the pellet to dis-solve the RNA. If the pellet is diffi cult to dissolve, add more water or warm to 37 ° C to facilitate the dissolution. It is important to resuspend the pellet com-pletely to obtain an accurate measure of the concentration of your RNA. If the sample is not clean, it can be purifi ed by the cleanup step suggested later. Al-though some protocols have suggested that performing an additional step of chloroform would clean the RNA samples (e.g., Accerbi et al., 2010 ), we found that an additional chloroform step decreases the RNA yield substantially.
A NanoDrop ND-1000 spectrophotometer (Thermo Scientifi c, Wilmington, Delaware, USA) can be used to verify the concentration and purity of the RNA obtained. To assess the presence and purity of extracted RNA, use the ratio of absorbance at 230 nm, 260 nm, and 280 nm. If the RNA is pure, we expect a 260/280 ratio around 2, although this ratio does not guarantee pure RNA (see below). If the ratio is appreciably lower, it is an indication of the presence of protein, phenol, or other contaminants that absorb strongly at or near 280 nm. The ratio 260/230 is expected to be around 2–2.2. If this value is appreciably lower, it is an indication that contaminants such as carbohydrates, EDTA, guanidine isothiocyanate, and phenol that absorb at 230 nm are present in the sample. Ratios lower than expected could indicate that additional cleaning is necessary and the optional cleanup should be followed. While a more accurate assessment of the quality will be determined with a bioanalyzer prior to se-quencing, this initial NanoDrop read will provide an indication of the presence of RNA, enabling the researcher to continue.
Optional cleanup — If the sample is not clean, the following modifi ed cleanup procedure will help to purify the total RNA. This protocol is adapted from that published for DNA cleanup by Rohland and Reich (2012) , using mag-netic beads to capture nucleotide material and permit additional washing steps that aid in the removal of undesirable metabolites. All stock solutions and re-agents must be prepared with RNase-free water.
Preparing Sera-Mag beads solution— Begin by preparing 50 mL of Sera-Mag beads. Add 9 g of PEG-8000, 10 mL of 5 M NaCl, 500 μ L of 1 M Tris-HCl (pH 8.0), and 100 μ L of 0.5 M EDTA (pH 8.0) to a 50-mL Falcon tube. Add 1 mL (9 g) of the carboxyl-modifi ed Sera-Mag Magnetic SpeedBeads (Thermo
downstream use of the extracted RNA (e.g., Berendzen et al., 2005 ). Between October 2011 and September 2013, a total of 53 references for plant RNA extraction methods are found in Web of Science. Of these methods, 90% have only been tested in one plant lineage (e.g., Kumar and Singh, 2012 ; Gudenschwager et al., 2012 ) and the majority are relatively straightforward modifi cations of the cetyltrimethylammonium bromide (CTAB) method with polyvinylpyrrolidone (PVP; e.g., Japelaghi et al., 2011 ; Yin et al., 2011 ).
Given the variability of extraction techniques and the quality of their products, the ability to perform comparative analyses of genomes or transcriptomes across plant lineages and tissue types remains a challenge for researchers interested in ques-tions of plant evolution. Here, we present a general protocol that can be used for a wide range of plant tissues and across a broad range of taxa. Our intention in presenting this protocol is not to discount other protocols that work well in particular cases, but rather to present a single protocol that works well across a broad variety of plant tissue types and plant species. This modi-fi ed protocol uses the proprietary Plant RNA Reagent from Life Technologies (Carlsbad, California, USA) that is composed in part of 2-mercaptoethanol (10–30% weight) and sodium azide (0.1–1.0% weight).
We demonstrate the effi cacy of this extraction pipeline in dif-ferent plant species including mosses, gymnosperms, and an-giosperms, and demonstrate its ability to generate successful sequencing product from different plant tissue types, including individual fl oral and reproductive organs as well as combined reproductive and vegetative tissues.
METHODS AND RESULTS
Sampling — Plant material was collected from greenhouses and botanical gardens ( Table 1 ) and either stored in RNA later (Ambion, Carlsbad, California, USA) or frozen immediately in liquid nitrogen. Preserved tissue was placed in long-term storage at −80 ° C. For storage in RNA later , approximately 3 × the volume of RNA later : tissue is used. Tissue stored in RNA later and frozen (−80 ° C) was defrosted just enough to remove the tissue from the RNA later prior to extraction.
Basic protocol — The following protocol was modifi ed from the manufac-turer’s provided instructions for effective use of the Plant RNA Reagent from Life Technologies. As indicated, all solutions are prepared with sterile RNase-free water, and all supplies and handling materials are cleaned with RNase AWAY (Ambion) prior to dissection and storage. This protocol is optimized to isolate RNA from approximately 0.1 g of plant tissue. If the amount of plant tissue is increased, reagent volumes must be scaled appropriately.
Grinding the tissue— One of the critical points to obtain high yield in the extraction of genetic material is the grinding. It is essential to grind the tissue as fi nely as possible, maintaining samples as cold as possible during grinding to avoid degradation. Selection of FastPrep or mortars/pestle depends on the hard-ness of the tissue being processed.
A FastPrep FP120 Homogenizer (Thermo Savant, Carlsbad, California, USA) was used for grinding fl oral organs and soft leaf tissue. Approximately 0.1 g of frozen fl oral organs, whole fl owers, and leaves or herbaceous stems were added to FastPrep 2-mL tubes (MP Biomedicals, Santa Ana, California, USA) 1/5 fi lled with bulk Lysing Matrix D (MP Biomedicals). FastPrep tubes containing the frozen tissue plus Lysing Matrix beads were shaken in the ho-mogenizer (FastPrep) for 40 s at speed 6 (6 m/s) without buffer at room temperature.
For grinding hard tissue or ligneous tissue, such as cone scales from gymno-sperms, the tissue was ground under liquid nitrogen in a mortar and pestle that was sterilized and baked (minimum 12 h at 150 ° C). The tissue was ground as fi nely as possible, and the powdered material was placed in a 1.5-mL tube. The
Applications in Plant Sciences 2013 1 ( 12 ): 1300070 Yockteng et al.—RNA extraction from diverse plant tissues doi:10.3732/apps.1300070
3 of 6http://www.bioone.org/loi/apps
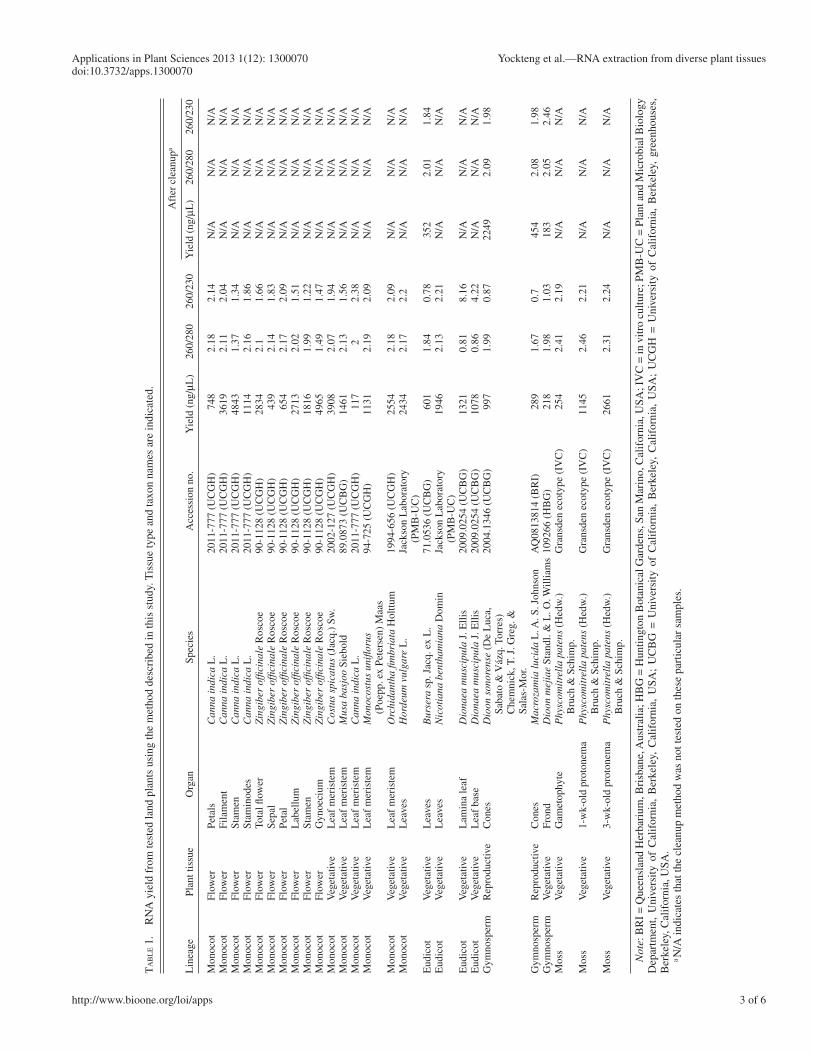
TA
BL
E 1
. R
NA
yie
ld f
rom
test
ed la
nd p
lant
s us
ing
the
met
hod
desc
ribe
d in
this
stu
dy. T
issu
e ty
pe a
nd ta
xon
nam
es a
re in
dica
ted.
Aft
er c
lean
up a
Lin
eage
Plan
t tis
sue
Org
anSp
ecie
sA
cces
sion
no.
Yie
ld (
ng/ μ
L)
260/
280
260/
230
Yie
ld (
ng/ μ
L)
260/
280
260/
230
Mon
ocot
Flow
erPe
tals
Can
na in
dica
L.
2011
-777
(U
CG
H)
748
2.18
2.14
N/A
N/A
N/A
Mon
ocot
Flow
erFi
lam
ent
Can
na in
dica
L.
2011
-777
(U
CG
H)
3619
2.11
2.04
N/A
N/A
N/A
Mon
ocot
Flow
erSt
amen
Can
na in
dica
L.
2011
-777
(U
CG
H)
4843
1.37
1.34
N/A
N/A
N/A
Mon
ocot
Flow
erSt
amin
odes
Can
na in
dica
L.
2011
-777
(U
CG
H)
1114
2.16
1.86
N/A
N/A
N/A
Mon
ocot
Flow
erTo
tal fl
ow
er Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)28
342.
11.
66N
/AN
/AN
/AM
onoc
otFl
ower
Sepa
l Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)43
92.
141.
83N
/AN
/AN
/AM
onoc
otFl
ower
Peta
l Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)65
42.
172.
09N
/AN
/AN
/AM
onoc
otFl
ower
Lab
ellu
m Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)27
132.
021.
51N
/AN
/AN
/AM
onoc
otFl
ower
Stam
en Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)18
161.
991.
22N
/AN
/AN
/AM
onoc
otFl
ower
Gyn
oeci
um Zi
ngib
er o
ffi ci
nale
Ros
coe
90-1
128
(UC
GH
)49
651.
491.
47N
/AN
/AN
/AM
onoc
otV
eget
ativ
eL
eaf
mer
iste
m C
ostu
s sp
icat
us (
Jacq
.) S
w.
2002
-127
(U
CG
H)
3908
2.07
1.94
N/A
N/A
N/A
Mon
ocot
Veg
etat
ive
Lea
f m
eris
tem
Mus
a ba
sjoo
Sie
bold
89.0
873
(UC
BG
)14
612.
131.
56N
/AN
/AN
/AM
onoc
otV
eget
ativ
eL
eaf
mer
iste
m C
anna
indi
ca L
.20
11-7
77 (
UC
GH
)11
72
2.38
N/A
N/A
N/A
Mon
ocot
Veg
etat
ive
Lea
f m
eris
tem
Mon
ocos
tus
unifl
orus
(P
oepp
. ex
Pete
rsen
) M
aas
94-7
25 (
UC
GH
)11
312.
192.
09N
/AN
/AN
/A
Mon
ocot
Veg
etat
ive
Lea
f m
eris
tem
Orc
hida
ntha
fi m
bria
ta H
olttu
m19
94-6
56 (
UC
GH
)25
542.
182.
09N
/AN
/AN
/AM
onoc
otV
eget
ativ
eL
eave
s H
orde
um v
ulga
re L
.Ja
ckso
n L
abor
ator
y (P
MB
-UC
)24
342.
172.
2N
/AN
/AN
/A
Eud
icot
Veg
etat
ive
Lea
ves
Bur
sera
sp.
Jac
q. e
x L
.71
.053
6 (U
CB
G)
601
1.84
0.78
352
2.01
1.84
Eud
icot
Veg
etat
ive
Lea
ves
Nic
otia
na b
enth
amia
na D
omin
Jack
son
Lab
orat
ory
(PM
B-U
C)
1946
2.13
2.21
N/A
N/A
N/A
Eud
icot
Veg
etat
ive
Lam
ina
leaf
Dio
naea
mus
cipu
la J
. Elli
s20
09.0
254
(UC
BG
)13
210.
818.
16N
/AN
/AN
/AE
udic
otV
eget
ativ
eL
eaf
base
Dio
naea
mus
cipu
la J
. Elli
s20
09.0
254
(UC
BG
)10
780.
864.
22N
/AN
/AN
/AG
ymno
sper
mR
epro
duct
ive
Con
es D
ioon
son
oren
se (
De
Luc
a,
Saba
to &
Váz
q. T
orre
s)
Che
mni
ck, T
. J. G
reg.
&
Sala
s-M
or.
2004
.134
6 (U
CB
G)
997
1.99
0.87
2249
2.09
1.98
Gym
nosp
erm
Rep
rodu
ctiv
eC
ones
Mac
roza
mia
luci
da L
. A. S
. Joh
nson
AQ
0813
814
(BR
I)28
91.
670.
745
42.
081.
98G
ymno
sper
mV
eget
ativ
eFr
ond
Dio
on m
ejia
e St
andl
. & L
. O. W
illia
ms
1092
66 (
HB
G)
218
1.98
1.03
183
2.05
2.46
Mos
sV
eget
ativ
eG
amet
ophy
te P
hysc
omit
rell
a pa
tens
(H
edw
.)
Bru
ch &
Sch
imp.
Gra
nsde
n ec
otyp
e (I
VC
)25
42.
412.
19N
/AN
/AN
/A
Mos
sV
eget
ativ
e1-
wk-
old
prot
onem
a P
hysc
omit
rell
a pa
tens
(H
edw
.)
Bru
ch &
Sch
imp.
Gra
nsde
n ec
otyp
e (I
VC
)11
452.
462.
21N
/AN
/AN
/A
Mos
sV
eget
ativ
e3-
wk-
old
prot
onem
a P
hysc
omit
rell
a pa
tens
(H
edw
.)
Bru
ch &
Sch
imp.
Gra
nsde
n ec
otyp
e (I
VC
)26
612.
312.
24N
/AN
/AN
/A
Not
e : B
RI
= Q
ueen
slan
d H
erba
rium
, Bri
sban
e, A
ustr
alia
; HB
G =
Hun
tingt
on B
otan
ical
Gar
dens
, San
Mar
ino,
Cal
ifor
nia,
USA
; IV
C =
in v
itro
cultu
re; P
MB
-UC
= P
lant
and
Mic
robi
al B
iolo
gy
Dep
artm
ent,
Uni
vers
ity o
f C
alif
orni
a, B
erke
ley,
Cal
ifor
nia,
USA
; U
CB
G =
Uni
vers
ity o
f C
alif
orni
a, B
erke
ley,
Cal
ifor
nia,
USA
; U
CG
H =
Uni
vers
ity o
f C
alif
orni
a, B
erke
ley,
gre
enho
uses
, B
erke
ley,
Cal
ifor
nia,
USA
. a N
/A in
dica
tes
that
the
clea
nup
met
hod
was
not
test
ed o
n th
ese
part
icul
ar s
ampl
es.
4 of 6
Applications in Plant Sciences 2013 1 ( 12 ): 1300070 Yockteng et al.—RNA extraction from diverse plant tissues doi:10.3732/apps.1300070
http://www.bioone.org/loi/apps
exon capture approach to identify markers for population ge-netic and phylogenetic analyses (e.g., Bi et al., 2012 ).
Quality and quantity — Experiments using a PCR-based ap-proach to sequence fragments of cDNA generated from RNA extractions typically only require checking the concentration of RNA via NanoDrop (or similar equipment) and running an ali-quot of the RNA on an agarose gel to verify the appearance of ribosomal RNA bands indicating a lack of degradation.
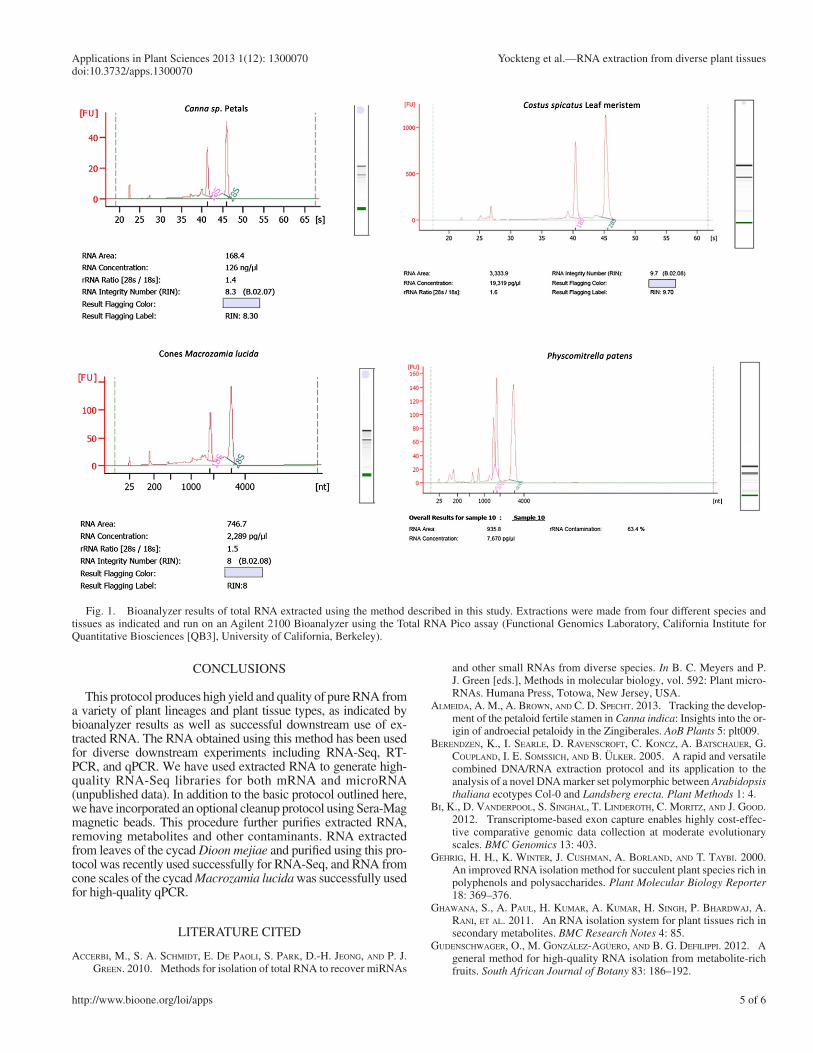
In RNA-Seq or ChIP-Seq experiments, both the quality and the quantity of the RNA are very important. We have found that it is necessary to start cDNA synthesis and subsequent sequenc-ing library preparation with at least 10 μ g of pure RNA that is determined to have good integrity based on the entire electropho-retic trace. An Agilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany) calculates the “RNA integrity number” (RIN) using all of the information present in the electrophoretic trace. In general, RNA with a value above 7 is required to pro-duce good results in a next-generation sequencing analysis.
Bioanalyzer results are shown for four samples extracted with our detailed method ( Fig. 1 ) . The two peaks of 18S and 28S ribosomal RNA appear as indicated, and the trace in the background corresponds to other types of RNA with less con-centration; i.e., the mRNA. All RIN values are above 8, indicat-ing no degradation of the RNA. These traces represent typical results obtained from our method, indicating high concentra-tion and good quality.
The purity of extracted RNA can depend on the amount of secondary metabolites produced by the plants being studied. Our extraction method consistently produced good yield for a diversity of plants; however, the recovered RNA was some-times not pure enough for subsequent cDNA synthesis and se-quencing library preparation. This was particularly true in plants such as Dionaea muscipula J. Ellis (Venus fl ytrap) where the presence of complex proteins affected the 260/280 ratio. In the case of Bursera sp. (Burseraceae) and in cone scales of cycads ( Dioon sonorense (De Luca, Sabato & Vázq. Torres) Chemnick, T. J. Greg. & Salas-Mor. and Macrozamia lucida ), the ratio 260/230 was affected by secondary metabolites and highly lignifi ed tissue, respectively, and the values fell below 1, indicating the presence of carbohydrates.
In these plants we tried other methods of RNA extraction such as the RNAqueous (Ambion) protocol, based on silica col-umns that are reported to isolate pure RNA, and the TRIzol Reagent (Ambion), which is reported to be effective with a broad range of samples. Neither of these methods were able to produce a good yield (e.g., RNA recovered using RNAqueous was only 8.6 ng/ μ L from fresh, frozen, or RNA later -pre-served leaves of Bursera spp.). Another recommendation to increase RNA purity commonly found across various protocols is to conduct two chloroform washes. After many attempts, we con-clude that adding a chloroform step in our protocol signifi cantly reduces the RNA yield without resulting in any improvement in RNA quality.
For recalcitrant samples with lignin and/or secondary me-tabolites, we propose several modifi cations to a method that has been previous published for cleaning DNA ( Rohland and Reich, 2012 ). This low-cost method uses carboxyl-modifi ed beads that have a high binding capacity for nucleic acids, per-mitting additional washes without signifi cant loss and enabling the isolation of RNA from other components obtaining high yield and pure extraction product.
Scientifi c, Waltham, Massachusetts, USA; cat. no. 09-981-123) to a 1.5-mL tube. Pellet the beads for 5 min with a magnetic stand designed for 1.5–2-mL tubes. Remove the storage buffer, leaving the tubes in the magnetic stand. Wash beads 1 × with 1 mL TE or water. Remove the tube from the magnetic stand and immediately resuspend the beads in an additional 1 mL TE or water.
Add the bead suspension to the prepared 50-mL Falcon tube and wrap the Falcon tube with aluminum foil. Store at 4 ° C for further use. The fi nal concen-tration of the bead solution is 0.1% carboxyl-modifi ed Sera-Mag Magnetic SpeedBeads, 18% PEG-8000 (w/v), 1 M NaCl, 10 mM Tris-HCl (pH 8.0), 1 mM EDTA (pH 8.0), and 0.05% Tween 20.
RNA cleanup— Bring a measured amount of prepared Sera-Mag beads solution to room temperature at least 30 min prior to use. For maximum binding, measure out 3 × the total RNA volume of the beads solution and place into a 1.5-mL tube along with the RNA. Place tubes on a rotator at room temperature for 10 min.
Spin the beads solution down before the beads dry permanently onto the wall of the tube. The beads must be spun down very quickly and at low speed to avoid bead precipitation; a mini benchtop centrifuge is recommended, with a spin so quick such that the lid does not need to be closed. Incubate without ro-tating at room temperature for an additional 10 min to maximize RNA binding. Place the 1.5-mL tubes in the magnetic stand for 5 min, then remove the buffer with a pipette and wash the beads 2 × with 500 μ L of fresh 80% ethanol. After the second wash, remove all ethanol and make sure that no ethanol is left in the samples. Remove the tubes from the magnetic stand and spin beads down from the sides of the wall before they dry. Air-dry the bead pellet for 10 min. Elute with RNase-free water using approximately the same volume as your original sample of RNA.
Vortex beads and water just to mix, and spin down quickly at low speed as before. Place tubes on a rotator at room temperature for 2–5 min for maximum RNA elution, and spin down quickly. Place tubes in the magnetic stand. Let stand for 5 min and recover eluted RNA in a separate tube.
A NanoDrop measure is recommended to check quality and yield of RNA.
DISCUSSION
Diversity of samples and tissue types — High-yield and pure RNA was obtained for different plant lineages ( Table 1 ), in-cluding mosses, gymnosperms, and various groups across angiosperms. Successful cDNA library construction was com-pleted from various plant tissue types, including individual fl o-ral and reproductive organs as well as combined reproductive and vegetative tissues ( Table 1 ).
In general, high yield was obtained even when the starting material measured less than 0.1 g. RNA obtained was used in several downstream experiments such as cDNA synthesis for RT-PCR and qPCR ( Yockteng et al., 2013 ; Almeida et al., 2013 ; Almeida et al., unpublished), and for generating tran-scriptome libraries and high-quality RNA-Seq data using TruSeq RNA sample preparation kits (Illumina, San Diego, California, USA).
RNA extracted from specifi c fl oral organs from Costus spi-catus (Jacq.) Sw., Musa basjoo Siebold, and Canna indica L. ( Table 1 ) was labeled with TruSeq RNA sample preparation kits to generate and sequence RNA-Seq libraries, which yielded high-quality sequence data that contributed to analyses of the gene expression differences underlying evolution of flower morphology across the Zingiberales ( Yockteng et al., 2013 ; Almeida et al., unpublished).
RNA extracted from leaf material and cone scales of cycad species Dioon mejiae Standl. & L. O. Williams and Macroza-mia lucida L. A. S. Johnson ( Table 1 ) was cleaned with the optional cleanup method presented here. RNA-Seq libraries were successfully sequenced and transcriptome assemblies are being used to design gene targets for a transcriptome-based
Applications in Plant Sciences 2013 1 ( 12 ): 1300070 Yockteng et al.—RNA extraction from diverse plant tissues doi:10.3732/apps.1300070
5 of 6http://www.bioone.org/loi/apps
and other small RNAs from diverse species. In B. C. Meyers and P. J. Green [eds.], Methods in molecular biology, vol. 592: Plant micro-RNAs. Humana Press, Totowa, New Jersey, USA.
ALMEIDA , A. M. , A. BROWN , AND C. D. SPECHT . 2013 . Tracking the develop-ment of the petaloid fertile stamen in Canna indica : Insights into the or-igin of androecial petaloidy in the Zingiberales . AoB Plants 5 : plt009 .
BERENDZEN , K. , I. SEARLE , D. RAVENSCROFT , C. KONCZ , A. BATSCHAUER , G. COUPLAND , I. E. SOMSSICH , AND B. ÜLKER . 2005 . A rapid and versatile combined DNA/RNA extraction protocol and its application to the analysis of a novel DNA marker set polymorphic between Arabidopsis thaliana ecotypes Col-0 and Landsberg erecta. Plant Methods 1 : 4 .
BI , K. , D. VANDERPOOL , S. SINGHAL , T. LINDEROTH , C. MORITZ , AND J. GOOD . 2012 . Transcriptome-based exon capture enables highly cost-effec-tive comparative genomic data collection at moderate evolutionary scales. BMC Genomics 13 : 403 .
GEHRIG , H. H. , K. WINTER , J. CUSHMAN , A. BORLAND , AND T. TAYBI . 2000 . An improved RNA isolation method for succulent plant species rich in polyphenols and polysaccharides. Plant Molecular Biology Reporter 18 : 369 – 376 .
GHAWANA , S. , A. PAUL , H. KUMAR , A. KUMAR , H. SINGH , P. BHARDWAJ , A. RANI , ET AL . 2011 . An RNA isolation system for plant tissues rich in secondary metabolites. BMC Research Notes 4 : 85 .
GUDENSCHWAGER , O. , M. GONZÁLEZ-AGÜERO , AND B. G. DEFILIPPI . 2012 . A general method for high-quality RNA isolation from metabolite-rich fruits. South African Journal of Botany 83 : 186 – 192 .
CONCLUSIONS
This protocol produces high yield and quality of pure RNA from a variety of plant lineages and plant tissue types, as indicated by bioanalyzer results as well as successful downstream use of ex-tracted RNA. The RNA obtained using this method has been used for diverse downstream experiments including RNA-Seq, RT-PCR, and qPCR. We have used extracted RNA to generate high-quality RNA-Seq libraries for both mRNA and microRNA (unpublished data). In addition to the basic protocol outlined here, we have incorporated an optional cleanup protocol using Sera-Mag magnetic beads. This procedure further purifi es extracted RNA, removing metabolites and other contaminants. RNA extracted from leaves of the cycad Dioon mejiae and purifi ed using this pro-tocol was recently used successfully for RNA-Seq, and RNA from cone scales of the cycad Macrozamia lucida was successfully used for high-quality qPCR.
LITERATURE CITED
ACCERBI , M. , S. A. SCHMIDT , E. DE PAOLI , S. PARK , D.-H. JEONG , AND P. J. GREEN . 2010 . Methods for isolation of total RNA to recover miRNAs
Fig. 1. Bioanalyzer results of total RNA extracted using the method described in this study. Extractions were made from four different species and tissues as indicated and run on an Agilent 2100 Bioanalyzer using the Total RNA Pico assay (Functional Genomics Laboratory, California Institute for Quantitative Biosciences [QB3], University of California, Berkeley).
6 of 6
Applications in Plant Sciences 2013 1 ( 12 ): 1300070 Yockteng et al.—RNA extraction from diverse plant tissues doi:10.3732/apps.1300070
http://www.bioone.org/loi/apps
JAPELAGHI , R. H. , R. HADDAD , AND G.-A. GAROOSI . 2011 . Rapid and ef-fi cient isolation of high quality nucleic acids from plant tissues rich in polyphenols and polysaccharides. Molecular Biotechnology 49 : 129 – 137 .
KUMAR , A. , AND K. SINGH . 2012 . Isolation of high quality RNA from Phyllanthus emblica and its evaluation by downstream applications. Molecular Biotechnology 52 : 269 – 275 .
MACRAE , E. 2007 . Extraction of plant RNA. In E. Hilario and J. Mackay [eds.], Methods in molecular biology, vol. 353: Protocols for nucleic acid analysis by nonradioactive probes, 2nd ed. Humana Press, Totowa, New Jersey, USA.
MENG , L. , AND L. FELDMAN . 2010 . A rapid TRIzol-based two-step method for DNA-free RNA extraction from Arabidopsis siliques and dry seeds. Biotechnology Journal 5 : 183 – 186 .
QI , G. , J. T. LI , Q. P. RUAN , J. YANG , AND Z. X. SU . 2009 . An optimized, small-scale preparation of high-quality RNA from dry seeds of Davidia involucrata. Phytochemical Analysis 20 : 139 – 142 .
ROHLAND , N. , AND D. REICH . 2012 . Cost-effective, high-throughput DNA sequencing libraries for multiplexed target capture. Genome Research 22 : 939 – 946 .
YIN , D. , H. LIU , X. ZHANG , AND D. CUI . 2011 . A protocol for extraction of high-quality RNA and DNA from peanut plant tissues. Molecular Biotechnology 49 : 187 – 191 .
YOCKTENG , R. , A. M. ALMEIDA , K. MORIOKA , E. R. ALVAREZ-BUYLLA , AND C. D. SPECHT . 2013 . Molecular evolution and patterns of duplication in the SEP/AGL6 -like lineage of the Zingiberales: A proposed mecha-nism for fl oral diversifi cation. Molecular Biology and Evolution 30 : 2401 – 2422 .