A NEW GEKKO FROM THE BABUYAN ISLANDS, NORTHERN PHILIPPINES RAFE M. BROWN 1,4 ,CARL H. OLIVEROS 1,2 ,CAMERON D. SILER 1 , AND ARVIN C. DIESMOS 3 1 Natural History Museum and Biodiversity Research Center, Department of Ecology and Evolutionary Biology, The University of Kansas, Lawrence, KS 66045-7561, USA 2 ISLA Biodiversity Conservation; 9 Bougainvillea St, Manuela Subdivision, Las Pinas City 1741 Philippines 3 National Museum of the Philippines, Rizal Park, Padre Burgos Ave. Ermita 1000, Manila, Philippines ABSTRACT: We describe a new species of gekkonid lizard on the basis of 21 recently acquired specimens from Babuyan Claro Island, Babuyan Islands group, northern Philippines. The new species differs from other Philippine Gekko by characteristics of external morphology, color pattern, and body size. The new species has been found low on trunks and buttresses of mature closed-canopy climax forest trees at low elevation near the island’s coast. It is known from only Babuyan Claro Island and is likely endemic to this single small, isolated landmass. The remaining gekkonid fauna of the Babuyans and Batanes island groups is understudied and in need of comprehensive review. Key words: Babuyan Claro; Babuyan Islands; Gekkonidae; New species; Philippines PHILIPPINE lizards of the family Gekkonidae are represented by 10 genera and 37 species: Cyrtodactylus (4), Gekko (9), Gehyra (1), Hemidactylus (5, including H. platyurus,a species formerly assigned to Cosymbotus), Hemiphylodactylus (1), Lepidodactylus (6), Luperosaurus (6), Pseudogekko (4), and Pty- chozoon (1) (Brown and Alcala, 1978; Brown et al., 2007; Gaulke et al., 2007; Taylor, 1922a,b). Although the lack of definitive characters distinguishing the genera Lupero- saurus and Gekko have clouded generic boundaries (Brown and Alcala, 1978; Brown et al., 2000a; Brown et al., 2007; Russell, 1979), most authors have applied the name Gekko to Philippine species that possess (1) moderate to large body size and longer, more slender limbs; (2) near or complete absence of interdigital webbing; (3) a lack of cutaneous expansions, save for ventrolateral adipose folds and very slight expansions bordering the posterior margins of the hind limbs; (4) convex to conical enlarged dorsal tubercles arranged in longitudinal rows; (5) enlarged, plate-like subcaudals; (6) differentiated, slen- der, and elongate postmentals; and (7) minute and non-imbricate rows of dorsal scales between tubercle rows (minute dorsals differ- entiated from the enlarged, imbricate scales of the venter (Brown and Alcala, 1978; Brown et al., 2000a; Brown et al., 2007). Philippine members of the genus Gekko include seven endemic species (G. athymus, G. ernstkelleri, G. gigante, G. mindorensis, G. palawanensis, G. porosus, and G. romblon; Brown and Alcala, 1978; Brown et al., 2007; Roesler et al., 2006; Taylor, 1922a), two species shared with neighboring countries (G. gecko and G. monarchus; Manthey and Grossman, 1997; Wermuth, 1965), and one species (G. hokouensis) that is probably included in the country’s gekkonid fauna in error (Brown and Alcala, 1978; Ota et al., 1989). Several additional Philippine gekkonid taxa presently await description, including multiple unnamed species from the northern Philip- pines. During recent biological inventory work in the Babuyan archipelago, we discov- ered a distinctive species of Gekko on Babuyan Claro Island, now represented in U.S. and Philippine collections by 21 speci- mens. In this paper we describe the new species and discuss morphological diversity, patterns of gekkonid endemism, and probable processes of evolutionary diversification in the Philippine island radiation of Gekko. MATERIALS AND METHODS We (RMB and CDS) collected data from fluid-preserved specimens deposited in U.S. and Philippine collections (see Acknowledg- ments). Sex was determined by inspection of gonads or by scoring of prominent secondary 4 CORRESPONDENCE: e-mail, [email protected]Herpetologica, 64(3), 2008, 305–320 E 2008 by The Herpetologists’ League, Inc. 305
Transcript
A NEW GEKKO FROM THE BABUYAN ISLANDS,NORTHERN PHILIPPINES
RAFE M. BROWN1,4, CARL H. OLIVEROS
1,2, CAMERON D. SILER1, AND ARVIN C. DIESMOS
3
1Natural History Museum and Biodiversity Research Center, Department of Ecology and Evolutionary Biology,The University of Kansas, Lawrence, KS 66045-7561, USA
2ISLA Biodiversity Conservation; 9 Bougainvillea St, Manuela Subdivision, Las Pinas City 1741 Philippines3National Museum of the Philippines, Rizal Park, Padre Burgos Ave. Ermita 1000, Manila, Philippines
ABSTRACT: We describe a new species of gekkonid lizard on the basis of 21 recently acquired specimensfrom Babuyan Claro Island, Babuyan Islands group, northern Philippines. The new species differs from otherPhilippine Gekko by characteristics of external morphology, color pattern, and body size. The new species hasbeen found low on trunks and buttresses of mature closed-canopy climax forest trees at low elevation near theisland’s coast. It is known from only Babuyan Claro Island and is likely endemic to this single small, isolatedlandmass. The remaining gekkonid fauna of the Babuyans and Batanes island groups is understudied and inneed of comprehensive review.
Key words: Babuyan Claro; Babuyan Islands; Gekkonidae; New species; Philippines
PHILIPPINE lizards of the family Gekkonidaeare represented by 10 genera and 37 species:Cyrtodactylus (4), Gekko (9), Gehyra (1),Hemidactylus (5, including H. platyurus, aspecies formerly assigned to Cosymbotus),Hemiphylodactylus (1), Lepidodactylus (6),Luperosaurus (6), Pseudogekko (4), and Pty-chozoon (1) (Brown and Alcala, 1978; Brownet al., 2007; Gaulke et al., 2007; Taylor,1922a,b). Although the lack of definitivecharacters distinguishing the genera Lupero-saurus and Gekko have clouded genericboundaries (Brown and Alcala, 1978; Brownet al., 2000a; Brown et al., 2007; Russell,1979), most authors have applied the nameGekko to Philippine species that possess (1)moderate to large body size and longer, moreslender limbs; (2) near or complete absence ofinterdigital webbing; (3) a lack of cutaneousexpansions, save for ventrolateral adiposefolds and very slight expansions borderingthe posterior margins of the hind limbs; (4)convex to conical enlarged dorsal tuberclesarranged in longitudinal rows; (5) enlarged,plate-like subcaudals; (6) differentiated, slen-der, and elongate postmentals; and (7) minuteand non-imbricate rows of dorsal scalesbetween tubercle rows (minute dorsals differ-entiated from the enlarged, imbricate scales ofthe venter (Brown and Alcala, 1978; Brown etal., 2000a; Brown et al., 2007).
Philippine members of the genus Gekkoinclude seven endemic species (G. athymus,G. ernstkelleri, G. gigante, G. mindorensis, G.palawanensis, G. porosus, and G. romblon;Brown and Alcala, 1978; Brown et al., 2007;Roesler et al., 2006; Taylor, 1922a), twospecies shared with neighboring countries(G. gecko and G. monarchus; Manthey andGrossman, 1997; Wermuth, 1965), and onespecies (G. hokouensis) that is probablyincluded in the country’s gekkonid fauna inerror (Brown and Alcala, 1978; Ota et al.,1989).
Several additional Philippine gekkonid taxapresently await description, including multipleunnamed species from the northern Philip-pines. During recent biological inventorywork in the Babuyan archipelago, we discov-ered a distinctive species of Gekko onBabuyan Claro Island, now represented inU.S. and Philippine collections by 21 speci-mens. In this paper we describe the newspecies and discuss morphological diversity,patterns of gekkonid endemism, and probableprocesses of evolutionary diversification in thePhilippine island radiation of Gekko.
MATERIALS AND METHODS
We (RMB and CDS) collected data fromfluid-preserved specimens deposited in U.S.and Philippine collections (see Acknowledg-ments). Sex was determined by inspection ofgonads or by scoring of prominent secondary4 CORRESPONDENCE: e-mail, [email protected]
Herpetologica, 64(3), 2008, 305–320
E 2008 by The Herpetologists’ League, Inc.
305
sexual characteristics (Brown, 1999; Brown etal., 1997, 2000a;) when dissection was notpossible. Measurements (to the nearest0.1 mm) were taken with Fowler digitalcalipers following character definitions byOta and Crombie (1989), Brown et al.(1997), Brown (1999), and Brown et al.(2007). Character abbreviations are: snout–vent length (SVL); tail length (TL); headlength, (HL); head width (HW); head depth(HD); snout length (SNL); eye diameter,(ED); eye–narial distance (END); auricularopening diameter (AO); inter-narial distance(IND); inter-orbital distance (IOD); axilla–groin distance (AGD); femur length (FL);tibia length, (TBL); Toe I length (TiL); Toe IVlength (TivL); tail width (TW); and tail depth(TD); number of supralabials (SUL) andinfralabials (IFL) to the center of the eyeand posteriorly to the point at which pointlabials are no longer differentiated; enlargedcircumorbitals dorsoanterior to orbit (CO);modified spiny circumorbitals (cilaria;) dorso-posterior to orbit (SC); differentiated preanalpore-bearing scales (PS); femoral pore-bear-ing scales (FS); differentiated subdigital scan-sors on fingers 1–5 (FS I–V); subdigitalscansors on toes 1–5 (TS I–V); midbodyventral transverse scale rows counted betweenlateral body folds (MBVS); midbody dorsaltransverse scale rows counted across dorsumbetween lateral body folds (MBDS); midbodytransverse tubercle rows counted across dor-sum between dorsolateral body folds (MBTR);paravertebrals between midpoints of limbinsertions (PVS); ventrals within AGD (count-ed midventrally between limb insertion; VS)paravertebral tubercles within AGD (countedalong right side of vertebral axis between limbinsertions; PVT); tail annuli (in complete tails;TA); and subcaudals (beneath complete tails;SC). For the recognition of the new species,we adopted the General Lineage SpeciesConcept (GLC) of de Queiroz (1998, 1999)as the natural extension of the EvolutionarySpecies Concept (Wiley, 1978). Application oflineage-based species concepts to islandendemics is straightforward because of theknown history of isolation of island popula-tions (Brown and Diesmos, 2002; Brown andGuttman, 2002). We consider as new speciesmorphologically diagnosable, isolated popula-
tions where the hypothesis of a distinctevolutionary lineage cannot be rejected.
SYSTEMATICS
Gekko crombota sp. nov.Figs. 2–5
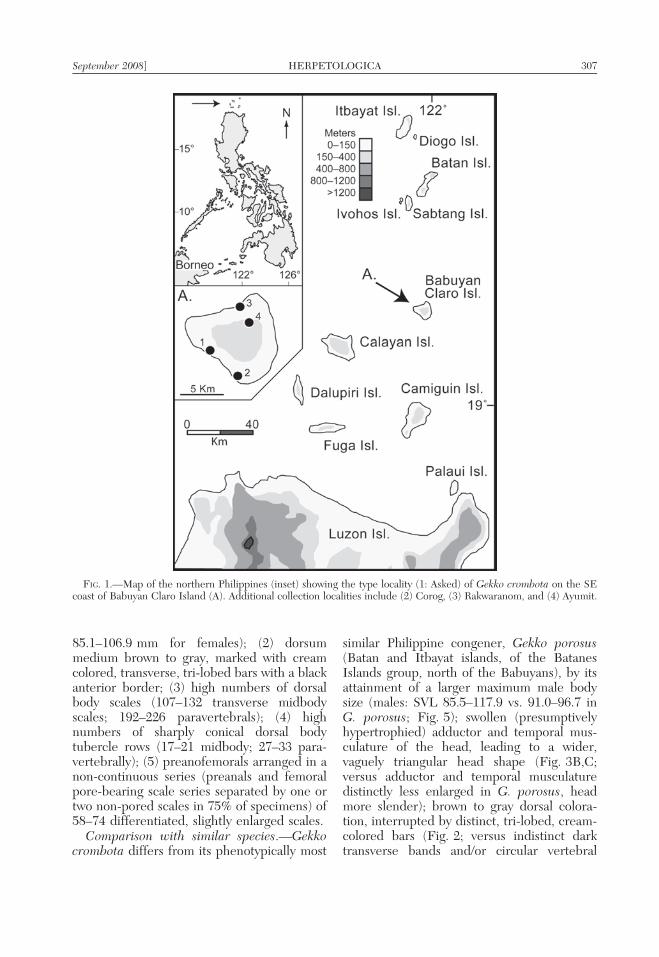
Holotype.—PNM 9280 (Field no. RMB5954; formerly KU 304833), an adult malecollected at night (2125 h on 13 March 2005)on the buttress of a large dipterocarp treeby RMB at an area known locally as‘‘Asked,’’ Barangay Babuyan Claro, Munici-pality of Calayan, Cagayan Province, Wcoast of Babuyan Claro Island, Philippines(19.5041u N, 121.9120u E; 20 m above sealevel).
Paratopotypes.—KU 304825 adult male;KU 304808, 304814, 304821, 304848, PNM9281–82 (formerly KU 304829, 304832), twomale juveniles and four juveniles of undeter-mined sex; KU 304807, subadult female; KU304809, 304826, 3043830, 304836, 304845,PNM 9283–84 (formerly KU 304847 and304849), adult females, collected by RMB,CO, Jason Fernandez and Boying Fernandez;same date, locality, and circumstances ofcapture (19:45–22:45 h) as holotype.
Other paratypes.—PNM 9095–96 (FieldNos. MGDP 110 and 113), adult females;and PNM 9097 (Field No. MGDP 111), adultmale, collected by CO and Marisol Pedigrosa,23 April 2004 at Ayumit, Municipality ofCalayan, Cagayan Province, Babuyan ClaroIsland (19.545u N, 121.958u E, 360 m); PNM9098, subadult male, collected by CO andMarisol Pedigrosa, 27 April 2004 at Corog,Municipality of Calayan, Cagayan Province,Babuyan Claro Island 19.489u N, 121.948u E,between sea level and 100 m; PNM 9090(Field No. MGDP 119), adult male, collectedby CO and Marisol Pedigrosa, 27 April 2004 atRakwaranom, Municipality of Calayan, Caga-yan Province, Babuyan Claro Island 19.561uN, 121.953u E, between sea level and 100 m.
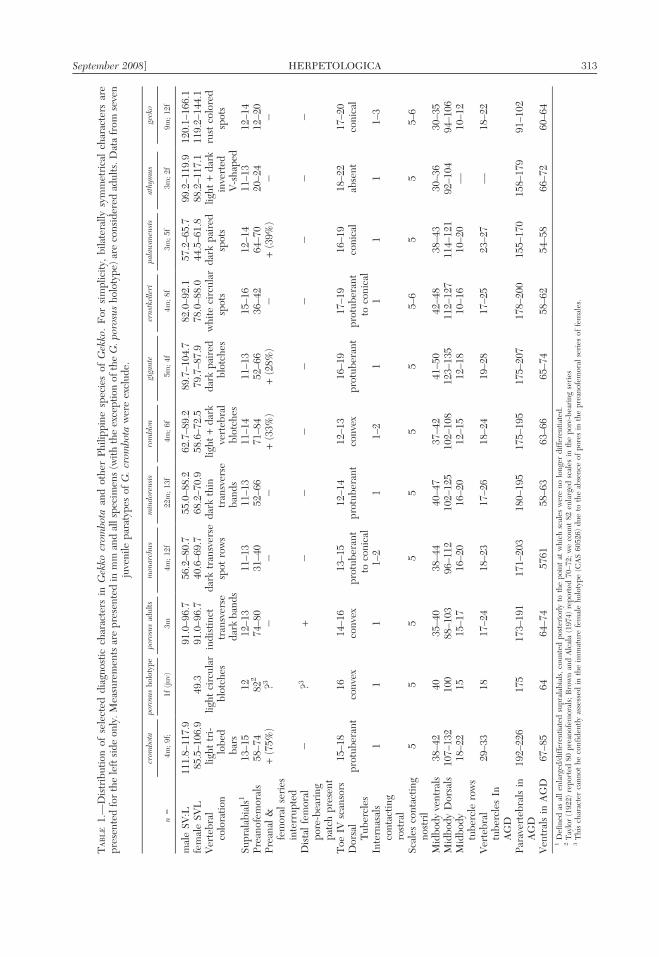
Diagnosis.—Gekko crombota differs fromall other species of Philippine Gekko (i.e., G.athymus, G. ernstkelleri, G. gecko, G. gigante,G. mindorensis, G. monarchus, G. palawa-nensis, G. porosus, and G. romblon) in thefollowing combination of features (1) largebody size (SVL 85.5–117.9 for adult males;
306 HERPETOLOGICA [Vol. 64, No. 3
85.1–106.9 mm for females); (2) dorsummedium brown to gray, marked with creamcolored, transverse, tri-lobed bars with a blackanterior border; (3) high numbers of dorsalbody scales (107–132 transverse midbodyscales; 192–226 paravertebrals); (4) highnumbers of sharply conical dorsal bodytubercle rows (17–21 midbody; 27–33 para-vertebrally); (5) preanofemorals arranged in anon-continuous series (preanals and femoralpore-bearing scale series separated by one ortwo non-pored scales in 75% of specimens) of58–74 differentiated, slightly enlarged scales.
Comparison with similar species.—Gekkocrombota differs from its phenotypically most
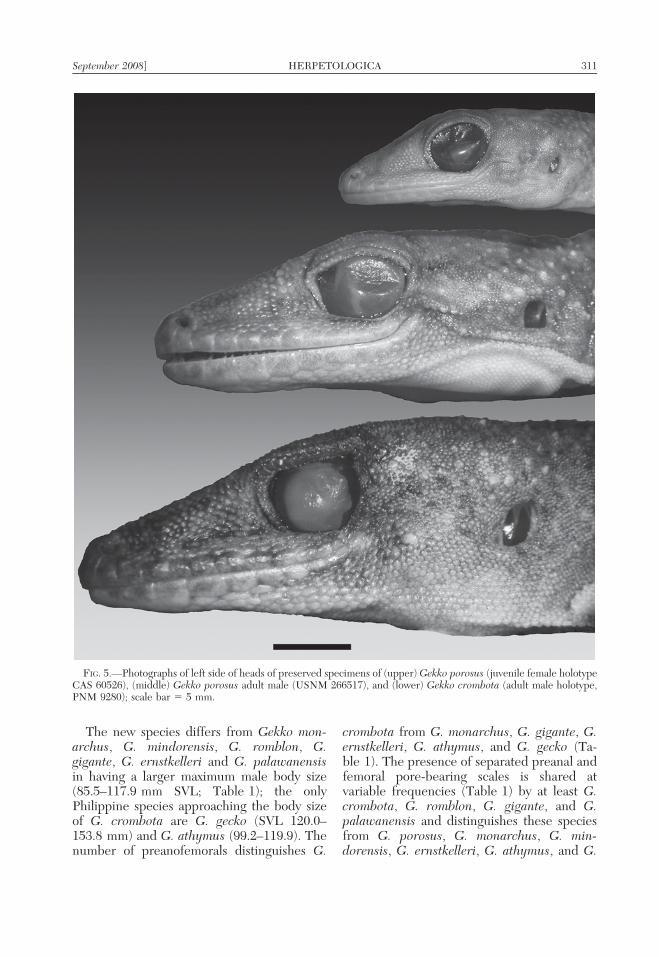
similar Philippine congener, Gekko porosus(Batan and Itbayat islands, of the BatanesIslands group, north of the Babuyans), by itsattainment of a larger maximum male bodysize (males: SVL 85.5–117.9 vs. 91.0–96.7 inG. porosus; Fig. 5); swollen (presumptivelyhypertrophied) adductor and temporal mus-culature of the head, leading to a wider,vaguely triangular head shape (Fig. 3B,C;versus adductor and temporal musculaturedistinctly less enlarged in G. porosus, headmore slender); brown to gray dorsal colora-tion, interrupted by distinct, tri-lobed, cream-colored bars (Fig. 2; versus indistinct darktransverse bands and/or circular vertebral
FIG. 1.—Map of the northern Philippines (inset) showing the type locality (1: Asked) of Gekko crombota on the SEcoast of Babuyan Claro Island (A). Additional collection localities include (2) Corog, (3) Rakwaranom, and (4) Ayumit.
September 2008] HERPETOLOGICA 307
blotches in G. porosus); a greater number(107–132) of transverse midbody dorsal scales(vs. 88–103 in G. porosus); a greater number(192–226) of paravertebral scales (vs. 173–191in G. porosus); the presence of sharplyprotuberant (vs. merely convex or slightlyraised) dorsal tubercles; a greater number(17–21) of midbody dorsal tubercle rows (vs.15–17 in G. porosus); and a greater number(27–33) of paravertebral tubercle rows (vs.
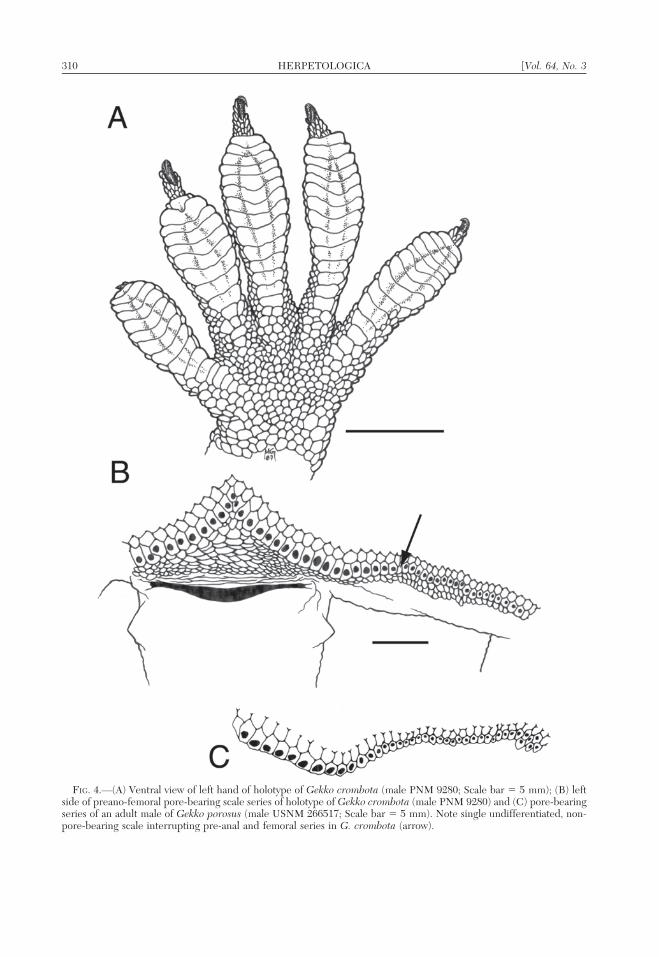
17–24); separation (1 or 2 undifferentiatedscales) between the preanal and femoral seriesof pore-bearing scales (in 75% of specimens;vs. absent in G. porosus); the presence offewer (58–74) preanofemoral pore-bearingscales (vs. 74–82 in G. porosus); and theabsence of a modified distal femoral pore-bearing patch (vs. present in G. porosus,composed of a short series of 2 or 3 rows ofpore-bearing scales (Fig. 4C).
FIG. 2.—Photographs of northern Philippine Gekko in life. (A, B) Adult female Gekko crombota (KU 304849; SVL 5111.8 mm); (C, D) juvenile G. crombota of undetermined sex (KU 304829; SVL 5 58.5 mm); (E) Adult presumablymale Gekko porosus and (F) juvenile G. mindorensis of undetermined sex; both photographed (specimens not collected)in Crystal Cave, Batan Island, 2006, by RMB.
308 HERPETOLOGICA [Vol. 64, No. 3
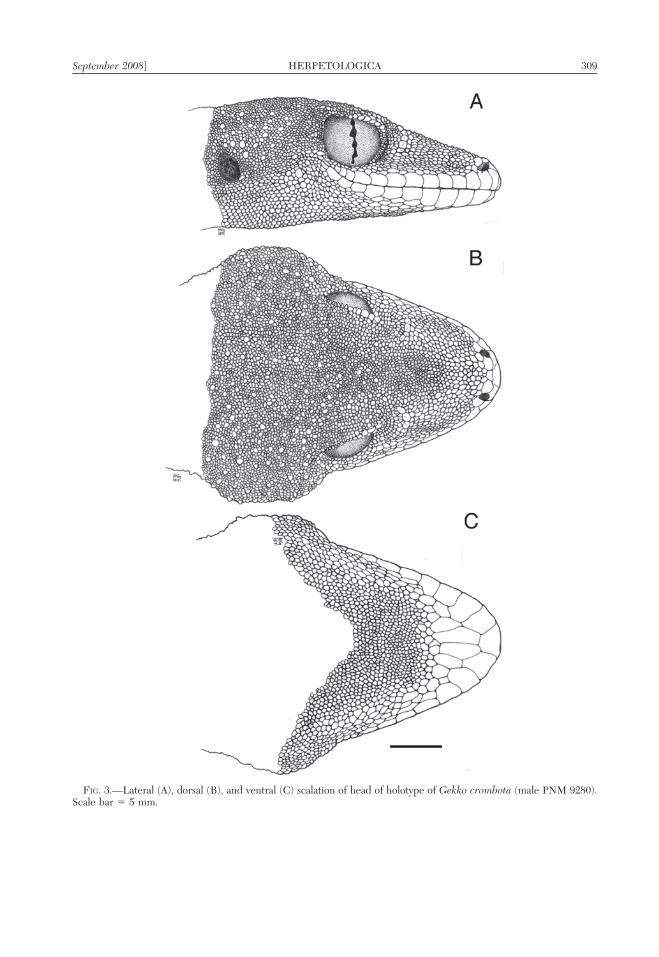
FIG. 3.—Lateral (A), dorsal (B), and ventral (C) scalation of head of holotype of Gekko crombota (male PNM 9280).Scale bar 5 5 mm.
September 2008] HERPETOLOGICA 309
FIG. 4.—(A) Ventral view of left hand of holotype of Gekko crombota (male PNM 9280; Scale bar 5 5 mm); (B) leftside of preano-femoral pore-bearing scale series of holotype of Gekko crombota (male PNM 9280) and (C) pore-bearingseries of an adult male of Gekko porosus (male USNM 266517; Scale bar 5 5 mm). Note single undifferentiated, non-pore-bearing scale interrupting pre-anal and femoral series in G. crombota (arrow).
310 HERPETOLOGICA [Vol. 64, No. 3
The new species differs from Gekko mon-archus, G. mindorensis, G. romblon, G.gigante, G. ernstkelleri and G. palawanensisin having a larger maximum male body size(85.5–117.9 mm SVL; Table 1); the onlyPhilippine species approaching the body sizeof G. crombota are G. gecko (SVL 120.0–153.8 mm) and G. athymus (99.2–119.9). Thenumber of preanofemorals distinguishes G.
crombota from G. monarchus, G. gigante, G.ernstkelleri, G. athymus, and G. gecko (Ta-ble 1). The presence of separated preanal andfemoral pore-bearing scales is shared atvariable frequencies (Table 1) by at least G.crombota, G. romblon, G. gigante, and G.palawanensis and distinguishes these speciesfrom G. porosus, G. monarchus, G. min-dorensis, G. ernstkelleri, G. athymus, and G.
FIG. 5.—Photographs of left side of heads of preserved specimens of (upper) Gekko porosus (juvenile female holotypeCAS 60526), (middle) Gekko porosus adult male (USNM 266517), and (lower) Gekko crombota (adult male holotype,PNM 9280); scale bar 5 5 mm.
September 2008] HERPETOLOGICA 311
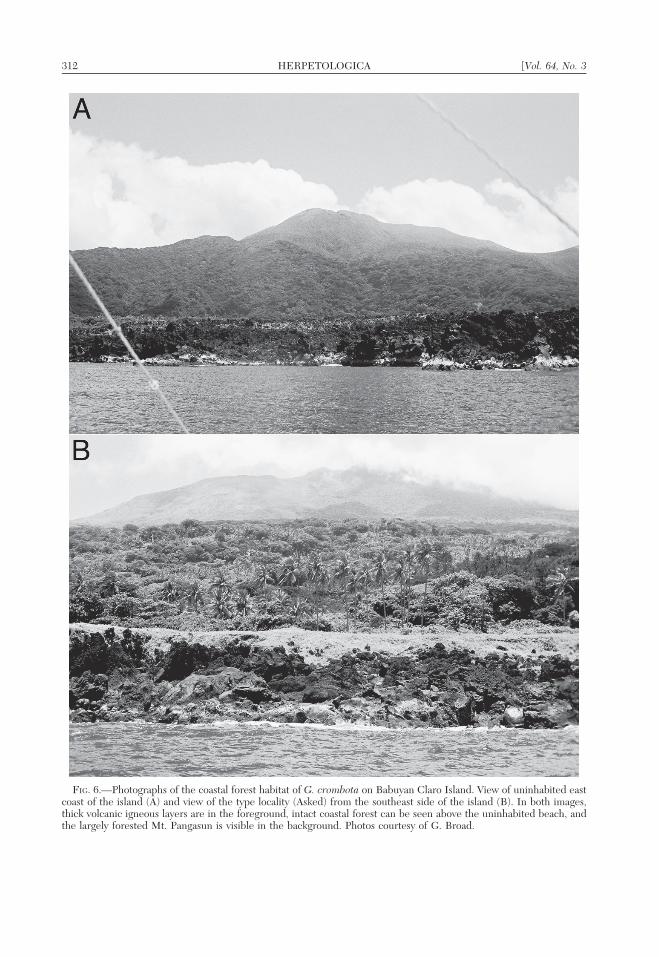
FIG. 6.—Photographs of the coastal forest habitat of G. crombota on Babuyan Claro Island. View of uninhabited eastcoast of the island (A) and view of the type locality (Asked) from the southeast side of the island (B). In both images,thick volcanic igneous layers are in the foreground, intact coastal forest can be seen above the uninhabited beach, andthe largely forested Mt. Pangasun is visible in the background. Photos courtesy of G. Broad.
312 HERPETOLOGICA [Vol. 64, No. 3
TA
BL
E1.
—D
istr
ibu
tion
ofse
lect
edd
iagn
osti
cch
arac
ters
inG
ekko
crom
bota
and
oth
erP
hili
pp
ine
spec
ies
ofG
ekko
.F
orsi
mp
licit
y,b
ilate
rally
sym
met
rica
lch
arac
ters
are
pre
sen
ted
for
the
left
sid
eon
ly.
Mea
sure
men
tsar
ep
rese
nte
din
mm
and
all
spec
imen
s(w
ith
the
exce
pti
onof
the
G.p
oros
ush
olot
ype)
are
con
sid
ered
adu
lts.
Dat
afr
omse
ven
juve
nile
par
atyp
esof
G.
crom
bota
wer
eex
clu
de.
n5
crom
bota
poro
sus
hol
otyp
epo
rosu
sad
ult
sm
onar
chus
min
dore
nsis
rom
blon
giga
nte
erns
tkel
leri
pala
wan
ensi
sat
hym
usge
cko
4m;
9f;
1f(j
uv)
3m4m
;12
f22
m;
13f
4m;
6f5m
;4f
4m;
8f3m
;5f
3m;
2f9m
;12
f
mal
eS
V:L
111.
8–11
7.9
91.0
–96.
756
.2–8
0.7
55.0
–88.
262
.7–8
9.2
89.7
–104
.782
.0–9
2.1
57.2
–65.
799
.2–1
19.9
120.
1–16
6.1
fem
ale
SV
L85
.5–1
06.9
49.3
91.0
–96.
740
.6–6
9.7
68.2
–70.
958
.6–7
2.5
79.7
–87.
978
.0–8
8.0
44.5
–61.
888
.2–1
17.1
119.
2–14
4.1
Ver
teb
ral
colo
rati
onlig
ht
tri-
lob
edb
ars
ligh
tci
rcu
lar
blo
tch
esin
dis
tin
cttr
ansv
erse
dar
kb
and
sdar
ktr
ansv
erse
spot
row
sd
ark
thin
tran
sver
seb
and
s
ligh
t+
dar
kve
rteb
ral
blo
tch
es
dar
kp
aire
db
lotc
hes
wh
ite
circ
ula
rsp
ots
dar
kp
aire
dsp
ots
ligh
t+
dar
kin
vert
edV
-sh
aped
rust
colo
red
spot
s
Su
pra
lab
ials
113
–15
1212
–13
11–1
311
–13
11–1
411
–13
15–1
612
–14
11–1
312
–14
Pre
anof
emor
als
58–7
482
274
–80
31–4
052
–66
71–8
452
–66
36–4
264
–70
20–2
412
–20
Pre
anal
&fe
mor
alse
ries
inte
rru
pte
d
+(7
5%)
?32
22
+(3
3%)
+(2
8%)
2+
(39%
)2
2
Dis
tal
fem
oral
por
e-b
eari
ng
pat
chp
rese
nt
2?3
+2
22
22
22
2
Toe
IVsc
anso
rs15
–18
1614
–16
13–1
512
–14
12–1
316
–19
17–1
916
–19
18–2
217
–20
Dor
sal
Tu
ber
cles
pro
tub
eran
tco
nve
xco
nve
xp
rotu
ber
ant
toco
nic
alp
rotu
ber
ant
con
vex
pro
tub
eran
tp
rotu
ber
ant
toco
nic
alco
nic
alab
sen
tco
nic
al
Inte
rnas
als
con
tact
ing
rost
ral
11
11–
21
1–2
11
11
1–3
Sca
les
con
tact
ing
nos
tril
55
55
55
55–
65
55–
6
Mid
bod
yve
ntr
als
38–4
240
35–4
038
–44
40–4
737
–42
41–5
042
–48
38–4
330
–36
30–3
5M
idb
ody
Dor
sals
107–
132
100
88–1
0396
–112
102–
125
102–
108
123–
135
112–
127
114–
121
92–1
0494
–106
Mid
bod
ytu
ber
cle
row
s18
–22
1515
–17
16–2
016
–20
12–1
512
–18
10–1
610
–20
—10
–12
Ver
teb
ral
tub
ercl
esIn
AG
D
29–3
318
17–2
418
–23
17–2
618
–24
19–2
817
–25
23–2
7—
18–2
2
Par
aver
teb
rals
inA
GD
192–
226
175
173–
191
171–
203
180–
195
175–
195
175–
207
178–
200
155–
170
158–
179
91–1
02
Ven
tral
sin
AG
D67
–85
6464
–74
5761
58–6
363
–66
65–7
458
–62
54–5
866
–72
60–6
41
Def
ined
asal
len
larg
ed/d
iffe
ren
tiat
edsu
pra
lab
ials
,co
un
ted
pos
teri
orly
toth
ep
oin
tat
wh
ich
scal
esw
ere
no
lon
ger
dif
fere
nti
ated
.2
Tay
lor
(192
2)re
por
ted
80p
rean
ofem
oral
s;B
row
nan
dA
lcal
a(1
974)
rep
orte
d70
–72;
we
cou
nt
82en
larg
edsc
ales
inth
ep
ore-
bea
rin
gse
ries
3T
his
char
acte
rca
nn
otb
eco
nfi
den
tly
asse
ssed
inth
eim
mat
ure
fem
ale
hol
otyp
e(C
AS
6052
6)d
ue
toth
eab
sen
ceof
por
esin
the
pre
anof
emor
alse
ries
offe
mal
es.
September 2008] HERPETOLOGICA 313
gecko. The new species is further distin-guished from G. athymus by the presence ofdorsal body tubercles (absent from G. athymus)and from G. porosus and G. romblon by thepresence of sharply conical dorsal tubercles (vs.tubercles only slightly convex in these species).Finally, the presence of distinct transverse lightcream-colored tri-lobed bars across the dorsumappears to distinguish G. crombota from thehighly variable suite of color patterns exhibitedby other Philippine species of Gekko. These andother differences among Philippine Gekkospecies are summarized in Table 1.
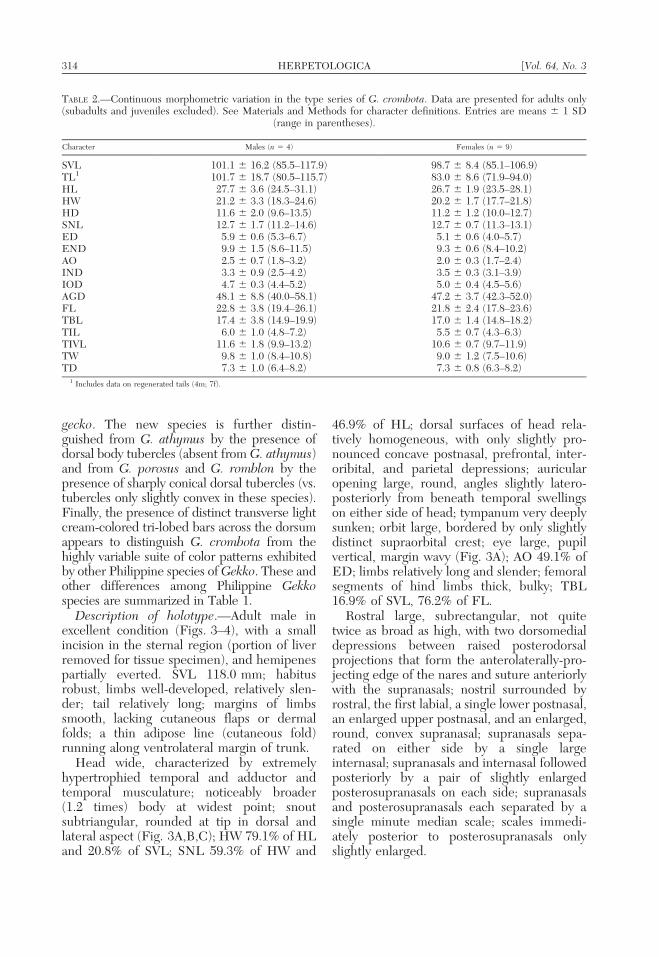
Description of holotype.—Adult male inexcellent condition (Figs. 3–4), with a smallincision in the sternal region (portion of liverremoved for tissue specimen), and hemipenespartially everted. SVL 118.0 mm; habitusrobust, limbs well-developed, relatively slen-der; tail relatively long; margins of limbssmooth, lacking cutaneous flaps or dermalfolds; a thin adipose line (cutaneous fold)running along ventrolateral margin of trunk.
Head wide, characterized by extremelyhypertrophied temporal and adductor andtemporal musculature; noticeably broader(1.2 times) body at widest point; snoutsubtriangular, rounded at tip in dorsal andlateral aspect (Fig. 3A,B,C); HW 79.1% of HLand 20.8% of SVL; SNL 59.3% of HW and
46.9% of HL; dorsal surfaces of head rela-tively homogeneous, with only slightly pro-nounced concave postnasal, prefrontal, inter-oribital, and parietal depressions; auricularopening large, round, angles slightly latero-posteriorly from beneath temporal swellingson either side of head; tympanum very deeplysunken; orbit large, bordered by only slightlydistinct supraorbital crest; eye large, pupilvertical, margin wavy (Fig. 3A); AO 49.1% ofED; limbs relatively long and slender; femoralsegments of hind limbs thick, bulky; TBL16.9% of SVL, 76.2% of FL.
Rostral large, subrectangular, not quitetwice as broad as high, with two dorsomedialdepressions between raised posterodorsalprojections that form the anterolaterally-pro-jecting edge of the nares and suture anteriorlywith the supranasals; nostril surrounded byrostral, the first labial, a single lower postnasal,an enlarged upper postnasal, and an enlarged,round, convex supranasal; supranasals sepa-rated on either side by a single largeinternasal; supranasals and internasal followedposteriorly by a pair of slightly enlargedposterosupranasals on each side; supranasalsand posterosupranasals each separated by asingle minute median scale; scales immedi-ately posterior to posterosupranasals onlyslightly enlarged.
TABLE 2.—Continuous morphometric variation in the type series of G. crombota. Data are presented for adults only(subadults and juveniles excluded). See Materials and Methods for character definitions. Entries are means 6 1 SD
Total number of differentiated supralabials12/12 (L/R; 9/8–9 to center of eye), bordereddorsally by one row of slightly differentiatedsnout scales; total number of differentiatedinfralabials 11/11 (8–9/9 to center of eye),bordered ventrally by one row of enlargedscales and 3 rows of only slightly differentiatedchin scales; mental triangular; mental and firstthree infralabials on left (first four on right)greatly enlarged and wrapping onto ventralsurfaces of chin, nearly twice the size ofindividual infralabials 4–11; mental followedby a pair of slender, elongate postmentals;postmentals bordered posterolaterally by asecondary pair, approximately one half thelength of first pair, and a tertiary pair of lateralpostmentals, one quarter the length of prima-ry postmentals; postmental scale series bor-dered posteriorly by a single series of slightlyenlarged scales; followed immediately by asharp transition to nondifferentiated chin andgular scales; postrictal scales slightly enlarged,2 or 3 times the size of gular scales; remainderof undifferentiated gular scales very small,round, nonimbricate, juxtaposed (Fig. 3C).
Dorsal cephalic scales highly heterogeneousand varied in shape, disposition, and distribu-tion; scales of rostrum enlarged, round, oval tosubrectangular, and convex to conical; post-nasal, prefrontal, and interorbital depressionspossess noticeably smaller scales; palpebralscales heterogeneous, with some scales assmall as adjacent interorbital region andothers as large and raised as rostral scales;undifferentiated posterior head scales granu-lar, flat to irregularly convex, reducing in sizeposteriorly, interspersed with numerousslightly enlarged conical tubercules, andmaking a smooth transition through nuchalregion to small, juxtaposed, flat trunk scalesinterspersed with enlarged, sharply conicalbody tubercles; throat and chin scales small,juxtaposed, nonimbricate; gular and pectoralregions with enlarged cycloid, imbricatescales, continuing to increase in size throughventral abdomen, becoming very enlarged andstrongly imbricate.
Ornamental cephalic scalation includesnumerous conical tubercles on posterolateralportions of head (temporal, supratympanic,and postrictal regions) and a short curvedseries of 4–5 enlarged, sharply conical (with
blade-like ridges) preorbital scales (Fig. 3A,B); 38/42 circumorbitals in total, differentiat-ed into the following distinct regions: (1)minute precircumorbitals, (2) enlarged, flat,squarish circumoribtals dorsoanterior to orbit(7 on left, 8 on right), (3) transverse elongationand modification into fringe-like points (spinyciliaria, 11 on left, 9 on right) acrossdorsoposterior margin of orbit, graduallyreducing to (4) minute postcircumorbitals; atotal of 35 interorbital scales (straight linedistance from center of each eye, across botheyelids).
Axilla–groin distance 49.2% of SVL; undif-ferentiated dorsal body scales round toirregularly octagonal, nonimbricate, relativelyhomogeneous in size, convex; dorsals sharplytransition to imbricate ventrals along theventrolateral adipose fold; dorsals lack sur-rounding interstitial granules but are inter-spersed with 18 irregularly transverse rows (33paravertebral rows) of highly enlarged andprotuberant, to strongly conical dorsal bodytubercles; each dorsal tubercle with a raised,thorn- to keel-like posteriorly-projectingpoint; each tubercle surrounded by a circleof very slightly enlarged adjacent dorsals; 109transverse midbody dorsals; 202 paraverte-brals between midpoints of limb insertions; 42transversely arranged ventrals; scales ondorsal surfaces of limbs larger than dorsals,with interspersed enlarged tubercles extend-ing down limbs and terminating at the dorsalsurfaces of hands and feet; enlarged patchesof distinct imbricate scales present on wrist,anterior (preaxial) surface of upper arm andthigh, on knee, and on distal ventral surface ofhind limb; scales on dorsal surfaces of handsand feet similar to dorsal limb scales (butlacking tubercles); ventral body scales flat,cycloid, strongly imbricate, much larger thanlateral or dorsal body scales, largest at mid-ventral line.
Seventy-four dimpled pore-bearing scales(Fig. 4B) in a near-continuous preanofemoralseries (18 preanals on left, 19 on right;separated from femorals by a single undiffer-entiated scale; 19 femorals on left; 18 on right)each punctured by pore bearing dark orange,exudate, arranged in a wavy, widely obtuse,inverted ‘‘V’’ formation and continuing to justbefore the patellar region; preanal pores 2–3
September 2008] HERPETOLOGICA 315
times the diameter of femoral pores; preanalssituated atop a substantial preanal bulge thatfolds over into precloacal region in preservedspecimen but was erect and protuberant inlife; preanals preceded by five similarlyenlarged but non-dimpled scale rows; pre-anals followed by five enlarged scales rows,roughly forming a triangular patch of scalesbefore vent; femoral series lacks preceding orfollowing enlarged scale rows; scales latero-posterior to preanofemoral series (i.e., alongventroposterior surfaces of hind limb) reducein size sharply to minute scales of theposterior edge of the hind limb.
Digits moderately expanded and covered onpalmar/plantar surfaces by bowed, unnotched,undivided scansors (Fig. 4A); digits lack in-terdigital webbing; subdigital scansors ofmanus: 11/12, 10/11, 14/15, 14/16, and 13/12on left/right digits I–V respectively; pes: 13/12,11/12, 16/15, 15/15, and 15/14 on left/rightdigits I–V respectively; subdigital scansors ofmanus and pes bordered proximally (onpalmar and plantar surfaces) by 1–4 slightlyenlarged scales that form a near-continuousseries with enlarged scansors; all digits clawed,but first (inner) claw greatly reduced; remain-ing terminal claw-bearing phalanges com-pressed, with large recurved claws, not risingfree at distal end until they extend beyonddilated hyperextensible portion of digit.
Tail base bordered by a single, greatlyenlarged conical postcloacal spur on each sideof vent; postcloacal swellings pronounced;hemipenes incompletely everted, their distalstructures not evident; tail long, 98.0% ofSVL; tail not depressed, subcylindrical, divid-ed into distinct fracture planes/autotomygrooves (5 whorls or annulations); distal tailportions not original; tail with clear autotomyscar and distally regenerated portion; 11caudal annulations before autotomy scar(22.2 mm), 12–13 annulations estimated inautotomized portion based on length(47.1 mm), for an estimated total possibleannulation count of 25–26; TD (not includingbasal postcloacal swelling) 75.9% of TW;dorsal tail (following description based onoriginal portions of tail) as heavily adorned asdorsum with tubercles; caudal tuberclesconcentrated along posterior edge of caudalannulation; caudals similar in size to dorsals;
subcaudals enlarged, plate-like, 3–4 rows perannulations; subcausals widely expanded tocover majority of ventral surface of tail, or splitinto a pair of subcaudals along posteriormargin of each annulation.
Variation.—Ranges of selected diagnosticmeristic characters are presented in Table 1.Morphometric variation in mensural charac-ters of adult paratypes is presented in Table 2.The type series contains two large maturemales, two small but presumably maturemales (with swollen preanal buldges andhemipenal tail base swellings), nine maturefemales (all gravid, with large white oblongeggs visible through skin of posteroventralbody wall) and eight juveniles/subadults, fourof which are males, with hemipenes partiallyeverted.
Coloration of holotype in ethanol.—Dorsalground coloration of head, body, tail, anddorsal surfaces of limbs medium gray withscattered indistinct light gray blotches andblack flecks; light tri-lobed bars traverse thebody in nuchal region, above limb insertions,and across torso to base of tail; anteriormargin of tri-lobed light gray dorsal barsbordered by faintly darker pigmentation;posterior edge of light gray transverse barscolored as dorsal trunk ground coloration.
Dorsal and lateral surfaces of head similarto dorsal ground coloration, but with distinctlight gray spots; a light cream bar extendsposteriorly from the orbit; palpebra dark gray;rostral and supralabials medium gray; infra-labials very light gray.
Limbs colored as torso, lacking transversebanding; dorsal surfaces of hands and feetlight gray; digits light gray with dark graysurfaces of expanded distal portions; tailmedium gray with dark gray bands corre-sponding to two caudal annuli; regeneratedportion of tail flat gray.
Ventral head, neck, and torso light creamventral surfaces of limbs slightly darker;ventral surfaces of digits (scansors) dark gray;preanofemoal region white with orange poreexudate; ventral surfaces of tail medium gray(both original and regenerated portions).
Coloration of holotype in life.—(from pho-tographs of holotype before preservation;Fig. 2A–D) Dorsal ground coloration darkpurplish-gray to yellowish-brown, with indis-
316 HERPETOLOGICA [Vol. 64, No. 3
tinct dark gray to black blotches; dorsum withsix light cream tri-lobed bars traversing theaxilla-groin region, each bordered anteriorlyby an accompanying thin transverse wavyblack band (the darkest above insertion ofhind limbs) and posteriorly by normal trunkground coloration.
Dorsal nuchal region and posterior portionsof head very similar to trunk coloration butwith distinct, round, cream-colored spots;similar cream bars radiate out from the orbit;postrictal region flat gray; labial scales pur-plish-gray with cream spots on every thirdlabial scale; darker black blotches and fleckscongregate on snout, interorbital region, andparietal region; infralabial region and chingray to light gray; snout scale purplish gray;gular region light gray to brownish-tan.
Dorsal surfaces of limbs light gray withnumerous dark brown flecks; dorsal surfacesof digits dark gray with slightly lighter claws;dorsal and lateral portions of tail bandedalternating dark gray and cream (correspond-ing to tail annuli); distal autotomy regrowthdark brown.
Ventral body and limbs yellow with scat-tered gray and dark brown flecks; preano-femoral region bright yellow with dark orangepores; palmar and plantar surfaces of manusand pes yellowish with light gray subdigitalscansors; ventral tail cream with browntransverse bars, and solid brown ventralcoloration distal to autotomy scar and subse-quent regrowth.
Color variation.—Our sample of four adultmales, a subadult male, two juvenile males,nine adult females, and five juveniles ofundetermined sex exhibits moderate colorvariation. The adult male paratype (KU304825) and two female paratypes (KU304809, 304836) have darker dorsal colorationthan that of the holotype, with very dark grayanterior margins of the light transverse bars.Two females (KU 304826, 3004849) havemuch darker dorsal ground coloration thando any of the other specimens. In thesespecimens, the tri-lobed light gray transversebars take on a more striking, contrastingappearance. Three adult female paratypes(KU 304845, 304847, 304851) have a light tomedium gray, nearly patterenless dorsum,with only faint vestiges of transverse banding.
Juveniles are patterned more brightly thanadults, with more intensely contrasting lightand dark dorsal and tail coloration. In somejuveniles (KU 304821) the tail bandingcontrast is nearly extreme black and white.Juveniles (KU 304814, 304821, 304829304832) also possess cream spots on thedorsal surfaces of articulations between adja-cent phalanges. This gives the fingers and toesa light and dark banded appearance. Thispattern is faint in the subadult paratype (KU304807) and absent in all adults.
Ventral coloration is nearly invariant, withthe exception that some specimens have verydark gray to black ventral tail surfaces (e.g.,KU 304809, 304826, 304849) while othershave very light gray ventral tail surfaces thatare only slightly darker than ventral bodycoloration (e.g., KU 304845, 304847). Only intwo juveniles is the black and white transversebanded caudal pigmentation strong enough towrap around on to the ventral surface of thetail (KU 304814, 304821).
Distribution and natural history.—The newspecies is known only from Babuyan ClaroIsland where it was collected low (,3 m) ontrunks and buttresses in primary dipterocarpforest at low elevations, close to the island’scoast. Given the island’s volcanic origin andgeological history of isolation (Marini et al.,2005; McDermott et al., 1993) we do notexpect G. crombota to be distributed on otherislands in the Babuyan Island group, theBatanes Island group, or mainland Luzon.Our impression is that the new species is verycommon at the type locality. No vocalizationsby this species were heard during our briefvisits to the type locality. The only othergekkonids encountered during our three nightstay on Babuyan Claro were Hemidactylusfrenatus, Cyrtodactylus philippinicus, andLuperosaurus macgregori (see Discussion inBrown et al., 2007).
Etymology.—The specific epithet crombotais a group of letters derived from the namesCrombie and Ota, and treated as a noun inapposition. We employ the amalgam crombotato jointly honor Ronald Crombie and Hide-toshi Ota in recognition of their collaborativesurvey efforts in the Babuyan Islands (Ota andCrombie, 1989; Ota and Ross, 1994) and theircontinued work on the gekkonid fauna of this
September 2008] HERPETOLOGICA 317
archipelago (H. Ota and R. Crombie, unpub-lished data). Suggested common name: Ba-buyan Claro Gecko.
DISCUSSION
The description of Gekko crombota bringsthe total number of endemic Philippine Gekkospecies to eight (ten total, when G. hokouensisis excluded and the nonendemic G. mon-archus and G. gecko are included). We arecertain that this number represents an under-estimate of species diversity and we enumer-ate the following series of unresolved taxo-nomic issues that need to be addressed beforea full realization of Philippine gekkoniddiversity can be achieved. First, the Babuyansand Batanes island groups require additionalsurvey work before we can be reasonablycertain that the total gekkonid fauna is known.We are aware of at least four additional,undescribed Gekko species in the Babuyansalone and we suspect that several additionalundescribed species in the Babuyans andBatanes await discovery on small isolatedislands surrounded by deep water. Second,although morphologically distinct species arestill being described from other parts of thearchipelago (Roesler et al., 2006), the majorityof the Philippines’ more subtle or possiblycryptic species diversity undoubtedly mas-querades under the widespread species Gekkomindorensis and Gekko monarchus. Prelimi-nary molecular sequence data (C. Siler, A.Diesmos, and R. Brown, unpublished data)suggest that these taxa may be comprised ofnumerous cryptic, unrecognized evolutionarylineages, and potentially are worthy of specificrank. At a minimum, the major PleistoceneAggregate Island Complexes of the Philip-pines (Brown and Diesmos, 2002; Brown etal., 2007; Gaulke et al., 2007) might each beexpected to harbor endemic species diversityin both of these species complexes. Third,numerous deep water islands (e.g., notconnected to adjacent islands during the lastglaciations; Brown and Diesmos, 2002) havenot been exhaustively surveyed for herpeto-fauna and are thus ripe for the potentialdiscovery of additional Gekko species. Islandsis this category are are Lubang, CamiguinNorte, Camiguin Sur, Calayan, Dalupiri,Fuga, Masbate, Siquijor, Dinagat, Siargao,
Sarangani, Maranduque, Coron, Busuanga,Burias, Ticao, Semira, Semirara, Maestre deCampo, Cuyo, Basilan, Jolo, Tawi-Tawi, andmany other similarly small, isolated landmass-es. Fourth, it is clear that comprehensiveefforts to survey gekkonid fauna of thePhilippines would do well to target isolatedlimestone karst areas and natural caves. Suchhabitats are not only patchily distributed (andthus can be expected to have promotedevolutionary divergence via isolation) but arealso heavily imperiled and increasingly underthreat from overexploitation by humans for avariety of natural resources (Clements et al.,2006). The discovery of G. ernstkelleri in anisolated limestone outcop on Panay Island (C.D. Siler, personal observations; Roesler et al.,2006) demonstrates that hidden gekkoniddiversity awaits field herpetologists willing totarget long-overlookecd limestone habitats.Similarly, G. gigante (endemic to the lime-stone landbridge Gigante island group; W.Brown and Alcala, 1978; R. Brown and Alcala,2000) is proof that geological isolation (e.g.,landmasses separated by deep water) may notbe necessary to promote gekkonid diversifica-tion if limestone habitats have been isolatedover geological timescales.
Finally, numerous isolated mountain rangeson larger islands (Luzon, Palawan, Samar-Leyte, Mindanao) can be expected to supportas yet undocumented gekkonid diversity.Recent discoveries from the Sierra Madremountain range of Luzon (Brown et al.,2000b, 2007; A. Diesmos, unpublished data)suggest that these remaining forested regionsall warrant extensive biodiversity surveys.
Whatever the circumstances, we are certainthat the diversity of Philippine Gekko speciesis substantially underestimated. Given theundeniable fact that destructive exploitationof Southeast Asian forests is partly related toan ignorance of their biodiversity, it is crucialthat future faunal inventories throughout thePhilippines pay careful attention to habitatsknown to harbor endemic gekkonids. Wewould not be surprised if the eventual numberof known endemic Philippine Gekko specieswere to double in the very near future.
Acknowledgments.—For the loans of specimens orassistance while visiting museum collections, we thankthe following individuals and their respective institutions
318 HERPETOLOGICA [Vol. 64, No. 3
(museum abbreviations follow Leviton et al., 1985): A.Resetar, M. Kearney, and H. Voris (FMNH); J. Vindum,R. Drewes, and A. Leviton (CAS); J. Simmons and L.Trueb (KU), A. Wynn, R. Wilson, R. Heyer, and K. deQueiroz (USNM) and R. Sison (PNM). Support forfieldwork was provided by the University of Kansas, andthe National Science Foundation. The Stearns Fellowshipof the California Academy of Sciences provided supportthat allowed RMB and ACD to undertake multiple visitsto CAS. We thank the Department of the Environmentand Natural Resources, and the Protected Areas andWildlife Bureau (especially C. Custodio, T. M. Lim, andA. Tagtag), for facilitating research and export permits forthis and related studies, KU IACUC for approvingresearch protocols, and M. Pedregosa, and M. A. Reyes,N. Antoque, B. Fernandez, and J. Fernandez for untiringassistance in the field. We also thank Municipal DENRauthorities of Calayan, Cagayan Province for logisticalsupport. Thanks are due to M. Garfield for scientificillustration and J. Weghorst, A. Bauer, and an anonymousreviewer for comments on previous versions of themanuscript.
LITERATURE CITED
BROWN, R. M. 1999. New species of parachute gecko(Squamata; Gekkonidae; Genus Ptychozoon) fromnortheastern Thailand and Central Vietnam. Copeia1999:990–1001.
BROWN, R. M., AND A. C. ALCALA. 2000. Geckos, cavefrogs, and small land-bridge islands in the Visayan sea.Haring Ibon 2:19–22.
BROWN, R. M., AND A. C. DIESMOS. 2002. Application oflineage-based species concepts to oceanic island frogpopulations: the effects of differing taxonomic philos-ophies on the estimation of Philippine biodiversity. TheSilliman Journal 42:133–162.
BROWN, R. M., AND S. I. GUTTMAN. 2002. Phylogeneticsystematics of the Rana signata complex of Philippineand Bornean stream frogs: reconsideration of Huxley’smodification of Wallace’s Line at the Oriental-Austra-lian faunal zone interface. Biological Journal of theLinnean Society 76:393–461.
BROWN, R. M., A. C. DIESMOS, AND M. V. DUYA. 2007. Anew Luperosaurus (Squamata: Gekkonidae) from theSierra Madre of Luzon Island, Philippines. RafflesBulletin of Zoology 55:167–174.
BROWN, R. M., J. W. FERNER, AND A. C. DIESMOS. 1997.Definition of the Philippine Parachute Gecko, Ptycho-zoon intermedium Taylor 1915 (Reptilia: Lacertilia:Gekkonidae): redescription, designation of a neotype,and comparisons with related species. Herpetologica53:357–373.
BROWN, R. M., J. SUPRIATNA, AND H. OTA. 2000a. Discoveryof a new species of Luperosaurus (Squamata; Gekko-nidae) from Sulawesi, with a phylogenetic analysis ofthe genus and comments on the status of L. serrati-caudus. Copeia 2000:191–209.
BROWN, R. M., J. A. MCGUIRE, J. W. FERNER, N.ICARANGAL, JR., AND R. S. KENNEDY. 2000b. Amphibiansand reptiles of Luzon Island, II: Preliminary report onthe herpetofauna of Aurora Memorial National Park,Philippines. Hamadryad 25:175–195.
BROWN, W. C., AND A. C. ALCALA. 1978. Philippine Lizardsof the Family Gekkonidae. Silliman University Press,Dumaguete City, Philippines.
CLEMENTS, R., N. SODHI, M. SCHILTHUIZEN, AND P. K. L.NG. 2006. Limestone karsts of Southeast Asia: Imper-iled arks of biodiversity. Bioscience 56:733–742.
DE QUEIROZ, K. 1998. The general lineage concept ofspecies, species criteria, and the process of speciation.Pp. 57–75. In D. J. Howard and S. H. Berlocher (Eds.),Endless Forms: Species and Speciation. Oxford Uni-versity Press, New York, New York, U.S.A.
DE QUEIROZ, K. 1999. The general lineage concept ofspecies and the defining properties of the speciescategory. Pp. 49–89. In R. A. Wilson (Ed.), Species:New Interdisciplinary Essays. Massachusetts Instituteof Technology Press, Cambridge, Massachusetts,U.S.A.
GAULKE, M., H. ROESLER, AND R. M. BROWN. 2007. A newspecies of Luperosaurus (Squamata: Gekkonidae) fromPanay Island, Philippines, with comments on thetaxonomic status of Luperosaurus cumingii (Gray,1845). Copeia 2007:413–425.
LEVITON, A. E., R. H. GIBBS, JR., E. HEAL, AND C. E.DAWSON. 1985. Standards in herpetology and ichthyol-ogy: Part I. Standard symbolic codes for institutionalresource collections in herpetology and ichthyology.Copeia 1985:802–821.
MANTHEY, U., AND W. GROSSMAN. 1997. Amphibien undReptilien Sudostasiens. Natur und Tier-Verlag, Berlin,Germany.
MARINI, J.-C., C. CHAUVEL, AND R. C. MAURY. 2005. Hfisotope compositions of northern Luzon lavas suggestinvolvement of pelagic sediments in their source.Contributions to Mineralogy and Petrology149:216–232.
MCDERMOTT, F., M. J. DEFANT, C. J. HAWKESWORTH, R. C.MAURY, AND J. L. JORON. 1993. Isotope and traceelement for three component mixing in the genesis ofnorth Luzon lavas (Philippines). Contributions toMineralogy and Petrology 113:9–23.
OTA, H., AND R. I. CROMBIE. 1989. A new lizard of thegenus Lepidodactylus (Reptilia: Gekkonidae) fromBatan island, Philippines. Proceedings of the BiologicalSociety of Washington 102:559–567.
OTA, H., AND C. A. ROSS. 1994. Four new species ofLycodon (Serpentes: Colubridae) from the northernPhilippines. Copeia 1994:159–174.
OTA, H. K.-Y. LUE, S.-H. CHEN, AND W. C. BROWN. 1989.Taxonomic status of the Taiwanese Gekko, withcomments on the synonymy of Luperosaurus amissusTaylor. Journal of Herpetology 23:76–78.
ROESLER, H., C. D. SILER, R. M. BROWN, A. D.DEMEGILLO, AND M. GAULKE. 2006. Gekko ernstkellerisp. n.—a new gekkonid lizard from Panay Island,Philippines. Salamandra 42:197–211.
RUSSELL, A. P. 1979. A new species of Luperosaurus(Gekkonidae) with comments on the genus. Herpeto-logica 35:282–288.
TAYLOR, E. H. 1922a. The lizards of the Philippine Islands.Philippine Bureau of Science, Manila, Philippines.
TAYLOR, E. H. 1922b. Additions to the herpetologicalfauna of the Philippine Islands, I. Philippine Journal ofScience 21:161–206.
September 2008] HERPETOLOGICA 319
WERMUTH, H. 1965. Gekkonidae, Pygopodidae, Xantusi-dae. Pp. 154–155. In R. Mertens, W. Hennig, and H.Wermuth (Eds.), Das Tierreich, Vol. 80. Walter deGruyter and Co., Berlin, Germany.
WILEY, E. O. 1978. The evolutionary species conceptreconsidered. Systematic Zoology 21:17–26.
.Accepted: 3 June 2008
.Associate Editor: Christopher Raxworthy
APPENDIX I
Comparative Material Examined
All specimens examined are from the Philippines.Numbers in parentheses indicate the number of speci-mens examined for each species and museum abbrevia-tions follow Leviton et al. (1985).
Gekko athymus.—(7) PALAWAN ISLAND, PALAWAN
PROVINCE, ca. 10 km WSW of Iwahig: CAS 137677; ca. 8–9 km S. of Balico: CAS-SU 23119 (holotype); ca. 20 kmSW of Iwahig: CAS-SU 23121 (paratype); Municipality ofBrooke’s Point, Barangay Samarinana; Mt. Mantalingahan:KU 309331–34.
Gekko ernstkelleri.—(10) PANAY ISLAND, ANTIQUE
PROVINCE, Municipality of Pandan, Barangay Duyong,Duyong Hillside (5 ‘‘Mt. Lihidian’’), 300 m.a.sl.: PNM9152–54; KU 300196–202.
Gekko gecko.—(13) LUBANG ISLAND, OCCIDENTAL
MINDORO PROVINCE, Municipality of Lubang, BarangayParaiso: KU 303960–72.
Gekko gigante.—(8) SOUTH GIGANTE ISLAND,ILOILO PROVINCE, Municipality of Carles, Tantangan: CAS124315–17 (paratypes); NORTH GIGANTE ISLAND,
ILOILO PROVINCE, Municipality of Carles: CAS 124866–67(Paratypes); Barangay Asloman: KU 305138–40.


















![Gekko Covers EN 1008 - Suuntons.suunto.com/Manuals/Gekko/Userguides/Gekko_userguide_EN.pdf · DIVE SURF O2% CEILING ASC TIME QUIT ... altitudes greater than 300 m [1000 ft] ... 3000](https://static.documents.pub/doc/80x56/5b1de8a97f8b9a397f8b7bb8/gekko-covers-en-1008-dive-surf-o2-ceiling-asc-time-quit-altitudes-greater.jpg)







