A STUDY OF UREASE ACTIVITY IN THE RUMEN lUCROFLORA OF SHEEP by Graham Alfred Jones A THESIS Submitted to the Faculty of Graduate Studies and Research, McGill University, in partial fulfilment of the requirements for the degree of Doctor of Philosophy Department of Agricultural Bacteriology, Macdonald College of McGUl University, Que bec. April, 196.3
Transcript
A STUDY OF UREASE ACTIVITY
IN THE RUMEN lUCROFLORA OF SHEEP
by
Graham Alfred Jones
A THESIS
Submitted to the Faculty of Graduate Studies and Research, McGill University, in partial fulfilment of the requirements
for the degree of Doctor of Philosophy
Department of Agricultural Bacteriology, Macdonald College of McGUl University, Que bec. April, 196.3
TABLE OF CONTENTS
Page
ACKNON!,EDGmŒN'IS •••••••••••••••••••••••••••••••••••••••••••••••••••• v
CLAIM OF CONTRIBUTION TO KNO\v.LEDGE •••.••••••••••••••••••••••••••••• vi
GENERAL INTRODUCTION •••••••••••••••••••••••••••••••••.•••••••••••••• 1
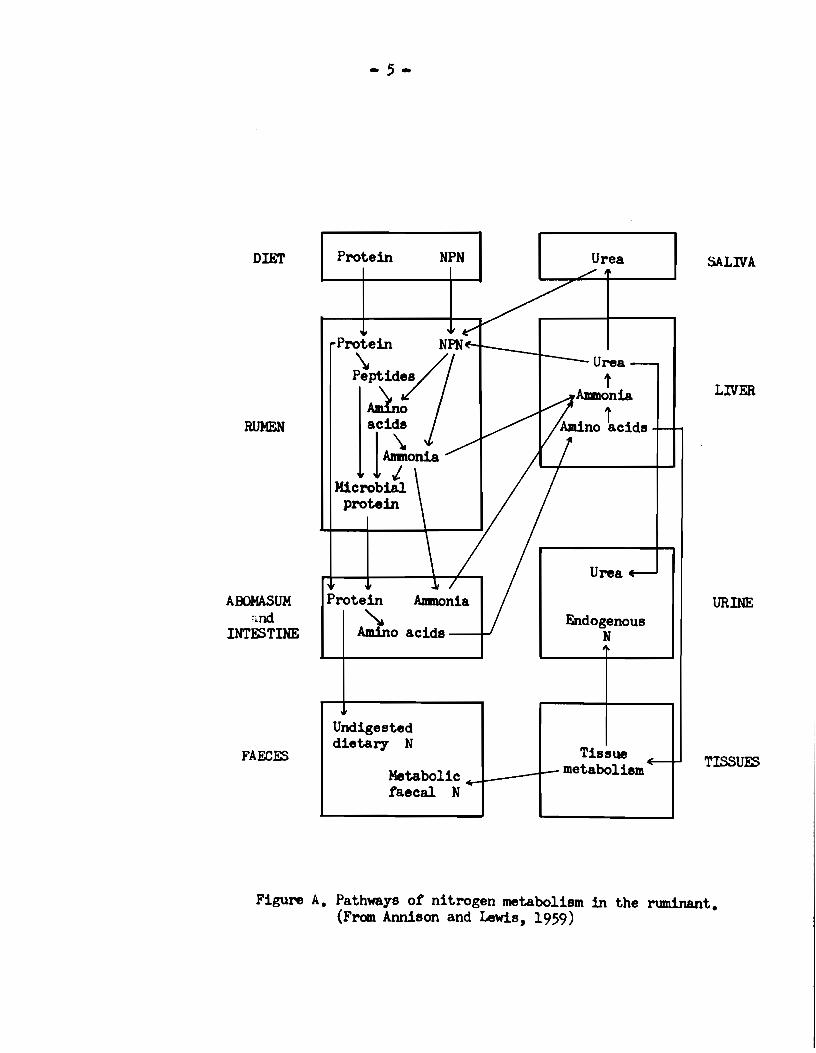
A. Function of the rlllll6n ••••••••••••••••••••••••••••••••••••••• 3 B. Nitrogen metabolism in the rumen •••••••••••••••••••••••••••• 4 C. The utUization of NPN feed supplements by ruminants •••••••• 9
PART I. THE ATT1!11PTED ISOLATION OF ANAEROBIC UREASB-PRODUCING BACTERIA FROM SHmP RUMEN CONTENTS
LITERATURE REVIEW •••••• ·······•·•·····•·•···•·•····••··••·•··••••· 14
A. HYdrolysis of urea in the rumen ••••••••••••••••••••••••••• 14 B. Enumeration of rumen bacteria ••••••••••••••••••••.•••••••• 16 C. Cultural detection of urease production by bacteria ••••••• 17 D. Urease-producing rumen bacteria ••••••••••••••••••••••••••• 20
B. Collection of rumen contents •••••••••••••••••••••••••••••• 25 a) Slleep 2 ••••••••••••••••••••••••••••••••••••••••••••• 25 b) Sheep 35 and 37 •••••••••••••••·••••••••••••••••••••• 25 c) Steers 1 and 2 ••••••••••••••••·•·••••••••••••••••••• 25
c. Preparation or rumen fluid ••••••••••••••••••••••••••••••• 25 D. Fractionation of rumen :tluid ••••••••••••••••••••••••••••• 26
a) Yicrobial and supernatant fractions ••••••••••••••••• 26 b) Subfractionation of the microbial fraction •••••••••• 26 c) Separation of rumen protozoal fraction •••••••••••••• 28
E. Measurement of urease activity ••••••••••••••••••••••••••• 29 F. Bacterial viable counts •.•••••••••••••••••••••••••••••••• 30
a) Total viable counts ••••••••••••••••••••••••••••••••• 31 b) Counts of viable ureolytic organisms •••••••••••••••• 32
- ii -
G. Urease-producing bacteria from rumen fluid ••••••••••••••• 33 a) Isolation ••••••••••.•••••••••••••.••••••••••••••••••• 33
RESUL1S
A. B. c.
i) Isolation from buffered reinforced clostridial broth ..•••.•..•.•••.•.•.•.........•...•........
ii) Isolations from supplemented rumen supernatant 33
liquor medium •••••••••••••••••••••••••••••••••• 34 iii) Isolations of rumen staphylococci by enrichmant
in a brain heart infusion - NaCl medium •••••••• 37 b) Identification of isolated organisms •••••••••••••••• 38 c) Urease activity of ureolytic isolates ••••••••••••••• 38
.......................................................... Urease activity of rumen fluid and its component fractions Proportion of ureolytic bacteria in rumen fluid •••••••••• Attampts at the isolation and characterization of ureaaeproducing bacteria from rumen fluid ••••••••••••••••••••••
39
39 44
a) Isolation of bacteria from cultures in buffered reinforced clostridial medium ••••••••••••••••••••••• 46
A. Factors affecting the activity of jack-bean urease ••••••• 67 a) Urease stimulating agents ••••••••••••••••••••••••••• 67 b) Urease inhibiting agents •••••••••••••••••••••••••••• 69
B. Properties of urease produced by non-rumen bacteria •••••• 70 C. Properties of rumen urease ••••••••••••••••••••••••••••••• 71
MATERIAIS AND METHODS ............................................ 72
A. B.
Collection and preparation of rumen fluid •••••••••••••••• 72 Preparation of ~d rumen ureaae •••••••••••••••••••••••• 72
a) Whole oeil preparations of rumen microorganisme ••••• 72 b) Acetone-dried powders of rumen microorganisme ••••••• 72 c) Cell-free extracts of rumen microorganisme •••••••••• 73
c. D. E.
RESULTS
iii
Preparation of jaek-bean urease •.•....•......••••...••... Preparation of ashed rumen supernatant liquor •••••••••••• Measurement of urease aetivity •••••••••••••••••••••••••••
a) Routin.e method •.••••••.•...•.•...•.•••••.•...•••.••. b) Urease activity in the presence of added cations ••••
D. Estimation of nitrogen in samples ••••••••••••••••••••••• 138 a) Ammonia in rumen supernatant liquor •••••••••••••••• 138 b) Estimation of total nitrogen ••••••••••••••••••••••• 139
E. Nl5 Analyses •••••..•••••••.•.•.•••••••.•.•.•.•••.••••.•• 141 a) Oxidation of ammonia-nitrogen to N2 ••••••••••••.•.• 141 b) Determination of N14JN15 ratio •·••••••••••••••••••• 141
A. Rumen fluid • . • • • • . . . • . • • . . . . . . • • • . • • • • . . . . . . . . . . • . • • • . • • 143 a) pH of rumen fluid •••••••••••••••••••••••••••••••••• 143 b) Ammonia content of rumen supernatant 1iquor • • • • • • • • 143 c) Total nitrogen content of rumen supernatant 1iquor
and its enrichment with N15 •••••••••••••••••••••••• 146 d) Total nitrogen content of the microbial fraction
of rumen fiuid and its enricbment with N15 • • • • • • • • • 149 B. Saliva: N15 enrichment •••••••••••••••••••••••••••••••••• 151 C. Urine: excretion of N15 in urine •·•••••••••••••••••••••• 153
of dilution fluid were nushed with sterile COz and shaken, and 1 ml.
volumes in duplica.te of ea.ch dilution from l/10 to l/lolO -were pla.ted
with the enrichment medium., "Which wa.s solidified with 1.5% agar. The
plates 'Were incuba.ted in Brewer anaerobie jars a.t 37°C for 48 hr. under
an atmosphere of COz and Hz· After incubation, colonies were picked
from the plates into 3 ml. volumes of the enrichment medium.. These
were la.ter transferred to agar slopes of the same medium., and the
liquid cultures "Were tested for the presence of urea.se-producing
bacteria in the manner described in section F(b) above.
iii) Isolations of rumen staphylococci by enricbment in
a brain heart infusion - Na.Cl medium.
Rumen sta.phylococci were isola.ted from the rumen contents of
sheep Z by a modification of the staphylococcus enrichment method of
Wil.son, Poter and I.ew:i.s (~959). One ml.. ot aseptically prepared rumen
f~uid was a.dded to 30 ml. of brain heart infusion (Difco) containing
7.5% NaCl. The suspension wa.s shaken for 9 hr. in a wa.terbath at
37°0 and streak inoculations were tban made on plates of sta.phylococcus
medium no. 110 (Difco). Atter incubation for 36 hr. a.t 37°C, colonies
were picked into brain heart infusion medium. and subsequently
characterized.
- 38-
b) Identification of isolated organisme
Praliminary classification of the isolates into thair
respective genera was made from tm rasults of tests recommended by
Skerman (1959). Subsequent species identification was made according
to Bergey (1957).
c) Urease activity of uraolytic isolates
Each organiam wa.s inoculated on to tryptone glucose 0.2%
yeast extract agar contained in Roux bottles. The inoculated bottles
were incubated at 37°C for 48 hr. The cella were then washed off the
agar with O.OaL-maleate buffer, pH 6.8, and the suspension of cells
was centrifu.ged. The supernatant liquid wa.s decanted and the cells
were wa.shed once by resuspending them in buffer, centrifu.ging the
suspension and again decanting tm supernatant liquid. The cells
from two Roux bottles were finally suspended in about 8 ml. of buffer,
and 3 ml. volumes of this suspension were used for manom.etric measura
ments in duplicata of the urease activity of tm cella. The number of
organisms in the suspension wa.s detennined by a standard count using
sterile 0.9% NaCl. as the dil.uting fl.uid and tryptone gl.ucose 0.2%
yeast extract agar as ths plating medium. Plates were incubated at
37°C for 48 hr.
RESULTS
A. Urease Activity of Rumen Fluid and its Compg!!!nt Fractions.
The urease activities of samples of whole rumen tluid fran
t:œ five experimental anima.ls used in this study are shown in Table I.
The range of activities found with sheep 2 represented a variation in
the amount of urea hydrolyzable to ammonia from 25 - 310 mg. per 100 ml.
of rumen fluid per hr., and therefore indicated a geœrally high leval.
of rumen urease activity. The average activity found with this urea
fed animal was sim.ilar to the levels of activity found in single det
em.ina tions wi th the othar four anima.ls, which were fed more natural
diets. It is evident from the resulta with sheep 2, however, that a
wide range of urease activities may be found from time to tim.e.
Clearly, therefore, a single determination of rumen urease activity
may not be indicative of the generally prevailing urea-hydrolyzing
capacity of rumen fluid from an individual animal.
Ali the experimenta subsequently reported in this section
"Were carried out with rumen fluid samples from sheep 2.
Table II shows the urease acti viti.es of the component
fractions of wbole rumen fluid, together with those of suspensions
of the hay and grain fed to the animal. The urease activity of the
whole fluid was almost quantitatively recovered in the protozoa-free
rumen tluid, tha.t is, in the bacterial fraction. (This fraction was
represented by whole tluid from which the protozoa had been removed
-40-
Table I. Urease activity of whole rumen fiuid from three sheep and two steers.
Sheep 2
Sheep 35
Sheep 37
Steer 1
Steer 2
Urease Activity (~ per ml.)
507lDE
33QlDf.
lEAverage of 30 determinations; range 130 - 1626 lilEResult from 1 determination
-41-
Table II. Urease activity of tractions or rumen tluid.
Fraction
Protozoal fraction
Protozoa-free rumen fiuid (bacteriai traction)
Rumen supernatant liquor
Dietary grain suspension
Dietary hay suspension
Whole rumen fiuid
Urease Activity (qNH_3 par ml.)
4
189
0
0
0
193
- 42-
by a sedimentation technique and subsequent centritugatio~ The
occurrence in the protozoal fraction of a small proportion of the
urease activity of 'Whole rwnen fluid wa.s probably due to the presence
in the suspension of small numbers of ureolytic bacteria. The absence
of ureolytic activity from the suspensions of hay and grain elim.inates
the presence of these components in the bacterial fraction as a poss
ible source of urease, and sinca no animal enzymes are sacreted by
the rumen epithelium (Dukes. 1955), the resulta show that the product
ion of urease in whole rumen fiuid is attributable entirely to the
rumen bacteria. Sinee there was no activity in the relatively cell
free rwnen supernatant liquor, i t is furthermore evident tha t the
enzyme activity assoeiated wi th the bacteria wasex clusively intra
cellula.r.
In order to find out whether urease activity was assoc:ia.ted
with a particular group of morphological types of rumen bacter:ia.,
subfractions of the rumen microbial population wera preparad by
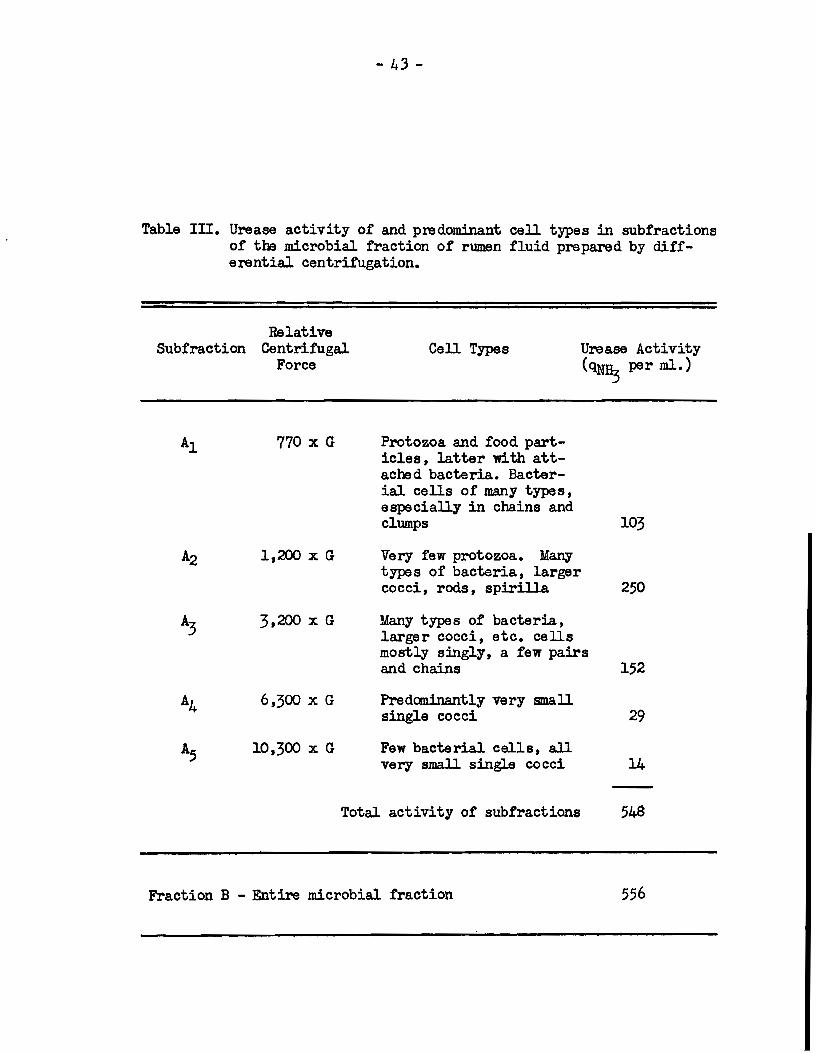
differential centrifugation. Suspensions of these subfractions in
buffer wsre round to have the compositions and urease activities
shown in Table Ill. The resulta show that the bacteria responsible
for about 65% of rumen urease aetivity 'Wera the larger organisme
which were thrown down when tl:e suspension of the entire population
was eentrifuged at 1,200 x G. When occurring in clum.ps or chains,
many of these organisme sedimented at 770 x G. The subfraction
sedimenting at 3,200 x G, which contained many types of smaJJer
- 4:3-
Table Ill. Urease activity of and predomjnant cali types in subfractions of the microbial fraction of rumen fluid prepared by differentia! centrifugation.
Relative Subfraction Centrifuga!
Force
Al 770 x G
A.2 1,200 x G
A.3 3,200 x G
A4 6,300 x G
A5 10,300 x G
Cell Types
Protozoa and food part-icles, latter with att-achad bacteria. Bacter-ial cella of many types, especially in chains and clum.ps
Very few protozoa. Many
Urease Activity {qN~ per ml.)
103
type a of bacteria, larger cocci, roda, apirilla 250
Many types of bacteria, larger cocci, etc. cella most ].y singly, a few pairs and chaina 152
Predaninantly very small single cocci 29
Few ba.cterial cells, ali very sma.ll single cocci 14
-Total activity of subfractions 548
Fraction B - Entire microbial fraction 556
- 44-
bacteria, bad associated with it about 2!7% of the total urease
activity of the rumen microbial population. -Onl.y a com.paratively
low activity was associated with the very small cocci which were
obtained upon centrifuging the suspension above 3,200 x G.
These resulta suggest that rumen urease activity was
associated with a relatively limited number of types of rumen bacteri.a,
which appeared to comprise the larger organisms :round in rumen nuid.
B. Proportion of Ureolrtic Bacteria in Rumen Fluid.
Evidence was obtained in the exper:iment described above that
organisms representing a l:imited group of rumen bacteria were mainly
responsible for rumen urease production. A review of the lite rature,
however, failed to reveal any estimate of the proportion of the total
numbers of rumen bacteria represented by urease-producing organisme.
An attempt was therefore made to estimate the proportion of ureolytic
bacteria in the rumen fluid of the urea-fed sheep. Using the method
of most probable numbers, the populations of viable bacteria and of
viable ureolyt.ic bacteria in rumen f'luid w:n-e determined on !our
different occasi0118 in a 10-week period. The results, which are
presented in Table IV, show tha.t the average proportion of rumen
bacteria :round to produce urease was 35%. Microscopie examination
of the cultures used in these determinations showed that non-m.otile,
Gram-negative cocci, arranged singly, in pairs and in short cha.ins,
were invariably present in predominating numbers in cultures with
- 4$-
Table IV. Total numbers of viable bacteria. and numbers of viable ureolytic bacteria in rumen fluid.
Test No.
1
2
3
4
Average
Total Viable Count
(MPN per ml.)
1.7 x 108
5.4 x 107
2.2 x 107 + 7 lE 3.6 - 1.9 x 10
Viable Ureolytic Count Ureolytic count as
(llPN per ml.) % of Total Count
3.5 x 107
3.5 x 107
2.2 x 107 + 63' 6.4 - 1.5 x 10
JI!.Mean of duplicata determinations on one set of serial dilutions
- 46-
urease activity. These resulte support the conclusion that the high
urease activity of rumen fluid from the urea-fed sheep was due to
the presence of large numbers of bacteria of relatively few types.
C. Attempts at the Isolation and Characterization of Urease-producing
Bacteria from Rumen Fluid.
Atte.m.pts were made to isolate in pure culture organisme
representing the predominating urease-producing flora in tœ rumen
fluid of the urea-fed animal, sheep 2.
a) Isolation of bacteria from cultures in buffered reinforced
clostridial medium
First attempts at isolations of ureolytic rumen bacteria
were made using the cultures in buffered :reinforced clostridial broth
employed for the enumeration of these organisme. The cultures were
streaked on buffe red reinforced clostridial agar. Of a total of Z7
colonies picked at random fran tbese plates, 13 consisted of organisme
which produced an alkaline reaction on urea agar within 12 hr. When
t.hese isol.at.es were transf'erred to tryptone gl.ucose 0.2% yeast e.xtract
brotht two types of growth developed. In tœ case of one, the bacter
ia, which were found to be short, Gram-negative rods, produced a
heavy mucoid pellicle and a light turbidity. In tm other, a unifozm
be avy turbidity was produced by smal1, Gram-negative cocci which re
sembled in morphology and arrangement the organisme sean in the BRCB
cultures. One organism of each type was selected at randan,
- 47-
characterized ani identitied. 'l'hase organisms were re.ferred to
respectively as isolates 1 and 2.
i) Isolate 1
This organiam, wt4ch was isolated from a rumen .fluid dilution
of lJ109 , was identitied as Pseudom.onas aeruginosa on the basis o.f the
.follow:i.ng characters.
Cella rod-shaped, 0.8 by 1.5 to 3.0 microns, arranged singly and in
pairs. Motile at 37°C, with one polar .flagellum, non-motile at
Anunonium. sulphate not used as sole source of nitrogen.
Methylene blue reduced.
Coagulase not produced.
Serum not dige sted.
Blood not haemolyzad.
Facultatively anaerobie.
Tha organism essent:ially satisfied the oonditions suggested
by Gall and Huhtanen (1951) for consideration as a true rumen organism.
It was, for exa.mple, facultati vely anaerobie, it occurred in rumen
fluid in num.bers exceeding 106 per ml. and brought about reactions,
such as the fermentation of soluble carbohydrates, known to occur in
the rumen. The organism. was tberefore considered to be a normal
canponent of the rumen microflora.
Attempts were made to reisolate it from the rumen contents
of sheep 2, eight months a:rter the original isolation. For this
purpose, a prima.ry enricbnent of the staphylococci in a sa.mple of
rumen fluid was made in a brain heart infusion - NaCl medium, and
!ive rumen staphylococci were subsequently isolated. Although none
was identical to isolate 2, all of them were very similar to it and
generally differed only with respect to the range of soluble
- 51 -
carbohydrates which were fermented. This failure to reisolate the
organism, however, was not considered to provide evidence against
tm conclusion that isolate 2 was a true rumen organism. Thus it
was further investigated to assess the possible contribution of the
organism to rumen urease activity.
iii) Urease activi ty of iaolate 2
Isolate 2 was grown on tryptone glucose 0.2% yeast e.x.tract
agar and the cells suspended in ma.leate buffer. The suspension wa.s
shown by a standard plate count to contain 1. 9 x 1011 cella per ml. ,
and its urease activity index found to be 19. Thus, the urease
activity of the organism was very law. The activity produced by an
organism when growing on an artificial medium, however, may not give
a valid indication of the activity it would produce when grown und.er
the environmental conditions prevailing in the rumen (Gibbons and
Doetsch, 1959).
In the following section a medium wbich provided enricbnent
cultures of rumen ureolytic bacteria is described. It consisted of
so~ rumen supernatant liquor supplemented with glucose, phytone and
urea (medium 17, Table V). In view of the hii?P urease activity
expressed by .mi:œd cultures of rumen ureolytic bacteria when grown
in this medium, it was thought probable that it .might support enhanced
urease production by isolate 2. Batches of the medium -were therefore
inoculated with the organism, and after incubation the cella were
harvested and suspended in maleate buffer. The suspension contaiœd
- 52 -
approx:imately 2 x lolO cella per ml. When its urease activity was
determined manometrically, however, it was found to have no œasurable
activity whatsoever.
It was therefore concluded from these experimenta that the
Staphylococcus sp. isolated from the rumen contents of sheep 2,
aJ.though weakly ureolytic when grown on artificial media, did not make
a significant contribution to rumen urease activity ~ !!!2·
b) Isolation of bacteria from cultures in a suppleœnted rumen
supernatant liquor medium.
Owing to the lack of auccess experienced in the attempted
isolation of rumen bacteria with a high specifie urease activity by . the method so far described, a second approach to the problem was
made. This involved the development of a medium which would provide
enrichment cultures of rumen ureolytic bacteria, and pennit subsequent
isolation of organisme fran these cultures.
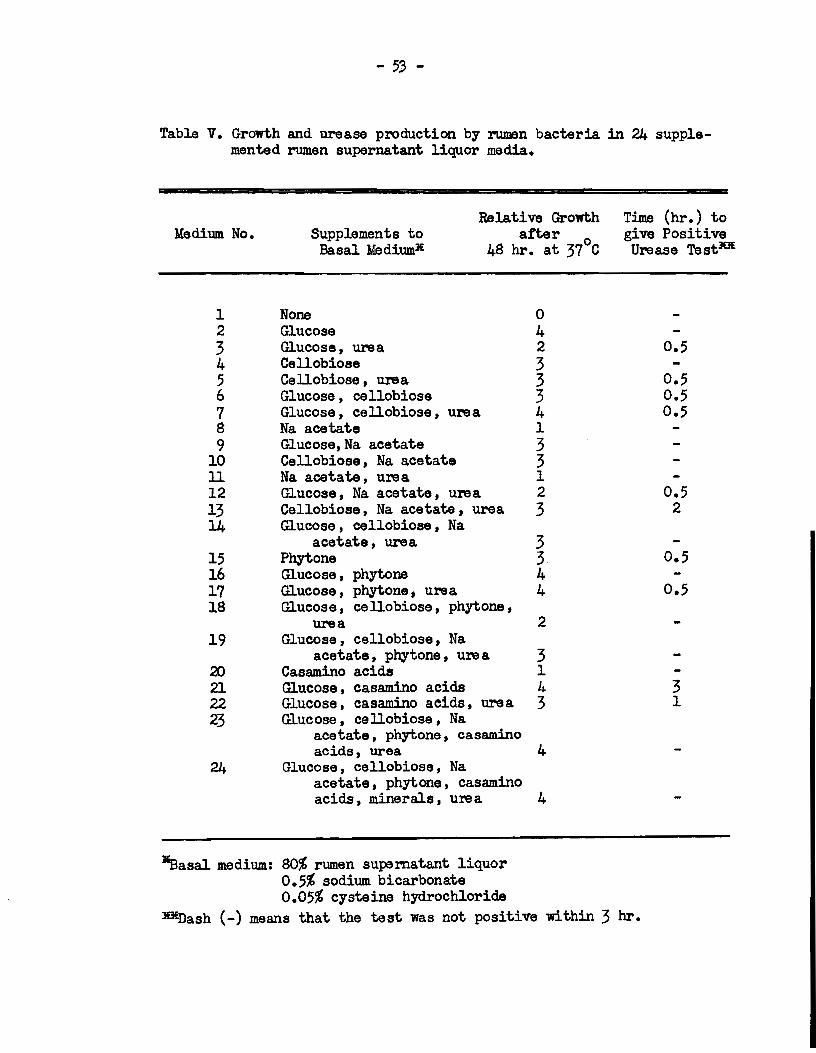
For this purpose, various supplements were added to a basal
medium eontaining 90% rumen supernatant l.iquor, and the rel.ative
abilities of twenty-four such media to support growth and urease
production by rumen bacteria in mixed culture 11ere determined. The
re sul ts are shown in Table V.
Growth developed to varying extents in ail media except
medium 1, which contained no supplements, but the greatest development
of urease activity occurred in those media which were supplemented
-53 -
Table V. Growth and urease production by rumen bacteria in 24 supplemented rumen supernatant liquor media.
Medium No.
1 2 3 4 5 6 7 8 9
10 ll 12 13 14
15 16 17 18
19
20 21 22 23
24
Supplements to Basal ll.ediumlE
Relative Growth a:tter
0 48 hr. at 37 C
None 0 Glucose 4 Glucose, ure a 2 Cellobiose 3 Cellobiose , ure a 3 Glucose, cellobiose 3 Glucose, cellobiose, ure a 4 Na acetate 1 Glucose,Na acetate 3 Cellobiose, Na acetate 3 Na acetate, ure a 1 Glucose, Na acetate, ure a 2 Cellobiose, Na acetate , urea 3 Glucose , cellobiose , Na
acetate, ure a 3 Phytone 3 Glucose, phytone 4 Glucose, phytone, ure a 4 Glucose, cellobiose, phytone,
ure a 2 Glucose , cellobiose, Na
acetate, phytone, ure a 3 Casam.ino acids 1 Glucose , casamino acids 4 Glucose, casamino acids, ure a 3 Glucose, cellobiose, Na
acetate , phytone, casamino acids, urea 4
Glucose, cellobiose, Na acetate, phytone, casamino acids, .m.inerals, ure a 4
*Ionie strength - o.06M Pre.r:aration: A - Whole rumen nuid
B - Organ181118 in mal.eate butter c- " " " • D- " " n •
E- " " n •
F- " " n •
+Na+ + K+ +Mg++ + ca++
F
- 85 -
differed slightly from animal to animal, the general affects
observed were the same with cella from ali three animals. It was
particularly noted that these effects did not differ significantly
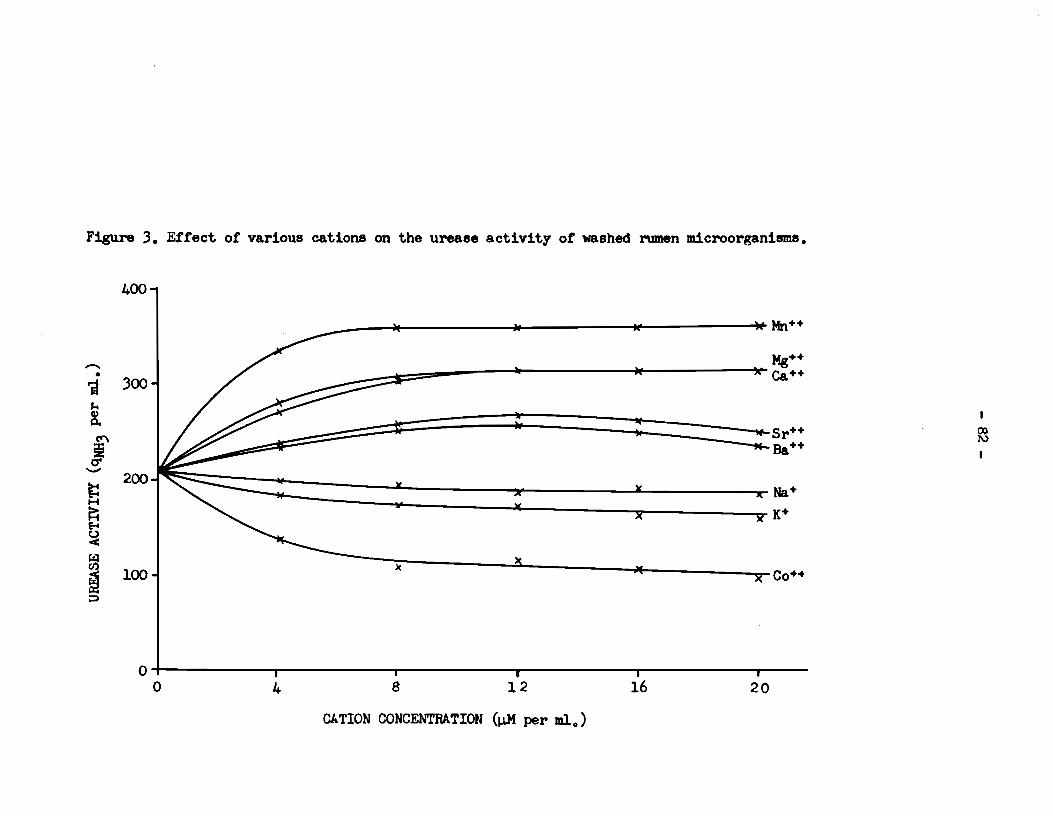
from those found with Mg++ in the case of sheep 2 (Figure 6). It
was therefore concluded that the capacity of Mg++ to stimulate the
activity of rumen urease was not influenced by the presence of urea
in the diet of sheep 2.
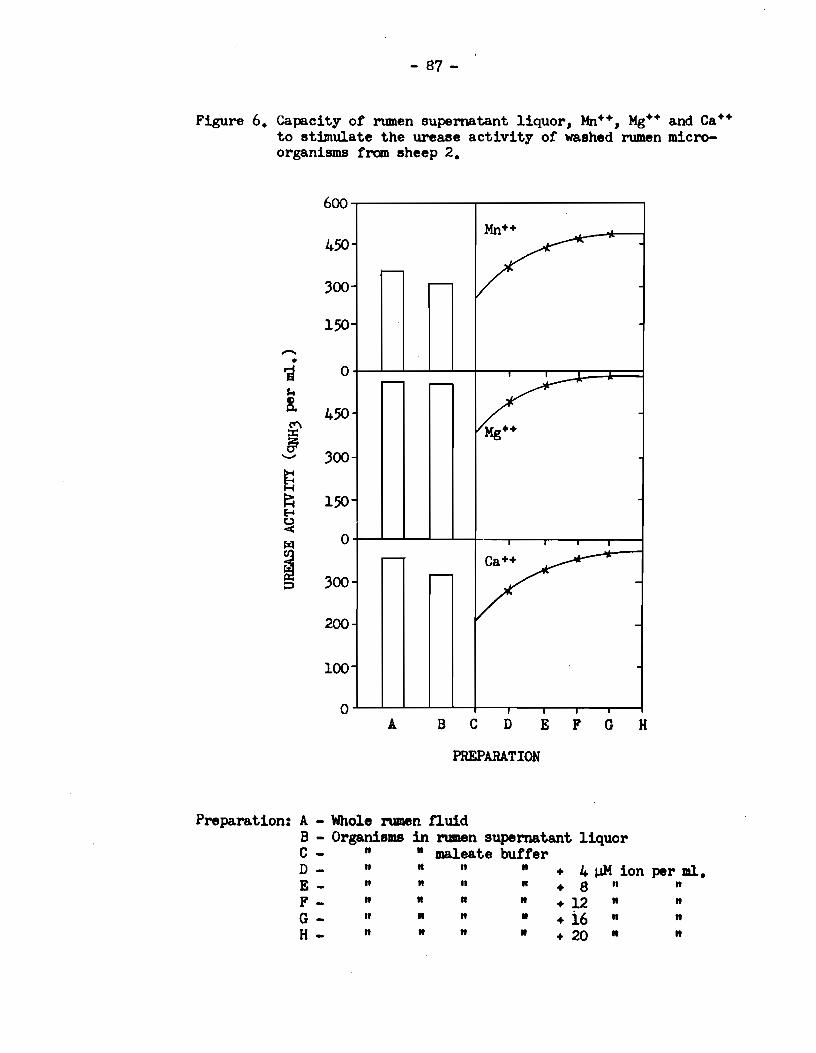
The resulta in Figure 6 show that the effect of Ca++ on
the urease activity of washed cella from sheep 2 was almost identical
to that of Mg++. Between the affects of Mn++ and Mg++, however, an
appreciable quantitative difference was observed. Thus, in the
presence of 1 pM of Mn++ per ml. of buffer, the urease activity of
the washed cella was the same as that gi ven by a suspension of similar
cells in rumen supernatant liquor. In the presence of 12 - 20 JiM of
Mn++ per ml. the activity was raised to a level 56% above that given
by the cell suspension in rumen supernatant liquor and 41% above that
of whole rumen fluid. Thus it was concluded that di valent ions,
probably Mn++, Mg++ and Ca++, were the factors present in rumen super
na tant liquor which were responsible for stimulation of the urease
activity of intact rumen bacteria.
It was not possible to draw a conclusion regarding the way
in whichureas.e stimulation was brought about by these ions, however,
Since the experimenta had been carried out using intact rumen micro
organisns in which the enzyme was entirely intracellular, the affects
-86-
Figure 5. Capacity or rumen su}iematant liquor and Mg++ to stimulate the urease aetivity of washed rumen mieroor8&nisms tram two sheep and one steer.
aoo r--
600 . Sheep 35 r--
400 ~
200
-• 'i1 0 ,..
Sheep 37 r--
!. 600 ('t'\
r--~ . j - 400
~ ~ 200 E-t
~
Steer 2
P:l 0
~ 600 :::>
400 r--
r--200
0 • A B C D E F G H
PREPARATION
Preparation: A - Whole rumen tl.uid B - Organisms in rumen supernatant liquor c - " " " ma1eate butter D - " n " " n + 4 ~ Mg++ per ml. E- n tt n n " + 8 u " F _ n n tt n 11 + 12 n n G _ n n n tt tt + 16 11 tt
H- tt 11 n n n +20 n n
- 87-
Figure 6. Capacity or rumen supematant liquor, Mn++, Mg++ and ca++ to stimulate the urease activity or washed rumen microorganisme from sheep 2.
600
450
300
150
-• i 0 $.e
!. 450 ~ g;
300 -E ~ 150 f-4 ~ !';il 0
1 300 p
200
100
0 A B c D E F G H
PREPARATION
Preparation: A - Who1e rumen fiuid B - OrganisiiiB in rumen supematant 11quor C - " • maleate butter D - " " 11
• + 4 J,J.M ion per ml. E- n n 11 11 + 8 11 n
F- n " n • +12 • " G - n • " 11 + i6 " " H- " 11
" • +20 " "
- 88 -
observed could have been due either to the influence of the ions on
the permeability of the cells to urea or to an absolute effect of
the ions on the intraeellular enzyme. In order to de.monstrate which
of these affects was in fact involved, a series of experimenta was
carried out using as sources of urease preparations in which the
permeability properties of the intact cells were respectively modi
fied and eliminated, nam.ely, by preparing an acetone-dried powder
and a cell-free extract of the mixed flora of rumen microorganisme.
b) Ex:pe riments wi th an acetone -dried powder of rumen micro
organisns
An aeetone-dried powder was prepa.red from washed rumen
microorganisns and determinations were made of the affects on its
urease activity of suspending the powder in va.rioua media. The
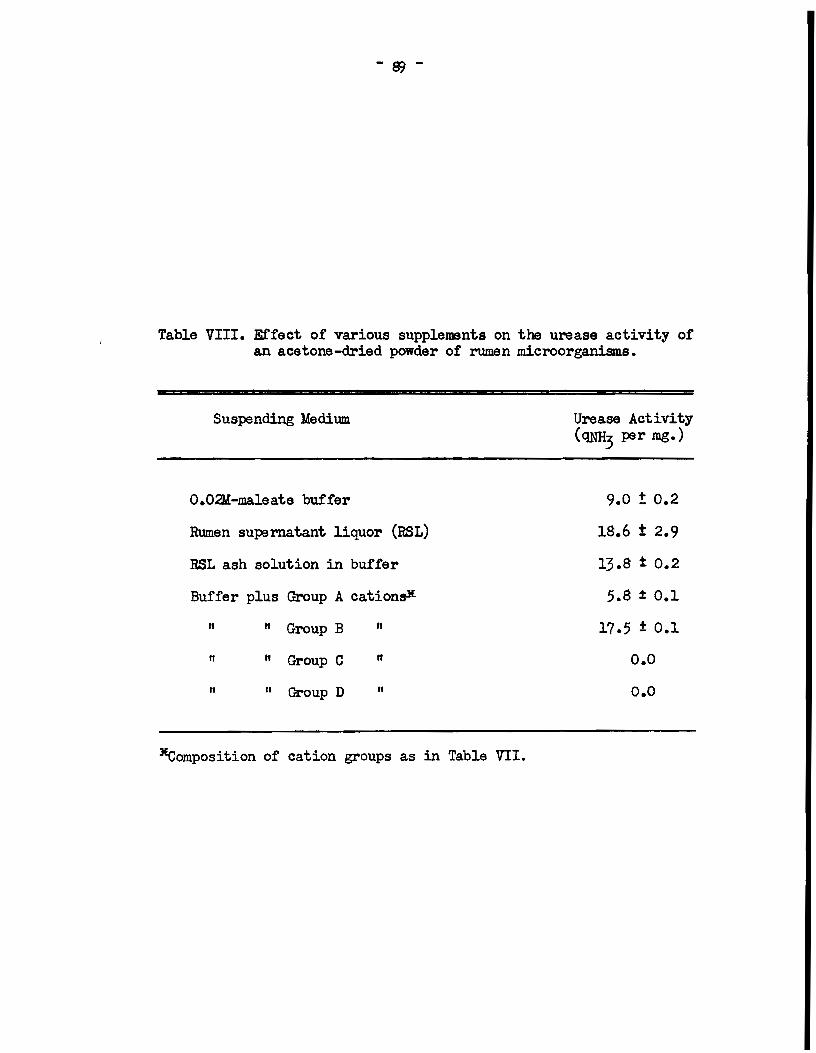
resulta presented in Table VIII show that tm urease activity of the
powder was influenced in a mannar similar to that of the intact cella.
Thus, when the powder was suspended in rumen supernatant liquor or in
a solution in buffer of the ash of rumen supernatant liquor the
urease activity of the powder was appreciably greater than 'When it
was suspendad in buffer alone, indicating that urease activity was
enhanced by an :i,norganic factor present in rumen supernatant liquor.
The combinations of cations previously deseribed were te sted .for
their affects on the enzyme. It is evident from the resulta in
Table VIII that the group of alkaline earth metal ions again enhanced
urease aetivity. It was concluded there.fore that the ureaae aotivity
- fJJ -
Table VIII. Etfact of various supplements on the urease activity of an acetone-dried powder of rumen .microorganism.s.
Suspending Medium
o.02M-maleata buffer
Rumen supernatant liquor (RSL)
RSL ash solution in buffer
Buffer plus Group A cationsx
Il " Group B tl
11 " Group C
tt 11 Group D Il
lEcomposition of cation groups as in Table VII.
Ureasa Activity ( <lliH3 per mg.)
9.0 ! 0.2
18.6 t 2.9
13.8 t 0.2
5.8 :!: 0.1
17.5 :t 0.1
o.o
o.o
- 90-
of an acetone-dried preparation of rumen microorganisms wa.s stimul
ated by divalent cations.
c) Ex:periment with a cell-free extract of rumen microorganism.s
Since studies have shown that treating cells with acetone
modifies their permeabllity properties (Gunsalus, 1955), the affect
of inorganic ions on the urease activity of whole cells would appear
not to be an affect on membrane permeability. The re is no information,
however, on the extent to which acetone treatment modifies the pe:nnea
bllity pro}:erties of rumen microorganisme. To establish beyond doubt
whether the e:f'fects of' the ions on the 11\hole cella were due to affects
on the urease enzyme or enzymes, cell-free extracts of the organisms
were prepared. This was done by subjecting a suspension of washed
rumen microorganisme in maleate buffer to sonic oscillation. The
preparation was centrifuged free of whole cells and debris. The
affects of rumen supernatant liquor and various concentrations of
Mg++ on the urease activity of the extract 118re then determined. The
resulta, which are presented in Figure 7, show that the enzyme
activity was strongly stimulated by both rumen supernatant liquor and
Mg++. The resulta obtained with this cell-free extract of rumen
microorganisme therefore resembled closely those obtained with whole
cella. It was concluded that divalent ions exerted an absolute
stimulating affect upon the urease enzyme and that the affects
observed with the whole cells were not due primarily to the influence
of the ions on the permeability of the cell membrane.
- 91 -
Figure 7. Capacitr of nunen su~rnatant 1iquor and Mg++ to stimula te the urease activitr of a cell-free extract of washed rumen microorganisme.
-• Et ... 8. 200
r;: §t -~ ~ f-4 ~ 100
1 :::::>
A B c D E F G H
DIWTION MEDD.JM
Dilution medium: A - Rumen supernatant 11quor B - Maleate butter c- " n + 2 IJ.M Mg++ per ml. D - lt lt + 4 n tt
B- " .. + 6 lt " F- tt n + 8 " n
G- n If + 10 n " H- n n + 12 Il n
- 92 -
B. Effect of Divalent Ions on the Activity of Jack-bean Urease.
In order to determine the affects of divalent ions on the
activity of jack-bean urease a solution of the enzyme in maleate
buffer was prepared and its activity in the presence of five ions
at concentrations of 12 uM per ml. of solution was measured. The
results are shown in Figure S. It is clear from these results that
the affects of the ions on jack-bean urease ware very different from
their affects on rumen urease. Thus, whereas the five ions ail
st.imulated rumen urease to varying extents, they all inhibited jack
bean urease, the sequence of inhibition being Ca++~sr++~;7Mg++ ~
Mn++/Ba++.
C. Localization of Urease Activity in Rumen Microbial Cells.
The results so far reported indicate that the stimulation
of the urease activity of intact rumen microorganisms by inorganic
ions was due prim.arily to the affects of the ions on the urease enzyme.
This suggested either that the urease enzyme was located on the cell
surface o:f these organisns or that the cell membrane of the intact
cells presented little or no obstacle to the passage of the ions into
the cell. In an effort to distinguish between the se two possibilities,
the following experimenta were perfonned.
When inorganic ions were added to suspensions of washed
rumen microorganisms it was observed that there was a t.ime lag before
the ions exerted their stimulating or inhibiting affects. This is
- 9:3 -
Figure s. Effect of diva1ent cations on the activity of jack-bean urease.
1200·
-• Et -.. !. "" 800 . J -E i:i E-4 c <
1 400 .
:::>
0._--~----~--~----._--~----~---------
A B c D B F
SUSPF.XSION Ml!DllJM
Suspension medium: A - Maleate butter B - " " + 12 ~ Ba++ C _ 1t 11 + n Mn++
D - " " + " Mg++ E - " " + " sr++ F - " " + " ca++
per ml. " lt .. "
- 94 -
shawn in Figure 9 with various ions capable of stimulating urease
activity. The shortest lag occurred with Mn++, the ion producing
the greatest stimulation. The longest time lag occurred with sr++.
Once the stimulation of enzyme activity had begun, the reaction
proceeded at a constant rate which varied wi th the stimula ting ion
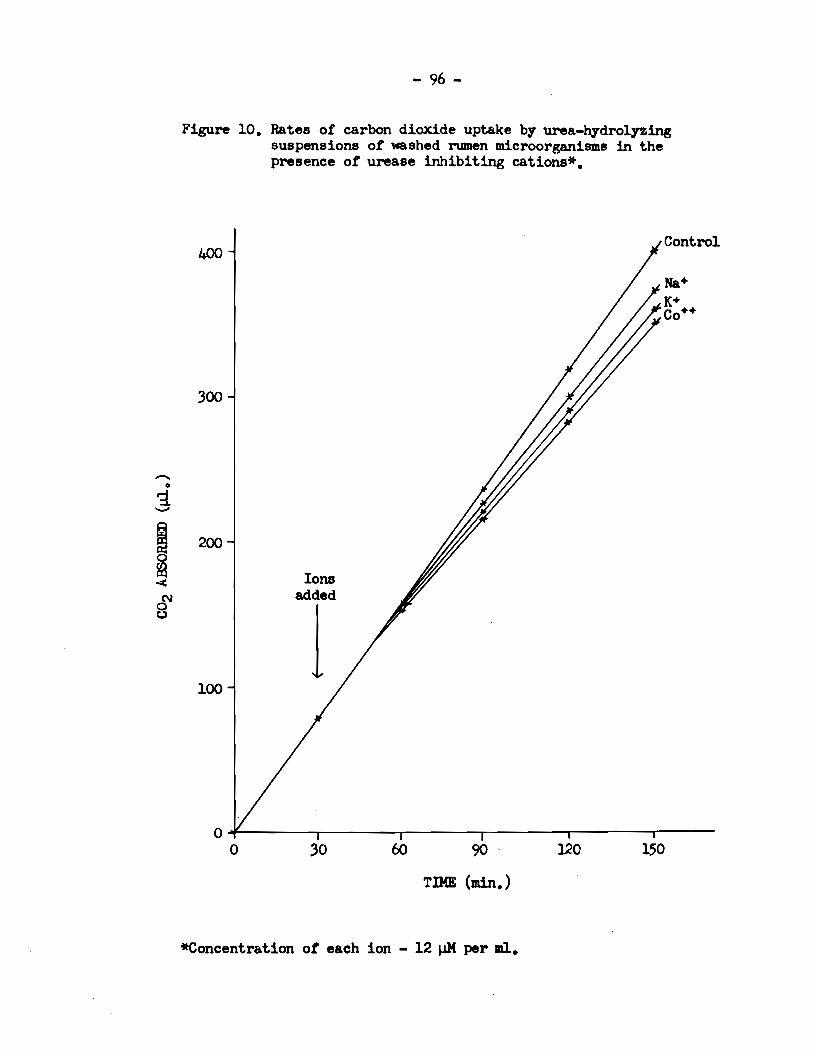
tested. Similarzy, inhibiting ions showed a time lag bef ore inhib
ition could be observed, Figure 10. The time lags observed before
the initiation of stimulation or inhibition could beat be explained
if one assumed that the urease enzyme was intracellular and that the
different ions took varying lengths of time to cross the membrane and
reach a concentration at which they could e:xert an effect in tbs cells.
If' this were so, one could conclude that the rate of penetration of
the urease stimulating ion decreased in the order of increase in the
lag period, namezy, Mn++ ?Mg++ :;::::=--ca++ 7 Ba.++> sr++, and tho se of
the inhibiting ions in the ord.er Co++/ K+ >Na+.
If' tm time lags observed were indeed due to barriere to
the read.y penetration of the ions to tm site of an intracellular
enzy.m.e, the tim.e l.ags shoul.d be modi.fi.ed by factors which woul.d be
expected to modify the permeability of the membranes. For this
reason the time talœn by various ions to initiate stimulating or
inhibiting affects on rumen urease activity were determined using an
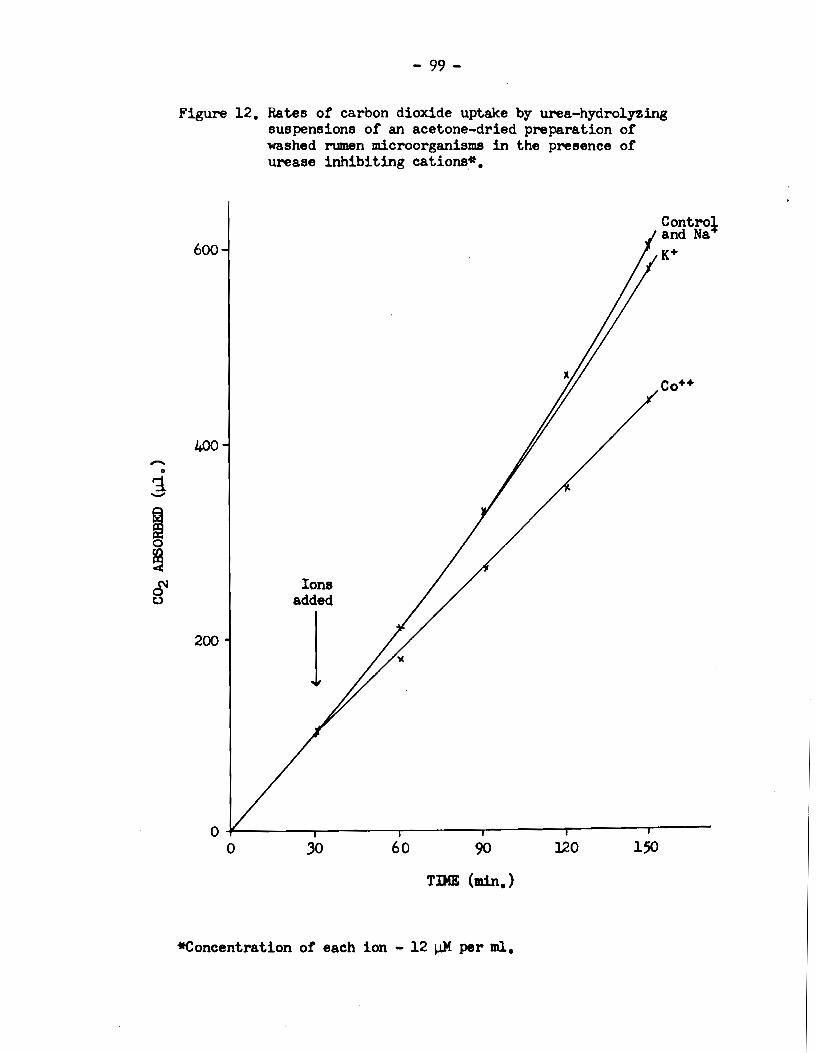
acetone powd.er of the rumen microbial cells. The resulta in Figures
11 and 12 show that even with the acetone powder there was a lag
before the effects of the stimulating and inhibiting ions expressed
themselves. This suggests that if the time lags were due to
-• '1 -1 < ON 0
- 95 -
Figure 9. Rates ot carbon dioxide upt,ake by urea-hydrolyzing suspensions ot washed rumen m.icroorganisms in the presence ot urease stimula ting cations*.
300
200
Ions ad.ded
100
l
Mn++
Mg++
ca++
:sa++ sr++
Control
0~----~------,-----~,-----.-------~---0 30 60 90
TIME (min.)
*Concentration ot each ion - 12 IJ.M per al.
120 150
-• 3. -1 fe < N
0 0
- 96 -
Figure 10. Ra tes of carbon dioxide uptake by urea-hydrolyzing suspensions of washed rumen microorganisme in the presence of urease inhibiting cations*.
variations in the rates of penetration of the ions the acetone
treated cella still reta.ined soma of the permeability properties
of the whole cella. The relative lengths of the time lags with
the various ions showed differences from those obta.ined with whole
cella, however, suggesting tha.t the permeability properties of the
acetone-treated cells bad in fact been modified. Cell penetration
by J.fn++, for exa.mple, was slower with the acetone-treated cells tha.n
with the whole cells, whereas the absence of a time la.g in the case
of Co++ suggested that the acetone-treated cella were freely perm
eable to this ion. The order of increasing time lag for the stimul
ating ions using the acetone powder wa.s Mg++ = Ca++c:::::..Mn++L. sr++<
Ba+•. That of tœ inhibiting ions was co••.c:::::.K+; Na+ exerted no
apparent effect on urease activity within 2 br. There were aga.in
differences between the affects of the various ions after the initial
lag period. The sequence of these affects for the stimulating ions,
in decreasing order of magnitude, was :Mg++/Ca++,?Mn•+:::::>sr+•:;::::..Ba++,
which is similar to the order in which they appeared to penetrate the
acetone-treated eells. It was different, however, from the sequence
previously found for the absolute affects of these ions on the urease
activity of whole cells.
To establish if the time lags observed were in fact due to
permeability affects, the action of tœ ions on a cell-free extract
of rumen microorganisms was studied. The resulta obtained, Figure 13,
show tha.t Mn,.,.., :Mg++ and Ca,.,.. all stimula.ted urease activity immediately
-• '1 -1 83 « 8N
- 98-
Figure 11. Rates of carbon dioxide uptake by urea-hydro1yzing suspensions of an acetone-dried preparation of w.ashed rumen microorganisms in the presence of urease stimulating cations*o
Figure 12. Rates ot carbon dioxide upta.ke by urea-hydro1yzing suspensions ot an acetone-dried preparation ot washed rumen microorganisms in the presence ot urease inhibiting cations*.
Figure 13. Rates of carbon dioxide uptake by a urea-hydrolyzing cell-free extract of washed rumen microorganisms in the presence of urease stimulating cations*.
400
Ions added
1 200
60 90
TlME (min.)
*Concentration of each ion - 12 ~ per ml.
120
Mn++ Mg++ ca++
sr++
Ba++
Control
-0
'3. ..._,
1 ~
C\1 0 0
- 101 -
Figure 14. Rates of carbon dioxide uptake by a urea-hydrol)'fiing cell-free extract of washed rumen microorganisme in the presence of urease inhibiting cations*.
500
Control Na+
K+
co•• 375
25
Ions added
12 l
0 30 f:IJ 120
TDΠ(min.)
*Concentration of each ion - 12 ~ per ml.
- 102-
upon addition of the ions to the urea-hydrolyzing extract.sr+-+- and
Ba+-+, on the otber hand, both stim.ulated the enzyme only a:tter a
lag period. In the case of inhibiting ions, Figure 14, Co++ and K+
both inhibited the enzyme immediately upon addition to the enzyme
system, while Nat"was effective only a:tter a 1ag period.
DISCUSSION
The resulta of this study have clearly shown that the rumen
solution contains inorganic components which have the capacity to
stimulate intracellular rumen urease, and evidence has been presented
that the alkaline earth metal ions and Un++ also have this capacity.
Of these ions, Ca"'+ and llgu are known to have important
fun ct ions in ruminant metabolism (Duncan, 1958; Rook and Storry, 1962).
Analyses of rumen liquor from sheep on various diets (Gerton, 1951)
have shown that the rumen solution may contain concentrations of Ca++
and Mg++ equivalent respectively to 2.5 - 5.0 JiM and 4.0 - 5.0 )fM per
ml. The significance and function of Un++ in the rumen, on the other
hand, is but little understood (Barnett and Reid, 1961) and a search
of the literature has failed to reveal a reference to the normal
concentration of soluble Mn++ in the rumen solution. From experimenta
on the requirements of ruminants for dietary Un++, which have generally
been found to be lesa than 50 p.p.m. of the ration (Bentley and Phillips,
1951; Embry, Ga.stler, Radabaugh and OJ.son, J.95S), it may be deduced,
however, that the rumen concentration of soluble Mn++ is considerably
lower than that of either ca++ or :Mg++. Neither sr++ nor Ba++ is
known normally to have any significance in rumen metabolism. From
a consideration of the concentrations of divalent ions found to
stimulate rumen urease in vitro, it seems reasonable to conclude
that Ca++ and Mg++ were probably the components of the rumen solution
which were primarily responsible for its stimulatory affects on the
- 104-
enzyme.
A search of the literature revealed one other reference to
the stimulation of urease by alkaline earth metal ions. This wa.s a
report by Shibata (1958). The original paper was not seetJ,however,
and neitmr the source of the urease used by the author nor the
concentrations of the cations found to be effective is lmown. The
present investigation has shovm. that the affect of these ions on
jack-bean urease at the concentration tested was one of inhibition
rather than stimulation. Jack-bean urease was also found to be
inhibited by Mn++, and this is in agreement with the findings of
Shaw (1954). This au thor has postulated that the mechanism of
urease inhibition by several cations, including Jln"''t', involves
a.ttachment of the ion to the sulphydryl groups of the enzyme, since
ali the cations involved f'orm insoluble sulphides.
Clearly a .mechanism of f'undamentally different character
must be involved in the case of' Jln++ and rumen urease , for here an
appreciable increase in the activity of the enzyme resulted. Wha.t
this mechanism might be, however, cannot a.t present be suggested,
nor can a mechanism by which the other di valent cations may stimu1-
a.te rumen urease be postulated. Rumen urea.se wa.s found to be
inhibited by Co'"+, Na++ and K+ and therefore in this respect resem
bled jack-bean urease which is also inhibited by these ions (Wall and
Laidler, 1953; Shaw, 1954).
The reduction in urease activity which wa.s found in tm
present study to accompany the washing of intact rumen microorganisms
- 105 -
with buffer indicated a reduction in the intracellular concentration
of di valent cations. Furthermore, the suspension of resting intact
cells in solutions of divalent ions resulted in uptake of the ions
by the cells, together with a subsequent enhancement of urease
activity. These observations show, firstzy, that the ions were not
bound to any appreciable extent intracellularly, and secondly, that
the ions were qui te readily diffusible into the cella in the re sting
state. This situation shows some differences from that found to
e.xist in other whole cell systems. Tsuyuki and Maclsod (1951), for
example, round that extracellular Mg+-+ and )ln++, which ions are
essential cofactors for several enzymes involved in glycolysis, did
not affect the rate of glycolysis by resting cell suspensions of
Lactobacillus arabinosus grown in a Mzl++-deficient medium, although
:Mn++ was required for growth by the organism. The authors interpreted
their resulta as indicating that Mn++ was bound intracellularly and
that during washing of the cella the concentration of the ion within
the cella remained sufficiently high so that further addition of
divalent cations to a suspension of the cella bad no sti.mulating
effect on metabolism.
A dependance of divalent cation absorption on metabolic
activity was shown for baker 1s yeast in isotopie studies by Rothstein
and his associates (Rothstein and Hayes, 1956; Rothstein, Hayes,
Jennings and Hooper, 1958; Jennings, Hooper and Rothstein, 1958).
If resting cella of the organism were placed in solutions containing
Mn++, Mg++, ca++ or sr++, a rapid and reversible binding of the ion
at specifie sites on the cell membrane oecurred to a li.mited extent.
- 106-
Beyond this surface binding thare was no absorption of tm ion, nor
did divalent cations leak out of tm cytoplasm, showing that thase
ions could not penetrate tm cell membrane. If phosphate and glucose
11ere added to the suspension, however, in addition to the surface
binding soma di valent ion was irreversibly absorbed by tm ce lis.
The sequence of absorption of divalent cations by metabolizing cells
of the yeast was Mg·f..f· /Mrl'""">Ca++:;::>SrH. In the present study it
was found that Mn,++ penetrated tm resting cella of rumen bacteria
more rapidly than :Mg++, but otherwise the seque nee was similar.
It is concluded from this investigation, therefore, that
rumen ureolytic bacteria differ from soma other biological systems
both with respect to pe:nneability of the cella to cations, and with
respect to the susceptibility of their urease to stimulation by
dival.ent cations. It is of interest to note, when considering tm
per.meability of the cella to cations and the affects of these cations
on urease acti vity, that the time lags which occurred be fore J4n++,
:Mg+"" and Ca+.,. stimulated the urease activity of intact cella, were
attributable entirely to the time taken for these ions to penetrate
the ce lis, whereas in the case of sr++ and Ba++ this was not so.
Even when these latter ions were in apparently free contact with the
enzyme, i.e. , in the case of the cell-free extract of rumen micro
organisme, stimulation did not occur imrnediately. This suggests
that an active complex between ion and enzyme was not imrnediately
formed.
- 107 -
Rumen bacteria, as a.lready mentioned, no:rma.lly exist in a
medium, na.mely the rumen solution, whieh conta.ins signi.ficant concen
trations of soluble Ca++ and :Mg++. These concentrations are contin
ua.lly ma.inta.ined by solubilization of constituants of the anima.l8
diet. The rumen solution also contains relatively large a.mounts of
Na1' and K"'. For example, Garton {1951) found each ion to be present
in tha rumen solution of sheep to the extent of 100 - 200 mg. per
100 nù.. Ail these ions have been shown in the present study to be
absorbed by resting cells of urease-producing rumen organisme and
subsequently to influence the urease activity of the organisme. If
a simila.r absorption occurs in the case of actively metabolizing
cella, it may be postula.ted that tha activity ma.nifested by urea.se
producing organisme in !!!.2 is the resultant of opposing stimulating
and inhibiting affects on the intracellular enzyme brought about by
ions which are present in the rumen solution.
A detailed study of the bacteria predomina.ntly responsible
for rumen urease production must await the isolation in pure culture
of the organisme concemed. This, as indicated in Part I, is a matter
which presents considerable problems. Fresh attempts to isolate such
organisme, however, would appear to be of potential value, for the
present investigation has shown that a study of their physiologica.l
properties may prove to be a matter of considerable interest from.
the viewpoint of comparative biochemistry.
SUMMAR.Y
A study was made of soma factors affecting the activity
of rumen urease, uaing preparations of .mi::xed rumen microorganisme
as a source of enzyme •
When whole cells o:f rumen microorganisms were washed with
maleate buf:fer there was a reduction in the urease activity o:f the
cells. Resuspension o:f the washed cells in rumen supernatant liquor,
however, brought about an enhancement of the ir urease activity. This
e:f:fect was due to the pre sance of an inorganic rumen urease stjmulat
ing factor in rumen superna.tant liquor.
When whole cella were washed in bu:f:fer there was a pro
gressive reduction in the urease activity of the cella as the number
of washings increased. Up to the third washing the activity o:f tœ
cells could be restored to the level prevailing in unwashed cells
by resuspending the washed cells in rumen supernatant liquor. A:fter
the sixth and ninth washings, on the other hand, the urease activity
of the cella could not be fully restored in this way.
Five divalent cations also had the capacity speci:fically
to stimulate the urease activity of washed whole cells of rumen
microorganisme. The degree of urease stimulation brought about by
these ions decreased in the sequence Mn,++~g++7Ca++:;:>sr++/Ba++.
The activity of the cella was reduced in the presence of Na+, K+
- 109-
and Co+t-, and tm enzyme was completely inhibited by Zn++, eu++ and
Fe+++.
Solutions of Mg++ had the capacity to replace rumen super
natant liquor in the restoration of urease aetivity to washed whole
ce ils of run.en microorganisme from a urea-fed sheep and fran two
sheep and a steer not receiving dietary urea. Solutions of Ca++ had
a similar capacity to Mg++ in restoring the activity of washed cells
from the urea-fed animal. The affect of solutions of Mn++, however,
was to raise the activity of washed cells from this animal to a level
approximately 40% greater than that prevailing in unwashed cells.
The urease activity of an acetone-dried powder of washed
rumen microorganisme was stimulated by inorganic factors present in
rumen supematant liquor and also by the group of divalent cations
Mg++, Ca++, sr++ and Ba++.
The urease activity of a cell-free extract of washed
rumen microorganisma was stimulated by rumen supernatant liquor. of
Solutions Mg..,.., could replace rumen supernatant liquor in stimulating
the activi ty of the ext.ract.
It was concluded that di valent ions, probably Un++, Mgt-t-
and Ca++, were the factors present in rumen supernatant liquor which
were responsible for the stimulation of rumen urease. Moreoever, the
stimulating effect of these ions on the urease activity of whole cella
was due primarily to the affects of the ions on the urease enzyme and
not to their effects on permeability of the cells to urea.
- 110-
The activity of jack-bean urease was reduced in the presence
of all divalent ions found to stimulate the activity of rumen urease.
The degree of inhibition of the enzyme brought about by these ions
in decreasing order was Ca""""> Sr"""".>Mg""""?M'n""""?Ba""""· Apparently,
therefore, the re exista a funda.mental difference between rumen urease
and jack-bean urease with respect to their response to divalent cations.
When inorganic ions were added individually to actively urea
hydrolyzing suspensions of washed rumen microorganiams, there were time
lags before the ions exerted their st:imulating or inhibiting effects.
The time lags with urease stimulating ions increased in the order
Mn""+~ :Mg++<: Ca+"< Ba++<:::: Sr"""", and those wi.th the urease inhibiting
ions in the order Co""~ K+<.. Na .......
Time lags were also observed when inorganic ions were added
to urea-hydrolyzing suspensions of an acetone-dried powder of washed
rumen microorganisms. In this case the lags increased in the orders
MgH = Ca++<Mn""+< Sr++.C:::::::. Ba++ and Co++< K+ with the stimula ting
and inhibiting ions respectively.
The re l'lere no time lags, however, be .tore the initiation of
effects on urease activity when Mn+ .. , Mg....,, Ca++, Co++ or K+ "Were
added to a urea-hydrolyzing cell-free extract of washed rumen micro
organisms, although in the case of sr++, Ba++ and Na+ such time lage
were observed.
The tim.e lags observed with the whole cella were explained
- lll-
in tenns of the varying lengths of time taken by different ions to
cross the cell membrane and reach a concentration at which they
could exert an affect on the intracellular urease enzyme. Modifi
cation of the perm.eability properties of the cells by treatment with
acetone also modified the rates of penetration of the ions. The tim.e
lags observed when sr++, Ba++ and Na"' were added to the cell-free
extract suggested that there was a delay in the formation of an
active complex between the ions and the enzyme.
The absolute affects of the different ions upon the urease
enzyme varied with the different preparations of rumen urease studied,
in a mannar generally sim.ilar to that of variations in the cell
penetration rates of the ions.
The differences which exist between rumen ureolytic bacteria
and sana other biological systems, both with respect to the pe:rmeability
of cella to cations and the susceptibility of urease to stimulation
by di valent cations, were discussed with reference to the rumen envir
onment.
BIBLIOGRAPHY
Abou Akkada, A.R., and Howard, B.H.
1960 The biochemistry of rumen protozoa. 3. The carbohydrate metabolism of Entodinium. Biocham. J., 76: 445-451.
Amdson, E.F., Chalmers, M.I., Marshall, S.B.M., and Synge, R.L.M.
1954 Ruminal anmonia formation in relation to the protein requirement of sheep. n. Ruminal amnonia formation with various diets. J. Agr. Sei., 44: 'Z/0-2:73.
Annison, E.F., and Iswis, D.
1959 Metabolism in the Rumen. Metheun andco:-;-London.
1959 Bacterial species of the rumen. Bacteriol. Rev. , 23: 125-153.
Chibnall, A. C.
1939 Protein Metabolism in tœ Plant. Yale Univ. Press, New Haven.
- ll4-
Christensen, W.B.
1946 Urea decomposition as a means of differentiating Proteus and paraco1on cultures from each otœr and from Salmonella and Shigella types. J. Bacteriol., 52: 461-466.
Dixon, M., and Webb, E.C.
1958 Enzy.mes. Longmans, Green and Co., London.
Doetsch, R.N., Robinson, R.Q., and Shaw, J.C.
1952 Techniques employed in cultural investigations of the bacterio1ogy of bovine rumen contents. J. Animal. Sei., li: 536-544 ..
1958 The interpretation of etudies of calcium and phosphorus balance in ruminants. Nutr. Abstr. Rev., 28: 695-715.
El.l.is, W.C., and Ptander, W.H.
1958 In vitro rumen microbial protein and nuc1eic acid metabolism .. J. Animal Sei., 17: ll92.
Embry, L.B., Gastler, G.F., Radabough, D. V., and Olson, O.E.
1958 Manganese requirements of grow:i.ng and fattening catt1e. J. Animal Sei., 17: 1204.
- 115 -
Fasm.an, G.D. , and Niemann, C.
1951 A reinvestigation of the kinetics of the urease-catalyzed hydrolysis of urea. I. The ac ti vit y of urease in the pre sen ce of sodium and potassium phosphate. J. Am. Cham. Soc., 73: 1646-1650.
Gall, L.s., and Huhtanen, C.N.
1951 Criteria for judging a tru.e rumen organisn and a description of five rumen bacteria. J. Dairy Sei., 34: 353-362.
Gall, L.S., Stark, C.N., and Loosli, J.K.
1947 The isolation and preli.minary stud.y of soma physiological characteristics of the predominating flora fran the rumen of cattle ani sheep. J. Dairy Sei., 30: 891-899.
Garton, G.A.
1951 Observations on the distribution of inorganie phosphorus, soluble calcium and soluble magnesium in the stomach of the sheep. J. Exptl. Biol., 28: 358-368.
Gibbons, R.J., and Doetsch, R.N.
1959 Physiological s~udy of an obligately anaerobie ureol~ic bacterium. J. Bacteriol., 77: 417-428.
Gibbons, R.J., and McCarthy, R.D.
1957 Obligately anaerobie urea-hydrolyzing bacteria in the bovine rumen. Maryland Agr. Expt1. Sta., :Mise. Pub1. No. 291: 12-16.
- 116-
Gray, F.V., Pilgrim, A.F., and Weiler, R.A.
1953 Conversion of plant nitrogen to microbial nitrogen in the rumen of the sheep. Nature, 172: 347-348.
Gunsalus, I.e.
1955 Extraction of enzymes from microorganisns. In Colowick, S.P., and Caplan, N.O. Methods .!n, Enzymologz, Vol. I, Academie Press Ine., New York.
Hamilton, T.S., Robinson, W.B., and Johnson, B.C.
1948 Furthar eomparison of the utilization of nitrogen of urea with that of some feed proteins by sheep. J. Animal Sei., 7: 26-34.
Harris, L.E., and Mitchell, H.H.
1941 The value of urea in the synthesis of protein in the pauneh of the ruminant. I. In maintenance. J. Nutr., 22: 167-182.
Hirseh, A., and Grinsted, E.
1954 Methods for the growth and enumeration of anaerobie spore -formsrs from chee se , with observations on the effect of nisîn. J. Dairy Res., 21: 101-110.
Hobson, P.N.
1961 Technique of eounting rumen microorganisns. In lewis, D., Ed. Digestive P}lysiologz and Nutrition of the Ruminant. Butterwortb;,:Lëndon.
Hobson, P.N.
1962 Personal communication.
- 117-
Hoskins, J.K.
1934 Most probable numbers for evaluation of coli-aerogenes tests by fementation tube method. Public Health Rept. (U.S.), 49: 393.
Houpt, T.R.
1959 Utilization of blood urea in ruminants. Am. J. Physiol., 197: 115-120.
Huhtanen, C.N., and Gall, L.s.
1955 Manametric estimation of rumen urease. J. Bacteriol., 69: 102-103.
Hungata, R.E.
1947 Studies on cellulose fer.mentation. III. The culture and isolation of cellulose-decomposing bacteria from the rumen of cat tle. J. Bacteriol., 53: 631-645.
195S The participation of phosphata in the for.mation of a carrier for the transport of magnesium and manganese ions into yeast cells. J. Gan. Physiol., 41: 1019-1026.
- 118-
Johanson, R., Moir, R.J., and Underwood, E.J.
1949 Sulphur-containing amino-acids in the rumen bacteria of sheep. Nature, 163: 101.
Johns, A.T.
1951 The mechanisms of propionic acid formation by Veil1onella gazogenes. J. Gen. Microbio1., 5: 326-336.
Jurtshuk, P., Doetsch, R.N., and Shaw, J.c.
1958 Anaerobie purine dissimilation by washed suspensions of bovine rumen bacteria. J. Dairy Sei., 41: 190-202.
Kistiakowsky, G.B., and Shaw, W.H.R.
1953 Ureo1ytic activity of urease at pH S.9. J. Am. Chem. Soc., 75: 2751-2754.
Laidler, K.J., and Hoare, J.P.
1949 The mo1ecular kinetics of the urea-urease system.. I. The kinetic 1aws. J. Am. Cham. Soc., 71: 2699-2702.
Larson, A.D., and Kallio, R.E.
1954 Purification and properties of bacterial urease. J. Bacterio1., 6S: 67~71.
lewis, D.
1951 The metabolism of nitrate and nitrite in the sheep. 2. Hydrogen donators in nitrate reduction by rumen microorganisns ia vitro. Biochem. J., 49: 149-153.
- 119-
Lewis, D.
1960 Ammonia toxieity in the ruminant. J. Agr. Sei., 55: lll-117.
Lewis, D., Hill, K.J., and Annison, E.F.
1957 Studies on the portal blood of sheep. 1. AbsorPtion of ammonia from the rumen of sheep. Bioehem. J., 66: 587-592.
lewis, D., and MeDonald, I.W.
1958 The inter-relationships of individual proteins and earbohydrates during fermentation in t:œ rumen of the sheep. I. The fermentation of casein in t:œ presence of starch or other earbohydrate materials. J. Agr. Sei., 51: 108-ll8.
Lister, A.J.
1956 The kineties of urease aetivity in Corypebaeterium renale. J. Gen. Microbiol., 14: 478-484.
Loosli, J. K., and Harris, L.E.
1945 Methionine inereases tœ value of urea for lambs. J. Animal Sei., 4: 435-437.
Loosli, J.K., Williams, H.H., Thomas, W.E. Ferris, F .H., and Maynard, L.A.
1949 Synthesis of amino aeids in the rumen. Science, llO: 144-145.
Lugg, J.W.H.
1949 Plant proteins. Advan. Protein Chem., 5: 229-304.
- 120-
MacKay, E.S.M., and Oxford, A.E.
1954 Soma facultatively anaerobie Gram-negative rods from the runen of the calf and t:œ sheep. J. Gen. Microbio1., 11: 472-476.
Mann, s.o., Masson, F.M., and Oxford, A.E.,
1954 Facultative anaerobie bacteria from the sheep's rumen. J. Gen. Microbiol., 10: 142-149.
Mam, s.o., and Oxford, A.E.
1955 Relationships between viable saccharolytic bacteria in rumen and abomasum of the young calf and kid. J. Gen. Microbio1., 12: 140-146.
Masson, F.M., and Oxford, A.E.
1951 The action of the ciliates of the sheep's rumen upon various water-solub1e carbohydrates, including polysaccharides. J •• Gen. Microbiol., 5: 664-672.
Mateer, J.G., and Marshall, E.K.
1916 The urease content of certain beans, with special reference to the jack-bean. J. Biol. Cham., 25: 297-305.
McDonald, I.w. 1948 The absorption of ammonia from the
rumen of the sheep. Biochem. J., 42: 584-587.
McNaught, M.L., Owen, E.C., Henry, K.M., and Kon, S.K.
1954 The uti1ization of non-protein nitrogen in the bovine rumen. S. The nutritive value of the proteins of preparations of dried rumen bacteria, rumen protozoa and brewer's yeast for rats. Biochem. J., 56: 151-156.
1950 The digestion of rumen microorganisms by the host animals. J. Dair,y Sei., 33: 565-572.
Prescott, s.e., Winslow, C.-E.A., and McCrad)!.l M,H~,
1946 Water Bacteriology. 6th ed. Wiley and Sons,
New York.
- 122 -
Qua.stel, J.H.
1933 The action of polyhydric phenols on urease; the influence of thiol compounds. Biochem. J., 27: ll16-1122.
Rook, J.A.F., and Storry, J.E.
1962 Magnesium in tm nutrition of farm. anim.a.ls. Nutr. Abstr. Rev., 32: 1055-1077.
Rothstein, A., and Hayes, A.
1956 The relationship of the cell surface to metabolism. XIII. The cation-binding properties of the yeast cell surface. Arch. Biochem. Biophys., 63: 87-99
Rothstein, A., Hayes, A., Jennings, D.H., and Hooper, D.C.
1958 The active transport of magnesium and manganese ions into the yeast cell. J. Gen. Physiol., 41: 585-594.
Rustigian, R., and Stuart, C.A.
1941 Decomposition of urea by Proteus. Proc. Soc. Exptl. Biol. Med., 47: 108-112.
Rys, R. , Gor ski, L. , and Styczynski, H.
1956 Roczniki Nauk Rolniczych: Ser. B, 70: 573-575. Cited in Nutr. Abstr. Rev., 70: 573(1958).
Shaw, W.H.R.
1954 The inhibition of urease by various metal ions. J. Am. Chem. Soc., 76: 2160-2163.
- 123-
Shaw, W.H.R.
1961 Cation toxicity and the stability of the transition~tal complexes. Nature, 192: 754-755.
Shaw, W.H.R., and Raval, D.N.
1961 The inhibition of urease by metal ions at pH 8.9. J. Am. Chem. Soc. , 83: 3184-3187.
The resulta show that the rumen ammonia concentration fluctuated
considerably throughout the experimental period. Wlthin the first
i hr. tba runen ammonia concentration rose to a level 25% greater
than its initial level, showing that the urea added to the rumen was
rapidly hydrolyzect · to ammonia. Thereafter, despite the fluctuations,
there was a trend towards a lower concentration during the following
7 - 8 hr., and then towards an increased concentration again during
the next 4 hr. period, by the end of which the concentration had
regained its initial levaL.
c) Total nitrogen content of rumen supernatant liquor and
its enricèment with Nl5.
The total nitrogen content of the rumen supernatant liquor
during the experimental period is shown in Appendix Figure 3. The
resulta represent the concentration of soluble nitrogen found in the
rumen fluid at each sampling time. The figure also shows the levels
of enrichment of this nitrogen fraction with Nl5. The degree of
enrichment is e:xpressed as atoms % excess N15 ovar the normal abundance
of 0.366 atom.s %. The results show that despite the fluctuations in
ammonia concentration of the rumen fluid which occurred during the
experimental period, and which were noted above, the soluble nitrogen
concentration remained relatively constant. There was a rapid decrease
following the 7th hour, when 1000 ml. of water Viere added to the rumen,
but apart from this the decrease in concentration during tha first 8 hr.
l'ms relatively small. It was followed by a slight increase in
-•
~~ 40
:Z:J.. ~8. g . e-.r -
-Il) Il)
~ 'bit
J ~ 2.0
U'\
~ 1.0
Appendix Figure 3. Changes in concentration and Nl5_enrichment of the total soluble nitrogen camponent ot rumen tluid after addition ot Nl5-labelled urea to the rumen.
concentration during the final 4 hrs. of the experimental period.
The level of enrichment of this nitrogen fraction rose very
sharply to 6 a toms % excess Nl5 within the first ! hr. after adminis
tration of the labelled urea. Thereafter tœ enrichm.ent decreased,
at first sharply and then more slawly. The initial sharp rise may be
attributed to the addition of a relatively large amount of Nl5-urea
to the rumen and its subsequent solution in the rumen fluid. The
rapid fall in enrichment indicates either tha.t the N15 -a.mmonia formed
by hydrolysis of the urea wa.s selecti vely absorbed fran the rumen
solution or that th9 added Nl5 wa.s rapidly diluted by unlabelled
nitrogen, which might have entered the rumen directly or indirectly
from the bloodstream. The former is probably the more likely poss
ibility, as evidencErl by the previously noted fall in tm ammonia
concentration in the rumen solution, which coincided with the decrease
in enrichment. Between the 5th and ?th hours of the experimental
period there was an increase in the enrichment of soluble nitrogen
with r5. This was probably caused by a sudden increase in tm amount
of Nl5 entering the rumen by way of the sali va or the rumen wall, since
there was a small increase in the total nitrogen content of the rumen
supernatant liquor during the sa.rœ period. A relatively steady state
in the leval of enrichment of soluble rumen nitrogen with Nl5 appeared
to be reached at the 7th hour. This indicates that Nl5 entering the
rumen was then balanced by that being removed.
- 149-
d) Total nitrogen content of the microbial fraction
of rumen fluid and its enricbnent with r5 Appendix Figure 4 shows tha total amount of microbial nitrogen
present in the rumen fluid samples collected during the experimental
period. It also shows the levels of enrichrnent of microbial nitrogen
with Nl5. Since the samples of the microbial fraction upon which
the determinations were made had been washed with saline during prep
aration, it was thought unlikely that there was a significant contamination
of the fractions with soluble nitrogen from the rumen fluid, and it was
therefore concluded that the nitrogen found in the samplas represented
incorporated nitrogen. However, the microbial fraction was frozen for
storage and later thawed and this may have resulted in sane degree of
cell lysis. Seme microbial nitrogen may therefore have been lost
bef ore the analyses were made. Apart from de cre ases whieh were brought
about by the addition of water to the rumen, the leval of microbial
nitrogen in the rumen fluid rose gradually throughout the experimental
period. This could be attributed either to the occurrence of micro
bial synthesis or to the absorption of water from tha rumen.
The lsv'els of enrichm.ent of microbial nitrogen with N15
indieate that a significant degree of incorporation of Nl5 into the
microbial nitrogen occurred, at least during tha first 8 hr. after
administration of the urea,. and thus they support the conclusion that
the utilization of ure a by the ruminant animal involves, at least in
part, the incorporation of urea-nitrogen into rumen microbial nitrogen.
-• i:i'if ~§ ~~ :z:s. ·~
~ . ~r ._,
-ID .... .,
= 'tslt.
8 +) cd -
1.1'\
~
Appendix Figure 4. Changes in concentration and Nl5-enrichment of the total microbial nitrogen eom.ponent of rumen fluid after addition of Nl5-l.abelled urea to the rumen.
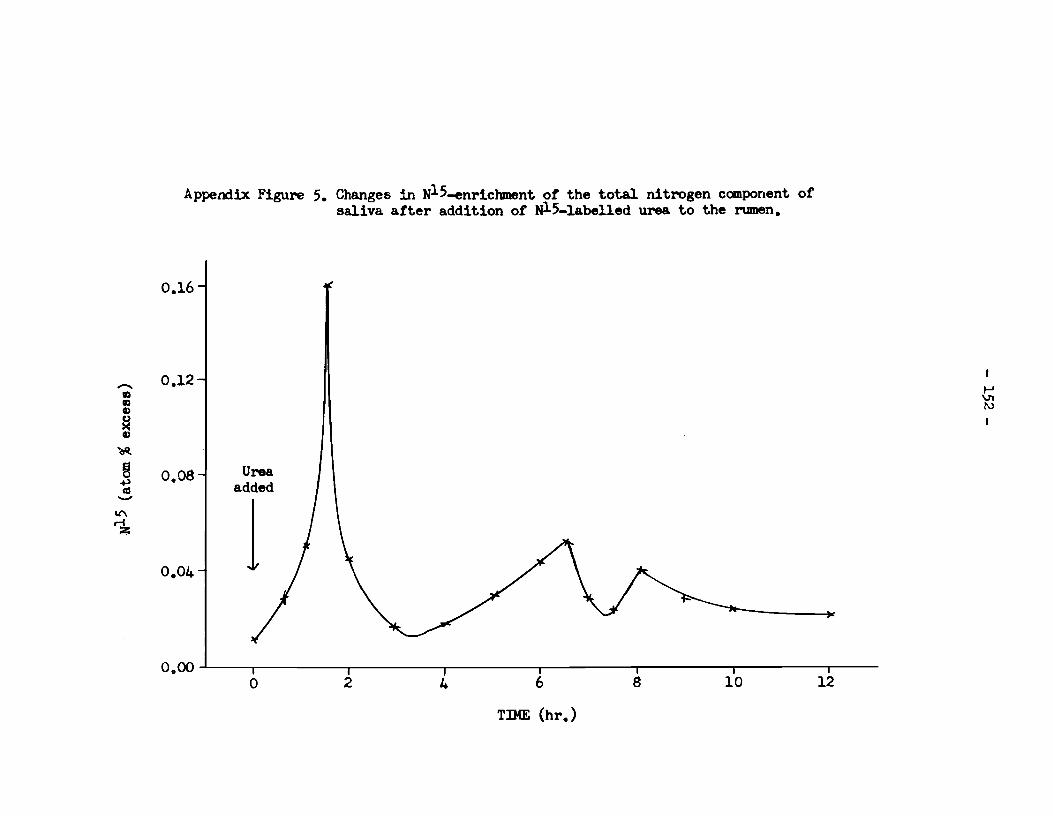
of enrichnent was not attained for a furthar 1 hr. It was thus
reached à hr. after the peak level of enrichment found in t:œ soluble
nitrogen fraction of rumen fiuid. Following the peak level tœre
was a rapid reduction in the enrichment of saliva-nitrogen with Nl5,
but a significant excess was present throughout t:œ experim~ntal
period. This indicates that N15 was continually entering tœ rumen
in saliva and tharefore, by implication, also entering and leaving
the rumen by diffusion through the rumen wall; tha resulta therefore
confirm that seme nitrogen absorbed from the rumen is recycled to that
organ xia the saliva.
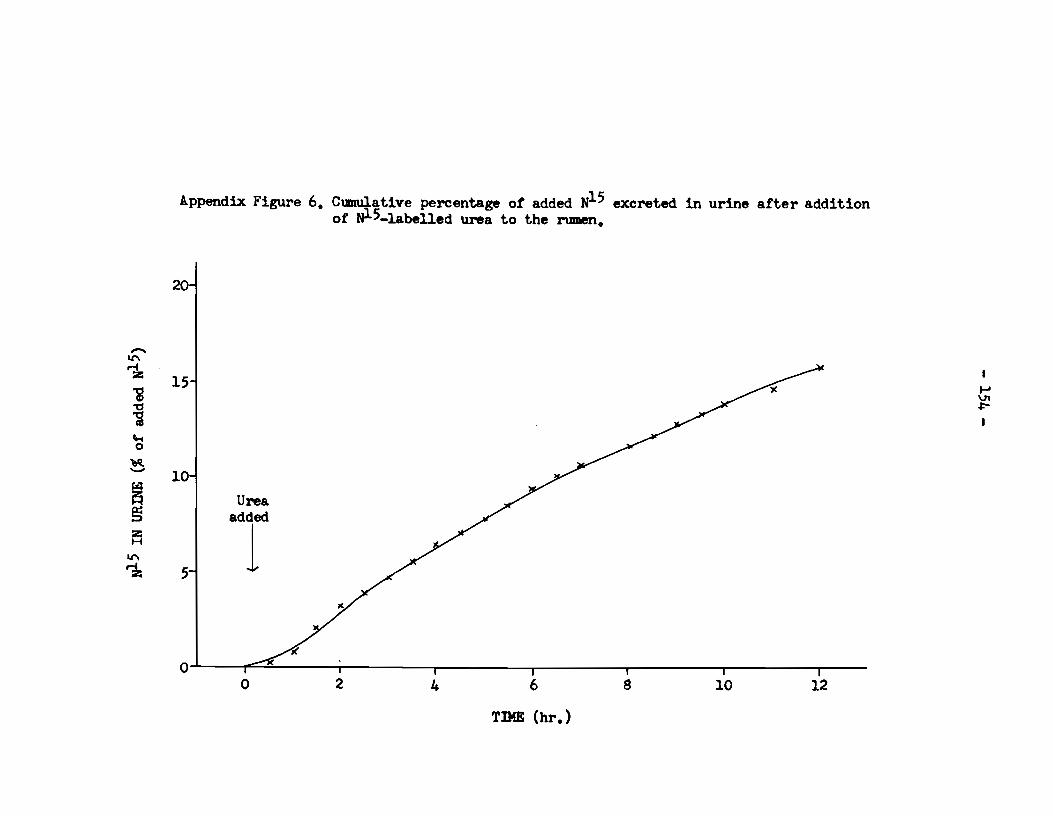
C. Urine: Excretion of Nl5 in Urine.
At each sampling time a m.easured volume of urine was
collected fran the animal and the total nitrogen content of the
sample was then determined. Subsequently the Nl5-enrichment of
this nitrogen component was measured. It was therefore possible to
calculate the total amount of N15 lost to tœ animal by excretion in
the urine at each sampling time. The resulta of these calculations
from samples obtained during the 12 hr. experimental period are shown
in Appendix Figure 6, where the cumulative quantity of Nl5 excreted
is expressed as a percentage of the Nl5 originally added to the rumen
as urea. The resulta show that there was an almost linear increase
wi th time in the total amount of N15 excreted by the animal between
the lst and 12th hours of the experiment. By the 12th hour after
administration of the urea, 16% of the added N15 had been excreted.
-Il'\ ~ i ~
al '-t 0
~ -~ § :z: H
Il'\
~
Appendix Figure 6. Cum.u,J.ative percentage of added W.5 exereted in urine after addition ot uJ-5-labelled urea to the rumen.
21
15
10
J Urea.
added
1
0 '"' 0 2 4 6
TIME (hr.)
s
~-
10 12
.......
"' ~ 1
- 155 -
S:imilar calculations were made on samples of urine
collected at the 30th, 78th and 102nd hours after administration
of the urea. The proportions of tœ added Nl5 which had been
excreted by these times were, respectively, 26%, 52% and 62%. By
the 102nd hour the degree of enrichment of urine-nitrogen with r5
had decreased to 0.03 atom % excess, suggesting that at this time
there was an almost insignificant excretion of N15.
DISCUSSION
The Prelimina.ry nature of tœ e.xperiment described, and the
reasons for some deficiencies in the resulta obtained from it, were
discussed in the Introduction. The chief factor limiting the value of
the resulta was the absence of reliable data for the changes in the
fluid volume of the rumen during the course of tœ e:xperiment which
could be used to convert the resulta obtained from the nitrogen analyses
to a "total rumen fluid volume" basis. Apparently the polyethylene glycol
solutions added to tha rumen did not become adequately mixed with the
rumen fluid, am subsequently measurements of the mG concentration in
rumen fluid samples gave an incorrect indication of the PEG distribution
volume and therefore also of the volume of liquid pre sent in the rumen.
For this reason, the resulta of thase determinations were discarded
and no attempt was made to put the resulta of the Nl5 analyses on an
absolute quantitative basie. Only the most general conclusions can
thus be drawn from tha resulta presented.
Within a very short time following the addition of labelled
ure a to the rumen of the sheep, all the nitrogen components analysed
were found to be enriched with N15 and furthermore, significant levels
of r5 enrichment occurred in them for at !east the succeeding 12 hr.
period. The enrichm.ent with Nl5 was found in the nitrogen of the rumen
microorganisme, and the sali va and urine of the animal. The re sul ts
therefore provide evidence in support of tha existence of some of the
pathways of nitrogen metabolism in the ruminant animal which have been
- 157 -
elucidated by various workers and which were illustrated in Figure A.
As already mentioned, the most important factor in the
utilization of dietary NPN by ruminants is the efficiency with which
it is incorp::>rated into microbial protein. Although resulta of the
present investigation clearly show that some labelled urea-nitrogen
was incorporated into the rumen microorganisms, they do not indicate
whether this occurred during synthesis of new bacterial protein or
whether the incorporation re sulted fran exchange between extracellular
Nl5 and intracellular nitrogen in the absence of synthesis. Nor do
the resulta indicate the proportion of added Nl5 which was utilized in
this way. An indication of the efficiency wi th 'Which tha animal utilized
added Ure!Hlitrogen can be obtained, however, from a consideration of
the amount of N15 excreted in tha urine. When 0.5 gm. urea was added
to the rumen the proportion of urea-nitrogen retained by the animal
within 12 hr. was 84%. By the 102nd hour after administration of the
urea, whan tha animal was again eating and drinking no:rmally, 62% of
the added N15 had been excreted in the urine. With respect to urine-
nitrogen excretion it is not possible to distinguish that of exogenous
origin although it is only this which representa a loss, through
inefficient utilization, of dietary nitrogen. Same nitrogen may also
be lost in the feces through inefficient absorption from the alimentary
tract, but no account was taken of fecal nitrogen in the present
investigation.
The figures obtained for urinary labelled nitrogen excretion
would undoubted.ly be subject to considerable variation if changes were
- 158 -
made in the experimental conditions. Nitrogen retention m.ight be
increased, for e.xampla, if a carbohydrate source such as glucose had
been added to the rumen at the same time as the urea, since this would
probably resulta in a greater incorporation of nitrogen into m.icrobial
protein. Retention might be reduced if a larger am.ount of urea were
added, since the rapid release of larger auantities of ammonia into
the rumen solution would result in more extensive absorption and
greater exogenous nitrogen excretion (Oyaert and Bouckaert, 1960).
This preliminary experiment has provided resulta which,
though requiring corroboration, would serve as a valuable starting
point for further, more extensive investigations on the fate of
Nl5_labelled urea in the rumen of the sbeep.
SlJIB[ARY
A preliminary experiment was carried out on the fate of
N15-labelled urea in the rumen of a sheep. Nl5-labelled urea was
introduced into the rumen of a fasting sheep, and changes in the pH
of the rumen solution and in the concentrations, and levels of ~5
enricbnent, of various nitrogen components in the rumen fluid, the
saliva and the urine of the animal -were followed during the succeed
ing 12 hr. period. Determinations of rumen volume were made during
the course of the experiment using polyethyleneglycol as a marker,
but the resulta were not reliable and were not reported.
The pH of tm rumen solution decreased gradually during the
first 7 - 8 hr. of the experiment and then increased again. The
ammonia concentration of the rumen solution showed considerable
fluctuations, but tended also to decrease to the 8th hour and then to
increase. The soluble nitrogen concentration of rumen fluid remained
relatively constant throughout the experimental period, but the level
of m.icrobial nitrogen gradually increasad.
Within i hr. following addition of the labelled urea to
the rumen and for the remainder of the experimental period, all the
nitrogen components analysed -were found to be enriched with r 5•
Generally the level of enrichment rose sharply within the first
i - 1 hr. and then decreased more slowly during the remainder of the
period. The leval of enrichment of m.icrobial nitrogen, ho-wever, rose
to o. 75 atom % excess Nl5 by the 8th hour after administration of the
- 160-
urea and than .feil dramatically to 0.1 atom % excess within 2 hr.
O.f t:œ total amount o.f Nl5 added to the rumen, 16% was
excreted in the urine o.f the animal by the 12th hour a.fter adminis
tration, and by the 102nd hour 62% had been excreted.
The resulta of this preliminary experiment provided
evidence in support o.f soma of the known pathways of nitrogen
metabolism. in the ruminant animal, and would be of value in planning
a more extensive investigation on the fate of urea-nitrogen in the
rumen.
BIBLIOGRAPHY
Annison, E.F. , and Lewis, D.
1959 Metabolisn in the Rumen. Metheun and. Co :-;-London.
Arias, C., Burroughs, w., Gerlaugh, P., and Bethke, R.M.
1951 The influence of different amounts and sources of energy upon in vitro urea utilization by rumen microorganisms. J. Animal Sei., 10: 683-692.
Belasco, I.J.
1954 New nitrogen feed compounds for ruminantsa laboratory evaluation. J. Animal Sei., 13: 601-610.
Belasco, I.J.
1956 The role of carbohydrates in urea utilization, cellulose digestion and fatty acid metabolism. J. Animal Sei. , 15: 496-508.
Hendrick, H.
1960 z. Tierphysiol. Tierernaehr. Futtermittelk., 15: 218-227. Cited in Cham. Abstr., 55: 4705(1961).
Hydén, S.
1961 Determination of the am.ount of fluid in the reticule-rumen of the sheep and its rate of passage to the omasum. Kungl. La.ntbrukshogskolans Ann., 27: 51-79.
1949 The nutritive value of nitrogenous compounds for ruminants. II. The formation of body protein from ure a labelled with the isotope Nl5. Sei. Agri., 29: 185-188.
Wegner, M.I., Booth, A.N., Bohstedt, G., and Hart, E.B.
1940 The nin vitro 11 conversion of inorganic nitro~n to protein by microorganisms from the cow's rumen. J. Dairy Sei., 23: 1123-1129.
Weiler, R.A., Gray, F. V., and Pilgrim, A.F.
1958 The conversion of plant nitro~n to microbial nitrogen in the rumen of the sheep. Brit. J. Nutr., 12: 421-429.







































































































































































![Research Article jmb RevieThe concept is to use the urease activity of microorganisms [9]. When microorganisms are in contact ... characterization, microorganism, Sporosarcina pasteurii](https://static.documents.pub/doc/80x56/6063fdf1646658391265f051/research-article-jmb-the-concept-is-to-use-the-urease-activity-of-microorganisms.jpg)











