This article was downloaded by: [Universite Laval] On: 04 July 2014, At: 14:10 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Vertebrate Paleontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ujvp20 A Middle Jurassic Sphenodon-like sphenodontian (Diapsida: Lepidosauria) from Huizachal Canyon, Tamaulipas, Mexico Victor-Hugo Reynoso a a Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria , Mexico, D.F., 04510 Published online: 24 Aug 2010. To cite this article: Victor-Hugo Reynoso (1996) A Middle Jurassic Sphenodon-like sphenodontian (Diapsida: Lepidosauria) from Huizachal Canyon, Tamaulipas, Mexico, Journal of Vertebrate Paleontology, 16:2, 210-221, DOI: 10.1080/02724634.1996.10011309 To link to this article: http://dx.doi.org/10.1080/02724634.1996.10011309 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
Transcript
This article was downloaded by: [Universite Laval]On: 04 July 2014, At: 14:10Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Vertebrate PaleontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ujvp20
A Middle Jurassic Sphenodon-like sphenodontian(Diapsida: Lepidosauria) from Huizachal Canyon,Tamaulipas, MexicoVictor-Hugo Reynoso aa Instituto de Geología, Universidad Nacional Autónoma de México, CiudadUniversitaria , Mexico, D.F., 04510Published online: 24 Aug 2010.
To cite this article: Victor-Hugo Reynoso (1996) A Middle Jurassic Sphenodon-like sphenodontian (Diapsida:Lepidosauria) from Huizachal Canyon, Tamaulipas, Mexico, Journal of Vertebrate Paleontology, 16:2, 210-221, DOI:10.1080/02724634.1996.10011309
To link to this article: http://dx.doi.org/10.1080/02724634.1996.10011309
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
VICTOR-HUGO REYNOSOInstituto de Geologia, Universidad Nacional Aut6noma de Mexico, Ciudad Universitaria, Mexico D.E 04510.
ABSTRACT-A new Middle Jurassic North American sphenodontian, Cynosphenodon huizachalensis, gen . et sp. nov.is described on the basis of several dentaries and maxillae. It is characterized by its small size and its striking similarityto Sphenodon. The presence of a deep posteromedial wear facet on the caniniform tooth, a small denticle in theadditional tooth series, and an anterior small groove dorsal and convergent to the Meckelian Canal, are unique characteristics of this species. In adult individuals the caniniform tooth placed posteriorly to an edentulous ridge, and thepresence of wear facets on teeth and jaws caused by propalinal jaw action, suggest that C. huiza chalensis is closelyrelated to Sphenodon; the wear facet on the posteromedial face of the caniniform tooth, however, may suggest a differentarrangement of the anterior palatal region.
RESUMEN-Con base en varios dentarios y rnaxilas, se describe Cynosphenodon huizachalensis, gen. et sp . nov. unnuevo esfenodonte del Jurasico Medio de Norte America. Se caracteriza par sus pequefias dimensiones y su gransimilitud a Sphenodon. La presencia de una faceta de desgaste en la parte posteromedia del diente caniniforme, de undentfculo en la serie dental adicional, y de un pequefio surco dorsal y convergente al Canal de Meckel, son caracteresunicos de esta especie. El diente caniniforme ubicado posteriormente a un margen edentado en adultos, y la presenciade facetas de desgaste en dientes y mandfbulas causados por movimientos mandibulares propalfneos, sugieren que C.huizachalensis esta cercanamente relacionado a Sphenodon ; la faceta de desgaste en la cara posteromedia del dientecaniniforme, sin embargo, sugiere un arreglo diferente de la region palatal anterior.
INTRODUCTION
Beginning in 1985, preliminary descriptions of a Middle Jurassic North American terrestrial vertebrate assemblage werepublished (Clark and Hopson, 1985; Fastovsky et aI., 1987;Clark et aI., 1991 ; Clark and Hernandez, 1994) . Close to onehundred sphenodontian specimens have been collected fromthis assemblage, making this unquestionably the most abundantgroup in the fauna. Much more material remains to be prepared.
The presence of sphenodontians in the Huizachal Canyonfauna was reported in 1987 (Fastovsky et aI., 1987 ; see alsoFraser, 1988, Clark et aI., 1991), but, no detailed descriptionhas yet been published. Reynoso (1992) considered the sphenodontian collection to include at least three different taxa ofsphenodontians. The best known and most abundant taxon isdescribed below. Descriptions of two other taxa will be published elsewhere.
Huizachal Canyon is located 15 km south west of CiudadVictoria, in the state of Tamaulipas, Mexico (Fig. I) . Thesphenodontian remains occur at different localities in the lower part of the La Boca Formation red beds. The fossils arevery small in size, green, and soft. Though some are crystallized or fragmented , most of them are very well preserved.They are enclosed in a very hard, silicified, hematitic mudstone, and preparation is a slow mechanical procedure usingneedles .
Sphenodontians were a highly diverse group in the Late Triassic to Late Jurassic of Europe and China. Almost 30 differentspecies have been described to date (Huene, 1952 ; CocudeMichel, 1963 ; Fabre, 1973, 1974; Evans, 1980 ; Fraser, 1982,1986 , 1988; Carroll, 1985; Fraser and Benton, 1989 ; Whiteside,1986 ; Wu, 1994) . The reported diversity of this group in NorthAmerica, however, is much lower, and fewer species have beendescribed: an unnamed genus from the Late Triassic of Con-
* Current address : Redpath Museum, McGill University, 859 Sherbrooke 51. West Montreal, Quebec, Canada H3A 2K6.
necticut (Sues and Baird, 1993); the Early Jurassic Clevosaurusbairdi (Sue s et aI., 1994); the Late Jurassic Opisthias rarus(Gilmore, 1909), Theretairus antiquus (Simpson, 1926), and Eilenodon robustus (Rasmussen and Callison, 1981); and the Early Cretaceous Toxolophosaurus cloudi (Throckmorton et aI.,1981). Realizing GUnther's (1867) prediction, many fossil species have proved to be closely related to Sphenodon, so theworld wide abundance of the Rhynchocephalia (sensu Gauthieret aI., 1988) makes this taxon very well known. Today onlytwo species of the genus Sphenodon survive (Daugherty et aI.,1990) .
Abbreviations-IGM: Instituto de Geologia, Ciudad Universitaria, Mexico; KU , Kansas University; RM Redpath Museum, McGill University.
SYSTEMATIC PALEONTOLOGY
LEPIDOSAURIA Dumeril et Bibron, 1839SPHENODONTIA Williston, 1925SPHENODONTIDAE Cope, 1869
SPHENODONTINAE Nopcsa, 1928
Diagnosis-Sphenodontians bearing caniniform dentitionposterior to an edentulous ridge at the anterior end of the dentary. Sphenodon, Theretairus (Simpson, 1926), and an unnamedsphenodontian from Kirtlington, Oxfordshire, England (Evans,1992) are included. Here the name Sphenodontinae is restrictedto Sphenodon and its closest relatives. Clevosaurus and Opisthias (see Fraser, 1986) are excluded.
Genus CYNOSPHENODON. gen. nov.
Type Species-C. huizachalensisEtymology-From the Greek cyno (=dog; cynodon = fang),
and Sphenodon. In reference to its anterior caniniform tooth andto its similar appearance (and close relationship) to Sphenodon.
Diagnosis-As for the type species.
210
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
REYNOSO-SPHENODON-LIKE SPHENODONTIAN 211
A
101
Huizachal Canyon
_101_
IN
ToCd. Mant.
5km'-- ....I.
La Boca OutcropsHuizBchal canyon
Federal tf'dlTNay
DirtRoad
Period Series FonnationI I I A
CII I I I A-ltl ZULOAGA I I IU
..J !I I I II / I /.....
U)LA JOVA Co
~l..
U) CII 0 ~
"C... )
"C o ,cts .- ... \:E - ' 0 -ca I
"""s: - ,,() . . .
LA BOCA ca .:::I \ , I I
N.- , . 0 •~
,\
"":) :I: ' . 1 •-- ------, , . , . ,FOSSILIFEROUS
, BEDS , .' 'I, ,IIIIII I, I I , I I
?lilli/I,);'1'1'1' , 1',/1• ",I'I;?/11/1, ,
.A ;' I';' ~:"/,I/
" V"B
FIGURE 1. A, Locatio n of Hu izachal Canyon, and B, stratigra ph ic rela tions hips of La Boca Forma tion outcrops, showing the level wheresphe nodo ntian fossi ls were co llected.
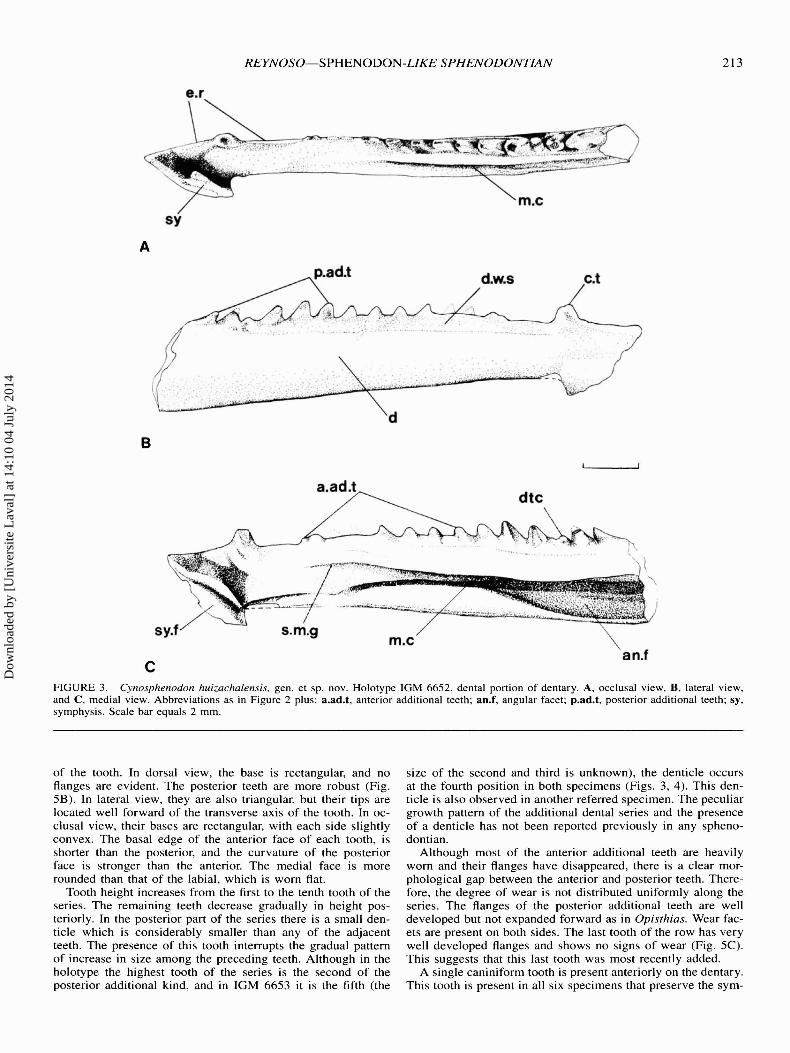
Species C YN OSPHEN ODON HUIZACHALENSIS. sp. nov.Fig. 2
Holotype-Cat. No. 10M 665 2. Dental port ion of the rightdent ary with the co mp lete tooth row.
Hypodigm-IOM 6653. poster ior right dentary; 10M 6654,anterior portion of a right maxill a; 10M 6655, midd le portio nof a right maxill a; 10M 6656. anter ior part of a right dentary ;10M 6657, anterio r port ion of a rig ht dentary; 10M 6658 symphysial region of a right dentary; 10M 6659; anterior part of aleft dentary; 10M 6660. posterior part of a left dentary withcoronoid process.
Referred Material-I 6 incomplete lower j aws.Distribution-Lower part of La Boca Formation ; Hui zachal
Canyon , Municipio de Ciudad Victori a, Tamaulipas, Mex ico.Age-Middle Jurassic (Fastov sky et al., 1995) .Etymology-From huizachal, (= place of mesqu ites) na
huat l. In reference to the name of the canyon where the taxonwas found.
Diagnosis-The lower jaw is simila r to that of Sphenodonbut much smalle r (Fig. 2). Un ique characters are : a sma llgroove dorsal but convergent to the Meckeli an Canal on theanterom ed ial surface of the dentar y; more posterior additio nalteeth become pro gressively sma ller than the middle ones andabo ut the same size as the anterior teeth ; a sma ll denticle intercalated within the posterior part of the chee k tooth series ;and a deep posteromedial wear face t on the ca niniform too thprob ably made by a large vomerine tooth or by the first toothof an anteri orly enlarged palat al tooth series.
DESCRIPTION
Dentary-Th e dentary is smaller and more slender than thatof Sphenodo n. On the basis of the holotype and IGM 6660 thelength of the dentary is es timated to have been 23 mm . Nospeci me n in the hypod igm is larger than the holo type. Me asuredbeneath the coronoid process the depth of the dentary fro m theventra l edge of the jaw to the base of the teeth ranges from 3.1mm to 1.1 mm in young indivi dua ls. In the holotype the lengthof the dental ma rgin anterior to the coronoid process is 14.9mm (tw o-th irds of the total length of the dentary). The depthof the jaw attai ns its maximum below the tenth tooth and isapproximate ly one- fifth of the total dentary length. Th e heightof the dentar y decreases anter iorly, reach ing its minimum sizej ust before the symphysial region (before the canin iform tooth ).At this po int it is almost half its maximum height (Fig. 3) .
Dor sally , the coronoid process of the dentary is about onesix th of the dentary length. The process is well developed andanteriorly is almost perpendicul ar to the dental series . In theposterior marg in an invag inatio n for the mandibular foramen ispre sent. Medi ally , the angular, articular, coro noid, pre articul ar,sura ngular, and dentary facets are present (Fig. 4). Apparentl ythe suture between both dentaries was not ossified as in othersphenodo ntians, so a ligamentou s connec tio n was probablypresent. Th e dorsal edge of the dentary is straight, but , posteri orto the corono id pro cess, it run s slightly oblique in relation tothe dental edge . The posteri or end of the dentar y tapers posteri orly as in Sphenodon . The artic ular, prearticular, and surangular are unknown.
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
212 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 16, NO .2, 1996
me .f
, ' .•\ .. ' :.;" . ~ : : I ,.. : '. : . '
p.pr.d
,~6'/'
//;:::==~--" --"'/ ..\ \ ,......'.
A
~~""'---7--~=~~~-------pa B
FIGURE 2. Cynosphenodon huizachalensis, gen . et sp . nov . Reconstruction of the lower jaw and maxilla. A, lateral view; 8, medial view . ad.t,additional teeth ; ad.fos, adductor fossa; an, angular; c.t, caniniform tooth; d , dentary bone; dtc, denticle; e.n, external nares; e.r, edentulous ridge;c.pr, coronoid process; me.f, mental foramina; h.t , hatchling teeth ; m , maxilla; m.c, Meckelian Canal ; 0, orbit; pa, prearticular; pm, premaxilla;p.pr.d, posterior process of dcntary ; sa , surangular; s.m.g, secondary medial groove; sy.f, symphysial facet. Premaxilla based upon materialtentatively referred to this species. Scale bar equals 5 mm .
In lateral view, the ventral border of the dentary is nearlystraight (Fig. 3), but beneath the coronoid process it curvesgently upward and extends straight to the posterior end of thedentary. Close to the mandibular symphysis, the ventral edgeof the dentary projects strongly downward. The dorsal marginof the jaw projects further forward than the ventral margin.
In dorsal view, the dentary is straight except where its anterior part turns medially at the symphysis (Fig. 3A). The occlusal margin bears an edentulous ridge in the symphysial region followed by the caniniform tooth. Between the caniniformand the cheek tooth series another edentulous ridge is aboutone-tenth the dental margin length. Immediately before the posterior edentulous ridge begins, the dorsal edge of the dentarybecomes very thin . Both edentulous ridges functioned as trituration surfaces and are heavily worn. In the smaller specimens,hatchling dentitions are present in these areas. Posterior to theedentulous ridge, the tooth row extends posteriorly to about thecoronoid process.
The external surface of the dentary is smooth (Fig. 3B) andfour to five mental foramina aligned close to the anterior endare present on 10M 6657 and 10M 6658, but are not evidentin the holotype. A slight ridge extends from below the posteriorpart of the caniniform tooth to slightly forward of the coronoidprocess . The surface above that flange is well polished by toothon-bone wear; no scorings or bite marks are evident.
On the medial surface, below the dental series, a slight ridgeand a polished surface similar to, but less conspicuous than, thelateral surface is observed. Posterior to them, the jaw bearsthree small foramina. Two are placed one below the other andthe third is slightly forward. All the sutural facets are clearlypreserved. The symphysial facet is suboval and bears a smallgroove (Fig. 3C). The angular facet is extensive and indicates
that the angular bone covered at least one-third of the dentarysurface below the tooth row. It approaches the ventral edge butis not exposed on the lateral side of the jaw (Fig. 4B). Thecoronoid and prearticular facets are above and posterior to theMeckelian Canal, respectively; and there is a facet for the surangular at the posterior end of the coronoid process. The articular facet is evident at the tip of the posterior process of thedentary.
The Meckelian Canal , as in all sphenodontians, is open medially and extends along the midline of the jaw. In Cynosphenodon, however, the anterior end of the canal angles downwardsrunning close to the ventral edge of the dentary. Anteriorly, itreaches the symphysis and is continuous with a small grooveover the symphysial facet. Below the fifth tooth, the canal doubles in diameter due to the fusion of a second smaller grooveplaced dorsally and convergent to the anterior part of the Meekelian Canal. Posteriorly, the canal becomes narrow again untilit reaches the adductor fossa. The transition between the Meekelian Canal and the adductor fossa was presumably covered bythe coronoid and articular, as in Sphenodon.
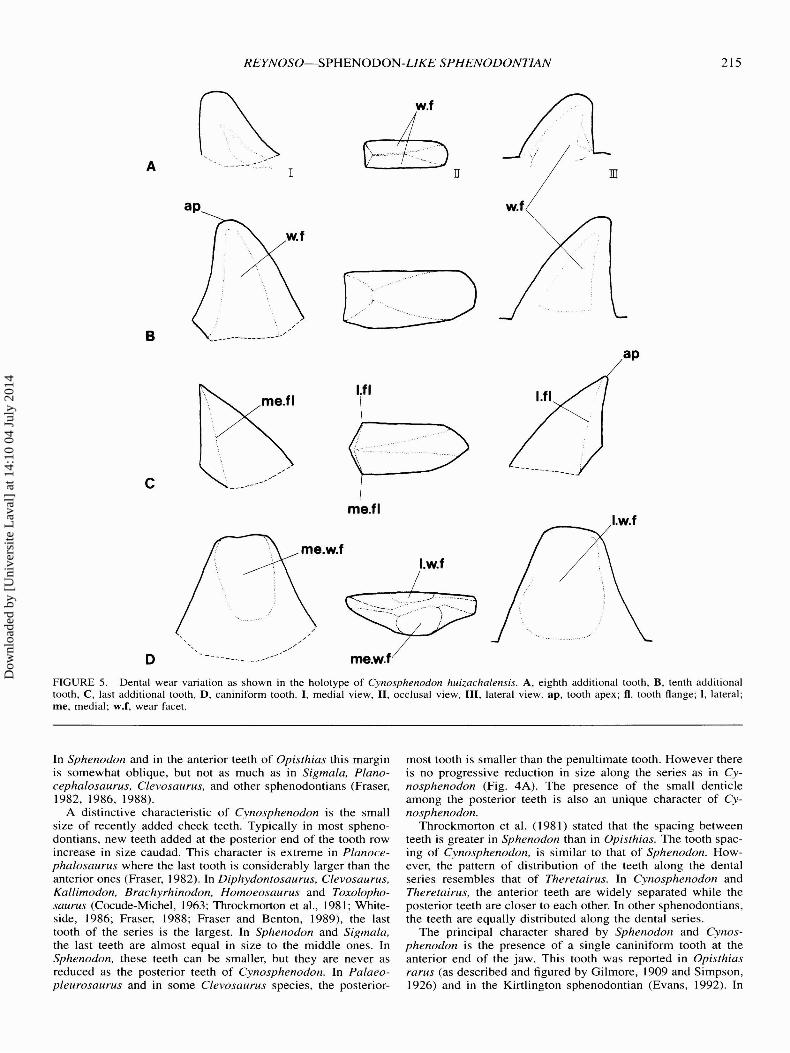
Dentition-The teeth are acrodont and subconical. One successional tooth and an additional tooth series (Robinson, 1975)can be identified. The additional teeth are spaced similar tothose of Sphenodon, but, the spacing is greater in the anteriorteeth. The holotype is the only specimen with a complete toothrow, bearing 14 teeth; however, the maximum possible numberof additional teeth remains unknown. In the holotype the firstand second teeth are almost worn away. The third and fourthteeth are broken (Fig. 3) .
There are two different kinds of teeth in the additional series.The anterior ones are narrower transversely and triangular inlateral view (Fig. 5A). Their tips are close to the transverse axis
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
e.r
REYNOSO-SPHENODON-LIKE SPHENODONTIAN 213
sy
A
B
c
s.m.gm.c
d.w.s C.t
an.f
FIGURE 3. Cynosphenodon huizachalensis, gen. et sp. nov. Holotype IGM 6652. dental portion of dentary. A. occlusal view. B. lateral view.and C, medial view. Abbreviations as in Figure 2 plus : a.ad.t, anterior additional teeth; an.f, angular facet ; p.ad.t, posterior additional teeth; sy,symphysis. Scale bar equals 2 mm .
of the tooth. In dorsal view, the base is rectangular, and noflanges are evident. The posterior teeth are more robust (Fig .5B) . In lateral view, they are also triangular, but their tips arelocated well forward of the transverse axis of the tooth. In occlusal view, their bases are rectangular, with each side slightlyconvex. The basal edge of the anterior face of each tooth, isshorter than the posterior, and the curvature of the posteriorface is stronger than the anterior. The medial face is morerounded than that of the labial, which is worn flat.
Tooth height increases from the first to the tenth tooth of theseries. The remaining teeth decrease gradually in height posteriorly. In the posterior part of the series there is a small denticle which is considerably smaller than any of the adjacentteeth. The presence of this tooth interrupts the gradual patternof increase in size among the preceding teeth. Although in theholotype the highest tooth of the series is the second of theposterior additional kind, and in 10M 6653 it is the fifth (the
size of the second and third is unknown), the denticle occursat the fourth position in both specimens (Figs. 3, 4). This denticle is also observed in another referred specimen. The peculiargrowth pattern of the additional dental series and the presenceof a denticle has not been reported previously in any sphenodontian.
Although most of the anterior additional teeth are heavilyworn and their flanges have disappeared, there is a clear morphological gap between the anterior and posterior teeth. Therefore, the degree of wear is not distributed uniformly along theseries. The flanges of the posterior additional teeth are welldeveloped but not expanded forward as in Opisthias. Wear facets are present on both sides. The last tooth of the row has verywell developed flanges and shows no signs of wear (Fig . 5C).This suggests that this last tooth was most recently added.
A single caniniform tooth is present anteriorly on the dentary.This tooth is present in all six specimens that preserve the syrn-
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
214
A
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. /6, NO. 2, /996
m.e
Bad.fos
FIGURE 4. Cynosphenodon huizachalensis, gen. et sp . nov. Posterior part of the dentary in medial view . A, IGM 6653, B, IGM 6660. Abbreviations as in Figure 2 plus: an.f, angular facet ; ar.f', articular facet ; c.f', coronoid facet ; ma.f', mandibular foramen ; pa.f' , prearticular facet ; sa.f,surangular facet. Scale bar equals 2 mm .
physis. The caniniform tooth is as tall as the tallest additionaltooth. In lateral view, it is triangular and has a truncated tip dueto wear (Fig. 5D). The lateral face of the tooth is flatter thanthe medial face . A deep wear facet is present on each side . Inlateral view, a vertical groove runs from the base to the tip ofthe tooth, where it fades away. A shallower wear facet is alsoobserved on the medial face , but it is placed posteriorly anddoes not extend to the base of the tooth .
Maxilla-Two gracile maxillae with dentition very similarto that of the dentary are assigned to this taxon (Fig. 6A , B) .These maxillae are quite distinct from other sphenodontianmaxillae in the Huizachal assemblage. Both elements are incomplete, so their overall shape cannot be determined. Only thelateral side is exposed in both elements, and the medial surfaceremains embedded in matrix .
The maxilla is high, but the extent of the anterior and suborbital processes are unknown. A small ridge extends parallelto the dental series, with several foramina above it. On thedorsal process of the maxilla, a small portion of the narial border is preserved anteriorly, and on the posterior end of the dorsal process are remains of the prefrontal facet.
The maxillary dentition is gracile, with little spacing betweenteeth. IGM 6654 represents a mature individual with all additional teeth completely worn away except for two anterior successional teeth . The taller tooth has a caniniform shape. It istriangular with the tip displaced anterior to the mid-transverseaxis. It has a lateral crest slightly anterior to the transverse axis.In the smaller specimen, IGM 6655, some hatchling teeth arepresent posterior to the caniniform tooth. In IGM 6654, however, the jaw margin forms an edentulous trituration surface.
DISCUSSION
The upper and lower jaw elements of Cynosphenodon huizachalensis are similar to those of Sphenodon. It also shares anumber of characters with Opisthias and Theretairus. Althoughthe presence of a propalinal jaw movement is also shared withEilenodon and Toxolophosaurus, these taxa differ considerablyin their laterally expanded teeth and very deep jaw. Cynosphenodon is not directly comparable with European and Asiaticsphenodontians since it lacks a precision bite, has a slenderlower jaw and an anterior caniniform tooth. Therefore the discussion and comparisons primarily focus on Opisthias, Theretairus, and Sphenodon : genera with slender jaws and a propalinal jaw action.
The dentition of Cynosphenodon is similar to that of Sphenodon as described by Robinson (1976) and Throckmorton et al.(1981) . Acrodont triangular teeth with a rounded shape in crosssection are present in both species and shared with most sphenodontians. The short anteroposterior length of the teeth in Cynosphenodon and Sphenodon, however, is not known in anyother sphenodontian. In lateral view, the triangular shape ismore of a straight triangle than an equilateral. In other sphenodontians (e .g., Clevosaurus, Planocephalosaurus. Kallimodon,and Sigmala) the posterior teeth are obtusely triangular and innone of them does the tip of each tooth extend anteriorly beyond its medial transverse axis. In unworn teeth of Opisthias,the anteriorly projected tip is stronger than in Sphenodon; however, it does not project as far as in Cynosphenodon. In Cynosphenodon the anterior face of almost every tooth is orientedperpendicular to the dorsal jaw margin as seen in lateral view.
FIGURE 5. Dental wear variation as shown in the holotype of Cynosphenodon huizachalensis. A, eighth additional tooth, B, tenth additionaltooth , C , last additional tooth, D, caniniform tooth. I , medial view, II , occlusal view, III, lateral view . ap, tooth apex ; fl. tooth flange; I, lateral;me, medial; w.f, wear facet.
In Sphenodon and in the anterior teeth of Opisthias this marginis somewhat oblique, but not as much as in Sigmala, Planocephalosaurus, Clevosaurus, and other sphenodontians (Fraser,1982, 1986, 1988) .
A distinctive characteristic of Cynosphenodon is the smallsize of recently added cheek teeth. Typically in most sphenodontians, new teeth added at the posterior end of the tooth rowincrease in size caudad. This character is extreme in Planocephalosaurus where the last tooth is considerably larger than theanterior ones (Fraser, 1982). In Diphydontosaurus, Clevosaurus,Kallimodon, Brachyrhinodon, Homoeosaurus and Toxolophosaurus (Cocude-Michel , 1963; Throckmorton et al., 1981; Whiteside , 1986 ; Fraser, 1988; Fraser and Benton, 1989), the lasttooth of the series is the largest. In Sphenodon and Sigmala,the last teeth are almost equal in size to the middle ones. InSphenodon, these teeth can be smaller, but they are never asreduced as the posterior teeth of Cynosphenodon. In Palaeopleurosaurus and in some Clevosaurus species, the posterior-
most tooth is smaller than the penultimate tooth. However thereis no progressive reduction in size along the series as in Cynosphenodon (Fig. 4A). The presence of the small denticleamong the posterior teeth is also an unique character of Cynosphenodon.
Throckmorton et al. (1981) stated that the spacing betweenteeth is greater in Sphenodon than in Opisthias. The tooth spacing of Cynosphenodon, is similar to that of Sphenodon. However, the pattern of distribution of the teeth along the dentalseries resembles that of Theretairus. In Cynosphenodon andTheretairus, the anterior teeth are widely separated while theposterior teeth are closer to each other. In other sphenodontians,the teeth are equally distributed along the dental series.
The principal character shared by Sphenodon and Cynosphenodon is the presence of a single caniniform tooth at theanterior end of the jaw. This tooth was reported in Opisthiasrarus (as described and figured by Gilmore, 1909 and Simpson,1926) and in the Kirtlington sphenodontian (Evans, 1992) . In
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
216 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 16, NO.2 . 1996
m.i.pr
A
rn.d.pr
C.t
pm
pm.f
m.f
FIGURE 6. Cynosphenodon huizachalensis, gen . et sp. nov. A, IGM 6654. lateral view of a right maxilla of an old specimen showing alladditional teeth completely worn . The two anterior teeth seem to be of the successional kind . B. IGM 6655. young right maxilla with apparenthatchling dentition posterior to the caniniform tooth. C, IGM 6661. anterior view of the right premaxilla. Abbreviations as in Figure 2 plus : m.f,maxillary foramina; m.d.pr, maxillary dorsal process; m.i.pr, maxillary infraorbital process; pm.f, premaxillary facet; pm.t, premaxillary teeth.Scale bars equal I mm.
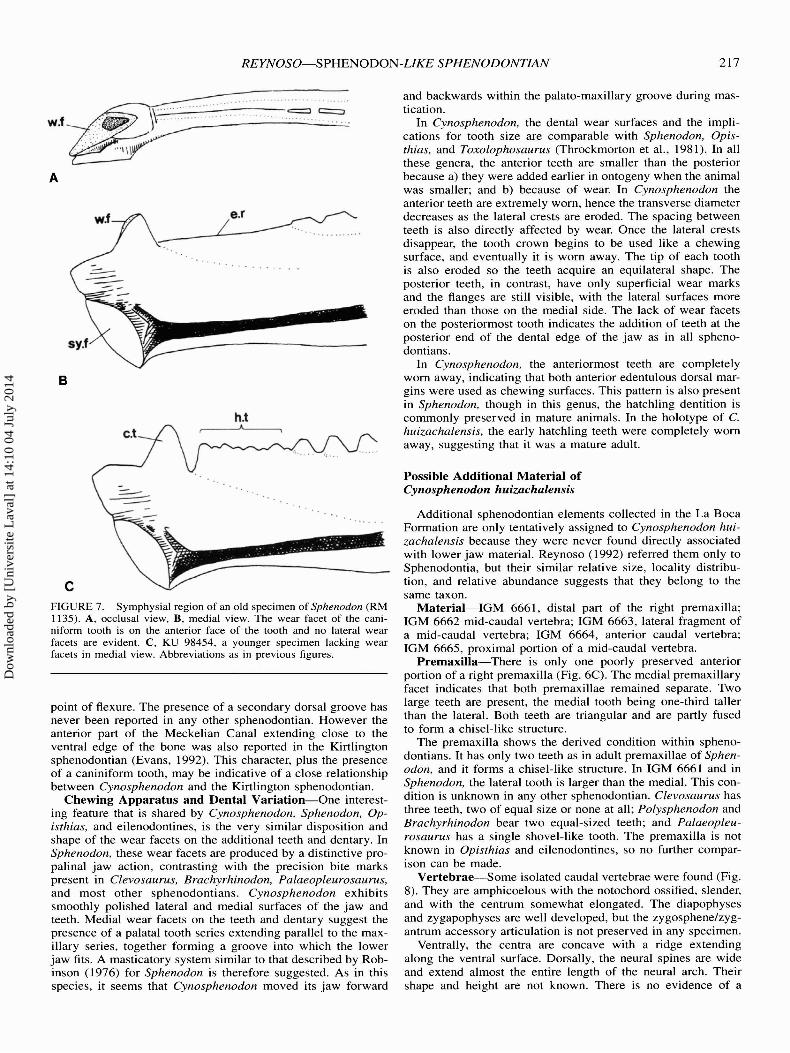
Opisthias, however, this tooth is not present in any of the paratypes, and its presence in the holotype is uncertain (Evans andFraser, 1992; pers. obs .). If the caniniform tooth is present inOpisthias, it would differ from that of Sphenodon in its position . In Opisthias, it would be located in the anteriormost partof the jaw, and in Cynosphenodon, Sphenodon, and the Kirt1ington sphenodontian (Evans, 1992), the caniniform is preceded by an edentulous ridge, and displaced further back in thejaw. According to Robinson (1976), in Sphenodon, hatchlingteeth are present on the anterior edentulous ridge early in ontogeny. This same condition is observed in the juveniles ofCynosphenodon.
A comparison of the distribution of the dental wear facets inthe caniniform tooth of Sphenodon and Cynosphenodon showsthem to be quite different. As seen in a very mature specimenof Sphenodon (RM 1135) , no lateral wear facets are evident,but a major facet is present along the anterior face of the tooth(Fig . 7) . This facet was made by the occlusion between theanterior part of the tooth and the posterior face of the lateralpremaxillary tooth. Such a facet, however, is not present in ayoung specimen of Sphenodon (KU 98454). As described before , the wear facets of the caniniform tooth of Cynosphenodonare on the lateral and medial surfaces. Though the lateral facetcan be explained by inferring occlusion with an anterior max illary tooth, the medial facet is more difficult to explain. In allsphenodontians, the palatine tooth row that runs parallel or subparallel to the maxillary tooth series is located posterior to theposition of the caniniform tooth of the dentary. In Sphenodon,the area anterior to the palatine tooth series, just above the
caniniform tooth position, is occupied by the internal nares . Thetwo possible explanations for the origin of the caniniform medial facet in Cynosphenodon is either the presence of huge vomerine teeth parallel to those of the premaxilla or the extensionof the palatine tooth series further anteriorly. Although, vomerine teeth are present in several sphenodontians such as Planocephalosaurus, Diphydontosaurus, Clevosaurus, and Brachyrhinodon (Fraser, 1982, 1988 ; Whiteside, 1986; Fraser and Benton, 1989), these teeth are not big enough to produce such alarge groove; on the other hand, no known sphenodontians havetheir palatine tooth series extending into the region of the internal nares. Consequently it is postulated that Cynosphenodon,had a rather different arrangement of the anterior palatal region.
While the teeth of Cynosphenodon resemble those of Sphenodon, the overall shape of the dentary is closer to that of Opisthias (Gilmore, 1909) . In Cynosphenodon and Opisthias, theventral edge of the dentary is straight in lateral view, exceptfor the symphysial region. In contrast, the ventral edge of thedentary in Sphenodon and all other sphenodontians describes agentle curve. The straight shape of the ventral edge of the dentary in Cynosphenodon makes the anteroventral expansion ofthe mandibular symphysis more exaggerated. This condition isshared by Opisthias, Cynosphenodon, and probably Sigmala,but not Sphenodon.
The Meckelian Canal of all sphenodontians usually extendsanteroposteriorly along the midline of the medial side of thedentary. The canal of Cynosphenodon is quite distinct in twoways. It angles downwards reaching the ventral edge of thedentary, and contacts a small secondary dorsal groove at the
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
REYNOSO-SPHENODON-LIKE SPHENODONTIAN 217
Additional sphenodontian elements collected in the La BocaFormation are only tentatively assigned to Cynosphenodon huizachalensis because they were never found directly associatedwith lower jaw material. Reynoso (1992) referred them only toSphenodontia, but their similar relative size, locality distribution, and relative abundance suggests that they belong to thesame taxon.
Material-IGM 6661, distal part of the right premaxilla;IGM 6662 mid-caudal vertebra; IGM 6663, lateral fragment ofa mid-caudal vertebra; IGM 6664, anterior caudal vertebra;IGM 6665, proximal portion of a mid-caudal vertebra.
Premaxilla-There is only one poorly preserved anteriorportion of a right premaxilla (Fig. 6C) . The medial premaxillaryfacet indicates that both premaxillae remained separate. Twolarge teeth are present, the medial tooth being one-third talIerthan the lateral. Both teeth are triangular and are partly fusedto form a chisel-like structure.
The premaxilla shows the derived condition within sphenodontians. It has only two teeth as in adult premaxillae of Sphenodon , and it forms a chisel-like structure. In IGM 6661 and inSphenodon, the lateral tooth is larger than the medial. This condition is unknown in any other sphenodontian. Clevosaurus hasthree teeth, two of equal size or none at all; Polysphenodon andBrachyrhinodon bear two equal-sized teeth; and Palaeopleurosaurus has a single shovel-like tooth. The premaxilla is notknown in Opisthias and eilenodontines, so no further comparison can be made.
Vertebrae-Some isolated caudal vertebrae were found (Fig .8). They are amphicoelous with the notochord ossified, slender,and with the centrum somewhat elongated. The diapophysesand zygapophyses are welI developed, but the zygosphene/zygantrum accessory articulation is not preserved in any specimen.
Ventrally, the centra are concave with a ridge extendingalong the ventral surface. DorsalIy, the neural spines are wideand extend almost the entire length of the neural arch. Theirshape and height are not known. There is no evidence of a
Possible Additional Material ofCynosphenodon huizachalensis
and backwards within the palato-maxillary groove during mastication.
In Cynosphenodon, the dental wear surfaces and the implications for tooth size are comparable with Sphenodon, Opisthias, and Toxolophosaurus (Throckmorton et al., 1981) . In allthese genera, the anterior teeth are smalIer than the posteriorbecause a) they were added earlier in ontogeny when the animalwas smalIer; and b) because of wear. In Cynosphenodon theanterior teeth are extremely worn, hence the transverse diameterdecreases as the lateral crests are eroded. The spacing betweenteeth is also directly affected by wear. Once the lateral crestsdisappear, the tooth crown begins to be used like a chewingsurface, and eventualIy it is worn away. The tip of each toothis also eroded so the teeth acquire an equilateral shape. Theposterior teeth, in contrast, have only superficial wear marksand the flanges are stilI visible, with the lateral surfaces moreeroded than those on the medial side. The lack of wear facetson the posteriormost tooth indicates the addition of teeth at theposterior end of the dental edge of the jaw as in all sphenodontians.
In Cynosphenodon, the anteriormost teeth are completelyworn away, indicating that both anterior edentulous dorsal margins were used as chewing surfaces. This pattern is also presentin Sphenodon, though in this genus, the hatchling dentition iscommonly preserved in mature animals. In the holotype of C.huizachalensis, the early hatchling teeth were completely wornaway, suggesting that it was a mature adult.
e.r
C.t
w.f
B
cFIGURE 7. Symphysial region of an old specimen of Sphenodon (RM1135) . A, occlusal view, B, medial view. The wear facet of the caniniform tooth is on the anterior face of the tooth and no lateral wearfacets are evident. C, KU 98454, a younger specimen lacking wearfacets in medial view. Abbreviations as in previous figures .
point of flexure . The presence of a secondary dorsal groove hasnever been reported in any other sphenodontian. However theanterior part of the Meckelian Canal extending close to theventral edge of the bone was also reported in the Kirtlingtonsphenodontian (Evans, 1992). This character, plus the presenceof a caniniform tooth, may be indicative of a close relationshipbetween Cynosphenodon and the Kirtlington sphenodontian.
Chewing Apparatus and Dental Variation-One interesting feature that is shared by Cynosphenodon, Sphenodon, Opisthias, and eilenodontines, is the very similar disposition andshape of the wear facets on the additional teeth and dentary. InSphenodon, these wear facets are produced by a distinctive propalinal jaw action, contrasting with the precision bite markspresent in Clevosaurus, Brachyrhinodon, Palaeopleurosaurus,and most other sphenodontians. Cynosphenodon exhibitssmoothly polished lateral and medial surfaces of the jaw andteeth. Medial wear facets on the teeth and dentary suggest thepresence of a palatal tooth series extending paralIel to the maxillary series, together forming a groove into which the lowerjaw fits. A masticatory system similar to that described by Robinson (1976) for Sphenodon is therefore suggested. As in thisspecies, it seems that Cynosphenodon moved its jaw forward
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
2 18 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 16, NO.2, 1996
FIG URE 8. M idd le an terior ca udal vertebrae of? Cyn ospheno do n huizachalens is. A, dorsal view , B, poste rior view, C , latera l view, D, anterior view , and E, ventral view. a. zy anterior zygapophysis; ce , centrum; ne.arc , neu ra l arch; ne.c, neural ca nal; ne.sp, neural spine; n.c ,notochordal canal ; p. zy posterior zygapop hysis ; tr.p , transverse process .Scale bar eq uals 1 mm .
SYSTE MAT IC AN D TAXO NOMIC REL ATIONSHIPS
Th e verte brae do not show any spec ial fea tures distingu ishingthem fro m any othe r sp henodo ntian vertebrae. Th ey are simi larto the anterio r ca uda l vertebrae of Sp henodon and Gephyrosaurus, although their neural arc hes not as wide dorsovent rall y asfigured by Evans ( 1981 :fig. 9c ) for Gep hyrosaurus, possiblybecause they co rrespond to a more posterior vertebra. The vertebrae referred to Cynosphenodon, however, are more graci lethan those anterior to the autoto mo us vertebrae of Sp henodon.
Th e phylogeneti c rel ation sh ips of the sphe nodo ntians havebee n studied by Whiteside ( 1986 ), Evans ( 1988) , Ga uthier etal. ( 1988), Fraser ( 1988), Fraser and Benton ( 1989) , Wu ( 199 1,1994) and Sues et al. (1994) . However, taxa closely re lated toSp heno don have often been left out of such studies becausemuch of the mat eri al is very inco mplete. Cynosp heno don is alsopoorly known, but several characters ca n be ob served or deduced fro m the ava ilable eleme nts , and these characters permita hypoth esis of the interre lationships of this taxon to be prese nted.
PAUP 3. 1.1 (Swofford, 1992) Bran ch and Bound search wasmade using Wu 's (1994) data matri x, mostly coded as in Sueset al. (1994) . So me change s were made (Table I and Appendix)Character I I and 12, and characters 22 , 23 and 24 were mergedand recoded as suggested by Sues et al ( 1994) . Character 13wa s coded aga in according to the index supratemporal fenestralength/ skull lengt h. Although Wu's index is easier to est imate,it does not reflect the relative size of the tem poral region , asproposed by Fraser and Ben ton ( 1989 :character 18). The indexsupratemporal fenes tra/orbit does not properly describe anyskull character. Th e derived condition of character 29, "presence of ex tens ive posterolingual flanges on so me maxillaryteeth," de pends upon the presence of flanges (character 26) , sothat there are two di ffe ren t derived states of the sa me character[charac ter 34 here]. Th e charac ter is newly coded as suggestedby Ga uthier et al. ( 1988 ). Th e cha racter state " all teeth acrodont " (Benton, 1985 ; Eva ns , 1988) is a clear sy napo mo rphydeterminabl e in most sphenodo ntians . It was incl uded as anex tra state of the cha rac ter " de nta l implantation" [character26 ]; the state " lateral tooth attac hme nt reduced (Gauthier et al.,1988) showing some degree of acrodo nty" (Fraser and Bent on ,1989) is also incl uded.
Several character s used by Benton ( 1985), Fraser ( 1988),Ga uthie r et al. (1988), and (or) Fraser and Bent on (1989) wererei ntroduced to the ana lysis. C haracters [20], [21 ], [22], [27 ],[29 ], [30], [31], [37 ], [40], [41] and [42] were ignore d by Wu
ne.c
p.zy
a.zy
ne.sp
A
c
suture be tween the neural arc h and the ce ntrum nor of subcentral fora mina .
Th e most anterior verte bra is co mpressed dorsoventrall y andthe tran sverse processes are present. In the mid-anterior caudalverte brae, the transverse processes are well devel oped and perpendicul ar to the longitudinal axis . Th eir bases are broad and,a lthough their total length is unknown, they see m to be qu itesho rt. Th e transverse processes gradually becom e sho rte r inmore poster ior vertebrae. No ne of the vertebrae show a septumfor autotomy, perh aps becau se no posterior ca uda l ve rtebrae arerepresented .
E
TABLE I. Character state ma trix used to estab lish phy logenetic re lationshi ps o f rhy nchocepha lia ns. Abbreviations: 0 = primitive ; I , 2, and 3 =derived states; ? = unknown; - = not applicable ; V = variable (0 & I ); X = variable ( I & 2) .
lJ) :::. lJ) lJ) c: .g 2 lJ) :::.2 ('(I Q e .glJ) lJ) 2 ('(I:::. s ('(I :::. a a 2 :::. c :::. lJ) a('(I .c:
~ c: .S ('(I a ('(I c: a c: cQ. lJ) c g. Ql -e :::.
~ "tl .c: Ql .g::;, e .g §- ('(I lJ) .g~
.c:.c: a a~ E u g. E lJ) Ql
.§ Ql lil- aE a Ql ~ a a c
.S' c: ~ ~u E .c: c: ~ a Ql
2- .9-~ ~ Ql
~lij- Ql s .c:::;, e & <3 ~ in ~ ~0 (!) o CO CJ)
UG
clevosaurs
Sphenodontidae
A
Sphenodontia
Rhynchocephalia
HGVRE 9. Most parsimonious tree obtained by the Branch andBound search function of PAVP. All characters are unordered and Opisthias was deleted. Tree length = 79, Consistency Index = 0.722, andRetention Index = 0.798 . Ambiguous characters are in brackets. Rhynchocephalia: 8, 14,23, [35]. Sphenodontia: 6,26,27, [32] . Node A: 21,24, 30, 39, [32(2), 40] . Sphenodontidae : 2, 8(0) , 26(2), 28, 33, 36 [22 ,34(2)]. Node B: 5, 18, 37, [15(0)]. Clevosaurs (sensu Wu 1994) : 1(2),3,4,7, [38] . Node C: 34. Node 0: 13, 16,41(2), [38(2)]. Node E: 10,II, [12]. Node F: I. Node G: 25, [19, 34]. Sphenodontinae: 31. NodeH = Eilenodontinae: 20, 29. Gephyrosaurus: [9] . Diphydontosaurus: I,[22]. Planocephalosaurus: [9, 15(0), 34, 38] . Palaeopleurosaurus: 10,II, 12(2) , 14(0), 17, [9,38(2)]. Polysphenodon: 2(0) ,42. Homoeosaurus: 2(0) ,25,42. Sphenodon: [3,7, 17,40(2),41(1)].
(1994). In character [40] the state " prominent posterior processof ischium" (Evans 1988: character K14), was also reconsidered. Although some of these characters may be dubious (e.g.,limb proportions), this study is attempting to incorporate allavailable evidence (Kluge 1989). Characters 21, 22 , and 23 ofFraser and Benton (1989), character 20 of Wu (1994) and characters 1,4, 9, 33, and 31 of Sues et al (1994) are autapomorphicand so were excluded.
Several outgroup states were miscoded by Wu (1994: characters 30, 31, and 32: tables I and 2). Squamates do not havea palatine tooth row, and they may have one pterygoid toothrow or none at all. These characters were coded as 'not applicable' in squamates, adding the state " none or one pterygoidtooth rows " (state 2) splitting and modifying Wu's state (1) ofthe character " number of pterygoid tooth rows" [38 here]. Topolarize conflicting and variable states in squamates, Younginawas included as a second outgroup. The inclusion of Young inareversed the polarity of character [IS] .
The resulting single most parsimonious tree (Fig. 9) has thesame basic topology as that published by Wu (1994:fig. 21)showing a single minor change in the sister-group relationshipsof " clevosaurs" (sensu Wu, 1994). According to this hypothesis
the sister relationship of Polysphenodon and Brachyrhinodon issupported by two characters: antorbital region/skull length lessthan one-fourth (not shared with any other taxa) and maxillaryteeth with a small medial flange. In Wu's hypothesis, the sistergrouping of Clevosaurus and Brachyrhinodon is supported bya single character: inferior temporal fenestra/skull length morethan one-fourth. If the sister-grouping of these taxa is forced,the tree length increases by one step.
According to the most parsimonious hypothesis, Cynosphenodon is the sister group of Sphenodon. It shares several synapomorphies with the Rhynchocephalia as defined by Gauthieret al. (1988). The presence of the posterior process of the den tary projecting beyond the coronoid process is unique to rhynchocephalians and is also present in Cynosphenodon. The pre sence of tooth replacement by adding teeth at the back of thejaw, places this genus within the Sphenodontia as defined byBenton (1985). The presence of a high coronoid process, abroad mandibular symphysis, anterior hatchling teeth, and anterolabial flanges on at least one dentary tooth, strongly supports the placement of Cynosphenodon within a clade of sphenodontians that excludes Diphydontosaurus (Node A, Fig. 9). Alarge mandibular foramen, fully acrodont dentition, well developed wear facets, and three or less premaxillary teeth forminga chi sel-like structure are characters shared by Cynosphenodonand all the sphenodontians except Diph ydontosaurus and Plan ocephalosaurus. In summary, derived states of characters [21] ,[22]. [23], [24] , [26], [27] , [28], [30], [32] , [33] , and [39]strongly support that Cyno sphenodon is a crown member of theSphenodontia. However additional resolution is only providedby a few characters.
One of the most studied features in sphenodontian morphology is the form and function of the jaw (Farlow, 1975 ; Robinson, 1976 ; Throckmorton et al., 1981; Rasmussen and Callison , 1981; Gorniak et al., 1982 ; Whiteside, 1986) . Of specialinterest is the propalinal jaw action present in several sphenodontians. Propalinal jaw action is related to other modificationsof the mandible: antero-posterior enlargement of the mandibulararticular surface, and the presence of smooth wear facets on thedentary and teeth (Gauthier et al., 1988) . The presence of thesecharacters supports the monophyly of Node G (Fig . 9). Thesefeatures are shared by Toxolophosaurus, Eilenodon, and Sphenodon. Smooth wear facets on the dentary and teeth are alsoundoubtedly present in Cynosphenodon. The reduction of theretroarticular process is also synapomorphic at this node. However, the character is not known in Toxolophosaurus and Cynosphenodon. Toxolophosaurus and Eilenodon are sister-taxasharing the presence of mediolateral expansion of the additionalteeth and a deep jaw.
The incorporation of Cynosphenodon leads to the addition ofthe new character "presence of a caniniform tooth posterior tothe anterior edentulous ridge in adults", once autapomorphic ofSphenodon. The presence of a caniniform tooth was previouslysuggested by Gauthier et al. (1988) and Evans (1988) as a synapomorphy of Opisthias and Sphenodon. This tooth, however,is not clearly present in Opisthias (Evans and Fraser, 1992 ; pers .obs. ) and no clear relation of this taxon to Sphenodon is supported. A caniniform tooth posterior to an edentulous marginis present in Sphenodon and Cynosphenodon, as well as inTheretairus (which has two caniniform teeth) and in an unnamed sphenodontian from England (Evans, 1992). This synapomorphy diagnoses the Sphenodontinae. The presence of caniniform teeth is shared by four taxa, and although the numberof taxa that share a character is not an argument to support thequality of the character and the group it defines, the hypothesisof it being acquired separately is much less parsimonious. TheSphenodontinae here described is not the same as Gauthier etal.'s (1988) informal group " sphenodonts" (Opisthias + Sphenodon). The possible lack of a caniniform tooth in Opisthias
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
220 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 16, NO.2, 1996
restricts it from the clade as here defined. Further studies onOpisthias will clarify its phylogenetic status (Fraser and Evans1992).
CONCLUSIONS
The morphotype of Sphenodon was considered a highlyprimitive reptile by early workers, and it has been used as amodel to establish polarity in squamate phylogeny (e.g ., Esteset al ., 1988). As result of recent studies (Carroll, 1977; Benton,1985; Whiteside, 1986; Evans, 1988; Fraser and Benton, 1989),the view of Sphenodon as a primitive lepidosaur must be modified . Within the rhynchocephalian lineage, Sphenodon is ahighly derived genus.
In spite of this new understanding, the morphology and ageof Cynosphenodon shows that derived features such as the presence of a caniniform tooth, shared with Sphenodon, are notrecent among sphenodontians. This structure arose at least inthe Middle Jurassic. On the other hand, Robinson (1976) suggested that propalinal movements arose relatively late in sphenodontian evolution. Throckmorton et al. (1981), however, suggested that such movements were present as early as the LateJurassic. The discovery of Cynosphenodon indicates that thisfunction was developed even earlier among sphenodontians.
ACKNOWLEDGMENTS
I especially would like to thank J. Clark who invited me toparticipate in the description of the Huizachal sphenodontians.I also want to thank M. Montellano, 1. Ferrusqufa, L. Espinosa,and O. Flores for reading the original Spanish version of thispaper, and to J. Clark, H. D. Sues and X. Wu for reading theEnglish version. I thank D. Fastovsky for introducing me to thegeology of the Huizachal Canyon and for providing me withaccess to unpublished information. I am indebted to R. Hernandez and J. Clark who collected many of the fossils describedhere and to R. Carroll, for permitting me to use his copy ofPAUP. Finally I deeply appreciate the hospitality of Sr. F. Moreno and all the people of the Ejido EI Huizachal.
The research and field work was supported by the DireccionGeneral de Asuntos del Personal Acadernico of the UniversidadNacional Autonorna de Mexico and the National Science Foundation, Grants EAR 8917368, 81917389, 9218871, 9218971,and a Grant of the National Geographic Society.
LITERATURE CITED
Benton, M. J. 1985 . Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society 84:97-164.
Carroll , R . L. 1977 . The origin of lizards; pp . 359-396 in S. M . Andrews, R . S. Miles and A. D. Walker (eds.), Problems in VertebrateEvolution. Linnean Society Symposium Series 4
---1985. A pleurosaur from the Lower Jurassic and the taxonomicposition of the Sphenodontida. Palaeontographica, Abteilung A,189: 1-28.
Clark, J. M ., and J . A. Hopson. 1985. Distinctive mammal-like reptilefrom Mexico and its bearing on the phylogeny of the Tritylodontidae. Nature 315:398-400.
---M. Montellano, J. A. Hopson, and R. Hernandez. 1991. Mammals and other tetrapods from the early Jurassic La Boca Formation, Northeastern Mexico. Journal of Vertebrate Paleontology II:23A.
--- and R. Hernandez. 1994. A new burrowing diapsid from theJurassic La Boca Formation of Tamaulipas, Mexico. Journal of Vertebrate Paleontology 14: 180-195.
Cocude-Michel, M. 1963 . Les rhynchocephales et les sauriens de calcaires lithographiques (Jurassique superieur) d'Europe occidentale.Nouvelles Archives du Museum d'Histoire Naturelle de Lyon 7:1187.
Daugherty, C. H ., A. Cree, J. M . Hay, and M . B. Thompson. 1990.Neglected taxonomy and continuing extintions of tuatara (Sphenodon) . Nature 347 :117-119.
Estes, R ., K. de Queiroz, and J. A . Gauthier. 1988 . Phylogenetic relationships within Squamata; pp. 119-183 in R. Estes and G. Pregill (eds.), Phylogenetic Relationships of the Lizard Families. Stanford University Press. Stanford.
Evans, S. E . 1980. The skull of a new eosuchian reptile from the LowerJurassic of South Wales. Zoological Journal of the Linnean Society,London 70 :203-264.
---1981. The postcranial skekeleton of the Lower Jurassic eosuchian Gephyrosaurus bridensis. Zoological Journal of the LinneanSociety 73 :81-116.
---1984. The classification of Lepidosauria. Zoological Journal ofthe Linnean Society. 82 :87-100.
---1988. The early history and relationships of the Diapsida; pp .221-260 in M. J. Benton (ed.) , The Phylogeny and Clasification ofthe Tetrapods, Vol. I . Clarendon Press, Oxford.
---1992. A sphenodontian (Reptilia: Lepidosauria) from the Middle Jurassic of England. Neues Jahrbuch fur Geologie und Palaontologie Mh . 1992:449-457.
--- and N. C. Fraser. 1992 . A sphenodontid jaw (Reptilia: Lepidosauria) from the Upper Jurassic of Dorset. Proceedings of theDorset Natural History Society 1992: 199-200.
Fabre, J . 1973. Un squelette d ' Homoeosaurus aff. solnhofensis (Rhynchocephalia) du Portlandien du Petit Plan de Canjuers (Var). Comptes Rendus de I' Academie des Sciences, Paris 276: 1139-1142.
---1974. Un squelette de Pleurosaurus ginsburgi nov . sp . (Rhynchocephalia) du Portlandien du Petit Plan de Canjuers (Var.) .Comptes Rendus de I' Academic des Sciences, Paris. Serie D 278 :2417-2420.
Farlow, J. O . 1975 . Observations on captive tuatara (Sphenodon punctatusy. Journal of Herpetology 9:353-355 .
Fastovsky, D. E., J. M. Clark, and J. A. Hopson. 1987. Preliminaryreport of a vertebrate fauna from an unusual paleoenvironmentalsetting, Huizachal Group, Early or Mid-Jurassic, Tamaulipas, Mexico. Fourth Symposium on Mesozoic Terrestrial Ecosystems 3:8287.
------ N. H. Strater, M. Montellano, R. Hernandez R., and J.A. Hopson. 1995. Depositional environments of a Middle Jurassicterrestrial vertebrate assemblage. Huizachal Canyon, Mexico. Journal of Vertebrate Paleontology 15:561-575.
Fraser, N. C. 1982. A new Rhynchocephalian from the British UpperTriassic. Palaeontology 25:709-725.
---1986. New Triassic sphenodontids from south-west England anda review of their classification. Palaeontology 29 :165-186.
---1988. The osteology and relationships of Clevosaurus (Reptilia:Sphenodontida). Philosophical Transactions of the Royal Society ofLondon, Series B 321 :125-172.
--- and M. J. Benton. 1989. The Triassic reptiles Brachyrhinodonand Polysphenodon and relationships of the sphenodontids . Zoological Journal of the Linnean Society 96:413-445.
Gauthier, J ., R. Estes, and K. de Queiroz. 1988 . A phylogenetic analysis of Lepidosauromorpha; pp . 15-98 in R. Estes and G. Pregill(eds.) , Phylogenetic Relationships of the Lizard Families. StanfordUniversity Press, Stanford.
Gilmore, C. W. 1909 . A new rhynchocephalian reptile from the Jurassicof Wyoming, with notes on the fauna of "Quarry 9" . United StatesNational Mu seum, Proceedings 37:35-42.
Gorniak, G. C; H. I. Rosenberg, and C. Gans. 1982. Mastication inthe Tuatara, Sphenodon pun ctatus (Reptilia: Rhynchocephalia).Structure and activity of the motor system. Journal of Morphology171:32 1- 353.
Gunther, A. 1867 . Contribution to the anatomy of Hatteria (Rhynchocephalus, Owen). Philosophical Transactions of the Royal Societyof London 157 :595-629.
Hoffstetter, R . 1955 . Rhynchocephalia; pp . 556-576 in J . Piveteau(ed .), Traite de Paleontologie, vol. 5 .
Huene, F. V. 1952. Revision der Gattung Pleurosaurus auf Grund neuerund alter Funde. Palaeontographica A1OI :167-200.
Kluge, A. G. 1989. A concern for evidence and phylogenetic hypothesis of relationships among Epicrates (Boidae, Serpentes). Systematic Zoology 38 :7-25.
Rasmussen, T. E., and G. Callison. 1981. A new herbivorous sphenodontid (Rhynchocephalia: Reptilia) from the Jurassic of Colorado.Journal of Paleontology 55 :1109-1116.
Reynoso, V. H. 1992. Descripcion de los esfenodontes (Sphenodontia,Reptilia) de la fauna del Canon del Huizachal (Jurasico Temprano-
Dow
nloa
ded
by [
Uni
vers
ite L
aval
] at
14:
10 0
4 Ju
ly 2
014
REYNOSO-SPHENODON-LIKE SPHENODONTIAN 221
Medio), Tamaulipas, Mexico. Master's Thesis, Universidad Nacional Aut6noma de Mexico.
Robinson, P. L. 1976. How Sphenodon and Uromastix grow their teethand use them; pp . 43-64 in A. d'Bellairs and C. B. Cox (eds.),Morphology and Biology of Reptiles . Academic Press, London.
Simpson, G. G. 1926. American terrestrial Rhynchocephalia. AmericanJournal of Science 5:12-16.
Swofford, D. L. 1993 . PAUP: Phylogenetic Analysis Using Parsimony,Version 3.1.1. Computer program distributed by The Illinois Natural History Survey, Champaign, llIinois.
Sues, H .-D ., and D . Baird. 1993 . A skull of a sphenodontian lepidosaurfrom the New Heaven Arkose (Upper Triassic: Norian) of Connecticut. Journal of Vertebrate Paleontology 13:370-372.
---N. H . Shubin, and P. E. Ol son. 1994. A new sphenodontian(Lepidosauria: Rhynchocephalia) from the McCoy Brook Formation (Lower Jurassic) of Nova Scotia, Canada. Journal of VertebratePaleontology 14:327-340.
Throckmorton, G. S., J. A. Hopson, and P. Parks. 1981. A redescriptionof Toxolophosaurus cloudi Olson, a Lower Cretaceous herbivoroussphenodontid reptile. Journal of Paleontology 55 :586-597.
Whiteside, D. I. 1986. The head skeleton of the Rhaetian sphenodontidDiphydontosaurus avonensis gen. et sp . nov. and the modernizingof a living fossil. Philosophical Transactions of the Royal Societyof London, Series B 312:379-430.
Wu, X.-c. 1991. The comparative anatomy and systematics of Mesozoic sphenodontidans. Doctoral thesis, Me Gill University.
---1994. ?Late Triassic-Early Jurassic sphenodontidans (Clevosaurus) from China and the Phylogeny of the Sphenodontida; pp. 3869 in N. C. Fraser and H.-D . Sues (eds.), In the Shadow of theDinosaurs: Early Mesozoic Tatrapods. Cambridge University Press,Cambridge and New York.
Received 13 July 1994; accepted 9 February 1995.
APPENDIX
Characters and character states used for the rhynchocephalian phylogenetic analysis . Character numbers in parentheses correspond tothose of Wu (1994).
Skull
1 (11 , 12 modified). Antorbital region/skull length: one-third or more(0) ; between one-fourth and one-third (I); one fourth or less (2) .
2 (13 modified). Supratemporal fenestra/skull length: one-fourth or less(0); more than one-fourth (I).
3 (14). Lower temporal fene stra/skull length: one-fourth or less (0) ; morethan one-fourth (I).
4 (I). Premaxillary process of maxilla: elongate (0); reduced (I).5 (2). Posterior portion of maxilla: gradually tapering off or very narrow
(0); broad (I).6 (3) . Lacrimal: present (0) ; absent (1).7 (9) . Dorsal process jugal: broad and short (0); narrow and elongate (I).8 (4) . Frontals: separated (0) ; fused (I).9 (5) . Parietals: separated (0); fu sed (1).
(I); or convex (2).13 (10). Parietal foramen position: posterior to the anterior margin of
supratemporal fenestra (0); level or anterior (I).14 (17). Lower temporal bar: aligned with the maxillary tooth row (0);
bowed away beyond the limit of the abductor chamber (I).15 (18 polarity reversed) . Lower temporal bar: complete (0); incom
plete (I).
Palate
16 (33). Palatine: tapers posteriorly (0) ; becomes relatively wide (I).17 (34). Central region of pterygoid between three rami: short (0) ; elon-
gate (1).18 (35). Pterygoid: borders suborbital fenestra (0); precluded from the
suborbital fenestra (I).
Mandible
19 (36). Jaw motion: orthal (0) ; propalinal (I) .20. Mandible: narrow (0) ; deep (1), (Rasmussen and Callison, 1981;
see also Benton, 1985)21. Mandibular symphysis breadth: slender (0) ; broad (1), (Benton,
1985: character Z13)] .22. Mandibular foramen: small (0) ; big (I), (Benton, 1985: character
ZI5).23 (15). Posterior process of dentary: ends anteriorly to coronoid pro
cess (0); ends posteriorly (I).24 (16). Coronoid process : absent or weak (0); pronounced (1) .25 (19). Retroarticular process size: pronounced (0); reduced (1).
Teeth
26 (21 modified) . Dental implantation: pleurodont (0) ; a degree of acrodonty (I); fully acrodont (2) .
27 . Tooth replacement: alternate (0) ; addition of teeth at the back of thejaw (I), (Benton, 1985: character Z8)] .
28 (27). Lateral and medial wear facets on marginal teeth: absent orpoorly developed (0); well established (I) .
29. Marginal teeth breadth: equal to length (0); mediolaterally expanded(I), (Fraser and Benton, 1989: character 27).
30. Dental regionalization with small juvenile teeth at anterior end ofmaxilla and dentary: absent (0) ; present (I), [Benton, 1985 : characterZ7)].
31. Anterior caniniform tooth on dentary and maxilla: absent (0) ; present (I), (Gauthier et al ., 1988: character 51).
32 (22 , 23, 24 merged). Number of premaxillary teeth: more than seven(0); seven to four (I); three or less (2) .
33 (25). Premaxillary teeth: separated (0) ; forming a chisel-like structure in mature individuals (1).
34 (26, 29 merged). Posterior maxillary tooth shape: simple cones (0);with posteromedial flanges in at least one tooth (I); with extensiveflanges (2) .
35 (30). Lateral palatine tooth row: small (0) ; enlarged (1) .36 (31). Number of palatine tooth rows: more than one (0) ; a single
lateral tooth row (1) .37. Flanges on palatal tooth row: absent (0); present on some (I), (Fra
ser and Benton, 1989: character 15) .38 (32 modified). Number of pterygoid tooth rows: three or more (0) ;
two rows (I); one or ab sent (2).39 (28). Anterolateral flanges on dentary teeth: absent (0); present on
at least one tooth (1) .
Postcranial
40. Posterior process of second sacral vertebra: small (0); prominent(1) ; absent (2), (Gauthier et al., 1988: character 88, modified).
41. Ischium: uninterrupted posterior edge extending from the acetabulum to the median symphysis (0); posterior process on posteriormargin (1); prominent posterior process present (2), (Evans, 1988 :character K14; Fraser and Benton, 1989: character 4, modified).
42. Limb proportions with respect to the presacral column: humerus:=:; 0.20, tibia es 0 .25, femur < 0 .30, radius < 0.15 (0) ; all measuresgreater than these values (1), (Fraser and Benton, 1989: character29).