HAL Id: hal-00895694 https://hal.archives-ouvertes.fr/hal-00895694 Submitted on 1 Jan 2009 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés. A simple and rapid method for the disruption of Staphylococcus aureus, optimized for quantitative reverse transcriptase applications: Application for the examination of Camembert cheese Wilfried Ablain, Sylvie Hallier Soulier, David Causeur, Michel Gautier, Florence Baron To cite this version: Wilfried Ablain, Sylvie Hallier Soulier, David Causeur, Michel Gautier, Florence Baron. A simple and rapid method for the disruption of Staphylococcus aureus, optimized for quantitative reverse transcriptase applications: Application for the examination of Camembert cheese. Dairy Science & Technology, EDP sciences/Springer, 2009, 89 (1), pp.69-81. hal-00895694
Transcript
HAL Id: hal-00895694https://hal.archives-ouvertes.fr/hal-00895694
Submitted on 1 Jan 2009
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
A simple and rapid method for the disruption ofStaphylococcus aureus, optimized for quantitative
reverse transcriptase applications: Application for theexamination of Camembert cheese
Wilfried Ablain, Sylvie Hallier Soulier, David Causeur, Michel Gautier,Florence Baron
To cite this version:Wilfried Ablain, Sylvie Hallier Soulier, David Causeur, Michel Gautier, Florence Baron. A simpleand rapid method for the disruption of Staphylococcus aureus, optimized for quantitative reversetranscriptase applications: Application for the examination of Camembert cheese. Dairy Science &Technology, EDP sciences/Springer, 2009, 89 (1), pp.69-81. �hal-00895694�
A simple and rapid method for the disruptionof Staphylococcus aureus, optimized for
quantitative reverse transcriptase applications:Application for the examination
of Camembert cheese
Wilfried Ablain1,2,3, Sylvie Hallier Soulier3, David Causeur4,Michel Gautier1,2, Florence Baron1,2*
1 Agrocampus Ouest, Laboratoire de Microbiologie Alimentaire, 65 rue de Saint-Brieuc,35042 Rennes Cedex, France
2 INRA, UMR1253, Laboratoire de Science et Technologie du Lait et de l’Œuf,65 rue de Saint-Brieuc, 35042 Rennes Cedex, France
3 Genesystems, 1 rue du Courtil, Centre CICEA, 35170 Bruz, France4 Agrocampus Ouest, Laboratoire de Mathématiques Appliquées, 65 rue de Saint-Brieuc,
35042 Rennes Cedex, France
Received 17 April 2008 – Accepted 21 October 2008
Abstract – Transcriptomic studies of microorganisms are dependent upon the efficiency of the RNAextraction procedure. In this study, we compared different methods used to disrupt bacterial cellsthat are frequently described in the literature, such as mechanical (sonication, bead beating) and en-zymatic (lysozyme or lysostaphin digestion) disruption. Factorial designs and ANOVA procedureswere used to compare statistically the efficiency of these protocols on Staphylococcus aureus. Theresults were assessed in terms of quality and quantity of RNA extract suitable for further quantitativereverse transcriptase PCR (qRT-PCR) analysis. We selected a simple, rapid (in less than four hours)and sensitive RNA extraction/purification protocol based on lysostaphin treatment, followed by abead-beating procedure. This method allowed an excellent recovery (> 85%) of 16S rRNA fromover a wide range of CFU (109 to 102 CFU·mL−1), efficient on different S. aureus strains in bothexponential and stationary growth phases in pure culture. Application of the protocol for the exam-ination of artificially contaminated Camembert cheese achieved sensitivities of 1.1 × 102 copies ofthe 16S rRNA gene·g−1 of cheese. This protocol constitutes an essential tool for gene expressionstudies of S. aureus in Camembert cheese.
Résumé – Mise au point d’une méthode rapide et simple de lyse de Staphylococcus aureus,optimisée pour une analyse par RT-PCR en temps réel : Application à l’examen de camem-bert. Le succès de l’étude du transcriptome d’un microorganisme dépend de l’efficacité de l’extrac-tion des ARN. Par cette étude, nous avons comparé différentes méthodes fréquemment rencontréesdans la littérature pour lyser les bactéries telles que les lyses mécaniques (sonication, bead beating)et enzymatiques (digestion au lysozyme ou à la lysostaphine). L’utilisation de plans d’expérience apermis de comparer statistiquement l’efficacité des différents protocoles sur Staphylococcus aureus.Les résultats sont comparés selon la qualité et la quantité des ARN extraits, par reverse transcrip-tase PCR quantitative (qRT-PCR). Nous avons sélectionné un protocole d’extraction/purificationdes ARN, simple, rapide (en moins de quatre heures) et sensible, basé sur une digestion à la lysos-taphine suivie d’un cassage par billes. Cette méthode permet une excellente récupération (> 85 %)de l’ARNr 16S à partir d’une large gamme d’UFC (109 à 102 UFC·mL−1), et est efficace sur diffé-rentes souches de S. aureus en culture pure, en phase exponentielle ou stationnaire de croissance.L’application du protocole pour l’examen d’un camembert artificiellement contaminé permet de dé-tecter jusqu’à 1.1×102 copies d’ARNr 16S·g−1 de fromage. Ce protocole constitue un outil essentielpour l’étude de l’expression des gènes de S. aureus en matrice camembert.
During the past few years, transcrip-tomic studies using quantitative reversetranscriptase PCR (qRT-PCR) have beendeveloped considerably to study bacterialcomportment, virulence and drug resis-tance.
One of the main hurdles to qRT-PCRis the recovery of total RNA directly fromthe sample. Indeed, if the efficient recov-ery of RNA extract in large amount is es-sential, the purity and integrity of RNA arecritical elements for the overall success ofRNA-based analyses [15]. The extractionand purification procedure of total RNA re-quires the recovery of high-quality RNAthat is undegraded, and free of: proteins,DNA, inhibitors for RT and real-time PCRreaction, and nucleases to further qRT-PCRstudies [9].
A variety of methods have been de-veloped for the preparation of cell-freeextracts, such as mechanical (sonication,
blending or bead beating) and enzymatic(lysozyme or lysostaphin digestion) dis-ruption of the bacterial cells [2, 4, 6, 12, 16,17, 21]. These procedures have been usedsuccessfully with a variety of microorgan-isms and culture conditions.
In our work, we evaluated the effectsof different methods, applied for bacte-rial lysis. The study was carried out onStaphylococcus aureus, which is knownto be very difficult to disrupt. More-over, Staphylococcus aureus is consideredan important pathogen due to a com-bination of toxin-mediated virulence, in-vasiveness and antibiotic resistance. Thisbacterium is involved in nosocomial infec-tions and in food poisoning. Consumptionof foods containing this microorganism isoften identified as the cause of illness.Intoxication is characterized by vomitingand diarrhea resulting from the ingestionof foods or beverages containing one ormore preformed enterotoxins produced byStaphylococcus aureus [9, 14].
RNA extraction of S. aureus for qRT-PCR 71
The aim of this study was to select asimple, rapid and efficient lysis protocolusable for the study of RNA content ofS. aureus by qRT-PCR.
2. MATERIALS AND METHODS
2.1. Bacterial strains, cultureconditions and samplepreparation
Ten S. aureus strains were used: 3 labo-ratory strains (RN 4220, Mu50 and N315),2 isolated from humans (CHU Rennes) and5 from milk products (milk and cheese).Only the RN4220 strain was used for RNAextraction optimizations.
All strains were stored at –80 ◦Cin Tryptone Soy Broth (TSB, AES,Combourg, France) supplemented with15% (vol/vol) glycerol. For all experi-ments, an overnight culture was preparedfrom frozen bacterial stock in 5 mL of TSBat 37 ◦C, and the subcultures were grownafter 1/100 dilution in TSB with shaking(250 rpm) at 37 ◦C. Population levels wereenumerated on Tryptone soy agar (TSA,AES) after 24 h of incubation at 37 ◦C.
For optimization of lysis procedures,2.5 mL of culture were added to 2 volumesof RNA protect bacteria reagent (QiagenInc., Courtabœuf, France). After vortexing(30 s), the mixture was incubated at roomtemperature (5 min) and then diluted ten-fold in sterile Tryptone salt (TS, (Tryptone1 g·L−1, Sodium Chloride 8.5 g·L−1), AESLaboratoires, Combourg, France).
One-hundred-microliter aliquots con-taining 106 bacteria were snap-frozen inliquid nitrogen and stored at –80 ◦C be-fore analysis. For the sensitivity studies,the aliquots were diluted in tryptone salt toachieve the required bacteria level beforelysis.
2.2. Food sample analysis
Twenty-five grams of French Camem-bert (local supermarket) were homoge-nized (45 s) with 225 mL of buffered-salinepeptone water (AES) in a Stomacher 400(Seward, London, UK). Fifty-milliliter-aliquots of homogenate were inoculatedwith 2 mL of varying dilutions of strainRN4220 (log phase) or TS (negative con-trol). After centrifugation at 5500× g for5 min at 4 ◦C, the pellet was washedthree times with Chelex 25% (w/v; Sigma-Aldrich Co., Missouri, USA). The fi-nal pellet (containing bacteria and Chelexbeads) was resuspended in 100 µL ofTE 1 X (TrisHCl 10 mmol·L−1; EDTA1 mmol·L−1) and then used for RNA orDNA extraction, as described below.
2.3. Lysis protocol
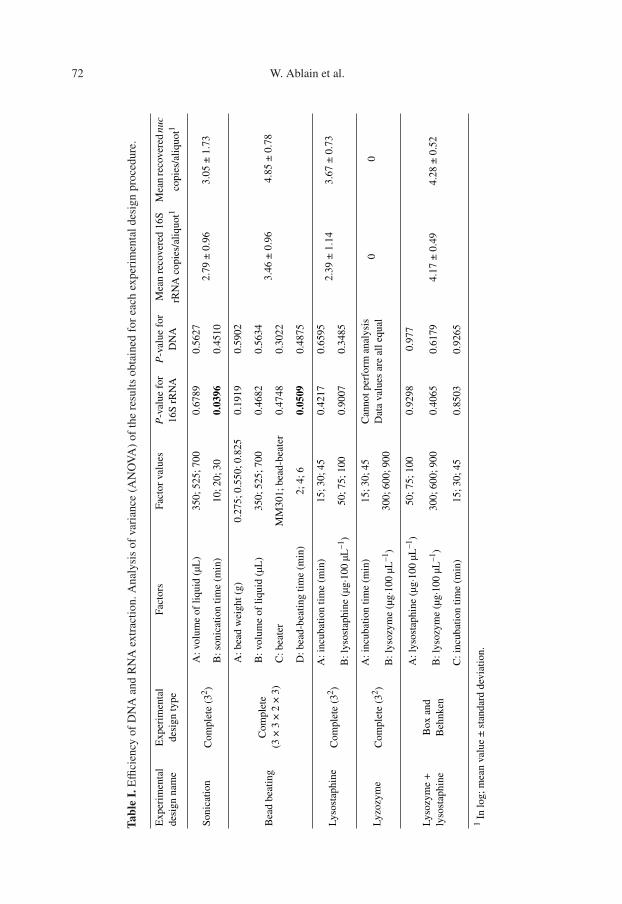
In this study, three extraction techniqueswere compared in terms of DNA and RNAextraction yield: enzymatic treatment, son-ication and bead-beating homogenization.The experiments were carried out accord-ing to the experimental design proceduresdescribed in Table I.
2.3.1. Enzymatic digestion
Incubation time and the enzymatic con-centration were tested for two different en-zymes (lysozyme and lysostaphin, Sigma-Aldrich Co.). Ten microliters of enzymaticsolution (concentrations are given in Tab. I)were added to 100 µL of frozen bacteriaaliquots and incubated at 37 ◦C for 15, 30or 45 min.
2.3.2. Sonication
Two parameters were evaluated: the vol-ume of cellular suspension and the sonica-tion time. Volumes of 350, 525 or 700 µLof lysis buffer (see DNA and RNA purifi-cation procedure) were added to 100 µL
72 W. Ablain et al.
Tabl
eI.
Effi
cien
cyof
DN
Aan
dR
NA
extr
acti
on.A
naly
sis
ofva
rian
ce(A
NO
VA
)of
the
resu
lts
obta
ined
for
each
expe
rim
enta
ldes
ign
proc
edur
e.
Exp
erim
enta
l de
sign
nam
eE
xper
imen
tal
desi
gn ty
peFa
ctor
sFa
ctor
val
ues
P-v
alue
for
16
S r
RN
AP
-val
ue f
or
DN
AM
ean
reco
vere
d 16
S
rRN
A c
opie
s/al
iquo
t1M
ean
reco
vere
d nu
c
copi
es/a
liqu
ot1
Son
icat
ion
Com
plet
e (3
2 ) A
: vol
ume
of li
quid
(µL
)35
0; 5
25; 7
000.
6789
0.56
27
B: s
onic
atio
n ti
me
(min
) 10
; 20;
30
0.03
960.
4510
Bea
d be
atin
gC
ompl
ete
(3 ×
3 ×
2 ×
3)
A: b
ead
wei
ght (
g)
0.27
5; 0
.550
; 0.8
250.
1919
0.59
02
B: v
olum
e of
liqu
id (
µL)
350;
525
; 700
0.46
820.
5634
C: b
eate
r
MM
301;
bea
d-be
ater
0.47
480.
3022
D: b
ead-
beat
ing
tim
e (m
in)
2; 4
; 60.
0509
0.48
75
Lys
osta
phin
eC
ompl
ete
(32 )
A: i
ncub
atio
n ti
me
(min
) 15
; 30;
45
0.42
170.
6595
B: l
ysos
taph
ine
(µg·
100
µL–1
)50
; 75;
100
0.90
070.
3485
Lyz
ozym
eC
ompl
ete
(32 )
A: i
ncub
atio
n ti
me
(min
)15
; 30;
45
Can
not p
erfo
rm a
naly
sis
D
ata
valu
es a
re a
ll eq
ual
00
B: l
ysoz
yme
(µg·
100
µL–1
)30
0; 6
00; 9
00
Lys
ozym
e +
ly
sost
aphi
neB
ox a
nd
Beh
nken
A: l
ysos
taph
ine
(µg·
100
µL–1
)50
; 75;
100
0.92
980.
977
B: l
ysoz
yme
(µg·
100
µL–1
)30
0; 6
00; 9
000.
4065
0.61
79
C: i
ncub
atio
n ti
me
(min
)
15; 3
0; 4
50.
8503
0.92
65
1 In
log;
mea
n va
lue
± s
tand
ard
devi
atio
n.
2.79
± 0
.96
3.05
± 1
.73
3.46
± 0
.96
4.85
± 0
.78
2.39
± 1
.14
3.67
± 0
.73
4.17
± 0
.49
4.28
± 0
.52
RNA extraction of S. aureus for qRT-PCR 73
of defrosted bacteria aliquots and soni-cated for 10, 20 or 30 min at 20 ◦C in a20-liter sonication bath (88170 BioblockScientific).
2.3.3. Bead beating
Four parameters were tested: the vol-ume of cellular suspension, the quantity ofbeads, the bead-beating time and the sup-plier of the beater. Volumes of 350, 525or 700 µL of corresponding lysis buffer(see DNA and RNA purification proce-dure) and 0.275, 0.550 or 0.825 mg of acid-washed zirconium beads (Sigma-AldrichCo.) were added to 100 µL defrosted bac-teria aliquot, then bead-beaten for 2, 4 or6 min. The preparations were treated byalternating 1-min cycles of beating withincubation on ice for 2 min. Two dif-ferent beaters were tested: the MM301(Qiagen) at 30 Hz, and the Beadbeater celldisrupter (Biospec Products; Bartlesville,Oklahoma), at 3200 rpm speed.
2.3.4. Experimental design
For each lysozyme and lysostaphin di-gestion, and for sonication and bead-beating protocols, the lysis was performedaccording to a complete factorial de-sign. For simultaneous lysostaphin andlysozyme digestion, a Box-Behnken de-sign [7] was used. The order of the exper-iments was randomized for each protocol.Each experimental design was performedonly once.
DNA and RNA quantity and qualitywere taken into account to evaluate the per-formance of the lysis protocols.
2.4. DNA and RNA purificationprocedure
Bacterial genomic DNA and totalRNA were purified using the Nucle-ospin DNA plant L kit or the Nucle-ospin RNA II minikit (Macherey-Nagel
GmbH & Co. KG, Düren, Germany) ac-cording to the manufacturer’s recommen-dations, but modified as follows, before thelysate clarification state: the crude lysateswere completed at 700 µL with C4 so-lution/EtOH (vol/vol) mix or with RA1solution/beta-mercapto-ethanol mix for theDNA and RNA protocol, respectively. Theelution was performed in a 200-µL or60-µL one-step procedure for DNA andRNA, respectively. After elution, the sam-ples were frozen immediately and stored at–80 ◦C until use. Contaminating genomicDNA was removed from total RNA usingthe DNA-free kit (Ambion, Austin, Texas,USA), according to the manufacturer’s rec-ommendations. The absence of DNA inRNA extracts was assessed by real-timePCR targeting of the 16S rRNA gene.
2.5. Oligonucleotide primersand probes used
S. aureus-specific oligonucleotides andfluorogenic probes (Tab. II), targeting thenuc single-copy gene and the 16S rRNA5-copy gene were designed on the basis ofthe Mu50 strain gene (accession numberID: 158878) with Primer Express, soft-ware version 2.0 (Applied Biosystems,Foster City, California, USA). Our primersand probes specifically distinguishedS. aureus strains from nontarget bac-teria, as assessed on 10 staphylococcal(strains RN4220; LMA1049; LMA1053;LMA1054; LMA1093; LMA1094;LMA1095; LMA1177; LMA1178; Mu50)and 13 “non-staphylococcal” strains(Pseudomonas aeruginosa; Escherichiacoli; Klebsellia oxytoca; Klebselliapneumoniae; Enterococcus faecalis;Enterococcus faecium CIP; Enterococcusdurans; Streptococcus bovis; Strepto-coccus equinus; Listeria monocytogenesEfGDe; Salmonella enteritidis; Bacil-lus cereus; Yersinia enterocolitica) and
74 W. Ablain et al.
Table II. Oligonucleotides used for both real-time PCR and qRT-PCR assays.
Target gene Accession n◦ Name Type Sequence
nuc gi|46623 nuc SaQfor Forward primer 5’-ATG GAC GTG GCT TAG CGT AT-3’
one “non-S. aureus” (Staphylococcusepidermidis) strain (data not shown).
2.6. Real-time PCR conditions
The assays were performed in duplicatewith a 20-µL real-time PCR amplificationmixture containing 15 µL of PCR mix (1 XTaq DNA polymerase buffer (Qiagen Inc.,Valencia, California, USA), 2.5 mmol·L−1
MgCl2 (Sigma-Aldrich), 200 µmol·L−1
dNTPs mix (Sigma-Aldrich), 0.4 µmol·L−1
forward and reverse primers, 0.2 µmol·L−1
fluorogenic FAM-TAMRA-labeled probe,0.1% BSA (w/w) (Sigma-Aldrich), 0.01%Tween 20 (w/w), 0.75 U Taq DNA poly-merase (Qiagen) and 5 µL DNA or cDNA.Reactions were done with an iCycler ap-paratus (Bio-Rad Laboratories, California,USA) with the following program: 4 minat 95 ◦C, followed by 46 cycles of 15 s at95 ◦C and 45 s at 57 ◦C (nuc gene phase)or 58 ◦C (16S rRNA gene phase).
The standard curves were generated byusing tenfold dilution series (2.44 × 100 to2.44× 104 genome units (GU)/well) of ge-nomic DNA. Purified genomic DNA solu-tion (3.108 GU·mL−1) was quantified us-ing PicoGreen (Molecular Probes, Eugene,Oregon, USA) in a luminescence spec-trometer LS50B (Perkin-Elmer, Norwalk,Connecticut, USA), and diluted to theworking concentration in double-distilledwater. The corresponding genome unitsnumber was calculated on the basis ofthe size of the S. aureus genome [13]:
one molecule of genomic DNA corre-sponded to 3.23 fg of DNA. All reac-tions were performed in duplicate. The95% confidence interval was calculatedfor each serial dilution, according to a bi-nomial distribution [11, 18] by using theSTATGRAPHICS Plus software, version5.0 (Chicago, Illinois, USA).
Quantitative RT-PCRs were conductedin two-step reactions using the iCycler(Bio-Rad). SuperScript II reverse tran-scriptase (Invitrogen, Carlsbad, California,USA) was used to produce single-strandcDNA from 5 µL of purified total RNAaccording to the supplier’s instructions.The cDNA synthesis was primed by the16S SaQrev primer (Tab. II) at 42 ◦C for50 min.
2.8. Statistical analysis
For each lysis protocol, RNA quan-tity and quality were measured by real-time RT-PCR, while DNA quantity andquality were measured by real-time PCR.The STATGRAPHICS Plus 5.0 software(Chicago, Illinois, USA) was used for allstatistical analyses for RNA and DNAquantities. Multiple group comparisonswere made using analysis of variance(ANOVA).
RNA extraction of S. aureus for qRT-PCR 75
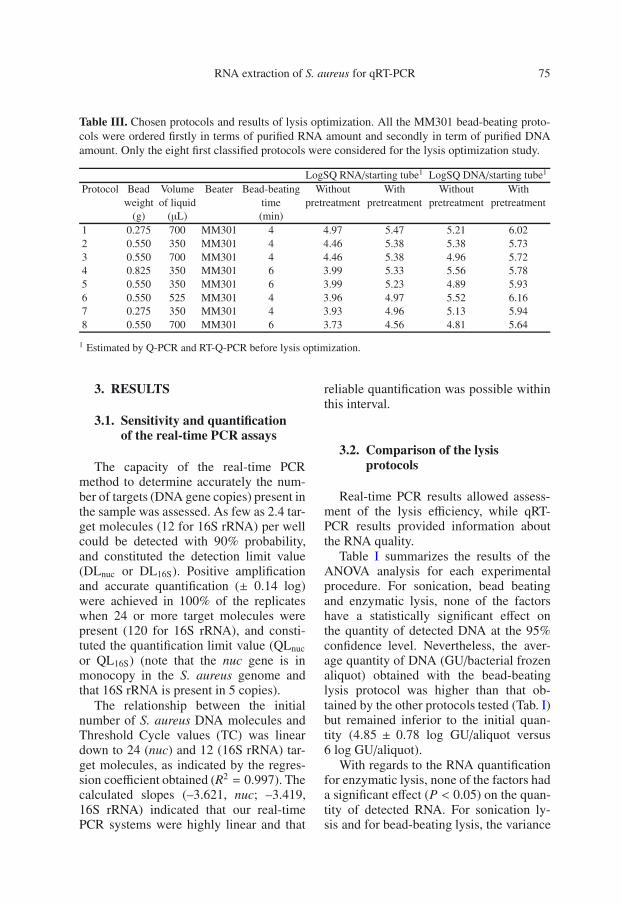
Table III. Chosen protocols and results of lysis optimization. All the MM301 bead-beating proto-cols were ordered firstly in terms of purified RNA amount and secondly in term of purified DNAamount. Only the eight first classified protocols were considered for the lysis optimization study.
LogSQ RNA/starting tube1 LogSQ DNA/starting tube1
Protocol Bead Volume Beater Bead-beating Without With Without Withweight of liquid time pretreatment pretreatment pretreatment pretreatment
1 Estimated by Q-PCR and RT-Q-PCR before lysis optimization.
3. RESULTS
3.1. Sensitivity and quantificationof the real-time PCR assays
The capacity of the real-time PCRmethod to determine accurately the num-ber of targets (DNA gene copies) present inthe sample was assessed. As few as 2.4 tar-get molecules (12 for 16S rRNA) per wellcould be detected with 90% probability,and constituted the detection limit value(DLnuc or DL16S). Positive amplificationand accurate quantification (± 0.14 log)were achieved in 100% of the replicateswhen 24 or more target molecules werepresent (120 for 16S rRNA), and consti-tuted the quantification limit value (QLnuc
or QL16S) (note that the nuc gene is inmonocopy in the S. aureus genome andthat 16S rRNA is present in 5 copies).
The relationship between the initialnumber of S. aureus DNA molecules andThreshold Cycle values (TC) was lineardown to 24 (nuc) and 12 (16S rRNA) tar-get molecules, as indicated by the regres-sion coefficient obtained (R2 = 0.997). Thecalculated slopes (–3.621, nuc; –3.419,16S rRNA) indicated that our real-timePCR systems were highly linear and that
reliable quantification was possible withinthis interval.
3.2. Comparison of the lysisprotocols
Real-time PCR results allowed assess-ment of the lysis efficiency, while qRT-PCR results provided information aboutthe RNA quality.
Table I summarizes the results of theANOVA analysis for each experimentalprocedure. For sonication, bead beatingand enzymatic lysis, none of the factorshave a statistically significant effect onthe quantity of detected DNA at the 95%confidence level. Nevertheless, the aver-age quantity of DNA (GU/bacterial frozenaliquot) obtained with the bead-beatinglysis protocol was higher than that ob-tained by the other protocols tested (Tab. I)but remained inferior to the initial quan-tity (4.85 ± 0.78 log GU/aliquot versus6 log GU/aliquot).
With regards to the RNA quantificationfor enzymatic lysis, none of the factors hada significant effect (P < 0.05) on the quan-tity of detected RNA. For sonication ly-sis and for bead-beating lysis, the variance
76 W. Ablain et al.
of time had a statistically significant ef-fect (P < 0.05) on the detected quantityof RNA (Tab. I). In fact, for sonication ly-sis, a procedure time of 10 min was signif-icantly (P < 0.05) more effective, while aprocedure time of 4 min was significantlymore effective (P < 0.05) for bead beating.The average quantity of RNA (16S rRNAcopies/bacterial frozen aliquots) obtainedfor a bead-beating time of 4 min (3.9 ±0.52 log) was higher than that obtainedwith 10 min of sonication (3.4 ± 0.39 log),suggesting that bead-beating lysis was bet-ter for RNA extraction (data not shown).
For lyzosyme lysis, regardless of thetested conditions, no DNA or RNA was de-tectable by real-time PCR or qRT-PCR, re-spectively, suggesting that lyzosyme wasnot effective for digestion of the staphylo-coccal cell wall.
Concerning both real-time PCR andqRT-PCR signals, bead-beating lysisappeared to be the most effective protocol,but did not allow 100% lysis efficiency.These results suggest that the condi-tions for this lysis procedure have to beimproved.
3.3. Effect of lysostaphinpretreatment on thebead-beating lysis efficiency
In order to improve the bead-beatingprocedure, we studied the effect of enzy-matic pretreatment on bead-beating lysisefficiency. Since the type of beater did nothave a statistically significant effect on thequantity of either RNA or DNA, we se-lected the MM301 beater for the study,which permitted more analyses to be con-ducted at the same time. Since none of thefactors tested for lysostaphin lysis had astatistically significant effect (P > 0.05)on the quantity of detected DNA andRNA (Tab. I), we chose both the low-est dose and the shortest incubation time.The chosen protocols are summarized in
Table III. ANOVA analysis showed thatlysostaphin pretreatment significantly (P =0.001) improved bead-beating lysis effi-ciency for all protocols tested (using 106
bacteria frozen aliquots). On average, theincrease in amount was approximately 0.7log for DNA (5.86 ± 0.18 log of nuccopies/aliquot with lysostaphin pretreat-ment versus 5.16 ± 0.31 log without)and 1 log for RNA (5.17 ± 0.27 log of16S rRNA copies/aliquot with lysostaphinpretreatment versus 4.21 ± 0.45 logwithout) (Tab. III). With regards to RNAquantities, protocol 2 appeared to be statis-tically the most efficient for RNA extrac-tion (Student’s t-test P = 0.05) (data notshown). Hence, this protocol was selectedfor the next study.
3.4. Protocol efficiency with differentstaphylococcal strainsand growth states
The efficiency of the selected protocolwas tested with 10 different staphylococ-cal strains (in the stationary state). For onestrain (RN4220), 2 different growth stateswere studied. The protocol was efficient forall the strains for both DNA and RNA ex-tractions (data not shown). The mean re-covery rates were 92.4 ± 12.5% for DNAand 67.4±19.8% for RNA, respectively (asdetermined by relating GU or 16S rRNAcopy numbers to CFU·mL−1). For strainRN4220, the DNA recovery rate obtainedfor exponential (103.2±1.1%) and station-ary (104.0 ± 1.3%) growth phases showedthat lysis was equally efficient for bothstates (P < 0.001). Nevertheless, for theRNA quantity, a significant (P < 0.05)difference (32.3 ± 4.7% versus 122.7 ±15.5%) could be observed, suggesting that16S rRNA was more abundant in the mi-crobial cell during its exponential statethan its stationary phase.
RNA extraction of S. aureus for qRT-PCR 77
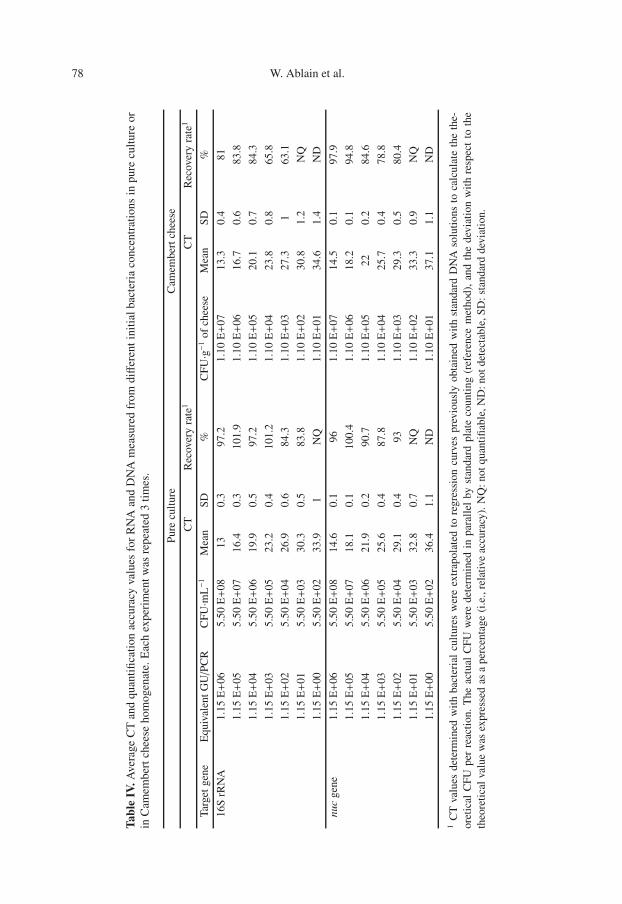
3.5. Sensitivity of the optimalprotocol
The sensitivity of the lysis protocolassay was investigated by using differentinitial quantities of strain RN4220 in theexponential phase. The results are summa-rized in Table IV. For the qRT-PCR as-says, the overall quantification limit was5.5×103 CFU·mL−1 (i.e. 1.15×101 equiv-alent GU/PCR), and the detection limitvalue was tenfold inferior. The linear re-gression analysis of CT values related tobacterial numbers in the reactions yieldeda R2 value of 0.999 with a slope sim-ilar (–3.45) to that obtained with puri-fied genomic DNA (–3.41), indicating thatour qRT-PCR assays accurately quantifiedS. aureus 16S rRNA on the interval. Fur-thermore, these results suggested that ourprotocol could be applied to gene expres-sion level studies with as little as 5.5 ×103 CFU·mL−1.
3.6. Food sample analysis
The ability of our procedure to de-tect S. aureus 16S rRNA in Camembertcheese was evaluated. The results showedsuccessful amplification in all the con-taminated cheese samples, while no sig-nal was detected in uninoculated foods(Tab. IV). The 16S rRNA amplicon was de-tected down to concentrations of 1.1 × 102
copies of 16S rRNA·g−1 of Camembert(i.e. 1.15 × 101 equivalent GU/PCR when5 g of cheese are analyzed), and 1.1 × 103
copies of 16S rRNA·g−1 were quantifiedwith 63% of accuracy.
4. DISCUSSION
In the literature, lysis protocols are usu-ally described for RNA extraction fromGram-positive bacteria, but the particularconditions of the tests are different from
those encountered in diagnostics. In orderto select the most efficient protocol, wetested different protocols using five screen-ing experimental design procedures: soni-cation, lysostaphin and/or lysozyme lysis,and bead beating in conditions frequentlydescribed in the literature. The resultsobtained by enzymatic lysis confirmedthat S. aureus was completely resistantto lysozyme, as previously demonstratedby Bera et al. [1, 3]. On the other hand,the lysostaphin lysis appeared largely effi-cient in releasing nucleic acids from S. au-reus, for both real-time PCR and qRT-PCRapplications. Nevertheless, the lysostaphinlysis was not complete. Indeed, the numberof nuc copies detected (4.67 log GU·mL−1)did not reach the number of cells subjectedto the disruption (7 log GU·mL−1).
For the combined lysozyme andlysostaphin lysis, the yield of extractednucleic acid was slightly better than thatobtained for lysostaphin lysis alone. Theseresults showed that lyzosyme potentatesthe lysostaphin anti-staphylococcal activ-ity, as previously observed by Cisani et al.,but the effects of lyzozyme are quite weak[10]. Two mechanical disruptions werealso tested: sonication and bead-beatingmethods. The sonication procedure wasnot very effective for either DNA or RNAextraction. The use of a more completeexperimental design, integrating sonica-tion generator power and the ultra-soundfrequency could allow for optimizationof this lysis [16]. For the bead-beatingprotocol, a preliminary study showed thatzirconium beads were more efficient thanglass beads. The bead-beating procedureappeared to be the most efficient forDNA and RNA extraction; however, lysiswas not complete, suggesting that thebead-beating protocol requires furtheroptimization. Note that both mechanicaldisruption methods presented an optimumrecovery rate for 16S rRNA, using theminimal experimental conditions (time,power, etc.). Longer disruption treatments
78 W. Ablain et al.
Tabl
eIV
.Ave
rage
CT
and
quan
tifi
cati
onac
cura
cyva
lues
for
RN
Aan
dD
NA
mea
sure
dfr
omdiff
eren
tini
tial
bact
eria
conc
entr
atio
nsin
pure
cult
ure
orin
Cam
embe
rtch
eese
hom
ogen
ate.
Eac
hex
peri
men
twas
repe
ated
3ti
mes
.
Pure
cult
ure
Cam
embe
rtch
eese
CT
Rec
over
yra
te1
CT
Rec
over
yra
te1
Targ
etge
neE
quiv
alen
tGU/P
CR
CFU·m
L−1
Mea
nSD
%C
FU·g−
1of
chee
seM
ean
SD%
16S
rRN
A1.
15E+
065.
50E+
0813
0.3
97.2
1.10
E+
0713
.30.
481
1.15
E+
055.
50E+
0716
.40.
310
1.9
1.10
E+
0616
.70.
683
.8
1.15
E+
045.
50E+
0619
.90.
597
.21.
10E+
0520
.10.
784
.3
1.15
E+
035.
50E+
0523
.20.
410
1.2
1.10
E+
0423
.80.
865
.8
1.15
E+
025.
50E+
0426
.90.
684
.31.
10E+
0327
.31
63.1
1.15
E+
015.
50E+
0330
.30.
583
.81.
10E+
0230
.81.
2N
Q
1.15
E+
005.
50E+
0233
.91
NQ
1.10
E+
0134
.61.
4N
D
nuc
gene
1.15
E+
065.
50E+
0814
.60.
196
1.10
E+
0714
.50.
197
.9
1.15
E+
055.
50E+
0718
.10.
110
0.4
1.10
E+
0618
.20.
194
.8
1.15
E+
045.
50E+
0621
.90.
290
.71.
10E+
0522
0.2
84.6
1.15
E+
035.
50E+
0525
.60.
487
.81.
10E+
0425
.70.
478
.8
1.15
E+
025.
50E+
0429
.10.
493
1.10
E+
0329
.30.
580
.4
1.15
E+
015.
50E+
0332
.80.
7N
Q1.
10E+
0233
.30.
9N
Q
1.15
E+
005.
50E+
0236
.41.
1N
D1.
10E+
0137
.11.
1N
D
1C
Tva
lues
dete
rmin
edw
ith
bact
eria
lcu
ltur
esw
ere
extr
apol
ated
tore
gres
sion
curv
espr
evio
usly
obta
ined
wit
hst
anda
rdD
NA
solu
tion
sto
calc
ulat
eth
eth
e-or
etic
alC
FUpe
rre
acti
on.
The
actu
alC
FUw
ere
dete
rmin
edin
para
llel
byst
anda
rdpl
ate
coun
ting
(ref
eren
cem
etho
d),
and
the
devi
atio
nw
ith
resp
ect
toth
eth
eore
tica
lval
uew
asex
pres
sed
asa
perc
enta
ge(i
.e.,
rela
tive
accu
racy
).N
Q:n
otqu
anti
fiabl
e,N
D:n
otde
tect
able
,SD
:sta
ndar
dde
viat
ion.
RNA extraction of S. aureus for qRT-PCR 79
decreased the 16S rRNA yield. This couldbe due to 16S rRNA degradation occuringby stronger lysis treatments.
Lysostaphin pretreatment applied be-fore the bead-beating procedure signif-icantly (P-value = 0.05) improved thenucleic acid yield. Moreover, the stud-ies using different staphylococcal strainsdemonstrated that our protocol was ef-fective for DNA and RNA recovery forall the strains tested. Our protocol wasequally effective for the two physiologi-cal states tested: cells collected from ex-ponential or stationary growth states. Fur-thermore, there were fourfold more copiesof 16S rRNA detectable in log phase bacte-ria than in stationary phase bacteria. Theseresults are in agreement with those ob-tained by Vandecasteele et al. [19, 20],who showed that the ratio cDNA/CFU for16S rRNA was 25 during the exponentialphase for S. epidermidis and was 3 duringthe stationary phase.
The most challenging aspect of design-ing methods for the detection of food-borne pathogens is the achievement of alow detection limit. This goal is of particu-lar importance for S. aureus, which is oftenpresent in low numbers in food products[1]. Transcriptomic studies of pathogenbacteria by qRT-PCR require extraction ofbacterial total RNA content. Actually, theaccuracy of gene expression evaluation isrecognized to be influenced by the quan-tity and quality of starting RNA. The qRT-PCR results were very similar to the quan-titative results obtained by the plate countmethod over a five log range of CFU.This indicated that our method allowedan accurate quantification of S. aureus16S rRNA at concentrations as low as 5.5×103 CFU·mL−1 (i.e. 1.15 × 101 equivalentGU/PCR) in broth cultures, while the sen-sitivity was reduced by ten in Camembertcheese (1.1 × 103 CFU·g−1 of Camembert,i.e. 1.15 × 102 equivalent GU/PCR). Notethat 50 mL of homogenate (correspond-ing to 5 g of cheese) was analyzed versus
only 100 µL for broth culture samples. Infact, problems in applying a RT-PCR as-say to foods can arise from various fac-tors [8] such as the presence of substrateschelating magnesium ions necessary forPCR, degradation of nucleic acids and/orprimers by RNases or DNases, and directinhibition of DNA polymerase and reversetranscriptase. The degree of inhibition isgreatly dependent on the type of food. Theremoval of food components such as pro-teins, fats and polysaccharides can posi-tively affect the yield and quality of ex-tracted nucleic acids [5]. Therefore, thelevel of contaminating substances was re-duced by the introduction of specific ad-ditional steps during the processes of ex-traction from cheese samples. The lowersensitivity of the assay for Camembert ver-sus pure culture was probably due to agreater loss of cells (and the consequentminor RNA yield), as a result of the morecomplex extraction procedure required forthis kind of sample. Nonetheless, theseresults demonstrate the suitability of RT-PCR for studying the expression of genesof S. aureus in Camembert; it could be suc-cessfully adopted for use with other foods.However, the method should also be testedon naturally contaminated samples, sincedistribution and physiology of S. aureusin naturally contaminated cheese might bevery different, compared with artificiallycontaminated samples.
Our procedure allows the detection andquantification of 16S rRNA in about 4 h.Using RNA extracts from dilutions ofbacteria, the experiments demonstrated agood linear correlation between the to-tal cell number and the fluorescence sig-nals. The accurate quantification of the16S rRNA obtained from food samplesshowed values lower than the values frompure cultures. However, this difference canbe attributed to two additional variables;namely, the food matrix effect and the pres-ence of endogenous microflora. Indeed, thechemical and genetic complexity of the
80 W. Ablain et al.
naturally contaminated food samples, oftenassociated with a large amount of bacteria,might affect either the quality or the yieldof total RNA, or both.
5. CONCLUSION
In conclusion, we have developed a sim-ple, rapid (in less than one hour) and sen-sitive lysis protocol applicable to differentS. aureus strains. This protocol allowed anextraction of both high quality and largequantity of DNA and RNA, for further real-time PCR and qRT-PCR applications. Thisprotocol could constitute an essential toolfor transcriptomic study of S. aureus, suchas the study of differential RNA content oflive or dead bacteria in Camembert.
Acknowledgements: We thank S. Chevalierfor competent technical assistance.
REFERENCES
[1] Alarcon B., Vicedo B., Aznar R., PCR-basedprocedures for detection and quantificationof Staphylococcus aureus and their applica-tion in food, J. Appl. Microbiol. 100 (2006)352–364.
[2] Aldous W.K., Pounder J.I., Cloud J.L.,Woods G.L., Comparison of six methods ofextracting Mycobacterium tuberculosis DNAfrom processed sputum for testing by quan-titative real-time PCR, J. Clin. Microbiol. 43(2005) 2471–2473.
[3] Bera A., Herbert S., Jakob A., Vollmer W.,Gotz F., Why are pathogenic staphylo-cocci so lysozyme resistant? The peptido-glycan O-acetyltransferase OatA is the ma-jor determinant for lysozyme resistance ofStaphylococcus aureus, Mol. Microbiol. 55(2005) 778–787.
[4] Bergh K., Stoelhaug A., Loeseth K.,Bevanger L., Detection of group B strep-tococci (GBS) in vaginal swabs usingreal-time PCR with TaqMan probe hy-bridization, Indian J. Med. Res. 119 Suppl.(2004) 221–223.
[5] Bleve G., Rizzotti L., Dellaglio F., TorrianiS., Development of reverse transcription
(RT)-PCR and real-time RT-PCR assays forrapid detection and quantification of viableyeasts and molds contaminating yogurts andpasteurized food products, Appl. Environ.Microbiol. 69 (2003) 4116–4122.
[6] Bourrain M., Achouak W., Urbain V., HeulinT., DNA extraction from activated sludges,Curr. Microbiol. 38 (1999) 315–319.
[7] Box G., Behnken D., Some new three-leveldesigns for the study of quantitative vari-ables, Technometrics 2 (1960) 455–475.
[8] Bustin S.A., Absolute quantification ofmRNA using real-time reverse transcriptionpolymerase chain reaction assays, J. Mol.Endocrinol. 25 (2000) 169–193.
[9] Bustin S.A., Nolan T., Pitfalls of quanti-tative real-time reverse-transcription poly-merase chain reaction, J. Biomol. Tech. 15(2004) 155–166.
[10] Cisani G., Varaldo P.E., Grazi G., Soro O.,High-level potentiation of lysostaphinanti-staphylococcal activity by lysozyme,Antimicrob. Agents Chemother. 21 (1982)531–535.
[11] Kay S., Van den Eede G., The limits of GMOdetection, Nat. Biotechnol. 19 (2001) 405.
[12] Kocagoz T., Yilmaz E., Ozkara S., KocagozS., Hayran M., Sachedeva M., ChambersH.F., Detection of Mycobacterium tuber-culosis in sputum samples by polymerasechain reaction using a simplified procedure,J. Clin. Microbiol. 31 (1993) 1435–1438.
[13] Kuroda M., Ohta T., Uchiyama I., Baba T.,Yuzawa H., Kobayashi I., Cui L., OguchiA., Aoki K., Nagai Y., Lian J., Ito T.,Kanamori M., Matsumaru H., MaruyamaA., Murakami H., Hosoyama A., Mizutani-Ui Y., Takahashi N.K., Sawano T., InoueR., Kaito C., Sekimizu K., Hirakawa H.,Kuhara S., Goto S., Yabuzaki J., KanehisaM., Yamashita A., Oshima K., Furuya K.,Yoshino C., Shiba T., Hattori M., OgasawaraN., Hayashi H., Hiramatsu K., Wholegenome sequencing of meticillin-resistantStaphylococcus aureus, Lancet 357 (2001)1225–1240.
[14] Le Loir Y., Baron F., Gautier M.,Staphylococcus aureus and food poisoning,Genet. Mol. Res. 1 (2003) 63–76.
[15] Lin S.L., Chuong C.M., Widelitz R.B., YingS.Y., In vivo analysis of cancerous gene ex-pression by RNA-polymerase chain reaction,Nucleic Acids Res. 27 (1999) 4585–4589.
[17] Rantakokko-Jalava K., Jalava J., OptimalDNA isolation method for detection of bac-teria in clinical specimens by broad-rangePCR, J. Clin. Microbiol. 40 (2002) 4211–4217.
[18] Van den Eede G., Kay S., Anklam E.,Schimmel H., Analytical challenges: bridg-ing the gap from regulation to enforcement,J. AOAC Int. 85 (2002) 757–761.
[19] Vandecasteele S.J., Peetermans W.E.,Merckx R., Van Eldere J., Quantification ofexpression of Staphylococcus epidermidis
housekeeping genes with Taqman quantita-tive PCR during in vitro growth and underdifferent conditions, J. Bacteriol. 183 (2001)7094–7101.
[20] Vandecasteele S.J., Peetermans W.E.,Merckx R., Van Ranst M., Van Eldere J.,Use of gDNA as internal standard for geneexpression in staphylococci in vitro andin vivo, Biochem. Biophys. Res. Commun.291 (2002) 528–534.
[21] Weinrick B., Dunman P.M., McAleese F.,Murphy E., Projan S.J., Fang Y., NovickR.P., Effect of mild acid on gene expressionin Staphylococcus aureus, J. Bacteriol. 186(2004) 8407–8423.