Page 1
Treatment of murine cerebral malaria by artemisone in combination with 1
conventional anti-malarial drugs: anti-plasmodial effects and immune 2
responses 3
4 5 W. Armand Guiguemde,a Nicholas H. Hunt,b Jintao Guo,b Annael 6
Marciano,c Richard K. Haynes,d Julie Clark,a R. Kiplin Guy,a Jacob 7
Golenserb,c,* 8
9
Department of Chemical Biology and Therapeutics, St Jude Children’s 10
Research Hospital, Memphis, Tennessee, USAa; Department of Pathology 11
and Bosch Institute, The University of Sydney, Sydney, Australiab; 12
Department of Microbiology and Molecular Genetics, The Kuvin Center 13
for the Research of Tropical and Infectious Diseases, The Hebrew 14
University of Jerusalem, Jerusalem, Israelc; Centre of Excellence for 15
Pharmaceutical Sciences, North-West University, Potchefstroom, South 16
Africad 17
18
*Corresponding author. Tel.: 97226758090; fax: 97226757425. 19
E-mail address: [email protected] 20
21
22
The decreasing effectiveness of anti-malarial therapy due to drug 23
resistance necessitates constant efforts to develop new drugs. Artemisinin 24
derivatives are the most recent drugs that have been introduced and are 25
considered the first line of treatment but there are already indications of 26
Plasmodium falciparum resistance to artemisinins. Consequently, drug 27
combinations are recommended for prevention of the induction of 28
resistance. The research here demonstrates the effects of novel 29
combinations of the new artemisinin derivative, artemisone, a recently 30
AAC Accepts, published online ahead of print on 9 June 2014Antimicrob. Agents Chemother. doi:10.1128/AAC.01553-13Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 2
described 10-alkylamino artemisinin derivative with improved anti-31
malarial activity and reduced neurotoxicity. We here investigate its ability 32
to kill P. falciparum in a high throughput in vitro assay and to protect mice 33
against lethal cerebral malaria caused by P. berghei ANKA when used 34
alone or in combination with established anti-malarial drugs. Artemisone 35
effects against P. falciparum in vitro were synergistic with halofantrine 36
and mefloquine, and additive with 25 other drugs, including chloroquine 37
and doxycycline. The concentrations of artemisone combinations that were 38
toxic against THP-1 cells in vitro were much higher than their effective 39
anti-malarial concentration. Artemisone, mefloquine, chloroquine or 40
piperaquine given individually mostly protected mice against cerebral 41
malaria caused by P. berghei ANKA but did not prevent parasite 42
recrudescence. Combinations of artemisone with any of the other three 43
drugs did completely cure most mice of malaria. The combination of 44
artemisone and chloroquine decreased the ratio of pro-inflammatory 45
(interferon-γ, tumour necrosis factor) to anti-inflammatory (IL-10, IL-4) 46
cytokines in the plasma of P. berghei-infected mice. Thus artemisone in 47
combinations with other anti-malarial drugs might have a dual action, both 48
killing parasites and limiting the potentially deleterious host inflammatory 49
response. 50
51
Keywords: 52
Malaria 53
Plasmodium 54
Cerebral malaria 55
Anti-malarial drugs 56
Drug combinations 57
Inflammation 58
59
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 3
Running title: 60
Artemisone combinations: effects on CM and immunity 61
62
63
The dwindling arsenal of drugs for treatment of malaria and the need for 64
developing and selecting new ones is a continuing problem that has been 65
extensively discussed. Most drugs that recently have been approved for 66
human use are artemisinin derivatives (1). However, some artemisinin 67
derivatives that have been introduced have already encountered various 68
degrees of resistance and consequently have been used in drug 69
combinations (2, 3, 4). 70
Artemisone is a recent artemisinin derivative, a semi-synthetic 10-71
alkylamino artemisinin that can be synthesized from dihydroartemisinin. It 72
is an attractive drug because, in comparison to current artemisinins, it is 73
not neurotoxic. In in vitro screens it elicits no cytotoxicity towards brain 74
stem cell cultures and neurofilaments at concentrations up to 25 μM, and 75
has no effect on the respiratory chain (5, 6). Lack of neurotoxicity was also 76
verified in various animal screens (7). 77
Artemisone was found to be highly effective in culture against 78
Plasmodium falciparum (8), in vivo against murine cerebral malaria (CM) 79
induced by P. berghei ANKA (4) and against P. falciparum in monkeys 80
(9). It has been used in a Phase IIa clinical trial for non-severe malaria in 81
humans (10). Artemisone can cure Toxoplasma gondii (11) and Neospora 82
caninum (12) in animal models. 83
We recently have shown in a mouse model of CM that artemisone could 84
prevent death even when administered at relatively late stages of cerebral 85
pathogenesis. No parasite resistance to artemisone was detected and co-86
administration of artemisone and chloroquine was more effective than 87
monotherapy with either drug, leading to complete cure (4). These results 88
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 4
suggest the use of artemisone for combination therapy. However, a 89
thorough study is needed to establish the efficacy of additional 90
combinations of artemisone with commercially available anti-malarial 91
drugs. For this purpose we used high throughput in vitro screening against 92
P. falciparum and a reliable CM model (P. berghei ANKA in C57Bl mice) 93
for in vivo validation (2). 94
When choosing a multiple testing procedure for screening combinatorial 95
drug libraries, natural products or any compound reservoir, the results 96
suggesting further investigation or rejection of a candidate drug often 97
ignore a possible significant effect on the outcome of treatment following 98
the use of these drugs: attenuation of immune responses may alleviate 99
clinical symptoms that are caused by immunopathology. In this context, 100
various forms of severe malaria including CM are the result of 101
immunopathology (13). Therefore, immunomodulators represent an 102
interesting new approach to CM treatment. Likewise, Fasudyl, a Rho 103
kinase inhibitor, was suggested as an adjunctive therapeutic agent in the 104
management of severe malaria (14, 15). IDR-1018, an adjunctive anti-105
inflammatory peptide, was partially protective against murine CM (16). 106
Moreover, anti-plasmodial drugs, including artemisinins, may affect 107
immune responses, in addition to exerting direct effect on the parasites 108
(17, 18). 109
In view of growing information on parasites resistant to artemisinin 110
derivatives, malaria treatments now recommended by the World Health 111
Organization are artemisinin-based combination treatments (ACT). These 112
are combinations of an artemisinin derivative and another structurally 113
unrelated and more slowly eliminated antimalarials (19). Such pairings 114
might include drugs that are not effective as a monotherapy but are useful 115
in the combination, for example Malarone® (combined atovaquone and 116
proguanil) is considered a useful malaria therapeutic agent. However there 117
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 5
are indications of resistance to malarone® (20), stressing the need for a 118
constant search for both new individual anti-malarial compounds and drug 119
combinations. Consequently, we decided to examine the effects of 120
artemisone, a recently discovered 10-alkylamino- artemisinin, alone and in 121
drug combinations, in P. falciparum cultures and in a mouse malaria 122
model, including an examination of the effect of a representative drug 123
combination on cytokine responses that are relevant to CM induction. 124
125
MATERIALS AND METHOD 126
Parasites. P. berghei ANKA was maintained in vivo by serial transfer of 127
parasitized erythrocytes (PE) from infected to naïve mice. Experimental 128
mice were infected by intraperitoneal (i.p.) injection of 5×104 PE from 129
peripheral blood of infected donor mice, an inoculum that caused fatal 130
experimental cerebral malaria (ECM) in at least 80% of infected 131
C57BL/6 mice. The link between early death and ECM in mouse models 132
has been discussed previously (2, 4): mice that died at a parasitemia of 133
20% or below, with accompanying neurological symptoms and drastic 134
reductions in body weight and temperature, were considered to have died 135
of ECM, which where possible was confirmed by the presence in the 136
CNS of hemorrhages, edema and intravascular leukocyte accumulation 137
upon histopathological analysis. Untreated mice that did not die from 138
ECM went on to succumb to severe anemia and hyperparasitemia, as has 139
been reported in all other cases where mice are resistant to ECM induced 140
by P. berghei ANKA (21, 22). 141
The 3D7 strain of P. falciparum (purchased from the American Type 142
Culture Collection, ATCC) was grown in culture as specified later. 143
Animals. C57Bl/6 mice (Harlan, Jerusalem, Israel; Animal Resources 144
Centre, Perth, Australia) aged 7-8 weeks were used in all experiments, 8 to 145
10 mice per group (as described). The mice were housed under standard 146
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 6
light and temperature conditions and provided with unlimited access to 147
water and food. All experiments were carried out in accordance with 148
institutional guidelines for animal care, by protocols approved by the 149
Animal Ethical Care Committee of The Hebrew University of Jerusalem 150
and in accordance with the guidelines under the Australian Code of 151
Practice for the Care and Use of Animals for Scientific Purposes and 152
approved by the University of Sydney Animal Ethics Committee. 153
Parasitemia was monitored microscopically in thin blood Giemsa stained 154
smears prepared from tail blood. Clinical score was evaluated and used for 155
scoring disease severity (Scoring Chart (4)). Mice were euthanized when 156
they reached a degree of disease severity that would inevitably have led to 157
their death. 158
Histology. Mice were deeply anaesthetized with isofluorane and sacrificed 159
by terminal intracardial perfusion with 10 ml ice-cold PBS. Organs were 160
removed and fixed overnight in 10% (v/v) neutral buffered formalin. 161
Paraffin embedded tissues were cut into 5-7 μm slices, deparaffinated, and 162
stained with hematoxylin and eosin before coverslipping. 163
Drugs. Dihydroartemisinin (DHA) and artesunate were purchased from 164
the Kunming Pharmaceutical Corporation. Artemisone was synthesized 165
from DHA and purified by flash column chromatography, followed by 166
recrystallization according to the procedure previously reported (8). 167
Piperaquine was donated by Cipla Ltd, Mumbai, India. It was dissolved in 168
double distilled water, adjusted to pH 3.5 with HCl and injected in 100 μl. 169
Dimethyl sulfoxide (DMSO) and chloroquine diphosphate were purchased 170
from Sigma-Aldrich, Ltd. All artemisinin derivatives were prepared in 171
DMSO according to the required dosage and administered in a volume of 172
20 μl by intraperitoneal injection. Chloroquine diphosphate (Sigma) was 173
dissolved in PBS and administered in a 50 μl volume by intraperitoneal 174
injection. Mefloquine (Sigma) was dissolved in DMSO and used in the 175
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 7
same way as the artemisinin derivatives. Drug structures have been shown 176
elsewhere (2). Artemisone and chloroquine were injected 6 times, twice on 177
days 6, 7 and 8. Piperaquine was injected once a day due to its longer half-178
life in mice (23). 179
Drug toxicity. Drug toxicity was determined in THP-1 cells (human 180
monocytes, ATCC, USA) as previously described (2) using Alamar blue 181
viability assay in 96-well flat-bottom plates (Nunc). The Alamar blue 182
method has been questioned concerning the use of redox active drugs 183
and/or adherent or fast growing cells. However, in our system where the 184
cells were exposed to the drugs before the addition of the indicator and the 185
cells were not adherent or fast growing, it is unlikely that there is a 186
significant aberration of the results. Percent growth inhibition of the cells 187
was calculated according to the equation: % inhibition = [(Fluorescence 188
control – Fluorescence test) / (Fluorescence control)] x 100. 189
Automated screening of in vitro anti-plasmodial activity. Automated 190
screening has been described elsewhere (2). Briefly, parasites were 191
incubated for 72 h and parasitemia was estimated by using a DNA dye 192
solution (SYBR green). Three-fold serial dilutions resulting in 10 different 193
concentrations were examined. Tests were run in triplicates in two 194
independent runs to determine IC50 against the 3D7 P. falciparum strain 195
for each drug. Synergy or antagonism was determined using Bliss 196
independence (24) and fractional inhibition concentration (FIC50)(25). 197
Drug combination were defined as non-additive when more than one 198
binary combination effect lay outside the predicted effect (Bliss); and 199
when more than one FIC50 was outside the 95% confidence interval of a 200
control FIC50 (using a compound against itself as a control). 201
Analysis of in vitro anti-plasmodial activity. The effects of piperaquine 202
and its combinations with artemisone against P. falciparum were evaluated 203
manually in vitro by a luciferin-luciferase bioluminescence assay (2). 204
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 8
Briefly, we used erythrocytic stages of P. falciparum stably expressing the 205
luciferase gene by the hrp2 promoter from a chromosomal locus (Pf:LUC) 206
in 96 well flat-bottom sterile plates. After 48 hours medium was removed 207
and the erythrocytes lysed by lysis buffer of the Bright-Glo Luciferase 208
Assay System (100µl/well). 50µl of the Bright-Glo substrate was added to 209
each well and the luminescence measured by a luminometer (Fluoroskan 210
Ascent FL, Thermo). 211
ELISA assay. ELISA assays for murine plasma cytokine analysis of 212
interleukin(IL)-10, interferon (IFN)γ, IL-4 and tumor necrosis factor 213
(TNF) were purchased from Biolegend, Israel. C57Bl/6 mice were injected 214
with P. berghei ANKA and treated with different drugs at days 6, 7 and 8 215
post infection (PI). On days 0, 5, 8, and 12 PI, mice were sacrificed and 216
blood samples were collected in heparin for plasma cytokine analysis of 217
IL-10, IFNγ, IL4 and TNF by ELISA assay according to the 218
manufacturer's instructions. 219
Statistics. When comparing parasitemia, p values were calculated using 220
Students t-test; for analysis of survival curves, the Kaplan-Meier test was 221
employed. In both cases, values below 0.05 were considered significant. 222
223
RESULTS 224
Evaluation of drug combinations in P. falciparum. Drug susceptibility 225
assays were performed in P. falciparum cultures, using HTS techniques. 226
To evaluate synergistic combinations, we used two orthogonal methods 227
(FIC50 and Bliss independence) and the combination of artemisone with 228
itself was used to define the additive background (Fig. 1). A summary of 229
the overall results of examining combinations of artemisone with 230
conventional anti-malarial drugs is shown in Fig. 2. Artemisone in 231
combination with most antimalarials currently used in the clinic was 232
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 9
additive, except with halofantrine and mefloquine where synergy was 233
identified. No antagonism was identified. 234
Manual analysis of in vitro anti-plasmodial activity revealed identical 235
results (data not shown). Artemisone-piperaquine combinations that were 236
examined only manually depicted synergistic effect. 237
Cytotoxicity assays. Drug toxicity was determined against THP-1 cells 238
using the Alamar blue viability assay. For positive control (maximal 239
growth inhibition) we used KuRei, a cell inhibitor (26). The assay was 240
performed in triplicates and the standard deviation of the activities was 241
within 10% of the mean for each drug. The IC50 values of artemisone and 242
chloroquine were >282 and 313 nM, respectively; the IC50 values for 243
mefloquine and piperaquine were >12.1 and 1.5 μM, respectively. These 244
data, and the results of drug combinations (Fig. 3), should be compared to 245
the effects of the drugs on P. falciparum cultures (2). The anti-plasmodial 246
effects were evident at much lower concentrations (about 80 fold lower). It 247
is obvious that the drug combinations had no synergistic cytotoxicity. 248
The effects of artemisone combination therapy on P. berghey infected 249
mice. Drug concentrations suitable for in vivo combination experiments 250
were selected based on the HTS screening and previous results (2, 4): 251
those that induced a temporary reduction in parasitemia and clinical score. 252
Combination therapy of infected mice with artemisone and either 253
mefloquine, chloroquine or piperaquine was applied to try to prevent the 254
cerebral symptoms and achieve complete cure. Fig. 4, 5a, 5b depict 255
resulting parasitemias, survival curves, body temperatures, weight and 256
clinical scores. Abrupt decline in temperature and loss of weight were 257
reflecting initial symptoms of the disease that were alleviated by the 258
drugs. However, all mice treated with the individual drugs, despite an 259
initial delay in parasitemia increase, eventually succumbed to the disease. 260
In contrast, the combinations prevented CM, delayed recrudescence and 261
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 10
prevented death in most mice. Artemisone-mefloquine combinations were 262
superior to the individual drugs but not as efficient as artemisone 263
combinations with chloroquine or piperaquine. 264
Histology. Brains were assessed at different intervals post infection (at 265
least four matching animals from each group). The results paralleled the 266
clinical score: in control untreated mice, hemorrhages and intravascular 267
leukocyte accumulation were abundant, but in drug treated animals 268
depicting low clinical score there were no such manifestations (data not 269
shown). 270
Analysis of cytokines involved in development of CM. The purpose of 271
this part of the study was to examine a possible correlation between the 272
outcome of the treatment and the immunological status of the treated 273
animals. We examined the effects of different therapies on the plasma 274
levels of representative cytokines in the malaria-infected mice treated at 275
days 6, 7 and 8 PI with artemisone (2x10mg/kg), chloroquine (2x15mg/kg) 276
or their combination. These concentrations were chosen to allow maximal 277
anti-malarial activity (without approaching toxic levels). TNF and IFNγ 278
represent pro-inflammatory cytokines (Th1 type). IL-4 and IL-10 represent 279
anti-inflammatory cytokines (Th2 type). All injected animals had shown 280
early CM symptoms (coat ruffled, hunched, slight decrease in body weight 281
and temperature) before treatment was started (e.g. ruffled coat, wobbly 282
gait; scoring chart,(4)). Cytokine plasma levels were estimated using 283
ELISA on days 0, 5, 8 and 12 PI. Control groups included uninfected 284
untreated, uninfected treated and infected untreated mice. 285
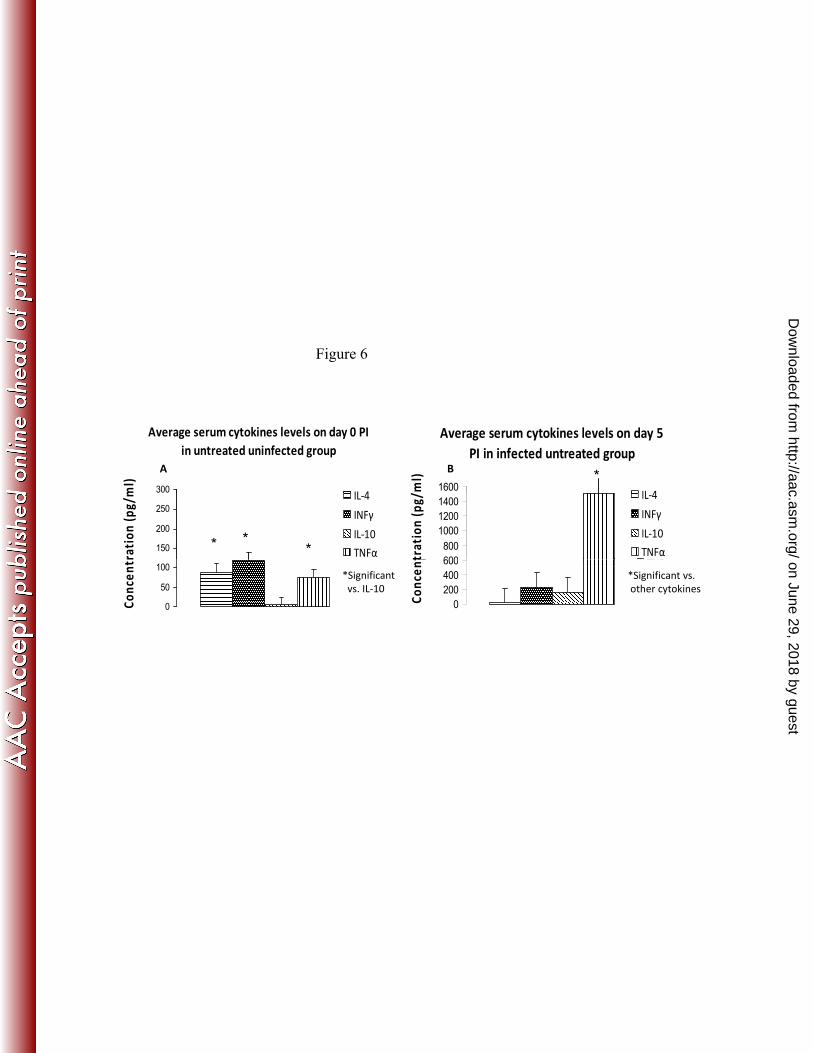
On day 0 (Fig. 6A), in uninfected untreated mice, the IL-10 level in the 286
plasma was lower than that of the other cytokines. On day 5 (Fig. 6B), in 287
infected untreated mice, the most striking events were a significant 288
increase in TNF and decrease in IL-4 levels, in comparison both with the 289
other cytokines and with their level in the control group on day 0. 290
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 11
On day 8 (Fig. 7), in chloroquine and combination treated mice, the plasma 291
level of the anti-inflammatory cytokine IL-4 was higher than in infected 292
untreated mice. The IL-10 level had increased in all treated groups 293
compared to that in untreated mice, where it was below detection. Plasma 294
IFNγ levels were comparable in all groups, although there was a slightly 295
lower level in the combination treated mice. It is interesting that infected 296
untreated mice displayed much higher TNF in plasma compared to all the 297
treated mice. These results are consistent with the involvement of pro-298
inflammatory cytokines such as IFNγ and TNF in CM. 299
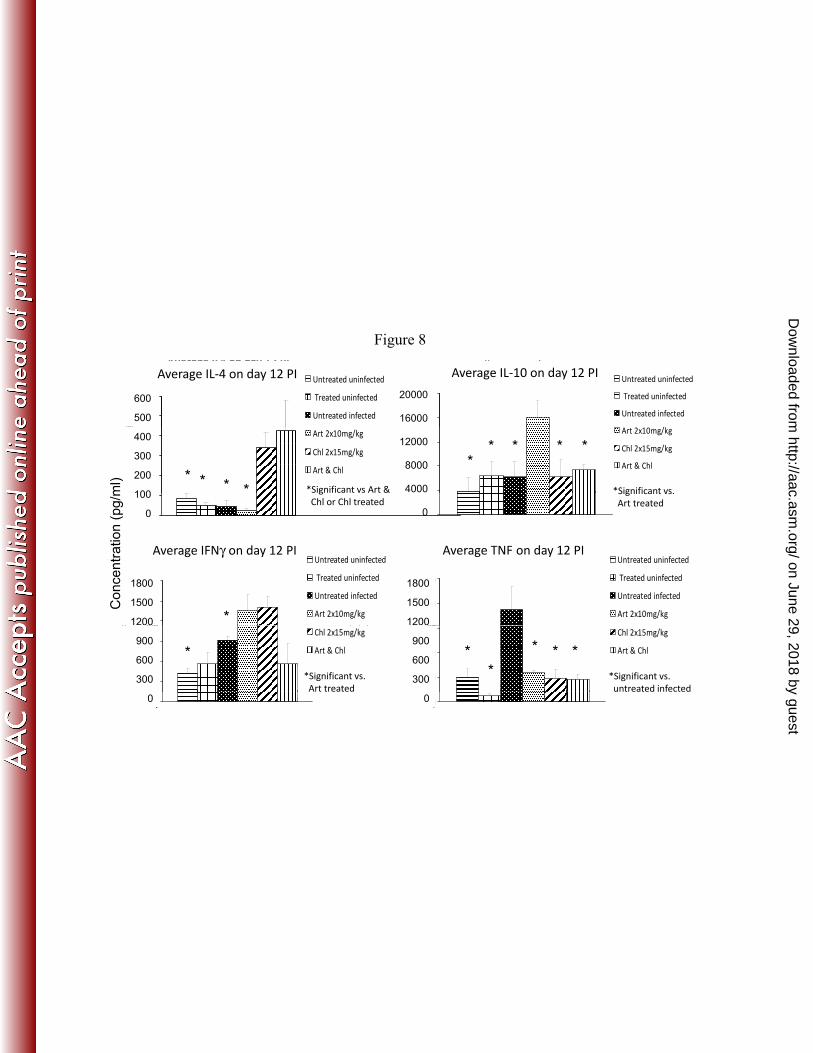
Fig. 8 depicts the plasma cytokine levels on day 12. Higher plasma IL-4 300
was seen in the chloroquine and combination treated mice, compared to 301
the other groups. Artemisone treated mice displayed higher plasma IL-10 302
level compared to the other groups. In the combination treated mice, there 303
was lower plasma IFNγ relative to mice treated by individual drugs, TNF 304
in plasma had declined significantly in all drug treated mice, while in 305
infected untreated mice it rose significantly. 306
307
DISCUSSION 308
Drug combinations are the current strategy in malaria treatment to contain 309
resistance to individual drugs or postpone its induction. Vivas et al. (27) 310
found by using isobolograms obtained with susceptible 3D7 and drug-311
resistant K1 P. falciparum strains at the IC50 level, slight antagonistic 312
trends between artemisone and chloroquine, amodiaquine, tafenoquine, 313
atovaquone or pyrimethamine. Additive to slight synergistic interactions 314
were seen with artemisone and mefloquine, lumefantrine or quinine. In 315
vitro automated screening allows for the examination of multiple drug 316
combinations in order to identify potential partner drugs to be used in the 317
clinic, or to avoid combinations that would be antagonistic (Fig. 1, 2). We 318
tested artemisone in combination with most antimalarials currently used in 319
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 12
the clinic and found that it was additive with all drugs tested except 320
halofantrine and mefloquine, where synergy was identified (Fig. 2). Most 321
importantly, no antagonism was identified, implying that artemisone could 322
be paired with current antimalarials for further in vivo combination studies. 323
We selected combinations of artemisone with mefloquine, piperaquine or 324
chloroquine for in vivo evaluations. This decision was partly based on the 325
in vitro synergistic effect of artemisone with mefloquine and additive 326
effect with piperaquine or chloroquine (Fig. 2), and also our experience 327
where piperaquine or chloroquine, in combination with artemiside (another 328
alkyl-amino-artemisinin) were found additive in vitro, and synergistic in 329
vivo (2). Vivas et al. (27) found slight antagonism between the in vitro 330
effects of artemisone and chloroquine, while we found additivity. The 331
difference in these in vitro results is not great and may have its origin in 332
the different experimental conditions. However, both groups report 333
synergistic in vivo effects. This is important because often in vitro results 334
are not translated into the in vivo domain (28). It is especially interesting 335
to examine artemisone (which is already in clinical trials) in combination 336
with chloroquine. The latter drug played an important role in malaria 337
eradication but it remains attractive because of its low cost, and the 338
possibility that when combined with other drugs, treatment success will be 339
significantly increased. Also, it is desirable to elucidate the use of 340
combination of drugs that are less effective alone because of parasite 341
resistance but are efficient together. A clear example is provided by 342
Malarone that comprises the combination of atovaquone and proguanil 343
(resistance to the individual drugs in this combination has been 344
unequivocally demonstrated). 345
Schmuck et al. (29) suggest that artemisone is embryotoxic. However, in 346
that paper there are no comparative experimental results - it focuses on 347
artemisone alone, ignoring other artemisinin derivatives. In a direct in vivo 348
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 13
comparison of artemisone and artesunate, artemisone was not found 349
neurotoxic in contrast with artesunate (7). Moreover, artesunate in low 350
doses was embryotoxic in rats (30). In addition, a fatal artesunate toxicity 351
was reported in a child (31). Yet, artesunate alone or in drug combinations 352
is considered a first line anti-malarial treatment. Indeed, in a limited study 353
involving pregnant women there was no evidence of artesunate 354
embriotoxicity (32). Artemisone is more effective than artesunate in vitro 355
and against murine models of malaria (4, 27). In our experiments, 356
artemisone combinations with the tested anti-malarial drugs were not 357
toxic, both in vitro (Fig. 3) and in vivo. The highest concentrations of 358
artemisone (10mg/kg), in drug combinations, when injected twice a day 359
for three days, had no visible effects on the mice. Overall, artemisone 360
might be considered for treatment of malaria especially in drug 361
combinations, where toxicity of individual drugs can be reduced. 362
In long term experiments we found that mice treated with the individual 363
drugs, despite an initial delay in parasitemia increase, eventually 364
succumbed to the disease. However, most mice treated with artemisone 365
combination therapy were completely cured (Fig. 4, 5). Vivas et al. (27) 366
described some in vivo artemisone–drug interactions in a rodent model by 367
using the Peter’s four day test. This method is inadequate for estimation of 368
the effect of drug treatment on severe malaria (e.g. CM), where 369
pathogenesis is most pronounced a week or more after infection and 370
pathology (or lack of pathology) is the result of a prolonged innate 371
immune response and early acquired immunity (4, 33). 372
The in vivo experiments were performed in a reliable mouse model of CM 373
(22, 34). The underlying mechanism of CM pathogenesis remains 374
incompletely understood but there is widespread agreement that cytokines 375
(and other components of the immune system) have a crucial role in CM 376
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 14
and severe malaria in general, in mice and in humans. An imbalance 377
between the release of pro-inflammatory and anti-inflammatory cytokines 378
has been associated with the central nervous system dysfunction found in 379
human and experimental CM (35). The current study in C57BL/6 mice 380
infected with P. berghei ANKA aimed to investigate whether the anti-381
plasmodial effect and prevention of CM correlate with an immunological 382
shift, expressed in plasma cytokine levels. The inflammatory cytokines 383
TNF, IFN-γ (Th1-type) and anti-inflammatory IL-4 and IL-10 (Th2-type), 384
were determined in plasma by ELISA. 385
In clinical studies of human CM, elevated serum IFN-γ is seen in acute 386
malaria infection in South-East Asian (36) and African (37) patients. 387
Murine and human studies strongly support a role for IFN-γ and 388
downstream immune system processes in the pathogenesis of CM (38, 39, 389
40). Evidence suggesting that TNF is another key element in the 390
pathogenesis of experimental CM has been reviewed extensively (41, 42). 391
This includes the observation of high serum levels of TNF at the onset of 392
CM (43) and the prevention of the neurological syndrome when TNF 393
levels are low (44, 45). Anti-inflammatory cytokines such as IL-10 seem to 394
have a host-protective role in murine malaria. For example, the clinical 395
scores of IL-10 deficient, infected mice were significantly higher when 396
compared with WT mice. In addition, in a susceptible mouse strain, 397
administration of IL-10 gave some degree of protection against CM 398
induced by P. berghei ANKA (46, 47). IL-4 is an anti-inflammatory 399
cytokine. Changes in plasma IL-4 have been reported to correlate with 400
severe malaria (48) but, conversely, increased levels of IL-4 have been 401
linked with reduced immunopathological symptoms (49, 50). 402
Anti-malarial drugs may induce immunological alterations in treated 403
patients and animals. Artemisinins can produce immunosuppression by 404
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 15
down regulating various cytokines of both the innate and acquired immune 405
systems. They induce an anti-inflammatory effect, neutropenia, and 406
reduction in macrophage number and functions, which may produce 407
immunosuppression. Artemisinins also have the ability to induce 408
immunosuppression by inhibiting delayed-type hypersensitivity, 409
lymphocyte proliferation and a rise in antibody level (51). Immune 410
deviation caused by chloroquine and other anti-malarial drugs has been 411
reported (52, 53). 412
We found that the plasma levels of the pro-inflammatory cytokines TNF 413
and IFN-γ increased in P. berghei ANKA-infected mice (Fig. 6 - 8). In 414
parallel with CM reduction, there was an attenuated increase in TNF 415
plasma levels in P. berghei ANKA-infected mice, after treatment with 416
chloroquine, artemisone or the combination of the two. Reduction of IFN-417
γ was achieved with the drug combination. These results agree with the 418
hypothesis that the CM syndrome is a result of a shift in the balance of 419
Th1/Th2 responses toward Th1. While many immune components 420
(cytokines, chemokines, effector cells) and metabolic pathways are 421
involved in processes leading to the expression of CM, one drug may 422
affect only some of these components while another one may affect others. 423
Thus, judicious selection of combination therapy may reduce parasitemia 424
by direct actions on the parasite and also inhibit the severe symptoms of 425
malaria through immunomodulatory actions. In general, all drug treatments 426
reduced inflammatory cytokines and increased anti-inflammatory 427
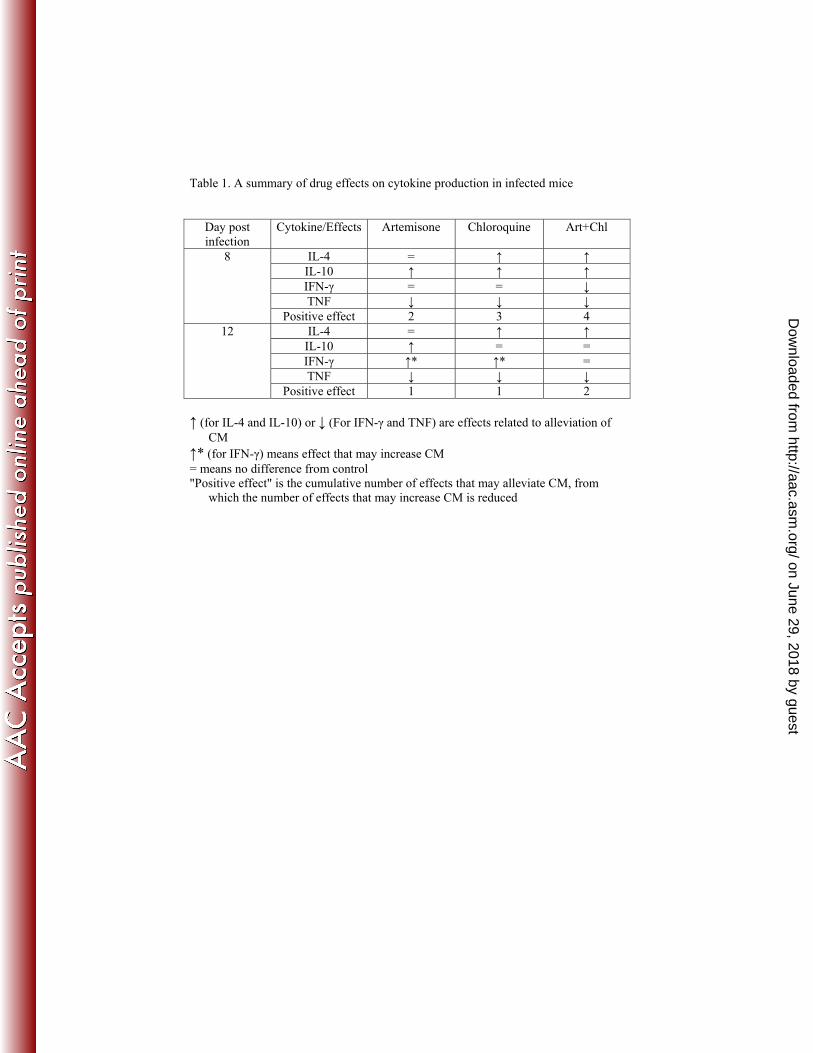
cytokines (positive effects). However, when examining carefully the data 428
of day 8 and 12 post infection, the positive effects induced by the drug 429
combination was more pronounced (Fig. 6-8; Table 1). 430
Overall, experiments are needed to determine how drugs that are 431
used in combination influence each other in terms of immunomodulation, 432
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 16
toxicity, pharmacokinetic, pharmacodynamic and even pharmacogenetic 433
aspects. 434
435
ACKNOWLEDGEMENTS 436
This work was supported by a grant from the Australian National 437
Health and Medical Research Council (to NH), the Sir Zelman 438
Cowen Universities Fund, the Israel Science Foundation and the 439
Deutsche Forschungsgemeinschaft (DFG) (to JG). We thank Cipla 440
Ltd, Mumbai, India for the kind donation of piperaquine. 441
442
REFERENCES 443
1. Dondorp, A.M., Yeung, S., White, L., Nguon, C., Day, N.P., Socheat, D., von 444 Seidlein, L. 2010. Artemisinin resistance: current status and scenarios for containment. 445 Nat Rev Microbiol. 8, 272-280. 446 447 2. Guo, J., Guiguemde, A.W., Bentura-Marciano, A., Clark, J., Haynes, R.K., 448 Chan, W.C., Wong, H.N., Hunt, N.H., Guy, R.K., Golenser, J. 2012. Synthesis of 449 artemiside and its effects in combination with conventional drugs against severe murine 450 malaria. Antimicrob Agents Ch. 56, 163-173. 451 452 3. Phyo, A.P., Nkhoma, S., Stepniewska, K., Ashley, E.A., Nair, S., McGready, R., 453 ler Moo, C., Al-Saai, S., Dondorp, A.M., Lwin, K.M., Singhasivanon, P., Day, 454 N.P., White, N.J.,Anderson, T.J., Nosten, F. 2012. Emergence of artemisinin-455 resistant malaria on the western border of Thailand: a longitudinal study. Lancet 379, 456 1960-1966. 457 458 4. Waknine-Grinberg, J.H., Hunt, N., Bentura-Marciano, A., McQuillan, J.A., 459 Chan, H.W., Chan, W.C., Barenholz, Y., Haynes, R.K., Golenser, J. 2010a. 460 Artemisone effective against murine cerebral malaria. Malaria J. 9, 227. 461 462 5. Schmuck, G., Haynes, R.K. 2000. Establishment of an in vitro screening model for 463 neurodegeneration induced by antimalarial drugs of the artemisinin-type. Neurotox 464 Res. 2, 37-49. 465 466 6. Schmuck, G., Roehrdanz, E., Haynes, R.K., Kahl, R. 2002. Neurotoxic mode of 467 action of artemisinin. Antimicrob Agents Ch. 46, 821-827. 468 469 7. Schmuck, G.; Temerowski, M.; Haynes, R. K.; Fugmann, B. 2003. Identification 470 of a non-neurotoxic artemisinin derivative in vivo and in vitro. Antimicrob Agents Ch. 471 3, 35-47 472 473
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 17
8. Haynes, R.K., Fugmann, B., Stetter, J., Rieckmann, K., Heilmann, H.D., Chan, 474 H.W., Cheung, M.K., Lam, W.L., Wong, H.N., Croft, S.L., Vivas, L., Rattray, L., 475 Stewart, L., Peters, W., Robinson, B,L,, Edstein, M.D., Kotecka, B., Kyle, D.E., 476 Beckermann, B., Gerisch, M., Radtke, M., Schmuck, G., Steinke, W., Wollborn, 477 U., Schmeer, K., Römer, A. 2006. Artemisone - A highly active antimalarial drug of 478 the artemisinin class. Angew Chem-Int Edit. 45, 2082-2088. 479
9. Obaldia, N. 3rd, Kotecka, B.M., Edstein, M.D., Haynes, R.K., Fugmann, B., 480 Kyle, D.E., Rieckmann, K.H. 2009. Evaluation of artemisone combinations in Aotus 481 monkeys infected with P. falciparum. Antimicrob Agents Ch. 53, 3592-3594. 482
10. Nagelschmitz, J., Voith, B., Wensing, G., Roemer, A., Fugmann, B., Haynes, 483 R.K., M. Kotecka, B.M., Karl H. Rieckmann, and Edstein, M.D. 2008. First 484 assessment in humans of the safety, tolerability, pharmacokinetics, and ex vivo 485 pharmacodynamic antimalarial activity of the new artemisinin derivative artemisone. 486 Antimicrob Agents Ch. 52, 3085–3091. 487 488 11. Dunay, R., Chan, W.C., Haynes, R.K., Sibley, L.D. 2009. Artemisone and 489 artemiside control acute and reactivated toxoplasmosis in a murine model. Antimicrob 490 Agents Ch. 53, 4450–4456. 491 492 12. Mazuz, M.L., Haynes, R., Shkap, V., Fish, L., Wollkomirsky, R., Leibovich, B., 493 Molad, T., Savitsky, I., and Golenser, J. 2012. Neospora caninum: in vivo and in 494 vitro treatment with artemisone. Vet Parasitol. 187, 99-104. 495 496 13. Grau, G.E., Craig, A.G. 2012. Cerebral malaria pathogenesis: revisiting parasite 497 and host contributions. Future Microbiol. 7, 291-302. 498 499 14. Taoufiq, Z., Gay, F., Balvanyos, J., Ciceron, L., Tefit, M., Lechat, P., Mazier, 500 D. 2008. Rho kinase inhibition in severe malaria: thwarting parasite-induced collateral 501 damage to endothelia. J Infect Dis. 197, 1062-1073. 502 503 15. Waknine-Grinberg, J.H., McQuillan, J.A., Hunt, N., Ginsburg, H., Golenser, 504 J. 2010b. Modulation of cerebral malaria by fasudil and other immune-modifying 505 compounds. Exp Parasitol. 125, 141-146. 506 507 16. Achtman, A.H., Pilat, S., Law, C.W., Lynn, D.J., Janot, L., Mayer, M,L, Ma, 508 S., Kindrachuk, J., Finlay, B.B., Brinkman, F.S., Smyth, G.K., Hancock, R.E. 509 Schofield, L. 2012. Effective adjunctive therapy by an innate defense regulatory 510 peptide in a preclinical model of severe malaria. Sci Transl Med. 4, 135ra64. 511 512 17. Li, T., Chen, H., Wei, N., Mei, X., Zhang, S., Liu, D.L., Gao, Y., Bai, S.F., Liu, 513 X.G., Zhou, Y.X. 2012. Anti-inflammatory and immunomodulatory mechanisms of 514 artemisinin on contact hypersensitivity. Int immunopharmacol. 12, 144-150. 515 516 18. Wang, Z., Qiu, J., Guo, T.B., Liu, A., Wang, Y., Li, Y., Zhang, J.Z. 2007. Anti-517 inflammatory properties and regulatory mechanism of a novel derivative of artemisinin 518 in experimental autoimmune encephalomyelitis. J Immunol. 179, 5958-5965. 519 520
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 18
19. Nosten, F , White, N.S. 2007. Artemisinin based combination treatment of 521 falciparum malaria. J. Am J Trop Med Hyg. 77, 181-192.31. 522
523 20. Farnert, A., Lindberg, J., Gil, P., Swedberg, G., Berqvist, Y., Thapar, M.M., 524 Lindegardh, N., Berezcky, S., and Bjorkman, A. 2003. Evidence of Plasmodium 525 falciparum malaria resistant to atovaquone and proguanil hydrochloride: case reports. 526 BMJ., 326, 628-629. 527 528 21. Engwerda, C., Belnoue, E., Gruner, A.C., Renia, L. 2005. Experimental models 529 of cerebral malaria, p 103-143. In Langhorne, J. (Ed.), Immunology and 530 Immunopathogenesis of Malaria. Springer-Verlag, Berlin. 531 532 22. Neill, A.L., and Hunt, N.H. 1992. Pathology of fatal and resolving P. berghei CM 533 in mice. Parasitology 105, 165-175. 534 535 23. Moore, B.R., Ilett, K.F., Page-Sharp, M., Jeffrey D. Jago, J.D., Batty, K.T. 536 2009. Piperaquine pharmacodynamics and parasite viability in a murine malaria model. 537 Antimicrob Agents Ch. 53, 2707–2713. 538 539 24. Bliss, C.I. 1999. The toxicity of poisons applied jointly. Ann Appl Biol. 26, 585-540 615. 541 542 25. Canfield, C.J., Pudney, M., Gutteridge, W.E. 1995. Interactions of atovaquone 543 with other antimalarial drugs against Plasmodium falciparum in vitro. Exp Parasitol. 544 80, 373-381. 545 546 26. Reichwald, C., Shimony, O., Dunkel, U., Sacerdoti-Sierra, N., Jaffe, C.L., and 547 Kunick, C. 2008. 2-(3-aryl-3-oxopropen-1-yl)-9-tert-butyl-paullones: a new 548 antileishmanial chemotype. J. Med Chem. 51, 3659-3665. 549 550 27. Vivas, L., Rattray, L., Stewart, L.B., Robinson, B.L., Fugmann, B., Haynes, 551 R.K., Peters, W., Croft, S.L. 2007. Antimalarial efficacy and drug interactions of the 552 novel semi-synthetic endoperoxide artemisone in vitro and in vivo. J Antimicrob 553 Chemother. 59, 658-665. 554 555 28. Golenser, J. Hunt, N. 2013. Neglected Aspects of Drug Discovery - 556 Microbiological Aspects. Curr Clin Pharmacol. 1, 73-80 557 558 29. Schmuck, G., Klaus, A.-M., Krotlinger, F., Langewische, F.W. 2009. 559 Developmental and Reproductive toxicity studies on artemisone. Birth Defects 560 Research. 86, 131–143. 561 562 30. Chung, M-K., Yu, W-J., Lee, J-S., Lee, J-H.. 2013. Embryotoxicity and 563 toxicokinetics of the antimalarial artesunate in rats. Toxicol Res. 29, 27-34. 564 565 31. Camposa, S., de la Cerdac,P., Rivera. A. 2008. Fatal artesunate toxicity in a 566 child. J. Pediatric Infec Dis. 3, 69–75. 567 568 32. McGrady, R., Cho, T., Keo, N.K., Thwai, K.L., Villegas, L., Looareesuwan, 569 S., White, N.J., Nosten, F. 2001. Artemisinin antimalarials in pregnancy: a 570
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 19
prospective treatment study of 539 episodes of multidrug-resistant Plasmodium 571 falciparum. Clin Infect Dis. 33, 2009–2016. 572 573 33. Hunt, N.H., Golenser. J., Chan-Ling, T., Parekh, S., Rae, C., Potter, S., 574
Medana, I.M., Miu, J., Ball, H.J. 2006. Immunopathogenesis of cerebral malaria. Int 575
J Parasitol. 36, 569-582. 576
34. Hunt, N.H., Grau, G.E., Engwerda, C., Barnum, S.R., van der Heyde, H., 577 Hansen, D.S., Schofield, L., and Golenser, J. 2010. Murine cerebral malaria: the 578 whole story. Trends Parasitol. 26, 272-274. 579
35. Torre, D., Speranza, F., Giola, M., Matteelli, A., Tambini, R., Biondi, G. 2002. 580 Role of Th1 and Th2 cytokines in immune response to uncomplicated Plasmodium 581 falciparum malaria. Clin Diagn Lab Immun. 9, 348-351. 582 583 36. Ho, M,, Sexton, M.M., Tongtawe, P., Looareesuwan, S., Suntharasamai, P., 584 Webster H.K. 1995. Interleukin-10 inhibits tumor necrosis factor production but not 585 antigen-specific lymphoproliferation in acute Plasmodium falciparum malaria. J Infect 586 Dis. 172, 838-844. 587 588 37. Ringwald, P., Peyron, F., Vuillez, J.P., Touze J.E., LeBras, J., Deloron, P. 589 1991. Levels of cytokines in plasma during Plasmodium falciparum malaria attacks. J 590 Clin Microbiol. 29, 2076-2078. 591 592
38. Grau GE, Heremans H, Piguet PF, Pointaire P, Lambert PH, Billiau A, 593 Vassalli P. 1989a. Monoclonal antibody against interferon gamma can prevent 594 experimental cerebral malaria and its associated overproduction of tumor necrosis 595 factor. Proc Natl Acad Sci USA. 86, 5572-5574. 596
39. Sanni, L,A,, Thomas, S,R,, Tattam, B,N,, Moore, D,E,, Chaudhri, G, Stocker, 597 R., Hunt, N.H. 1998. Dramatic changes in oxidative tryptophan metabolism along the 598 kynurenine pathway in experimental cerebral and noncerebral malaria. Am J Pathol. 599 152, 611-619. 600 601 40. Yanez, D.M., Manning, D.D., Cooley, A.J., Weidanz, W.P., van der Heyde, 602 H.C. 603 1996. Participation of lymphocyte subpopulations in the pathogenesis of 604 experimental murine cerebral malaria. J. Immunol. 157, 1620–1624. 605 606 41. Clark, I.A., Rockett, K.A. 1994. The cytokine theory of human cerebral malaria. 607 Parasitol Today. 10, 410–412. 608 609 42. Grau, G.E., Piguet, P.F., Vassali, P., Lambert, P.H. 1989b. Tumor necrosis 610 factor and other cytokines in cerebral malaria: experimental and clinical data. Immunol 611 Rev. 112, 49-70. 612
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 20
43. Lyke, K.E., Burges, R., Cissoko, Y., Sangare, L., Dao, M., Diarra, I., Kone, A., 613 Harley, R., Plowe, C.V., Doumbo, O.K., Sztein, M.B. 2004. Serum levels of the 614 proinflammatory cytokines interleukin-1 beta (IL-1β), IL-6, IL-8, IL-10, tumor 615 necrosis factor alpha, and IL-12(p70) in malian children with severe Plasmodium 616 falciparum malaria and matched uncomplicated malaria or healthy controls. Infect. 617 Immun. 72, 5630-5637. 618
44. de Miranda, A.S., Lacerda-Queiroz, N., de Carvalho Vilela, M., Rodrigues, 619 D.H., Rachid, M.A., Quevedo, J., Teixeira, A.L. 2011. Anxiety-like behavior and 620 proinflammatory cytokine levels in the brain of C57BL/6 mice infected with 621 Plasmodium berghei (strain ANKA). Neurosci Lett. 491, 202-206. 622
45. Rudin, W., Eugster, H.P., Bordmann, G., Bonato, J., Müller, M., Yamage, M., 623 Ryffel, B. 1997. Resistance to cerebral malaria in tumor necrosis factor-alpha/beta-624 deficient mice is associated with a reduction of intercellular adhesion molecule-1 up-625 regulation and T helper type 1 response. Am J Pathol. 150, 257-266. 626
46. Golenser J., McQuillan J., Hee L., Mitchell A.J., Hunt NH. 2006. Conventional 627 and experimental treatment of cerebral malaria. Int J Parasitol. 36, 583-593. 628
47. Kossodo,S., Monso, C., Juillard P., Velu, T., Goldmani, M., Grau, G.E. 1997. 629 Interleukin-10 modulates susceptibility in experimental cerebral malaria. Immunology 630 91,536-540. 631
48. Cabantous, S., Poudiougou, B., Oumar, A.A., Traore, A., Barry, A., Vitte, J., 632 Bongrand, P., Marquet, S., Doumbo, O., Dessein, A.J. 2009. Genetic evidence for 633 the aggravation of Plasmodium falciparum malaria by interleukin 4. J Infect Dis. 200, 634 1530-1539. 635 636 49. Angulo, I., and Fresno, M. 2012. Cytokines in the pathogenesis of and protection 637 against malaria. Clin Vaccine Immunol. 9, 1145-1152. 638 639 50. de Kossodo, S., Grau, G.E. 1993. Profiles of cytokine production in relation with 640 susceptibility to cerebral malaria. J Immunol. 151, 4811-4820. 641 642 51. Shakir, L Hussain, M., Javeed, A., Ashraf, M., Riaz, A. 2011. Artemisinins and 643 immune system. Eur J Pharmacol. 668, 6-14. 644 645 52. Fryauff, D.J., Church, L.W., Richards, A.L., Widjaja, H., Mouzin, E., 646 Ratiwayanto, S., Hadiputranto, H., Sutamihardja, M.A., Richie, T.L., Subianto, 647 B.,Tjitra, E., Hoffman, S.L. 1997. Lymphocyte response to tetanus toxoid among 648 Indonesian men immunized with tetanus-diphtheria during extended chloroquine or 649 primaquine prophylaxis. J. Infect Dis. 176, 1644-1648. 650 651 53. Ramos-Avila, A., Ventura-Gallegos, J.L., Zentella-Dehesa, A., Machuca-652 Rodriguez, 653 C., Moreno-Altamirano, M.M., Narvaez, V., Legorreta-Herrera, M. 2007. 654 Immunomodulatory role of chloroquine and pyrimethamine in Plasmodium yoelii 655 17XL infected mice. Scand J Immunol. 65, 54-62. 656 657
658
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 21
Figure legends 659
660
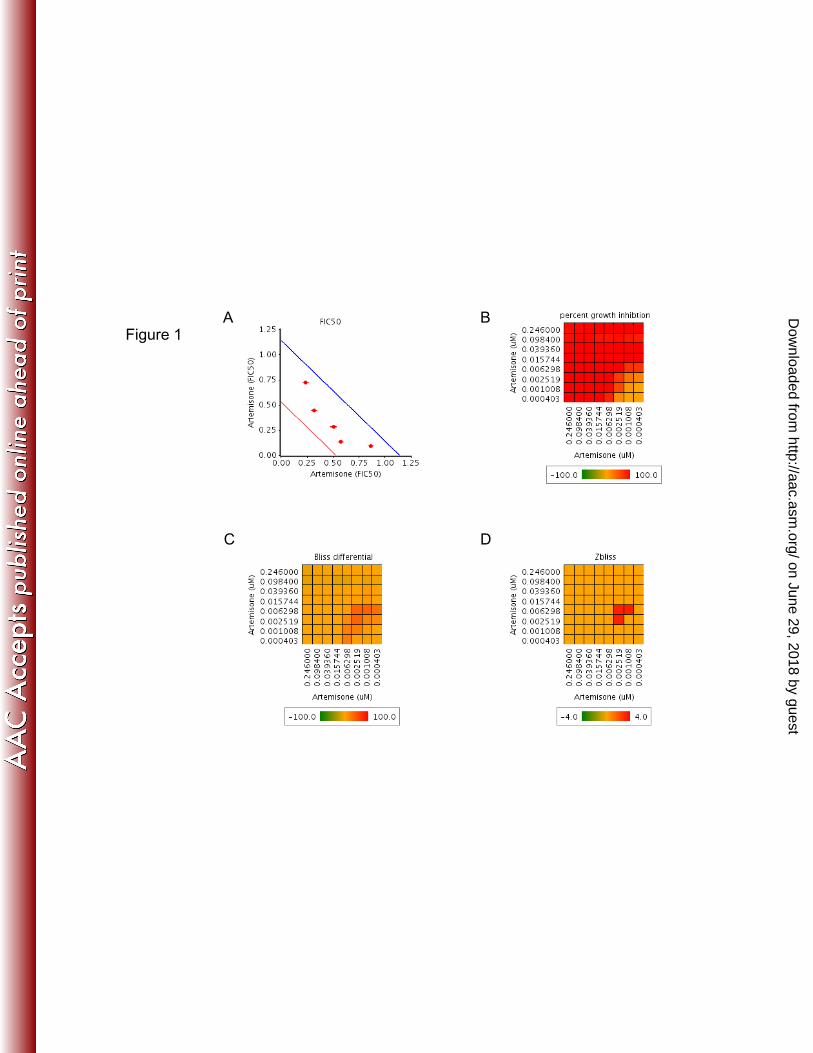
FIG 1. Activity per pair of drug combinations. 661
Artemisone pairwise combinations were examined in vitro. Artemisone 662
combined with itself was used as a control for additivity. (A) Fractional 663
IC50 of artemisone in combination with itself. The surface between the red 664
and the blue lines defines the additivity area. The distance between these 665
two lines equals 95% of the FIC50 confidence interval. (B) Heat map 666
representing the percentage of the growth inhibition of the tested drug 667
combinations. (C) Heat map representing the Bliss differential of the drug 668
combinations, the difference in growth inhibition between observed and 669
predicted values. (D) Zbliss is derived from data shown in panel c using Z 670
scores, considering all Bliss differential values as additive values. Values 671
in panels b, c, and d are averages from four dependent replicates. 672
673
FIG 2. Artemisone pairwise combinations examined in vitro 674
Color coded summary of the pairwise combinations examined by the high 675
throughput system. For each pair, FIC50s and Bliss values were calculated 676
(as shown in Figure 3) and the degree of synergy was estimated according 677
to defined criteria based on Z-scores (Zs). 678
679
FIG 3. Cytotoxicity assays of artemisone combinations with standard 680
antimalarial drugs, chloroquine (Ch), mefloquine (Mef) and 681
piperaquine (P). 682
Drug toxicity was determined against THP-1 cells using Alamar blue 683
viability assay. The numbers in the columns represent the experimental 684
results (average of three). The numbers in brackets are the theoretical 685
additive values. For example, artemisone 1 (Art1, 25nM), chloroquine 2 686
(Chl2, 110nM) and their combination inhibit 13%, 28%, and 38% of 687
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 22
untreated THP-1 growth, respectively. The theoretical additive value is 688
41% (13+28). 689
Compound concentrations in the experiment (*nM, **mM): 690
Art1 25*, Art2 75*, Art3 225* 691
Ch1 37*, Ch2 110*, Ch3 330* 692
Mef1 1.3**, Mef2 4.0**, Mef3 12.0** 693
P1 0.38**, P2 0.75**, P3 1.50** 694
695
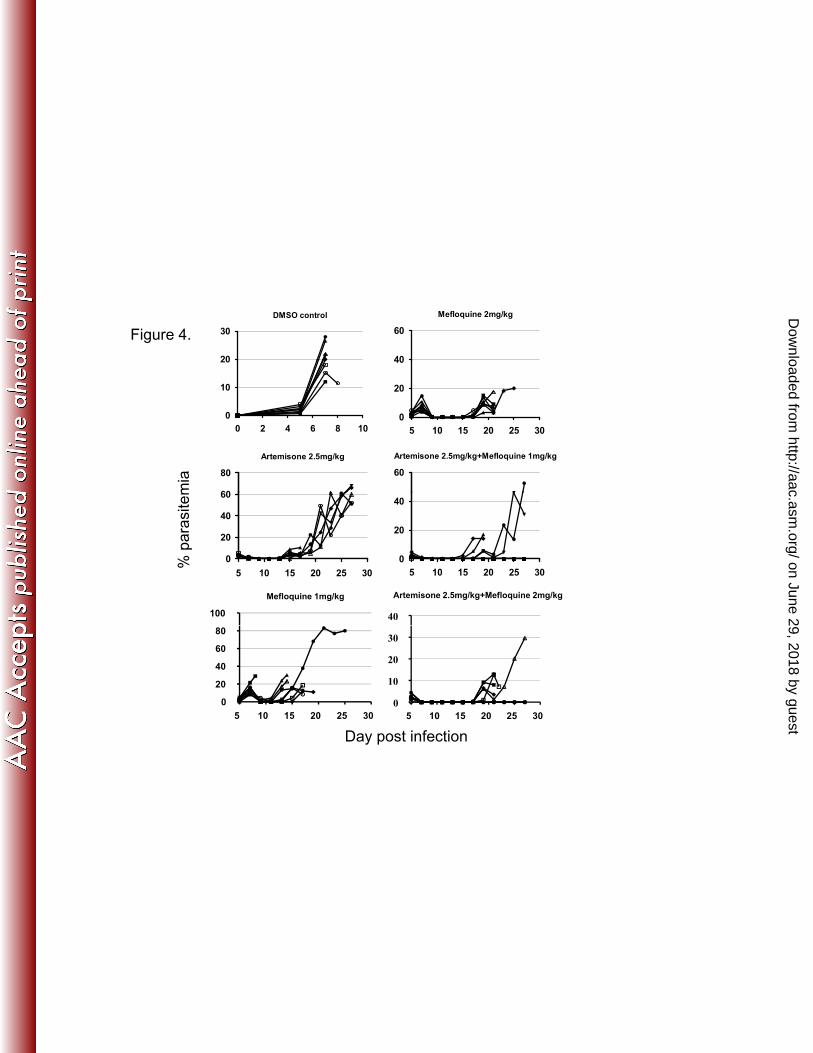
FIG 4. Parasitemias of infected mice, treated with artemisone or 696
mefloquine or their combinations. 697
Each line represents a single mouse. Infected mice were treated twice a 698
day on days 6-8 post infection. N = 8/group. 699
700
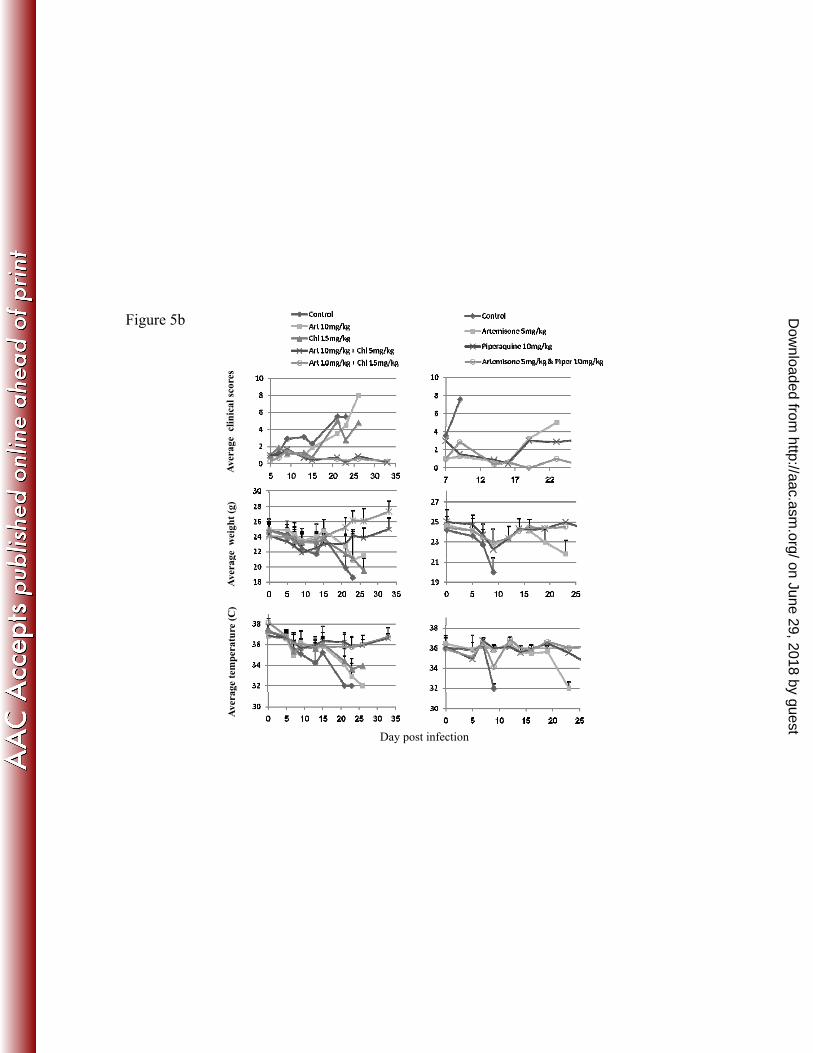
FIG 5. Effect of artemisone, chloroquine and piperaquine on P. 701
berghei ANKA infection. 702
Infected mice were treated on days 6-8 post infection with artemisone (art; 703
twice each day) and chloroquine (chl; twice each day) or artemisone (twice 704
each day) and piperaquine (piper; once per day). 705
5a. Data are parasitemias and survival curves of infected mice. 706
5b. Data are clinical score, weight, and body temperature of infected mice. 707
Values are mean ± SD; n = 8-10/group. 708
709
FIG 6. Plasma cytokine levels in control (A) and infected untreated 710
mice on day 5 (B). 711
Columns and vertical bars represent mean ± SD; n = 8-10/group. 712
713
FIG 7. Effect of artemisone and chloroquine on 714
cytokines in infected mice on day 8 post infection 715
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 23
Plasma cytokine levels in mice treated on days 6-8 (twice each day) 716
post infection by artemisone (Art), chloroquine (Chl) or their 717
combination. Columns and vertical bars represent mean ± SEM; n = 718
8-10/group. 719
720
FIG 8. Effect of artemisone and chloroquine on cytokines 721
in P. berghei ANKA infection on day 12 post infection 722
Plasma cytokine levels in mice treated on days 6-8 (twice each day) 723
post infection by artemisone (Art), chloroquine (Chl) or their 724
combination. Columns and vertical bars represent mean ± SEM; n = 725
8-10/group. 726
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 24
A BFigure 1
C D
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 25
FIG 1. Activity per pair of drug combinations. A t i i i bi ti i d i it A t iArtemisone pairwise combinations were examined in vitro. Artemisone combined with itself was used as a control for additivity. (A) Fractional IC50 of artemisone in combination with itself. The surface between the red and the blue lines defines the additivity area. The distance between ythese two lines equals 95% of the FIC50 confidence interval. (B) Heat map representing the percentage of the growth inhibition of the tested drug combinations. (C) Heat map representing the Bliss differential of the drug combinations the difference in growth inhibition betweenthe drug combinations, the difference in growth inhibition between observed and predicted values. (D) Zbliss is derived from data shown in panel c using Z scores, considering all Bliss differential values as additive values. Values in panels b, c, and d are averages from four p gdependent replicates.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 26
Test drugFigure 2
AdditiveAdditiveSynergistic: one drug combinations has a ZsBlis differential activity >3 and ZsFIC>3
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 27
FIG 2. Artemisone pairwise combinations examined in vitro. Color coded summary of the pairwise combinations examined by theColor coded summary of the pairwise combinations examined by the high throughput system. For each pair, FIC50s and Bliss values were calculated (as shown in Figure 3) and the degree of synergy was estimated according to defined criteria based on Z-scores (Zs). on June 29, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 28
Figure 3.
13 13
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 29
FIG 3. Cytotoxicity assays of artemisone combinations with standardFIG 3. Cytotoxicity assays of artemisone combinations with standard antimalarial drugs, chloroquine (Ch), mefloquine (Mef) and piperaquine (P).Drug toxicity was determined against THP-1 cells using Alamar blue viability assay. The numbers in the columns represent the experimental results (average of three). The n mbers in brackets are the theoretical additi e al es For e ampleThe numbers in brackets are the theoretical additive values. For example, artemisone 1 (Art1, 25nM), chloroquine 2 (Chl2, 110nM) and their combination inhibit 13%, 28%, and 38% of untreated THP-1 growth, respectively. The theoretical additive value is 41% (13+28).( )Compound concentrations in the experiment (*nM, **mM):Art1 25*, Art2 75*, Art3 225*Ch1 37*, Ch2 110*, Ch3 330*Mef1 1 3** Mef2 4 0** Mef3 12 0**Mef1 1.3 , Mef2 4.0 , Mef3 12.0P1 0.38**, P2 0.75**, P3 1.50**
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 30
DMSO control
20
30
Mefloquine 2mg/kg
40
60Figure 4.
0 2 4 6 8 100
10
20
0
20
40
0 2 4 6 8 10
Artemisone 2.5mg/kg
60
80
mia
Artemisone 2.5mg/kg+Mefloquine 1mg/kg
60
5 10 15 20 25 30
0
20
40
60
% p
aras
item
0
20
40
5 10 15 20 25 300%
5 10 15 20 25 300
Artemisone 2.5mg/kg+Mefloquine 2mg/kg
40
Mefloquine 1mg/kg
100
10
20
30
20
40
60
80
Day post infection5 10 15 20 25 30
05 10 15 20 25 30
0
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 31
FIG 4. Parasitemias of infected mice, treated with artemisone orFIG 4. Parasitemias of infected mice, treated with artemisone or mefloquine or their combinations.Each line represents a single mouse. Infected mice were treated twice a day on days 6-8 post infection. N = 8/group. Mortality and survival results
i d i t bl 1are summarized in table 1.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 32
Figure 5a
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 33
FIG 5. Effect of artemisone, chloroquine and piperaquine on P. bergheiANKA infection.
f fInfected mice were treated on days 6-8 post infection with artemisone (art; twice each day) and chloroquine (chl; twice each day) or artemisone (twice each day) and piperaquine (piper; once per day).5a. Data are parasitemias and survival curves of infected mice.5a. Data are parasitemias and survival curves of infected mice.5b. Data are clinical score, weight, and body temperature of infected mice.Values are mean ± SD; n = 8-10/group.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 34
Figure 5b
linic
al sc
ores
Aver
age
cge
wei
ght (
g)e
(C)
Aver
agag
e te
mpe
ratu
reAv
era
Day post infection
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 35
FIG 5. Effect of artemisone, chloroquine and piperaquine on P. bergheiANKA infection.Infected mice were treated on days 6-8 post infection with artemisoneInfected mice were treated on days 6 8 post infection with artemisone (art; twice each day) and chloroquine (chl; twice each day) or artemisone (twice each day) and piperaquine (piper; once per day).5a. Data are parasitemias and survival curves of infected mice.5b D t li i l i ht d b d t t f i f t d5b. Data are clinical score, weight, and body temperature of infected mice.Values are mean ± SD; n = 8-10/group.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 36
Figure 6g
Average serum cytokines levels on day 0 PI in untreated uninfected group
300ml)
IL 4
A
Average serum cytokines levels on day 5 PI in infected untreated group
1600ml)
IL 4ml) *B
150
200
250
rati
on (p
g/m IL-4
INFγ
IL-10
TNFα* *
*600800
100012001400
trat
ion
(pg/
m
IL-4INFγIL-10TNF
IL-4
INFγ
IL-10
TNFαtrat
ion
(pg/
0
50
100
Conc
entr
*Significant vs. IL-10
0200400600
Conc
ent TNFα
Conc
ent
*Significant vs. other cytokines
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 37
FIG 6. Plasma cytokine levels in control (A) and infected untreated y ( )mice on day 5 (B).Columns and vertical bars represent mean ± SD; n = 8-10/group.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 38
Figure 7
Average IL4 on day 8 PI
700
800Infected un
Art 2x10m/ml)
*Infected untreated
/
Average IL-4 on day 8 PI
800700 20000
Infected untreated
Art 2x10mg/kg
20000 Art 2x10mg/kg**20000 Infected untreated
Art 2x10mg/kg
Average IL-10 on day 8 PI
* **
200
300
400
500
600Art 2x10m
Chl 2x15m
Art & Chl
once
ntra
tion
(pg/
*
*Significant vs
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
600500400300200
15000
10000
l)
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
5000
10000
15000g/ g
Chl 2x15mg/kg*
*Significant vs
15000
10000
5000
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
**
0
100
200Co *significant vsuntreated and
Significant vs. untreated and Art
Average INFγ on day 8 PIAverage INFγ on day 8 PI
200100
0
5000
atio
n (p
g/m
0
5000 Art & Chl*Significant vs.
untreated5000
0
Average TNF on day 8 PI
Infected untreated
Art 2x10mg/kg
hl /k
Average INFγ on day 8 PI
2000
2500
3000 InfecteduntreatedArt**
* Infected untreated
Art 2x10mg/kg
3000
2500
2000
Average INFγ on day 8 PI
Con
cent
ra
Infected untreated
Art 2x10mg/kg 8001000 Infected
untreated
1000
800
Average TNF on day 8 PI
Infected untreated
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
500
1000
1500
20002x10mg/kg
non significant vs. Art&Chl.
*
*Significant vs.untreated
Chl 2x15mg/kg
Art & Chl
2000
1500
1000
500
Chl 2x15mg/kg
Art & Chl
0200400600 untreated
Art 2x10mg/kg significant vs. 600
400
200 *Significant vs. untreated
Chl 2x15mg/kg
Art & Chl* * **
0s t&C
0 00
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 39
FIG 7. Effect of artemisone and chloroquine on cytokines in infected mice on day 8 post infection. y pPlasma cytokine levels in mice treated on days 6-8 (twice each day) post infection by artemisone (Art), chloroquine (Chl) or their combination. Columns and vertical bars represent mean ± SEM; n = 8-10/group.
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 40
Average IL10 on day 12 PIAverage IL4 on day 12 PIFigure 8
g y
16000
20000
20000
16000
Average IL4 on day 12 PI
500
600
Untreated uninfected
Treated uninfected
Untreated infected
/ml)
/ml)
/ml)
/ml)
/ml)
/ml)
g/m
l)
Average IL-4 on day 12 PI Untreated uninfected
Treated uninfected
Untreated infected
Average IL-10 on day 12 PI
600
500
4000
8000
12000 * * ***
ml)
12000
8000
4000l)
100
200
300
400
* * **
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
*Significant vs Art &
Conc
entr
atio
n (p
g/Co
ncen
trat
ion
(pg/
Conc
entr
atio
n (p
g/Co
ncen
trat
ion
(pg/
Conc
entr
atio
n (p
g/Co
ncen
trat
ion
(pg/
ncen
trat
ion
(pg
Art 2x10mg/kg
Chl 2x15mg/kg
Art & Chl
*Significant vs.
400
300
200
100ml)
** * * *
* * * *0
4000
ratio
n (p
g/m
0
atio
n (p
g/m
l
Average INFγ on day 12 PI
0
100g
Chl or Chl treatedCon
Average INFγ on day 12 PIU t t dUntreated uninfected
Average IFNγ on day 12 PI
Significant vs. Art treated
100
0
tratio
n (p
g/m
Average TNF on day 12 PIUntreated uninfected
Con
cent
rC
once
ntra
*600
800
1000
120015001800
Untreateduninfected Treated
i f t d*
Untreated uninfected
Treated uninfected
Untreated infected
Art 2x10mg/kg
on (p
g/m
l)on
(pg/
ml)
on (p
g/m
l)on
(pg/
ml)
on (p
g/m
l)on
(pg/
ml)
n (p
g/m
l)1800
1500
1200
Con
cent
600
800
1000Untreated uninfected
Treated uninfected
Untreated infected
Art 2x10mg/kg
1800
1500
1200*
*
200
400
600
300600900
1200 uninfectedUntreated* significant vs. Art .
*Chl 2x15mg/kg
Art & Chl
*Significant vs. Art treated
Conc
entr
atio
Conc
entr
atio
Conc
entr
atio
Conc
entr
atio
Conc
entr
atio
Conc
entr
atio
Conc
entr
atio
900
600
300 200
400
600
***
*
*Chl 2x15mg/kg
Art & Chl
*Significant vs. untreated infected
900
600
300
**
* * **
000 00
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Page 41
FIG 8. Effect of artemisone and chloroquine on cytokines in P. bergheiANKA infection on day 12 post infectionANKA infection on day 12 post infection.Plasma cytokine levels in mice treated on days 6-8 (twice each day) post infection by artemisone (Art), chloroquine (Chl) or their combination. Columns and vertical bars represent mean ± SEM; n = 8-10/group. on June 29, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 42
Table 1. A summary of drug effects on cytokine production in infected mice
Day post infection
Cytokine/Effects Artemisone Chloroquine Art+Chl
8 IL-4 = ↑ ↑ IL-10 ↑ ↑ ↑ IFN-γ = = ↓ TNF ↓ ↓ ↓
Positive effect 2 3 4 12 IL-4 = ↑ ↑
IL-10 ↑ = = IFN-γ ↑* ↑* = TNF ↓ ↓ ↓
Positive effect 1 1 2 ↑ (for IL-4 and IL-10) or ↓ (For IFN-γ and TNF) are effects related to alleviation of
CM ↑* (for IFN-γ) means effect that may increase CM = means no difference from control "Positive effect" is the cumulative number of effects that may alleviate CM, from
which the number of effects that may increase CM is reduced
on June 29, 2018 by guesthttp://aac.asm
.org/D
ownloaded from