A new specimen of Acrocanthosaurusatokensis (Theropoda, Dinosauria) from theLower Cretaceous Antlers Formation (LowerCretaceous, Aptian) of Oklahoma, USA
Philip J. CURRIERoyal Tyrrell Museum of Palaeontology, Box 7500, Drumheller,
Currie P. J. & Carpenter K. 2000. — A new specimen of Acrocanthosaurus atokensis(Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous,Aptian) of Oklahoma, USA. Geodiversitas 22 (2) : 207-246.
The data matrix is available at http://www.mnhn.fr/publication/matadd/g00n2a3.html
ABSTRACTA new skeleton of Acrocanthosaurus atokensis is the most complete specimencollected and has the only known complete skull. Aspects of the new skeletonare described in detail, with special attention directed to the morphology ofthe skull and forelimb. Although unquestionably one of the largest theropodsever found, it is smaller than Carcharodontosaurus, Giganotosaurus andTyrannosaurus. Comparison with other theropods suggests thatAcrocanthosaurus bears a strong resemblance to these taxa because of charac-ters that are size determinate, and the evidence suggests Acrocanthosaurus ismore closely related to Allosauridae than to Carcharodontosauridae. Threefamilies (Allosauridae, Carcharodontosauridae, Sinraptoridae) are recognizedin the Allosauroidea.
KEY WORDSDinosaurs, theropods,
Early Cretaceous, USA.
INTRODUCTION
Acrocanthosaurus atokensis was described byStovall & Langston (1950) on the basis of twopartial skeletons (one with part of a skull) fromthe Lower Cretaceous Antlers Formation (TrinityGroup, Aptian-Albian) of Atoka County,Oklahoma. The holotype is OMNH 10146(MUO 8-0-S9 of Stovall & Langston 1950) andthe paratype is OMNH 10147 (MUO 8-0-S8).Acrocanthosaurus Stovall & Langston, 1950 wasthe top predator of one of the richest faunasknown from Early Cretaceous times (Cifelli et al.1997). Although it seems to have been widelydistributed (Lipka 1998) in North America, itwas not the only large theropod of Aptian-Albiantimes (Harris 1998b).Another partial skull and skeleton ofAcrocanthosaurus was “discovered” in 1990 (thespecimen was actually noticed for the first timemore than 40 years earlier) near Weatherford,Texas in the Twin Mountain Formation (TrinityGroup), which correlates with the lower part ofthe Antlers Formation (Harris 1998a). It was col-lected by a crew from Southern MethodistUniversity. SMU 74646 includes about 70% of askeleton (Harris 1998a).
Parts of another specimen (two posterior cervicalor anterior dorsal centra, ischial fragment, distalend of a femur), described in this paper, were col-lected by the Oklahoma Museum of NaturalHistory and were catalogued as OMNH 10168.Most of the specimen, however, was collected byCephis Hall & Sid Love, who retrieved the skull,about two dozen vertebrae, ribs, chevrons, most ofthe front limbs, pelvic fragments, parts of bothfemora and tibiae, and most of the bones of thefoot. This, the most complete skeleton ofAcrocanthosaurus known, also comes from theAntlers Formation (Trinity Group) of McCurtainCounty, Oklahoma. The unprepared specimenwas eventually acquired by Allen Graffham ofGeological Enterprises, Inc., Ardmore, Oklahoma,who in turn arranged for the Black Hills Institutein Hill City, South Dakota, to prepare the speci-men. The preparation was completed at the end ofthe summer of 1996, and the original skeletonwent the following year to the North Carolina StateMuseum of Natural Sciences.The total length of the skeleton (Fig. 1) as mountedfor display by the Black Hills Institute is 11.5 m.This would make it one of the largest knowntheropods, comparable in total length withTyrannosaurus rex Osborn, 1905 (the mountedskeletons of BHI 3033 and MOR 555 are 11.5 m)
Currie P. J. & Carpenter K.
208 GEODIVERSITAS • 2000 • 22 (2)
RÉSUMÉUn nouveau specimen d’Acrocanthosaurus atokensis (Theropoda, Dinosauria)du Crétacé inférieur de la Formation Antlers (Crétacé inférieur, Aptien) del’Oklahoma, États-Unis.Nous décrivons le squelette d’Acrocanthosaurus atokensis le mieux conservéjusqu’ici et possédant un crâne complet. La morphologie des structures incon-nues sont décrites ; l’accent est mis sur la morphologie du crâne et du membreantérieur. Bien que ce théropode soit l’un des plus grands, ses dimensions restentinférieures à celles de Carcharodontosaurus, Giganotosaurus et Tyrannosaurus.Il présente un fort degré de ressemblance morphologique avec ces taxons,dont la nature serait liée à leur taille importante. Une analyse de parcimonieplace Acrocanthosaurus comme plus proche parent des Allosauridae que desCarcharodontosauridae. Trois familles sont identifiées au sein des Allosauroidea :les Allosauridae, les Carcharodontosauridae et les Sinraptoridae.
MOTS CLÉSDinosaures, théropodes,
Crétacé inférieur, États-Unis.
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
209GEODIVERSITAS • 2000 • 22 (2)
and Giganotosaurus Coria & Salgado, 1995 (themounted skeleton of MUCPv-CH1 is 12.2 m).We have not attempted to describe the entireskeleton of Acrocanthosaurus in this paper, buthave focused on new information available fromNCSM 14345. This is especially true for theskull, pectoral girdle and forelimb and, to a lesserextent, the hindlimb. The reader is referred topapers by Stovall & Langston (1950) and Harris(1998a) for complementary information on thepalate, braincase, vertebral column and pelvis. Amore detailed description of the braincase of theholotype has been prepared by Welles et al.Acrocanthosaurus (Fig. 1) was assigned to theAllosauridae by Stovall & Langston (1950). Thiswas generally accepted by other authors until itwas reassigned to the Carcharodontosauridae(Sereno et al. 1996). A more thorough analysis byHarris (1998a) supported the inclusion ofAcrocanthosaurus within the Carcharodonto-sauridae. Information from the new specimenwill be used in this paper to re-examine the rela-tionships of Acrocanthosaurus.
ABBREVIATIONS
BYU Brigham Young University, Provo;FMNH Field Museum of Natural History,
Chicago;
GI Paleontological Museum of theGeological Institute of Mongolia,Ulaan Baatar;
MACN-CH Museo Argentino de CienciasNaturales “Bernardino Rivadavia”,Buenos Aires;
MOR Museum of the Rockies, Bozeman;MUCPv-CH Museo de la Universidad Nacional del
Comahue, El Chocón collection,Neuquén, Argentina;
NCSM North Carolina State Museum ofNatural Sciences, Raleigh;
NMMNH New Mexico Museum of NaturalHistory, Albuquerque;
OMNH Oklahoma Museum of NaturalHistory, University of Oklahoma,Norman;
PIN Paleontological Institute of the RussianAcademy of Sciences, Moscow;
SMU Southern Methodist University,Dallas;
TMP Royal Tyrrell Museum of Palaeon-tology, Drumheller;
USNM United States National Museum ofNatural History, Smithsonian Ins-titution, Washington;
UUVP University of Utah, VertebratePaleontology Collection, Salt LakeCity.
FIG. 1. — Acrocanthosaurus atokensis. Reconstruction of skeleton. Scale bar: 1 m.
MATERIAL. — North Carolina State Museum ofNatural Sciences, NCSM 14345, nearly completeskull and most of postcranium.
LOCALITY AND AGE. — NCSM 14345 was recoveredfrom the Antlers Formation (Trinity Group, LowerCretaceous) of McCurtain County, Oklahoma.
FIG. 2. — Acrocanthosaurus atokensis NCSM 14345; A, skull in left view; B, skull in right view. Scale bar: 10 cm.
A
B
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
211GEODIVERSITAS • 2000 • 22 (2)
REVISED DIAGNOSIS. — Large theropod with elongateneural spines that are more than 2.5 times correspon-ding presacral, sacral and proximal caudal lengths ofthe centra. Lacrimal contacts postorbital; supraoccipi-tal expands on either side of the midline to protrude asa double boss behind the nuchal crest; pleurocoelousfossae and foramina pronounced on all presacral andsacral vertebrae; cervical neural spines have triangularanterior processes that insert into depressions beneathoverhanging processes on preceding neural spines;accessory transverse processes on mid-caudal vertebrae.
DESCRIPTION
NCSM 14345 was found lying on its side, par-tially disarticulated. There has been some crush-
ing and distortion of the skull, mostly towardsthe back (Fig. 2). For example, the left postor-bital was broken in at least two places, and hadpulled away from its suture with the frontal, theventral part rotating medially. Minor crushingand distortion are also evident in the posteriorhalf of the jaws. The skull (Figs 2; 3) is almost 129 cm long (pre-maxilla to quadrate) with a preorbital length of85 cm. The height of the skull in front of theorbit is 47 cm. Femur length (Table 1) is a rela-tively stable standard of measurement intheropods, and can be used to assess proportional
FIG. 3. — Acrocanthosaurus atokensis. Reconstruction of skull; A, dorsal view; B, left lateral view; C, occipital condyle; D-G, denti-cles of maxillary teeth. Scale bars: A, B, 10 cm; C, 2 cm.
D E F G
C
A
B
A, B
differences of the skull. The skull/femur ratio is1.17, which compares well with CeratosaurusMarsh, 1884 (1.00, Gilmore 1920), sinraptorids(0.92-1.17, Currie & Zhao 1993) and tyran-nosaurids (0.97-1.17). The skull of Allosaurus isapparently relatively short (0.76 in USNM 4734and 1.03 in AMNH 5753).In addition to the five major openings on eachside of the skull (external naris, antorbital fenes-tra, orbit, upper and lateral temporal fenestrae),there are two accessory openings in the maxilla(Figs 2; 3). The external naris is relatively smalland elongate, with a maximum length of 16 cm(anteroventral to posterodorsal axis). The naris isbound entirely by the premaxilla and nasal, andthe maxilla is excluded from the opening. This isthe plesiomorphic state found in primitivetheropods like Herrerasaurus Reig, 1963 (Sereno& Novas 1992), in contrast with AllosaurusMarsh, 1877 (Madsen 1976), MonolophosaurusZhao & Currie, 1993 and some Cretaceoustheropods (Currie 1985). The triangular antor-bital fenestra is enormous, covering almost half(42 cm) the preorbital skull length and two thirds(63%, 29 cm) of the height. The jugal forms partof the boundary in Acrocanthosaurus. In somereconstructions of Allosaurus (Madsen 1976), thejugal is excluded from the margin of the fenestra,although it does reach the antorbital fenestra in atleast some specimens (Bakker, pers. comm.1993). The keyhole shaped orbit is high (34 cm)but relatively short anteroposteriorly (12 cm).The outline of the lateral temporal opening ispear-shaped and is bordered above by a shortintertemporal bar.Anterior to the orbit, the side of the skull is a rel-atively flat, vertical face, clearly set off from the nar-row skull roof as in most carnosaurs. A sharplydefined ridge separates the dorsal and lateral sur-faces of the nasal and lacrimal bones. The ridge,which overhangs the face by as much as 1.5 cm atthe anteriormost nasal-lacrimal contact, is rela-tively straight in profile and is only slightly rugoseas in Allosaurus (Madsen 1976). There are no nasal,lacrimal, frontal or postorbital horns as there arein Allosaurus, Ceratosaurus, CarnotaurusBonaparte, Novas & Coria, 1990, Monolo-
phosaurus and most tyrannosaurs, and in this char-acter Acrocanthosaurus is similar to Tyrannosaurus.The paired premaxillae form the sharp, narrowtip of the skull (Fig. 3A). Each is pierced by morethan half a dozen foramina (Fig. 3B) for branchesof the medial ethmoidal nerve and subnarialartery. The maxillary body is higher than long asin Ceratosaurus, Yangchuanosaurus Dong, Zhou& Zhang, 1983 and Torvosaurus Galton &Jensen, 1979, whereas it is longer than high inAllosaurus (Britt 1991) and Sinraptor Currie &Zhao, 1993. In contrast with Allosaurus, the ante-rior margin slopes posterodorsally. There is asloping, posteriorly concave (in lateral aspect)contact with the maxilla, which tapers into a pos-terodorsal (subnarial) process to contact the sub-narial process of the nasal. As in Allosaurus, thesubnarial foramen passes between the main bod-ies of the premaxilla and maxilla.The maxilla is more than 82 cm long. It is excludedfrom the border of the external naris by the pre-maxilla and nasal, but forms most of the anteriormargin of the antorbital opening. A maxillary fen-estra (7 cm by 3.5 cm) pierces the maxilla betweenthe external naris and the antorbital opening. Asin most theropods, the smaller promaxillary fen-estra (3.5 cm high) is tucked under the rim thatforms the anteroventral corner of the antorbitalfossa. The right maxilla has 15 teeth (counting oneempty position), which is close to the counts forSinraptor (15, Currie & Zhao 1993) andYangchuanosaurus (14-15, Dong et al. 1983). Asin Allosaurus, Sinraptor, Yangchuanosaurus andmost other large theropods, the lateral surface ofthe maxilla is rugose only along its anterior edgeand immediately above the tooth row. It is defi-nitely not rugose in the way that the maxillae ofAbelisaurus, Carcharodontosaurus, Carnotaurus andGiganotosaurus are. Most of the lateral surface ofthe maxilla is smooth and somewhat indentedwhere it defines the extent of the antorbital fossa.The maxillae of Carcharodontosaurus (Stromer1931; Sereno et al. 1996), Carnotaurus (Bonaparteet al. 1990), Ceratosaurus (Gilmore 1920),Giganotosaurus (MUCPv-CH-1), IndosuchusChatterjee, 1978 and Torvosaurus (Britt 1991) arequite different in that most of the lateral surfaces
Currie P. J. & Carpenter K.
212 GEODIVERSITAS • 2000 • 22 (2)
are rugose, showing that the antorbital fossae arenot as well developed.The ventral margin of the promaxillary fenestrain the anteroventral apex of the antorbital fossa isat about the same level as the floors of the narisand the antorbital fenestra. This fenestra is foundin most theropods; sometimes (e.g. Ceratosaurus)it is not visible in lateral aspect, and sometimes itis a relatively large opening (Witmer 1997). Thelarger maxillary fenestra is not consistentlyexpressed in theropods, but tends to be present inadvanced forms, including Afrovenator (Sereno etal. 1994), Allosaurus (Witmer 1997), tyran-nosaurids and most small theropods. It is absentin Abelisaurus Bonaparte & Novas 1985,Carnotaurus (Bonaparte et al. 1990), Ceratosaurus(Gilmore 1920), Carcharodontosaurus Stromer,1931 (Sereno et al. 1996 identify a maxillary fen-estra, but it is in the position normally occupiedby the promaxillary fenestra), Giganotosaurus(MUCPv-CH-1), Indosuchus Chatterjee 1978,Majungatholus Sues & Taquet, 1979 (Sampson etal. 1998) and Torvosaurus (Britt 1991), and isexpressed as a variable complex of openings insinraptorids (Currie & Zhao 1993; Witmer1997). The posterodorsal process of the maxillapasses between the nasal bone and the antorbitalfenestra, and bifurcates distally to embrace theanteroventral process of the lacrimal.The nasal is 85 cm in length, more than four timesthe length of the frontal. The dorsal surface is flatand relatively narrow for most of its length, and isclearly delimited by a sharp inflection from thenasal contribution to the antorbital fossa as it is inAllosaurus, Sinraptor and Yangchuanosaurus. Thisis different in abelisaurids (Bonaparte & Novas1985; Bonaparte et al. 1990), Monolophosaurus andtyrannosaurids, where at least half of the nasal hasa convex and highly rugose dorsal surface. Likemost theropods, the median internasal suture is notfused. The nasal is split anteriorly where the pos-terior tip of the premaxilla overlaps a shallowtrough for a length of 11 cm. As in Allosaurus,Ceratosaurus, Giganotosaurus, Monolophosaurus,Sinraptor and Yangchuanosaurus, the nasal formsthe anterodorsal rim of the antorbital fossa, andeven extends slightly onto the medial wall. It
appears as if pneumatic openings may have invadedthe nasal along the margin of the antorbital fossain the way they do in Allosaurus (Madsen 1976;Gilmore 1920), Giganotosaurus (R. Coria, pers.comm. 1996), Monolophosaurus (Zhao & Currie1993) and Sinraptor (Currie & Zhao 1993). As inmost other large theropods, there is a groove in theventrolateral margin of the nasal for the maxillaryarticulation that would have permitted some lat-eral-medial rotation of the maxilla. Above the pos-terior end of the suture with the maxilla, the nasaloverlaps the anterior tip of the lacrimal. Each nasalmeets a prefrontal posterolaterally, and overlaps ananteroposteriorly elongate shelf of the frontal(Fig. 3A).The lacrimal is 35 cm long and 37 cm high. Thedorsolateral margin forms a low ridge at the frontof the orbit comparable with Carcharodontosaurus,Giganotosaurus (R. Coria, pers. comm. 1996),Sinraptor (Currie & Zhao 1993) and Yangchuano-saurus (Dong et al. 1978). Allosaurus, Ceratosaurus(Gilmore 1920), the smaller genera of tyran-nosaurids, and many other theropods have devel-oped “horns” in this region. The lacrimal has a lim-ited dorsal exposure. Anteriorly, there areinterdigitating sutures with the maxilla and nasal.Its lateral surface is sculptured posterodorsally forclose association with the skin. The lacrimal sendsa narrow posterior extension along the ventrolat-eral margin of the prefrontal and seems to contactthe postorbital beneath a thin lateral lappet of theprefrontal (Fig. 3B). In lateral aspect (Figs 2A; 3B),the posterior margin of the ventral (preorbital)process of the lacrimal has a double concavity sep-arated by a short process (for attachment of theLigamentum suborbitale) that marks the anteroven-tral extent of the eyeball itself (diameter was about11 cm). Although less obvious in the holotype(Stovall & Langston 1950), the process is similarto those of Abelisaurus (Bonaparte & Novas 1985),Majungatholus (Sampson et al. 1998), Mono-lophosaurus, Sinraptor and Yangchuanosaurus. Theposterior margin of the preorbital bar of other largetheropods tends to be uniformly concave in lateralview. Below this low process of the lacrimal inAcrocanthosaurus, the preorbital bar narrows to64 mm in lateral aspect. The posteroventral end of
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
the lacrimal expands somewhat for its contact withthe jugal. Much of the anterolateral surface of thepreorbital bar is shallowly excavated for the antor-bital fossa, which extends ventrally onto the jugal.Ventrally, the lacrimal is overlapped by the jugal,and does not contact the maxilla. In contrast, thelacrimal may contact the maxilla externally inAllosaurus and Ceratosaurus (Gilmore 1920).As in other large theropods, there is a lateral, pneu-matic opening into a vacuity in the main pos-terodorsal body of the lacrimal. The opening hasa diameter of 2.7 cm in this specimen. It wouldhave housed a pneumatic diverticulum of the nasalcavity (Osmolska 1985; Witmer 1997). The pres-ence of two large openings in the lacrimal of theholotype (Stovall & Langston 1950) shows thereis some variation of this feature, which is not sur-prising considering how variable pneumatic fea-tures are (Currie & Zhao 1993).Posteromedial to the lacrimal, there is a relativelylarge, triangular prefrontal (11 cm long, 6 cm
broad) as in Allosaurus, Sinraptor and othercarnosaurs. It extends posteriorly to contact thepostorbital and helps to roof over the orbit. Athin lappet appears to have extended over thelacrimal to reach the orbital margin.The dorsal surface of the frontal is flat, as it is inAllosaurus and Sinraptor (Currie & Zhao 1993).The surface of the bone is smooth as in themajority of large theropods, but in contrast withthe ornamented surface in Abelisaurus (Bonaparte& Novas 1985) and Majungatholus (Sampson etal. 1998). As in the holotype, the interfrontalsuture is more or less completely fused. Like thefrontals of the holotype, there is a slightly raisedrugose bump along the midline near the fron-toparietal suture. A shallow trough, emphasizedanteriorly by a raised ridge, leads into a foramenat the junction of the frontal, prefrontal and pos-torbital (Fig. 3B), although the latter two bonescontact each other to exclude the frontal from themargin of the orbital rim. To an extent, this is a
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
size specific characteristic that is shared by thelargest theropods, including Carcharodontosaurus,Giganotosaurus, Tarbosaurus and Tyrannosaurus.The frontal forms the anterior rim of thesupratemporal fossa, which it floors for a distanceof almost 5 cm. At the dorsal limit of the uppertemporal fenestra, the fused frontoparietal sutureis strengthened by additional bone depositionthat protrudes laterally into the fenestra as asharply defined ridge. This has also been noted inGiganotosaurus (Coria & Currie in prep.) andSinraptor (Currie & Zhao 1993). The frontopari-etal suture can be followed laterally to the pointwhere both bones contact the postorbital. The parietal has limited dorsal exposure along themidline (about 3 cm) and compares well with theparietals of carcharodontosaurids and sinrap-torids. However, the dorsal surface is almost per-pendicular to the occiput (Stovall & Langston1950: pl. II-2), which itself is almost vertical.This angle is distinctly obtuse in carcharodon-tosaurids and sinraptorids. Correlated with this,each posterolateral wing of the parietal forms analmost vertical posterior wall for the supratempo-ral fossa, whereas it strongly slopes in carcharo-dontosaurids and sinraptorids. The posterolateralwing extended along the top of the paroccipitalprocess, tapering to a point that ends only 5 cmfrom the distal end of the process. There is nosharp sagittal crest along the midline of the pari-etal and, in this sense, Acrocanthosaurus is asprimitive as all theropods except tyrannosauridsand troodontids. The nuchal crest is low anddoes not extend more than a few millimetresabove the supraoccipital. Again, this situation isvery different in tyrannosaurids and troodontids.The orbital margin of the postorbital extends for-ward to contact the lacrimal and prefrontal, andhas thickened into a prominent horizontal orbitalboss as in abelisaurids, carcharodontosaurids andthe largest tyrannosaurids. These are size-depend-ent characteristics that have uncertain taxonomicsignificance. The rugose rim of the postorbital is4 cm thick dorsoventrally. The intertemporalramus of the postorbital is relatively short and isoriented almost as much ventrally as it is posteri-orly (Fig. 3B). The postorbital bar of the postor-
bital is relatively wide in lateral view, reaching amaximum anteroposterior length of 9.7 cmbehind the orbit and has an anterior processbeneath the orbit (Fig. 2A). This process is alsoevident in the holotype (Stovall & Langston1950), and similar processes are found inCarcharodontosaurus, Giganotosaurus, GorgosaurusLambe, 1917, Tarbosaurus Maleev, 1974 andTyrannosaurus. Presence of the process is in partsize-related, and therefore has limited utility forassessing taxonomic relationships.The jugal is similar to those of most other largetheropods in that it separates the maxilla andlacrimal to take part in the rim of the antorbitalfenestra. The long, sloping anteroventral edge sitsin a shallow trough on the posterodorsal margin ofthe maxilla. The lateral surface is depressed ante-riorly where it contributes to the antorbital fossa.This region has been damaged somewhat, but therewere almost certainly pneumatic openings into thebody of the jugal as in other specimens ofAcrocanthosaurus (Stovall & Langston 1950;Harris 1998a), some specimens of Allosaurus(USNM 4734, UUVP 1403, UUVP 3894,UUVP 3981), Monolophosaurus, Sinraptor andtyrannosaurids. The postorbital process of the jugal(NCSM 14345, OMNH 10146) is a huge trian-gular plate of bone in lateral view, and has a longsloping contact with the postorbital bone that onlyreaches two thirds of the way up the postorbitalbar. The lateral surface of this part of the jugal isshallowly concave as in Daspletosaurus Russell,1970, Tarbosaurus and Tyrannosaurus. Similar toAllosaurus , Ceratosaurus , Sinraptor andDeinonychus Ostrom, 1969, the jugal forms theventroposterior margin of the orbit. Like alltheropods except Sinraptor dongi, the quadratoju-gal process of the jugal splits posteriorly into two.The uppermost prong of the quadratojugal processis much shorter than the ventral prong as in all largetheropods except abelisaurids (Bonaparte et al.1990; Sampson et al. 1998), Ceratosaurus and tor-vosaurs (Bakker et al. 1992).The holotype of Acrocanthosaurus (Stovall &Langston 1950) has an inferior process on thesquamosal that expands moderately ventrally forits contact with the quadratojugal. The two
Currie P. J. & Carpenter K.
216 GEODIVERSITAS • 2000 • 22 (2)
bones then invade the lateral temporal fenestra toa greater extent than is seen in Allosaurus and sin-raptorids, but not as much as in tyrannosaurids.The inferior process of the squamosal is indentedby a deep quadrate cotylus, and the head of thequadrate would have been exposed laterally.Posteroventrally, the squamosal extended alongthe dorsal surface of the paroccipital process, cov-ering up the ventrolateral process of the parietaland wrapping around onto the anterodorsal sur-face of the distal end of the paroccipital process.The squamosal was inclined in such a mannerthat it would have roofed over the posterior tem-poral musculature.A conspicuous midline ridge on the supraoccipi-tal is comparable with those of carcharodon-tosaurids, sinraptorids and Allosaurus (Madsen1976). Dorsally the supraoccipital expands oneither side of the midline until it protrudes in adouble boss 5 cm behind the nuchal crest. Thedorsal surfaces of these knobs are rugose, and areonly a few millimetres below the highest point onthe parietal. Stovall & Langston (1950) reportedthat the supraoccipital was excluded from themargin of the foramen magnum by the exoccipi-tals, although fusion makes it difficult to be surethat this is the case. The lateral margins of thesupraoccipital are nearly vertical, and are moresimilar to those of Allosaurus (Madsen 1976) thanto Sinraptor.The paroccipital process is relatively wide but shal-low, in contrast with the deeper tyrannosaurid con-dition. Distally the process ends in a vertical sheetof bone 2.5 cm thick and 11 cm high. The paroc-cipital process turns downwards distally (Stovall &Langston 1950) as in Allosaurus and Sinraptor, anddoes not project as strongly backwards (Fig. 3A) asit does in Carcharodontosaurus (Sereno et al. 1996),Giganotosaurus (MUCPv-CH-1) and Sinraptor(Currie & Zhao 1993). In the holotype, at leasttwo branches of the twelfth cranial nerve emergethrough a depressed area between the occipitalcondyle and the paroccipital process.The quadrate is a relatively tall bone (31 cm) asin almost all theropods except Allosaurus andtyrannosaurids, and is only shallowly concaveposteriorly. The condyles of the jaw articulation
are at least 11 cm across. There is a quadrate fora-men with a diameter of 28 mm. Except for a nar-row slit, it is completely surrounded by thequadrate as in Allosaurus (Madsen 1976) andSinraptor (Currie & Zhao 1993). The gap isclosed by the quadratojugal, which also curvesonto the occiput where it overlaps the quadratesomewhat along the dorsolateral margin of theforamen.The occipital condyle (Fig. 3C) is nearly circularin appearance, which contrasts with the flattenedoval appearance of the occipital condyles ofGiganotosaurus, Piveteausaurus Taquet & Welles,1977 and Tyrannosaurus. The shape of the occip-ital condyle is characteristic of theropods thatcarry their skulls at the end of an extended neck.In spite of the large size of the specimen, thesutures between the exoccipital and basioccipitalare still visible in the occipital condyle.Palatal elements are generally obscured by theother bones of the skull. The palatine andectopterygoid of Acrocanthosaurus have beendescribed for SMU 74646 (Harris 1998a).There are four premaxillary, 15 maxillary and anunknown number of dentary tooth positions. Thethird premaxillary tooth is the largest on the leftside, attaining a dorsoventral length of 85 mm anda basal diameter of 27 mm. It is 72% the length ofthe largest maxillary tooth. Allosaurus andNeovenator Hutt, Martill & Barker, 1996 have fivepremaxillary teeth, Ceratosaurus and Torvosaurus(BYU 4882) have three, and most other theropods,including abelisaurids, Eustreptospondylus Walker,1964, Herrerasaurus, Marshosaurus Madsen, 1976,Monolophosaurus, Proceratosaurus Woodward1910, Sinraptor, tyrannosaurs and Yang-chuanosaurus, have four. Because the premaxillaemeet at an acute angle, the premaxillary teeth arenot D-shaped in section as they are in tyran-nosaurids (Currie et al. 1990). In fact, even in thefirst premaxillary tooth of Acrocanthosaurus, theanterior carina is positioned on the midline of thetooth at the tip. However, it curves onto the antero-medial surface closer to the root as in allosauridsand dromaeosaurids, but in contrast withTorvosaurus (Britt 1991). Ceratosaurus has no ante-rior carina on premaxillary teeth (Bakker, pers.
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
217GEODIVERSITAS • 2000 • 22 (2)
comm. 1993). There are 15 denticles per 5 mm onthe anterior carina, and 13 on the posterior. Thelongest crown (fourth from the front) in the leftmaxilla is 118 mm high, with a FABL (fore-aft baselength) of 41 mm. This is comparable in lengthwith the longest tooth of a similar sized tyran-nosaurid (GI 107/2, Tarbosaurus, has a skull lengthof 122 cm and a maximum maxillary tooth lengthof 115 mm), with the exception of Tyrannosauruswhich seems to have had more elongate teeth(AMNH 5027 has a skull length of 136 cm and amaxillary tooth length of 142 mm). Although themaxillary teeth are narrower and more blade-likethan those of tyrannosaurids, they are not as later-ally compressed as the teeth of carcharodon-tosaurids (Sereno et al. 1996). For example, theratio of length to width of a cross-section of thebase of an Acrocanthosaurus tooth described byHarris (1998a) is 0.63, compared with 0.43 in aselection of carcharodontosaurid teeth (casts inTyrrell Museum of privately owned specimensfrom Morocco). Furthermore, the teeth ofAcrocanthosaurus do not have the wrinkles in theenamel that are characteristic of carcharodon-tosaurid teeth (Sereno et al. 1996; Harris 1998a).In spite of the large size of the teeth inNCSM 14345, the denticles are minute (the sec-ond premaxillary tooth has 12.5 per 5 mm on theposterior carina; the fourth maxillary tooth has17.5 per 5 mm on the anterior carina, and from12.5 to 17.5 on the posterior edge). This is evenless than the denticle size of Carcharodontosaurus,where there are 10 serrations per 5 mm in a toothwith a FABL of 36 mm (Russell 1996). Harris(1998a) pointed out that the denticles extendacross the tips of Acrocanthosaurus teeth, and sug-gested this is unusual. However, it is the same sit-uation in a diverse assemblage of theropods thatincludes carcharodontosaurids, velociraptorinedromaeosaurids, and tyrannosaurids, so this fea-ture has limited taxonomic utility.
Lower jawThe lower jaw (132.5 cm long) is well-preservedon both sides, although crushing has damagedsome of the thinner bones, especially along theintramandibular articulation and around the
margins of the external mandibular fenestra. Theexternal mandibular fenestra is preserved on bothsides, although it is difficult to precisely definethe margins. It appears to have been about 20 cmlong, which is significantly larger than that ofAllosaurus (Madsen 1976). It is positioned at thejunction of the dentary, angular and surangular.The jaw is shallow anteriorly, and deep posterior-ly. The depth has been emphasized somewhat bycollapse of the adductor fossa.As in almost all theropods (Bakker et al. 1988),including Herrerasaurus (Sereno & Novas 1992),there is an intramandibular joint. A posteriorlyprojecting lateral process of the dentary near thedorsal margin, referred to by Currie & Zhao(1993) as the intramandibular process of the den-tary, fits into a slot in the labial side of the suran-gular. An anteriorly projecting tongue, the intra-mandibular process of the surangular, extendsventrolateral to the intramandibular process of thedentary to form a combination sliding and pivot-ing joint. Ventrally, the anterior end of the angu-lar had a sliding joint along the top of a shelf formedby the dentary and the splenial.The dentary is relatively long and low, with gen-erally concave upper and lower margins in lateralview. It is unusual in having a distinctive ventralprocess at the bottom of the symphysis, whichgives the front of the jaw (10 cm high) an angu-lar, squared off appearance. This is even moreconspicuous in Giganotosaurus, but is also presentto a lesser degree in large tyrannosaurids, suggest-ing that it may be a size-dependent character.The lateral surface, especially at the front of thebone, is pierced by a dorsal row of relatively largemental foramina for innervation and vasculariza-tion of the skin and lips. The smooth surface ofthe bone contrasts with the ornamented surfacein Abelisaurus (Bonaparte & Novas 1985). Thefront of the jaw is relatively shallow and is only96 mm deep at mid-length. For comparison, thesame measurement in AMNH 5881, a specimenof Tyrannosaurus rex with a slightly shorter jaw, is170 mm. Posteriorly, the dentary is deep (19 cmat the front of the intramandibular articulation,and almost 10 cm deeper at the back). Its posteri-or margin slopes posteroventrally from the lock-
Currie P. J. & Carpenter K.
218 GEODIVERSITAS • 2000 • 22 (2)
ing intramandibular joint. Two notches in theback of the dentary on the lower, left side areprobably the result of postdepositional damage,but are fortuitous in that they expose part of thesliding joint with the angular.The splenial presumably wrapped around the ven-tromedial margin of the angular, thereby formingpart of the sliding intramandibular joint. However,it does not wrap around far enough to be exposed inlateral view as in Herrerasaurus (Sereno & Novas1992), Ceratosaurus (USNM4735), dromaeosaurids(Currie 1995) and other primitive theropods.The anterior surangular foramen of the surangu-lar is continuous anteriorly with a groove as inAllosaurus, Monolophosaurus, Sinraptor and othertheropods. The lower margin of the groove isformed anteriorly by a finger-like process thatextends laterally over the labial surface of the den-tary. A similar finger-like process of the dentarycontacts the medial and dorsal margins of thegroove. There is an extensive squamose suture forthe angular, and posterolaterally the surangularcovers part of the prearticular. Stovall & Langston(1950) suggested that the posterior surangularforamen of OMNH 10146 was relatively large, asit is in tyrannosaurs. However, it is relatively small
(diameter of 12 mm) in NCSM 14345, where thisregion is better preserved. It is found anterolateralto the glenoid beneath a powerful lateral ridge thatoverhangs the lateral surface of the jaw by about5 cm. This ridge is more powerfully developed thanin any other known large theropod other thanGiganotosaurus (MUCPv-CH-1), although it isalso pronounced in abelisaurids (MACN-CH 894). Harris (1998a) noted the presence of aconspicuous knob on the surangular shelf close tothe jaw articulation. As in other theropods, thesurangular of Acrocanthosaurus forms part of thelateral mandibular glenoid (Harris 1998a).The angular is a shallowly curved plate of bonestrengthened by a thick ventral margin. An exten-sive part of the bone is overlapped laterally by thedentary, but the contact is smooth, and slidingmovement was possible. Posteriorly, the angularlaterally overlaps the surangular. The ventral edgeof the angular is thickest where it forms the ven-tral margin of the jaw behind the intramandibularjoint. Unlike Allosaurus but like most othertheropods, the posterior end of the angular termi-nates anterior to the posterior surangular foramen.The dorsal edge of the supradentary can be seenbehind the last mandibular tooth extending pos-
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
219GEODIVERSITAS • 2000 • 22 (2)
FIG. 4. — Acrocanthosaurus atokensis NCSM 14345, caudal vertebrae; A, preserved series in left lateral view; B, mid-caudal (seventhone in preserved series) vertebra with haemal arch; C, ninth caudal in preserved series; D, distal caudal (twelfth in preserved series)with preceding haemal spine. Scale bars: A, 10 cm; B-D, 5 cm.
B-D
teriorly across the intramandibular joint. It can-not be determined whether it was fused to thecoronoid or not, although it was presumablycapable of restricted sliding movement againstthe surangular laterally and the prearticular medi-ally as in Monolophosaurus (Zhao & Currie1993). The posteroventral margin of the preartic-ular is exposed in lateral view where it extends tothe back of the jaw.The medial glenoid and most of the interglenoidridge for the jaw articulation are found on thearticular. Harris (1998a) reported that the inter-glenoid ridge of SMU 74646 is relatively lowcompared with that of Allosaurus. Overall, thearticular looks relatively short in lateral viewbecause of the posteromedial orientation of theretroarticular process, but is in fact much largerthan it is in Allosaurus (Gilmore 1920; Madsen1976) and tyrannosaurids (Lambe 1917; Osborn1912; Molnar 1991).
Axial skeletonAlmost two dozen partial vertebrae were recoveredfrom the skeleton, but, with the exception of threecaudals, none are complete (Fig. 4). From what canbe seen of the vertebrae and a number of isolatedneural spines, vertebrae of NCSM 14345 did notdiffer in any substantial way from those found withthe holotype (Stovall & Langston 1950) andSMU 74646 (Harris 1998a). A fragment of a pos-terior cervical or anterior dorsal centrum is piercedby two pneumatopores as in OMNH 10146 andSMU 74646. The internal pneumatization is ofthe complex type (Britt 1993), in contrast with thesimple pneumatization found in Allosaurus andSinraptor. This has also been noted in SMU 74646(Harris 1998a). The neural spines in the middleand distal parts of the tail are restricted to the pos-terior region of the vertebrae. As in the caudal ver-tebrae described by Stovall & Langston (1950) andHarris (1998a), there is a “cranial process” above
Currie P. J. & Carpenter K.
220 GEODIVERSITAS • 2000 • 22 (2)
FIG. 5. — Acrocanthosaurus atokensis NCSM 14345; A, posterior cervical rib; B-D, dorsal ribs. Scale bar: 10 cm.
the anterior insertion region for the interspinousligaments (Fig. 4B, C). Mid-caudal centra areabout 16 cm long, 10 cm wide and 11.5 cm high,whereas the most distal one preserved is 12 by 7.5by 7.5 cm. The distal caudal has a moderately elon-gate prezygopophysis (Fig. 4D).Six chevrons (Fig. 4) from the mid-caudals aredirectly comparable in overall shape with those ofAllosaurus (Madsen 1976), and in spite of elonga-tion of the neural spines, do not appear to be anymore elongate than in the Jurassic theropod.One elongate cervical rib (Fig. 5A) has a prominentknob projecting anterolaterally from between thetuberculum and capitulum. Most of the ribs fromthe dorsal region are represented in the collection,but fewer than half a dozen are complete. They aredirectly comparable with those of Allosaurus(Madsen 1976), and show that Acrocanthosaurushad a rather high, narrow body. There were alsomany fragmentary gastralia recovered, but there isnothing to suggest that the gastral basket was differ-ent in any substantial way from other specimens ofAcrocanthosaurus (Harris 1998a).
Appendicular skeletonAll elements of the pectoral girdle (Fig. 6) andfront limb (Fig. 7) are represented except forsome of the unguals. The arm is relatively short.The sum of the lengths of the humerus, ulna(excluding the olecranon process), ulnare,metacarpal II and digit II is 105 cm, which isshorter than either the scapulocoracoid (118 cm)or the femur (128 cm). In contrast, Allosaurus hasa relatively longer arm. That of MOR 693 is94 cm long, which is 30% longer than the scapu-locoracoid. The arm of Acrocanthosaurus is 81%the length of the femur, whereas the arm ofAllosaurus is 26% longer than the femur inMOR 693 and 12% longer in USNM 4734. Thedifferences are less evident in the humerus thanthey are in the forearm and hand, where speci-mens of Allosaurus (MOR 693, TATE 11,USNM 4734) that are 20% smaller than Acro-canthosaurus have radii, ulnae and metacarpalsthat are absolutely longer than the same elementsin Acrocanthosaurus. This shows that the differ-ences in arm length between the taxa cannot be
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
221GEODIVERSITAS • 2000 • 22 (2)
attributed to allometric scaling. Tyrannosaurids,on the other hand, have even shorter arms thanAcrocanthosaurus. The humerus is about 30% the
FIG. 6. — Acrocanthosaurus atokensis NCSM 14345, left scapu-locoracoid. Scale bar: 10 cm.
outside curve) long. It is only 9.5 cm wide at itsnarrowest point when seen in lateral aspect, andthe distal end is only moderately expanded. Theacromial process is pronounced and is sharplyoffset from the anterodorsal margin of the scapu-lar blade as in Allosaurus (Madsen 1976),Sinraptor (Currie & Zhao 1993) and tyran-nosaurids (Maleev 1974). The scapular blades ofCeratosaurus (Madsen 1976), Carnotaurus(Bonaparte et al. 1990), Edmarka Bakker et al.,1992 and Torvosaurus (Bakker et al. 1992), andMegalosaurus Buckland, 1824 (Walker 1964) aremore robust, and the anterior margin gradessmoothly into the acromion process.The outline of the coracoid (Fig. 6) is also closerto that of Allosaurus than it is to Ceratosaurus(Madsen 1976) or megalosaurs (Bakker et al.1992). The coracoid foramen is a small but con-spicuous opening close to the centre of the bone.The coracoid extends posteroventrally wellbeyond its contribution to the glenoid as inadvanced theropods plus abelisaurids.The humerus (Fig. 8) is a rather massive element37 cm long. The proximal end is expanded tosuch a degree that its maximum width is half thelength of the bone. The deltopectoral crest islarge (9.5 cm) and well-developed, protruding6.5 cm from the shaft (Fig. 8E). UnlikeAllosaurus, which has a deltopectoral crest thattapers steadily away from the shaft, that ofAcrocanthosaurus is quadrilateral in outline.There is a well-developed scar for the humero-radialis muscle (Fig. 8E). The transverse shaftdiameter is only about a third the width of theproximal end, and slightly less than half of thedistal expansion. The distal end is rotated out-ward from the plane of the proximal end(Fig. 8B), and the deltopectoral crest is orientedat right angles to the distal end of the bone(Fig. 8G) as in Allosaurus. There is a deep pit onthe anteroventral side proximal to the radialcondyle and the ulnar groove (Fig. 3C). Theentepicondyle is a small (1.5 cm) but pronouncedtubercle (Fig. 8D) separated from the radialcondyle by a groove of finished bone, althoughfinished bone continues from the distal end ofthe humerus up along the crest of the tubercle. In
Currie P. J. & Carpenter K.
222 GEODIVERSITAS • 2000 • 22 (2)
length of the femur in both Acrocanthosaurus andTyrannosaurus, but the forearm is relatively short-er in the latter. The front limb of Tarbosaurus(GI 107/2) is 65 cm in length, which is 93% thelength of the scapulocoracoid and 60% thelength of the femur, whereas that of oneTyrannosaurus rex (FMNH PR2081) is 89 cmlong, which is less than 70% of either scapuloco-racoid or femur length. The scapula is a long, slender, gently curved ele-ment (Fig. 6), 97 cm (103 cm measured on the
FIG. 7. — Acrocanthosaurus atokensis NCSM 14345, re-construction of right arm. Scale bar: 10 cm.
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
223GEODIVERSITAS • 2000 • 22 (2)
most characters, the humerus of Acrocanthosaurusis like that of Allosaurus, and most of the differ-ences seem to be allometric changes related to thelarger size of the Cretaceous form. However, theshaft is not as twisted along its length as it is inAllosaurus.The ulna is a robust bone with a total length of25.5 cm, almost 15% of which is composed ofthe olecranon process (Fig. 9). The proximal con-cavity that contacts the proximal end of theradius has neither articular facets nor strong liga-mentous attachments. However, there is a well-defined distal articulation for the radius, whichconsists of a convex surface of unfinished boneelevated almost a centimetre from the general
contour of the ulna by a pedestal of bone(Fig. 9C, lower right). Amongst describedtheropods, the ulna most closely resembles thatof Allosaurus (Madsen 1976), although it is rela-tively shorter (69% the length of the humerus,compared with 85% the length in USNM 4734).The radial notch of the ulna is not as deep orprominent as in Allosaurus.The radius (Fig. 9) is a curved, slender elementthat is difficult to measure. The straight linelength is 22 cm, whereas the distance between theclosest points of the articular surfaces is only20 cm. It is therefore 55% to 60% of the lengthof the humerus, whereas the radius of Allosaurusis more than 70% of humerus length (Gilmore
FIG. 8. — Acrocanthosaurus atokensis NCSM 14345, right humerus; A, medial aspect; B, proximal aspect; C, posterodorsal aspect;D, distal aspect; E, lateral aspect; F, anteroventral aspect; G, outlines of proximal (bold) and distal (light) ends. Scale bar: 10 cm.
➝
➝➝
Currie P. J. & Carpenter K.
224 GEODIVERSITAS • 2000 • 22 (2)
FIG. 9. — Acrocanthosaurus atokensis NCSM 14345; A-F, right ulna; A, medial view; B, proximal view; C, anterior view; D, distalview; E, lateral view; F, posterior view; G-L, radius; G, medial view; H, proximal view; I, anterior view; J, distal view; K, lateral view;L, posterior view; M, outlines of proximal (bold) and distal (thin) ends of articulated ulna (upper) and radius (lower) with the arrowpointing towards the anterior side of the forearm. Scale bar: 10 cm.
➝➝
➝
➝
➝
1920). The proximal articulation is trapezoidal(Fig. 9H) with a maximum mediolateral width of6 cm and an anteroposterior length of 5.2 cm. It issaddle-shaped with strong anterior and posteriorridges. The trough-like articulation continuesmedially onto the ulna. The edge of the proximalend is rugose where it contacts the ulna. Distal tothe proximal articulation, the shaft of the radiuscurves anteromedially (Fig. 9I) as it twists aroundthe front of the ulna, closely following the curva-ture of that bone. A well-defined, twisting ridgewith a rugose surface (Fig. 9I) lies close to the ante-rior margin of the ulna and presumably marks theline of ligamentous attachments with its neighbor.The lower half of the bone has an almost straightshaft, although the distal end is set off at an angleto it (Fig. 9I, K). There is an extensive elevatedarea (about 4 cm wide and 2 cm high) posterome-dially near the distal articulation for contact withthe ulna. Its rugose surface, and that of the corre-sponding region of the ulna, suggests that the liga-mentous attachments were strong and movementbetween the bones was limited. This also wouldhave restricted rotation of the wrist. The distalarticular surface is a shallow concavity that wouldhave articulated with the radiale.Three carpals were recovered with the specimen(Figs 10; 11). One of these elements was preservedin articulation with the metacarpals of the righthand. Furthermore, it fits perfectly across the headof metacarpal I and extends about a quarter of theway across the head of metacarpal II. This elementhas been identified as the intermedium in othertheropods, including Allosaurus (Gilmore 1920),Gorgosaurus (Lambe 1917) and Tarbosaurus(Maleev 1974), although Madsen (1976) identi-fied it as distal carpal I (with carpal II fused into itin 14 out of 18 specimens of Allosaurus), as didOsborn (1917) for Struthiomimus, Russell &Dong (1993) for Alxasaurus and Padian &Chiappe (1997) for maniraptorans. Colbert iden-tified the carpal in this position in CoelophysisCope, 1889 as distal carpal I + II. In this paper, werefer to it as distal carpal I because it occupies theprimitive position (Sereno 1993) of this elementabove the contact between metacarpals I and II.However, we also recognize that it was probably
formed by the fusion of several elements as inAllosaurus. Regardless of what name should be puton this element, it is morphologically similar tothe same unit in Allosaurus, therizinosaurids andtyrannosaurids.Distal carpal I is a complex element that is closelyappressed to metacarpals I and II. This close asso-ciation shows that little movement would havebeen possible amongst these three elements, all ofwhich were tightly integrated. At its thickestpoint above the posterior junction of meta-carpals I and II, it is 16.2 mm thick proximodis-tally. Lateromedially it measures 60.8 mm closeto the extensor surface and is 52.4 mm antero-posteriorly. In comparison, one of the larger dis-tal carpals from the Cleveland-Lloyd collection ofAllosaurus elements is thicker (24 mm), narrower(50 mm at the extensor surface) and anteroposte-riorly longer (54 mm). The anterior (extensor)surface (Figs 10C; 11B) is a thin edge of finishedbone (it is tall in Allosaurus, Madsen 1976) thataligns with the anterior margin of metacarpal I.This surface is rugose and pitted like the proxi-moanterior surface of the first metacarpal, sug-gesting there were strong ligamentousattachments between these bones. Lateral to thejunction of metacarpals I and II, the distal surface(Fig. 10B) becomes concave, rising up to coverthe proximal articular surface of the secondmetacarpal. Concurrent with this, the carpalthins laterally and withdraws from the extensorsurface to leave part of the proximal articular sur-face of metacarpal II exposed anteromedially(Fig. 11A). The posterior (flexor) surface of thecarpal is partially composed of a surface of fin-ished bone above the junction of the twometacarpals (Figs 10D; 11D). Medial to this, thecarpal thins and withdraws from the flexor sur-face (Fig. 11A) to wrap around the margin of adepression in the proximoposterior surface of thefirst metacarpal. This is where the troughlike“pulley” articulation of the carpal passes onto themetacarpal. Medial to the depression in themetacarpal, the posterior edge of the carpalapproaches the flexor surface of the metacarpal,although it remains a thin edge between convexproximal and distal articular surfaces (Fig. 11C).
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
225GEODIVERSITAS • 2000 • 22 (2)
Currie P. J. & Carpenter K.
226 GEODIVERSITAS • 2000 • 22 (2)
The proximal surface has a distinct trough thatstarts from the emarginated posteromedial region(Fig. 10A) and extends anterolaterally to the lat-eral surface (Fig. 10F). A less prominent trough-like depression isolates the convex anteromedialregion of the proximal surface. The distal surfaceof the carpal (Fig. 10B) has a prominent ridgethat follows the junction of the first and secondmetacarpals. The more lateral articular surface forthe second metacarpal consists of shallow anteriorand posterior depressions separated by a weak lat-eromedial ridge (emphasized anteriorly by a shal-low trough). The lateral articular surface fitsperfectly on a pair of low convex surfaces on thesecond metacarpal. The medial articulation on
the distal surface is more complex. It contactedthe first metacarpal in three areas. The mostmedial one is the largest, and is a flattened saddle-shaped articulation. This is separated from theother articulations by a channel that seems tohave been utilized by blood vessels or nerves. It isdeep and well-defined on the distal surface of thecarpal where it enters posteriorly the region ofarticulation with the first metacarpal from theflexor surface, but becomes less distinct as itdivides into two troughs. The more medial oneexits the extensor surface via a well-marked canalin the proximal surface of the first metacarpal.The other branch turns to become the shallowtrough anterior to the low mediolateral ridge that
FIG. 10. — Acrocanthosaurus atokensis NCSM 14345, carpals; A-F, distal carpal I; A, dorsal view; B, ventral view; C, anterior view;D, posterior view; E, medial view; F, lateral view; G-L, ulnare; G, dorsal view; H, ventral view; I, medial view; J, lateral view; K, ante-rior view; L, posterior view; M-R, radiale; M, dorsal view; N, ventral view; O, medial view; P, lateral view; Q, anterior view; R, posteri-or view. Scale bar: 2 cm.
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
227GEODIVERSITAS • 2000 • 22 (2)
divides the lateral articulation into anterior andposterior components. Overall, then, the distalarticular surface has a total of five contacts withthe first and second metacarpals.
FIG. 11. — Acrocanthosaurus atokensis NCSM 14345, reconstruction of the right wrist; A, dorsal aspect; B, anterior aspect; C, medi-al aspect; D, posterior aspect. Scale bar: 2 cm.
➝
The ulnare closely resembles that of Allosaurus(Madsen 1976). In proximal and distal aspects(Fig. 11G, H), the bone is quadrangular, althoughthe anterolateral side is so short that it looks trian-
radialeA
B
C
D
radiale
distal carpal
distal carpal
ulnare
ulnare
radius
ulna
Currie P. J. & Carpenter K.
228 GEODIVERSITAS • 2000 • 22 (2)
gular. The anterolateral face has a convex surfaceand an oval outline (Fig. 10K). The longitudinalaxis of the oval is 18 mm long. The posterior sideof the carpal (Fig. 10L) is triangular (41 mm wideand 23 mm high) and has an almost flat surface.The proximal surface is saddle-shaped (in sectionit is concave along the anteroposterior axis, andconvex perpendicular to that axis) and the distalsurface (Fig. 10H) is mostly convex.The smallest of the carpals is the radiale (Figs 10;11), which is 4 cm across and 2.8 cm deep. Mostsurfaces are convex, and the bone had a sliding
anteromedial-posterolateral contact with distalcarpal I. There are three powerfully built metacarpals inAcrocanthosaurus (Fig. 12). With the exception ofthe broadly expanded proximal and distal ends,the metacarpals resemble those of Allosaurus(Madsen 1976), Sinraptor (Currie & Zhao 1993)and other carnosaurs. Szechuanosaurus zigongensis(Gao 1993) also has broadly expanded ends onsome of its metacarpals. However, there are manyhighly significant differences showing this animalis not closely related to Acrocanthosaurus, which
FIG. 12. — Acrocanthosaurus atokensis NCSM 14345, right manus. Metacarpals (A, E, H) and phalanges (B, C, D, F, G, I) in proximal(a, e, h), medial (far left), anterior (‘), lateral (‘‘) and posterior (‘‘‘, far right). Scale bar: 5 cm.
➝
➝
229GEODIVERSITAS • 2000 • 22 (2)
suggests that the expanded metacarpal ends prob-ably have functional significance.The first metacarpal is 62 mm in length. Its prox-imal and distal heads expand to 50 mm, while theshaft is a flattened oval in section (36 mm trans-versely, 24 mm anteroposteriorly). In compari-son, one of the larger first metacarpals from theCleveland-Lloyd collection of Allosaurus elementsis 68 mm long with a proximal expansion of 47,shaft diameters of 45 × 36, and a distal expansionof 53 mm. The proximal end in Acrocanthosaurusis closely applied to the lateral surface of the sec-ond metacarpal. The roughly triangular proximalarticular surface (50 mm lateromedially, 47 mmanteroposteriorly compared with 45 × 50 mm inAllosaurus) is complex (Fig. 12A). The lateral halfslopes lateroventrally, especially where it articu-lates with the second metacarpal. This articula-tion consists mainly of a deep depression close tothe flexor surface, but it extends anteriorly to theextensor surface as well. The posteromedial face is
emarginated for a distinct, smooth-walled depres-sion that is continuous with an emargination onthe posteromedial edge of the first distal carpal.This is continuous with the “trough” of the pul-ley-like surface of the distal carpal. It may repre-sent the end of a sliding articulation of the“pulley-like” wrist joint. As pointed out, a canalfor a blood vessel or nerve enters the regionbetween these two bones on the lateral side ofthis depression. This canal becomes more obvi-ous on the metacarpal as it passes forward andsubdivides, forming a deep trough that cuts intothe proximal surface of the metacarpal on theanterior (flexor) surface where the bone is emar-ginated. The Y-shaped system divides the proxi-mal surface into three articular surfaces for thefirst distal carpal. The deep lateral depression forthe second metacarpal and the posteromedialdepression for the end of the “trough” of the“pulley” are characteristic of all avetheropodans(= carnosaurs + coelurosaurs).
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
FIG. 12. — Acrocanthosaurus atokensis NCSM 14345, right manus. Metacarpals (A, E, H) and phalanges (B, C, D, F, G, I) in proximal(a, e, h), medial (far left), anterior (‘), lateral (‘‘) and posterior (‘‘‘, far right). Scale bar: 5 cm.
There are prominent tubercles with rugosities forthe attachment of ligaments in the proximolateral(Fig. 12H”) and distomedial regions of the exten-sor surface (Fig. 12H’). The distal condyles areseparated by a deep sulcus (Fig. 12H’). The artic-ular surfaces extend high on the extensor surface,allowing the digit to hyperextend until its shaft isalmost perpendicular to the shaft of themetacarpal. In contrast, the first phalanx couldnot flex much more than 30 degrees. Thecondyles are asymmetrical so that the first pha-lanx would have rotated somewhat during exten-sion, when the tip of the claw would have turnedmore medially. In the degree of rotation and theasymmetry of the condyles, Acrocanthosaurusseems to be different from Allosaurus (Madsen1976). There is a deep ligament pit distolaterally(Fig. 12H”), but there is not one medial to themedial condyle (Fig. 12H).The second metacarpal is 116mm long, with a max-imum proximal breath of 74.5 mm, a transverseshaft diameter of 32 mm, and a distal width of 69.5.In comparison, one of the larger second metacarpalsfrom the Cleveland-Lloyd collection of Allosauruselements is 120 mm long with a proximal expan-sion of 58, a shaft width of 34 and a distal expan-sion of 56mm. In proximal outline (Figs11A; 12E),the lateral, medial and flexor edges are concave, andthe extensor margin is mostly convex. The proxi-mal articular surface extends onto the medial sur-face posteriorly (Fig. 12E) to form a surprising smallfacet that articulates with the first metacarpal. Onthe lateral surface, there are two rugose surfaces onthe proximal end for a firm, almost suture-like con-tact with metacarpal III (Fig. 12E”). There is a3 mm wide canal between the two surfaces thatundoubtedly was used by blood vessels and/ornerves (Fig. 12E”). The course of the canal starts ina deep notch in the anteromedial surface of the thirdmetacarpal (Fig. 12A’), continues distally in thecanal on metacarpal II, and exits on the flexor sur-face distal to the contact between metacarpals II andIII, just proximal to a prominent ligament tubercle(Fig. 12E’’’). Shallow depressions are all that repre-sent the colateral ligament pits on the medial andlateral condyles. These are highly developed inAllosaurus (Madsen 1976), Sinraptor (Currie &
Zhao 1993) and most other large theropods. Thecondyles are highly asymmetrical (Fig. 12E’), thelateral one having a much greater anteroposteriorsurface. The joint allows the first phalanx of the sec-ond digit to hyperextend as much as it flexes (about40 degrees), and turns the digit so that the tip ofthe claw would have rotated medially during flex-ion and laterally during hyperextension. As in thefirst metacarpal, the disparity in distal condyle sizeand function is much greater than it is in Allosaurus.The third metacarpal is 89 mm long, about53 mm across proximally, has a shaft diameter of22 mm, and a distal width of 39 mm. The equiva-lent measurements in a large Allosaurus are 105(length), 38 (proximal), 12 (shaft diameter) and29 mm (distal width). The bone has a pronouncedposteromedial expansion (Fig. 12A) where it con-tacts the second metacarpal in what must havebeen a virtually immobile joint. There is a suture-like contact with the adjacent metacarpal on theposteromedial expansion, bound anteriorly by awell-defined canal for a blood vessel or nerve(Fig. 12A). The canal divides, part of it passingdistoanteriorly into the sutural contact, and theother branch transferring distomedially onto thelateral surface of the second metacarpal(Fig. 12E”). When articulated, the long axis of thethird metacarpal is rotated posteriorly so that thecentre of the distal end of the bone is about 3 cmbehind the centre of metacarpal II.The length of the phalanges of the second digit isalmost 90% the length of the humerus. The thirddigit is 93% as long as the first, which in turn is73% the length of the second. These proportionsare very close to what is found in Allosaurus(USNM 4734, Gilmore 1920), which suggests thatthe stocky appearance of the hand ofAcrocanthosaurus is a function of allometricchanges. As in Allosaurus, the longest phalanx(excluding the unguals) is I-1, followed by II-2,II-1 and III-3. Phalanges I-1 and II-1 have gingly-moid articulations with their metacarpals, whereasIII-1 has a shallowly concave, undivided articula-tion. Collateral ligament pits are shallow andpoorly defined on the manual phalanges (Fig. 12).Although the femur is incomplete (Fig. 13), it wasat least 110 cm long. The length of the femur can
Currie P. J. & Carpenter K.
230 GEODIVERSITAS • 2000 • 22 (2)
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
231GEODIVERSITAS • 2000 • 22 (2)
be estimated as 128 cm by comparison with previ-ously described specimens of Acrocanthosaurus(Stovall & Langston 1950; Harris 1998a). As in othercarnosaurs, the relatively straight shaft of the femuris pierced by a nutrient foramen proximal to thefourth trochanter. Like Allosaurus, Sinraptor andother carnosaurs, the fourth trochanter is representedby a low but prominent ridge next to the depressionfor the M.caudifemoralis longus. The minimum trans-verse diameter of the shaft is 15 cm, which is less thanthe same dimension in adult specimens ofTyrannosaurus (BHI 3033, MOR 555,TMP 81.12.1). This indicates that Acrocanthosaurus
was a lighter animal than Tyrannosaurus, even thoughthe overall length of the body was similar. The distalend of the femur has a ridge along the medial mar-gin of the anterodorsal surface (Fig. 13B) similar toAllosaurus (Madsen 1976), but not as sharply definedas those of Giganotosaurus (MUCPv-CH-1) andSinraptor (Currie & Zhao 1993). The ridge boundsthe adductor fossa medially. The fossa is continuouswith the intercondylar trough of the distal end. Thereare well-developed distal condyles (Fig. 13C), the lat-eral one associated with a distinct crista tibiofibularis.The tibia is incomplete. However, similarity in sizeof the distal end of the tibia and the lengths of the
FIG. 13. — Acrocanthosaurus atokensis NCSM 14345, right hindlimb elements; A-C, femur ; A, lateral view; B, anterior view; C, distalview; D, tibia, astragalus and calcaneum in anterior aspect; E, proximal head of tibia in medial view. Scale bar: 10 cm.
Currie P. J. & Carpenter K.
232 GEODIVERSITAS • 2000 • 22 (2)
FIG. 14. — Acrocanthosaurus atokensis NCSM 14345, right pes; A, proximal aspect of metatarsus; B, anterior view of preserved ele-ments; C, anteromedial view of fifth metatarsal; D, medial views of digit IV; E, medial views of digit III; F, medial views of digit II.Scale bar: 10 cm.
➝
second metatarsals suggests that it would have beenclose to the same length as that of OMNH 10147(96 cm). There is a well-developed cnemial crest(Fig. 13D, E) and a fibular crest. The tibia flaresdistally in anterior view (Fig. 13D) where it con-tacts the astragalus and calcaneum. The anteriorsurface is flat to accommodate the ascendingprocess of the astragalus. Only the medial portionof the main body of the astragalus was recovered.Overall it appears to have been quite similar to thatof Allosaurus with a relatively high, proximolater-ally sloping ascending process. The calcaneum isrelatively large disk of bone (122 mm anteropos-teriorly, 66 mm mediolaterally and 99 mm high)with a distinct process that plugged into a depres-sion in the lateral margin of the astragalus as inAllosaurus and Sinraptor.The first, second, and fifth metatarsals are com-plete, but only the proximal ends of the third andfourth ones were recovered (Fig. 14). The secondmetatarsal (41 cm) is 99% of the length of that ofOMNH 10147, so the total length of metatarsal IIIcan be estimated as 44 cm. Only the smallest spec-imens of tyrannosaurids have such short thirdmetatarsals. For example, TMP 91.36.500 is a5.1 m long skeleton of Gorgosaurus libratus that hasa 46 cm long metatarsal III. The length themetatarsus does, however, fall within the rangeexpected for all other large theropods. Generallythe foot is closely comparable to those of Allosaurusand Sinraptor, but is more robust. In proximalview, the outlines of the metatarsals (Fig. 14A) arealmost identical to those of Allosaurus.Digit I of the pes, which is complete, is 15 cmlong. The first two phalanges of pedal digit III are16.0 and 11.5 cm, suggesting that the total lengthof the toe would have been about half a metre.The fourth toe is missing the last two phalanges.The unguals have triangular cross-sections andresemble the pedal unguals of other carnosaurs.
DISCUSSION
Acrocanthosaurus is one of the largest theropodsknown. Amongst the four described specimens,NCSM 14345 is larger than either the holotype
(OMNH 10146) or SMU 74646. It appears to havebeen about the same size as OMNH 10147, whichhas a 958 mm long tibia (Stovall & Langston 1950).In total length, NCSM 14345 was comparable withGiganotosaurus (Coria & Salgado 1995) andTyrannosaurus. Femur length, a relatively stablemeasurement for gauging overall size (Christiansen1998), suggests it was bigger than the largest knownspecimens of Carcharodontosaurus (Stromer 1931),Saurophaganax (Chure 1996), Suchomimus Serenoet al., 1998 and Tarbosaurus (GI 107/2), but wassmaller than Giganotosaurus (MUCPv-CH-1) andTyrannosaurus (at least five specimens). In terms ofskull size (the premaxilla to quadrate length ofNCSM 14345 is 123 cm), Acrocanthosaurus is sur-passed by one specimen of Tarbosaurus (PIN 551-1has a skull 135 cm long), five specimens ofTyrannosaurus (with skull lengths of 127 to 153cm),Carcharodontosaurus (estimated length of 160 cm,Sereno et al. 1996) and Giganotosaurus (estimatedlength of MUCPv-CH-1 is 160 cm). Finally, thecircumference of the femur (425 mm inNCSM 14345) can be used to estimate weight(Anderson et al. 1985), suggesting that the new spec-imen of Acrocanthosaurus was heavier at 2.40 met-ric tons than most specimens of Tarbosaurus and allbut one (NMMNH P-26083, Williamson & Chure1996) of Allosaurus, weighed a little less thanSaurophaganax (Chure 1996), and was lighter thanCarcharodontosaurus, Giganotosaurus (4.16 metrictons) and Tyrannosaurus (weight estimate ofMOR 555 is also 4.16 metric tons).The total length of Acrocanthosaurus is comparablewith Tyrannosaurus because like Allosaurus, it has rel-atively longer vertebrae when compared with the skullor femur. For example, the 19th presacral vertebralcentrum of Tyrannosaurus (MOR 555) is 148 mmlong and 180 mm wide. The widths of the centrashould be proportional to the weights of the animals,and should scale to the two thirds power of lengths(Currie 1978). Because the femur of SMU 74646(Acrocanthosaurus) is 0.85 the length of the femur ofthe Tyrannosaurus, one would expect that the widthof the dorsal vertebra of the smaller animal should be0.852/3 (= 0.78) that of the width of the larger ani-mal. The actual width of the 19th centrum ofAcrocanthosaurus is 138 mm, which is 0.77 the width
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
233GEODIVERSITAS • 2000 • 22 (2)
of the Tyrannosaurus centrum, very close to the pre-dicted value. The length of the vertebra should scalelinearly with femur length, and therefore should be0.85 time (126 mm) the length of the Tyrannosauruscentrum. In fact it is 135 mm long in Acrocantho-saurus, showing that the dorsal vertebrae are signifi-cantly longer than in tyrannosaurs. Another way ofdemonstrating the relatively longer lengths ofAcrocanthosaurus vertebrae is by examining their rel-ative proportions. In large tyrannosaurs, the presacralvertebrae are shorter than they are wide, whereas incarnosaurs they are always longer than wide. Overall, Acrocanthosaurus compares most closelymorphologically with Allosaurus. Acrocanthosauruswas assigned to the Allosauridae by Stovall &Langston (1950). It is more primitive thanAllosaurus in that it has only four premaxillary teethlike most other theropods, whereas allosaurids havefive. The jugal takes part in the border of the antor-bital fenestra, a character listed by Holtz (1994) asa derived condition. However, it is far more likelythat this is the primitive condition because it ispresent in Herrerasaurus and most theropods. InAllosaurus, the jugal is excluded from the border ofthe antorbital fenestra by the maxilla and lacrimal.Acrocanthosaurus is also more primitive thanAllosaurus in having a tall quadrate, in lacking adownturned paroccipital process, and in lackingthe neomorph antarticular (Madsen 1976).Furthermore, the surangular of Acrocanthosaurusdoes not contact the angular in front of the exter-nal mandibular fenestra. Unlike Allosaurus but likemost other theropods, the posterior end of theangular terminates anterior to the posterior suran-gular foramen, which is relatively small. The neu-ral spines of the vertebrae are relatively muchhigher than in any other theropod exceptSpinosaurus. Pleurocoels are found in at least thefirst 21 presacral vertebrae (Harris 1998a), but areonly found in the cervicals and first four or fivedorsals of Allosaurus. The arms of Acro-canthosaurus, especially the forearm and metacar-pus, are shorter than those of Allosaurus.Acrocanthosaurus was assigned to the Carcharo-dontosauridae by Sereno et al. (1996), a viewsupported by a more extensive analysis by Harris(1998a). However, Harris (1998a) went on to
point out problems with paleogeographic inter-pretations if Acrocanthosaurus is related to Car-charodontosaurus and Giganotosaurus. In fact, there are few characters that can be used tounite Acrocanthosaurus with Carcharodontosaurusand Giganotosaurus. Sereno et al. (1996) used eightcharacters to define the Carcharodontosauridae. Ofthese, the broad postorbital-lacrimal contact is prob-ably size-related. Although both Acrocanthosaurusand carcharodontosaurids have pronounced shelvesof bone over the orbits, this is also true of abelisauridsand big tyrannosaurids. In Acrocanthosaurus, thisshelf is formed by both the prefrontal and lacrimal,which extend backwards over the eye to contact thepostorbital. In the carcharodontosauridGiganotosaurus (R. Coria, pers. comm. 1998) andpossibly Tyrannosaurus (N. Larson, pers. comm.1997), the gap is bridged by a new bone, possiblysimilar to the ornithischian palpebral. This suggestscarcharodontosaurids are roofing over the orbit in adifferent way than Acrocanthosaurus. The suborbitalf lange found on the postorbital ofCarcharodontosaurus can also be seen inGiganotosaurus (Coria & Salgado 1995), abelisaurids(Bonaparte 1991; Sampson et al. 1998), some tyran-nosaurids, and to a lesser extent in CryolophosaurusHammer & Hickerson, 1994. The holotype ofAcrocanthosaurus has a postorbital flange (Stovall &Langston 1950), but this feature is not evident in thenew specimen. The ventral extension of the basisphe-noid that unites Giganotosaurus withCarcharodontosaurus is not present inAcrocanthosaurus. A pronounced process thatextends the ventral end of the intermandibular sym-physis is supposedly a character shared by the threetaxa. However, there are no jaws ofCarcharodontosaurus that are complete enough toshow this, and it is no more pronounced inNCSM 14345 than it is in large specimens ofTyrannosaurus rex. The mid-cervical vertebrae of car-charodontosaurids are supposed to be at least 20%broader than tall. This may be true forCarcharodontosaurus (Sereno et al. 1996), but not forGiganotosaurus (MUCPv-CH-1) which has almostround cervical centra in end view. Furthermore, thecervical centra of Acrocanthosaurus are only slightlywider than they are high (Stovall & Langston 1950;
Currie P. J. & Carpenter K.
234 GEODIVERSITAS • 2000 • 22 (2)
Harris 1998a). The degree of elevation of the ante-rior intervertebral articulation in midcervical verte-brae is determined by neck posture, and is widelyvariable throughout the Theropoda. Although thereare depressions in the lateral surfaces of the proximalcaudal centra, they are not pneumatic inAcrocanthosaurus (Harris 1998a). Pleurocoels havebeen reported in the caudal vertebrae ofCarcharodontosaurus and Bahariasaurus Stromer,1931 (Rauhut 1995), but are not present in the cau-dals of Giganotosaurus (R. Coria, pers. comm.). Thepubic boot of carcharodontosaurids, according toSereno et al. (1996), is 30% of the total length of thebone. Similar pubic proportions are found in a widerange of theropods, including Allosaurus,ornithomimids, oviraptorids and tyrannosaurs. The analysis by Harris (1998a) unitedAcrocanthosaurus with the Carcharodontosauridaeon the basis of the reniform cervical centra (at least20% wider than tall), but did state that the featureis much more accentuated in Carcharodontosaurus.Russell (1996) set up a new genus (Sigilmassasaurus)for a Moroccan animal with reniform cervical cen-tra, and described a posterior cervical ofCarcharodontosaurus with an essentially round cen-trum in posterior view. Furthermore, as pointed outpreviously, the cervical centra of Giganotosaurus areround in posterior view, not reniform. This charac-ter, therefore, is a very weak one for placingAcrocanthosaurus in the Carcharodontosauridae.There are many differences between Acrocantho-saurus and the two unquestionable carcharodon-tosaurids — Carcharodontosaurus and Giganoto-saurus. Acrocanthosaurus lacks the pronouncedsculpturing on the maxilla, and has an antorbital fossathat extends farther beyond the bounds of the antor-bital fenestra than the situation in carcharodon-tosaurids. Carcharodontosaurids lack the maxillaryfenestra that is found in most tetanurine theropods,including Acrocanthosaurus. The braincase of Acro-canthosaurus (Welles et al. in prep.) is similar to thatof Allosaurus, and strikingly different from the highlymodified braincases of carcharodontosaurids(Larsson 1996; Coria & Currie in prep.). The lateraltemporal fenestrae of Acrocanthosaurus andAllosaurusare similar in outline, and contrast stronglywith the relatively larger openings of carcharodon-
tosaurids and other more primitive theropods.Correlated with this, the quadrates of carcharodon-tosaurids are positioned much farther behind theocciput than they are in Acrocanthosaurus. This is themajor reason that carcharodontosaurid skulls are somuch longer than the skulls of other theropods,including Acrocanthosaurus. The occiput ofAcrocanthosaurus is nearly vertical in contrast withthe strongly sloping posterior occipital region of car-charodontosaurids. There is a double boss on thesupraoccipital of Acrocanthosaurus, compared withthe more normal medial boss of a carcharodon-tosaurid supraoccipital. There are also many differ-ences in the postcranial skeleton, although few are sostriking as those of the femur. Whereas the femur ofAcrocanthosaurus is closely comparable with that ofAllosaurus (Stovall & Langston 1950), femora ofGiganotosaurus (MUCPv-CH-1) and Carcharodon-tosaurus (Stromer 1931) are more primitive inappearance with the heads angled upwards from theshaft (not perpendicular) and lesser trochanters thatare well below proximal head level. Novas (1997)pointed out other features in carcharodontosauridskeletons that may indicate that this family is moreclosely related to Abelisauridae than Allosauridae.We set up a data matrix (Appendix 2) based mostlyon that used by Harris (1998a) in his analysis of car-charodontosaurids. Some corrections and additionswere made to his character list, mostly because of thenew information provided by NCSM 14345 andnew specimens of Giganotosaurus from Argentina.The matrix consisted of 42 cranial , 24 axial and 44 appendicular characters(Appendix 1). We also included some different taxain our analysis. Herrerasaurus was retained as the out-group to all other theropods, and the Abelisauridaewas included in the analysis as a second outgroup.The data matrix was analyzed using the beta versionof PAUP 4.0b2 (Swofford 1998). The Branch-and-Bound search method produced three equally parsi-monious trees (tree length = 235, C.I. = 0.638, R.I. =0.600, R.C. = 0.382). Both accelerated and delayedtransformations (Acctran & Deltran) were per-formed for character state optimization. MacClade3.07 (Maddison & Maddison 1992) was used toproduce Figure 15. The results of this analysis showthat, in all three trees, Allosaurus is the closest thero-
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
235GEODIVERSITAS • 2000 • 22 (2)
Currie P. J. & Carpenter K.
236 GEODIVERSITAS • 2000 • 22 (2)
pod to Acrocanthosaurus, and that Giganotosaurusand Carcharodontosaurus form a second clade. Thetrees differ only in inter-relationships of theAllosauridae, Carcharodontosauridae andSinraptoridae. Padian et al. (1997) defined theAllosauroidea as all descendants of the most recentcommon ancestor of Allosaurus and Sinraptor. In thepresent analysis, it could not be determined if car-charodontosaurids should be included within theAllosauroidea.
AcknowledgementsThe authors were given access to the original spec-imen by Allen Graffham of Geological Enterprises,Inc. (Ardmore, Oklahoma), and were assisted intheir research by staff of the Black Hills Institutefor Geological Research, Inc. (Hill City, SouthDakota). Peter Larson was particularly helpful inHill City. We are very grateful to Dr Thomas Holtz(University of Maryland), Dr Wann Langston, Jr.
(University of Texas, Austin), and Dr Dale A.Russell (North Carolina State University) whoreviewed various drafts of this paper. Dr LouisJacobs (Southern Methodist University) and DrRichard Cifelli (University of Oklahoma) werekind enough to provide access to specimens in theircare. With the exception of Figure 2 (provided bythe Black Hills Institute), all of the illustrationswere done by the second author. We thank CliveCoy and Dr Eva Koppelhus (Tyrrell Museum) forall of their help in completing the manuscript.
REFERENCES
Anderson J. F., Hall-Martin A. & Russell D. A. 1985. —Long-bone circumference and weight in mammals,birds and dinosaurs. Journal of Zoology (A) 207: 53-61.
Bakker R. T., Kralis D., Siegwarth J. & Filla J. 1992. —Edmarka rex, a new, gigantic theropod dinosaur fromthe middle Morrison Formation, Late Jurassic of theComo Bluff outcrop region. Hunteria 2 (9):1-24.
Herre
rasa
urus
Abeli
saur
idae
Sinr
apto
ridae
Acroc
anth
osau
rus
Allosa
urus
Gigan
otos
auru
sCar
char
odon
tosa
urus
Mon
olop
hosa
urus
Tyra
nnos
aurid
aeOvir
apto
rosa
uria
Drom
aeos
aurid
ae
FIG. 15. — Strict consensus of three trees generated in analysis of phylogenetic relationships of Acrocanthosaurus using 110 mor-phological characters.
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
237GEODIVERSITAS • 2000 • 22 (2)
Bakker R. T., Williams M. & Currie P. J. 1988. —Nanotyrannus, a new genus of pygmy tyrannosaur,from the latest Cretaceous of Montana. Hunteria 1(5): 1-30.
Bonaparte J. F. 1991. — The Gondwanian theropodfamilies Abelisauridae and Noasauridae. HistoricalBiology 5: 1-25.
Bonaparte J. F. & Novas F. E. 1985. — Abelisaurus com-ahuensis, n. gen, n. sp., Carnosauria del CretacicoTardio de Patagonia. Ameghiniana 21: 259-265.
Bonaparte J. F., Novas F. E. & Coria R. A. 1990. —Carnotaurus sastrei Bonaparte, the horned, lightlybuilt carnosaur from the Middle Cretaceous ofPatagonia. Natural History Museum of Los AngelesCounty, Contributions in Science 416: 1-41.
Britt B. B. 1991. — The theropods of the Dry MesaQuarry (Morrison Formation), Colorado: withemphasis on the osteology of Torvosaurus tanneri.Brigham Young University, Geology Studies 37: 1-72.
Britt B. B. 1993. — Pneumatic Postcranial Bones inDinosaurs and Other Archosaurs. PhD thesis,University of Calgary, Calgary, Canada, 383 p.
Chatterjee S. 1978. — Indosuchus and Indosaurus,Cretaceous carnosaurs from India. Journal ofPaleontology 52: 570-580.
Christiansen P. 1998. — Biomechanical and BiometricAspects of the Appendicular Skeleton of Non-AvianTheropods (Dinosauria: Theropoda) in comparisonwith Extant Mammalia. PhD thesis, University ofCopenhagen, Dept of Historical Geology andPalaeontology, Copenhagen, The Netherlands.
Chure D. J. 1996. — A reassessment of the gigantictheropod Saurophagus maximus from the MorrisonFormation (Upper Jurassic) of Oklahoma, USA :103-106 in Sun S. & Wang Y. (eds), Sixth Symposiumon Mesozoic Terrestrial Ecosystems and Biota, ShortPapers. China Ocean Press, Beijing.
Cifelli R. L., Gardner J. D., Nydam R. L. & BrinkmanD. L. 1997. — Additions to the vertebrate fauna ofthe Antlers Formation (Lower Cretaceous), south-eastern Oklahoma. Oklahoma Geology Notes 57 (4):124-131.
Coria R. A. & Currie P. J. in review. — The braincaseof Giganotosaurus carolinii (Dinosauria: Theropoda)from the Upper Cretaceous of Argentina.
Coria R. A. & Salgado L. 1995. — A new giant car-nivorous dinosaur from the Cretaceous of Patagonia.Nature 377: 224-226.
Currie P. J. 1978. — The orthometric linear unit.Journal of Paleontology 52: 964-971.
Currie P. J. 1985. — Cranial anatomy of Steno-nychosaurus inequalis (Saurischia, Theropoda) and itsbearing on the origin of birds. Canadian Journal ofEarth Sciences 22: 1643-1658.
Currie P. J. 1995. — New information on the anatomyand relationships of Dromaeosaurus albertensis(Dinosauria: Theropoda). Journal of VertebratePaleontology 15: 576-591.
Currie P. J., Rigby K. Jr & Sloan R. E. 1990. —Theropod teeth from the Judith River Formation ofsouthern Alberta, Canada : 107-125, in Carpenter K.& Currie P. J. (eds), Dinosaur Systematics: Approachesand Perspectives. Cambridge University Press, NewYork.
Currie P. J. & Zhao X. J. 1993. — A new large thero-pod (Dinosauria, Theropoda) from the Jurassic ofXinjiang, People’s Republic of China. CanadianJournal of Earth Sciences 30: 2037-2081.
Dong Z. M., Zhang Y. H., Li X. & Zhou S. W. 1978. —Note on a new carnosaur Yangchuanosaurusshangyuensis gen. et sp. nov. from the Jurassic ofYangchuan District, Sichuan Province. KexueTongbao 23: 298-302 (in Chinese).
Dong Z. M., Zhou S. W. & Zhang Y. H. 1983. — Thedinosaurian remains from Sichuan Basin, China.Palaeontologia Sinica, New Series C 23, 162, 145 p.,43 plates (in Chinese).
Gao Y.-H. 1993. — A new species of Szechuanosaurusfrom the Middle Jurassic of Dashanpu, Zigong,Sichuan. Vertebrata PalAsiatica 31: 308-314.
Gilmore G. W. 1920. — Osteology of the carnivorousDinosauria in the United States National Museum,with special reference to the genera Antrodemus(Allosaurus) and Ceratosaurus. United States NationalMuseum, Bulletin 110: 1-159.
Hammer W. R. & Hickerson W. J. 1994. — A crestedtheropod dinosaur from Antarctica. Science 264: 828-830.
Harris J. D. 1998a. — A reanalysis of Acrocanthosaurusatokensis, its phylogenetic status, and paleobiogeo-graphic implications, based on a new specimen fromTexas. New Mexico Museum of Natural History andScience, Bulletin 13, 75 p.
Harris J. D. 1998b. — Large, Early Cretaceoustheropods in North America. New Mexico Museumof Natural History and Science, Bulletin 14: 225-228.
Holtz T. R. Jr. 1994. — The phylogenetic position ofthe Tyrannosauridae: implications for theropod sys-tematics. Journal of Paleontology 68: 1100-1117.
Hutt S., Martill D. M. & Barker M. J. 1996. — Thefirst European allosaurid dinosaur (LowerCretaceous, Wealden Group, England). NeuesJahrbuch für Geologie und Paläontologie Monatshefte10: 635-644.
Lambe L. M. 1917. — The Cretaceous theropodousdinosaur Gorgosaurus. Geological Survey of Canada,Memoir 100: 1-84.
Larsson H. C. E. 1998. — Cranial morphology of theAfrican theropod, Carcharodontosaurus saharicus(Allosauroidea). Journal of Vertebrate Paleontology 16:47A.
Lipka T. R. 1998. — The affinities of the enigmatictheropods of the Arundel Clay facies (Aptian),Potomac Formation, Atlantic coastal plain ofMaryland. New Mexico Museum of Natural Historyand Science, Bulletin 14: 229-234.
Maddison W. P. & Maddison D. R. 1992. —MacClade:Analysis of Phylogeny and Character Evolution, version3.01. Sinauer Associates, Sunderland, Massachusetts.
Madsen J. H. 1976. — Allosaurus fragilis: a revised oste-ology. Utah Geological and Mineral Survey, Bulletin,109 xii + 163 p.
Maleev E. A. 1974. — Gigantic carnosaurs of the fam-ily Tyrannosauridae. Joint Soviet-MongolianPaleontological Expedition, Transactions 1: 132-191(in Russian).
Molnar R. E. 1991. — The cranial morphology ofTyrannosaurus rex. Paleontographica A 217: 137-176.
Novas F. E. 1997. — Abelisauridae : 1-2, in Currie P. J.& Padian K. (eds), Encyclopedia of Dinosaurs.Academic Press, San Diego.
Osborn H. F. 1912. — Crania of Tyrannosaurus andAllosaurus. American Museum of Natural History,Memoirs, New Series, 1: 1-30.
Osborn H. F. 1917. — Skeletal adaptations ofOrnitholestes, Struthiomimus, Tyrannosaurus.American Museum of Natural History, Bulletin 35:733-771.
Osmólska H. 1985. — Antorbital fenestra of archosaursand its suggested function. Vertebrate Morphology 30:159-162.
Padian K. & Chiappe L. M. 1997. — Bird origins: 71-79, in Currie P. J. & Padian K. (eds), Encyclopedia ofDinosaurs. Academic Press, San Diego.
Padian K. & Hutchinson J. 1997. — Allosauroidea: 6-9, in Currie P. J. & Padian K. (eds), Encyclopedia ofDinosaurs. Academic Press, San Diego.
Rauhut O. W. M. 1995. — Zur systematischen Stellungder afrikanischen Theropoden CarcharodontosaurusStromer 1931 und Bahariasaurus Stromer 1934.Berliner geowissenshaft Abhandlungen E16: 357-375.
Russell D. A. 1996. — Isolated Dinosaur bones fromthe Middle Cretaceous of the Tafilalt, Morocco.Bulletin du Muséum national d’Histoire naturelle, série4, C 18 (2-3) : 349-402.
Russell D. A. & Dong Z. M. 1993. — The affinities ofa new theropod from the Alxa Desert, InnerMongolia, People’s Republic of China. CanadianJournal of Earth Sciences 30: 2107-2127.
Sampson S. D., Witmer L. D., Forster C. A., KrauseD. W., O’Connor P. M., Dodson P. & RavoavyF. 1998. — Predatory dinosaur remains fromMadagascar: implications for the Cretaceous bio-geography of Gondwana. Science 280: 1048-1051.
Sereno P. C. 1993. — The pectoral girdle and forelimbof the basal theropod Herrerasaurus ischigualastensis.Journal of Vertebrate Paleontology 13: 425-450.
Sereno P. C., Beck A. L., Dutheil D. B., Gado B.,Larsson H. C. E., Lyon G. H., Marcot J. D., Rauhut
O. W. M., Sadleir R. W., Sidor C. A., VarricchioD. D., Wilson G. P. & Wilson J. A. 1998. — A long-snouted predatory dinosaur from Africa and the evo-lution of spinosaurids. Science 282: 1298-1302.
Sereno P. C., Dutheil D. B., Iarochene S. M., LarssonH. C. E., Lyon G. H., Magwene P. M., Sidor C. A.,Varricchio D.J. & Wilson J.A. 1996.—Late Cretaceousdinosaurs from the Sahara. Science 272: 986-991.
Sereno P. C. & Novas R. E. 1992. — The completeskull and skeleton of an early dinosaur. Science 258:1137-1140.
Sereno P. C., Wilson J. A., Larsson H. C. E., DutheilD. B. & Sues H.-D. 1994. — Early Cretaceousdinosaurs from the Sahara. Science 266: 267-271.
Stovall J. W. & Langston W. L. Jr. 1950. —Acrocanthosaurus atokensis, a new genus and speciesof Lower Cretaceous Theropoda from Oklahoma.American Midland Naturalist 43: 696-728.
Stromer E. 1931. — Ergebnisse der ForschungsreisenProf. E. Stromers in den Wüsten Ägyptens. II.Wirbeltier-Reste der Bajarîjestufe (unterstesCenoman). 10. Ein Skelett-Rest von Car-charodontosaurus nov. gen. Abhandlungen derBayerischen Akademie der Wissenschaften Mathematik-Naturwissenschaftern 9: 1-23.
Swofford D. L. 1998. — PAUP 4.0, beta version. SinauerAssociates, Sunderland, Massachusetts.
Taquet P. & Welles S. P. 1977. — Redescription ducrâne de dinosaure théropode de Dives (Normandie).Annales de Paléontologie (Vertébrés) 63: 191-206.
Walker A. D. 1964. — Triassic reptiles from the Elginarea: Ornithosuchus and the origin of carnosaurs.Philosophical Transactions (B) 248: 53-134.
Welles S. P., Langston W. Jr. & Currie P. J. in prep. —The braincase of the Early Cretaceous theropod
dinosaur Acrocanthosaurus atokensisStovall & Langston.Williamson T. E. & Chure D. J. 1996. — A large
allosaurid from the Upper Jurassic Morrison Formation(Brushy Basin Member), West-central New Mexico.Museum of Northern Arizona Bulletin 60: 73-79.
Witmer L. M. 1997. — The evolution of the antorbitalcavity of archosaurs: a study in soft-tissue recon-struction in the fossil record with an analysis of thefunction of pneumaticity. Society of VertebratePaleontology Memoir 3, 73 p.
Woodward A. S. 1910. — On a skull of Megalosaurusfrom the Great Oolite of Minchinhampton(Gloucestershire). Quarterly Journal of the GeologicalSociety of London, 66: 111-115
Zhao X. J. & Currie P. J. 1993. — A large crested thero-pod from the Jurassic of Xinjiang People’s Republicof China. Canadian Journal of Earth, Sciences 30:2027-2036.
Submitted for publication on 26 November 1998;accepted on 3 June 1999
Currie P. J. & Carpenter K.
238 GEODIVERSITAS • 2000 • 22 (2)
Acrocanthosaurus atokensis from Antlers Formation, Oklahoma, USA
239GEODIVERSITAS • 2000 • 22 (2)
.1. Premaxilla, shape of symphysis from below:0 - V-shaped.1 - U-shaped.
6. Facial bones (maxilla, nasal), sculpturing:0 - moderate.1 - heavily sculptured to edge of antorbital fenestra.
7. Nasal, participates in antorbital fossa:0 - no or slightly.1 - broadly.
8. Nasals, fused on midline:0 - no.1 - yes.
9. Prefrontal:0 - large.1 - reduced.2 - absent.
10. Postorbital, ventral end above ventral margin of orbit:
0 - yes.
1 - no.
11. Postorbital-lacrimal contact:
0 - absent.
1 - present.
12. Postorbital, suborbital flange:
0 - absent.
1 - small.
2 - large.
13. Lacrimal pneumatic recess:
0 - absent.
1 - present.
14. Lacrimal horn:
0 - non-existent.
1 - low crest or ridge.
2 - high-pointed cone.
15. Jugal pneumatic:
0 - no.
1 - yes.
16. Jugal, foramen on medial surface:
0 - absent.
1 - present.
17. Jugal, expressed on rim of antorbital fenestra:
0 - no.
1 - yes.
18. Jugal, qj process, length of upper prong to lower:
0 - subequal.
1 - upper shorter.
2 - upper longer.
APPENDIX 1
Morphological characters (all unordered) used in this paper, most of which are derived from Holtz (1994) and Harris (1998). “0” represents the primitive state.
Currie P. J. & Carpenter K.
240 GEODIVERSITAS • 2000 • 22 (2)
19. Preorbital bar, suborbital process:
0 - not present.
1 - present.
20. Quadrate short:
0 - no.
1 - yes.
21. Quadrate fenestra:
0 - none.
1 - between quadrate and quadratojugal.
2 - surrounded by quadrate.
22. Orbit, expanded and circular:
0 - no.
1 - yes.
23. Braincase, pneumatism:
0 - apneumatic.
1 - moderately.
2 - highly pneumatic.
24. Basioccipital participates in basal tubera:
0 - yes.
1 - no.
25. Occiput:
0 - nearly vertical.
1 - slopes posteroventrally.
26. Paroccipital processes downturned distally:
0 - no.
1 - moderate.
2 - distal ends below level of foramen magnum.
27. Exoccipital-opisthotic, posteroventral limit of con-tact with basisphenoid separated from basal tuberaby notch:
0 - no.
1 - yes.
28. Trigeminal nerve, separation of ophthalmic branch:



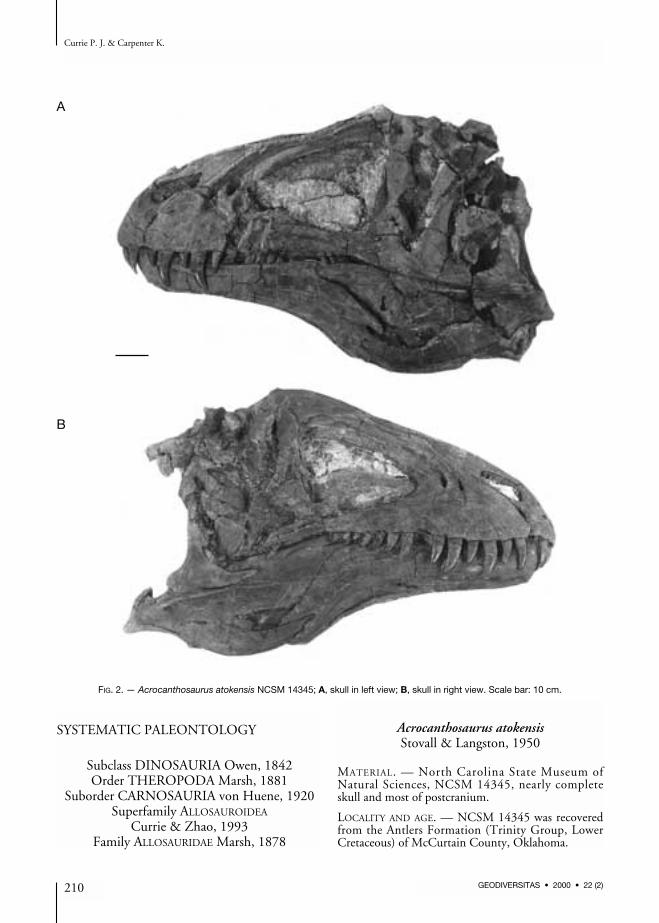



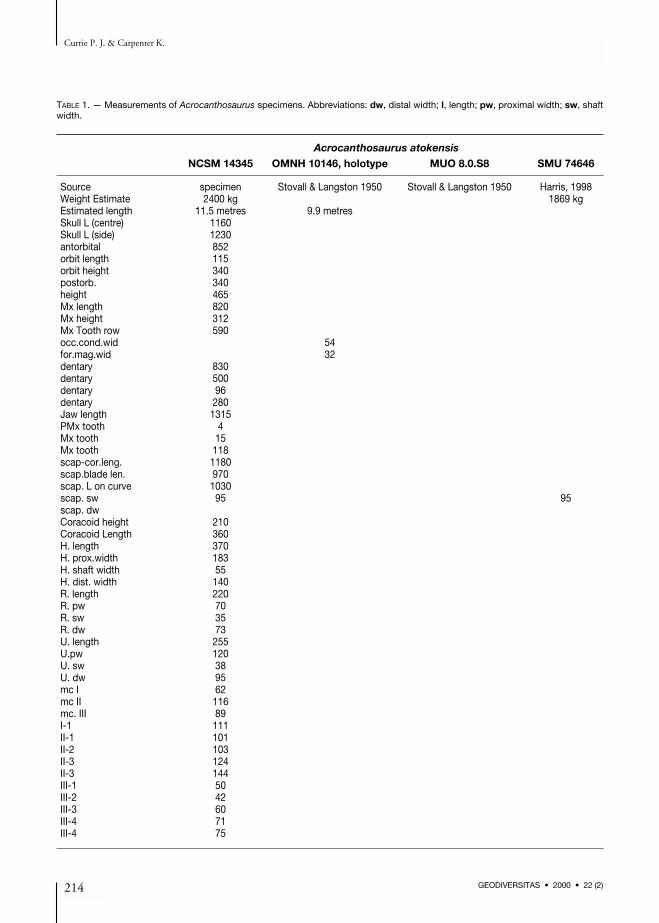






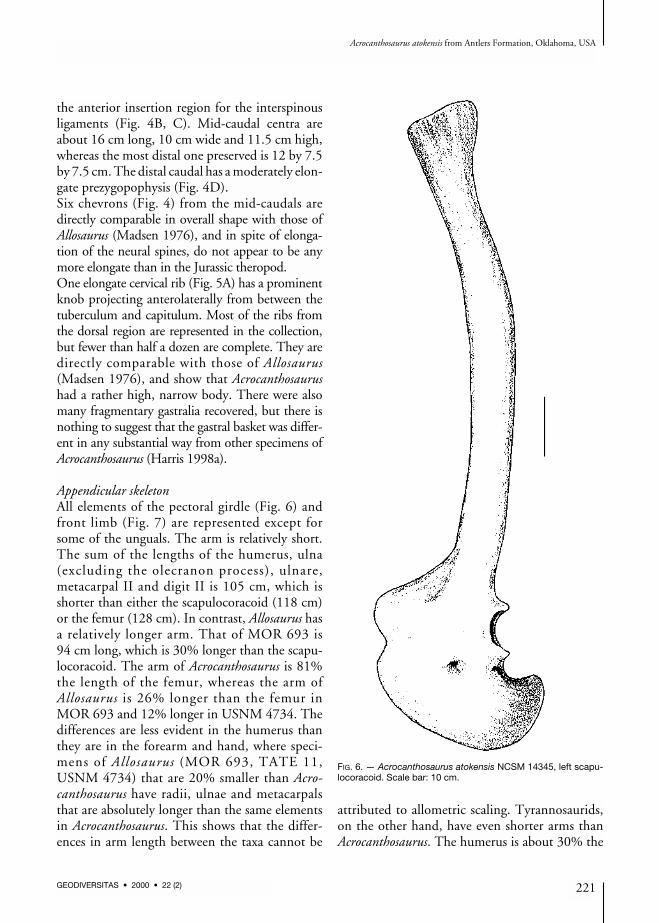
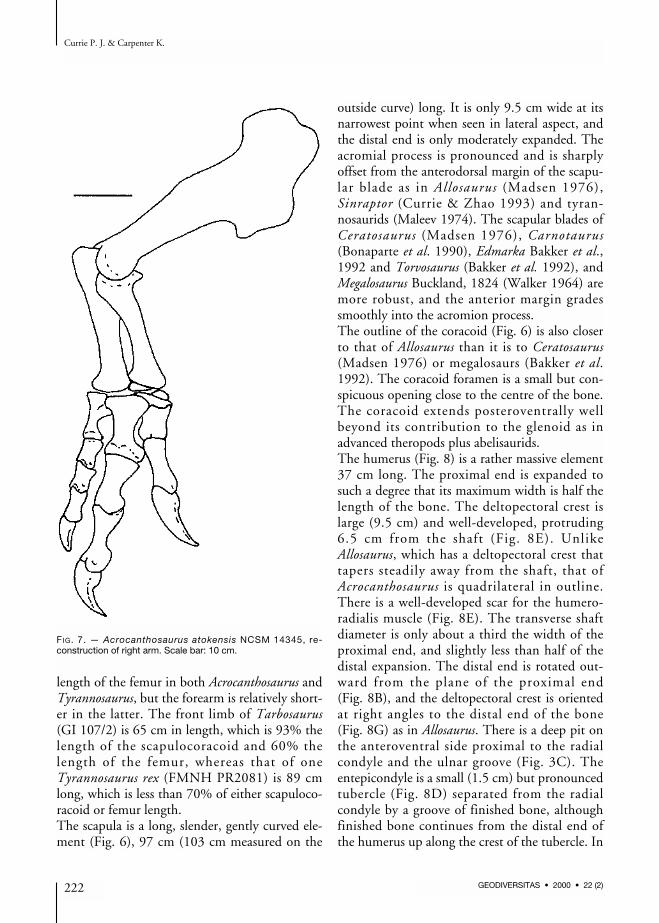





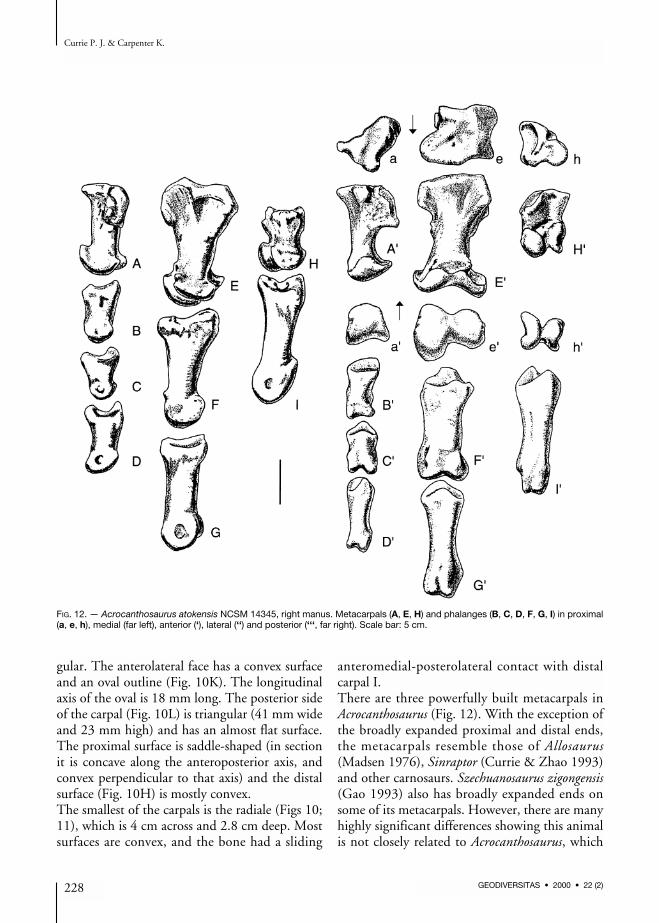
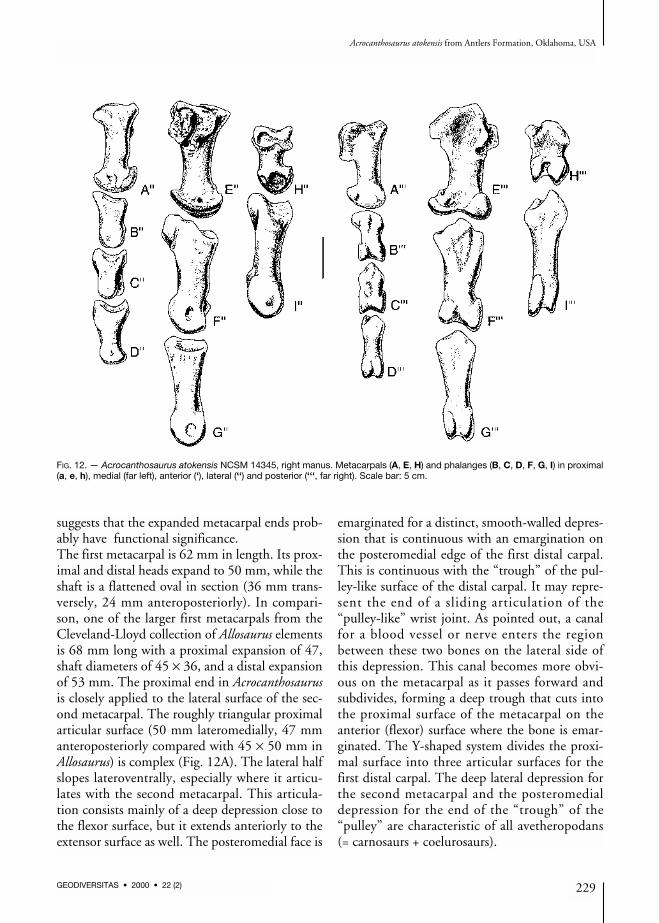






















![A densely feathered ornithomimid (Dinosauria: Theropoda) from … · 2016. 3. 20. · Ornithomimus] has oblique carbonaceous markings on the ulna and radius that are interpretedbyZelenitskyetal.](https://static.documents.pub/doc/80x56/6035283303337a5a604dd63e/a-densely-feathered-ornithomimid-dinosauria-theropoda-from-2016-3-20-ornithomimus.jpg)










