Page 1
11510
1
Temporal Reconsolidation of Appetitive Behaviour
A Window in Time: The Effect of Temporal Reconsolidation Boundaries on the Persistence of Appetitive Behaviour
Word count: 6,364
Candidate number: 11510
Page 2
11510
2
Temporal Reconsolidation of Appetitive Behaviour
Abstract
There is ample evidence that the loss of operant behaviour seen in extinction does not reflect
a loss of memory or even behaviour (Bouton 1984, 1988, 2002; Bouton & Bolles, 1979;
Acosta, Boynton, Kirschner & Neisewander, 2005). However, reconsolidation provides a
means of updating the original conditioning memory, altering the pervasiveness of previously
reward-paired cues (Nader, 2003). Previous research has implicated context in incentive
motivation for appetitive behaviours (Crombag & Shaham, 2002). We used the
extinction/reinstatement model to examine the effects of reconsolidation on reinstatement of
extinguished sucrose-seeking behaviour in 32 mice. Animals were trained to self-administer
sucrose and subsequently underwent extinction. The conditioning memory was then
reactivated either 30 minutes or 6 hours before they were tested twice for cue and context-
primed reinstatement of the extinguished appetitive behaviour. The results of the study
indicated the renewal of behaviour as a function of context but reconsolidation was not
shown. The potential explanations for this, such as boundary conditions, are explored.
Page 3
11510
3
Temporal Reconsolidation of Appetitive Behaviour
Nomenclature:
NCSPP: non-conditioned stimuli period; CSPP: conditioned stimulus presentation period;
CS: conditioned stimulus; US: unconditioned stimulus; RET30min: conditioned group
reactivated thirty minutes before test; RET6hr: conditioned group reactivated six hours before
test.
Acknowledgements:
With particular thanks to Dr. Hans Crombag; firstly, for his initial suggestion of this subject
as an area for experimentation and secondly, for his continued help and support throughout
this project‟s completion.. I would like to thank the support technicians in the animal research
facility at University of Sussex for their help and David Mawer for supervising us within that
facility. Finally, I would like to thank Thomas Ridley-Siegert, with whom I jointly collected
the data for this project, for being a superb dissertation partner.
Page 4
11510
4
Temporal Reconsolidation of Appetitive Behaviour
A Window in Time: The Effect of Temporal Reconsolidation Boundaries on the Persistence of Appetitive Behaviour.
Consolidation is a time-dependent stabilization process through which short-term memories
(STM) are converted into persistent or long-term memories (LTM) (McGaugh, 1966, 2000;
Glickman, 1961). There is evidence to suggest that, once consolidated, memories cannot be
disrupted using amnestic agents such as protein-synthesis inhibitors or hypoxia (Dudai,
2004). However, consolidated memory traces can be altered when reactivation returns them
to a transient state (Nader, 2003; Sara, 2000; Gordon & Spear, 1973). This instigates memory
processes similar to those seen in new learning: the memory is briefly labile before being
merged back into the long term memory (LTM); this phenomenon is called “reconsolidation”
(Nader & Hardt, 2009). This brief period when the memory is mutable means that
reconsolidation can disrupt the initial memory trace, permanently altering it (Nader, 2003).
Reconsolidation research has been conducted in several classes of animals including
Insecta (honeybees: Eisenhardt, Menzel, & Stollhoff, 2008), Nematodes (round worms: Rose
and Rankin, 2006) Malacotraca (crabs: Pedreira, Pérez-Cuest, & Maldonado, 2002), and
Mammalia (rats: Nader et al, 2000) (mice: Chevere, et al., 2002) (humans: Gomez, Hardt,
Hupbach & Nadel, 2007). The fact that reconsolidation has been illustrated in an array of
species suggests that it is vitally important for updating learnt behaviours and memories.
Reconsolidation may do this by integrating common elements from related memories,
thereupon, reducing cognitive load (Walker, Brakefield, Hobson, & Stickgold, 2003).
A number of studies have used instrumental fear conditioning to study reconsolidation
of associative memories (Dębiec, Doyére, Nader & LeDoux, 2005; Schiller, Levy, Niv,
LeDoux & Phelps, 2008; Monfils, Cowansage, Klann & LeDoux, 2009). In this model, a
conditional stimulus (CS, for example, a tone) is paired with a negative unconditional
stimulus (US, for example, foot shock), producing a behaviour (for example, freezing). This
research has demonstrated that immediate (but not delayed) extinction, after the
commencement of reconsolidation, leads to attrition of responding (Schiller et al. 2008;
Monfils et al., 2009).
Research focussing on the extinction of operant appetitive learning, when the CS is
coupled with a rewarding US (for example, drugs) when an animal performs a behaviour (for
example pressing a lever), is limited. As with fear conditioning, learned responses can be
extinguished by presenting the uncoupled CS repeatedly (Bouton & Swartzentruber, 1991;
Page 5
11510
5
Temporal Reconsolidation of Appetitive Behaviour
Bouton, 2002; Bouton & Moody, 2004). A number of factors (for example context) can
reinstate responding (Bouton, 1984, 1988; Bouton & Bolles, 1979; Acosta et al., 2005). One
suggested reason for this is that testing often occurs shortly after extinction making the
extinction memory more retrievable than the acquisition memory; as the time between
extinction and testing increases the extinction memory becomes harder to retrieve, inducing
spontaneous recovery (Bouton, 1993). This suggests that extinction does not destroy the
acquisition memory but instead is “new learning” which is stored along with the initial
association. This extinction memory inhibits the expression of the conditioning memory
(Pavlov, 1927) and the instrumental action achieves an additional “meaning”; rendering the
signal‟s significance context specific.
As the initial conditioned behaviour reappears in a new context it has been proposed
that conditioning associations are temporally more stable than extinction associations
(Bouton & Bolles, 1979; Bouton, 1993). Bouton (2002) advances that the reason for this that
extinction is the second thing learnt about the CS. In order for extinction to overshadow
conditioning associations, it would have to occur repeatedly, in a number of different
contexts consequently encouraging the pervasiveness of the association (Bouton, 2006).
Ideally, such “new learning” would need to be ubiquitous to all new contexts (Bouton, 2002).
However, the initial memory trace would remain, thus, it is imperative to make the distinction
between the absence of behaviour and the absence of memory (Abel & Lattel, 2004). The
most effective means of altering behaviour is to modify the initial memory rather than
produce a second association.
Stollhoff et al. (2008) argue that for extinction to become the dominant association the
original conditioning memory must be updated using reconsolidation. This would mean that
extinction would supplant, rather than replace, the original memory (Quirk et al., 2000;
Akirav, Maroun & Raizel, 2006).
Research into reconsolidation is important for a number of reasons. Firstly, anxiety
disorders, for example phobias, could be treated by altering the initial memory (Bernstein &
Victor, 2009; Weisberg, 2009). An estimated lifetime prevalence of anxiety disorders is 18%
(Alonso et al., 2004; Chiu, Demler, Kessler, Merikangas & Walters, 2005) producing a huge
burden on the economy and healthcare systems. Exposure based therapies for anxiety
disorders operate on the principle of extinction and, therefore, it seems logical that relapse is
reinstatement of the conditioning association in a new context (Bruce et al., 2005; Bouton,
Gutiérrez, Moody & Zilski 2006). Therefore, a treatment which alters the original fear
Page 6
11510
6
Temporal Reconsolidation of Appetitive Behaviour
association would be invaluable as a behavioural therapy (Schiller, 2010). To this end,
research into reconsolidation of fear conditioning has already been carried out with some
success (Monfils et al., 2009; Nader, 2003).
Moreover, reconsolidation research would be of inestimable value for drug addiction
treatment. Drug addiction is defined as a progressive, compulsive disorder characterized by
recurrent craving and relapse even after long periods of abstinence (Olausson, Quinn Taylor
& Torregrossa, 2009).
According to Gordon, Godfrey, Parrott & Tinsley (2006) the annual economic and
social costs of drugs is approximately £15.4 billion in the UK. Drugs with high abuse
potentials, such as heroin and crack cocaine, also put huge strain on the NHS. There are
approximately 320,000 heroin and/or crack cocaine users in England (National Treatment
Agency Media Release 2009) an estimated 48% of which seek treatment each year
(Department of Health and the National Treatment Agency 2010). However, relapse rates are
high; 40-60% of addicts treated resume drug abuse within a year (McLellan & McKay,
1998). This suggests that the extinction method of drug-related cues in rehabilitation clinics is
ineffective (Conklin & Tiffany, 2002).
A possible reason for this high lapse rate is that associations acquired in the
rehabilitation centres are not applicable post-discharge. In animal models, reward associated
cues can reinstate responding (Bouton, 1984, 2002; Rescorla, 2001; Acosta et al., 2005;
Olausson et al., 2009; Crombag et al., 2002b). Stewart, de Wit, & Eikelboom (1984)
suggested a similar mechanism occurs in humans and that drug paired stimuli acquire
reinforcing and motivational effects. Neuroimaging studies have shown that drug related cues
activate the same meso-cortico-limbic circuitry as drugs (Contoreggi et al., 1996; Volkow et
al., 2006). Context has a marked effect on both extinction and reinstatement of conditioned
behaviour (Denniston, Gunther & Miller, 1998); as both discrete conditioned stimuli (for
example tone) and discriminative stimuli (for example smells) can be used by laboratory
animals as indicators of reward availability (Ettenberg &McFarland 1997; Dankiewicz,
Gracy, Koob & Weiss, 2000; Aicardi, Burattini, Janak & Zironi, 2006). When returned to
their drug conditioned context post-extinction these cues can reinstate drug seeking behaviour
(Crombag & Shaham, 2002a; Crombag, Grimm, & Shaham, 2002b; Bossert, Gray, Lu, &
Shaham, 2006; Bouton & Ricker, 1994). This is direct evidence that the context in which an
association is learnt is key to expressions of maladaptive behaviour (Robinson & Yager,
2010).
Page 7
11510
7
Temporal Reconsolidation of Appetitive Behaviour
Although extinction is highly context specific (Bouton, 2002), reconsolidation
directly alters the salience of the initial association (Nader, 2003). Dudai (2004) describes
reconsolidation as an update system wherein new information revises the initial memory
using “synaptic consolidation” stabilizing memories in a matter of hours (Dudai & Morris,
2000). This revision occurs through two major mechanisms: training induced updating and
updating instigated by dissonance between the training and reactivation memories (Morris &
Wang, 2010). Consequently, reconsolidation could allow for the salience of drug related
stimuli to become diminished across multiple dimensions.
In the past, research into reconsolidation has used a number of paradigms to disrupt
the reactivated memory. These include electric shock (Lewis, Miller & Misanin, 1968),
hypoxia (Sara, 1973), hypothermia (Mactutus, Smith & Riccio, 1980) and protein synthesis
inhibitors (Anokhin, Tiunova, Rose, 2002). These studies have shown the brief window in
which reconsolidation occurs is immediately following reactivation. For example, the protein
synthesis inhibitor anisomycin (ANI) has been shown to produce amnestic effects when
administered anywhere between thirty minutes before to six hours post-training (Mark &
Watts, 1971; Bjorklund et al., 1986; Freeman, Rose & Scholey, 1995; Debiec, LeDoux, &
Nader, 2002). Such treatments have been shown to stop working after six hours indicating
that reconsolidation occurs within a six hour window (Schafe & Ledoux, 2000). The temporal
sensitivity of reconsolidation has also been shown to effect the reactivation period, which
must be kept brief (under ten minutes) in order to stop extinction (Frankland et al., 2004). A
recent study by Monfils et al. (2009) exploited the transient reconsolidation window in a fear
conditioning study. Their reason for this is that the amnestic treatments that are currently in
use are not viable for therapeutic treatment. Instead, this paradigm involved a single ten
minute retrieval session prior to testing and results showed enhanced extinction.
Since Monfils et al. (2009) study there has yet to be an appetitive study examining the
effect of context on reconsolidation using temporal reconsolidation. Like most abused drugs,
palatable food releases dopamine into the nucleus accumbens-shell (Avena, Hoebel & Rada,
2005) suggesting a neurochemical similarity between intermittent sucrose and drug bingeing.
Furthermore, food cues acquire incentive motivational properties in a similar way to abused
drugs (Robinson & Yager, 2010) supporting the assertion that research using natural rewards
can further our understanding of abused drugs (Kelley & Berridge, 2002). If reconsolidation
can be effectively achieved, then this paradigm could be used therapeutically to reduce
maladaptive behaviours. We hypothesis that appetitive behaviour will be subject to renewal
Page 8
11510
8
Temporal Reconsolidation of Appetitive Behaviour
in animals tested six hours after memory reactivation. Conversely, animals tested thirty
minutes after reactivation will show diminished appetitive behaviours.
Method
Subjects:
The subjects were 32 C57BL mice housed in plastic and stainless steel cages in a
climate controlled facility (temperature: mean= 21.7⁰C, max= 22.6⁰C, min= 21.2⁰C. Relative
Humidity: mean= 47, max= 57, min= 46) in groups of 4 littermates. Mice had an initial mean
weight of 21.4 grams and maintained on a 24 hour circadian cycle. The animals were food
deprived throughout the study and maintained at 90% of their initial body weight. Water was
ad libitum for the duration of the experiment. The procedures were conducted in accordance
with the UK 1986 Animals (Scientific Procedures) Act 9 (project licence PPL 707072).
Apparatus:
The experiment was conducted in two sets of operant chambers. The following was
the same in for each context: Skinner boxes, measuring 27 x 25.8 x 30.4 cm in length, width,
and height respectively, were constructed from two metal and two clear acrylic walls. The
active lever (5.28cm²) was located (4cm) to the left of the sucrose receptacle
(counterbalanced to the right for half of the mice). The receptacle (width= 1.8cm, height=
2.3cm, depth= 2.9cm) contained an infra-red beam that recorded head-entries. The 10%
sucrose (approx. 50µl) was coloured with 10 drops (per 500ml) of “silver spoon” green food
colouring and by 5ml plastic syringes and a compressed-air pump.
A computer running (MET-PC-IV) was interfaced with the chambers and recorded
lever presses (active and inactive), head entries into the receptacle and the times at which the
behaviours occurred.
Operant chambers were located in wooden cabinets on one side of the experimental
room. The average background noise varied within each cabinet; the CS tone was set at 10db
above the background noise for each chamber. The CS was delivered by a speaker, mounted
on the cabinet roof, above the chamber.
All equipment was washed with fragrance-free soap and water between sessions and
wiped with 70% ethanol between days.
Page 9
11510
9
Temporal Reconsolidation of Appetitive Behaviour
The boxes from context A and B differed in terms of their tactile, visual and olfactory
background stimuli to keep them discriminably different. The skinner boxes used for context
A were scented with clove, lit by a house light mounted on the wooden cabinet and the floor
of each Skinner box was left uncovered.
Context B boxes were scented using peppermint, lit by a panel light above the food
receptacle and the floors were covered with metal grids.
The boxes were scented by putting 5 drops of either “Julia Lawless Aqua Oleum
Peppermint Oil” or “Julia Lawless Aqua Oleum Clove Bud Oil” in the waste tray, this oil was
replaced every session after the tray was cleaned (Bouton & King, 1983).
Design:
This study examined the effect of the temporal reconsolidation window on contextual
renewal of appetitive behaviours. The experiment was a (2 x 2) mixed measures design. The
dependent variables were active/inactive lever responses, head entries and time spent in the
receptacle during CS presentation period (CSPP) and non-CS periods (NCSP) and
cumulative CS presentation periods. The independent variables were the reactivation group
(RET30min and RET6hr) and the context (either ABA or ABB).
Acquisition of the operant behaviour is shown by an increase in dependent variables
across conditioning sessions. Extinction is shown by a decline in all appetitive behaviours
across sessions.
During testing the animals were either be returned to their conditioning context or
remained in their extinction context. For renewal to be shown, the ABA condition would
have shown higher rates of responding than in the ABB condition; there would be a
significant effect of context but not reactivation group. Conversely, for reconsolidation to be
shown the 30min group would have had lower rates of appetitive responding than the 6hr
group, with RET30min ABB showing the lowest rate of responding and RET 6hr ABA
showing the highest level of responding.
Procedure:
In order to reduce expressions of neophobia, the mice undertook brief (40 minutes
for 2 days) receptacle approach training before the experiment began. In this training, mice
were placed in the Context A (counterbalanced to Context B for half of the mice) with a
Page 10
11510
10
Temporal Reconsolidation of Appetitive Behaviour
syringe pump activated on a RI30 schedule [min=15seconds, max=45seconds] set to
deliver a small, of sucrose solution [approximately 50µl] into the receptacle.
Stage one: conditioning: For days 3 to 14, mice were placed in context A
(counterbalanced to context B for half of the mice) and partook in conditioning sessions.
Initially, these were 40 minutes in length but were reduced to 30 on day 5. During these
sessions, a tone was paired with 10% sucrose delivery (CS+) in 5 second intervals if the
active lever was pressed (active/inactive levers were counterbalanced). During the 5
second sucrose delivery period any additional lever pressers were ineffective. This was
then followed by a 5 second interval of no noise but active lever presses remained
ineffective. This 10 second interval will be referred to as the CS presentation period
(CSPP) and the remaining proportion of the session will be called the non-CS period
(NCSP).
Stage two: reactivation and extinction: From days 15 to day 21 the mice were separated
into two groups. Both groups experienced a 10 minutes retrieval trial in the extinction
context (Frankland et al., 2004). Group 1 (RET30min) then were returned to their homes
cages for 30 minutes, whereas Group 2 (RET6hr) were returned to their home cages for 6
hours, before they then conducted a 30 minute extinction session in context B
(counterbalanced to context A for half of the mice). During extinction, each lever press of
the active lever (counterbalanced for half of the mice) elicited CS+ presentation in the
absence of sucrose.
Stage three: test.: On day 22, the reactivation groups (RET6hr and RET30min) were
further divided into two groups; one of these groups was then given a conditioned
reinforcement test in the same context as their extinction occurred (ABB) [for example,
RET6hr ABB and RET30min ABB]. The other group was given a conditioned
reinforcement test in the same context as their conditioning occurred (ABA) [RET6hr
ABA and RET30min ABA].
Page 11
11510
11
Temporal Reconsolidation of Appetitive Behaviour
Figure 1: Summary of experimental procedure. Contexts A and B illustrated here were
counterbalanced for half the animals. (Illustrations adapted from Abel and Lattal, 2001)
Results
Mauchley‟s test revealed that the assumption of sphericity had been violated a number of
main effects for conditioning, reactivation and extinction and so was adjusted using either
Greenhouse-Geisser or Huynh-Feldt estimates of sphericity
Stage One: Conditioning
Figure 2: Mean rate (per minute) of reinforced lever presses during CS presentation
periods across conditioning sessions (i.e. 1-12) Error bars ± 1 SE. As the 12 conditioning
sessions progressed there was an increase in the mean rate of active lever presses per
minute followed by a head entry into the receptacle.
Figure 2 shows an increase in reinforced lever presses throughout conditioning. This was
confirmed by a repeated measures ANOVA showing a significant main effect of session on
0
0.5
1
1.5
2
2.5
3
3.5
1 2 3 4 5 6 7 8 9 10 11 12
Rate
of r
espo
nse/
min
Session
CS Presentations
Page 12
11510
12
Temporal Reconsolidation of Appetitive Behaviour
the rate of reinforced lever presses per minute, F(5.29, 158.59)= 44.52, p<.001. A post hoc
contrast revealed that the rate of CS presentations were significantly higher, F(1, 30)=155.71,
p<.001, r = .92, in the last session than the first. There was no difference between the
reactivation groups in the rate of reinforced lever presses, F(1, 30) =.006, p=.94.
Figure 3 Mean percentages of head entries into the receptacle during the CS presentation
period (CSPP) and the non-CS period (NCSP) across all conditioning sessions. Error bars ± 1
SE. This illustrates a change in behaviour conditioning as a function of training. From
session 8 onwards the animals entered the receptacle during the CSPP over 60% of the
time. Conversely, the number if head entries during the NCSP declined throughout the
sessions.
Figure 3 shows that session had a differential effect on head entries during the CSPP and
NCSP indicating that behaviour changed as a function of training. A two-way repeated
measures ANOVA revealed that there was no significant main effect of session, F(1.47,
44.10) =1.71, p=.20, suggesting session did not have the same effect on responding during
CSPP and NCSP. There was a significant main effect of stimulus presence (CSPP compared
to NCSP), F(1, 30)=15.05, p=.001. Therefore, the percentage of head entries during the
CSPP was significantly higher than NCSP head entries.
There was no significant difference between the 6hr and 30minute groups in their
percentage of head entries, F(1, 30) =13.45, p=.001.
0
10
20
30
40
50
60
70
80
90
1 2 3 4 5 6 7 8 9 10 11 12
Perc
enta
ge o
f hea
d en
trie
s/m
in
Session
CS presentation period Non-CS presentation period
Page 13
11510
13
Temporal Reconsolidation of Appetitive Behaviour
Importantly, there was a significant interaction between CSPP and session (1-12),
F(4.76, 142.87)=48.56, p<.001, indicating training altered the CS. A post hoc contrast
illustrated that as conditioning sessions progressed the percentage of head entries during the
CSPP was significantly higher than NCSP head entries, F(1, 30)=162.79, p<.001, r = .92
These results suggest across conditioning the animals differentiated between the
CSPP and NSPP; entering the receptacle during CSPP more than during NCSP.
Figure 4: Mean press rates per minute of active lever and inactive lever across
conditioning sessions (1-12). Error bars ± 1 SE. Portrays the change the rate of lever
pressings throughout the conditioning sessions (i.e. 1-12) for both the active and inactive
levers. Rate of active lever presses increased throughout the sessions whereas the rate of
pressing for the inactive lever did not change across sessions.
Figure 4 indicates that the rate of compressions on both levers changes as a function of
conditioning. A two-way repeated measures ANOVA revealed a significant main effect of
session, F (4.64, 139.08) =37.60, p<.001, suggesting session effected CSPP and NCSP
equivalently.
There was a significant main effect on press rate (per minute) of active and inactive
lever, F(1, 30) =159.37, p<.001. There was no significant difference between the reactivation
groups, F(1, 30) =.013, p=.91. Suggesting the active lever was pressed significantly higher
than the inactive lever by both RET30min and RET6hr.
There was a significant interaction between the session number and the rate of lever
pressing, F(4.93, 147.80) =36.22, p<.001. This suggesting both number and rate of lever
compressions varied with conditioning session. A post hoc contrast revealed as sessions
Page 14
11510
14
Temporal Reconsolidation of Appetitive Behaviour
progressed the active lever press rate was significantly higher than the inactive lever press
rate, F(1, 30)=158.62, p<.001, r = .92.
These results suggest that, with training, the levers could elicit differential responses and that
the animals pressed the active lever significantly more than the inactive lever indicating the
acquisition of the operant behaviour.
Stage Two: Reactivation and Extinction:
A: Reactivation
Following stage one (conditioning) the animals were split into two counter-balanced
conditions (RET6hr and RET30min). Each group underwent a brief ten minute reactivation
session either six hours (RET6hr) or thirty minutes (RET30min) before thirty minutes
extinction session (see figure 1 for a schematic).
Figure 5: Mean rate of lever presses on previously active lever per minute during CS
presentation period (CSPP), for reactivation groups RET6hr and RET30min, across
reactivation sessions (1-7). Error bars ± 1 SE. For both the RET60hr and RET30min group
the number of responses decreased throughout the sessions.
The rate of responding on the active lever during CSPP decreased as reactivation (sessions 1-
7) progressed; initially both reactivation groups show the same rate of responding but rate of
responding for group RET30min decreased more rapidly, and was consistently lower, than
RET6hr. This observation was confirmed by a two-way mixed measures ANOVA which
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
1 2 3 4 5 6 7
Rate
of r
espo
nse/
min
Reactivation Session
6hr 30min
Page 15
11510
15
Temporal Reconsolidation of Appetitive Behaviour
revealed there was a significant main effect of session on the rate of reinforced lever presses
during CS presentations periods, F(2.94, 64.73)= 15.74, p<.001. A post hoc contrast revealed
that the rate of reinforced lever presses during CSPP were significantly lower, F(1, 22)
=44.94, p<.001, r = .82, in the last session than the first.
There was no significant interaction effect between the session and the reactivation
group, F(2.94, 64.73) =.834, p=.48. However, there was a significant main effect of group on
the rate of reinforced lever presses during CSPP, F(1, 22) =5.36, p<.05. This suggests that
the 30min group responded at a significantly lower rate than the 6hr group
Figure 6: Shows the percentage of head entries of the reactivation groups RET6hr and
RET30min during the CS presentation period (CSPP) and non-CS period (NCSP). Error bars ±
1 SE. Throughout the reactivation sessions the number of head entries in the CS
presentation period decreases. This decrease is more rapid in the RET6hr than the
RET30min group. For NCSP behaviour, in session one there is disparity in the behaviour
between the RET6hr and the RET30min groups. Both groups show a decline in this
behaviour as a function of reactivation.
Figure 6 shows the decrease in head entries during the CSPP as a function of the reactivation
sessions (1-7). A two-way mixed measures ANOVA which revealed that there was a
significant main effect of session on the percentage of head entries into the receptacle during
the CSPP, F(6, 132)= 5.304, p<.001. A post hoc contrast revealed that the percentage of head
entries during the CSPP was significantly lower, F(1,22)=25.76, p<.001, r = .73, in the last
0
20
40
60
80
100
120
1 2 3 4 5 6 7
% o
f hea
d en
trie
s int
o th
e Re
cept
acle
Reactivation Sessions
% of entries in CSPP RET6hr % of entries in CSPP RET30min
% of entries in NCSP RET6hr % of entries in NCSP RET30min
Page 16
11510
16
Temporal Reconsolidation of Appetitive Behaviour
session than the first; suggesting that during stage two both reactivation groups equally
decreased in the percentage of head entries during the CSPP indicating the extinction of the
operant behaviour.
Throughout reactivation the percentage of head entries during the NCSP increases
for both reactivation groups. This observation was confirmed by a two-way mixed measures
ANOVA which showed a significant main effect of session on percentages of head entries
into the receptacle during non-CS period for both RET6hr and RET30min, F(6, 132)= 7.97,
p<.001. A post hoc contrast revealed a significant increase in this behaviour between the first
and last session, F(1, 22) =23.55, p<.001, r = .72.
The mean percentage of NCSP head entries declined equally for both reactivation
groups; suggesting the mice were becoming less accurate in their responding.
There was no significant effect of group on the percentage of NCSP, F(1, 22) =.67,
p=.42 or CSPP, F (1, 22) =.33, p=.57, head entries. Furthermore, there was no significant
interaction effect between the session and the reactivation group for NCSP, F(6, 132) =.90,
p=.50, or CSPP, F(6, 132) =.56, p=.76, head entries.
Figure 7: Mean rate (per minute) of lever presses on the previously active lever and
inactive lever during CS presentation period (CSPP), for reactivation groups RET6hr and
RET30min, across reactivation sessions (1-7). Error bars ± 1 SE. For both the RET60hr and
RET30min group the number of responses decreased throughout the sessions.
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
1 2 3 4 5 6 7
Rate
of L
ever
Pre
sses
per
Min
ute
Reactivation Session
Inactive RET6hr Inactive RET30min Active RET6hr Active RET30min
Page 17
11510
17
Temporal Reconsolidation of Appetitive Behaviour
In session one active lever press rates for both groups are similar but reactivation
progressed; the rate of responding for group RET30min decreased more rapidly, and was
consistently lower, than RET6hr. This observation was confirmed by a two-way mixed
ANOVA which revealed a significant main effect of session on the press rates of the
previously active lever during CSPP for both reactivation groups, F(2.29, 50.42)= 15.80,
p<.001. A post hoc contrast revealed that the lever press rates of the (previously) active lever,
for reactivation groups were significantly lower, F(1,22)=40.34, p<.001, r=.80, in the last
session than the first.
Both reactivation groups show a consistently low response rate on the inactive lever.
There is a small increase in this behaviour in session 5 but this quickly decreases by session
6. This observation was confirmed by a two-way mixed measures ANOVA that revealed
there was no significant main effect of session on the press rates of inactive lever, F(2.20,
48.43)=2.45, p=.09.
There was no significant main effect of reactivation group on the press rates of
inactive, F(1, 22) =.37, p=.55 or active lever, F(1, 22) =2.12, p=.16. There was no significant
interaction effect between the session and the reactivation group for inactive, F(2.20, 48.43)
=1.43, p=.25, or active lever press rates, F(2.20, 48.43) =1.43, p=.25.
The results from reactivation show a reduction in operant behaviour since condition
indicating the extinction of the appetitive behaviour.
B: Extinction.
Page 18
11510
18
Temporal Reconsolidation of Appetitive Behaviour
Figure 8: Mean rate of reinforced lever presses (per minute) of the previously-active lever
during CS presentations (CSPP) for RET6hr and RET30min across extinction sessions. Error
bars ± 1 SE. Both reactivation groups show a decline in behaviour but throughout the
sessions responding of the RET6hr group was lower than that of RET30min.
Figure 8 shows a gradual decrease in the rate (per minute) of compressions of the previously-
active lever during CS presentation across the extinction sessions for both reactivation
groups. A two-way mixed measures ANOVA confirmed that there was a significant main
effect of session on the rate of CS Presentations, F(3.38, 101.43) = 29.24, p<.001. A post hoc
contrast revealed that the rate of CS presentations was significantly lower, F(1, 30) =62.93,
p<.001, r = .82, in the last session than the first for both reactivation groups. This suggests
that although in the conditioning sessions the CS presentation rate was high the behaviour
was now becoming extinct as the extinction sessions (1-7) progressed.
There was a significant effect of group on the rate of CS presentations, F(1, 30)
=4.68, p<.05. The rate of presses (per minute) on the previously active lever during the CSPP
was significantly lower for RET6hr than RET30min. However, there was no significant
interaction effect between the session and the reactivation group, F(3.38, 101.43) =1.76,
p=.15.
0
0.5
1
1.5
2
2.5
1 2 3 4 5 6 7
Rate
of R
espo
nse/
min
Session
6hr 30min
Page 19
11510
19
Temporal Reconsolidation of Appetitive Behaviour
Figure 9: Shows the percentage of head entries during the CS presentation period (CSPP)
and non-CS period (NCSP) across extinction sessions for RET6hr and RET30min. Error bars ±
1 SE. The percentage of head entries during the CS interval decreased from the first
session indicating that the behaviour was changed as a function of extinction training (i.e.
sessions 1 -7).
Figure 9 indicates that head entry behaviour during CSPP was altered across the extinction
sessions. Both reactivation groups show a decline in the behaviour and whilst there was a
difference between these groups in session 1 it was gone by the last session, 7. A two-way
mixed measures ANOVA which revealed that there was a significant main effect of session
on the percentage of head entries into the receptacle during the CSPP, F(5.27, 157.97) =
4.56, p=.001. A post hoc contrast revealed that the percentage of CS head entries was
significantly lower for both groups, F(1, 30) =12.82, p=.001, r = .54, in the last session than
the first. This suggests during session 1 the mice were making head entries during the CSPP
but that as extinction carried on this behaviour diminished.
This figure also shows that the percentage of head entries into the receptacle during
NCSP changed as a function of extinction training; both reactivation groups show a
consistent increase throughout extinction. This observation was confirmed by a two-way
mixed measures ANOVA which revealed that there was a significant main effect of session
on the percentage of NCSP head entries into the receptacle, F(6, 180)= 4.72, p<.001. A post
hoc contrast revealed that the percentage of NCSP head entries was significantly higher, F(1,
30) =12.82, p=.001, r = .55, in the last session than the first for both reactivation groups.
0102030405060708090
100
1 2 3 4 5 6 7
Perc
enta
ge o
f Hea
d En
trie
s
Extinction Session
% of entries in NCSP RET6hr % of entries in NCSP RET30min
% of entries in CSPP RET6hr % of entries in CSPP RET30min
Page 20
11510
20
Temporal Reconsolidation of Appetitive Behaviour
This suggests that both reactivation groups showed a reduced accuracy in their
operant behaviour between the first and last extinction sessions.
There was no significant difference between the reactivation groups for NCSP,
F(1,30)=1.55, p=.22, or CSPP, F(1,30)=1.44, p=.24, on the percentage of head entries. There
was no significant interaction effect between the session and reactivation groups for the
percentage of head entries made during NCSP, F(6, 180)=1.20, p=.31, or CSPP, F(5.27,
157.97)=1.52, p=.18.
Figure 10: Mean compression rates (per minute) of previously active and inactive levers
across extinction sessions for RET6hr and RET30min. Error bars ± 1 SE. Both RET6hr and
RET30min, maintain low press rates per minute of the inactive lever until session 5 when
it increases marginally only for it to decrease in session 6. For the active lever, the press
rates per minute of both reactivation groups rapidly decreased until session 5 (when they
both increased) and then decreased rapidly again after that session
As shown in figure 10, the mean lever press rate per minute of the previously active lever
changed as a function of training (i.e. sessions 1 to 7) but there was little difference between
RET30min and RET6hr. This observation was confirmed by a two-way mixed measures
ANOVA which revealed that there was a significant main effect of session on the rate of
active lever presses per minute, F(2.14. 64.12)= 26.12, p<.001. A post hoc contrast revealed
that the rate of active lever presses per minute were significantly lower, F(1, 30) =46.92,
p<.001, r = .78, in the last session than the first session for both reactivation groups.
0
1
2
3
4
5
6
1 2 3 4 5 6 7
Leve
r Pre
ss R
ate/
min
Extinction Session
Inactive RET6hr Inactive RET30min Active RET6hr Active RET30min
Page 21
11510
21
Temporal Reconsolidation of Appetitive Behaviour
The press rate (per minute) for the inactive lever changed as a function of extinction
training (i.e. sessions 1 to 7) but this did not differ between reactivation groups; a two-way
mixed measures ANOVA confirmed there was a significant main effect of session on the rate
of inactive lever presses per minute, F(3.37, 101.01)= 12.67, p<.001. A post hoc contrast
revealed that the rate of inactive lever presses per minute was not significantly different, F(1,
30) =3.25, p=.08, r=.31, in the last session than the first session for RET30min and RET6hr.
This suggests that both reactivation groups pressed the inactive lever at an equal, low rate
across sessions.
There was no significant effect of reactivation group on the rate of active lever
presses per minute, F(1, 30) =2.97, p=.10, or the rate of inactive lever presses per minute,
F(1, 30) =1.09, p=.31. There was no significant interaction effect between the session and the
reactivation group for either active lever press rate, F(2.14, 64.12) =1.47, p=.24, or the rate
of inactive lever presses (per minute), F(3.37, 101.01) =1.70, p=.17.
The results from stage 2 show a decline in appetitive behaviours (i.e. lever presses and head
entries) as a function of training.
Stage 3: Tests.
Stage 3 involved RET6hr and RET30min being split; half the animals were returned to their
original conditioning context (ABA) and the other half were returned to their extinction
context (ABB) (for a schematic see Figure 1). Reconsolidation will have occurred if
RET30min show lower rates of appetitive behaviours than the RET6hr group. Renewal
would be shown if animals in ABA would show higher levels of appetitive behaviour than
ABB mice.
Page 22
11510
22
Temporal Reconsolidation of Appetitive Behaviour
Figure 11: The mean rate of reinforced lever presses per minute during CS presentation
periods (CSPP) for reactivation groups RET6hr and RET30min in context ABB and ABA.
Error bars ± 1 SE.
Figure 11 shows the mean rate of active lever presses during CSPP was higher in context
ABA than in context ABB, however, there is no difference in rates of active lever presses
between RET6hr and RET30min; suggesting renewal. This observation was confirmed by a
two-way mixed measures ANOVA which revealed that there was a significant difference
between contexts ABA and ABB, F(1,30)=18.69, p=.001, but no significant effect of
reactivation group, F(1,30)=.001, p=.97. This suggests renewal of active lever pressing
during CSPP for both reactivation groups but that reconsolidation did not occur. There was
no significant interaction effect between context and the reactivation group, F(1, 30) =.44,
p=.05.
0
0.5
1
1.5
2
2.5
6hrs 30min
Rate
/min
Reactivation Group
CS Presentations ABB CS Presentations ABA
Page 23
11510
23
Temporal Reconsolidation of Appetitive Behaviour
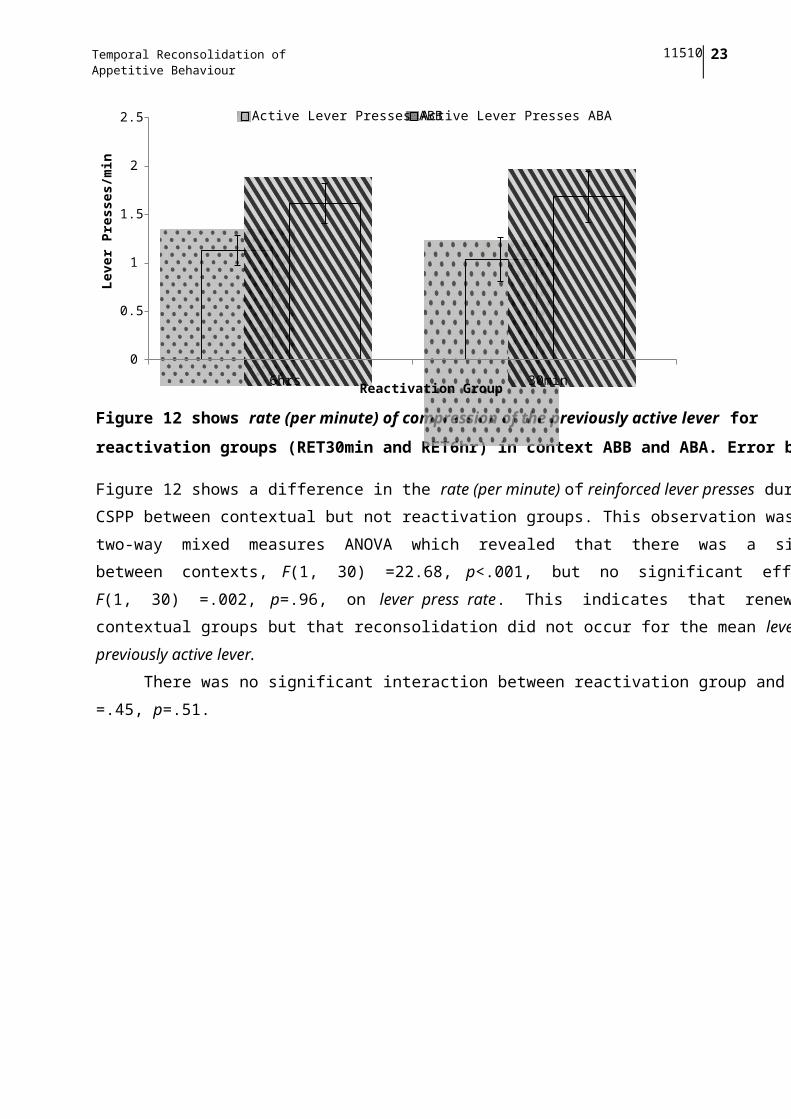
Figure 12 shows rate (per minute) of compression of the previously active lever for
reactivation groups (RET30min and RET6hr) in context ABB and ABA. Error bars ± 1 SE.
Figure 12 shows a difference in the rate (per minute) of reinforced lever presses during the
CSPP between contextual but not reactivation groups. This observation was confirmed by a
two-way mixed measures ANOVA which revealed that there was a significant difference
between contexts, F(1, 30) =22.68, p<.001, but no significant effect of reactivation group,
F(1, 30) =.002, p=.96, on lever press rate. This indicates that renewal occurred for both
contextual groups but that reconsolidation did not occur for the mean lever press rates of the
previously active lever.
There was no significant interaction between reactivation group and context, F(1, 30)
=.45, p=.51.
0
0.5
1
1.5
2
2.5
6hrs 30min
Leve
r Pre
sses
/min
Reactivation Group
Active Lever Presses ABB Active Lever Presses ABA
Page 24
11510
24
Temporal Reconsolidation of Appetitive Behaviour
Figure 13: Mean rate of inactive lever presses per minute for reactivation groups RET6hr
and RET30min, in context ABB and context ABA. Error bars ± 1 SE.
Figure 13 suggests there is no difference in rate of compressions across contextual groups or
reactivation group. This observation is confirmed by a two-way mixed measures ANOVA
which revealed that there was no significant difference between contexts ABA and ABB,
F(1,30)=.082, p=.78 and no significant effect of reactivation group, F(1,30)=1.48, p=.23.
This indicates that there was no renewal or reconsolidation effect on the inactive lever
suggesting that only behaviour that mediates the CS is subject to renewal. Furthermore, there
was no interaction effect between context and the reactivation group, F(1, 30)=.60, p=.45.
0
0.5
1
1.5
2
2.5
6hrs 30min
Leve
r Pre
sses
/min
Reactivation Group
Inactive Lever Presses ABB Inactive Lever Presses ABA
0
10
20
30
40
50
60
70
80
90
100
6hrs 30min
Pere
cent
age
of h
ead
entr
ies t
o m
agaz
ine
Reactivation Group
Magazine Entries During CSPP ABB Magazine Entries During CSPP ABA
Page 25
11510
25
Temporal Reconsolidation of Appetitive Behaviour
Figure 14: Mean percentage of head entries to the receptacle made during CS presentation
period (CSPP) for RET6hr and RET30min in context ABB and ABA. Error bars ± 1 SE. This
graph suggests a difference in contextual but not reactivation group.
Figure 14 shows the percentage of head entries into the receptacle during the CSPP were
higher in ABA than in ABB, suggesting renewal, but no apparent difference between
reactivation groups. This observation was confirmed by a two-way mixed measures ANOVA
which revealed that there was a significant difference between contexts ABA and ABB,
F(1,30)=11.75, p<.01 but no significant effect of reactivation group, F(1,30)=.11, p=.74. This
suggests that the renewal effect occurred for head entries during the CSPP for both
contextual groups but also that reconsolidation did not occur for head entries during the
CSPP. There was no significant interaction effect between context and the reactivation group,
F(1, 30) =.82, p=.37.
Figure 15 Mean percentages of entries to the receptacle made during non-CS period
(NCSP) in context ABB and ABA for reactivation groups RET6hr and RET30min. Error bars ±
1 SE.
Figure 15 shows that the percentage of head entries into the receptacle during the NCSP was
higher for group ABB than ABA but this behaviour did not differ across reactivation groups.
This observation was confirmed by a two-way mixed measures ANOVA which revealed that
there was a significant difference between contexts ABA and ABB, F(1,30)=14.69, p<.001
but no significant effect of reactivation group, F(1,30)=.38, p=.54. This suggests that the
0
10
20
30
40
50
60
70
80
90
100
6hrs 30min
Pere
cent
age
of h
ead
entr
ies t
o m
agaz
ine
Reactivation Group
Magazine Entries NCSP ABB Magazine Entries NCSP ABA
Page 26
11510
26
Temporal Reconsolidation of Appetitive Behaviour
renewal effect occurred for head entries during non-CS periods for both contextual groups
but that reconsolidation did not occur for head entries during NCSP.
There was no significant interaction effect between context and the reactivation
group, F(1, 30) =.23, p=.64.
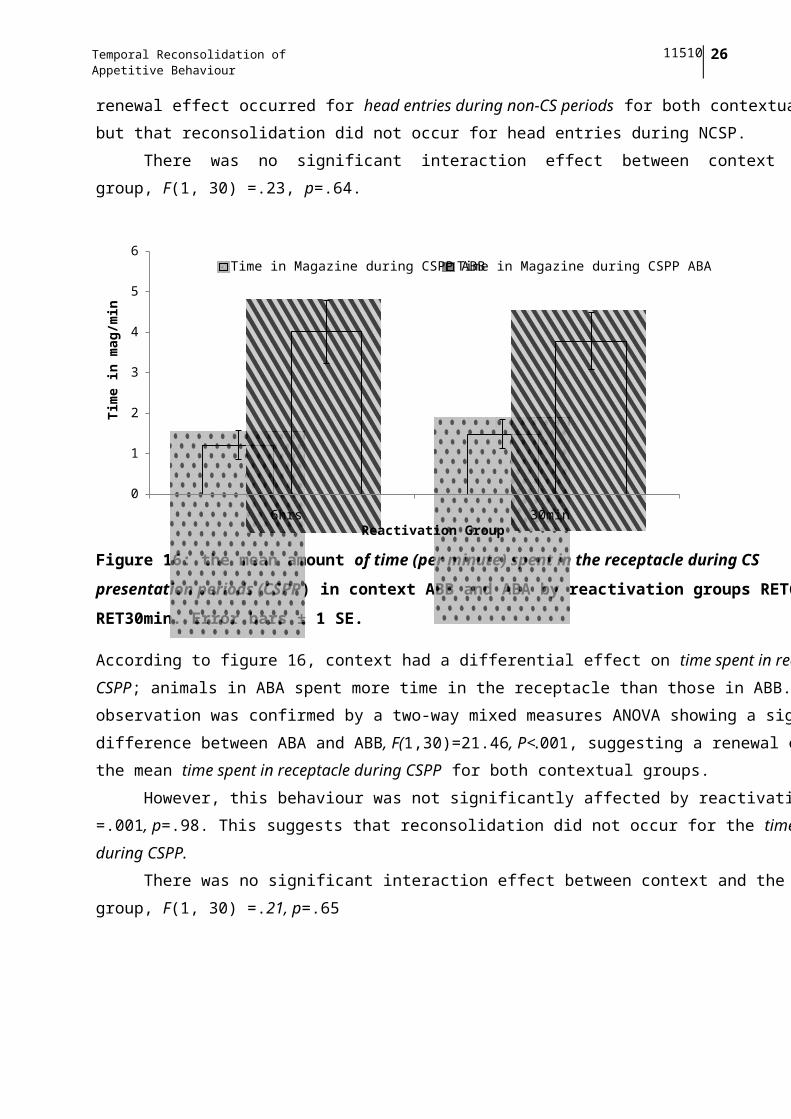
Figure 16: the mean amount of time (per minute) spent in the receptacle during CS
presentation periods (CSPP) in context ABB and ABA by reactivation groups RET6hr and
RET30min. Error bars ± 1 SE.
According to figure 16, context had a differential effect on time spent in receptacle during the
CSPP; animals in ABA spent more time in the receptacle than those in ABB. This
observation was confirmed by a two-way mixed measures ANOVA showing a significant
difference between ABA and ABB, F(1,30)=21.46, P<.001, suggesting a renewal effect for
the mean time spent in receptacle during CSPP for both contextual groups.
However, this behaviour was not significantly affected by reactivation group; F(1, 30)
=.001, p=.98. This suggests that reconsolidation did not occur for the time spent in receptacle
during CSPP.
There was no significant interaction effect between context and the reactivation
group, F(1, 30) =.21, p=.65
0
1
2
3
4
5
6
6hrs 30min
Tim
e in
mag
/min
Reactivation Group
Time in Magazine during CSPP ABB Time in Magazine during CSPP ABA
Page 27
11510
27
Temporal Reconsolidation of Appetitive Behaviour
Figure 17 compares the mean amount of time (per minute) spent in the receptacle in non-
CS periods (NCSP) in context ABB and ABA by reactivation groups RET6hr and RET30min.
Error bars ± 1 SE.
The time spent in the receptacle during NCSP was low and stable for both contextual and
reactivation conditions. A two-way mixed measures ANOVA confirmed no significant
difference in time spent in receptacle during NCSP between contexts ABA and ABB,
F(1,30)=1.17, p=.29 and no significant effect of reactivation group, F(1,30)=.02, p=.88. This
suggests that neither renewal nor reconsolidation occurred for the time spent in receptacle
during NCSP. There was no significant interaction effect between context and the
reactivation group, F(1, 30) =3.66, p=.07.
The results reflect a difference of contextual but not reactivation conditions, meaning renewal
but not reconsolidation was shown. Renewal was shown for reinforced lever presses during
CSPP, press rate of (previously) active lever, head entries during the CSPP and NCSP, and
time spent in receptacle during the CSPP. Reactivation conditions did not affect any results
meaning reconsolidation was not shown for behaviours.
0
1
2
3
4
5
6
6hrs 30min
Tim
e in
mag
/min
Reactivation Group
Time in Magazine NCSP ABB Time in Magazine NCSP ABA
Page 28
11510
28
Temporal Reconsolidation of Appetitive Behaviour
Discussion
During stage one; there was an increase in rate of active lever presses per minute and
head entries during CSPP. Conversely, the percentage of head entries into the receptacle and
active lever presses during NCSP dropped. The animals demonstrated discrimination
between the active and inactive levers: as a function of training active lever presses increased
but inactive lever presses decreased. There was no significant difference between reactivation
or contextual groups at any point during conditioning showing that behaviour was changing
only as a function of training. These results suggest that the mice were discriminating
between the CSPP and NCSP and between the inactive and active levers. This indicates they
had developed the operant behaviour as a function of conditioning training.
During stage two, behavioural changes included a decrease in the number of head
entries and lever presses during CSPP and NCSP. This suggests the mice were having their
previously acquired behavioural repertoire extinguished, reducing the accuracy of their
responding. Throughout reactivation there was no significant effect of contextual or
reactivation groups indicating behaviour was changing only as a function of training.
Stage three of the experiment indicated a renewal of the following conditioned
appetitive behaviours: rate of head entries and lever press rates of the previously active lever
during the CSPP, time spent in the receptacle during CSPP. Response rates during NCSP
remained low as did inactive lever press rates (per minute).
The renewal of the operant behaviour conditioned in stage one supports previous
research (Bossert et al 2006) and confirms the hypothesis that context can mediate appetitive
behaviours.
However, results indicated no significant difference of reactivation groups; the fact
that the behaviours did not alter as a function of time indicates that reconsolidation was not
shown. This result is contrary to our second prediction that a reactivation of conditioning
contexts would affect appetitive behaviours post-extinction. This finding is opposed to
previous research which used temporal reactivation as an amnestic treatment in
reconsolidation (Monfils et al. 2009).
Although this study reactivated, disrupted and tested the memory within the labile
period, thus fulfilling the criteria for reconsolidation described by Lewis et al. (1968),
reconsolidation may still have been disrupted by boundary conditions (Lewis, 1969; Gordon
& Spear, 1973). According to Nader & Einarsson (2010), boundary conditions are explicit
parameters which stop the reconsolidation of memories in physiological or experimental
Page 29
11510
29
Temporal Reconsolidation of Appetitive Behaviour
conditions in which it would ordinarily occur. Examples of boundary conditions, explored by
other studies, include: memory age (Alberini & Milekic 2002, Hardt & Nader, 2009; Tronson
& Taylor, 2007; Morris & Wang, 2010), intensity of conditioning (Frankland et al., 2004)
and level of extinction memory consolidation (Eisenberg, Kabilo, Bernam & Dudai 2003;
Pedreira & Maldonado, 2003).
According to Debeic, LeDoux & Nader (2003), extinction can be thought of as a
boundary condition: reconsolidation only occurs when extinction is not induced (Eisenberg et
al.2003, Nader 2003). It has been suggested that only a few or weak retrieval trials, that do
not result in extinction, lead to reconsolidation (Eisenberg et al., 2003, Pedreira &
Maldonado, 2003; Berlau et al 2000; Frankland et al 2004, Fischer et al. 2004).
Our reactivation sessions occurred over a number of days and showed a gradual
decrease in the operant behavioural repertoire. Furthermore, it might be proposed that, after a
prolonged time of no reward availability in either context, animals are no longer able to
remember which environment was associated with reward, and that the responding at test
reflects spontaneous recovery of the extinguished responding (Rescorla 2004). Moreover, it
has been suggested that consolidated memories might be triggered for reconsolidation as part
of memory updating i.e. when the current information is discrepant to the stored memory
trace (Morris &Wang 2010). Earlier studies show that in animal studies novelty detection is
high in new contexts (Barnes, Good, Honey, McGregor & Staal, 2007). We can assume that
short reactivation periods would increase the novelty detection during reactivation sessions
and so potentially elicit memory updating.
Another possible reason for a failure to show reconsolidation in this experiment is the
lack of emotional arousal during training and extinction. Mueller & Cahill (2010) assert that
nor-adrenaline strengthens consolidation in both appetitive and aversive learning via β-
receptor signalling. On a neuronal level, nor-adrenaline increases excitability which in turn
enhances synaptic plasticity. Administration of adrenaline has been shown to enhance
memory consolidation in animals and humans (Gold, van Buskirk, & Haycock, 1977).
Furthermore, context induced sucrose seeking has been shown to be disrupted by β-
adrenergic antagonists (Diegaarde, Schoffelmeer, & De Vries 2006). Previous research has
shown that anxious individuals are more sensitive to context and have greater contextual
awareness than calmer counterparts (Grillon et al., 2002). As Monfils et al., (2009) study was
done using aversive/shock conditioning this would have lead to a stress response and, thus,
nor-adrenaline release. The paradigm used in this study used no aversive stimuli, meaning no
Page 30
11510
30
Temporal Reconsolidation of Appetitive Behaviour
stress, and consequently no adrenaline or β-adrenergic response was induced; potentially
explaining why reconsolidation failed to take place.
One final criticism of reconsolidation research in general is that there is no universal
reactivation protocol which satisfactorily induces plasticity of consolidated memories (Nader
2007). Therefore, we can assume that slight parametric differences could influence
disruption of reconsolidation (Tronson & Taylor 2006). It would be beneficial for a
procedural criterion to be amalgamated with those of Lewis et al. (1968). Furthermore,
research exploring the effect of nor-adrenaline on reconsolidation (potentially Yohimbine
which induces a stress response) would be invaluable; particularly for research exploring
anxiety disorders. Finally, it is unclear whether reconsolidation represents a loss of memory
or difficulty with retrieval (Nader & Einarsson, 2010). Wang, de Oliveira Alvares and Nader
(2009) suggest that in order to truly change memory we must first understand the inhibitory
effect of boundary conditions on a molecular level, and thus, be able to harness them
therapeutically.
In conclusion, although this study failed to explicitly demonstrate reconsolidation of
appetitive behaviours it provides valuable insight into areas of future research and
mechanisms that, potentially, mediate reconsolidation.
Page 31
11510
31
Temporal Reconsolidation of Appetitive Behaviour
References:
Abel, T. & Lattel, K. M. (2004) Behavioural impairments caused by injections of the protein
synthesis inhibitor anisomycin after contextual retrieval reverse with time
Proceedings of the National Academy of Sciences of the United States of America,
101, 13 4667–4672
Acosta, J., Boynton, F. Kirschner, K.& Neisewander, J. (2005) Stimulation of 5-HT1B
receptors decreases cocaine- and sucrose-seeking behaviour Pharmacology,
Biochemistry and Behaviour, 80, 297–307
Aicardi, Burattini, Janak & Zironi (2006) Context is a Trigger for Relapse to Alcohol
Behavioural Brain Research, 167,150–155
Akirav, I., Marou, M.& Raizel, H. (2006) Enhancement of Conditioned Fear Extinction by
Infusion of the GABAAagonist Muscimol into the Rat Prefrontal Cortex and
Amygdala European Journal of Neuroscience,23,3, 758–764,
Alberini, C. M& Milekic, M. H. (2002) Temporally graded requirement for protein synthesis
following memory reactivation. Neuron 36, 521–525
Alonso J, Angermeyer MC, Bernert S, Bruffaerts R, Brugha IS, Bryson H, de Girolamo G, de
Graaf R, Demyttenaere K, Gasquet I, Haro JM, Katz SJ, Kessler RC, Kovess V,
Lepine JR, Ormel J, Polidori G, Russo LJ, Vilagut G, Almansa J, rbabzadeh-
Bouchez S, Autonell J, Bernal M, Buist-Bouwman MA, Codony M, Domingo-
Salvany A, Ferrer M, Joo SS, Martinez- Alonso M, Matschinger H, Mazzi F,
Morgan Z, Morosini R, Palacin C, Romera B, Taub N, ,lebergh WAM.
(2004)Prevalence of mental disorders in Europe: results from the European Study of
the Epidemiology of Mental Disorders (ESEMeD) project. Acta Psychiatrica
Scandinavica 109: 21–27
Page 32
11510
32
Temporal Reconsolidation of Appetitive Behaviour
Anokhin, K. V., Tiunova, A. A., & Rose, S. P. R. (2002). Reminder Effects- Reconsolidation
or Retrieval Deficit? Pharmacological Dissection with Protein Synthesis Inhibitors
Following Reminder for a Passive-Avoidance Task in Young Chicks. European
Journal of Neuroscience, 15(11), 1759-1765.
Avena, N. M., Hoebel, B.G. and Rada, P (2005) Daily Bingeing on sugar repeatedly releases
Dopamine into the accumbens shell , Neuroscience 134 737-744
Barnes P., Good MA, Honey R.C., McGregor, A. & Staal V (2007) Context- but not
familiarity-dependent forms of object recognition are impaired following excitotoxic
hippocampal lesions in rats. Behavioural Neuroscience 121:218–23
Bernstein GA. & Victor AM (2009) Anxiety disorders and posttraumatic stress disorder
update. Psychiatric Clinics of North America 32: 57– 69
Berlau, D. McGaugh, J.L, Power, A. and Steward, O. (2006) Anisomycin infused into the
hippocampus fails to block “reconsolidation” but impairs extinction: The role of re-
exposure duration Learning and Memory 13:27-34
Bjorklund, A., Fischer, W., Gage F.H., Peterson, G.M., Varon, S., Wictorin, K. &
Williams, L.R.(1986) Continuous infusion of nerve growth factor prevents basal
forebrain neuronal death after fimbria fornix transaction Proceedings of the
National Academy of Science of the United States of America , 88, 23 9281-9285
Bouton, M. E. (1984). Differential Control by Context in the Inflation and Reinstatement
Paradigms. Journal of Experimental Psychology: Animal Behaviour Processes, 10,
56-74.
Bouton, M.E., (1988) context and ambiguity in the extinction of emotional leaning:
Implications for exposure therapy Behaviour Research and Therapy,26, 2, 137- 139
Bouton, M. E., (1993) Context, time, and memory retrieval in the interference paradigms of
Pavlovian learning. Psychological Bulletin ,14, 1; 80-99
Page 33
11510
33
Temporal Reconsolidation of Appetitive Behaviour
Bouton ME.(2002) Context, ambiguity, and unlearning: sources of relapse after behavioral
extinction. Biological Psychiatry;52:976–86
Bouton, M.E., Vurbic, D., and Woods, A. (2006) D-Cycloserine facilitates context-specific
fear extinction learning Neurobiology of Learning and Memory 90, 3, 504-510
Bouton ME (2011) Learning and the persistence of appetite: Extinction and the motivation to
eat and overeat Physiology & Behavior 103 51–58
Bouton, M. E, & Bolles. R. C. (1979). Contextual Control of the Extinction of Conditioned
Fear. Learning and Motivation, 10(4), 445-466.
Bouton, M.E & King, D., (1983) Contextual Control of the Extinction of Conditioned Fear:
Tests for the Associative Value of the Context Journal of Experimental Psychology,
Animal Behavior Processes, 9, 3, 248-265
Bouton, M.E., Gutiérrez, Moody & Zilski (2006), Extinction in multiple contexts does not
necessarily make extinction less vulnerable to relapse Behaviour Research and
Therapy; 44, 983–994
Bouton, M.E., and Moody, E. (2004) Memory processes in classical conditioning
Neuroscience and Biobehavioral Reviews 28, 663–674
Bouton, M. E., & Ricker, S. T. (1994). Renewal of Extinguished Responding in a Second
Context. Animal Learning and Behavior, 22(3), 317-324.
Bouton, M.E. &Swartzentruber, D. (1986) Associative and Occasion-Setting Properties of
Contexts Participating in a Pavlovian Discrimination Journal of Experimental
Psychology: Animal Behavior Processes, 12, 333-350
Page 34
11510
34
Temporal Reconsolidation of Appetitive Behaviour
Bouton, M.E. &Swartzentruber, D. (1991) Source of Relapse after extinction in Pavlovian
and Instrumental Learning Clinical Psychology Rmm, ,. 11, 123-140,
Bossert, J.M., Gray, S.M., Lu, L., Shaham, Y., (2006) Activation of Group II Metabotropic
Glutamate Receptors in the Nucleus Accumbens Shell Attenuates Context-Induced
Relapse to Heroin Seeking Neuropsychopharmacology 31, 2197–2209
Bruce SE, Yonkers KA, Otto MW, Eisen JL, Weisberg RB, Pagano M, Shea MT, KellerMB
(2005) Influence of psychiatric comorbidity on recovery and recurrence in
generalized anxiety disorder, social phobia, and panic disorder: a 12-year
prospective study. American Journal of Psychiatry 162:1179 –1187.
Cahill, S. and Mueller, D. (2010) Noradrenergic modulation of extinction learning and
exposure therapy Behavioural Brain Research 208; 1–11
Chevere, I., Josselyn, H., Kida,S., Kogan,J., Masushige, S., Peña de Ortiz, S., & Silva A.
(2002) CREB required for the stability of new and reactivated fear memories.
Nature Neuroscience; 5: 348–355.
Chiu WT, Demler O, Kessler RC, Merikangas KR, Walters EE.(2005) Prevalence, severity,
and comorbidity of 12-month DSM-IV disorders in the National Comorbidity
Survey Replication. Archives of General Psychiatry 62: 617–627
Collingwood, Jamieson, Macdonald, Pudney & Tinsley (2005) Measuring the harm from
illegal drugs using the Drug Harm Index. Home Office Online Report 24/05
Conklin, C. A., & Tiffany, S. T. (2001). The Impact of Imagining Personalized Versus
Standardizes Urge Scenarios on Cigarette Craving and Automatic Reactivity.
Experimental and Clinical Psychopharmacology, 9(4), 399-408
Contoreggi, C., Kimes, A.S., London, E.D., Grant, S., Liu, X., Margolin, A., Newlin, D.B.,
Phillips, R.L. (1996) Activation of memory circuits during cue-elicited cocaine
craving Proceedings of the National Academy of Sciences of the United States of
America 93, 21 12040-12045
Page 35
11510
35
Temporal Reconsolidation of Appetitive Behaviour
Crombag, H., Grimm, J. W., & Shaham, Y. (2002). Effect of Dopamine Receptor
Antagonists on Renewal of Cocaine Seeking by Reexposure to Drug-Associated
Contextual Cues. Neuropsychopharmacology, 27, 1007-1016.
Crombag, H. S., & Shaham, Y. (2002). Renewal of Drug Seeking by Contextual Cues After
Prolonged Extinction in Rats. Behavioral Neuroscience, 116, 169-173
Dankiewicz LA, Gracy KN, Koob GF, Weiss F (2000). Heroinspecific stimuli reinstate
operant heroin-seeking behavior in rats after prolonged extinction. Pharmacology
Biochemical Behaviour 65: 489–494.
Dębiec, J., Doyère, V., Nader, K., & LeDoux, J. E. (2006). Directly Reactivated, But Not
Indirectly Reactivated, Memories Undergo Reconsolidation in the Amygdala.
Proceedings of the National Academy of Sciences of the United States of America,
103(9), 3428-3433.
Debiec, J., LeDoux, J.E., and Nader, K. (2002). Cellular and systems reconsolidation in the
hippocampus. Neuron 36, 527–538
Denniston JC Gunther LM, Miller RR (1998). Conducting exposure treatment in multiple
contexts can prevent relapse. Behavioural Research Therapy 36: 75–91.
Department of Health and the National Treatment Agency (2010) Statistics from the National
Drug Treatment Monitoring System (NDTMS) 1 April 2009 – 31 March 2010
Diergaarde, L., Schoffelmeer, A.N.M., and De Vries, T.J. (2006) Adrenoreceptor mediated
inhibition of long-term reward-related memory reconsolidation. Behavioural Brain
Research. 170: 333–336.
Dudai, Y. (2004) The Neurobiology of Consolidations or how stable is the engram? Annual
Review in Psychology. 55:51–86
Page 36
11510
36
Temporal Reconsolidation of Appetitive Behaviour
Dudai, Y., and Morris, R.G.M. (2000). To consolidate or not to consolidate: what are the
questions? In: Brain, Perception, Memory. Advances in Cognitive Sciences, J.J.
Bulhuis, ed. (Oxford: Oxford University Press), pp. 149–162
Duvarci S, LeDoux JE, Nader K. (2008) De novo mRNA synthesis is required for both
consolidation and reconsolidation of fear memories in the amygdala. Learning and
Memory 15: 747–755,
Einarsson, E.Ö. & Nader, K. (2010) A bridge over troubled water: reconsolidation as a link
between cognitive and neuroscientific memory research traditions Annual review of
psychology, 61, 141-67
Eisenberg, M., Kobilo, T., Berman, D. E., & Dubai, Y. (2003). Stability of Retrieved
Memory: Inverse Correlation with Trace Dominance. Science, 301(5636), 1102-
1104.
Eisenhardt, D., Menzel, R. & Stollhoff,. (2008) One retrieval trial induces reconsolidation in
an appetitive learning paradigm in honeybees (Apis mellifera) Neurobiology of
Learning and Memory 89, 419–425
Ettenberg A and McFarland K, (1997). Reinstatement of drug-seeking behavior produced by
heroin-predictive environmental stimuli. Psychopharmacology 131: 86–92.
Everitt, B.J,. Milton, A.M. & Lee, J.L. (2006) Reconsolidation and Extinction of Conditioned
Fear: Inhibition and Potentiation, The Journal of Neuroscience, 26(39) 10051-10056
Fischer A, Sananbenesi F, Schrick C, Spiess J, Radulovic J. (2004) Distinct roles of
hippocampal de novo protein synthesis and actin rearrangement in extinction of
contextual fear. The Journal of Neuroscience, 24:1962–66
Page 37
11510
37
Temporal Reconsolidation of Appetitive Behaviour
Frankland, P.W.& Bontempi, B. (2005) The organization of recent and remote memories.
National Review of Neuroscience, 6:119–130
Frankland, P., Josselyn, .S., Kida, S Masushige, S., Silva, A.& Suzuki, A., (2004) Memory
Reconsolidation and Extinction Have Distinct Temporal and Biochemical Signatures
The Journal of Neuroscience, 24(20):4787– 4795
Freeman, F.M., Rose, S.P.R. and Scholey A.B. (1995). Two Time Windows of Anisomycin-
Induced Amnesia for Passive Avoidance Training in the Day-Old Chick,
Neurobiology of Learning and Memory , 63, 3, 291-295
Glickman, S. (1961) Perseverative neural processes and consolidation of the memory trace.
Psychological Bulletin, , 58(3), 218-233
Gold, P.E., van Buskirk, R., and Haycock, J.W. 1977. Effects of post-training epinephrine
injections on retention of avoidance training in mice. Behaviour and Biology. 20:
197–204.
Gomez,R..Hardt,O, Hupbach,A.R.& Nadel.L. (2007).Reconsolidation of episodic memories:
a subtle reminder triggers integration of new information. Learning and Memory.
14: 47–53.
.
Gordon, W. C. & Spear, N. E. (1973).The effects of strychnine on recently acquired and
reactivated passive avoidance memories. Physiology of Behaviour. 10, 1071–1075
Gordon, l., Tinsley, l., Godfrey, C. and Parrott, S. (2006) the economic and social costs of
Class a drug use in england and Wales, 2003/04, In Singleton, N., Murray, R. and
Tinsley, L. (eds) „Measuring different aspects of problem drug use: methodological
developments.‟ Home Office Online Report 16/06
Grillon, C. (2002) Startle reactivity and anxiety disorders: aversive conditioning, context, and
neurobiology Biological psychiatry, 52, 10. 958-975
Page 38
11510
38
Temporal Reconsolidation of Appetitive Behaviour
Kelley, A. E., & Berridge, K. C. (2002). The Neuroscience of Natural Rewards: Relevance to
Addictive Drugs. The Journal of Neuroscience, 22(9), 3306-3311.
Lewis, D., R.R. Miller, and J.R. Misanin. 1968. Control of retrograde amnesia. Journal of
Comparative & Physiological Psychology 66: 48–52.
Mactutus, C.F., R.L. Smith, and D.C. Riccio. 1980. Extending the ACTH-induced memory
reactiviation in an amnestic paradigm. Physiology and Behaviour 24: 541–546.
Mark, R.F. and Watts, M.E. (1971) Drug Inhibition of Memory Formation in Chickens. II.
Short-Term Memory Proceedings of the Royal Society ,. 178 no. 1053 455-464
McGaugh JL. 1966. Time-dependent processes in memory storage. Science 153:1351–58
McGaugh, J. L. (2000) Memory--a Century of Consolidation Science 14, ,. 287 no. 5451 pp.
248-251
Nader K,. & Hardt, O. (2009).A single standard for memory: the case for reconsolidation.
National Review Neuroscience 10: 224–234,
McLellan, A. T., & McKay, J. R. (1998). Components of Successful Treatment Programmes:
Lessons from the Research Literature. In A. W. Graham & T. K. Schultz (Eds.),
Principles of Addiction Medicine (pp.327-343): American Society of Addiction
Medicine.
Misanin JR, Miller RR, Lewis DJ (1968) Retrograde amnesia produced by electroconvulsive
shock after reactivation of a consolidated memory trace. Science 160:554 –555.
Monfils, M-H., Cowansage, K. K., Klann, E., & LeDoux, J. E. (2009). Extinction-
Reconsolidation Boundaries: Key to Persistent Attenuation of Fear Memories.
Science, 324(5929), 951-955
Page 39
11510
39
Temporal Reconsolidation of Appetitive Behaviour
Morris and Wang (2010) Hippocampal-neocoritcak interactions in memory formation,
consolidation and reconsolidation, Annual Review of Psychology, 62: 49-79
Nader, K. (2003). Memory Traces Unbound. Trends in Neurosciences, 26(2), 65-72.
Nader, K. (2007) A single Standard for Memory; the case for Reconsolidation Debates in
Neuroscience , 1 , 1 2-16
Nader, K., Schafe, G. E., & LeDoux, J. E. (2000). Fear Memories Require Protein Synthesis
in the Amygdala for Reconsolidation After Retrieval. Nature, 406, 722-726
National Treatment Agency Media Release (2009) Moves to provide greater protection to
children living with drug addicts
Nelson JB. (2002) Context specificity of excitation and inhibition in ambiguous stimuli.
Learning and Motivation;33:284–310.
Olausson,P., Quinn, J., Taylor, J. & Torregrossa, M. (2009) Targeting extinction and
reconsolidation mechanisms to combat the impact of drug cues on addiction
Neuropharmacology 56 186–195
Pavlov, I. P. (1927). Conditioned Reflexes. Oxford, UK: Oxford University Press.
Pedreira, M. E., Pérez-Cuesta, L. M., & Maldonado, H. (2002). Reactivation and
Reconsolidation of Long-Term Memory in the Crab Chasmagnathus: Protein
Synthesis Requirement and Mediation by NMDA-Type Glutamatergic Receptors.
The Journal of Neuroscience, 22(18), 8305-8311
Pederia, M.E. &H.Maldonado. (2003). Protein synthesis subserves reconsolidation or
extinction depending on reminder duration. Neuron 38: 863–869.
Quirk GJ, Russo GK, Barron JL, Lebron K: (2000) The role of ventromedial prefrontal
cortex in the recovery of extinguished fear. Journal of Neuroscince, 20:6225-6231.
Page 40
11510
40
Temporal Reconsolidation of Appetitive Behaviour
Rescorla, R. (2001). Experimental extinction. In R. R. Mowrer, & S. B. Klein (Eds.),
Handbook of contemporary learning theories (pp. 119–154). Mahwah, NJ: Erlbaum.
Rescorla RA. (2004) Spontaneous recovery. Learning and Memory;11:501–9.
Robinson and Yager (2010) Cue-induced reinstatement of food seeking in rats that differ in
their propensity to attribute incentive salience to food cues, Behavioural Brain
Research 214 30–34
Rose, J. and Rankin, C. (2006) B locking Memory Reconsolidation Reverses Memory-
Associated Changes in Glutamate Receptor Expression The Journal of
Neuroscience, 8, 26(45): 11582-11587
Sara, S.J. 1973. Recovery from hypoxia and ECS induced amnesia after a single exposure to
the training environment. Physiology and Behaviour 9: 85–89.
Sara, S. (2000). Retrieval and reconsolidation: toward a neurobiology of remembering.
Learning and Memory. 7, 73–84.
Schafe, G. E., & LeDoux, J. E. (2000). Memory Consolidation of Auditory Pavlovian Fear
Conditioning Requires Protein Synthesis and Protein Kinase A in the Amygdala. The
Journal of Neuroscience, 20:RC96, 1-5.
Schiller ,D. , Vicentic, A., Monfil, M., Paré, D., Quirck, G. And Herry, C. (2010) Erasing
Fear Memories with Extinction Training The Journal of Neuroscience,
30(45): 14993-14997
Schiller, D., Levy, I., Niv, Y., LeDoux, J., Phelps, E. (2008) From Fear to Safety and Back:
Reversal of Fear in the Human Brain, The Journal of Neuroscience, 28(45): 11517-
11525
Page 41
11510
41
Temporal Reconsolidation of Appetitive Behaviour
Stewart J, de Wit H, Eikelboom R (April 1984). "Role of unconditioned and conditioned
drug effects in the self-administration of opiates and stimulants". Psychological
Review 91 (2): 251–68
Tronson, N.C & Taylor, J.R (2006) molecular mechanisms of memory reconsolidation,
Nature reviews Neuroscience 8 262-275
Volkow, N. D., Wang, G-J., Telang, F., Fowler, J.S., Logan, J., Childress, A. R, Jayne, M.,
Ma, Y., & Wong, C. (2006) Cocaine Cues and Dopamine in Dorsal Striatum:
Mechanism of Craving in Cocaine Addiction The Journal of Neuroscience,
26(24):6583-6588
Walker, M. P., Brakefield, T., Hobson, J. A. & Stickgold, R., (2003) Dissociable stages of
human memory consolidation and reconsolidation, Nature 425, 616–620
Wang, S.H., L. de Oliveira Alvares & K. Nader. 2009. Cellular and systems mechanisms of
memory strength as a constraint on auditory fear reconsolidation. Nature
Neuroscience. 12: 905–912.
Weisberg RB.(2009) Overview of generalized anxiety disorder: epidemiology, presentation,
and course. Journal of Clinical Psychiatry 70: 4 –9
Page 42
11510
42
Temporal Reconsolidation of Appetitive Behaviour
SCHOOL OF PSYCHOLOGY
ETHICS FORM FOR RESEARCH WITH ANIMALS PART A
Your name
Kate Doran
Supervisor
Hans Crombag
Project Title
Using Reconsolidation to Enhance
Contextual Extinction
Degree programme (for example BSc,
MRes)
Psychology BSc
Please answer the following questions: In addition to indicating “yes” or “no” for each question, please type in additional
explanatory information where appropriate
1. Please specify the animal species and number to be observed
(please add rows as required).
Species Name Common Name Wild or Captive Number
Mus musculus Mice Captive 16
* if numbers are not known please indicate an estimate or expected number
Page 43
11510
43
Temporal Reconsolidation of Appetitive Behaviour
2. Does any of the species listed above have special protection/endangered
status?
NO
If yes, provide details of the permits/permissions that have been
sought/obtained:
3. Where will the study be carried out (please provide details of the location of the
study, or state that it will be carried out in the University’s animal holding
facility):
The study will be carried out at the University of Sussex Animal Research Laboratory.
Where appropriate, I have contacted the owner/ manager of the location and
I have attached evidence to confirm their approval for the project.
4. Observation studies with minor interference: Can you confirm that: “Animals are
not interacted with or, where there is interaction, it would not be expected to
compromise the animal’s welfare any more than normal handling, feeding etc.
There is no pain, suffering or discomfort involved.”
YES
If the answer to 4 is “no” go to:
question 5 for work with wild or domestic animals
question 6 for work with laboratory animals.
Page 44
11510
44
Temporal Reconsolidation of Appetitive Behaviour
5. Studies involving wild or domestic animals: Do you or your supervisor believe
there are special ethical issues with the proposed study that you would like
reviewed by the ethics committee?
NO
If yes, what are they?
6. Studies involving laboratory animals: Can you confirm that the work is covered
by a project license under the animals scientific procedures act 1986?
YES
If yes, please specify which procedures require a project license?
Variation in the composition, constituents, quantity and/or availability of the diet
and/or drinking water which may cause pain, suffering, distress or lasting harm.
If yes, can you confirm that an individual who holds a personal license
under the animals scientific procedures act 1986 will be carrying out these
procedures for you?
Page 45
11510
45
Temporal Reconsolidation of Appetitive Behaviour
YES
IF YOUR STUDY INVOLVES BOTH HUMANS AND ANIMALS (for example , you
are monitoring human responses to animal behaviour) you must also complete the
ethics form for research with human participants, which is available at:
http://www.sussex.ac.uk/psychology/1-3-14.html
Page 46
11510
46
Temporal Reconsolidation of Appetitive Behaviour
DEPARTMENT OF PSYCHOLOGY
ETHICS FORM FOR RESEARCH WITH ANIMALS PART B
Summary sheet for projects: Please complete each section.
Title:
Give your project a title, even though you may decide to change it later
The Effect of Reconsolidation on Contextual Extinction.
Investigators:
Your name
Kate Doran
Your email address
[email protected]
Supervisor
Hans Crombag
Background (no more than 150 words):
Explain the aims of your project, and refer very briefly to relevant previous research.
Provide a rationale for your research, and explain what contribution it is likely to
make. State your hypotheses, with an appropriate justification.
Page 47
11510
47
Temporal Reconsolidation of Appetitive Behaviour
The aim of the experiment is to establish the extent to which reconsolidation can be
used to enhance extinction of a behaviour in multiple contexts. Research (Crombag &
Shaham, 2002; Crombag, Grimm, & Shaham, 2002; Bossert, Liu, Lu, & Shaham, 2004) has
shown that when re-exposed to a drug-associated context, rats reengage in drug-seeking
behaviour for a wide array of drugs. However, in regards to drug-taking context, this
suggests that any behavioural extinction that occurs in the rehabilitation clinic will only be
relevant to that context leaving the original drug-taking behaviour unaffected in previous
drug-taking contexts (Rescorla, 2001), or even a novel context (Gunther, Denniston, &
Miller, 1998).
Recent research (Monfils, Cowansage, Klann, & LeDoux, 2009) has suggested that
using reconsolidation prior to an extinction trial (within 6 hours after the isolated retrieval
trial [Nader, Schafe, & LeDoux, 2000]) enhances extinction so that the behaviour is
permanently altered (contrasting with standard extinction which leaves the original memory
unmodified [Pavlov, 1927]). If this finding can be applied to drug rehabilitation contexts, this
enhanced extinction could be used to reduce relapse when addicts return the old drug-
taking context (Childress, Hole, Ehrman, Robbins, McLellan, & O’Brien, 1993) or a novel
context. Currently, reconsolidation research has investigated areas such as fear (Monfils,
Cowansage, Klann, & LeDoux, 2009; Nader, 2003) and taste aversion (Eisenberg, Kobilo,
Berman, & Dubai, 2003) but, to our knowledge, an appetitive study has yet to be conducted.
A study into natural rewards (such as food) could enhance understanding and application of
findings to non-natural rewards (for example addictive drugs.) (Kelley & Berridge, 2002).
Based on previous research (Monfils, Cowansage, Klann, & LeDoux, 2009; See Crombag,
Bossert, Koya, & Shaham, 2008 for review) we hypothesise that those mice experiencing an
isolated retrieval trial and then receiving the extinction session will re-evaluate the value of
the CS leading to increased extinction being displayed in the previous conditioning setting.
Method summary: a) Describe your study population/subjects and study site/housing conditions.
Page 48
11510
48
Temporal Reconsolidation of Appetitive Behaviour
The animals used in this study would be 16 Mus musculus, mice. They would be maintained
on a normal 24hour cycle consisting of 12 hours light and darkness; experiments occurred in
the day portion of the cycle. In hours outside of experimentation the animals would be
housed in groups of 2-4 littermates. The animals would be food deprived throughout the
study; their weight being maintained at 90% of their initial body weight. Animals would
have access to water was ad libitum for the throughout the experiment.
b) What is your design?
The design of the experiment is a 2-way between –within measures design.
c) Give a description of any materials.
During the experimental portion the mice will be trained in 3 discrminably different contexts
(skinner boxes). The three different contexts would be provided by three sets of operant
chambers. In the following aspects these boxes would be the same: each Skinner box would
be constructed of unpainted metal walls and measuring 27 x 25.8 x 30.4 cm in length, width,
and height.
The boxes from context A, B and C will differ in terms of their tactile, visual and olfactory
background stimuli. The skinner boxes used for context A would be scented with vinegar,
the lighting would be altered using red acetate. The floor of each skinner box would be lined
with woodchips. Conversely, in context B the boxes would be scented using “vapour rub”,
the bulb would be left uncovered and the grids of the skinner box would also be left
uncovered.
Page 49
11510
49
Temporal Reconsolidation of Appetitive Behaviour
The Skinner boxes from Context C will be scented using essential oils, the light will be
coloured using green acetate and the floor of the Skinner box will be lined with a plastic
sheet. In order to further differentiate the environments the mice will be placed in each
context at different times of the day. Context A exposure will be in the morning, context B
around Noon and Context C in the afternoon.
These contexts will be referred to as A, B and C with A being the context in which training
and self-administration of sucrose took place, B as the extinction and C as the
alternative/testing context.
20% (w/v) sucrose solution made fresh daily
Eight mice per experimental group
A computer running (insert details of programs here) and interfaced with the conditioning
chambers
Conditioning chambers
Plastic syringes and hypodermic needles
Retractable levers
Liquid delivery receptacle
Auditory stimulus modules to present CS tone
Light stimulus to present alternative CS tone
Decibel (dB) meter for calibration of auditory stimuli (10 dB above background)
d) Provide a brief description of the procedure you will use.
Page 50
11510
50
Temporal Reconsolidation of Appetitive Behaviour
Firstly, the mice would engage in a brief (20 minutes for 2 days) period of receptacle
approach training. This involves the mice being placed in a training context [context A] with
a syringe pump which is activated on a RI30 schedule [min=15seconds, max=45seconds]
which is set to deliver a small volume of sucrose solution [approximately 50µl] into the
receptacle cup.
For the next eleven days the Mice would be placed in Context A and receive 20
conditioning trials to which a cue light (this would be counterbalanced with an auditory tone
for half of the mice) was paired with 10% sucrose delivery (CS+) and 20 trials in which an
auditory cue (again this would be counterbalanced using a light cue for half of the mice)
was presented with no sucrose delivered (CS-). Cues would be presented in 10second
durations. Throughout these (CS+) trials, 50µl sucrose solution was infused into the food-
cup receptacle 5 second succeeding cue onset.
The population of mice will be halved on day 16. These groups would both take part
in a retrieval trial in Context B. The first group, RET10min, would then be returned to their
homes cages for 10minutes before the 60 minute extinction session in Context B was
initiated. Following ach lever press of the right lever (counterbalanced for half of the mice)
the CS+ was presented in the absence of any sucrose solution being delivered. The second
group, RET6hr, will undergo the retrieval trial and then be returned to their homes cages for
6 hours before they take part in the 60 minute extinction session.
The mice will then be returned to their home cages and 24 hours later all mice would
be tested for extinction in Context C (allowed to lever press in Context A) in short 20 minute
trials.
e) Include a statement on ethical issues. – Even if there are none, you need to demonstrate
that you have thought about this, and justify your conclusion that there are no issues.
Page 51
11510
51
Temporal Reconsolidation of Appetitive Behaviour
Variation in the composition, constituents, quantity and/or availability of the diet and/or
drinking water which may cause pain, suffering, distress or lasting harm.
The justification for the use of animals in this study is the extensive research into appetitive
behaviour using classical or Pavlovian conditioning that has already taken place.. The effect
of context within this framework has also been researched at length. This provides a
invaluable baseline for comparison, which simply does not exist for human participants,
allowing us to examine the full effect of a relatively new methodology.
An ethical issue, concerned with our study, will be the stress induced due to our procedure.
This stress will be caused by two factors. Firstly, the mice will be stressed due to handling
the animals when transferring them from their housing to the conditioning and testing
contexts. Secondly in some mouse strains or in mice with genetic mutations limiting food
can be poorly tolerated. Therefore, it will very important to monitor the health of the
animals carefully throughout the period of food restriction particularly during the first few
days. Mice are also known to show a level of neophobia when exposed to novel food (Amico
et al., 2005). Therefore, to reduce this it would be beneficial to expose the animals to the
reinforcer initially in their home environment.
f) Indicate briefly how you will analyse your data.
A between-Within subjects (independent measures) two-tailed t-test and a within subjects
two-tailed t-test will be used to analyse the data.