Page 1
1
A Novel Inhibitor of Dengue Virus Replication that Targets the Capsid Protein 1
2
Chelsea M. Byrd1,*, Dongcheng Dai1, Douglas W. Grosenbach1, Aklile Berhanu1, Kevin 3
F. Jones1, Kara B. Cardwell1, Christine Schneider1, Kristin A. Wineinger1, Jessica M. 4
Page1, Chris Harver1, Eric Stavale1, Shanthakumar Tyavanagimatt1, Melialani A. 5
Stone1, Ralf Bartenschlager2, Pietro Scaturro2, Dennis E. Hruby1, and Robert Jordan1+ 6
7
1SIGA Technologies, Inc., Corvallis, OR 97333 8
+Current address: Gilead Sciences, Inc., Foster City, CA 94404 9
2 Department of Infectious Diseases, Molecular Virology, University of Heidelberg, 10
Heidelberg, Germany 11
12
* Corresponding author: Mailing address: 4575 SW Research Way, Corvallis, OR 13
97333. Phone: 541-753-2000. Fax: 541-753-9999. E-mail: [email protected] 14
15
Running title: A dengue virus inhibitor 16
Keywords: 17
Flavivirus 18
Flavivirus Capsid protein 19
Capsid 20
Antiviral 21
Dengue 22
23
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Antimicrob. Agents Chemother. doi:10.1128/AAC.01429-12 AAC Accepts, published online ahead of print on 15 October 2012
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 2
2
ABSTRACT 24
Dengue viruses (DENV) infect 50-100 million people worldwide per year, of 25
which 500,000 develop severe life-threatening disease. This mosquito-borne illness is 26
endemic in most tropical and sub-tropical countries, and has spread significantly over 27
the last decade. While there are several promising vaccine candidates in clinical trials, 28
there are currently no approved vaccines or therapeutics available for treatment of 29
dengue infection. Here we describe a novel small-molecule compound, ST-148, that is a 30
potent inhibitor of all four serotypes of DENV in vitro. ST-148 significantly reduced 31
viremia and viral load in vital organs, and tended to lower cytokine levels in the plasma 32
in a non-lethal model of DENV infection in AG129 mice. Compound resistance mapped 33
to the DENV capsid (C) gene, and a direct interaction of ST-148 with C protein is 34
suggested by alterations of the intrinsic fluorescence of the protein in the presence of 35
compound. Thus, ST-148 appears to interact with the DENV C protein and inhibits 36
distinct step(s) of the viral replication cycle. 37
38
39
40
41
42
43
44
45
46
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 3
3
INTRODUCTION 47
Dengue fever (DF) is an acute, febrile illness caused by infection with dengue 48
virus (DENV). DENV circulates in populations as one of four distinct serotypes and is 49
transmitted to humans via the bite of the Aedes mosquito. While DF is a debilitating 50
illness, most cases resolve without sequelae (9). In a small percentage of cases, 51
individuals develop a severe capillary leakage syndrome referred to as dengue 52
hemorrhagic fever (DHF), which can lead to a more severe disease called dengue 53
shock syndrome (DSS) (9, 12). An estimated 50 – 100 million individuals are infected 54
with DENV each year mostly in tropical and subtropical areas of Southeast Asia, 55
resulting in nearly 500,000 severe life-threatening illnesses and 25,000 deaths. The 56
incidence of dengue disease is growing as the mosquito vector spreads due to 57
urbanization, population growth, increased international travel, a decrease in mosquito 58
control efforts, and global warming (11). 59
The existence of four distinct serotypes has made DENV vaccine development 60
challenging. While serotype-specific immunity reduces the rate of reinfection, immunity 61
does not provide complete protection from infection by the other three virus serotypes 62
(18). In fact, a second infection with a different virus serotype can potentially increase 63
the risk of severe disease. This enhanced risk is thought to be due to a combination of 64
viral genetics and heterotypic, non-neutralizing antibodies which enhance virus infection 65
(10). Disease severity has been linked to viral load, and patients with DHF or DSS have 66
viral titers in the blood 10-1000 fold higher than patients with DF (28). Thus, an antiviral 67
drug administered early during the course of infection that inhibits viral replication and 68
decreases viral load might be expected to reduce severity of disease. 69
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 4
4
DENV belongs to the Flaviviridae family and can be cultured in several 70
transformed cell lines to produce robust cytopathic effects. Upon entry of the virus into 71
the host cell, the positive, single stranded RNA genome is translated into a single 72
polyprotein that is proteolytically processed to produce three structural proteins, capsid 73
(C), pre-membrane (prM) and envelope (E), and seven non-structural proteins, NS1, 74
NS2A, NS2B, NS3, NS4A, NS4B and NS5. The non-structural proteins form the viral 75
replicase that is found within vesicles derived from virus-modified ER membranes (31). 76
Full length, positive stranded viral RNA genomes are synthesized from a negative 77
stranded intermediate (7). The newly synthesized RNA genomes are thought to exit 78
through pores that connect the vesicles to the cytosol (31). The viral core (C) protein 79
associates with the genomic RNA to form the nucleocapsid which buds into the ER 80
lumen to produce the immature virus particle containing viral prM and E glycoproteins 81
(36). The immature virus particles traffic via the secretory pathway and are processed in 82
the late Golgi compartment by a furin protease that cleaves the prM protein to produce 83
infectious virus particles that are released from the cell (34, 35). 84
A number of antiviral compounds have been identified that inhibit DENV 85
replication in vitro and in vivo (reviewed in (20)). Virus-specific inhibitors have been 86
identified that target the viral envelope (22), methyl transferase (16), protease (26), 87
NS4B protein (33), polymerase (6, 19), and virus-specific RNA translation (30). In 88
addition, compounds that target host enzymes such as ER glucosidases (4, 5, 8, 32), 89
dihydroorotate dehydrogenase (30), and an intracellular cholesterol transporter (21) 90
have been shown to have antiviral activity. Although these compounds appear to be 91
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 5
5
effective at inhibiting DENV replication, there is still no approved antiviral therapeutic for 92
the treatment of DENV infection in humans. 93
To identify potential antiviral therapeutics to treat DENV infection, a high 94
throughput screening assay was developed that measured virus-induced cytopathic 95
effects. This assay was used to screen a chemical compound library composed of over 96
200,000 unique small molecules, to identify inhibitors of DENV replication. A novel 97
compound series with activity against all four DENV serotypes was identified. The lead 98
compound in this series, ST-148, inhibited DENV replication in multiple cell types and 99
reduced viral load in a mouse model of DENV replication. Drug resistance was mapped 100
to the capsid coding region of the virus genome and recombinant DENV containing 101
mutations in this region showed reduced susceptibility to ST-148. The compound 102
altered the intrinsic fluorescence of purified wild-type C protein as well as a mutant C 103
protein containing amino acid changes associated with reduced compound 104
susceptibility. These data suggest that ST-148 inhibits the DENV replication cycle by 105
targeting the C protein. 106
107
108
109
110
111
112
113
114
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 6
6
METHODS AND MATERIALS 115
116
Compound synthesis 117
ST-148 was purchased from ChemBridge (San Diego, CA) and was dissolved in 118
dimethyl sulfoxide (DMSO) (Sigma Aldrich; St. Louis, MO) to a concentration of 10 mM. 119
120
Cells and viruses 121
Vero, BSC40, BHK, C6/36, L929, MDCK and MDBK cells were obtained from 122
American Type Culture Collection (ATCC; Manassas, VA), and Huh-7 cells were 123
obtained from Japan Health Sciences Foundation (Tokyo, Japan). Vero, BSC40, BHK, 124
and L929 cells were maintained at 37°C with 5% CO2 in Minimal Essential Medium 125
(MEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2 mM L-126
glutamine and 10 µg/ml gentamicin (all culture reagents from Invitrogen; Grand Island, 127
NY). C6/36 cells were maintained at 28°C with 5% CO2 in MEM as described above. 128
MDBK cells were maintained in MEM as described above and supplemented with 10% 129
horse serum (HS) (Invitrogen) instead of FBS. MDCK cells were maintained in Eagle’s 130
Modified Essential Medium (EMEM) (ATCC) with 10% FBS and 100U/ml Penicillin and 131
100 µg/ml Streptomycin (Invitrogen). Dengue-1 (DENV-1) strain TH-Sman (VR-344), 132
dengue-2 (DENV-2) strain New Guinea C (NGC) (VR-1255), dengue-3 (DENV-3) strain 133
H87 (VR-1256), and dengue-4 (DENV-4) strain H241 (VR-1257) were purchased from 134
ATCC. DENV-2 K0049 was obtained from Dr. Rebecca Rico-Hesse (Texas Biomedical 135
Research Institute; San Antonio, TX). DENV-2 S221 was obtained from Dr. Sujan 136
Shresta (La Jolla Institute for Allergy and Immunology; La Jolla, CA). The DENV were 137
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 7
7
propagated on Vero or C6/36 cell cultures. Bovine viral diarrhea virus (BVDV) strain 138
NADL (VR-534), Modoc virus strain M544 (VR-415), influenza A strain A/PR/8/34 (VR-139
1469), Sindbis virus strain AR-339 (VR-1585), and herpes simplex virus (HSV) strain 140
KOS (VR-1493) were purchased from ATCC. Recombinant vaccinia virus expressing 141
the green fluorescent protein (vvGFP) is described elsewhere (3). All cell culture 142
incubations, except for C6/36 cell culture, were performed at 37°C with 5% CO2. 143
144
High throughput screening assay 145
Vero cell monolayers were seeded on 96-well plates at 4.0 x 103 cells per well 146
and incubated overnight. The cells were then infected with DENV-2 at a multiplicity of 147
infection (moi) of 0.1 and incubated for 5 days with 5 µM compound added by a 148
Multiprobe II high-throughput system (PerkinElmer; Waltham, MA). Infected cell 149
monolayers were fixed with 5% glutaraldehyde (JT Baker; Central Valley, PA) in 150
phosphate-buffered saline (Fisher; Pittsburgh, PA), and stained with 0.1% crystal violet 151
(Sigma) in 5% methanol for 30 min. Virus-induced cytopathic effects (CPE) were 152
quantified by measuring absorbance at 570 nm on an EnVision Multilabel Reader 153
(PerkinElmer). 154
155
Inhibitory potency 156
Vero cell monolayers were seeded in 96-well plates at 4.0 x 103 cells per well 157
and incubated overnight. Dose-response curves were generated by measuring virus-158
induced CPE in the presence of a range of compound concentrations. Eight compound 159
concentrations (25, 8, 2.5, 0.8, 0.25, 0.08, 0.025, and 0.008 µM) were used to generate 160
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 8
8
inhibition curves suitable for calculating the EC50 from virus-induced CPEs. Compound 161
dilutions were prepared in DMSO prior to addition to the cell culture medium. The final 162
DMSO concentration in all samples was 0.5%. Cell monolayers were infected with 163
DENV-2 at a moi of 0.1. At 5 days post-infection (dpi), the assay was terminated by 164
fixing and staining the cells as described above. The level of CPE was quantified by 165
measuring absorbance at 570 nm. EC50 values were calculated by fitting the data to a 166
four-parameter logistic model (variable-slope, nonlinear regression model) to generate a 167
dose-response curve using XLfit 4.1 (IBDS; Emeryville, CA). From this curve, the 168
concentration of compound that inhibited virus-induced CPE by 50% was calculated. 169
170
Cytotoxicity assays 171
For measurement of compound cytotoxicity, Vero, BSC40, and Huh-7 cells were 172
seeded at either 3.5 x 103 or 1.0 x 104 cells per well in MEM supplemented with 5% FBS 173
in 96-well plates, incubated overnight, and then incubated with various concentrations of 174
ST-148 for 48 hr. Cell viability was measured using a metabolic activity assay where the 175
absorbance at 570/600 nm was measured on an EnVision Multilabel Reader at 5-6 hr 176
post-addition of 470 µM resazurin (Sigma) at 10% of the assay volume and incubation 177
at 37°C with 5% CO2. BHK cells were seeded at 5.0 x 103 cells per well, and the 178
remainder of the procedure was performed as above. MDBK cells were seeded at 5.0 x 179
103 cells per well in MEM supplemented with 2% HS, incubated overnight as above, 180
and then incubated with various concentrations of ST-148 for 96 hr. Cell viability was 181
measured as above. C6/36 cells were seeded at 4.0 x 104 cells per well, incubated 182
overnight at 28°C with 5% CO2, and then incubated with various concentrations of ST-183
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 9
9
148 for 48 hr at 28°C with 5% CO2. Cell viability was measured as above. For the five 184
day cytotoxicity assay, the assay was performed as above with the following 185
modifications: Vero cells were seeded at 3.5 x 103 cells per well in 96 well plates, and 186
incubated for 5 days after compound addition. Cell viability was measured using the 187
CellTiter-Glo® Luminescent Cell Viability Assay (Promega; Madison, WI) according to 188
manufacturer’s protocol. Luminescence was measured on an EnVision Multilabel 189
Reader at 700 nm with a 0.1 sec integration time. 190
191
AMES assay for genotoxicity 192
To evaluate the mutagenic potential of ST-148, the compound was tested in the 193
Bacterial Reverse Mutation Assay and the Muta-Chromo Plate Assay (EBPI; 194
Mississauga, Canada). All bacterial tester strains were purchased from Molecular 195
Toxicology, Inc. (Boone, NC). For the Bacterial Reverse Mutation Assay, 0.1 ml of 196
overnight cultures of E. coli tester strain WP-2 uvrA were added to molten top agar (with 197
0.5% NaCl, 0.05 mM histidine, and 0.05 mM biotin) in the presence and absence of 198
10% (v/v) Aroclor-1254-induced rat liver S9 (with 0.1 M phosphate buffer pH 7.4, 4 mM 199
NADP, 6 mM G-6-P, 33 mM MgCl2, and 8 mM KCl) (Molecular Toxicology) and various 200
concentrations of ST-148 with appropriate controls. The top agar mixture was overlaid 201
onto minimal glucose agar plates (Vogel-Bonner medium E with 2% glucose) (Molecular 202
Toxicology), allowed to harden, and incubated at 37°C for 48 to 72 hr. After the 203
incubation, any revertant colonies were scored. The Muta-Chromo Plate Assay was 204
carried out according to manufacturer’s instructions using Salmonella typhimurium 205
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 10
10
tester strains TA-98, TA-100, TA-1535, and TA-1537, Aroclor-1254-induced rat liver S9 206
with co-factors and various concentrations of ST-148. 207
208
Viral titer reduction assays 209
A viral titer reduction assay was performed to measure the antiviral efficacy of 210
ST-148 in cell culture. Cells (Vero for DENV or Modoc virus, L929 for MHV) were 211
seeded at 1 x 105 cells per well in 12-well plates and incubated overnight. Cells were 212
infected with DENV-1, DENV-2, DENV-3, DENV-4, DENV-2 S221, or Modoc virus at a 213
moi of 0.1 in the presence of various concentrations of ST-148 and incubated for 1.5 hr. 214
The inoculum was removed and replaced with MEM supplemented with 2% FBS 215
containing various concentrations of ST-148 and incubated for 48 hr. Supernatant was 216
harvested and serially diluted across Vero (L929 for MHV) cells seeded at 3.0 x 105 217
cells per well in 6-well plates. After a 1.5 hr infection, plates were overlaid with a 1:1 mix 218
of SeaPlaque agarose (Lonza; Allendale, NJ) and 2X MEM. Plates were incubated (3 219
days for MHC, 5 days for Modoc, 7 days for DENV-2 and DENV-4, 8 days for DENV-2 220
S221, and 10 days for DENV-1 and DENV-3). Plates were fixed and stained as 221
described above with the exception of a shorter stain period (10 seconds) to visualize 222
plaques. 223
224
Time-of-drug-addition assay 225
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 11
11
One day prior to infection, 3 x 104 Vero cells were seeded in 24-well plates and 226
incubated overnight. Cells were then infected with DENV-2 at a moi of 1.0. After 1.5 hr, 227
the virus inoculum was replaced with MEM supplemented with 1% FBS. ST-148 at a 228
concentration of 25 µM was added to the assay medium at either 2 hr prior to infection 229
or at 0 (i.e. at the time of infection), 2, 4, 6, 8, 12, 24, or 48 hpi. At 48 hpi the 230
supernatant was harvested and plaque titrated on Vero cells as described above. 231
232
Antiviral specificity 233
All assays (except for influenza) were carried out in the appropriate medium 234
containing 2% FBS (HS for MDBK cells). Ninety-six well cell culture plates were seeded 235
24 hr prior to use with 2.0 x 104 (Vero for HSV, BSC40 for Sindbis and vvGFP), 1.5 x 236
104 (Vero for DENV-2 K0049) or 1.0 x 104 (MDBK for BVDV, MDCK for influenza) cells 237
per well. Compound was added to duplicate wells of cells at final concentrations of 50, 238
25, 12.5, 6.3, 3.1, 1.6, 0.8, and 0 µM. The final concentration of DMSO in the assays 239
was 0.5%. Cells were infected at the cell culture infectious dose causing > 90% CPE at 240
1.6 (Sindbis), 2 (HSV or vvGFP), 3 (influenza), or 4 (BVDV) dpi or at titers that 241
generated an enzyme-linked immunosorbent assay (ELISA) signal of 2.5-fold over 242
background for DENV-2 K0049 at the end of the incubation period. For the influenza 243
assay, the cell culture medium (EMEM, supplemented with 10% FBS and Penicillin-244
Streptomycin) was replaced with serum free, Ultra MDCK Chemically Defined, Serum-245
free Renal cell medium with L-glutamine (Lonza) containing 5 µg/ml TPCK-treated 246
trypsin (Amersham; Pittsburgh, PA). Influenza virus was added to the culture medium 247
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 12
12
and virus-induced CPE was measured at 72 hr post-infection. As controls, uninfected 248
cells and cells receiving virus without compound were included on each assay plate. At 249
the end of the incubation period, virus-infected cells were fixed and stained as 250
described above. DENV-2 K0049 ELISA was carried out using a 1:100 dilution of 251
mouse monoclonal antibody to DENV prM glycoprotein (Abcam; Cambridge, MA) and 252
1:100 dilution of goat anti-mouse IgG (H+L)-HRP conjugate (BioRad; Hercules, CA) as 253
a secondary antibody. 254
255
Selection of drug resistant virus variants 256
DENV variants with reduced susceptibility to ST-148 were isolated by plating 257
wild-type DENV-2 on Vero cell monolayers in the presence of 5 µM ST-148. The 258
inoculum was removed after 2 hr incubation, replaced with media containing ST-148 and 259
incubated at 37°C with 5% CO2. At 6 dpi, the supernatant was harvested and applied to 260
Vero cells in the presence of ST-148, and the infection and incubation were repeated as 261
above. Virus isolates exhibiting reduced susceptibility to ST-148 were plaque purified 262
three times in the presence of compound prior to large-scale stock preparation. The 263
purified virus isolate (v148R) was amplified and viral RNA was extracted from v148R 264
using the QIAamp Viral RNA Mini Kit (Qiagen; Valencia, CA). Viral RNA was reverse-265
transcribed into cDNA using the SuperScript III One-Step RT-PCR System with 266
Platinum Taq (Invitrogen). The entire cDNA genome was sequenced at the Oregon 267
State University Center for Genome Research and Biocomputing Core Laboratories 268
(Corvallis, OR) and analyzed for mutations. 269
270
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 13
13
Plaque reduction assay 271
A plaque reduction assay was performed to compare the level of resistance of 272
v148R and wild-type DENV-2 against ST-148. Vero cells were seeded at 3.0 x 105 cells 273
per well in 6-well plates and incubated overnight. Growth media was aspirated from the 274
plates and cells were infected with DENV-2 or v148R at 100 pfu per well in MEM 275
supplemented with 1% FBS in the presence of 5 µM ST-148 or an equal volume of 276
DMSO. After 1.5 hr incubation, the inoculum was aspirated and the cells were overlaid 277
with a 1:1 mix of SeaPlaque agarose and 2X MEM in the presence of 5 µM ST-148 or 278
an equal volume of DMSO. The plates were incubated for 7 days, and fixed and stained 279
as described above with the exception of a shorter stain period (10 seconds) to 280
visualize plaques. 281
282
Multi-cycle growth curve 283
A multi-cycle growth curve was performed to compare the replication fitness of 284
v148R and wild-type DENV-2. Vero cells were seeded at 3.0 x 104 cells per well in 12-285
well plates and incubated overnight. Growth media was aspirated from the plates and 286
cells were infected with DENV-2 or v148R at a moi of 0.1. After 1.5 hr incubation, the 287
inoculum was removed, replaced with fresh media, and incubated. The supernatant was 288
harvested at various time points and virus levels quantified by plaque assay on Vero 289
cells. 290
291
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 14
14
Reverse genetics 292
DENV-2 cDNA-Tonga/74 and Escherichia coli BD1528 ung-1 nadB7 (BD1528) 293
cells were provided by Dr. Steve Whitehead at the National Institutes of Health 294
(Bethesda, MD). The S34L mutation identified in v148R was reverse engineered into 295
cDNA and transfected to obtain an infectious virus (vcDNA-148R) to confirm the 296
presence of the resistance-generating mutation. The S34L mutation was generated in 297
DENV-2 cDNA-Tonga/74 using QuikChange II XL Site-Directed Mutagenesis Kit 298
(Agilent Technologies; Santa Clara, CA) and primers DenMut-Cap-S34L-F (5’ TTGACA 299
AAGAGATTCTTACTTGGAATGCTGCAG) and DenMut-Cap-S34L-R (5’ CTGCAGCAT 300
TCCAAGTAAGAATCTCTTTGTCAA) following the manufacturer’s instructions, 301
transformed into competent BD1528 cells and plated on LB plates containing 10 µg/ml 302
tetracycline. Colonies were picked and cultured and cDNA was extracted using the 303
QIAquick MiniPrep Kit (Qiagen). The cDNA was sequenced to confirm the presence of 304
the mutation of interest. The cDNA-S34L was linearized and transcribed with the 305
AmpliCap SP6 High Yield Message Maker Kit (Epicentre; Madison, WI) and purified 306
with RNeasy Mini Kit (Qiagen). In 6-well plates 24 hr prior to infection, Vero cells were 307
seeded at 3.0 x105 cells per well in MEM with 10% FBS. Five hr prior to infection, the 308
medium was removed, washed three times with Dulbecco’s Phosphate Buffered Saline 309
without calcium and magnesium (DPBS) (Cellgro; Manassas, VA), and replaced with 310
Opti-Pro Serum Free Media (Invitrogen). Transcribed RNA (1 µg) diluted in 20 mM 311
HEPES/saline was added to diluted DOTAP Liposomal Transfection Reagent (Roche; 312
Indianapolis, IN) and HEPES/saline in a polystyrene tube, and incubated for 10 min at 313
room temperature. The DOTAP mixture was added to 1 ml of Opti-Pro media on Vero 314
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 15
15
cells and incubated for 48 hr, after which 2 ml of Opti-Pro was added. After six days, the 315
transfection was serially diluted on Vero cells as above, incubated for 7 days, and then 316
fixed and stained as above to determine the presence of infectious vcDNA-148R viral 317
particles. 318
319
C protein purification and expression 320
The DENV mature capsid domain (position 1 to 101) was amplified by PCR using 321
cDNA-Tonga/74 (2) as a template and primers DenCap (5’phosphorylated- 322
AATAACCAACGGAAAAAGGCGAG) and DenCapRevHindIII 323
(5’TCAGTCAAGCTTCATACAGTTCTACGTCTCCTGTTTAAG). The PCR product was 324
digested with StuI and HindIII and cloned into pQE-30Xa vector (Qiagen). The PCR-325
amplified region was verified by sequencing. Plasmids containing the correct insert were 326
transformed into the E. coli M15 [pREP4] expression system (Qiagen) according to the 327
manufacturer’s protocol. Cultures were incubated at 37°C with shaking until they 328
reached an OD of 0.75, were induced with the addition of IPTG to a final concentration 329
of 1 mM, and incubated at 37°C for 5 hr with shaking. Cultures were harvested by 330
centrifugation at 8,000 x g for 20 minutes, and cell pellets were frozen overnight. Pellets 331
were resuspended in a pH 7.8 binding buffer (20 mM imidazole, 2 mM TCEP, 0.5% 332
CHAPS, 1 M NaCl and 20 mM HEPES with a complete EDTA-free Protease Inhibitor 333
Cocktail Tablet (Roche)), microfluidized, centrifuged at 20,000 x g for 30 minutes, and 334
filtered. Supernatant was passed through a pre-equilibrated 5 ml HisTrap HP column 335
(GE Healthcare; Waukesha, WI) and eluted in the same buffer as above except 336
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 16
16
containing 500 mM imidazole. Fractions containing histidine-tagged dimerized C protein 337
were frozen at -80°C. 338
339
Intrinsic fluorescence 340
Fluorescence measurements were taken using a Fluorolog-3 fluorescence 341
spectrophotometer (HORIBA Jobin Yvon; Edison, NJ) equipped with a mercury-arc 342
lamp. A scan was performed on C protein samples diluted in DPBS using an excitation 343
wavelength of 295 nm and emission wavelengths from 310 to 380 nm in the presence 344
and absence of compound. C protein was diluted until the peak fluorescence count 345
measured was between 1 and 2 million counts which were not saturating for the 346
detection system. Background DPBS and drug fluorescence measurements were also 347
performed and the signals were subtracted from each sample so that the specific C 348
protein signal was measured in the presence and absence of compound. Samples were 349
prepared in 500 µl volumes for use in a quartz microcuvette. Each experiment was 350
performed in triplicate. Data analysis and collection was performed using DataMax 351
software, version 2.20 installed on a SpectraAcq processor (HORIBA Jobin Yvon). 352
353
In vivo studies 354
All studies involving vertebrate animals were approved by the Institutional Animal 355
Care and Use Committee of Oregon State University, a fully AAALAC-accredited facility, 356
and followed all federal guidelines as outlined in the Guide for the Care and Use of 357
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 17
17
Laboratory Animals (National Research Council, 2011). Strain 129 (129S2/SvPasCrl) 358
mice were obtained from Charles River Laboratories (Wilmington, MA) and were 359
approximately 6 weeks of age for the pharmacokinetic studies. AG129 mice (IFN-α/β 360
and -γ receptor deficient on the Strain 129 background) were purchased from B & K 361
Universal (Hull, UK) and were 5 to 6 weeks of age for antiviral efficacy studies. 362
363
Pharmacokinetic analysis 364
ST-148 was formulated for oral (p.o.), intraperitoneal (i.p.), and intravenous (i.v.) 365
dosing of mice (Strain 129), as an aqueous solution in 32% hydroxypropyl-beta-366
cyclodextrin (HPβCD) (Roquette; Lestrem, France). ST-148 concentrations were 367
measured in plasma harvested from mice at approximately 0.08, 0.25, 0.5, 1, 2, 3, 4, 6, 368
8, and 24 hr after i.v. administration and 1, 2, 3, 4, 6, 8, and 24 hr after p.o. and i.p. 369
administration. Compound concentrations were measured by liquid chromatography-370
tandem mass spectrometry (LC-MS/MS) using a 3200 Q TRAP LC-MS/MS system 371
(Applied Biosystems / MDS Sciex; Foster City, CA). LC was performed with a 372
Phenomenex Synergi JAX-RP column (4-µm particle size; 50 mm by 2 mm) at a flow 373
rate of 0.5 ml/min with a mobile phase containing 20% acetonitrile, 80% 10 mM 374
ammonium formate, pH 3.5 in water. WinNonlin software (Pharsight; Sunnyvale, CA) 375
was used to estimate pharmacokinetic values. 376
377
Antiviral efficacy in dengue murine model 378
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 18
18
DENV-2 strain NGC was used in the model and was propagated in C6/36 379
mosquito cells. Five to six-week old female AG129 mice were housed appropriately in 380
labeled cages (4 mice per cage), identified by ear-notching, and weighed prior to any 381
other procedures. Mice were injected i.p. with 0.5 ml of virus suspension in PBS (1.3 x 382
106 pfu per mouse). Mice were dosed immediately post-challenge with either 50 mg/kg 383
of ST-148 or the same volume of vehicle. Weight measurements and clinical 384
observations were made daily. On day 3 post-infection, mice were sacrificed by CO2 385
asphyxiation. Spleen and liver samples were taken from each animal and transferred to 386
pre-weighed homogenization tubes (FastPrep-24 Lysing Matrix C with additional ¼ inch 387
ceramic sphere added [MP Biomedical; Solon, OH]) with 0.5 ml of DPBS. Blood was 388
collected into eppendorf tubes containing sodium citrate for plasma isolation. Plasma 389
viremia and tissue viral load were determined by plaque-titration. Plasma cytokine (IL-6, 390
IL-10, IL-12p70, IFN-γ, TNF-α, and MCP-1) levels were assessed using the BD CBA 391
Mouse Inflammation Kit per the manufacturer’s instructions (BD Biosciences, San Jose, 392
CA). 393
394
Statistical analysis 395
For comparison of viral load in the plasma, spleen, and liver as well as cytokine 396
levels in the plasma among vehicle and once daily (QD) and twice daily (BID) ST-148-397
treated groups, one-way analysis of variance (ANOVA) with a Holm-Sidak correction 398
was used. For comparison of these parameters between vehicle and TID ST-148-399
treated groups, an unpaired Student t test was used and P-values <0.05 were 400
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 19
19
considered statistically significant. SigmaPlot for Windows Version 11.0 was used for 401
performing all statistical analyses (Systat Software, Inc., San Jose, CA). 402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
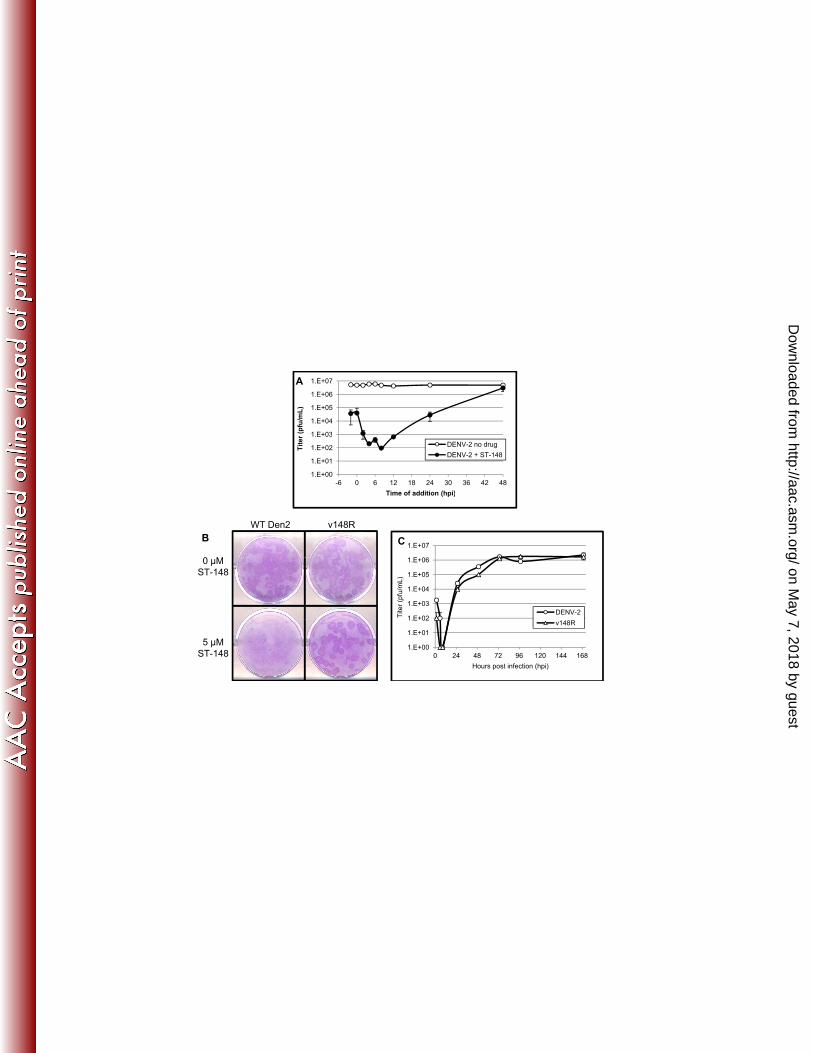
Page 20
20
RESULTS 424
425
ST-148 is a potent inhibitor of DENV replication in vitro 426
ST-148 (Figure 1) was identified through a high throughput screening (HTS) 427
assay of approximately 200,000 chemically diverse compounds. The HTS assay was 428
developed to identify inhibitors of DENV replication as measured by inhibition of virus-429
induced CPE in a 96-well plate format. ST-148 inhibited DENV-2 in a viral titer reduction 430
assay with an EC50 value (the concentration of compound necessary to inhibit 50% of 431
the virus titer) of 0.016 µM (Table 1) and an EC90 value of 0.125 µM. While ST-148 was 432
active against other dengue serotypes, the compound exhibited a broad range of 433
potencies with EC50 values of 0.512, 1.150, and 2.832 for DENV-3,-4, and -1 434
respectively. Analogs of ST-148, with improved potency, were more active against all 435
four serotypes and the range of EC50 values more narrow, suggesting that these 436
analogs were better optimized to interact with the viral target. ST-148 had potent activity 437
against all four serotypes of DENV in various cells types including Vero, C6/36, Huh-7, 438
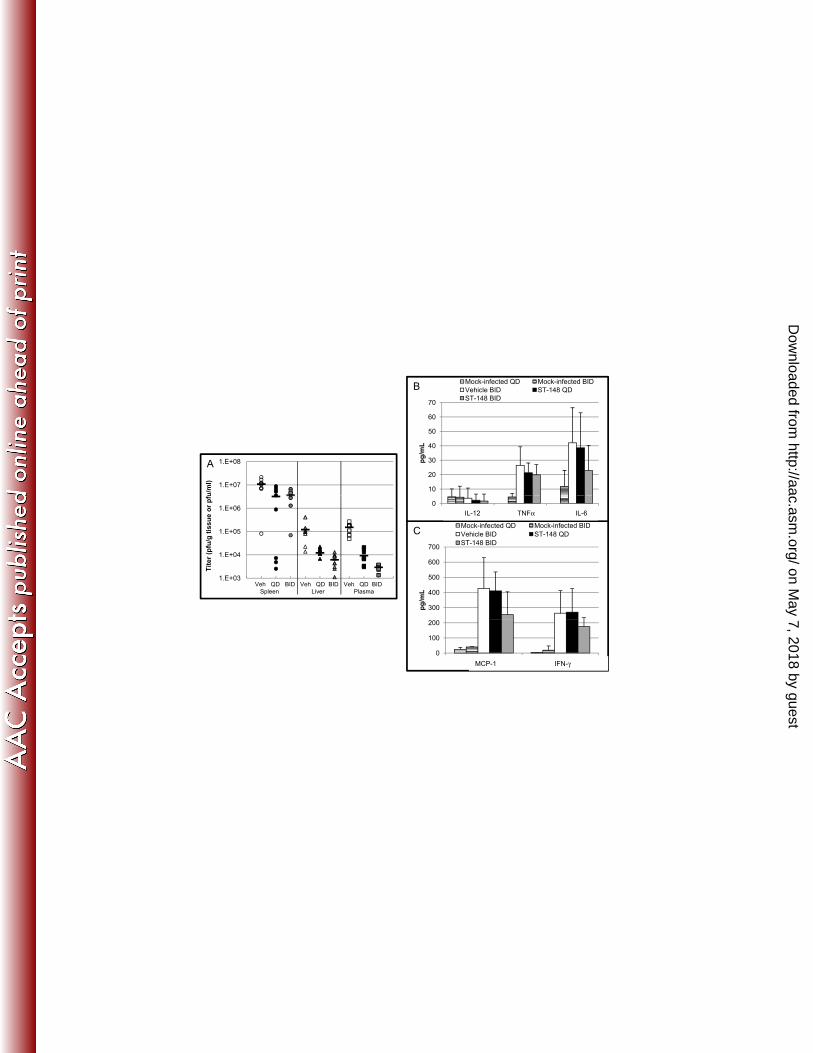
BHK, and L929 cells. ST-148 was not cytotoxic in any of the cell lines tested (Table 2). 439
440
ST-148 selectively inhibits flaviviruses 441
To determine the spectrum of antiviral activity, ST-148 potency was measured 442
against a variety of viruses from different virus families. As shown in Table 1 and Table 443
3, ST-148 was active against all DENV serotypes and Modoc virus, a murine flavivirus, 444
but showed less activity against other members of the Flaviviridae family, and did not 445
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 21
21
effectively inhibit viruses outside the Flaviviridae family. These results indicated that ST-446
148 selectively inhibits DENV within the Flavivirus genus. 447
448
ST-148 is not mutagenic 449
To evaluate the mutagenic potential of ST-148, the compound was tested in the 450
Bacterial Reverse Mutation Assay and the Muta-Chromo Plate Assay using Salmonella 451
typhimurium tester strains TA-98, TA-100, TA-1535, and TA-1537 as well as 452
Escherichia coli tester strain WP-2 uvrA in the presence and absence of Aroclor-453
induced rat liver S9. No mutagenic potential was observed with any of these conditions. 454
455
Time-of-drug-addition studies 456
To investigate which stage of the viral life cycle is affected by ST-148, a time-of-457
drug-addition experiment was conducted where 25 µM of ST-148 was added to DENV-2 458
infected Vero cells either just before the time of infection or at several time points after 459
infection. At 48 hr post-infection, viral yield was quantified by plaque assay. The data 460
indicate that ST-148 inhibits virus replication up to 12 hours after infection, consistent 461
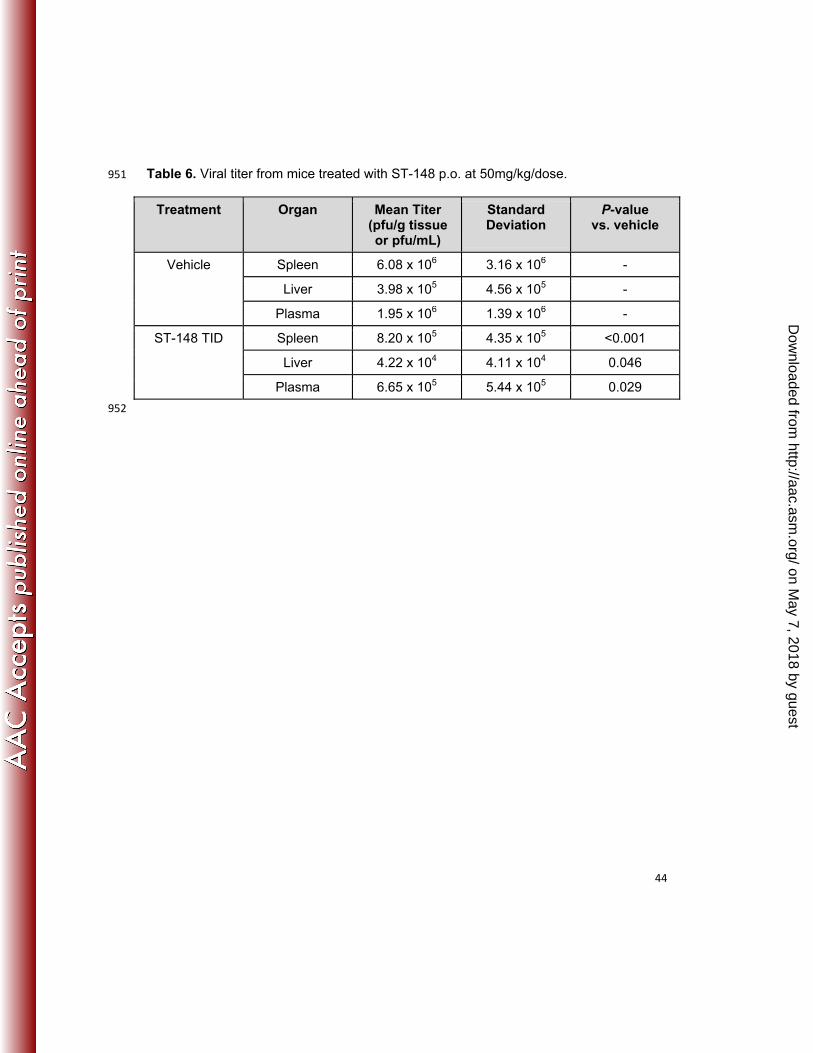
with a post-entry mechanism of action (Figure 2A). 462
463
Isolation of drug-resistant virus variants 464
To identify determinants of ST-148 sensitivity, virus variants resistant to the 465
inhibitory effects of the compound were generated by ten serial passages of virus in the 466
presence of ST-148. Virus capable of growing in the presence of 5 µM ST-148 467
(corresponding to a 312-fold higher EC50) was isolated and plaque-purified three times 468
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 22
22
in the presence of compound. A plaque reduction assay was conducted to measure the 469
ST-148 sensitivity of this variant (designated v148R) relative to wild-type virus. In the 470
presence of ST-148, plaque formation of the wild-type virus was greatly reduced, both in 471
size and number, while equal numbers of plaques, similar in size to the non-drug 472
treated samples, were observed for v148R in the presence of compound (Figure 2B). 473
The plaques are visualized by crystal violet staining with the stain in contact with the 474
monolayer for a very short duration (just a few seconds). The cytopathic effects 475
resulting from a dengue virus replication appear to enhance dye binding relative to the 476
uninfected monolayer resulting in plaques binding more dye and staining darker. This 477
rapid staining technique is used to visualize plaques formed by many types of virus that 478
fail to form obvious plaques on cell monolayers. In multi-cycle growth experiments the 479
replication rate and peak titer of v148R in the absence of compound was similar to wild-480
type virus suggesting that mutations affecting drug susceptibility had no measurable 481
effect on viral fitness (Figure 2C), at least in vitro. A viral titer reduction assay was used 482
to determine the level of susceptibility of the resistant virus to ST-148. The EC50 for 483
inhibition of the resistant variant (8.92 µM) was over 550-fold higher than the EC50 for 484
inhibition of wild-type DENV2 as determined in Vero cells (0.016 µM) (Table 1). 485
486
ST-148 resistance maps to the capsid coding region 487
Genomic RNA from plaque-purified resistant virus was isolated and amplified by 488
reverse transcription PCR to generate complementary DNA. The nucleotide sequence 489
of overlapping segments of the cDNA from the resistant variant was determined and 490
compared to wild-type DENV sequence to identify the changes in the viral genome that 491
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 23
23
correlated with reduced compound susceptibility. A single nucleotide change (nucleotide 492
101 in the capsid gene, cytosine to thymine) was found in the resistant virus genome 493
that resulted in a predicted serine (Ser) to leucine (Leu) change at amino acid position 494
34 in the capsid gene (Figure 3). Each of the DENV serotypes, but not Modoc virus, has 495
a serine at position 34. This change was reengineered into wild-type DENV-2 cDNA in 496
order to confirm that the single mutation found was responsible for conferring resistance 497
to ST-148. Infectious virus recovered from the engineered cDNA clone (vcDNA-148R) 498
showed reduced susceptibility to ST-148 with an EC50 for inhibition of 8.68 µM, similar 499
to the parental resistant isolate (Table 1). These results support the conclusion that the 500
S34L mutation is sufficient to confer resistance to ST-148. 501
502
ST-148 interacts with C protein 503
Compounds that bind proteins can alter the dielectric properties of aromatic 504
amino acid residues causing a change in the intrinsic fluorescence of the protein. To 505
determine if ST-148 interacts with C protein, its intrinsic fluorescence was measured 506
from 310 to 380 nm in the presence and absence of ST-148. In the presence of ST-148, 507
a dose-dependent decrease in the intrinsic fluorescence was observed consistent with a 508
direct interaction of the compound with C protein (Figure 4A). As controls, the intrinsic 509
protein fluorescence was measured in the presence of DMSO only, or in the presence 510
of ST-610, a DENV inhibitor compound discovered at SIGA that has a different chemical 511
structure and targets the helicase enzyme (manuscript in preparation) (Figure 1 and 512
4C). Neither DMSO nor ST-610 had a substantial effect on the intrinsic protein 513
fluorescence suggesting that the effects on fluorescence induced by ST-148 were 514
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 24
24
specific. Similar results were observed with a C protein variant isolated from ST-148-515
resistant virus (Figure 4B and D). These results suggest that ST-148 interacts with C 516
protein but amino acid changes that correlate with reduced compound susceptibility do 517
not alter this interaction. 518
519
Pharmacokinetic analyses and tolerability 520
The pharmacokinetic parameters of ST-148 in mice were determined after 521
formulating the compound as an aqueous solution in 32% hydroxypropyl-beta-522
cyclodextrin (HPβCD). Oral or intraperitoneal administration of ST-148 at 50 mg/kg/day 523
in Strain129 mice generated peak plasma concentrations of 3,138 or 52,283 ng/ml (7.44 524
or 124.04 µM) respectively (Table 4), which are 468- and 7,750-fold above the in vitro 525
EC50 value. Intravenous administration of ST-148 at 20 mg/kg/day generated a peak 526
plasma concentration of 41,133 ng/ml (97.58 µM), 6,093-fold above the EC50. The area 527
under the plasma concentration-time curve (AUC) was greatest in mice administered 528
ST-148 via the i.p. route. The absolute oral bioavailability of ST-148 was 9.1%. The 529
compound was rapidly absorbed after oral dosing with the time to maximum plasma 530
concentration measured at 1.0 hr. Intraperitoneal administration produced higher 531
plasma concentrations with rapid absorption, and peak plasma drug concentrations at 532
1.0 hr after dosing. There was no overt toxicity, or changes in physical signs including 533
body weights, observed in mice after repeated oral or i.p. ST-148 administration of 50 534
mg/kg/day for 3 days. These data indicate that ST-148 has limited oral bioavailability 535
and fairly rapid clearance, but good systemic availability following i.p. administration. 536
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 25
25
537
In vivo antiviral efficacy 538
A murine viremia model (14, 15, 25) using AG129 mice, which are deficient for 539
IFN-α/β and -γ receptors, was used to evaluate the in vivo efficacy of ST-148. Infection 540
of AG129 mice with DENV-2 (strain NGC) via intraperitoneal inoculation with 1.3 x 106 541
pfu produces a non-lethal infection with peak of viremia occurring on day 3 post-542
infection and an elevation in cytokine levels that are characteristic of DF. Immediately 543
after infection, ST-148 or vehicle at 50 mg/kg/dose was administered either once (QD) 544
or twice daily (BID) for three days. There was no additional weight changes in the ST-545
148 treated mice compared to vehicle-treated mice during the course of the experiment. 546
On average, BID treatment with ST-148 reduced peak plasma viremia 52-fold and 547
reduced viral load in the spleen and liver 3- and 20-fold respectively (Figure 5A). This 548
difference in viremia relative to vehicle-treated mice was statistically significant as 549
determined by one-way ANOVA (Table 5). An increase in the level of inflammatory 550
cytokines has been observed in the blood of patients who have DF (13) with a 551
contributing role in vascular leakage and disease severity. In the AG129 model, levels 552
of TNFα, IL-6, MCP-1, and IFN-γ are elevated, and a trend towards lower levels was 553
observed in ST-148 BID mice (Figure 5B and C), which was not statistically significant. 554
We further tested whether ST-148 was active in this model when administered 555
orally. Due to ST-148 p.o. pharmacokinetics that results in plasma levels of ST-148 556
above the in vitro EC90 only for approximately eight hours after administration, the 557
compound was administered three times per day (TID) at 8-hr intervals at 50 558
mg/kg/dose for three consecutive days. On average, treatment with ST-148 TID 559
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 26
26
reduced peak viremia 3-fold and reduced viral load in the spleen and liver 7- and 9-fold 560
respectively (Figure 6A). This difference in viremia relative to vehicle-treated mice was 561
statistically significant as determined by one-way ANOVA (Table 6). ST-148 562
administered orally showed a trend toward reduced levels of TNF-α, MCP-1, and IL-6, 563
but not IFN-γ, which were not statistically significant (Figure 6B and C). 564
Taken together, these results illustrate that ST-148 can decrease viremia, viral 565
load in tissues, and inflammatory cytokine levels when administered either i.p or orally. 566
567
DISCUSSION 568
A high throughput screen was conducted to identify inhibitors of DENV-induced 569
cytopathic effects. Characterization of compounds from this screen identified a chemical 570
series that inhibited replication of all four DENV serotypes. The lead compound in this 571
series, ST-148, exhibited strong DENV-2 antiviral activity with an EC50 of 16 nM and a 572
CC50 value of > 50 µM in several transformed cell types. ST-148 was inactive against 573
Japanese encephalitis virus and BVDV, but showed substantial activity against Modoc 574
virus (EC50 = 0.36 µM) and weak activity against YFV (EC50 = 6.97 µM) and HCV (EC50 575
= 39.2 µM). The weak activity against HCV was likely non-specific since this assay was 576
conducted using a subgenomic replicon system that lacks C protein. ST-148 was not 577
active against representative viruses from other virus families. The compound was not 578
genotoxic in Ames tests and was well tolerated in mice but exhibited modest oral 579
bioavailability (9.1%). Administration of ST-148 to mice, lacking interferon receptors, via 580
either oral gavage or intraperitoneal injection at 50 mg/kg reduced the viremia and viral 581
load in the liver and spleen by 1 log, suggesting that the compound is active in vivo. 582
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 27
27
DENV variants with reduced susceptibility to ST-148 were isolated and 583
compound susceptibility determinants mapped to the region of the genome encoding 584
the C protein. An amino acid substitution from serine to leucine at position 34 of the C 585
protein correlated with a loss of compound susceptibility. These results suggest that ST-586
148 interacts with the C protein to inhibit DENV replication. 587
A direct interaction of ST-148 with the C protein was measured by compound-588
induced effects on the intrinsic protein fluorescence. The reduction in intrinsic 589
fluorescence of C protein in the presence of ST-148 was not observed in the presence 590
of an unrelated compound, ST-610, that also inhibits DENV replication but targets the 591
helicase protein. While these data suggest a specific interaction of ST-148 with the C 592
protein, similar results were observed using a mutant C protein that contained the S34L 593
change associated with reduced compound susceptibility. Furthermore, ST-148 showed 594
substantial antiviral activity against Modoc virus, a natural flavivirus pathogen of mice, 595
and yellow fever virus. A comparison of the amino acid residues of DENV C protein with 596
selected flaviviruses including Modoc virus and yellow fever virus did not provide an 597
obvious explanation as to why these viruses would be susceptible to ST-148 while other 598
flaviviruses were not. It is possible that the interaction of ST-148 with the mutant C 599
protein may induce a conformational change that blocks the normal function of the C 600
protein, but not its intrinsic fluorescence properties. It’s also possible that ST-148 601
interacts with another region of the protein to affect intrinsic protein fluorescence and 602
the S34L amino acid change prevents inhibition by ST-148 without blocking this 603
interaction. Taken together, these data suggest that ST-148 interacts with the C protein 604
and blocks an essential activity required for virus replication. 605
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 28
28
Time-of-drug-addition experiments suggest that ST-148 is maximally effective 606
when added before or at the time of infection (Figure 2A), which might suggest inhibition 607
of uncoating which is consistent with an earlier report on the entry kinetics of DENV 608
(27). Given that the level of DENV released from infected cells is still somewhat reduced 609
when ST-148 is added up to 24 hrs after infection, it is also possible that ST-148 610
interaction with the C protein may have additional effect on later stages of viral 611
morphogenesis, such as nucleocapsid assembly. Alternatively, the assay may be 612
measuring virus spread, as an inhibition event at an early stage may be amplified 613
because of multiple rounds of virus infection and spread. But as it is likely the C protein 614
has multiple functions in uncoating, replication and assembly of progeny virions, more 615
work is needed to understand the overall effect of ST-148. 616
The mature form of the C protein is a basic, 12 kDa protein that forms 617
homodimers in solution consisting of four alpha helices (α1 to α4). They are arranged in 618
the dimer to produce a concave shaped molecule (17). The floor of the C protein dimer 619
is composed of the highly basic α4- α4’ helices which are thought to interact with RNA. 620
At the top of the concave structure, the α2−α2′ and α1−α1′ have been proposed to 621
interact with membranes (17). Based upon this structure, it has been proposed that 622
positively charged residues on α1-α1’ are involved in membrane interactions (17). The 623
amino acid change (S34L) associated with compound susceptibility resides at the end 624
of α1−α1’ (Figure 3B). Perhaps ST-148 interacts with this region of the protein and 625
prevents membrane association or other essential functions required for viral 626
replication. Although the C protein is the least conserved of the flavivirus proteins, the 627
structural properties are very similar and the charge distribution is well conserved. 628
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 29
29
The C protein binds RNA consistent with a proposed role in encapsidation of 629
newly replicated RNA into virions. A fraction of the cytoplasmic C protein in DENV-630
infected cells accumulates on the surface of ER-derived organelles named lipid droplets 631
(23). A similar phenotype was observed in cell lines expressing HCV C protein (1). 632
Substitutions of amino acid L50 or L54 in the DENV C protein disrupted lipid droplet 633
targeting and impaired viral particle formation. Likewise, mutations that disrupt lipid 634
droplet association of HCV C protein reduced particle formation (24). Moreover, one 635
study proposed that DENV infection increased the number of lipid droplets per cell (23). 636
Taken together, these results suggest that there is a link between lipid droplet 637
metabolism and viral replication (23). It is possible that ST-148 alters the interaction 638
between lipid droplets and the C protein, thereby inhibiting viral replication. While the 639
S34L change did not appear to alter interaction between C protein and ST-148 based 640
upon the results of the intrinsic protein fluorescence experiments, this amino acid 641
change could bypass the inhibitory effects of the compound by facilitating membrane 642
association. Future studies are ongoing to elucidate the mechanism of ST-148 643
inhibition. 644
645
646
647
648
649
650
651
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 30
30
ACKNOWLEDGEMENTS 652
This work was supported by NIH grants 1R43AI079937-01A1 and 1 653
R01A1093356-01. R. Bartenschlager is supported by grants from the European Union 654
(Silver (grant no. 260644) and Euvirna (grant no. 264286)). This work was performed by 655
employees of SIGA Technologies incorporated who hold stock in the company. We 656
would like to thank Dr. Steve Whitehead for the cDNA, Dr. Sujan Shresta for the S221 657
virus, Dr. Rebecca Rico-Hesse for the K0049 virus, and Dr. Johan Neyts for HCV 658
specificity testing. We would also like to thank Dr. Kady Honeychurch for DENV-specific 659
primer design, Andrew Wieczorek and Thuan Tran for HTS support, Candace Lovejoy 660
for project management, and Dr. Yali Chen for carrying out the AMES assay. Thanks to 661
Dr. Sean Amberg for critical review of the manuscript. 662
663
664
665
666
667
668
669
670
671
672
673
674
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 31
31
REFERENCES 675
676
1. Barba, G., F. Harper, T. Harada, M. Kohara, S. Goulinet, Y. Matsuura, G. Eder, Z. 677
Schaff, M. J. Chapman, T. Miyamura, and C. Brechot. 1997. Hepatitis C virus core 678
protein shows a cytoplasmic localization and associates to cellular lipid storage droplets. 679
Proc Natl Acad Sci U S A 94:1200-5. 680
2. Blaney, J. E., Jr., C. T. Hanson, K. A. Hanley, B. R. Murphy, and S. S. Whitehead. 681
2004. Vaccine candidates derived from a novel infectious cDNA clone of an American 682
genotype dengue virus type 2. BMC Infect Dis 4:39. 683
3. Byrd, C. M., T. C. Bolken, A. M. Mjalli, M. N. Arimilli, R. C. Andrews, R. Rothlein, T. 684
Andrea, M. Rao, K. L. Owens, and D. E. Hruby. 2004. New class of orthopoxvirus 685
antiviral drugs that block viral maturation. J Virol 78:12147-56. 686
4. Chang, J., W. Schul, T. D. Butters, A. Yip, B. Liu, A. Goh, S. B. Lakshminarayana, 687
D. Alonzi, G. Reinkensmeier, X. Pan, X. Qu, J. M. Weidner, L. Wang, W. Yu, N. 688
Borune, M. A. Kinch, J. E. Rayahin, R. Moriarty, X. Xu, P. Y. Shi, J. T. Guo, and T. 689
M. Block. 2011. Combination of alpha-glucosidase inhibitor and ribavirin for the 690
treatment of dengue virus infection in vitro and in vivo. Antiviral Res 89:26-34. 691
5. Chang, J., W. Schul, A. Yip, X. Xu, J. T. Guo, and T. M. Block. 2011. Competitive 692
inhibitor of cellular alpha-glucosidases protects mice from lethal dengue virus infection. 693
Antiviral Res 92:369-71. 694
6. Chen, Y. L., Z. Yin, J. Duraiswamy, W. Schul, C. C. Lim, B. Liu, H. Y. Xu, M. Qing, A. 695
Yip, G. Wang, W. L. Chan, H. P. Tan, M. Lo, S. Liung, R. R. Kondreddi, R. Rao, H. 696
Gu, H. He, T. H. Keller, and P. Y. Shi. 2010. Inhibition of dengue virus RNA synthesis 697
by an adenosine nucleoside. Antimicrob Agents Chemother 54:2932-9. 698
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 32
32
7. Cleaves, G. R., T. E. Ryan, and R. W. Schlesinger. 1981. Identification and 699
characterization of type 2 dengue virus replicative intermediate and replicative form 700
RNAs. Virology 111:73-83. 701
8. Courageot, M. P., M. P. Frenkiel, C. D. Dos Santos, V. Deubel, and P. Despres. 702
2000. Alpha-glucosidase inhibitors reduce dengue virus production by affecting the initial 703
steps of virion morphogenesis in the endoplasmic reticulum. J Virol 74:564-72. 704
9. Deen, J. L., E. Harris, B. Wills, A. Balmaseda, S. N. Hammond, C. Rocha, N. M. 705
Dung, N. T. Hung, T. T. Hien, and J. J. Farrar. 2006. The WHO dengue classification 706
and case definitions: time for a reassessment. Lancet 368:170-3. 707
10. Dejnirattisai, W., A. Jumnainsong, N. Onsirisakul, P. Fitton, S. Vasanawathana, W. 708
Limpitikul, C. Puttikhunt, C. Edwards, T. Duangchinda, S. Supasa, K. 709
Chawansuntati, P. Malasit, J. Mongkolsapaya, and G. Screaton. 2010. Cross-710
reacting antibodies enhance dengue virus infection in humans. Science 328:745-8. 711
11. Gubler, D. J. 2002. Epidemic dengue/dengue hemorrhagic fever as a public health, 712
social and economic problem in the 21st century. Trends Microbiol 10:100-3. 713
12. Halstead, S. B. 2007. Dengue. Lancet 370:1644-52. 714
13. Halstead, S. B. 1988. Pathogenesis of dengue: challenges to molecular biology. 715
Science 239:476-81. 716
14. Johnson, A. J., and J. T. Roehrig. 1999. New mouse model for dengue virus vaccine 717
testing. J Virol 73:783-6. 718
15. Kyle, J. L., P. R. Beatty, and E. Harris. 2007. Dengue virus infects macrophages and 719
dendritic cells in a mouse model of infection. J Infect Dis 195:1808-17. 720
16. Lim, S. P., L. S. Sonntag, C. Noble, S. H. Nilar, R. H. Ng, G. Zou, P. Monaghan, K. Y. 721
Chung, H. Dong, B. Liu, C. Bodenreider, G. Lee, M. Ding, W. L. Chan, G. Wang, Y. 722
L. Jian, A. T. Chao, J. Lescar, Z. Yin, T. R. Vedananda, T. H. Keller, and P. Y. Shi. 723
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 33
33
2011. Small molecule inhibitors that selectively block dengue virus methyltransferase. J 724
Biol Chem 286:6233-40. 725
17. Ma, L., C. T. Jones, T. D. Groesch, R. J. Kuhn, and C. B. Post. 2004. Solution 726
structure of dengue virus capsid protein reveals another fold. Proc Natl Acad Sci U S A 727
101:3414-9. 728
18. Monath, T. P. 1994. Dengue: the risk to developed and developing countries. Proc Natl 729
Acad Sci U S A 91:2395-400. 730
19. Niyomrattanakit, P., Y. L. Chen, H. Dong, Z. Yin, M. Qing, J. F. Glickman, K. Lin, D. 731
Mueller, H. Voshol, J. Y. Lim, S. Nilar, T. H. Keller, and P. Y. Shi. 2010. Inhibition of 732
dengue virus polymerase by blocking of the RNA tunnel. J Virol 84:5678-86. 733
20. Noble, C. G., Y. L. Chen, H. Dong, F. Gu, S. P. Lim, W. Schul, Q. Y. Wang, and P. Y. 734
Shi. 2010. Strategies for development of Dengue virus inhibitors. Antiviral Res 85:450-735
62. 736
21. Poh, M. K., G. Shui, X. Xie, P. Y. Shi, M. R. Wenk, and F. Gu. 2012. U18666A, an 737
intra-cellular cholesterol transport inhibitor, inhibits dengue virus entry and replication. 738
Antiviral Res 93:191-8. 739
22. Poh, M. K., A. Yip, S. Zhang, J. P. Priestle, N. L. Ma, J. M. Smit, J. Wilschut, P. Y. 740
Shi, M. R. Wenk, and W. Schul. 2009. A small molecule fusion inhibitor of dengue 741
virus. Antiviral Res 84:260-6. 742
23. Samsa, M. M., J. A. Mondotte, N. G. Iglesias, I. Assuncao-Miranda, G. Barbosa-743
Lima, A. T. Da Poian, P. T. Bozza, and A. V. Gamarnik. 2009. Dengue virus capsid 744
protein usurps lipid droplets for viral particle formation. PLoS Pathog 5:e1000632. 745
24. Shavinskaya, A., S. Boulant, F. Penin, J. McLauchlan, and R. Bartenschlager. 2007. 746
The lipid droplet binding domain of hepatitis C virus core protein is a major determinant 747
for efficient virus assembly. J Biol Chem 282:37158-69. 748
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 34
34
25. Shresta, S., J. L. Kyle, H. M. Snider, M. Basavapatna, P. R. Beatty, and E. Harris. 749
2004. Interferon-dependent immunity is essential for resistance to primary dengue virus 750
infection in mice, whereas T- and B-cell-dependent immunity are less critical. J Virol 751
78:2701-10. 752
26. Steuer, C., C. Gege, W. Fischl, K. H. Heinonen, R. Bartenschlager, and C. D. Klein. 753
2011. Synthesis and biological evaluation of alpha-ketoamides as inhibitors of the 754
Dengue virus protease with antiviral activity in cell-culture. Bioorg Med Chem 19:4067-755
74. 756
27. van der Schaar, H. M., M. J. Rust, C. Chen, H. van der Ende-Metselaar, J. Wilschut, 757
X. Zhuang, and J. M. Smit. 2008. Dissecting the cell entry pathway of dengue virus by 758
single-particle tracking in living cells. PLoS Pathog 4:e1000244. 759
28. Vaughn, D. W., S. Green, S. Kalayanarooj, B. L. Innis, S. Nimmannitya, S. 760
Suntayakorn, T. P. Endy, B. Raengsakulrach, A. L. Rothman, F. A. Ennis, and A. 761
Nisalak. 2000. Dengue viremia titer, antibody response pattern, and virus serotype 762
correlate with disease severity. J Infect Dis 181:2-9. 763
29. Vrolijk, J. M., A. Kaul, B. E. Hansen, V. Lohmann, B. L. Haagmans, S. W. Schalm, 764
and R. Bartenschlager. 2003. A replicon-based bioassay for the measurement of 765
interferons in patients with chronic hepatitis C. J Virol Methods 110:201-9. 766
30. Wang, Q. Y., R. R. Kondreddi, X. Xie, R. Rao, S. Nilar, H. Y. Xu, M. Qing, D. Chang, 767
H. Dong, F. Yokokawa, S. B. Lakshminarayana, A. Goh, W. Schul, L. Kramer, T. H. 768
Keller, and P. Y. Shi. 2011. A translation inhibitor that suppresses dengue virus in vitro 769
and in vivo. Antimicrob Agents Chemother 55:4072-80. 770
31. Welsch, S., S. Miller, I. Romero-Brey, A. Merz, C. K. Bleck, P. Walther, S. D. Fuller, 771
C. Antony, J. Krijnse-Locker, and R. Bartenschlager. 2009. Composition and three-772
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 35
35
dimensional architecture of the dengue virus replication and assembly sites. Cell Host 773
Microbe 5:365-75. 774
32. Whitby, K., T. C. Pierson, B. Geiss, K. Lane, M. Engle, Y. Zhou, R. W. Doms, and M. 775
S. Diamond. 2005. Castanospermine, a potent inhibitor of dengue virus infection in vitro 776
and in vivo. J Virol 79:8698-706. 777
33. Xie, X., Q. Y. Wang, H. Y. Xu, M. Qing, L. Kramer, Z. Yuan, and P. Y. Shi. 2011. 778
Inhibition of dengue virus by targeting viral NS4B protein. J Virol 85:11183-95. 779
34. Yu, I. M., W. Zhang, H. A. Holdaway, L. Li, V. A. Kostyuchenko, P. R. Chipman, R. J. 780
Kuhn, M. G. Rossmann, and J. Chen. 2008. Structure of the immature dengue virus at 781
low pH primes proteolytic maturation. Science 319:1834-7. 782
35. Zhang, Y., J. Corver, P. R. Chipman, W. Zhang, S. V. Pletnev, D. Sedlak, T. S. 783
Baker, J. H. Strauss, R. J. Kuhn, and M. G. Rossmann. 2003. Structures of immature 784
flavivirus particles. Embo J 22:2604-13. 785
36. Zhang, Y., B. Kaufmann, P. R. Chipman, R. J. Kuhn, and M. G. Rossmann. 2007. 786
Structure of immature West Nile virus. J Virol 81:6141-5. 787
788
789
790
791
792
793
794
795
796
797
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 36
36
LEGENDS TO FIGURES 798
Figure 1. Structures of ST-148 and ST-610. 799
800
Figure 2. (A) Time-of-drug-addition study. DENV-2 infected Vero cells were treated with 801
25 µM ST-148 at various time points and virus levels quantified by plaque assay at 48 802
hpi. The data are mean values ± SD for three independent experiments. (B) Plaque 803
formation of wild-type DENV and ST-148-resistant virus in the presence and absence of 804
ST-148. Vero cell monolayers were infected with DENV-2 or v148R in the presence and 805
absence of 5 µM ST-148 and covered with an agarose overlay. At 7 days post-infection 806
the cultures were fixed in 5% glutaraldehyde and stained with crystal violet. (C) Multi-807
cycle growth curve of wild-type DENV and ST-148-resistant virus in the absence of ST-808
148. Vero cell monolayers were infected with virus, harvested at various time points and 809
plaque titrated. The data are mean values ± SD for two observations. 810
811
Figure 3. (A) A sequence comparison of capsid orthologs. Graphic representation of 812
the DENV genome, illustrating the location of resistance to ST-148 (capsid S34L) and 813
an amino acid sequence comparison of orthologs from the capsid region of other 814
flaviviruses showing the degree of sequence conservation. (B) Structure of the 815
homodimer DENV-2 C protein. Subunit A – light blue, subunit B – dark blue. The 816
location of the serine residue at position 34 is indicated in red for both chains. Protein 817
structure modified using CN3D software (17). 818
819
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 37
37
Figure 4. The effect of ST-148 on the intrinsic fluorescence of C protein. Intrinsic 820
fluorescence measurements in arbitrary units (a.u.) were made by combining protein 821
and compound, and reading absorbance at an excitation of 295 nm and scanning 822
emissions between 310 and 380 nm. (A) C protein in the presence of ST-148, (B) ST-823
148-resistant mutant C protein (S34L) with ST-148, (C) C protein with ST-610, and (D) 824
mutant C protein with ST-610. 825
826
Figure 5. In vivo efficacy of ST-148 delivered i.p. AG129 mice (8 mice/ treatment 827
group) were infected with 1.3 x 106 pfu DENV-2 (strain NGC) i.p. on day 0. The mice 828
were treated with 50 mg/kg ST-148 i.p. either once (QD) or twice per day (BID). On day 829
3 post-infection (the peak of viremia), the viral load and viremia were quantified by 830
plaque assay. (A) Viral load and viremia. White symbols represent vehicle treated mice, 831
black symbols represent mice treated with ST-148 QD, and gray symbols represent 832
mice treated with ST-148 BID. Circles represent spleen titer, triangles represent liver 833
titer, and squares represent plasma titer. Solid bars represent the arithmetic mean. (B 834
and C) Inflammatory cytokine levels in the plasma. The lower limit of quantification for 835
the CBA assay is 20 pg/ml. The data are mean values ± SD for three observations. 836
837
Figure 6. Oral In vivo efficacy of ST-148. AG129 mice (8 mice/treatment group) were 838
infected with 9 x 106 pfu DENV-2 (strain NGC) i.p. on day 0. The mice were treated with 839
50 mg/kg ST-148 p.o. three times per day (TID). On day 3 post-infection (the peak of 840
viremia), the viral load and viremia were quantified by plaque assay. (A) Viral load and 841
viremia, open symbols represents vehicle-treated mice, filled symbols represent ST-842
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 38
38
148-treated mice. Circles represent spleen titer, triangles represent liver titer, and 843
squares represent plasma titer. Solid bars represent the arithmetic mean. (B and C) 844
Inflammatory cytokine levels in the plasma. The lower limit of quantification for the CBA 845
assay is 20 pg/ml. The data are mean values ± SD for three observations. 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
865
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 39
39
Table 1. Antiviral activity of ST-148 against DENV 866
Cell Type Virus EC50(µM)a Vero DENV-1 2.832 ± 1.13 DENV-2 0.016 ± 0.01 DENV-3 0.512 ± 0.42 DENV-4 1.150 ± 0.14 DENV-2 K0049 0.551 ± 0.40 DENV-2 S221 0.023 ± 0.01 v148R 8.918 ± 0.80 vcDNA-148R 8.684 ± 0.75 C6/36 DENV-2 0.039 ± 0.01 Huh-7 DENV-2 0.012 ± 0.01 BHK DENV-2 0.073 ± 0.08 L929 DENV-2 0.016 ± 0.01
aCells were infected with indicated viruses at a moi of 0.1 and treated with ST-148. 867
Culture medium was collected at 48 h post-infection and the viral titer determined by 868
plaque assay. For K0049, activity was measured by ELISA assay. v148R is the tissue 869
culture passaged, drug-resistant DENV-2 isolate, and vcDNA-148R is the virus 870
recovered from the engineered cDNA clone. The data are mean values ± SD for three 871
independent experiments. 872
873
874
875
876
877
878
879
880
881
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 40
40
Table 2. Cell toxicity of ST-148 882
Cell Type CC50 (µM)a Vero (2d) >100 Vero (5d) >100 BSC40 >100 C6/36 >100 Huh-7 >100 BHK >100 MDBK >100 L929 >50
aCells were incubated with ST-148 at a range of concentrations for 48 hr (also 5 days 883
for Vero; 96 hr for MDBK cells). Cell viability was measured using a resazurin assay 884
where the absorbance was measured on an EnVision Multilabel Reader at 5 hr post-885
addition of resazurin and incubation at 37°C. For the 5 day assay, viability was 886
measured using Cell Titer-Glo Luminescent Cell Viability Assay. The data are mean 887
values for three independent experiments. 888
889
890
891
892
893
894
895
896
897
898
899
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 41
41
Table 3. ST-148 antiviral spectrum of activity and selectivity 900
Virus Family Genus Group EC50
(µM) Modoc virus Flaviviridae Flavivirus +ssRNA 0.36 Yellow fever virusa Flaviviridae Flavivirus +ssRNA 6.97 Japanese encephalitis virusa Flaviviridae Flavivirus +ssRNA >50 Bovine viral diarrhea virus Flaviviridae Pestivirus +ssRNA >50 Hepatitis C virusb Flaviviridae Hepacivirus +ssRNA 39.2 Sindbis virus Togaviridae Alphavirus +ssRNA >50 Influenza A virus Orthomyxoviridae InfluenzavirusA -ssRNA >25 Vaccinia virus Poxviridae Orthopoxvirus dsDNA >50 Herpes simplex virus 1 Herpesviridae Simplexvirus dsDNA >50 a: In vitro testing done by Integrated Biotherapeutics via a CPE based assay in Vero 901
cells. b:In vitro testing done by Dr. Neyts laboratory at the Rega Institute using a 902
subgenomic HCV replicon (genotype 1b) in Huh 5.2 cells(29). 903
904
905
906
907
908
909
910
911
912
913
914
915
916
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 42
42
Table 4. Pharmacokinetic parameters of ST-148 in Strain 129 mice 917
Route Dosage (mg/kg)
Tmax
(hr) Cmax (ng/ml)
AUC (hr*ng/ml)
p.o. 50 1.0 3138 5826 i.p. 50 1.0 52283 77011 i.v. 20 0.1 41133 25523 Plasma drug concentrations were measured by LC-MS/MS over a 24-hr time period. 918
Tmax, time of maximal concentration; Cmax, maximal concentration. 919
920
921
922
923
924
925
926
927
928
929
930
931
932
933
934
935
936
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 43
43
Table 5. Viral titer from mice treated with ST-148 i.p. at 50mg/kg/dose. 937
Treatment
Organ Mean Titer (pfu/g tissue or pfu/mL)
Standard Deviation
P-value vs. vehicle
Vehicle Spleen 1.08 x 107 6.48 x 106 -
Liver 1.21 x 105 1.23 x 105 -
Plasma 1.52 x 105 7.19 x 104 -
ST-148 QD Spleen 3.16 x 106 3.47 x 106 0.002
Liver 1.21 x 104 6.83 x 103 0.004
Plasma 9.20 x 103 6.81 x 103 <0.001
ST-148 BID Spleen 3.59 x 106 2.14 x 106 0.004
Liver 6.12 x 103 4.06 x 103 0.006
Plasma 2.93 x 103 8.33 x 102 <0.001
938
939
940
941
942
943
944
945
946
947
948
949
950
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 44
44
Table 6. Viral titer from mice treated with ST-148 p.o. at 50mg/kg/dose. 951
Treatment Organ Mean Titer (pfu/g tissue or pfu/mL)
Standard Deviation
P-value vs. vehicle
Vehicle Spleen 6.08 x 106 3.16 x 106 -
Liver 3.98 x 105 4.56 x 105 -
Plasma 1.95 x 106 1.39 x 106 -
ST-148 TID Spleen 8.20 x 105 4.35 x 105 <0.001
Liver 4.22 x 104 4.11 x 104 0.046
Plasma 6.65 x 105 5.44 x 105 0.029
952
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 45
SN
NN
N
ON
N
O
NN S
O
O N
ST-610
SS
NN
ST-148
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 46
1 E+03
1.E+04
1.E+05
1.E+06
1.E+07
(pfu
/mL)
A
1.E+00
1.E+01
1.E+02
1.E+03
-6 0 6 12 18 24 30 36 42 48
Tite
r (
Time of addition (hpi)
DENV-2 no drugDENV-2 + ST-148
Time of addition (hpi)
1.E+07
WT Den2 v148R
0 M
B C
1 E+02
1.E+03
1.E+04
1.E+05
1.E+06
Tite
r (pf
u/m
L)
DENV-2
0 µMST-148
1.E+00
1.E+01
1.E+02
0 24 48 72 96 120 144 168Hours post infection (hpi)
v148R
5 µMST-148
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 47
5’ UTR 3’ UTRC prM E NS1 NS2A NS2B NS3 NS4A NS4B NS5
Capsid aa1 114
A B
NRVSTVQQLT KRFLLGMLQG RGP-LKLFMANRVSTVQQLT KRFSLGMLQG RGP-LKLFMANRVSTVQQLT KRFSLGMLQG RGP-LKLFMA
Capsid aa1-114
S34L
v148R DEN2 NGC DEN2 cDNA
1 NRVSTVSQLA KRFSKGLLSG QGP-MKLVMANRVSTGSQLA KRFSRGLLNG QGP-MKLVMANRVSTPQGLV KRFSTGLFSG KGP-LRMVLA
KKKKNAGRNG KEVPGLALVM GV------IHRLERGKMKIV PKESEKDSKT KPPDATIVVEQDVKFPGGGQ IVGGVYLLPR RGPRLGVRAT
DEN1 TH-Sman DEN3 H87 DEN4 H241
MODOC M544 BVDV NADL HCV 1b/DK1 Q Q
PRVLSLIGLK RAMLS-LIDG KGP-IRFVLA--VRSLSNKI KQKTKQIGNR PGP-SRGVQG
/
WNV KN3829 YFV Uga48
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 48
1400000
1600000
1800000
(a.u
.)
WTCapsidST-148, 500nMST-148, 2500nM
1400000
1600000
1800000
a.u.
)
WTCapsidDMSOST-610, 2500nM
A C
800000
1000000
1200000
Fluo
resc
necn
e
800000
1000000
1200000Fl
uore
scne
cne
(a
400000
600000
300 320 340 360 380Wavelength (nm)
400000
600000
300 320 340 360 380Wavelength (nm)
1400000 1400000B D
1000000
1200000
1400000
nce
(a.u
.)
MutantCapsidST-148, 500nMST-148, 2500nM
1000000
1200000
1400000
ce (a
.u.)
Mutant CapsidDMSOST-610, 2500nM
B D
600000
800000
Fluo
resc
en
600000
800000
Fluo
resc
en
400000300 320 340 360 380
Wavelength (nm)
400000300 320 340 360 380
Wavelength (nm)
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 49
50
60
70
Mock-infected QD Mock-infected BIDVehicle BID ST-148 QDST-148 BID
B
10
20
30
40
50pg
/mL
1.E+07
1.E+08
u/m
l)
A
700
Mock-infected QD Mock-infected BIDVehicle BID ST-148 QDST-148 BID
0IL-12 TNFa IL-6
1.E+04
1.E+05
1.E+06
r (pf
u/g
tissu
e or
pfu
C
IL-12 TNFα IL-6
300
400
500
600
pg/m
L
1.E+03
Tite
r
Veh QD BID Veh QD BID Veh QD BIDSpleen Liver Plasma
0
100
200
MCP-1 IFN-gMCP-1 IFN-γ
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 50
140160180
Mock-infected Vehicle ST-148B
406080
100120
pg/m
L
1.E+07
1.E+08
pfu/
mL)
A
020
IL-12 TNFa IL-6
800Mock-infected Vehicle ST-1481.E+05
1.E+06
Tite
r (pf
u/g
or p
C
IL-12 TNFα IL-6
300
400
500
600
700
800
pg/m
L
1.E+04Veh ST-148 Veh ST-148 Veh ST-148
Spleen Liver Plasma
0
100
200
300
MCP-1 INF-y
p
MCP-1 IFN-γ
on May 7, 2018 by guest
http://aac.asm.org/
Dow
nloaded from