NEW RESEARCH Abnormal Functional Activation and Connectivity in the Working Memory Network in Early-Onset Schizophrenia Marinos Kyriakopoulos, M.D., Ph.D., Danai Dima, Ph.D., Jonathan P. Roiser, Ph.D., Richard Corrigall, M.D., Gareth J. Barker, Ph.D., Sophia Frangou, M.D., Ph.D. Objective: Disruption within the working memory (WM) neural network is considered an integral feature of schizophrenia. The WM network, and the dorsolateral prefrontal cortex (DLPFC) in particular, undergo significant remodeling in late adolescence. Potential interac- tions between developmental changes in the WM network and disease-related processes for schizophrenia remain unclear. The aim of this study was to determine whether DLPFC activation and functional connectivity are impaired during WM in patients with early-onset schizophrenia (EOS; age of onset 18 years). Method: We used functional magnetic resonance imaging and psychophysiological interaction analysis to respectively measure blood oxygenation level– dependent signal and to derive functional connectivity estimates in response to the two-back WM task from 25 youths with EOS and 20 matched healthy adolescents. Results: Compared with healthy adolescents, patients with EOS showed re- duced engagement of the DLPFC, the anterior cingulate cortex (ACC), and frontal operculum, and had reduced DLPFC connectivity within the WM network. Patients with EOS showed abnormal reduction in the coupling of the DLPFC with the ACC, the inferior parietal lobule, and the middle occipital gyrus. In contrast to healthy adolescents, patients with EOS expres- sed age-related decrease in the activity of the DLPFC and an increase in its connectivity with the ACC. Conclusions: Patients with EOS show dysfunctional engagement and reduced integration within the WM neural network. The pattern of abnormal age-related correlations in DLPFC activity and connectivity suggests that schizophrenia-related processes have an impact on brain regions that show significant late developmental changes. J. Am. Acad. Child Adolesc. Psychiatry, 2012;51(9):911–920. Key Words: early onset schizophrenia, psycholog- ical interaction, PPI, fMRI, working memory D isruption in working memory (WM) is considered an integral feature of schizo- phrenia thought to underlie both the cog- nitive and clinical pathology observed in patients with this disorder. 1,2 This notion is further rein- forced by evidence showing that WM deviance is a feature of the schizophrenia prodrome and may predict transition to psychosis. 3,4 Working memory functions are supported by a distributed network of cortical and sub- cortical regions. 5 Within this network, task performance in healthy individuals has been crucially linked to the level of activation within the dorsolateral prefrontal cortex (DLPFC) and posterior parietal cortices. 5 Conversely, schizo- phrenia has been reliably associated with ab- normalities in DLPFC function 6-8 and reduced functional integration (i.e. coordinated interac- tion) within the WM network. 9 Several mecha- nistic explanations have been proposed to ac- count for these findings. These include reduced neuronal connectivity within the DLPFC 10 , im- paired synchronization of pyramidal cells in the DLPFC resulting from altered regulation by parvalbumin-positive GABA neurons 11 , abnor- mal anatomical connectivity between prefron- tal and posterior brain regions 12-14 and bot- tom-up deficits in WM driven by impairments Supplemental material cited in this article is available online. JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY VOLUME 51 NUMBER 9 SEPTEMBER 2012 911 www.jaacap.org
Transcript
NEW RESEARCH
Abnormal Functional Activation andConnectivity in the Working Memory
Network in Early-Onset SchizophreniaMarinos Kyriakopoulos, M.D., Ph.D., Danai Dima, Ph.D., Jonathan P. Roiser, Ph.D.,
Richard Corrigall, M.D., Gareth J. Barker, Ph.D., Sophia Frangou, M.D., Ph.D.
Objective: Disruption within the working memory (WM) neural network is considered anintegral feature of schizophrenia. The WM network, and the dorsolateral prefrontal cortex(DLPFC) in particular, undergo significant remodeling in late adolescence. Potential interac-tions between developmental changes in the WM network and disease-related processes forschizophrenia remain unclear. The aim of this study was to determine whether DLPFCactivation and functional connectivity are impaired during WM in patients with early-onsetschizophrenia (EOS; age of onset �18 years). Method: We used functional magneticresonance imaging and psychophysiological interaction analysis to respectively measure bloodoxygenation level–dependent signal and to derive functional connectivity estimates inresponse to the two-back WM task from 25 youths with EOS and 20 matched healthyadolescents. Results: Compared with healthy adolescents, patients with EOS showed re-duced engagement of the DLPFC, the anterior cingulate cortex (ACC), and frontal operculum,and had reduced DLPFC connectivity within the WM network. Patients with EOS showedabnormal reduction in the coupling of the DLPFC with the ACC, the inferior parietal lobule,and the middle occipital gyrus. In contrast to healthy adolescents, patients with EOS expres-sed age-related decrease in the activity of the DLPFC and an increase in its connectivity withthe ACC. Conclusions: Patients with EOS show dysfunctional engagement and reducedintegration within the WM neural network. The pattern of abnormal age-related correlationsin DLPFC activity and connectivity suggests that schizophrenia-related processes have animpact on brain regions that show significant late developmental changes. J. Am. Acad. ChildAdolesc. Psychiatry, 2012;51(9):911–920. Key Words: early onset schizophrenia, psycholog-ical interaction, PPI, fMRI, working memory
pnft
mt
D isruption in working memory (WM) isconsidered an integral feature of schizo-phrenia thought to underlie both the cog-
nitive and clinical pathology observed in patientswith this disorder.1,2 This notion is further rein-forced by evidence showing that WM deviance isa feature of the schizophrenia prodrome and maypredict transition to psychosis.3,4
Working memory functions are supportedby a distributed network of cortical and sub-cortical regions.5 Within this network, taskperformance in healthy individuals has been
Supplemental material cited in this article is available online.
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
crucially linked to the level of activation withinthe dorsolateral prefrontal cortex (DLPFC) andposterior parietal cortices.5 Conversely, schizo-
hrenia has been reliably associated with ab-ormalities in DLPFC function6-8 and reducedunctional integration (i.e. coordinated interac-ion) within the WM network.9 Several mecha-
nistic explanations have been proposed to ac-count for these findings. These include reducedneuronal connectivity within the DLPFC10, im-paired synchronization of pyramidal cells inthe DLPFC resulting from altered regulation byparvalbumin-positive GABA neurons11, abnor-
al anatomical connectivity between prefron-al and posterior brain regions12-14 and bot-
tom-up deficits in WM driven by impairments
911www.jaacap.org
apttt(d
ehPi
KYRIAKOPOULOS et al.
in basic perceptual processes within primarysensory brain regions.15
Another important consideration relates to thedevelopmental course of WM deficits in schizo-phrenia. Normally developing adolescents showsignificant performance gains in WM tasks.16
This improvement has been associated with in-creased recruitment of frontoparietal regions,17,18
enhanced white-matter structural connectivitybetween the superior frontal and parietal corti-ces19 and greater functional integration of theWM network.20 These findings suggest that dur-ing adolescence the WM network undergoes sig-nificant remodelling and strengthening.20 Theopposite may be the case in schizophrenia. Usingprospective longitudinal data from the Dune-din Multidisciplinary Health and DevelopmentStudy, Reichenberg and colleagues (2010)21 haveshown that WM deviance in future schizophreniacases shows progressive increase beginning inearly adolescence. The neural correlates of thesebehavioural changes are not known but are likelyto reflect interactions between developmentaland disease-related processes within the WMnetwork. In this context the investigation of theneural correlates of WM in patients with EarlyOnset Schizophrenia (EOS; age of onset �18years) is particularly pertinent.
Three studies to date have examined the neu-ral correlates of WM in patients with EOS com-pared with normally developing youths,22-24 butreduced DLPFC engagement was observed onlyin one.23 This raises questions as to the develop-mental course of DLPFC deficits. Two hypothe-ses have been put forward. Prefrontal deficits inschizophrenia may emerge post-adolescence as adownstream consequence of developmentalchanges.24 Alternatively, the magnitude of pre-frontal deficit may increase with age, thus be-coming more reliably detectable in the post-adolescent period.25
To address these questions, we investigatedthe neural correlates WM in EOS by acquiringfunctional magnetic resonance imaging (fMRI)data during the two-back task. This task has twoparticular advantages over other WM para-digms. First, it has been widely used in schizo-phrenia,6-9 thus providing a robust reference basefor our study. Second, the task requires manipu-lation of information in WM, which is moresensitive to developmental20 and schizophrenia-related processes.26 Two main analyses were
conducted. First, we tested whether functional
JOURN
912 www.jaacap.org
bnormalities within the WM neural network areresent in EOS and whether these are differen-
ially affected by developmental age. During WMasks, connectivity deficits relating to the integra-ion of the DLPFC with the anterior cingulateACC) and parietal cortices have been reliablyemonstrated in adult schizophrenia.6-9 Our sec-
ond analysis used psychophysiological interac-tion (PPI) analysis27 to test whether disruption inthe functional coupling of the DLPFC with otherbrain regions is present in EOS and whether it isinfluenced by developmental age.
METHODSubjectsTwenty-five patients were recruited from the SouthLondon and Maudsley NHS Foundation Trust Childand Adolescent mental health services, based on thefollowing criteria: fulfillment of DSM-IV criteria forschizophrenia; age 12 to 19 years; onset of schizophre-nia before the 18th birthday; no co-morbid Axis Idiagnosis; and IQ � 70. Twenty healthy individualsmatched for age, sex, and parental socioeconomicstatus, who had no personal history of psychiatricdisorder and no family history of psychosis in theirfirst-degree relatives, were recruited from the samegeographic area via advertisements. Exclusion criteriafor all participants were as follows: history of headinjury; lifetime history of substance dependence asdefined by the DSM-IV; substance abuse as defined bythe DSM-IV within the 6 months preceding studyentry; any concomitant medical condition; and historyof hereditary neurological disorders. All participantswere assessed by qualified child psychiatrists usingthe Structured Clinical Interview for DSM-IV Axis IDisorders.28,29 Psychopathology was assessed in pa-tients with the Positive and Negative Syndrome Scale(PANSS).30 Current and total cumulative antipsychoticexposure was calculated in chlorpromazine equiva-lents (CPZE). In all participants, the Wide RangeAchievement Test (WRAT3)31 was used to obtain anstimate of general intellectual ability, and the Annetteandedness scale was used to asses hand dominance.arental socioeconomic status was determined accord-
ng to the Standard Occupational Classification.32 Pa-tients were monitored so as to ensure that they weretested when clinically stable defined as scoring �3 oneach item of the PANSS positive syndrome subscalefor 2 consecutive weeks before scanning.
The study was approved by the Joint South Londonand Maudsley and Institute of Psychiatry ResearchEthics Committee. All participants and their parents orguardians, as appropriate, provided written informedconsent or assent after detailed description of the
study.
AL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
tdai(vrsocrtastcrtctarttwcwt(
WORKING MEMORY AND SCHIZOPHRENIA
NeuroimagingTwo-Back Task. The two-back task was administeredas a block design, incorporating alternating active andcontrol conditions. Stimuli consisted of letters dis-played one at the time at the center of a screenpresented to participants visually by means of a pris-matic mirror. At the beginning of each condition,participants were explicitly instructed to respond bybutton press to the target letter. In the control (zero-back) condition, the designated target letter was “X.”In the active condition (two-back), the target letter wasdefined as any letter that was identical to the onepresented two trials back. The order of the letters waspseudo-randomized to avoid any systematic ordereffects. In each condition, a series of 14 letters waspresented visually, with each letter displayed for 2seconds. Responses were monitored via a magneticresonance imagining (MRI)–compatible button boxheld in the subject’s dominant hand. There were 12epochs in all, each lasting 30 seconds, with totalexperiment duration of 6 minutes. The ratio of target tonon-target letters presented per block ranged from 2:12to 3:11. Response time to target letters and number ofcorrect responses (accuracy) were recorded.Task Performance. Before scanning, all participants re-ceived training on a training set of the zero-back andtwo-back conditions until they achieved 80% of thetargets in the zero-back condition to ensure that theyunderstood the task instructions. Group differencesin task performance during scanning were examinedusing repeated-measures analyses with accuracy or re-sponse time as dependent variables, condition (zero-backor two-back) as the within-subject factor, and diagnosisas the between-subject factor.Image Acquisition. fMRI and anatomical data wereacquired during the same session on a 1.5-T imagingsystem (Signa; GE Medical Systems, Milwaukee, Wis-consin) using a gradient-echo echoplanar imaging se-quence and a 3D T1-weighted Inversion Recoveryprepared Spoiled GRASS sequence, respectively. Atotal of 270 T2*-weighted EPI brain volumes depictingblood oxygenation level–dependent (BOLD) contrastwere acquired at each of 16 axial planes (echo time �40 milliseconds, repetition time � 2 seconds, voxeldimensions � 3.75 � 3.75 � 7 mm3, interslice gap � 0.7mm, matrix size � 64 � 64, flip angle � 70o), the firstfour of which were discarded to allow the scanner toreach a steady state in T1 contrast. Structural imageswere acquired using a 3D axial T1-weighted InversionRecovery prepared Spoiled GRASS sequence from 128slices (echo time � 5.1 milliseconds, repetition time �2 seconds, inversion time � 450 milliseconds, voxeldimensions � 0.9375 � 0.9375 � 1.5 mm3, matrix size �256 � 192, field of view � 240 � 180 mm2, flip angle �20°, number of excitations � 1).Image Analysis. Image processing and analysis wereimplemented using Statistical Parametric Mapping
(SPM8) software (www.fil.ion.ucl.ac.uk/spm/). Pre- o
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
processing involved realignment, transformation intostandard stereotactic Montreal Neurological Institute(MNI) space, and smoothing with an isotropic Gauss-ian kernel of 8-mm full-width-at-half-maximum. Nosubjects were excluded due to excessive interscanmotion (defined as �4-mm translation, �4° rotation),and there were no group differences in task registra-tion parameters. A first-level fixed effect model wascomputed for each participant. fMRI responses weremodeled using a canonical hemodynamic responsefunction (hrf) convolved with the vectors of interest;accuracy per block was added as a parametric modu-lator. Movement parameters were entered as nuisancecovariates. Statistical contrast images of the two-backversus zero-back condition were produced for eachsubject and entered in an independent-sample t test atthe second-level random effects analysis. Statisticalsignificance was set at p � .05 using family-wise error(FWE) correction. Regions of interest (ROIs) werecreated from suprathreshold clusters where an effectof diagnosis was noted, and weighted parameter esti-mates were extracted from each subject using theMarsBaR toolbox (marsbar.sourceforge.net). Thesewere used in further analyses in SPSS 15.0 (SPSS Inc,Chicago, IL) to examine the effect of age, WRAT3scores, antipsychotic medication exposure, illness du-ration, and PANSS scores.Psychophysiological Interaction Analysis. The aim ofhe PPI analysis was to examine DLPFC connectivityuring the two-back task. The PPI analysis consists ofdesign matrix with three regressors: the “psycholog-
cal variable,” representing the experimental taskhere, two-back versus zero-back); the “physiologicalariable,” representing the neural response in the seedegion (here the DLPFC); and a third variable repre-enting the interaction between the first and the sec-nd variables. The coordinates of the seed regionsorresponded to the local maxima within the left andight DLPFC detected in the within-group analysis ofhe two-back � zero-back contrast and thresholded atn uncorrected p � 0.05 to ensure its presence in allubjects. For each individual, the first eigenvariateime was extracted from a sphere of 6-mm radiusentered on the peak height coordinates for the left andight DLPFC for each group. A variable representinghe interaction between each time series and the psy-hological variable (two-back versus zero-back con-rast) was constructed for each subject. Voxel-wise PPInalyses were conducted within each group sepa-ately. Subject-specific contrast images were then en-ered into a random-effects analysis using one-sampletests (thresholded at p � .0001 uncorrected voxel-ise with p � .05 cluster level correction). To directly
ompare group differences in functional connectivity,e examined the interaction between the PPI (interac-
ion between DLPFC activity and memory) and grouppatients with EOS and healthy participants) (thresh-
For both fMRI and PPI analyses, stereotactic coor-dinates were converted from MNI spatial array to thatof Talairach and Tournoux (www.mrc-cbu.cam.ac.uk/Imaging/mnispace.html) and corresponding anatomi-cal regions and Brodmann areas (BA) were identifiedwith the Talairach Daemon Client (www.talairach.org).
RESULTSParticipantsDemographic and clinical information about thesample are shown in Table 1. There were nogroup differences in age (p � .35), sex (p � .7),parental socioeconomic status (p � .41), or
TABLE 1 Demographic and Clinical Characteristics andBehavioral Data for Study Subjects
Clinical CharacteristicsAge of onset (y) 14.8 (1.6) —PANSS positive
symptoms score8.9 (2.1) —
PANSS negativesymptoms score
12.4 (3.4) —
PANSS total score 44 (5.6) —Task Performance in Two-
Back TaskZero-Back accuracy (%
correct)0.95 (0.7) 0.99 (0.0)
Two-Back accuracy (%correct)
0.86 (0.1) 0.91 (0.2)
Zero-Back responsetime (sec)
0.50 (0.8) 0.49 (0.1)
Two-Back responsetime (sec)
0.67 (0.2) 0.57 (0.1)
Note: Except for sex, handedness, and socioeconomic status, data arepresented as mean (SD). PANSS � Positive and Negative SyndromeScale; sec � seconds; WRAT3 � Wide Range Achievement Test;y � years.
aThree condensed categories (1 � professional or managerial, 2 �
skilled or partly skilled, 3 � unskilled or unemployed) derived from theStandard Occupational Classification (Office of Population Censuses
oand Surveys, 2010).32
JOURN
914 www.jaacap.org
RAT3 scores (p � .8). All patients included inhe analysis received atypical antipsychotic med-cation (risperidone, n � 12; olanzapine, n � 4;ripiprazole, n � 5; amisulpiride, n � 2). Theean comparable daily antipsychotic dose at the
ime of scanning was 285.9 mg (range, 50-1200g) CPZE. No patient was prescribed anticholin-
rgic medication or benzodiazepines.
ehavioral Dataetails on task performance are shown in Table
. There no significant effect of diagnosis onccuracy and response time (p � .1).
MRI Analysisffect of Task. Both healthy and EOS participantsctivated similar networks encompassing the bi-ateral DLPFC, the ACC, the inferior parietalobule (IPL), the superior parietal lobule (SPL),nd the cerebellum (Table S1, available online).ffect of Diagnosis. In the two-back � zero-backontrast, patients with EOS showed reduced ac-ivation compared with healthy adolescents inhree clusters centred in the left DLPFC (middlerontal gyrus; BA9; x � �28, y � 32, z � 33;
cluster sixe 192; Z-value � 4.48), the ACC bilat-erally (BA 24/32; x � �2, y � 30, z � 19; Z-value �4.24. and x � 4, y � 32, z � 20; Z-value � 3.98;cluster size � 294) and the frontal operculum(inferior frontal gyrus; BA 44/45; x � �52, y � 9,z � 16; cluster size � 71; Z-value � 3.96) (Figure). Patients with EOS did not show greater acti-ation than healthy adolescents in any brainegion.
No significant correlations were observed be-ween mean signal change in suprathresholdlusters and WRAT3 scores, illness duration,urrent and cumulative medication exposure,nd PANSS scores in the EOS group (all p �
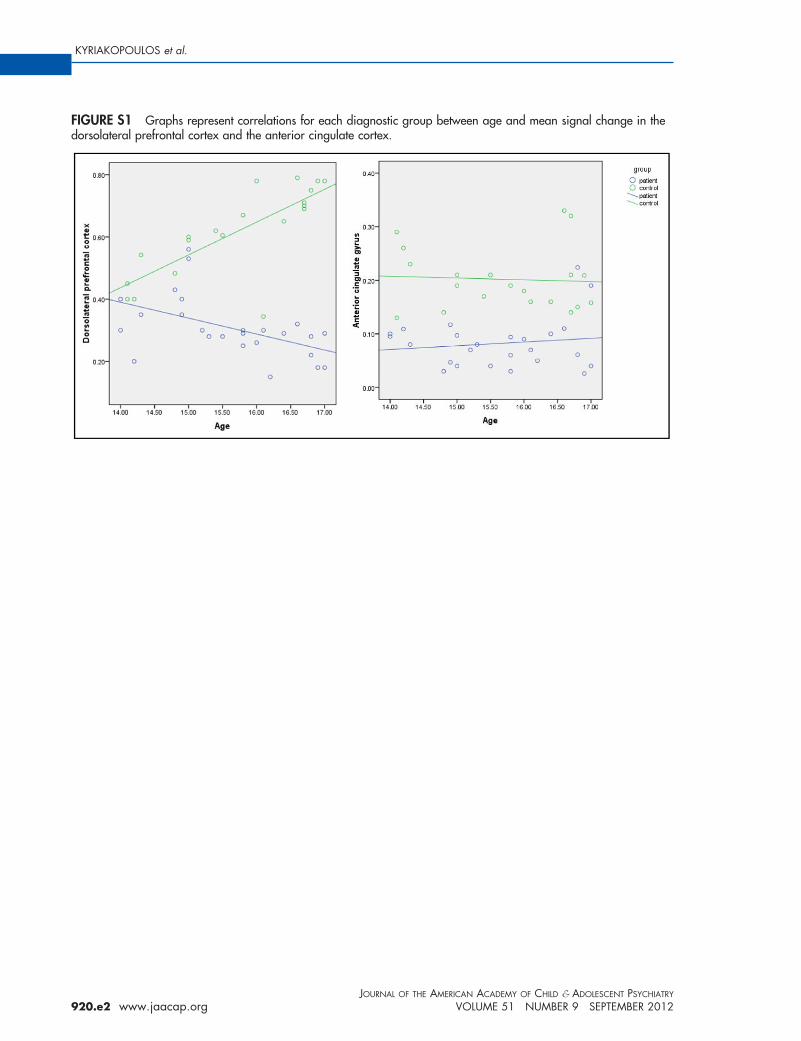
0.15). Significant correlations with age were iden-tified for mean signal change in left DLPFCwhich were positive for healthy adolescents (r �0.48, p � 0.03) and negative for patients with EOS(r � �0.55, p � 0.05); the slope of this age-relatedhange was significantly different between thewo groups (z � 2.79, p � 0.005) (Figure 1 andlso in Figure S1, available online). However, nouch differences in age-related changes betweenhe groups were identified in the ACC or frontal
perculum.
AL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
Psychophysiological Interaction AnalysisBased on the results of the whole brain within-group analysis, the left and right DLPFC seedvoxels were centred on the peak co-ordinates ofthe corresponding suprathreshold clusters foreach group. The Talairach coordinates for the leftDLPFC seed were x � �46, y � 19, z � 23(z-value � 6.45) in healthy participants and x ��44, y � 40, z � 18 (z-value � 4.64) in patientswith EOS (Table S1, available online). The Ta-lairach coordinates for the right DLPFC seed
FIGURE 1 Group differences in regional brain activatio(top) and z � 22 (bottom) respectively showing the clustepatients (n � 25) compared with healthy adolescents (n �anterior cingulate cortex (ACC). Graphs represent correlasignal change in each of the corresponding clusters. Theactivation of each group is displayed in the graphs purelywere identified for mean signal change in the left DLPFC0.03) and negative for patients with EOS (r � �0.55, pwas significantly different between the two groups (z � 2
were x � 44, y � 21, z � 32 (z-value � 4.55) in D
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
ealthy participants and x � 48, y � 23, z � 34z-value � 5.48) in patients with EOS (Figure S1,vailable online).
Within-group PPI analyses showed significantnteraction in healthy adolescents between WMtwo-back � zero-back) and the left and rightLPFC in several regions of the WM system
Table 2). In patients with EOS, no significantnteractions were observed between the leftLPFC and any other brain region, whereas onlyne interaction was observed for the right
ring the two-back task. Note: Axial sections at z � 33reduced activation in Early Onset Schizophrenia (EOS)) in the left dorsolateral prefrontal cortex (DLPFC) andfor each diagnostic group between age and meanle was divided in three age groups and the meanease of visualization. Significant correlations with ageh were positive for healthy adolescents (r � 0.48, p �05); the slope of these age-related changes in activationp � 0.005).
n durs of
20tionssamp
forwhic� 0..79,
LPFC (Table 2).
915www.jaacap.org
hopg
KYRIAKOPOULOS et al.
Group comparisons revealed a significant dif-ference in functional connectivity during two-back � zero-back between the left DLPFC seedand three regions, the ACC, the inferior parietallobule, and the middle occipital gyrus (Table 3;Figure 2), and between the right DLPFC seed andthe left DLPFC. For all regions, healthy adoles-cents showed increased connectivity comparedwith patients.
TABLE 2 Regions Expressing Functional Connectivity Betand Working Memory in Healthy Adolescents and Patien
TABLE 3 Regions Expressing Increase in Functional ConWorking Memory Task in Healthy Adolescents Compared
Region Gyrus Laterality
Seed Middle Frontal LeftFrontal Anterior cingulate gyrus Left
Inferior parietal lobule RightMiddle occipital gyrus Right
Seed Middle frontal RightFrontal Middle frontal gyrus Left
JOURN
916 www.jaacap.org
We then investigated the effect of age on thefunctional connectivity of the DLPFC. A signifi-cant positive correlation was identified for thefunctional coupling between DLPFC-ACC andage in patients (r � 0.68, p � .01) but not in the
ealthy adolescents (r � �0.05, p � .82); the slopef these age-related changes in DLPFC-ACC cou-ling was significantly different between the tworoups (z � 2.08, p � .04).
Left and Right Dorsolateral Prefrontal Cortical Activityith Early-Onset Schizophrenia
AL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
WORKING MEMORY AND SCHIZOPHRENIA
DISCUSSIONIn this study, we used fMRI and PPI to examineneural responses and functional connectivitywithin the WM network in patients with EOScompared with normally developing youths. Ouraim was to examine the effect of diagnosis ontask-specific neural activation maps as well as inthe patterns of functional connectivity underly-ing WM. Our key findings were that patientswith EOS compared with healthy adolescentsshowed reduced engagement of the DLPFC,ACC and frontal operculum and had reducedDLPFC connectivity within the WM network. Inaddition, patients with EOS had an age-relateddecrease in the activity of the DLPFC, and anage-related increase in the modulation of itsconnectivity with the ACC by WM.
Reduced Engagement of the WM Networkin EOSOur results of reduced activation within thelateral prefrontal cortex extending from theDLPFC to the operculum are in line with the richneuroimaging literature demonstrating a reliableassociation between reduced lateral prefrontalengagement and the clinical syndrome of schizo-
FIGURE 2 Regions of significant group differences in ptask and activity of the dorsolateral prefrontal cortex.
phrenia.6-9 Neuroimaging studies of adults with
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
schizophrenia suggest that the relative task diffi-culty between patients and healthy individualsmay account for whether DLPFC activation infound to be decreased or increased in patients.33
In patients with schizophrenia peak activation ofthe working memory system may be reached at alower memory load than in healthy individuals.Thus, patients with schizophrenia either engagegreater prefrontal resources to maintain theirperformance or may fail to sustain prefrontalengagement, resulting in underperformance.34,35
In this study, patients’ performance was numer-ically, although not statistically, lower than thatof healthy youths, particularly in the two-backtask, which may account for the observed reduc-tion in DLPFC activity.
However, Haenschel et al.22 and White et al.,24
who also examined WM in patients with EOS,did not find an effect of diagnosis on prefrontalactivation. This discrepancy is, in our view, at-tributable to significant differences in methodol-ogy. Haenschel et al.22 used a visual delayeddiscrimination task in conjunction with event-related potentials and reported early visual per-ceptual processing deficits in EOS. They thenexamined fMRI data from the occipital lobes
ophysiological interaction between the working memory
sych
focusing on functional abnormalities in visual
917www.jaacap.org
sttt
IoNspWgdm
ysAhtlyet
acwaalnsmo
KYRIAKOPOULOS et al.
cortices. The study is therefore not informativewith regard to the effect of diagnosis on otherWM network regions. White et al. used a versionof the Sternberg Item Recognition Paradigm(SIRP). This is a delayed match-to-sample taskthat requires participants to memorize a set ofnumbers and to indicate whether subsequentlypresented single numbers were included in thememorised set. The SIRP and N-back task havesimilar requirements in terms of informationmaintenance. However, the N-back task includesthe additional component of continuous WMupdating, which poses greater demands on theDLPFC36 and hence elicits more reliable activa-tion in this region than delayed match-to-sampletasks.8 Both in our study and in a similar inves-tigation by Pauly et al.23 the use of the N-backparadigm revealed reduced DLPFC engagementin EOS. Therefore it could be argued that, in thisage group, decrements in DLPFC function maybe more reliably elicited either in response togreater task difficulty or to the specific manipu-lation requirements of the N-back task.
In contrast, reduced ACC engagement in EOSappears to be present irrespective of WM para-digm.23,24 A broadly supported view is that theACC is engaged in performance monitoringacross a number of tasks and it modulatesDLPFC to implement post-error behavioral ad-justment.37,38 The ACC hypoactivation in pa-tients with EOS during WM is consistent withfindings in adult cases6,8 and suggests that cin-gulate dysfunction is a robust correlate of diseaseexpression in schizophrenia across age groups.
Reduced Prefrontal Functional Connectivity DuringWM in EOSIn healthy adolescents, the left DPLFC showedsignificant functional interactions with corticaland cerebellar regions throughout the WM net-work, in line with previous reports.20,39 As pre-dicted, patients with EOS expressed abnormalDLPFC connectivity similar to the pattern re-ported in adults with schizophrenia.9,40 There aretwo other studies that examined connectivity inpatients with EOS. White et al.24 found reducedconnectivity in patients with EOS compared withhealthy youths in several brain networks, which,however, did not include prefrontal regions. Incontrast, Zhou et al.41 reported reduced sponta-neous functional coupling of prefrontal areaswith posterior brain regions in patients with EOS
during the resting state. Our results therefore
JOURN
918 www.jaacap.org
upport the notion that abnormalities in prefron-al connectivity in EOS are present both spon-aneously and during cognitively demandingasks.
mplications for the Neurodevelopmental Modelf Schizophreniaormally developing youths in our sample
howed a significant correlation between age andrefrontal but not ACC activation during theM task. These findings are in line with a
rowing literature showing that late adolescentevelopment is associated with increased andore focal recruitment in prefrontal regions,42,43
whereas developmental changes in the ACCreach adult levels earlier.44 In addition, healthy
ouths showed a negative, albeit not statisticallyignificant, correlation between age and DLPFC–CC coupling. Studies in healthy individualsave shown a gradual and monotonic decrease in
he functional connectivity of the ACC withateral prefrontal regions from mid-childhood tooung adulthood.45 This has been interpreted asvidence of transition to more efficient organiza-ion of brain networks.
In patients with EOS, there was evidence ofn interaction between these developmentalhanges and disease-related mechanisms. First,e observed an inverse correlation between age
nd DLPFC activation, suggesting greater devi-nce in participants who underwent scanning inate adolescence. These functional changes reso-ate with anatomical findings from longitudinaltudies in EOS documenting progressive grayatter loss in prefrontal regions throughout ad-
lescence and into young adulthood.25,46,47 Sec-ond, the interaction between disease processesand late development appears to be region spe-cific. In contrast to DLPFC, activation within theACC showed no correlation with age and wassimilarly reduced in patients compared withhealthy adolescents across the age range exam-ined. This implies that age-related changes inEOS are greater in regions, such as the DLPFC,that undergo active reorganization during ado-lescence. Third, a similar inference may be drawnfrom the overall reduction of modulation ofDLPFC connectivity by WM within the WMnetwork, and abnormal positive correlation be-tween age and functional coupling of the DLPFCwith the ACC. These findings imply that interac-tions between late development and disease pro-
cess extend beyond specific regions to have an
AL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
WORKING MEMORY AND SCHIZOPHRENIA
impact on the functional organization of WMnetworks.
Methodological ConsiderationsThere are several methodological issues to beconsidered. All patients with EOS were treatedwith atypical antipsychotic medication, whichmay have affected both neural responsivenessand functional connectivity. Current evidencesuggests that treatment with atypical antipsy-chotics in schizophrenia has a “normalising”influence on cognitive function,48 neural engage-ment,49 and functional connectivity.50 It is alsoreassuring that we did not observe any signifi-cant effect of medication in any of the analyses.We consider the lack of statistically significantgroup differences in behavioral performance as aparticular strength of the study, as it removedconfounds relating to poor motivation or nonspe-cific difficulties in adhering to task instructions.Both functional and connectivity abnormalitiespersisted despite comparable performance in pa-tients and healthy adolescents, and this confirmsprevious observations relating to the increased sen-sitivity of fMRI compared with behavioural mea-sures. Our data also provide direction with regardto further research. First, PPI analysis tests forcorrelations, and not causal influences, and there-fore examination of effective connectivity betweenWM network regions may prove informative. Sec-ond, differential age-related changes in patientswith EOS and adolescents suggest the presence of
ongoing interactions between late brain develop-
7. Kim DI, Manoach DS, Mathalon DH, et al. Dysregulation ofworking memory and default-mode networks in schizophrenia
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
ment and disease processes that can be directlytested only in longitudinal studies.
In sum, we found that, during WM tasks,patients with EOS have reduced activation inprefrontal and anterior cingulate cortices com-pared with healthy adolescents, and abnormalDLPFC functional connectivity throughout theentire WM network. The pattern of abnormalage-related correlations in DLPFC activity andconnectivity suggests that the function and in-tegration of regions that show significant latedevelopmental changes are also affected byschizophrenia-related disease processes. &
Accepted June 27, 2012.
This article was reviewed under and accepted by Deputy Editor EllenLeibenluft, M.D.
Dr. Kyriakopoulos is with the Institute of Psychiatry, King’s CollegeLondon and Child and Adolescent Mental Health Services, MaudsleyHospital, UK. Drs. Dima and Frangou are with the Institute ofPsychiatry, King’s College London. Dr. Roiser is with the Institute ofCognitive Neuroscience, University College London, UK. Dr. Corrigallis with Child and Adolescent Mental Health Services, MaudsleyHospital, UK. Dr. Barker is with the Centre for Neuroimaging Sci-ences, Institute of Psychiatry, King’s College London, UK.
This study was supported by a Brain and Behavior Research Founda-tion Young Investigator Award (M.K.).
Disclosure: Dr. Frangou has received honoraria for advisory panelsand continuing medical education (CME) activities from Lundbeck,Janssen-Cilag, and BMS-Otsuka. Drs. Kyriakopoulos, Dima, Roiser,Corrigall, and Barker report no biomedical financial interests orpotential conflicts of interest.
Correspondence to Sophia Frangou, M.D., Ph.D., Section ofNeurobiology of Psychosis, Department of Psychosis Studies, Instituteof Psychiatry, King’s College London, De Crespigny Park, LondonSE5 8AF, UK; E-mail: [email protected]
REFERENCES1. Goldman-Rakic PS. Working memory dysfunction in schizophre-
nia. J Neuropsychiatry Clin Neurosci. 1994;6:348-357.2. Weinberger DR, Berman KF. Speculation on the meaning of
cerebral metabolic hypofrontality in schizophrenia. SchizophrBull. 1998;14:157-168.
3. Pukrop R, Ruhrmann S, Schultze-Lutter F, Bechdolf A, Brock-haus-Dumke A, Klosterkötter J. Neurocognitive indicators for aconversion to psychosis: comparison of patients in a potentiallyinitial prodromal state who did or did not convert to a psychosis.Schizophr Res. 2007;92:116-125.
4. Seidman LJ, Thermenos HW, Poldrack RA, et al. Altered brainactivation in dorsolateral prefrontal cortex in adolescents andyoung adults at genetic risk for schizophrenia: an fMRI study ofworking memory. Schizophr Res. 2006;85:58-72.
5. Champod AS, Petrides M. Dissociation within the frontoparietalnetwork in verbal working memory: a parametric functionalmagnetic resonance imaging study. J Neurosci 2010;30:3849-3856.
6. Glahn DC, Ragland JD, Abramoff A, Barrett J, Laird AR, BeardenCE. Beyond hypofrontality: a quantitative meta-analysis of func-tional neuroimaging studies of working memory in schizophre-nia. Hum Brain Mapp. 2005;25:60-69.
using independent component analysis, an fBIRN and MCICstudy. Hum Brain Mapp. 2009;30:3795-3811.
8. Minzenberg MJ, Laird AR, Thelen S, Carter CS, Glahn DC.Meta-analysis of 41 functional neuroimaging studies of executivefunction in schizophrenia. Arch Gen Psychiatry. 2009;66:811-822.
9. Meda SA, Stevens MC, Folley BS, Calhoun VD, Pearlson GD.Evidence for anomalous network connectivity during workingmemory encoding in schizophrenia: an ICA based analysis. PLoSOne. 2009;4:e7911.
10. Selemon LD, Rajkowska G, Goldman-Rakic PS. Abnormally highneuronal density in the schizophrenic cortex. A morphometricanalysis of prefrontal area 9 and occipital area 17. Arch GenPsychiatry. 1995;52:805-818.
11. Lewis DA, Hashimoto T, Volk DW. Cortical inhibitory neuronsand schizophrenia. Nat Rev Neurosci. 2005;6:312-324.
12. Ellison-Wright I, Bullmore E. Meta-analysis of diffusion tensorimaging studies in schizophrenia. Schizophr Res. 2009;108:3-10.
13. Kyriakopoulos M, Bargiotas T, Barker GJ, Frangou S. Diffusiontensor imaging in schizophrenia. Eur Psychiatry. 2008;23:255-273.
14. Sugranyes G, Kyriakopoulos M, Dima D, et al. Multimodalanalyses identify linked functional and white matter abnormali-
ties within the working memory network in schizophrenia.Schizophr Res. 2012;138:136-142.
15. Javitt DC. When doors of perception close: bottom-up models ofdisrupted cognition in schizophrenia. Annu Rev Clin Psychol.2009;5:249-275.
16. Crone EA, Wendelken C, Donohue S, van Leijenhorst L, BungeSA. Neurocognitive development of the ability to manipulateinformation in working memory. Proc Natl Acad Sci USA.2006;103:9315-9320.
17. Klingberg T, Forssberg H, Westerberg H. Increased brain activityin frontal and parietal cortex underlies the development ofvisuospatial working memory capacity during childhood. J CognNeurosci. 2002;14:1-10.
18. Kwon H, Reiss AL, Menon V. Neural basis of protracted devel-opmental changes in visuo-spatial working memory. Proc NatlAcad Sci USA. 2002;99:13336-13341.
19. Nagy Z, Westerberg H, Klingberg T. Maturation of white matteris associated with the development of cognitive functions duringchildhood. J Cogn Neurosci. 2004;16:1227-1233.
20. Finn AS, Sheridan MA, Kam CL, Hinshaw S, D’Esposito M.Longitudinal evidence for functional specialization of the neuralcircuit supporting working memory in the human brain. J Neu-rosci. 2010;30:11062-11067.
21. Reichenberg A, Caspi A, Harrington H, et al. Static and dynamiccognitive deficits in childhood preceding adult schizophrenia: a30-year study. Am J Psychiatry. 2010;167:160-169.
22. Haenschel C, Bittner RA, Haertling F, et al. Contribution ofimpaired early-stage visual processing to working memory dys-function in adolescents with schizophrenia: a study with event-related potentials and functional magnetic resonance imaging.Arch Gen Psychiatry. 2007;64:1229-1240.
23. Pauly K, Seiferth NY, Kellermann T, et al. Cerebral dysfunctionsof emotion-cognition interactions in adolescent-onset schizophre-nia. J Am Acad Child Adolesc Psychiatry. 2008;47:1299-1310.
24. White T, Schmidt M, Kim DI, Calhoun VD. Disrupted functionalbrain connectivity during verbal working memory in childrenand adolescents with schizophrenia. Cereb Cortex. 2011;21:510-518.
25. Thompson PM, Vidal C, Giedd JN, et al. Mapping adolescentbrain change reveals dynamic wave of accelerated gray matterloss in very early-onset schizophrenia. Proc Natl Acad Sci USA.2001;98:11650-11655.
26. Cannon TD, Glahn DC, Kim J, et al. Dorsolateral prefrontal cortexactivity during maintenance and manipulation of information inworking memory in patients with schizophrenia. Arch GenPsychiatry. 2005;62:1071-1080.
27. Friston KJ, Buechel C, Fink GR, et al. Psychophysiological andmodulatory interactions in neuroimaging. NeuroImage. 1997;6:218-229.
28. First MB, Spitzer RL, Gibbon M, Williams JBW. StructuredClinical Interview for DSM-IV-TR Axis I Disorders, ResearchVersion, Non-patient Edition. (SCID-I/NP). New York: Biomet-rics Research, New York State Psychiatric Institute; 2002.
29. First MB, Spitzer RL, Gibbon M, Williams JBW. StructuredClinical Interview for DSM-IV-TR Axis I Disorders, ResearchVersion, Patient Edition. (SCID-I/P). New York: Biometrics Re-search, New York State Psychiatric Institute; 2002.
30. Kay SR, Fiszbein A, Opler LA. The Positive and Negative Syn-drome Scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13:261-276.
31. Wilkinson GS. WRAT3 Administration Manual. Delaware: WideRange, 1993.
JOURN
920 www.jaacap.org
32. Standard Occupational Classification. London: Office of Popula-tion Censuses and Surveys, Her Majesty’s Stationary Office; 2010.
33. Manoach DS. Prefrontal cortex dysfunction during workingmemory performance in schizophrenia: reconciling discrepantfindings. Schizophr Res. 2003;60:285-298.
34. Karlsgodt KH, Glahn DC, van Erp TG, et al. The relationshipbetween performance and fMRI signal during working memoryin patients with schizophrenia, unaffected co-twins, and controlsubjects. Schizophr Res. 2007;89:191-197.
35. Potkin SG, Turner JA, Brown GG, et al. Working memory andDLPFC inefficiency in schizophrenia: the FBIRN study. SchizophrBull. 2009;35:19-31.
36. Glahn DC, Kim J, Cohen MS, Poutanen VP, Therman S, Bava S.Maintenance and manipulation in spatial working memory: dis-sociations in the prefrontal cortex. Neuroimage. 2002;17:201-213.
37. Botvinick M, Nystrom LE, Fissell K, Carter CS, Cohen JD. Conflictmonitoring versus selection-for-action in anterior cingulate cor-tex. Nature. 1999;402:179-181.
38. Kerns JG, Cohen JD, MacDonald AW 3rd, Cho RY, Stenger VA,Carter CS. Anterior cingulate conflict monitoring and adjust-ments in control. Science. 2004;303:1023-1026.
39. Scherf KS, Sweeney JA, Luna B. Brain basis of developmentalchange in visuospatial working memory. J Cogn Neurosci. 2006;18:1045-1058.
40. Meyer-Lindenberg A, Poline JB, Kohn PD, et al. Evidence forabnormal cortical functional connectivity during working mem-ory in schizophrenia. Am J Psychiatry. 2001;158:1809-1817.
41. Zhou B, Tan C, Tang J, Chen X. Brain functional connectivity offunctional magnetic resonance imaging of patients with early-onset schizophrenia. Zhong Nan Da Xue Xue Bao Yi Xue Ban.2010;35:17-24.
42. Konrad K, Neufang S, Thiel CM, et al. Development of attentionalnetworks: an fMRI study with children and adults. NeuroImage.2005;28:429-439.
43. Durston S, Davidson MC, Tottenham N, et al. A shift from diffuseto focal cortical activity with development. Dev Sci. 2006;9:1-8.
44. Bunge SA, Crone EA. Neural correlates of the development ofcognitive control. In: Neuroimaging in Developmental ClinicalNeuroscience. Rumsey JM, Ernst M (eds). Bethesda, MD: Cam-bridge University Press. 2009:22-37
45. Kelly AM, Di Martino A, Uddin LQ, et al. Development ofanterior cingulate functional connectivity from late childhood toearly adulthood. Cereb Cortex. 2009;19:640-657.
46. Giedd JN, Jeffries NO, Blumenthal J, et al. Childhood-onsetschizophrenia: progressive brain changes during adolescence.Biol Psychiatry. 1999;46:892-898.
47. Arango C, Moreno C, Martínez S, et al. Longitudinal brainchanges in early-onset psychosis. Schizophr Bull. 2008;34:341-353.
48. Keefe RS, Silva SG, Perkins DO, Lieberman JA. The effects ofatypical antipsychotic drugs on neurocognitive impairment inschizophrenia: a review and meta-analysis. Schizophr Bull. 1999;25:201-222.
49. Honey GD, Bullmore ET, Soni W, Varatheesan M, Williams SC,Sharma T. Differences in frontal cortical activation by a workingmemory task after substitution of risperidone for typical antipsy-chotic drugs in patients with schizophrenia. Proc Natl Acad SciUSA. 1999;96:13432-13437.
50. Stephan KE, Magnotta VA, White T, Arndt S, Flaum M, O’LearyDS, Andreasen NC. Effects of olanzapine on cerebellar functional
connectivity in schizophrenia measured by fMRI during a simplemotor task. Psychol Med. 2001;31:1065-1078.
AL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012
e; x � sagittal plane; y � coronal plane; z � axial plane.
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012 920.e1www.jaacap.org
KYRIAKOPOULOS et al.
FIGURE S1 Graphs represent correlations for each diagnostic group between age and mean signal change in thedorsolateral prefrontal cortex and the anterior cingulate cortex.
JOURNAL OF THE AMERICAN ACADEMY OF CHILD & ADOLESCENT PSYCHIATRY
VOLUME 51 NUMBER 9 SEPTEMBER 2012920.e2 www.jaacap.org