Page 1
ABSTRACT
KILEY, ERIN REBECCA. Impact of Ruminant Digestion on Germination of Ingested Seeds
of Native Warm-Season Grasses and Agronomic Weed Species in the Southeast United
States. (Under the direction of Dr. Lori Unruh-Snyder).
Dissemination of seeds by livestock has been discussed as a means of spreading both
desired and problematic plant species, across grasslands, pastures and agronomic areas
through various means. Excretion of seeds and spread of manure as a fertility amendment can
account for colonization of new plants. The dispersal of high nutritional-quality native
grasses is of value to grassland conservation and restoration, and to livestock producers as
use for forage. Likewise, problematic weed species can be spread through means of livestock
fertility amendments and supplemental feed. Seeds which retain viability subsequent to
ruminant digestive tract exposure can be vital to the process of introducing new desirable
forage species and enhancing the spread of established species. The objective of this study
was to examine the effects of rumen and intestinal digestion on seed germination of native
warm-season grasses and particularly problematic agronomic weed species in the Southeast
U.S. affecting crop yield. Two experiments were conducted to determine how ruminant
digestion influences seed viability and germination on native warm-season grasses for
potential forage, biomass and restoration use and on problematic weeds in agronomic
settings. Experiment one was to determine the impact of rumen exposure and digestion on
seed viability and germination. Seeds were treated to in situ (live animal) rumen digestion in
a cannulated steer for 48 hours. Experiment two was to examine the effect of small intestine
exposure and digestion to determine impact on seed viability and germination and potential
Page 2
points of digestion which cause or impact seed mortality. Seeds were treated to in vitro small
intestine digestion for 24 hours. Seed germination varied by plant species and response to
respective treatment. The combination of rumen phase and small intestine procedure (SIP)
exposure (abomasal-simulated digestion) resulted in highest seed mortality. Native-warm
season grass seed, such as switchgrass (Panicum virgatum L.) and big bluestem (Andropogon
gerardii L.), retained viability but germination after digestion exposure was reduced more
than agronomic weed species. Switchgrass mean germination percentage was reduced by
86.70% and big bluestem mean germination percentage was reduced by 79.59% with
exposure to rumen phase plus SIP. Troublesome agronomic weed seed, such as Palmer
amaranth (Amaranthus palmeri L.) and Texas panicum (Panicum texanum L.), retained
viability but germination declined with digestion exposure. Palmer amaranth mean
germination percentage was reduced by 53.77% and Texas panicum mean germination
percentage was reduced by 56.83% with exposure to rumen phase plus SIP, which was the
highest rate of germination. Herbivory is a key process in range and grassland ecosystems
and it is imperative to consider the role of herbivores as part of a holistic system and utilize
this understanding to improve management approaches. Further research is needed in this
area to determine and to understand how grazing animals, including cattle and other species,
can impact the dissemination of seeds and establishment of plant populations.
Page 3
© Copyright 2014 by Erin R. Kiley
All Rights Reserved
Page 4
Impact of Ruminant Digestion on Germination of Ingested Seeds of Native Warm-Season
Grasses and Agronomic Weed Species in the Southeast United States
by
Erin Rebecca Kiley
A thesis submitted to the Graduate Faculty of
North Carolina State University
in partial fulfillment of the
requirements for the degree of
Master of Science
Crop Science
Raleigh, North Carolina
2014
APPROVED BY:
_______________________________ ______________________________
Dr. Lori Unruh-Snyder Dr. Wesley J. Everman
Committee Chair
________________________________
Dr. Randy Wells
Page 5
ii
DEDICATION
Thank you to my wonderful family: Richard, Rebecca and Shannon and for the support from
my friends, community and colleagues.
I dedicate this work to the generations of farmers that have preceded me: my father and
grandfather. Thank you to my father for his unwavering love, support and guidance, making
this pursuit and all others possible. Thank you to my grandfather for serving as my first
beautiful example of how to be a positive steward of the land.
Thank you to Dr. Charles Sydnor for generously providing me the opportunity to start
farming, nurture my interest in regenerative land management, break conventions, know my
own strength, to find a home at Braeburn Farm and to fall in love with cows.
Page 6
iii
BIOGRAPHY
Erin Kiley is from Chesapeake, Virginia and is the eldest daughter of Richard and Rebecca
Kiley and sister to Shannon. Erin’s grandparents, Richard and Barbara Kiley, own and
operate a beef cattle farm in northeast Missouri, where she first was introduced to
sustainable, holistic agriculture. In 2009, Erin graduated from the University of Virginia in
Charlottesville, Virginia with a Bachelor of Arts degree in Foreign Affairs. Following
graduation, Erin worked for United States Agency for International Development project,
Health Systems 20/20, working to strengthen international health systems in developing
countries and managing the Tanzania portfolio. Erin continues to maintain involvement in
the field of international development as a consultant for the Global Fund to Fight AIDS,
Tuberculosis and Malaria based in Geneva, Switzerland. She moved to North Carolina in
August of 2011 to live and work on a sustainable grass fed beef farm in Alamance County
utilizing intensive nature-mimicking rotational grazing and holistic management. Erin
decided to pursue her M.S. degree under the direction of Dr. Lori Unruh-Snyder in Crop
Science with a focus in Grassland Ecology and hopes to pursue a career in sustainable,
holistic land management and regenerative resource conservation.
Page 7
iv
ACKNOWLEDGEMENTS
Thank you to my committee chair, Dr. Lori Unruh-Snyder for providing me the research
opportunity and to my advisory committee Dr. Wesley Everman and Dr. Randy Wells for
their expert advice and guidance. A special thank you to Dr. Michelle Schroeder-Moreno for
believing in my potential from the beginning and for her continued role as mentor. Thank
you to North Carolina faculty and staff: Brenda Penny, Dr. Paul Mueller, Dr. Jean-Marie
Luginbuhl, Dr. Matt Poore and Jordan Bakerman.
Page 8
v
TABLE OF CONTENTS
LIST OF TABLES .................................................................................................................. vii
LIST OF FIGURES ............................................................................................................... viii
INTRODUCTION .................................................................................................................... 1
REVIEW OF LITERATURE ................................................................................................... 4
Herbivory and Impact on Grassland and Pasture Ecology ................................................... 4
Plant and Animal Interactions and Impact on Seed Dispersal .............................................. 4
Seed Germination and Dormancy ......................................................................................... 6
Seed Passage Through Ruminant Gastrointestinal Tract and Impact on Seed Viability and
Germination .......................................................................................................................... 9
Seed Physiology and Impact on Ruminant Digestive Tract Passage .................................. 12
Effect of Ruminant Digestive Tract on Native Warm-Season Grasses .............................. 15
Native Warm-Season Grass Species ................................................................................... 18
Switchgrass ..................................................................................................................... 18
Big Bluestem ................................................................................................................... 20
Effect of Ruminant Digestive Tract on Agronomic Crops ................................................. 23
Weed Species ...................................................................................................................... 24
Palmer Amaranth ............................................................................................................ 24
Texas Panicum ................................................................................................................ 27
Literature Cited ................................................................................................................... 30
IMPACT OF RUMINANT DIGESTION ON GERMINATION OF INGESTED SEEDS OF
NATIVE WARM-SEASON GRASSES IN THE SOUTHEAST UNITED STATES .......... 38
Introduction ......................................................................................................................... 40
Materials and Methods ........................................................................................................ 44
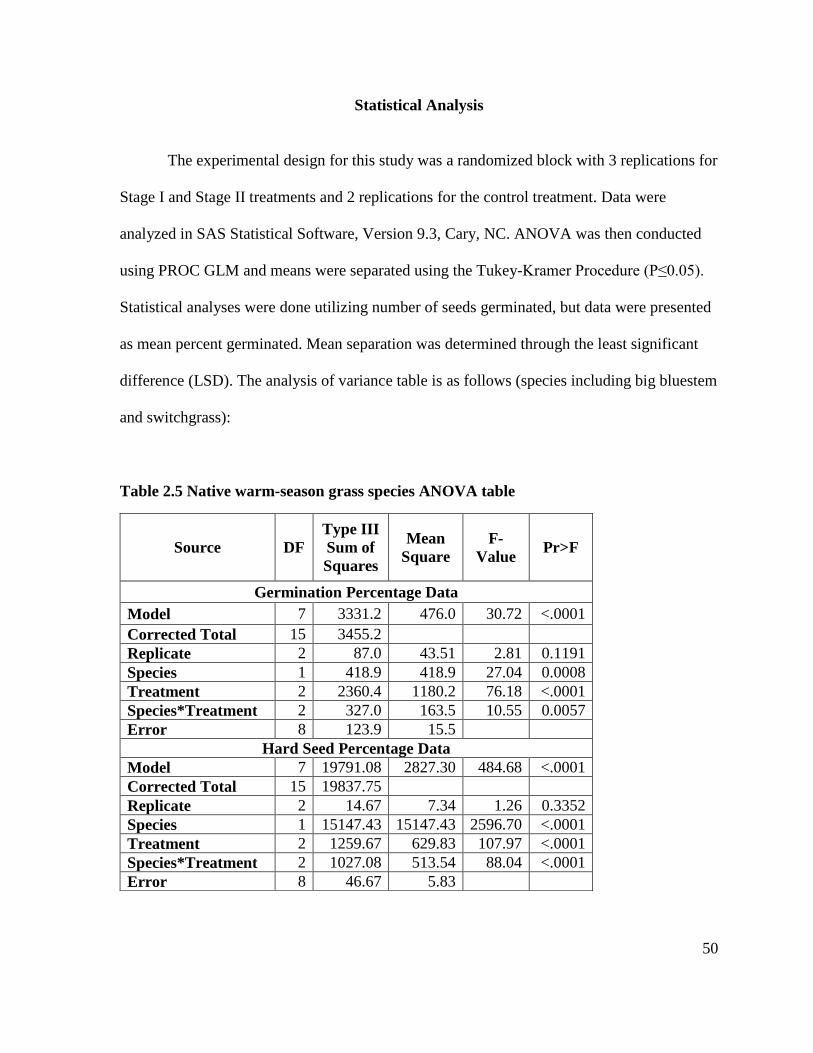
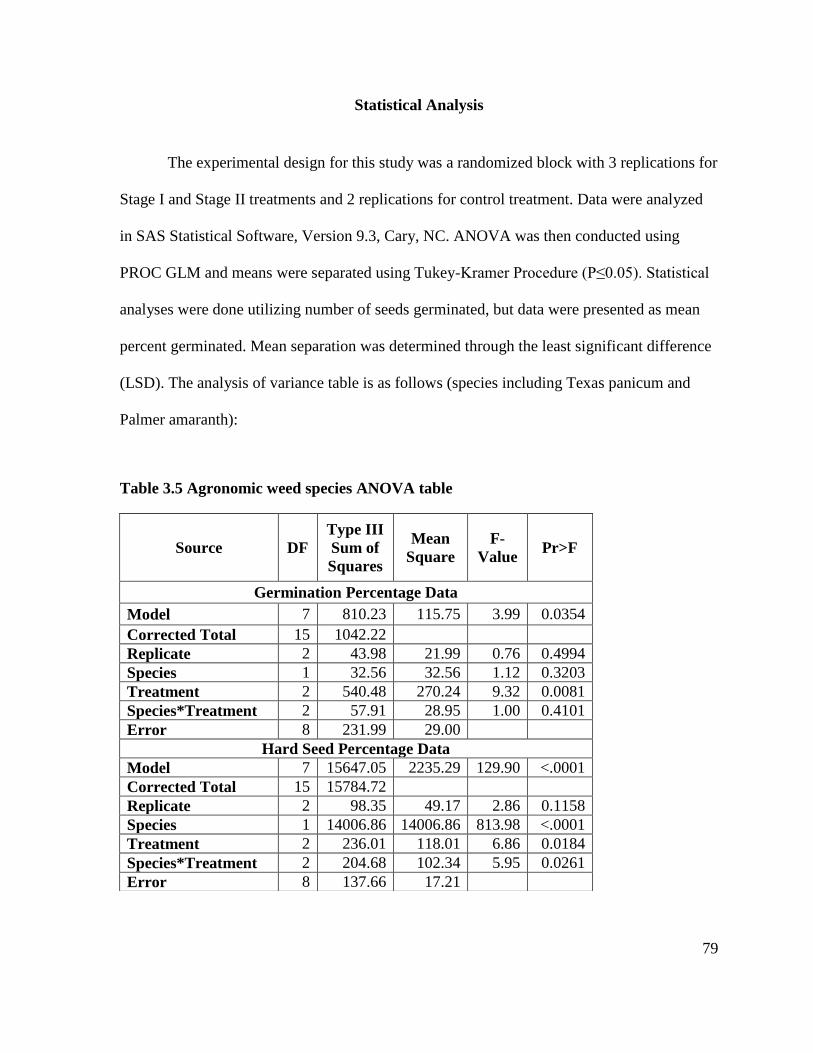
Statistical Analysis .............................................................................................................. 50
Results ................................................................................................................................. 51
Treatment Main Effect .................................................................................................... 52
Discussion ........................................................................................................................... 56
Page 9
vi
Management Implications ............................................................................................... 60
Literature Cited ................................................................................................................... 63
IMPACT OF RUMINANT DIGESTION ON GERMINATION OF INGESTED SEEDS OF
AGRONOMIC WEED SPECIES IN THE SOUTHEAST UNITED STATES ..................... 67
Introduction ......................................................................................................................... 69
Materials and Methods ........................................................................................................ 73
Statistical Analysis .............................................................................................................. 79
Results ................................................................................................................................. 80
Discussion ........................................................................................................................... 84
Management Implications ............................................................................................... 85
Literature Cited ................................................................................................................... 90
APPENDICES ........................................................................................................................ 95
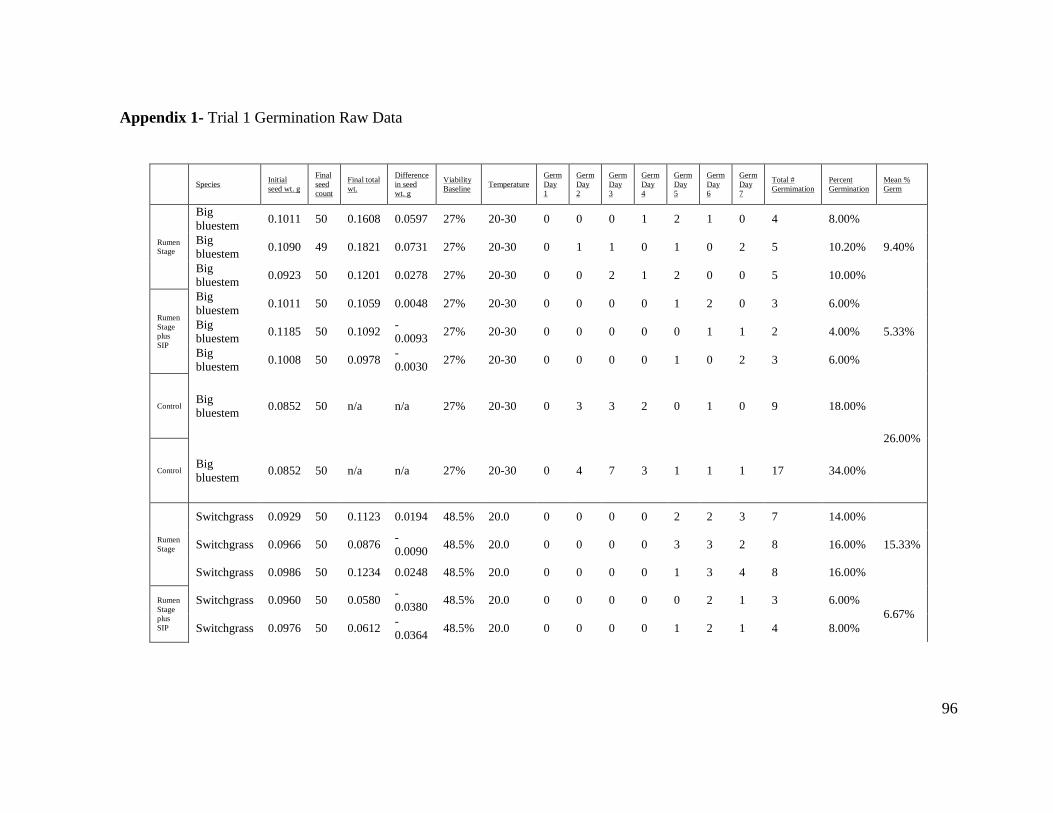
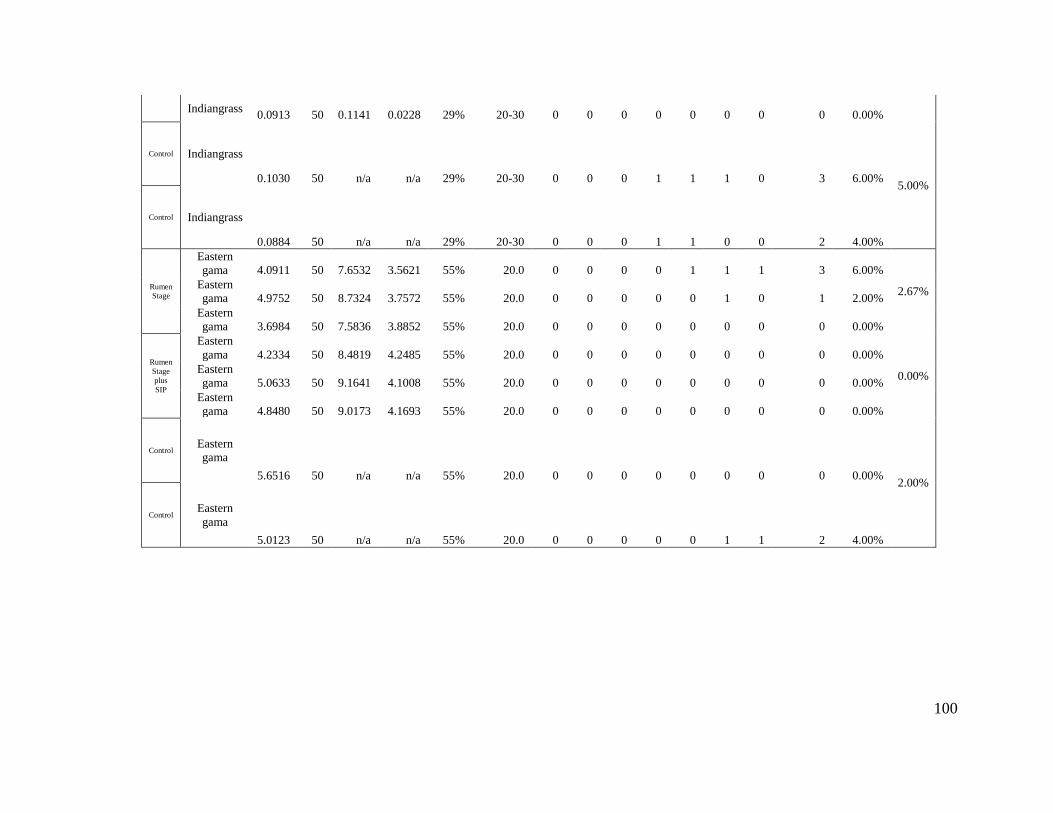
Appendix 1- Trial 1 Germination Raw Data....................................................................... 96
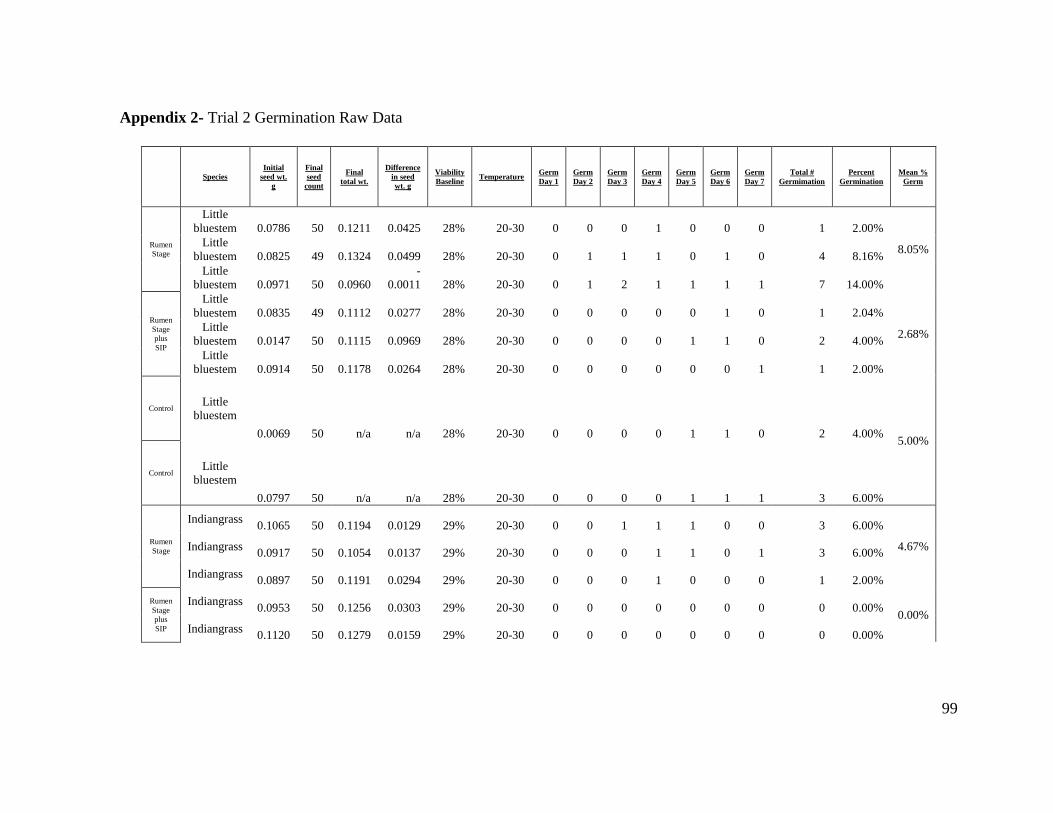
Appendix 2- Trial 2 Germination Raw Data....................................................................... 99
Page 10
vii
LIST OF TABLES
Table 2.1 Native warm-season grass species germination temperature and seed size ........... 44 Table 2.2 Thirty year averages and 2013 growing season averages for air temperature and
total precipitation in Upton, Kentucky.................................................................................... 44
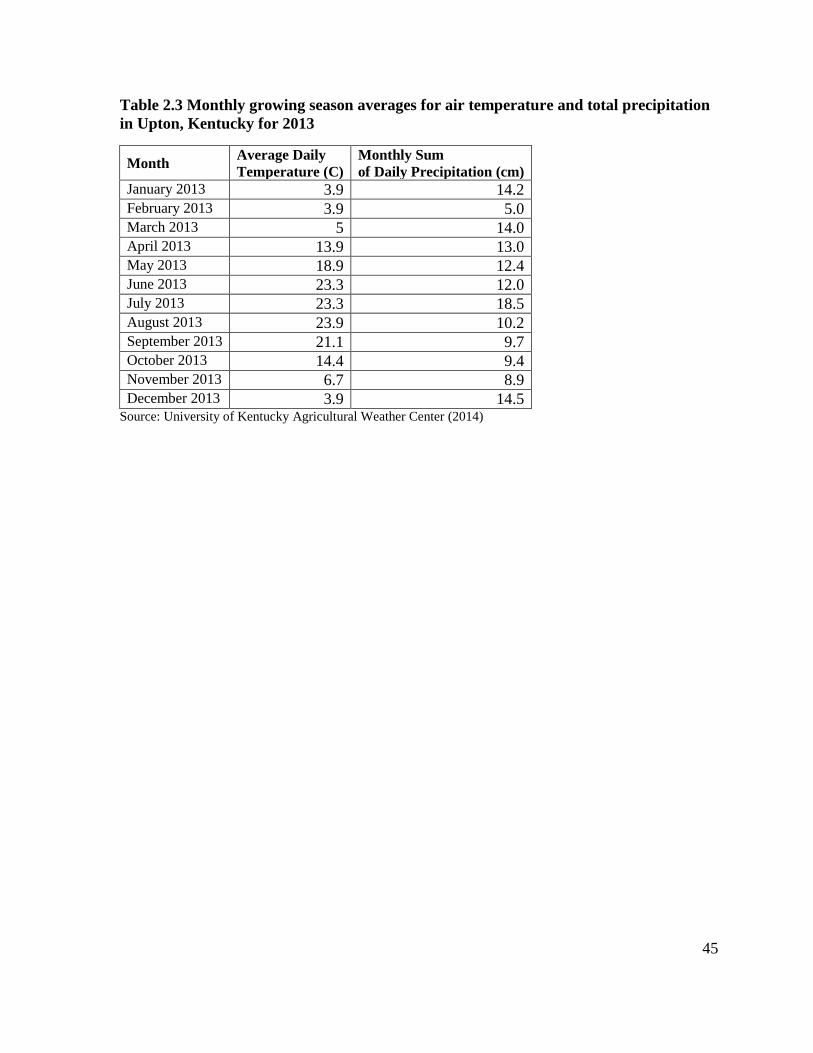
Table 2.3 Monthly growing season averages for air temperature and total precipitation in
Upton, Kentucky for 2013 ...................................................................................................... 49 Table 2.4 Switchgrass and big bluestem seed viability, hard seed, dormant seed and control
germination mean.................................................................................................................... 49 Table 2.5 Native warm-season grass species ANOVA table .................................................. 50
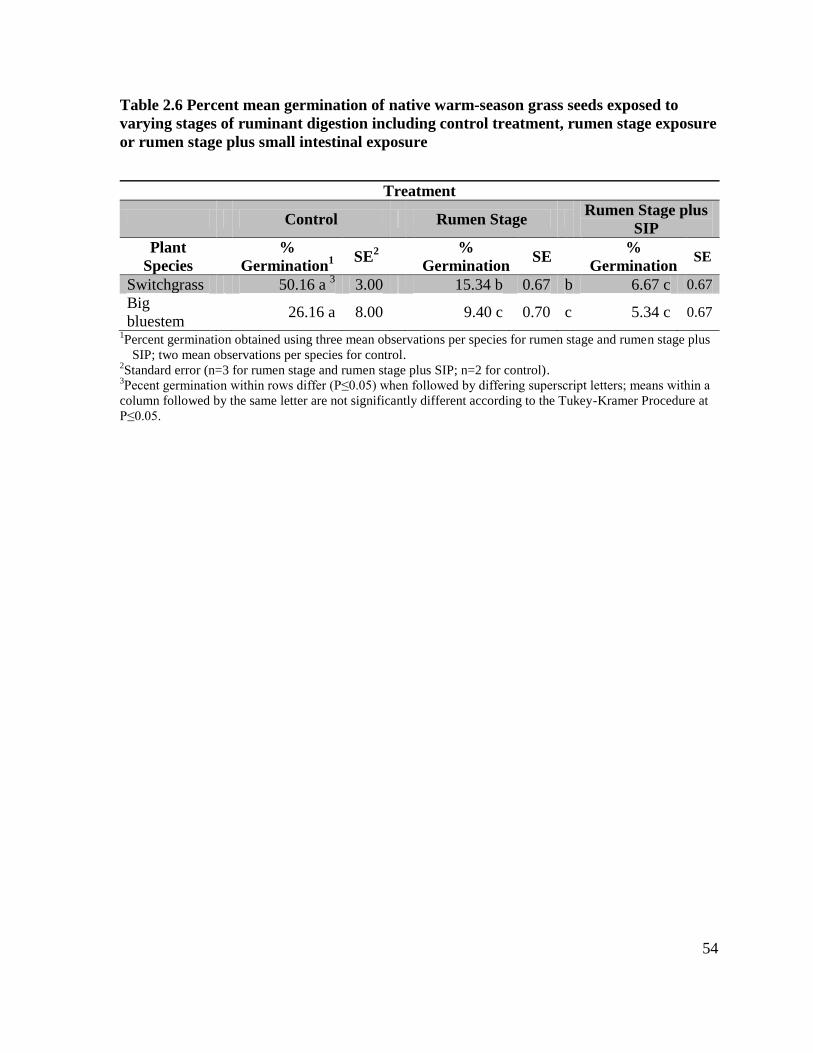
Table 2.6 Percent mean germination of native warm-season grass seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure ................... 55
Table 3.1 Agronomic weed species germination temperature and seed size .......................... 73
Table 3.2 Thirty year averages and 2013 growing season averages for air temperature and
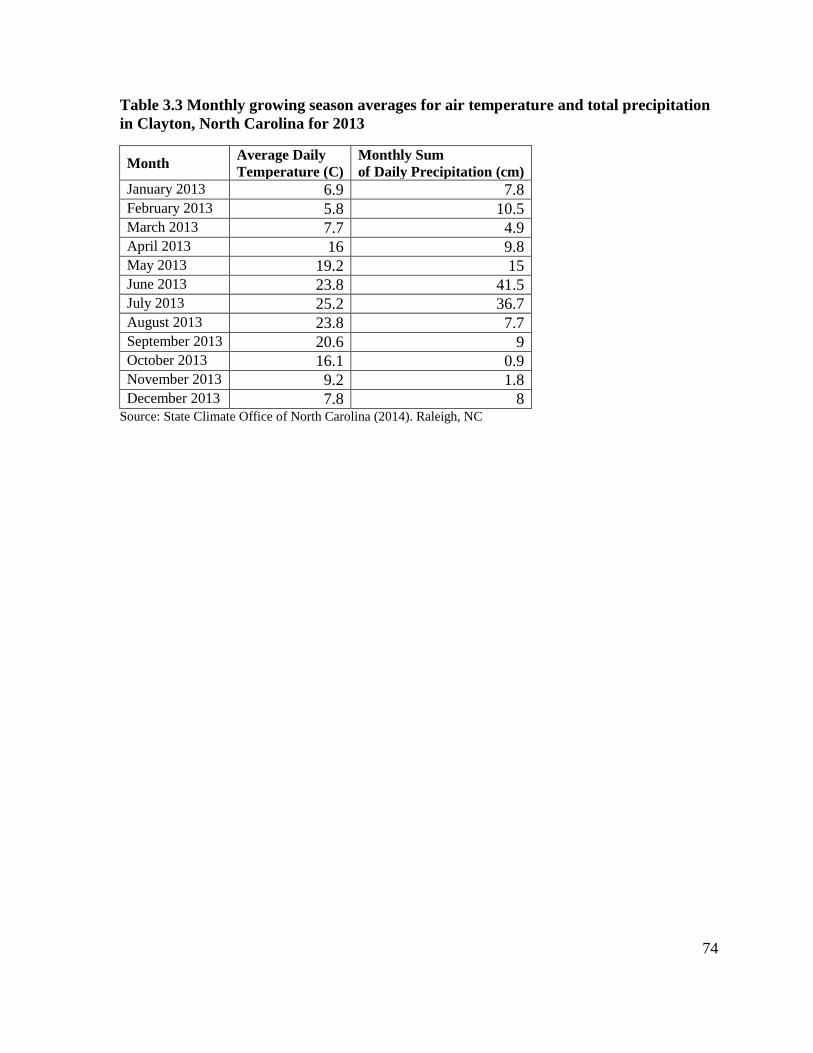
total precipitation in Clayton, North Carolina ........................................................................ 73 Table 3.3 Monthly growing season averages for air temperature and total precipitation in
Clayton, North Carolina for 2013 ........................................................................................... 74 Table 3.4 Texas panicum and Palmer amaranth seed viability, hard seed, dormant seed and
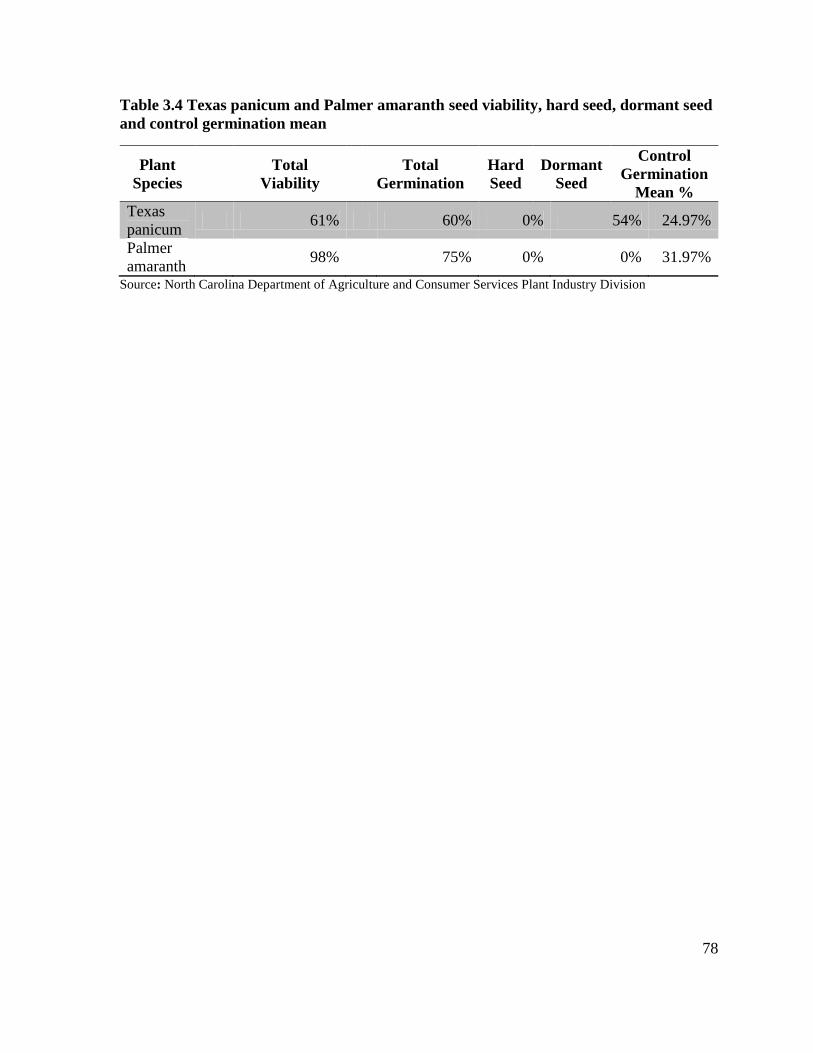
control germination mean ....................................................................................................... 78
Table 3.5 Agronomic weed species ANOVA table ................................................................ 79 Table 3.6 Percent mean germination of agronomic weed seeds exposed to control treatment,
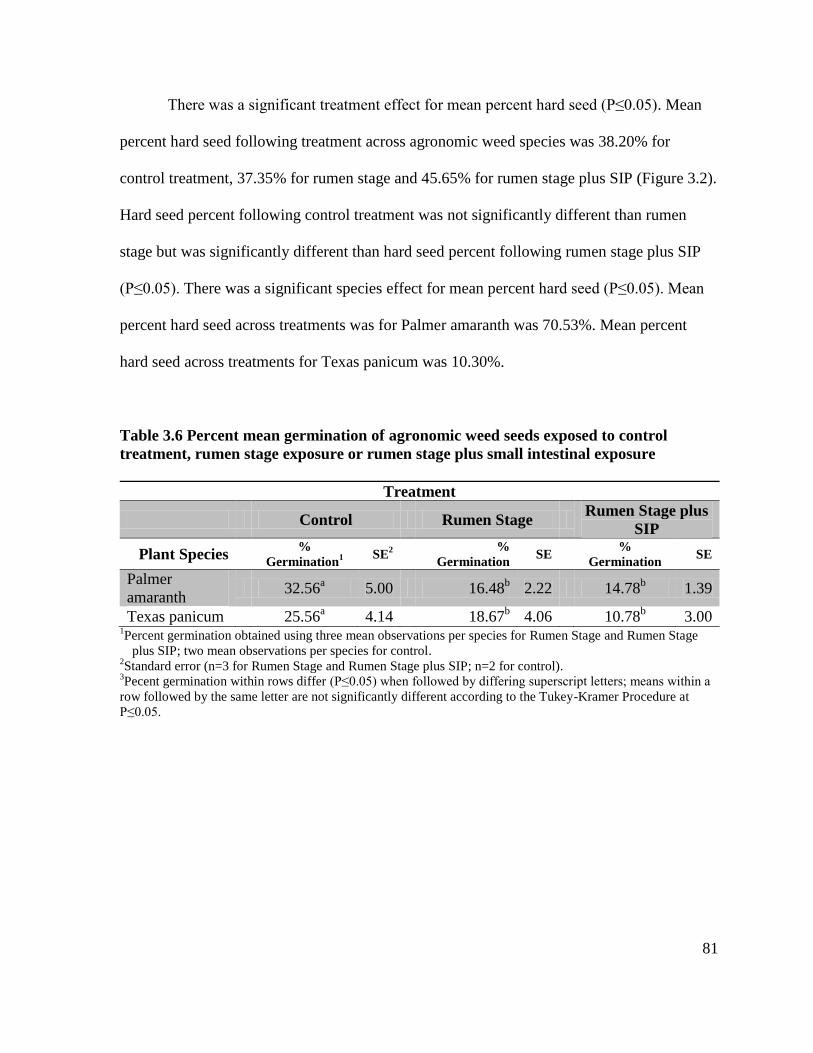
rumen stage exposure or rumen stage plus small intestinal exposure .................................... 81
Page 11
viii
LIST OF FIGURES
Figure 2.1 Diagram of longitudinally bisected seed and embryo exposed to tetrazolium
testing .......................................................................................................................................49
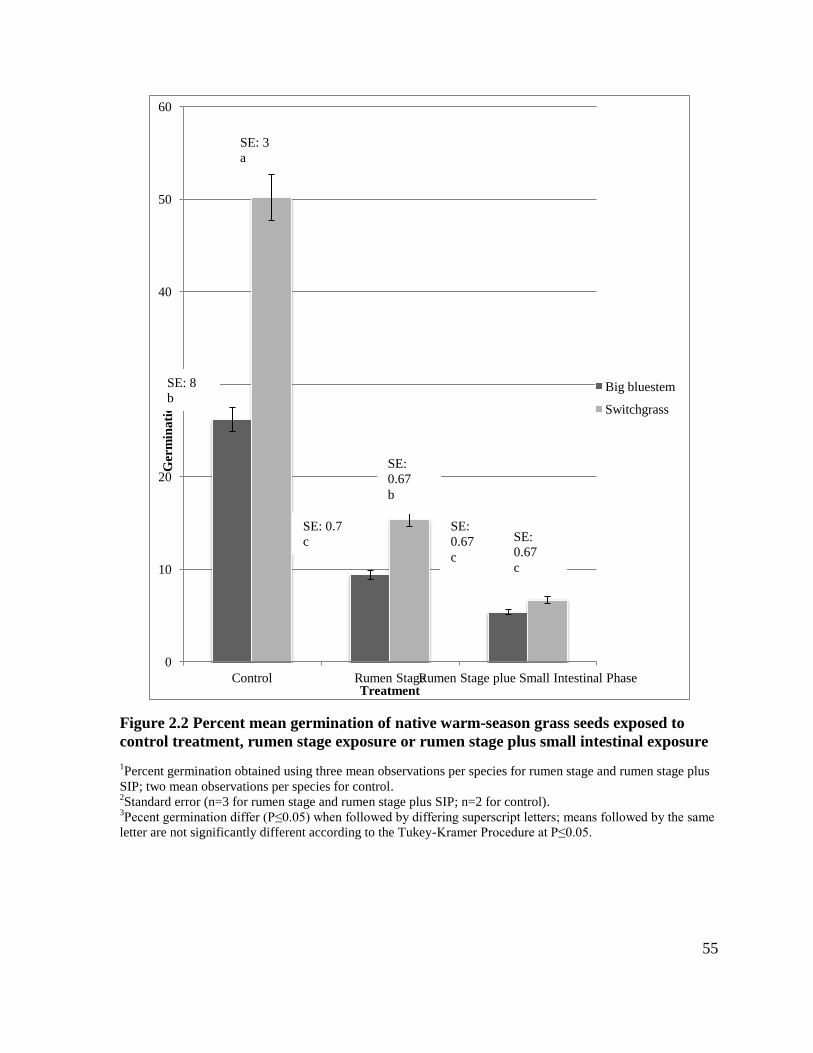
Figure 2.2 Percent mean germination of native warm season-grass seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure ....................55
Figure 2.3 Percent mean hard seed of native warm-season grass seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure ....................56
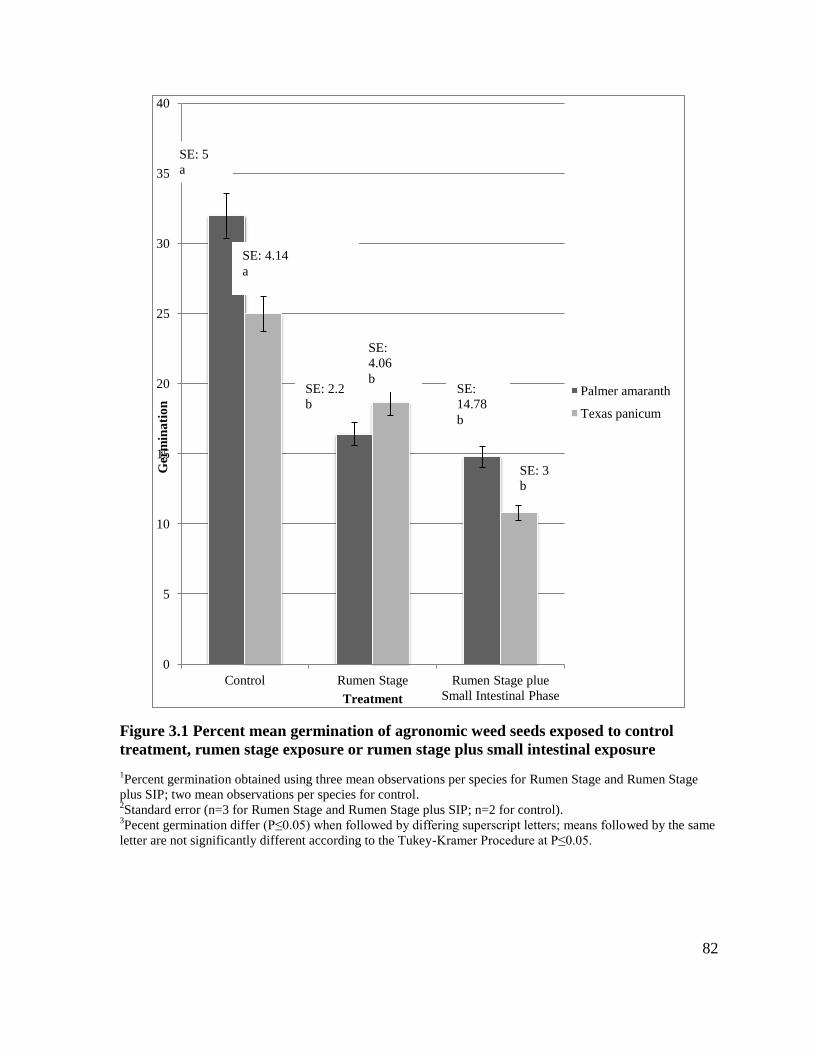
Figure 3.1 Percent mean germination of agronomic weed seeds exposed to control treatment,
rumen stage exposure or rumen stage plus small intestinal exposure .....................................82
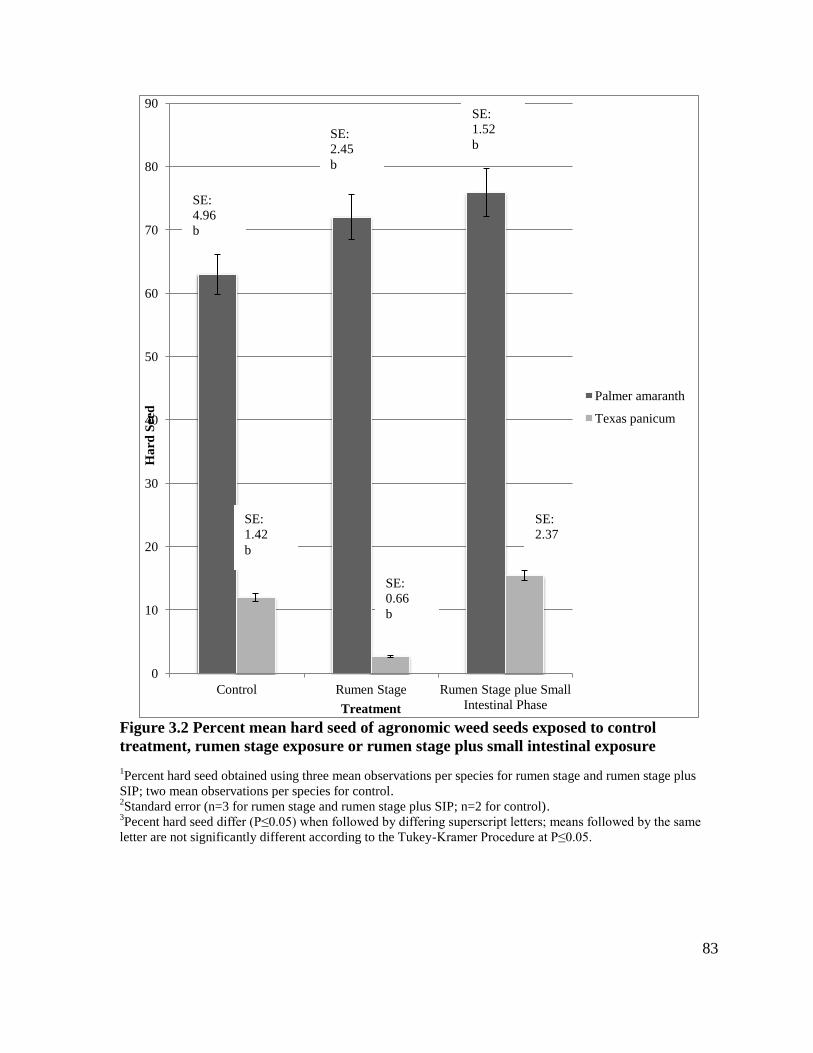
Figure 3.2 Percent mean hard seed of agronomic weed seeds exposed to control treatment,
rumen stage exposure or rumen stage plus small intestinal exposure .....................................83
Page 12
1
INTRODUCTION
Dissemination of seeds by livestock has been discussed as a means of spreading both
desired and problematic plant species, across grassland pastures and unexposed agronomic
areas through various means (Grubb, 1977). Excretion of seeds and spread of manure as a
fertility amendment can account for colonization of new plants (Grubb, 1977; Lowry, 1996).
Seeds which retain viability subsequent to ruminant digestive tract exposure can be vital to
the process of introducing new desirable forage species and enhancing the spread of
established species (Blackshaw and Rode, 1991; Lowry, 1996). Further research is needed in
this area to determine and to understand how grazing animals, including cattle, can impact
the dissemination of seeds and establishment of plant populations.
The dispersal of high nutritional-quality native grasses, often characterized by high
rates of seed dormancy, is of value to grassland, rangeland and prairie conservation and
restoration, and livestock producers as sward components and forage (Janzen, 1984).
Likewise, problematic weed species can similarly be spread through means of livestock
manure amendments and supplemental feed, such as cotton (Gossypium hirsutum L.) gin
trash or corn (Zea mays L.) and soybean (Glycine max L.) products (Norsworthy, et al.,
2009), negatively impacting yield of agronomic crops grown in subsequently contaminated
areas (Byrd et al., 2004). When exposed to ruminant ingestion, digestion, and excretion,
seeds viability, germination and successful disperals are influenced (Norsworthy, et al.,
2009). Some seeds will remain viable, or germinable, post-ruminant ingestion and digestion,
however, viability can vary widely across species (Blackshaw and Rode, 1991). According to
Page 13
2
Gardener et al. (1993), differences in survival rates and seed dissemination success after
digestion are related to specific seed characteristics, including length of time spent in the
digestive tract with less dense seeds having the longest retention time in cattle. Seed species
examined by Gardener et al. (1993) included signal grass (Brachiaria decumbens Stapf. cv.
Basilisk), green panic grass (Panicum maximum Jacq. var. trichoglume Eyles cv. Petrie ) and
glycine (Neonotonia wightii (Arn.) Lackey cv. Tinaroo). Over a 160 hour period, seeds
became especially fragmented after 70 hours in the ruminant digestive tract.
The objective of this study was to examine the effects of rumen and intestinal
digestion on seed germination of dormancy-prone native warm-season grasses and
particularly problematic agronomic weed species. Two experiments were conducted to
determine how ruminant digestion influences seed viability and germination on native warm-
season grasses for potential forage, biomass, restoration and on problematic weeds in
agronomic settings. An experiment was conducted to determine the impact of rumen
exposure and digestion on seed viability and germination. Seeds were treated in situ (live
animal) for rumen digestion in a cannulated steer for 48 hours and compared to a control
treatment (Blackshaw and Rode, 1991). The effect of small intestine exposure and digestion
on seed viability and germination and potential points of digestion which cause or impact
seed mortality was also investigated. Seeds were treated to in vitro small intestine digestion
for 24 hours (Calsamiglia and Stern, 1995; Unruh Snyder et al., 2011). It was hypothesized
that exposure to the ruminant digestive tract would diminish viability and germination, but
still allow for a viable means of seed transport and plant community spread. It was
hypothesized that exposure to the ruminant digestive tract would cause a decrease in
Page 14
3
germination rates of native warm-season grasses and increase germination rates of agronomic
weed species.
Page 15
4
REVIEW OF LITERATURE
Herbivory and Impact on Grassland and Pasture Ecology
Herbivory is a key process in range and grassland ecosystems. Herbivores are defined
by derivation of all required nutrients from plants, and thus have direct impact on these plants
(Pieper, 1994). With consideration to livestock grazing, it is imperative to consider the role
of herbivores as part of a holistic system and utilize this understanding to improve
management approaches (Pieper, 1994). Herbivores, specifically ruminants, influence plants,
soil and grassland ecology, through the process of grazing both in a positive and negative
mechanism (Janzen, 1984). Ruminants are earth’s dominant herbivores; a fundamental
reason for this is the evolution of a digestion method which utilizes microorganisms to
breakdown plant material. However, ruminant-produced enzymes cannot fully breakdown
these materials (Janzen, 1984). Many forage grasses are more successful when regularly
grazed which suggests co-evolution with grazing herbivores (Owen, 1980). Grazing
ruminants influence pasture composition and plant communities through seed dispersal by
ingestion, digestion, excretion and dissemination of seed (Gardener et al., 1993).
Plant and Animal Interactions and Impact on Seed Dispersal
Historically, herbivory and inherent ingestion of plant seeds has occurred for millions
of years. Evidence of this appeared as far back as the period of the Miocene epoch as
demonstrated by grass seed discovered in the fossilized digestive system contents of extinct
species, Teloceras major (Jansen, 1984). Distribution of viable seeds following digestion has
Page 16
5
been observed across a vast variety of ruminants and other mammals, including cows, sheep,
goats, bats, lemurs, gorillas, foxes, rhinoceros and elephants. Other seed-dispersers include
tortoises, lizards, fish, and avian species, thrushes, waxwings and warblers (Herrera, 2002).
Many plants and forage grasses have evolved adaptations which can utilize animals as
a method of dispersal, both internally and externally, benefiting plants by reducing parent
plant-offspring competition for resources and improved germination conditions (Gardener et
al., 1993; Stiles, 1992; Herrera, 2002). With consideration to the high numbers and
concentration of livestock on range and grassland, utilizing livestock as a means of seed
dispersal should be a considered method. Ungulates ingest and disperse seeds in sync with
the ingestion of leaf foliage (Janzen, 1984) However, there are relatively few attempts by
land managers to utilize livestock as a resource of dissemination for desirable plant
establishment and community expansion (Archer and Pyke, 1991).
Seed characteristics that contribute to seed ingestion and survivability post-digestion
include plants of high palatability, digestibility (low neutral detergent fiber [NDF]) and
nutrient content (crude protein [CP]), seeds located within close proximity to leaf matter and
small, hard seeds (Janzen, 1984; Ball et al., 2001). Welch (1985) found 662 seedlings of 24
plant species germinating from a single pat of cow manure in greenhouse conditions.
Included in these species was sweet vernal grass (Anthoxanthum odoratum L.), velvet grass
(Holcus lanatus L.), annual bluegrass (Poa annua L.) and Kentucky bluegrass (Poa pratensis
L.). Studies reported high numbers of germinated seeds and seedlings of Prosopis spp. and
Acacia spp. which emerge from manure of ungulates. This was observed on four continents
and spread of these plant species provided evidence in support of the effectiveness of
Page 17
6
livestock in seed dispersal (Archer and Pyke, 1991). Results from Simao Neto et al. (1987)
demonstrate that cattle excrete the highest number of undigested seeds when compared to
sheep or goats.
Among plant species spread through means of ruminant ingestion and excretion are
noxious and invasive weeds, or species having the potential to invade and degrade U.S.
landscapes (USDA, 2013). Germination rates and area of spread, in some cases, may be
enhanced through digestive tract exposure for weed species including redroot pigweed
(Amaranthus retroflexus L.), common lambsquarters (Chenopodium album L.) and wild
buckwheat (Polygonum convolvulu L.), (Simao Neto et al., 1987; Blackshaw and Rode,
1991). Subject to speed of passage through the digestive system, seeds can be disseminated
across area in which animals cover over a span of several days (Al-Mashilchi, 1993). Viable
seeds have been reported to have been recovered in manure pats 6 days post-ingestion with
some species viable for up to 30 months (McCully, 1951; Simao Neto et al., 1987; Gardener
et al., 1993).
Seed Germination and Dormancy
Growth of an embryo, or germination, is dependent on several external and internal
factors. Important external factors include presence of water, oxygen, and environmental
temperature. Germination requires the seed to imbibe water to proceed with metabolic
activities necessary for growth. Enzymes within the seed become activated and others
synthesized to allow for utilization of stored carbohydrates, lipids and proteins which support
cell division and growth. With the imbibition of water, the seed swells and pressure develops
Page 18
7
and causes the seed coat to rupture, thus allowing the seed to access oxygen for aerobic
respiration. In an aqueous environment such as the rumen, the seed may be prevented to
access the necessary amount of oxygen and prevent growth (Gardener et al., 1993). Seed
dormancy is the failure of a viable seed to germinate successfully, despite favorable
conditions (Bewley 1997). Baskin and Baskin (2004) suggest dormant seed to lack the
capacity to germinate within a specified period of time under normal environmental
conditions which are otherwise favorable for germination.
According to Gardener et al. (1993) embryo physiological immaturity can cause seed
dormancy and prevent germination, despite favorable conditions. Inherently dormant seeds
may need to undergo enzymatic and biochemical processes prior to germination, often
triggered by low temperatures (vernalization) or passage through digestive tracts. As an
evolutionary benefit, dormancy ensures favorable external environmental conditions for
germination. Seeds which are consumed by ruminants are subjected to imbibition of water.
Ruminal enzymes may additionally be absorbed by seeds causing adverse effects to the seed
embryo. The anaerobic environment of the rumen does not allow for seed absorption of
oxygen and gaseous exchange and can cause dormancy or non-viability (Simao Neto and
Jones, 1986). Seed dormancy is noted as a major challenge associated with restoration
plantings of native warm-season grasses (NWSG) (USDA, 2007). Grassland and prairie
restoration experts suggest that plant species native to an area should ideally be utilized in
conservation projects to achieve long-term sustainability (USDA, 2007).
Warm-season grasses are often subject to dormancy occurring in one or more of the
tissues within the seed or embryo. The seed coat can prevent successful germination through
Page 19
8
inhibition of water uptake, reduced gas exchange, diminished embryo expansion or
production of germination inhibitors (Adkins et al., 2002). To counteract dormancy caused
by seed coat impermeability, natural systems provide mechanisms to weaken coat and seed
tissues, including saprophytic fungi, fire, environmental conditions or scarification through
chemical and mechanical action (Adkins et al., 2002). Embryonic dormancy is thought to be
an evolutionary adaptation to grasses to enhance survival rates by avoiding unfavorable
germination conditions (Simpson, 1990). Germination processes frequently undergo
temporarily cessation and related seed dormancy ensues when exposed to drought, non-ideal
temperatures, fire, frost or flooding (Finch-Savage and Leubner-Metzger, 2006). Seed of big
bluestem (Andropogon gerardii Vitman) and switchgrass (Panicum virgatum L.) exhibit
dormancy post-harvest, defined by an increase in germination rate following seed treatment.
There has been substantial research conducted on dormancy-breaking and stratification
techniques for NWSG, including switchgrass (Sanderson et al., 1996). Traditional methods
of breaking seed dormancy for species characterized by inherent dormancy include
prechilling, storage and dormant seeding. Prechilling is the process of breaking seed
dormancy through exposing seed to artificially produced cold, wet treatment conditions in
order mimic natural winter conditions and processes (Wolfe and Fiske, 2009). According to
Beckman et al. (1993), moistened and wet-chill (4° C) treatments enhanced seedling
emergence of switchgrass by 35 and 150%, respectively. Indiangrass (Sorghastrum nutans
L.) exposed to moist-chill treatment for 4 weeks at 4° C resulted in near maximum
germination (Emal and Conard, 1973; Olivier, 2006). Ambient temperature storage used to
break dormancy can take 2 to 4 years to achieve adequate germination rates and could result
Page 20
9
in reduced seed viability after time. Unstratified seed of NWSG stored for 2 years have the
highest percentage of germinated seed (Owsley, n.d.). Subjection to 24 months of dry, cool
condition storage (13° C) increased switchgrass germination. Seeds treated with sodium
hypochlorite (NaClO) increased switchgrass germination rates by 61% (Haynes et al., 1997).
Planting dormant seed in November or December will allow seed to naturally be exposed to a
cold, wet treatment. Issues with this method include environmental condition variability and
subsequent inconsistent dormancy release and germination, as well as high weed competition
(Beckman et al., 1993).
Seed Passage Through Ruminant Gastrointestinal Tract and Impact on Seed Viability
and Germination
The process of seed dispersal and spread of plant communities can be important to
consider for matters of plant community restoration, management and control of weed
populations and is impacted by ingestion and the digestive tract of grazing ruminants
(Gardener et al., 1993). According to Gardener et al. (1993), grazing herbivores can be
utilized to initiate the propagation of desirable species more sustainably into areas unsuitable
or unreachable through conventional sowing processes. Conversely, grazing herbivores also
influence the spread of undesirable or invasive weeds to uncontaminated areas through
manure. Previous research has confirmed that seeds of species Brachiaria decumbens L.,
Axonopus afinis L., Neonotonia wightii cv. Tinaroo, Trgfolium semipilosum cv. Safari,
Stylosanthes hamata cv. Verano and S. scabra cv. Seca are capable of maintaining viability
and germinating following ruminant digestion and excretion, and in some cases, may be a
Page 21
10
crucial tool for expanding and maintaining plant communities (Simao Neto and Jones, 1986;
Simao Neto et al., 1987). Damage to ingested seeds passing through the digestive tract has
been monitored and it has been concluded that a high percentage of damage or deterioration
occurs through rumination in the rumen and to a lesser extent in the abomasum (Simao Neto
and Jones, 1986; Gardener et al., 1993). According to Simao Neto and Jones (1986), signal
grass seed viability was reduced to 0% after 96 hours of rumen exposure. Seed retention time
within the rumen and rate of passage may additionally impact viability depending on degree
of imbibition with subsequent exposure to ruminal conditions with respect to differing seed
coat hardness (Blackshaw and Rode, 1991; Gardener et al., 1993). Hardness of seed coat
influences digestion survivability with hard seeds less likely to imbibe threatening levels of
water or be influenced by microbes and acidic ruminal conditions, suggesting that smaller,
harder seeds may allow for higher rates of survivability after digestion (Gardener et al., 1993;
Janzen, 1984). Seeds which have impermeable seed coats are referred to as hard seed.
Impermeable seeds are common Fabaceae, Cannaceae, Chenopodiaceae, Convallariaceae,
Convolvulaceae, Geraniaceae, Malvaceae and Solanaceae families (Harrington, 1916).
Hard-seeded species, redroot pigweed, passed through cattle digestive tract showed 36, 29, 4
and 3 percent survival after 47, 59, 73 and 97 hours of exposure, respectively (Atkeson et al.
1934). Viability of Ornithopus compressus L. soft seed was impaired following storage at
room humidity levels of 76% with a constant temperature of 20°C (Barrett-Lennard and
Gladstones, 1964). Soft-seeded grass species, signal grass and carpet grass (Axonopus affinis
L.) exposed to prolonged storage in manure pats for 21 days at temperatures of 35°C resulted
in loss of all viability (Simao Neto and Jones, 1986).
Page 22
11
The impact of exposure to the ruminant digestive tract on seed survival and
germination has been examined in previous studies with various grass and weed species,
however more information is needed on the impact on NWSG and certain key troublesome
weed species influencing agronomic crop yields (Blackshaw and Rode, 1991; Gardener et al.,
1993). When seed is ingested by a ruminant, it first undergoes processes of chewing,
grinding and mastication with subsequent exposure to the complete digestive tract (Gardener
et al., 1993). Differences in ruminant digestive tracts among animal species also impact the
rate of seed viability loss and germination rates with seed survival typically declining with
increased time spent in the digestive tract (Janzen, 1984; Blackshaw and Rode, 1991). Some
weed species seeds, including redroot pigweed experienced enhanced germination after 24
hours of exposure to sheep rumen and, while seed germination declined following prolonged
exposure, a greater than 50% survival rate was found after 96 hours of rumen exposure
(Haidar et al., 2010). Studies have suggested that seed hardness is a major factor responsible
for seed survival during digestive processes (Gardener et al., 1993; Haidar et al., 2010).
According to Blackshaw and Rode (1991), redroot pigweed was chosen as a species of note
for its hard seed coat when looking at the impact of rumen exposure on germination rates.
The hard seed coat of redroot pigweed was thought to aid in enhanced survival rates when
compared with other weed species.
The rumen is the largest chamber of the ruminant stomach and is a site of
fermentation. Ingested food is held in the rumen reticulum and is subjected to extensive
microbial digestion and fermentation (Gardener et al., 1993). Microbes which inhabit the
digestive tracts of ruminants are the primary agents for breaking down complex
Page 23
12
carbohydrates in forage materials. Microbes include bacteria, protozoa, yeast and fungi and
reflect the nature of the diet consumed by the animal, with diets high in cellulose and low in
starch supporting high percentages of cellulolytic and saccharolytic bacteria, while high
starch and low cellulose diets support amylolytic bacteria (Van Soest, 1994). Seeds are
exposed to anaerobic conditions and 6 to 7 pH in the rumen and large intestine followed by
exposure to an acidic environment of 2.5 pH containing proteolytic, amylolytic and lipolytic
enzymes when passed to the abomasum and small intestine. Potential abrasion or crushing
action occurs in the rumen, abomasum and small intestine (Gardener et al., 1993; Van Soest,
1994). Ingesta (ingested food), moving from the rumen through to the lower gastrointestinal
tract, is influenced by the reduction of particle size through the rumination process (Van
Soest, 1994).
Seed Physiology and Impact on Ruminant Digestive Tract Passage
Seeds are impacted by the time spent within the digestive tract and begin to be
damaged after a certain time threshold through both physical and chemical processes
(Blackshaw and Rode, 1991; Gardener et al., 1993; Van Soest , 1994). Janzen (1984)
supported that the rumen digestive system reduces the germination of soft seed and
chemically scarifies hard seed coats, reducing hardness of seed coat through ingestion and
exposure to stomach and intestine, thus increasing germination capability for hard seeds
(Russi et al., 1992). When comparing in sacco and in vivo studies, results suggest that a high
percentage of seed damage occurs in the rumen and some damage in the abomasum as a
result of pepsin digestion (Simao Neto and Jones, 1986). Carpinelli et al. (2005) suggested
Page 24
13
that exposure to ruminal-only exposure over 0, 48 and 96 hours affected perennial
pepperweed (Lepidium latifolium L.) germination with germination 17 and 15 times greater
for 48- and 96-hour incubation treatments as compared to control, respectively. Germination
for seeds exposed to total digestive tract germination was 19 times greater than control
treatment. Archer and Pyke (1991) suggested that soft-seeded plant species may lose viability
and germinability at a more rapid rate than hard-seeded plant species, however, seeds passed
at too rapid of a rate through the rumen and gastrointestinal system may fail to be effectively
scarified and suffer comparably low germinability; additional time spent in the rumen post-
scarification may also adversely affect viability and germinability. A harder seed coat
permits for higher rates of survivability post-passage through the ruminant gastrointestinal
tract, with softer seeds characterized by higher susceptibility to damage by digestive
compounds and mastication or rupture through imbibition of water (Atekson et al., 1934;
Blackshaw and Rode, 1991; Gardener et al., 1993). According to Gardener et al. (1993), the
amount of viable seed (hard and germinable) found within faeces of cattle was significantly
correlated with the hard seed content of the original seed, with the amount of non-viable
(rotten) seed dependent on the amount of soft seed in the original seedlot, and influencing
whether non-viable seed disintegrated in the digestive tract and was lost. Simao Neto et al.
(1987) similarly found soft-seeded species as the most permeable and to have a higher
likelihood of being adversely affected by the digestion process in comparison to hard seeds
which were more resistant to the digestion process when looking at the average effects of in
vitro digestion in nylon bags (in sacco); both in sacco and in vitro digestion treatments
damaged selected hard seed species approximately less than 20%. Grass seed viability
Page 25
14
reduces linearly with the increased time of exposure to rumen environment, with signal grass
seed completely inviable after 96 hours treatment (Simao Neto and Jones et al., 1987).
With consideration to seed size, Russi et al. (1992) found that small seeds are more
likely to avoid damaging effects of mastication and more quickly and easily pass through the
gastrointestinal tract than seeds of larger size. Following sheep digestion, highest recovery
percentage of intact clover seed from 150-200g seed lots was 59% from field clover
(Trifolium campestre Schreb.) characterized by 1.1 mm seed length. Lowest recovery
percentage was 23% from star clover (Trifolium stellatum L.) characterized by 1.9 mm seed
length. Large seeds are more likely to be refused during the ingestion process and to have
longer resident periods within the rumen and digestive tract (Janzen, 1984).
Grass seed damaged during passage through the digestive tract frequently only
suffered loss of the lemma and palea and many seeds were viable and germinable with
treated signal grass, Stylosanthes hamata cv. Verano and S. scabra cv. Seca seeds
germinating at higher percentages than untreated seed (Simao Neto et al., 1987). Seed
survival of grass species such as barnyard grass [Echinochloa crus-galli L.], downy brome
(Bromus tectorum L.), green foxtail [Setaria viridis L.) and foxtail barley (Hordeum jubatum
L.) exposed to rumen digestion tends to be lower than broadleaf species lambsquarter and
redroot pigweed. Germination of common lambsquarter was reduced by 17% by passage
through cattle digestive system, with downy brome, foxtail barley and barnyard grass losing
complete viability (Blackshaw and Rode, 1991; Unruh Snyder et al., 2011). Blackshaw and
Rode (1991) suggest that loss of seed viability is often not a gradual process. With some
Page 26
15
seeds, there occurs an initial lag phase in which degradation of the seed coat is likely,
followed by a rapid decline in embryo viability.
Effect of Ruminant Digestive Tract on Native Warm-Season Grasses
Native warm-season grasses are indigenous and characterized by a growth period
during the warm season of the year. The majority of utilized forages in the mid-south of the
US are naturalized species from the European [tall fescue, orchardgrass (Dactylis glomerata
L.)], African [bermudagrass, crabgrass (Digitaria spp.)] or South American [dallisgrass
(Paspalum dilatatum L.), bahiagrass (Paspalum notatum L.)] continents (Harper et al., 2007).
Warm-season grasses are often difficult or slow to establish due to characterization of chaffy,
hairy seeds that are difficult to disseminate and plant using conventional grain drills. Big
bluestem seed coat is uneven and chaffy, disallowing use of conventional drills. Seed can be
debearded, a process which removes chaff and hair from the seed coat of big bluestem and
indiangrass, and allows the seed to be dispersed through conventional equipment (USDA,
2013). Native warm-season grasses are frequently characterized by low vigor seedlings,
unable to compete successfully with weeds, and seed dormancy (Henning, 1993). Vigor is
defined as seed properties which influence rate of growth, uniform emergence and normal
development of seedlings under a wide range of environmental field conditions (AOSA,
2007).
Summer months coincide with a period of reduced animal performance in the
Southeast U.S. as a result of cool season species-dominated pastures (most commonly tall
fescue [Festuca arundinacea L.]) and associated dormant period in combination with grazing
Page 27
16
of lower quality warm season perennials, including bermudagrass (Cynodon dactylon L.).
Throughout much of the Great Plains and southeast regions of the U.S., big bluestem,
indiangrass, little bluestem (Andropogon scoparius L.) and switchgrass are recognized as
valuable forage and conservation species (Pitman, 2000). Native warm-season grasses
growth patterns make them suited well for grazing in the southeastern U.S. Cool-season
perennial grasses in forage production, characterized by limited summer yield, cause a
reduction in performance and daily gains in grazing cattle and overgrazing of tall fescue
pastures; NWSG can provide an alternative quality forage source for producers and a
combination of cool- and warm-season pastures can be successfully managed to support a
continuous supply of high-quality forage for production (Henning, 1993; Harper et al., 2007).
When NWSG are utilized and managed as a mixed-grass stand, species mature at different
dates allowing for a wider range of quality forage harvest and a varied sward structure useful
for wildlife habitat and cover. For hay production purposes, NWSG can be more effective
species choices as compared to many cool-season perennials such tall fescue, as conditions
are frequently more favorable for hay production during NWSG growth season (Harper et al.,
2007).
Utilizing NWSG in a dual purpose management system for both forage and biomass
production for ethanol is gaining interest. In the U.S., bioenergy crops are estimated to
exceed 55 million acres by the year 2030, producing 60 billion gallons of ethanol and
biodiesel (Adler and Sanderson, 2006; USDA Statistics Service, 2013; McIntosh, 2013). This
increasing demand for bioenergy crops could have an important influence on the livestock
industry and forage production (Adler and Sanderson, 2006). To address the issue of finite
Page 28
17
land resources and meeting needs of both forage and biomass production, a dual purpose
management system through the utilization of grazing animals should be explored further. A
portion of the harvest yield is used as early forage with the remaining growth purposed as
biomass harvest. Mosali et al. (2013) suggested that switchgrass can provide forage for cattle
in spring and through the warm season while still allowing for biomass harvest in the fall.
Optimum yields have been generated in systems producing a spring harvest for forage use
and a subsequent fall harvest for biomass (Sanderson et al., 1996). Big bluestem and
indiangrass are high nutritional quality forage species providing high amounts of biomass in
summer months and have additional potential for ethanol production; big bluestem has a CP
content of 16-18% is maintained from May through August (Mitchell et al., 2001). A
management system which allows producers to harvest both forage and biomass from NWSG
within the same field has the potential for increased profitability and sustainability (Mulkey
et al., 2008).
Native warm-season grasses are characterized by a C4 photosynthetic system which
produces four-carbon acids (malate and aspartate) from the carbon fixation process instead of
the three-carbon product produced by the C3 photosynthetic system. C4 grasses have
relatively high photosynthesis at low nitrogen concentrations and produce high biomass
yields (Harper et al., 2007). In contrast to C3 cool-season grasses, C4 grasses have more cell
wall constituents and less cell contents causing a high concentration and low digestibility of
fiber and restricted intake (Waramit, 2010).
Page 29
18
Native Warm-Season Grass Species
Switchgrass
Switchgrass is a perennial C4 native warm season bunch grass that is widely adapted
to varying soil types and fertility throughout the United States and Southeast U.S.
characterized by CP levels up to 20% and broad spectrum functionality, including forage use
and for conservation and restoration of rangeland. It is characterized by average height of
approximately 1 to 1.5 meters and spreads through rhizomes and seed production; culm is
round in shape and ranges from red to tan in color. Inflorescence is open panicle in shape and
seed is approximately 3 to 6 millimeters in length (USDA, 2013). The majority of growth
occurs during summer months, initiating in April and continue through late September or
October (USDA, 2013).
Switchgrass has a native range throughout the majority of the continental United
States in prairies, open oak and pine forests and is commonly found in close proximity to
rivers, streams and in brackish marshes, growing most successfully in sandy to loam soils
and can tolerate shallow or droughty soils (USDA, 2013). Due to range diversity, switchgrass
has developed two major ecotypes: upland and lowland, with the lowland ecotype producing
high levels of biomass, often as high as 8 tons per acre. Switchgrass is highly adaptable and
can grow successfully in extreme soil pH (5.0-8.0), low soil P (>10 ppm or 9 kg/A), low soil
K (>90 ppm or 82 kg/A) (USDA, 2013). Switchgrass is important for forage production as it
is one of the earliest warm season grass species to break winter dormancy; growth initiates in
early March in much of the southeastern U.S. and has the potential to produce as much as
twice the yield as tall fescue per acre. Growth produced during the vegetative stage is
Page 30
19
approximately 60-70% total digestible nutrients (TDN) and up to 17% CP (USDA, 2013).
Switchgrass provides structure necessary for nesting, brooding and winter cover and
protection for wildlife species including upland game birds, turkeys, doves, songbirds and
small mammals. As switchgrass is generally adapted to a wide range, alternative uses can
include soil erosion prevention in windbreaks, riparian areas, strip-mine spoils, sand dunes,
dikes, gullies and other critical areas. Switchgrass yields up to 8 tons of biomass per acre
with low input requirements, such as water and inorganic fertilizer amendments, and is
compatible with conventional harvesting equipment (Harper et al., 2007). These
characteristics have lead to the use of switchgrass in several bioenergy conversion processes,
including cellulosic ethanol production, biogas, and direct combustion for thermal energy
applications (USDA, 2013). It is possible for producers to utilize switchgrass for both biofuel
production and for grazing or haying; to achieve this, the highest quality early stage growth
during April and May can be utilized for grazing and the later, mature growth harvested after
frost for biomass production during its highest stage of fiber production (Burns et al., 1997).
Studies suggest that ethanol produced from switchgrass offers 540% of the energy used to
produce it, compared to 25% more energy returned by ethanol produced from corn (Vogel et
al., 2008). Ocumpaugh et al. (1996) examined differences between seed dissemination
methods of switchgrass by comparing seed fed to cattle and excreted in manure to broadcast-
seeded plots. Results suggested that while seed germinability was reduced after passage
through the ruminant digestive tract compared to broadcast seed (52-62% germinability for
digestive tract treatment compared to 85-91% germinability of broadcast seed), recruitment
of switchgrass from passed seed in manure was equal to or higher than broadcast treatment in
Page 31
20
establishment (frequency of occurrence and density) and in plant growth and plant size
(Ocumpaugh et al., 1996). Ocumpaugh et al. (1991) suggested that 26% of switchgrass seed
fed to cattle avoided the process of mastication and full digestion and was viable post-
digestive tract exposure and excretion. Switchgrass plots containing digestive tract-exposed
treatment seeds excreted with manure resulted in 62 to 100% of plots contained emerging
plants, whereas in broadcast seed plots 2 to 40% contained emerging seed. Plots were
evaluated 1 year after spring trials and frequency results were again higher for digestive tract
and manure treatments versus broadcast treatments (56 vs. 4% in May 1990, P<0.05; and 90
VS. 51% in May 1991, P<0.01). Results comparing plant recruitment suggested that manure-
seeded plots were higher, despite broadcast-seed treatment plots receiving 1.5 to 1.7 times
more pure live seed (multiplying the percentage of viable seed or percent germination by the
percent of pure seed) (USDA, 2013) and plant size score (based on visual observations of
height, culm density and biomass) was greater in manure-seeded plots (Ocumpaugh et al.,
1996). Results of these studies suggest the benefit of considering non-traditional methods of
seeded, including fecal seeding, when looking at emergence, establishment and growth.
Big Bluestem
Big bluestem is a native, perennial, warm season bunchgrass. Plants are characterized
as glaucous, with robust purplish culms and frequently growing in large clumped
communities. Averaging 1 to 2 m in height, big bluestem is often sparingly branched toward
the top with lower sheaths and blades sometimes villous, flat and elongate at 5 to 10 mm in
length. Tillers are the most prominent morphological characteristic of big bluestem (Owsley,
n.d.). Seed heads generally consist of 3 racemes, often referred to as a “turkey foot”.
Page 32
21
Flowering is photoperiod dependent and occurs from July through October; seed are chaffy,
oblong and approximately 0.25 inches in length (Maze and Scagel, 1982). Cultivated and
maintained stands of big bluestem consistently produce 150 to 200 pounds of seed per acre
(Smoliak et al., n.d.).
Big bluestem is a common species in tall grass prairies, valley bottomland, and open
woods in the central and eastern areas of the United States, Canada and Mexico; it is a major
component of understory vegetation in longleaf pine forests and plantations in the
southeastern U.S. (USDA, 2013). Overgrazing and overuse of this grass caused significant
reduction on abundance in its native range since the late 1800s (Smoliak et al., n.d.). Big
bluestem is one of 20 species belonging to the genus Andropogon. Big bluestem is
characterized by the C4 photosynthesis system and thus grows primarily and rapidly during
summer months (Gould and Shaw, 1983). Growth of big bluestem seedlings reduces
significantly at 20° C when compared to temperatures of 25° C and 30° C (Hsu et al., 1985).
Big bluestem is often unsurpassed by other prairie grasses in terms of nutritional quality in
crude protein and palatability or biomass produced. Big bluestem is an important species for
livestock and wildlife, providing both food and cover. As a forage species, big bluestem is
palatable, tolerant of poor soil quality, droughty conditions and is of high quality
nutritionally; big bluestem CP levels in early summer are frequently twice that of cool-season
species at up to 18% (USDA, 2013). Big bluestem is an important forage species as it can
withstand substantial grazing pressure, however continuous grazing to a height point lower
than 15.24 centimeters will cause competitive pressure from other species, as a result, big
bluestem is more effectively grazed in a rotational management system (Stubbendieck et al.,
Page 33
22
1985). Grazing big bluestem as a management technique can allow for significantly higher
rates of photosynthesis than either clipped or control plants (Stubbendieck et al., 1985). Big
bluestem is upright and tall in structure, providing ideal nesting and brooding cover for the
greater prairie chicken, lesser prairie chicken, ring-necked pheasant, quail, white-tailed deer,
bison and small mammals (Jacobson et al., 1985). In Iowa, ‘Kaw’ big bluestem produced 4
tons/ha of dry matter with 0 applied N and over 7 tons/ha when 150 kg N/ha was applied
(USDA, 2013); this yield suggests that big bluestem has potential as a biomass fuel producer.
Big bluestem exhibits germination issues which can be attributed to seed predation,
fungus or bacteria during wet periods and destruction by a widespread insect midge,
Contarinia wattsi which can reduce seed yield by up to 50%. Some big bluestem ecotypes
grown in areas of the southeastern U.S. produce seed with higher dormancy levels as
compared to northern ecotypes (Owsley, n.d.). A greenhouse study utilizing solid matrix seed
priming (SMP) treatments increased big bluestem germination by 18%; in field treatments,
seedling emergence from untreated big bluestem seed was similar to or higher than the
treated seed. Solid matrix seed priming is an osmoconditioning process in which seeds are
mixed with a solid matrix, provided minimum necessary water and aerated (Beckman et al.,
1993). Hsu et al. (1985) demonstrated that chilling increased big bluestem germination rate.
There are several cultivars of big bluestem; ‘Kaw’ big bluestem was the cultivar chosen for
this study as it is the most commonly used and successful cultivar adapted to the southeastern
U.S. (USDA, 2013).
Studies examining germination rates of seeds from the Poaceae family and
Andropogon genus following passage through the digestive tract of cattle, goats and sheep
Page 34
23
have been conducted by Jolaosho et al. (2006). It was found that the number and viability of
seeds recovered from excreted faeces varies depending on seed species and characteristics,
ruminant species, and animal diet (Simao Neto and Jones, 1986; Jolaosho et al., 2006). It is
suggested that the factors influencing germination of naturally occurring Andropogon
tectorum L. and Panicum maximum L. in this study also affect the spread of plants to new
areas by process of animal ingestion and subsequent excretion (Gardener et al., 1993;
Jolaosho et al., 2006). It was found that germination of seeds from sheep and goat faeces was
higher than rates of seed recovered from cattle (28, 32 and 5%, respectively) with more than
50% of germinated seeds as annuals, including grasses and sedges (Jolaosho et al., 2006).
Seed recovered from cattle faeces as compared to sheep and goat were characterized by
lower germination rates potentially due to mastication of length of resident time in rumen
(Simao Neto and Jones, 1986; Jolaosho et al., 2006).
Effect of Ruminant Digestive Tract on Agronomic Crops
The National Cotton Council of America estimates cotton production to account for
approximately 11.3 million acres in the U.S. for 2014, an 8.2% increase from 2013 (USDA,
2013). More than 2 million metric tons of cotton gin trash is available across the cotton belt
(Holt et al., 2000). By-product and waste remaining after the cotton ginning process,
including leaf and stem fragments, lint, dust, and seeds from cotton and weeds, is known as
gin trash (Mittal et al., 1979). Disposal of gin trash can be a challenging problem for gin
operations; one method is to feed gin trash to livestock in pelletized form or as raw material
(Bader et. al, 1998; Kennedy and Rankins, 2008; Norsworthy et al., 2009). Dairy and beef
Page 35
24
cows are frequently fed cotton seed hulls, cotton gin trash or other by-product related feed
stocks derived from agronomic crops exposed to Palmer amaranth as supplement (Legleiter,
2013). While cotton gin trash has relatively low digestibility, it does have much higher
protein level (12.2% crude protein), as compared to similar roughage supplements such as
corn stover (Holt et al., 2000; Rogers et al., 2002; Poore, 2008). Additional supplemental
materials produced as byproducts of cotton production, including gin motes, reginned mote
waste, and cotton textile mill waste are palatable and useful as cow feed and supplement
(Rogers et al., 2002). Poore (2008) suggests cotton gin trash as a suitable emergency feed,
especially for gestating brood cows, or for use in total mixed ration diet for cows.
Some herbicide-resistant weeds have the capability to be dispersed by means of
cattle, especially when cattle are supplemented with weed seed contaminated gin trash (Byrd
et al., 2004; Norsworthy, et al., 2009). Subsequent to cattle and livestock transportation,
resistant weed seed in their digestive tracts would likewise be transported to non-infested
areas and potentially across states in a similar fashion to tropical soda apple (Solanum viarum
Dunal) (Byrd et al., 2004; Norsworthy, et al., 2009). Dastgheib (1989) suggests that
spreading sheep manure is likely to disseminate as many as 10 million weed seeds ha-1
.
Weed Species
Palmer Amaranth
Palmer amaranth is an annual broadleaf forb with growth potential of greater than 2
meters in height; it is native to the southwest U.S. and Mexico and has become a problematic
invasive weed species in the Midwest and Southeast U.S., most significantly affecting cotton
Page 36
25
and soybean crop yield. The genus Amaranthus belongs to the family Amaranthaceae
containing 75 species, with Palmer amaranth as one of 10 dioecious (separate male and
female plants) species within the genus. Most typically, Palmer amaranth is characterized by
a central reddish-green stem up to 2 m in height with many lateral branches (Sauer, 1955).
Ovate, hairless leaves often have a darker chevron pattern on the upper side. Palmer
amaranth began to spread in range beginning in the early 20th
century, likely via methods
including human activity and agricultural expansion, with the species first reported in the
state of Virginia in 1915 and in South Carolina in 1957 (Culpepper et al., 2006; Sauer, 1957).
Palmer amaranth is an obligate outcrosser as a dioecious species (Ward et al., 2013).
Accounts of agamospermy have been documented in female Palmer amaranth plants
pollinated by common waterhemp (A. tuberculatus Moq. J.D. Sauer, formerly A. rudis L.)
(Ward et al., 2013) as well as when isolated from all pollen sources (Ribeiro et al., 2012)
which is key in determining population genetics and subsequent implications for herbicide
resistance (Ward et al., 2013). Palmer amaranth seeds are 2 to 4 mm in length, smooth and
round (Sauer, 1955), are ill-adapted to be dispersed by average wind conditions and are
commonly spread by means of gravity, irrigation and water runoff, birds and mammals,
agronomic practices such as combining, plowing and mowing, and potentially through cotton
gin trash, compost and manure inputs (Norsworthy et al., 2009). Palmer amaranth produces
up to 600,000 seeds per plant when growing without competition and 211,000 seeds when
grown in competition with soybean rows spaced 97 cm apart in South Carolina (Keeley et al.,
1987; Jha et al., 2008). In North Carolina, maximum germination for Palmer amaranth
occurred at 26° C (Wright et at., 1999). Palmer amaranth has shown high capability at
Page 37
26
evolving herbicide resistance and is currently characterized by resistance to five herbicide
mechanisms of action (MOAs), including ALS-inhibiting herbicides, dinitroanilines,
triazines, glyphosate, and HPPD inhibitors with some populations possessing resistance to
multiple MOAs (Culpepper et al., 2006; Norsworthy et al., 2008; Wise et al., 2009).
Palmer amaranth is listed in the top 10 most troublesome weeds for corn and soybean
production (ranked #7 for corn in 2009, unlisted in 1994; ranked #2 for soybean in 2010, #23
in 1995) and is currently one of the most economically damaging glyphosate-resistant weeds
in the U.S. (Beckie, 2011; Ward et al., 2013). Bensch at al. (2003) reported that infestation of
soybean fields by Palmer amaranth reduced yield by up to 79%. Cotton lint yield is reduced
5.2 to 9.3% for each increase of 1 kg of Palmer amaranth weed biomass plot-1
(Rowland et
al., 1999). Palmer amaranth is characterized by allelopathic properties or biochemicals,
which can significantly influence crop, or alternate species growth when weed residue is
incorporated into soil through tillage. Following residue incorporation of Palmer amaranth
residue, subsequent crop yield of carrots (Daucus carota L.) was reduced by 49%, onion
(Allium cepa L.) reduced by 68%, grain sorghum (Sorghum spp.) and cabbage (Brassica
oleracea L.) was also reduced (Menges, 1987; Rowland et al., 1999).
According to Norsworthy, et al., (2009), Palmer amaranth was the most prevalent
broadleaf weed found in gin trash with densities of up to 4,070 germinable seeds per metric
ton of trash sampled. Germinable Palmer amaranth seed has found in the surface layer (0 to
25 cm in depth) of gin trash compost piles both 1 and 2 years after composting was initiating
with several gin operators and producers stating Palmer amaranth as highly problematic in
fields where gin trash was applied. Based on this research, weed seed contamination, and
Page 38
27
most prevalently, Palmer amaranth contamination, is common in gin trash, including
composted gin trash (Norsworthy et al., 2009).
Texas Panicum
Texas panicum is a monocotyledonous (single cotyledon in seed) warm season annual
grass weed belonging to the Poaceae family and is also commonly referred to as buffalo
grass and Texas millet (USDA, 2013). In the 1800s, Texas panicum was a frequently utilized
species for hay production and forage in the southern U.S. and grew prevalently in prairies.
Texas panicum is now among the top 10 most common and most problematic weeds
affecting agronomic crops in the southeastern U.S. (Webster, 2004; Webster and Nichols,
2012). It is a bunch grass and develops and grows rapidly, causing issues of inadequate
control in conventional agronomic settings; Texas panicum is characterized by herbicide-
resistance and produces an average of 23,010 seeds per plant (Chandler and Satelmann,
1969).
Texas panicum is extremely competitive and can grow to more than 1 meter in height,
forming dense communities in agronomic fields (Vangessel and Johnson, n.d.). Plant
characteristics include smooth, erect culms, leaves approximately 15-25 centimeters in length
with pubescence on the leaf underside and plants are able to grow from soil depth of 8
centimeters (USDA, 2013). In competition-free field environment, Texas panicum produces
more than 500 tillers and 2000 leaves after 18 weeks. Auricles are not present and the ligule
is membranous with pubescence. The seedhead is a compressed panicle with alternately erect
rays. Texas panicum seed can remain viable for more than 5.5 years after soil submersion
(Prostko et al., 2006; Egley and Chandler, 1983). Shroeder et al. (1990) found that Texas
Page 39
28
panicum is able to thrive in a variety of soil conditions, including droughty and course-
textured soil, which suggests the species’ extreme competiveness.
Annual grasses account for some of the most problematic weeds in agronomic peanut
crops in the U.S. (Webster and Nichols, 2012). Annual grasses act to reduce yield of peanut
through excessive harvest loss; Fall panicum (Panicum dichotomiflorum L.) has been found
to be highly competitive with peanut crops, causing yield reductions of up to 25% with a
density of one weed per 4.9 m (York and Coble, 1977; Johnson and Mullinix, 2005).
Texas panicum has demonstrated tolerance to the use of preemergence herbicides in
both cotton and peanut production. Moving agricultural equipment is a common method of
unintentional spread of Texas panicum within a field or from one field to another. Post-
harvest practices and control methods such as mowing, tillage and some pesticides can act to
limit the number of seeds produced. Glyphosate and some other postemergence herbicides
are effective on Texas panicum (Prostko et al., 2006). According to Prostko, et al., in
herbicide trials, pendimenthalin applied sequentially early, 10 to 12 days after planting, acted
to control Texas panicum at a rate of less than 35% late in the season and produced reduced
corn yield and net returns. Glyphosate applied at 21 to 24 days after planting and repeated at
35 to 38 days after planting, acted to control Texas panicum at a rate of 82 to 94% late in the
season compared with 43 to 80% control by nicosulfuron and foramsulfuron. Potential
ineffectiveness of pendimenthalin on Texas panicum is problematic as, when used in
combination with atrazine, this option of control is inexpensive and historically has provided
acceptable control for early season. Additionally, chloroacetamide herbicides have shown to
be less affective on Texas panicum in the southeastern U.S. than other regions (Prostko et al.,
Page 40
29
2006). Organic materials being transferred or spread can act as an agent of spreading seed,
including mulch, straw, hay or livestock manure. Currently, there are no satisfactory
broadcast treatments available for controlling Texas panicum in permanent pasture or
grassland settings and it is unclear what impact livestock ruminants have on viability and
spreading of viable seed to unexposed areas (Prostko et al., 2006).
Ocumpaugh et al. (1991) examined seed germination and rumen influence on seeds
contained within manure after 12 to 60 hours of being fed to cattle on other grass species
within the genus Panicum and suggested that 26% of Panicum virgatum L. seed fed to cattle
were likely to avoid mastication and full digestion, allowing for viability post-digestive tract
exposure and excretion. Ocumpaugh et al. (1996) studied differences in seed dissemination
methods of Panicum virgatum L. by comparing plots seeded with seed fed to cattle and
excreted in manure to broadcast-seeded plots. Results of the study comparing seed
germinability of Panicum virgatum L. after passage through the ruminant digestive tract and
broadcast seed suggested that digestive tract exposure reduced germinability, however,
recruitment of Panicum virgatum L. from passed seed present in manure was equal to or
higher than broadcast treatment in terms of establishment (frequency of occurrence and
density) and in plant growth and size (Ocumpaugh et al., 1996). Results from these studies
suggest potentially higher emergence, establishment and growth rates of Panicum species
when seed is ingested and excreted from ruminants as compared to alternative methods of
seed dissemination. Research is needed on the impact of rumen and ruminant small intestine
digestion on germination of Texas panicum.
Page 41
30
Literature Cited
Adkins, S.W., S.M. Bellairs and D.S. Loch. 2002. Seed dormancy mechanisms in warm
season grass species. Euphytica 126: 13-20.
Adler, P.R., M.A. Sanderson, A.A. Boateng, P.J. Weimer and H.G. Jung. 2006. Biomass
yield and biofuel quality of switchgrass harvested in fall or spring. Agronomy Journal
98: 1518-1525.
Al-Mashilchi, M. S. 1993. Influence of the ruminant digestive process on the germinability
of range forage species. M. S. Thesis, Utah State Univ.
Association of Official Seed Analysts (AOSA). 2007. Rules for testing seeds. 54 (2).
Archer, S. and D. A. Pyke. 1991. Plant-animal interactions affecting plant establishment and
persistence on revegetated rangeland. J. Range Manage. 44:558-565.
Atkeson, F. W., H. W. Hulbert, and T. R. Warren. 1934. Effect of bovine digestion and
manure storage on the viability of weed seeds. J. Amer. Soc. Agron. 26:390-397.
Bader, M. J., R. K. Bramwell, R. L. Stewart, and G. M. Hill. 1998. Gin trash studies
conducted in Georgia. Pages 1698-1699 in P. Dugger and D. Richter, eds.
Proceedings of the Beltwide Cotton Conference. Memphis, TN: National Cotton
Council of America.
Ball, D.M., M. Collins, G.D. Lacefield, N.P. Martin,D.A. Mertens, K.E.Olson,D.H. Putnam,
D.J. Undersander, and M.W.Wolf. 2001. Understanding Forage Quality. American
Farm Bureau Federation Publication 1-01, Park Ridge, IL.
Barrett-Lennard, R.A., Gladstones, J.S. 1964. Dormancy and hard-seededness in Western
Australian serradella (Ornithopus compressus L.). Australian Journal of Agricultural
Research 15, 895–904.
Barrow, J.R and K.M. Havstad. 1992. Recovery and germination of gelatin-encapsulated
seeds fed to cattle. J. Arid Environ. 22395-399.
Baskin, J.M. and C.C. Baskin. 2004. A classification system for seed dormancy. Seed
Science Research 14: 1-16.
Beckie, H. J. 2011. Herbicide-resistant weed management: focus on glyphosate. Pest Manag.
Sci. 67:1037–1048.
Page 42
31
Beckman, J.J., L.E. Moser, K. Kubik and S.S. Waller. 1993. Big bluestem and switchgrass
establishment as influenced by seed priming. Agronomy Journal 85:199-202.
Bensch, C. N., M. J. Horak, and D. Peterson. 2003. Interference of redroot pigweed
(Amaranthus retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A.
rudis) in soybean. Weed Sci. 51:37–43.
Bewley, D.J. 1997. Seed germination and dormancy. Plant Cell 9: 1055-1066.
Blackshaw, R. E. and L. M. Rode. 1991. Effect of ensiling and ruminant digestion by cattle
on weed seed viability. Weed Sci. 39:104-108.
Burns J.C., K.R. Pond, D.S. Fisher, and J.M. Luginbuhl. 1997. Changes in forage quality,
ingestive mastication, and digesta kinetics resulting from switchgrass maturity. J
Anim Sci 75:1368-1379.
Byrd, J. D., C. T. Bryson, and R. G. Westbrooks. 2004. Tropical soda apple (Solatium viarum
Dunal) identification and control. Published by the Mississippi Department of
Agriculture and Commerce.
Calsamiglia, S., and M. D. Stern. 1995. A three-step in vitro procedure for estimating
intestinal digestion of protein in ruminants. J. Anim. Sci. 73:1459-1465.
Carpinelli, M.F., Schauer, C.S., Bohnert, D.W., Hardegree, S.P., Falck, S.J., Svejcar, A.J.
2005. Effect of ruminal incubation on perennial pepperweed germination. Rangeland
Ecology and Management. 58:632-636.
Chandler, J. M. and P. W. Santelmann. 1969. Growth characteristics and herbicide
susceptibility of Texas panicum. Weed Sci. 17:91–93.
Culpepper, A. S., T. L. Grey, W. K. Vencill, J. M. Kichler, T. M. Webster, S. M. Brown, A.
C. York, J. W. Davis, and W. W. Hanna. 2006. Glyphosate-resistant Palmer amaranth
(Amaranthus palmen) confirmed in Georgia. Weed Sci. 54:620-626.
Dastgheib, F. 1989. Relative importance of crop seed, manure and irrigation water as sources
of weed infestation. Weed Res. 29:113-116.
Egley, G. H. and J. M. Chandler. 1983. Longevity of weed seeds after 5.5 years in the
Stoneville 50-year buried seed study. Weed Sci. 31:264–270.
Emal, J.G., and E.C. Conard. 1973. Seed dormancy and germination in indiangrass as
affected by light, chilling, and certain chemical treatments. Agronomy Journal
65:383-386.
Page 43
32
Finch-Savage, W.E. and G. Leubner-Metzger. 2006. Seed dormancy and the control of
germination. New Phytologist 171: 501-523.
Gardener, C. J., J. C. Mclvor, and A. Jansen. 1993. Passage of legume and grass seeds
through the digestive tract of cattle and their survival in feces. J. Appl. Ecol. 30:63-
74.
Gould, F.W. and R.B. Shaw. 1983. Grass systematics. 2nd edition. Texas A&M University
Press, College Station, Texas.
Grubb, P. J. 1977. The maintenance of species-richness in plant communities: the importance
of the regeneration niche. Biol. Rev. 52:107-145.
Haidar, M.A., C. Gharib, and F.T. Sleiman. 2010. Survival of weed seeds subjected to sheep
rumen digestion. Weed Res. 50:467–471.
Harper, C.A., G.E. Bates, M.P. Hansbrough, M.J. Gudlin, J.P. Gruchy and P.D. Keyser.
2007. Native warm-season grasses: identification, establishment and management for
wildlife and forage production in the mid-south. University of Tennessee Extension.
Institute of Agriculture. Knoxville, Tennessee.
Harrington, G.T. 1916. Agriculture Value of Hard Seeds. J. Agr. Res., 6: 761 - 795.
Haynes, J.G., W.G. Pill, and T.A. Evans. 1997. Seed treatments improve the germination and
seedling emergence of switchgrass (Panicum virgatum L.). Horticultural Science 32
(7): 1222-1226.
Holt, G.A., G.L. Barker, R.V. Baker and A. Brashears. 2000. Characterization of cotton gin
byproducts produced by various machinery groups used in the ginning operation.
Transactions of the ASAE 43:1392-1400.
Henning, J.C. 1993. Big bluestem, indiangrass and switchgrass. University of Missouri.
Department of Agronomy.
Herrera, C.M. 2002. Seed dispersal by vertebrates. Plant-animalinteractions: an evolutionary
approach. (ed. by C.M. Herreraand O. Pellmyr), pp. 185-208. Blackwell Publishing,
Oxford.
Hsu, F.H., C.J. Nelson, and A.G. Matches. 1985. Temperature effects on seedling
development of perennial warm-season forage grasses. Crop Sci. 25: 249-255.
Maze, J. and R.K. Scagel. 1982. Morphogenesis of the spikelets and inflorescence of
Andropogon gerardii. Canadian Journal of Botany. 60(6): 806-817.
Page 44
33
Jacobson, ET.; C.M. Taliaferro; C.L. Dewald. 1985. New and old world bluestems. In:
Carlson, Jack R.; Mcarthur, E. Durant, chairmen. Range plant improvement in
western North America: Proceedings of a symposium at the annual meeting of the
Society for Range Management; 1985 February 14; Salt Lake City, UT. Denver, Co:
Society for Range Management: 40-50.
Janzen, D. H. 1984. Dispersal of small seeds by big herbivores: foliage is the fruit. Amer.
Natur. 123:338-353.
Jha, P., J. K. Norsworthy, W. Bridges, and M. B. Riley. 2008. Influence of glyphosate timing
and row width on Palmer amaranth (Amaranthus palmeri) and pusley (Richardia
spp.) demographics in glyphosate-resistant soybean. Weed Sci. 56:408–415.
Johnson, W.C., B. Mullinix. 2005. Texas panicum (Panicum texanum) interference in peanut
(Arachis hypogaea) and implications for treatment decisions. Peanut Science. 32:68-
72.
Jolaosho, A.O., J.A. Olanite, B.O. Oduguwa and E.A.O. Adekunle. 2005. Comparative
effects of three seasons on seed population and viability in cattle, sheep and goat
dung in southwest Nigeria. Nigerian Journal of Animal Production 32(1): 94-101.
Published by Nigerian Society for Animal Production.
Keeley, P. E., C. H. Carter, and R. J. Thullen. 1987. Influence of planting date on growth of
Palmer amaranth (Amaranthus palmeri). Weed Sci. 35:199–204.
Kennedy, J. B. and D. L. Rankins, Jr. 2008. Comparison of cotton gin trash and peanut hulls
as low-cost roughage sources for growing beef cattle. Prof. Anim. Sci. 24:40-46.
Legleiter, T., B. Johnson. 2013. Palmer Amaranth Biology, Identification, and Management.
Purdue Weed Science.
Lowry, A. 1996. Influence of Ruminant Digestive Processes on Germination of Ingested
Seeds. Master's Thesis, Oregon State University.
Massinga, R. A., R. S. Currie, M. J. Horak, and J. Boyer, Jr. 2001. Interference of Palmer
amaranth in corn. Weed Science 49: 202–208.
Maze, J. and R.K. Scagel. 1982. Morphogenesis of the spikelets and inflorescence of
Andropogon gerardii. Canadian Journal of Botany. 60(6): 806-817.
McCully, W. G. 1951. Recovery and viability of Macartney rose seeds fed to cattle. J. Range
Manage. 4:101-106.
Page 45
34
McIntosh, D. W. 2013. Forage and Biomass Dual-Purpose Harvest System Using Native
Warm-Season Grasses. Master's Thesis, University of Tennessee.
Menges, R. M. 1987. Allelopathic effects of Palmer amaranth (Amaranthus palmeri) and
other plant residues in soil. Weed Science 35: 339–347.
Mitchell, R., J. Fritz, K. Moore, L. Moser, K. Vogel, D. Redfearn, and D. Wester. 2001.
Predicting forage quality in switchgrass and big bluestem. J. Agron. 93 (1):118-124.
Mittal, D. K., R. E. Fornes, R. D. Gilbert, and P. E. Sasser. 1979. Chemical composition of
cotton dusts part I: analysis of cotton plant parts, gin trash, and weed samples. Textile
Res. J. 49:364-368.
Mosali, J., J. T. Biermacher, B. Cook, and J. Blanton. 2013. Bioenergy for cattle and cars: A
switchgrass production system that engages cattle producers. J. Agron. 105 (4):960-
966.
Mulkey, V. R., V. N. Owens, and D. K. Lee. 2008. Management of warm-season grass
mixtures for biomass production in South Dakota USA. Bioresour. Technol. 99
(3):609-617.
Newman, P.R. and L.E. Moser. 1988. Seedling root development and morphology of cool-
season and warm-season forage grasses. Crop Sci. 28: 148-151.
Norsworthy, J. K., K. L. Smith, L. E. Steckel, and C. H. Koger. 2009. Weed seed
contamination of cotton gin trash. Weed Technol. 23:574–580.
Ocumpaugh, W.R., J.W. Stnth, and S. Archer. 1991. Using cattle to distribute pasture seeds.
p. F40-F49. In: Texas A&M Univ. Agr. Res. Sta. at Beeville. Information Report 91-
1.
Ocumpaugh, W.R., S. Archer, and J.W. Stuth. 1996. Switch-grass recruitment from
broadcast seed fed to cattle. Journal of Range Management, 49:368–371.
Olivier, T. 2006. Effects of Temperature and Storage Regimes on the Germination Rates of
Three Native Warm-Season Grasses. Master's Thesis, Nicholls State University.
Owen, D. F. 1980. How plants may benefit from the animals that eat them. Oikois 35:230-
235.
Owsley, C.M. n.d. Review of big bluestem. USDA-NRCS. Jimmy Carter Plant Materials
Center. Americus, Georgia.
Page 46
35
Pieper, R. D. 1994. Ecological implications of livestock grazing. p. 177-211. In: M. Vavra,
W. A. Laycock, and R. D. Pieper (eds.), Ecological implications of livestock
herbivory in the West. Soc. Range Manage., Denver, Colo.
Pitman, W.D. 2000. Adaptation of tall-grass prairie cultivars to west Louisiana. Journal of
Range Management 53: 47-51.
Poore, M H. 2008. Alternative feeds for beef cattle during periods of low forage availability.
North Carolina State University, North Carolina Cooperative Extension Service,
Department of Animal Science.
Prostko, E. P., T. L. Grey, and J. W. Davis. 2006. Texas panicum (Panicum texanum) control
in irrigated field corn (Zea mays) with foramsulfuron, glyphosate, nicosulfuron, and
pendimethalin. Weed Technol. 20:961–964.
Ribeiro, D. N., Z. Pan, F. E. Dayan, S. O. Duke, V. K. Nadula, D. R. Shaw, and B. S.
Baldwin. 2012. Apomixis involvement in inheritance of glyphosate resistance in
Amaranthus palmeri from Mississippi. Abstracts of the Weed Science Society of
America 2012 Annual Meeting.
Rogers, G.M., M.H. Poore and J.C. Paschal. 2002. Feeding cotton products to cattle. Chapter
5 In: G.M. Rogers and M.H. Poore (eds) Alternative feeds for beef cattle. Vet. Clin.
Food Anim. 18:267-294.
Rowland, M. W., D. S. Murray, and L. M. Verhalen. 1999. Full-season Palmer amaranth
(Amaranthus palmeri) interference with cotton (Gossypium hirsutum). Weed Sci.
47:305–309.
Russi, L., R. S. Cocks, and E. H. Roberts. 1992. The fate of legume seeds eaten by sheep
from a Mediterranean grassland. J. Appl. Ecol. 29:772-778.
Sanderson, M. A., R. L. Reed, S. B. McLaughlin, S. D. Wullschleger, B. V. Conger, D. J.
Parrish, D. D. Wolf, C. Taliaferro, A. A. Hopkins, and W. R. Ocumpaugh. 1996.
Switchgrass as a sustainable bioenergy crop. Bioresour. Technol.. 56 (1):83-93.
Sauer, J. D. 1955. Revision of the dioecious amaranths. Madrono 13:5–46.
Sauer, J. D. 1957. Recent migration and evolution of the dioecious amaranths. Evolution
11:11–31.
Schroeder, J., C.C. Dowler, and J.R. Stansell. 1990. Texas panicum (Panicum texanum)
growth as affected by irrigation management and planting date. Weed Sci. 38:374-
378.
Page 47
36
Simao Neto, M. and R. M. Jones. 1986. The effect of storage in cattle dung on viability of
tropical pasture seeds. Trop. Grassi. 20:180-183.
Simao Neto, M. , R. M. Jones, and D. Ratcliff. 1987. Recovery of pasture seed ingested by
ruminants. 1. Seed of six pasture species fed to cattle, sheep, and goats. Aust. J. Exp.
Agr. 27:239-246.
Smoliak, S., R.L. Ditterline, J.D. Scheetz, L.K. Holzworth, J.R. Sims, L.E. Wiesner, D.E.
Baldridge, and G.L. Tibke. Big bluestem. Montana Interagency Plant Materials
Handbook.
Steckel, L. E., C. L. Main, A. T. Ellis, and T. C. Mueller. 2008. Palmer amaranth
(Amaranthus palmen) in Tennessee has low level glyphosate resistance. Weed
Technol. 22:119-123.
Stubbendieck, J.; J.T. Nichols; K.K. Roberts. 1985. Nebraska range and pasture grasses
(including grass-like plants). E.C. 85-170. Lincoln, NE: University of Nebraska,
Department of Agriculture, Cooperative Extension Service.
Unruh Snyder, L.J., K. Burger, J. Santini, N. Baird, and S. Lake. 2011. The effect of rumen
digestion on viability of common Indiana weed seeds. American Forage & Grassland
Council Proceedings, June 12-15th
French Lick, IN. pp. 5.
Simpson, G.M. 1990. Seed dormancy in grass. Cambridge University Press, Cambridge.
Stiles, E. W. 1992. Animals as seed dispersers. In: Fenner, M (ed.), Seeds: the ecology of
regeneration in plant communities. CAB International, Wallingford, Oxon, UK.
USDA, NRCS. 2013. Natural Resources Conservation Service. The PLANTS database.
National Plant Data Center, Baton Rouge, LA 70874-4490 USA.
http://plants.usda.gov
USDA. 2007. Plant Press: A Louisiana Native Plant Initiative publication. Volume. 1 Issue 1.
USDA, Statistics Service. 2013. Acreage Report. Accessed Jan. 2013 on usda.gov.
Vangessel, M.; Q. Johnson. Texas Panicum Control in Field Crops and Commercial
Vegetables. Newark, DE: University of Delaware, Department of Agriculture,
Cooperative Extension Service.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. Cornell Univ. Press, Ithaca, New
York.
Page 48
37
Vogel, K.P., R. Mitchell, , G. Sarath. 2008. Development of Switchgrass Into a Biomass
Energy Crop. p. 64. In: R.S. Zalesny, Jr., R. Mitchell, and J. Richardson (eds.).
Proceedings of the Short Rotation Crops International Conference, Bloomington,
Minnesota, August 19-21.
Ward, S.M., T.M. Webster, and L.E. Steckel.. Palmer amaranth: A review. Weed
Technology. 27:12-27.
Waramit, N. 2010. Native warm-season grasses: Species, nitrogen fertilization, and harvest
date effects on biomass yield and composition. Graduate Dissertation, Iowa State
University.
Webster, T. M. and H. D. Coble. 1997. Changes in the weed species composition of the
southern United States: 1974 to 1995. Weed Technol. 11:308–317.
Webster, T. M. 2004. Weed survey— southern states. Proc. South. Weed Sci. Soc. 57:404–
426.
Webster, T. M. and R. L. Nichols. 2012. Changes in the prevalence of weed species in the
major agronomic crops of the Southern United States: 1994/1995 to 2008/2009. Weed
Sci. 60:145–157.
Welch, D. 1985. Studies in the grazing of heather moorland in north-east Scotland. IV. Seed
dispersal and plant establishment in dung. J. Appl. Ecol. 22:461-472.
Wise, A. M., T. L. Grey, E. P. Prostko, W. K. Vencill, and T. M. Webster. 2009. Establishing
the geographical distribution and level of acetolactate synthase resistance of Palmer
amaranth (Amaranthus palmeri) accessions in Georgia. Weed Technol. 23:214–220.
Wright, S. R., H. D. Coble, C. D. Raper, Jr, and T. W. Rufty, Jr. 1999. Comparative
responses of soybean (Glycine max), sicklepod (Senna obtusifolia), and Palmer
amaranth (Amaranthus palmeri) to root zone and aerial temperatures. Weed Sci.
47:167–174.
Wolfe, D., and D. A. Fiske. 2009. Planting and Managing Switchgrass for Forage, Wildlife,
and Conservation. Virginia Cooperative Extension. Virginia Tech University.
York, A.C., and H.D. Coble. 1977. Fall panicum interference in peanuts. Weed Sci. 25:43-
47.
Page 49
38
IMPACT OF RUMINANT DIGESTION ON GERMINATION OF INGESTED SEEDS
OF NATIVE WARM-SEASON GRASSES IN THE SOUTHEAST UNITED STATES
Erin Kiley, Dr. Lori Unruh-Snyder, Dr. Wesley Everman and Dr. Randy Wells
---Formatted for Rangeland Ecology & Management---
Dissemination of seeds by livestock has been discussed as a means of spreading desired plant
species, across grassland and pastures through various means (Grubb, 1977). Seeds which
retain viability subsequent to ruminant digestive tract exposure can be vital to the process of
introducing new desirable forage species and enhancing the spread of established species
(Blackshaw and Rode, 1991; Lowry, 1996). Further research is needed in this area to
determine and to understand how grazing animals, including cattle, can impact the
dissemination of seeds and establishment of plant populations. The scope of this study was to
examine the effects of rumen and intestinal digestion on seed germination of both dormancy-
prone native warm-season grasses including switchgrass (Panicum virgatum L.) and big
bluestem (Andropogon gerardii L.) with the purpose of addressing cattle as a means of
transport and ruminal influence on germination of valuable forage and potential biomass
crops in the Southeast U.S.
___________________________________________________________________________
*First, second, third, and fourth authors: Graduate Research Assistant, Assistant Professor,
Assistant Professor and Professor. Crop Science Department, P.O. Box 7620, North Carolina
State University, Raleigh, NC 27695-7620.
Corresponding authors email: [email protected]
Page 50
39
Two experiments were conducted to determine how ruminant digestion influences seed
viability and germination on native warm-season grasses for potential forage, biomass and
restoration use. Experiment one was to determine the impact of rumen exposure and
digestion on seed viability and germination. Seeds were treated to in situ (live animal) rumen
digestion in a cannulated steer for 48 hours. Experiment two was to examine the effect of
small intestine exposure and digestion to determine impact on seed viability and germination
and potential points of digestion which cause or impact seed mortality. Seeds were treated to
in vitro small intestine digestion for 24 hours. Seed germination varied by plant species and
response to respective treatment. The combination of rumen phase and small intestine (SIP)
procedure exposure (abomasal-simulated digestion) resulted in highest seed mortality.
Switchgrass mean germination percentage was reduced by 86.70% and big bluestem mean
germination percentage was reduced by 79.59% with exposure to rumen phase plus SIP.
Herbivory is a key process in range and grassland ecosystems and it is imperative to consider
the role of herbivores as part of a holistic system and utilize this understanding to improve
management approaches.
Page 51
40
Introduction
Agricultural crop rotations, livestock overgrazing, lack of regular fire and subsequent
competition from cool-season grasses have caused native warm-season grasses (NWSG) to
become less prominent species in pasture settings. Reintroduction and successful
management of NWSG for grazing, hay production, biomass production and habitat
restoration is occurring across the U.S. and providing highly palatable and nutrient-dense
forage to livestock during traditionally unproductive summer months with proper
management (Henning 1993). The deterioration of rangeland through loss of native grasses
has resulted in loss of valuable forages, wildlife habitat, soil, and water resources across the
U.S. and in the southeast, specifically (Harper et al. 2007). Revegetation through artificial
reseeding of marginal pasture, grassland and range by mechanical methods is often
economically expensive and unsuccessful. Development of sustainable alternative methods
requiring less capital, labor and mechanical inputs is needed. Through integration of
livestock with land management, long term and sustainable restoration and conservation of
range, grassland and agricultural pastures may be achieved (Barrow and Havstad 1992;
McIntosh 2013). A need for high quality warm-season grass species has provided a catalyst
for renewed interest in restoration of pastures and prairies through sustainable utilization of
native component species or NWSG already adapted to the southeastern U.S. Research
continues to suggest that NWSG are viable forage for grazing and hay production for
livestock (Harper et al. 2007).
Page 52
41
Ingestion, digestion, and excretion are all processes which can influence seed
viability and germination and successful dispersal of plant communities (Norsworthy, et al.
2009). The digestive tract is an anaerobic environment containing microbes which secrete
proteolytic and cellulolytic enzymes and attach to the seed coat in the rumen and improve
fiber digestion (Gardener et al. 1993). Seeds are exposed to an acidic environment (pH 2.5)
in the abomasum and small intestine (Gardener et al. 1993). Some seeds ingested by grazing
ruminants will remain viable, or germinable, post-ingestion and digestion, however survival
and success can vary widely across species (Blackshaw and Rode 1991).
Switchgrass is a perennial C4 native warm season bunch grass that is widely adapted
to varying soil types and fertility throughout the United States and Southeast U.S. and is
receiving renewed attention for its functionality in diverse uses, including prairie
conservation and restoration, wildlife habitat, and forage. Switchgrass provides high quality
forage in summer months with crude protein (CP) levels ranging from 14 to 20% late April
though late May (USDA 2013), making it an important species to consider within a forage
production system (Burns et al. 1984). A study comparing seedling morphology of both cool-
season and warm-season grass species found Andropogon species to achieve third leaf
emergence, or boot stage approximately 3 to 5 days earlier than other warm-season grasses
and approximately 3 to 15 days earlier than cool-season grasses (Newman and Moser 1988).
As a forage species, big bluestem is palatable, tolerant of poor soil quality, droughty
conditions and is of high quality nutritionally; big bluestem CP levels in early summer are
frequently twice that of cool-season species at up to 18% and dry matter digestibility at
approximately 50% higher (USDA 2013). Ocumpaugh et al. (1996) studied differences in
Page 53
42
seed dissemination methods of grass seed, including switchgrass, by comparing seed fed to
cattle and excreted in manure to broadcast-seeded plots where the results suggested that
while seed germinability was reduced after passage through the ruminant digestive tract
compared to broadcast seed (52-62% germinability for digestive tract treatment compared to
85-91% germinability of broadcast seed), recruitment of switchgrass from passed seed in
manure was equal to or higher than broadcast treatment in establishment (frequency of
occurrence and density) and in plant growth and plant size (Ocumpaugh et al. 1996).
Ocumpaugh et al. (1991) suggested that 26% of switchgrass seed fed to cattle avoided the
process of mastication and full digestion and was viable post-digestive tract exposure and
excretion. These results suggest benefits of fecal seeding over conventional broadcast
seeding methods when looking at emergence, establishment and growth. Survival, viability
and success of seeds when exposed to ruminant digestive tracts varies with animal and plant
species and both weed and forage species dispersed by livestock should be considered by
land managers (Janzen 1983, 1984; Gardener et al. 1993).
Therefore, the objective of this study was to conduct two experiments to determine
how ruminant digestion influences seed viability and germination on native warm-season
grasses for potential forage, biomass and restoration use and on problematic weeds in
agronomic settings. In this study, the first experiment was to determine the impact of rumen
exposure and digestion on seed viability and germination. Seeds were treated to in situ (live
animal) rumen digestion in a cannulated steer for 48 hours and compared to a control
treatment (Blackshaw and Rode 1991). The second experiment was to examine the effect of
small intestine exposure and digestion to determine impact on seed viability and germination
Page 54
43
and potential points of digestion which cause or impact seed mortality. Seeds were treated to
in vitro small intestine digestion for 24 hours (Calsamiglia and Stern 1995; Unruh Snyder et
al. 2011). It was hypothesized that exposure to the ruminant digestive system would decrease
germination rates of native-warm season grass species, big bluestem and switchgrass.
Page 55
44
Materials and Methods
Research was conducted in 2013 and 2014 at North Carolina State University in
Raleigh, NC. Native warm-season grass seed for big bluestem (‘Kaw’ cultivar) and
switchgrass (‘Alamo’ cultivar) lots were donated by Roundstone Native Seed, LLC in Upton,
Kentucky and were overnight shipped to ensure limited exposure or contamination.
Table 2.1 Native warm-season grass species germination temperature and seed size
Scientific Name Common
Name
Germination
Temperature1
Seed Size
Native warm-season grass species
Andropogon gerardii Vitman Big bluestem 20-30° C 5 to 10 mm
Panicum virgatum L. Switchgrass 20° C 2 to 6 mm
Source: Association of Official Seed Analysts (AOSA) 2007. Rules for testing seeds.
Table 2.2 Thirty year averages and 2013 growing season averages for air temperature
and total precipitation in Upton, Kentucky
30 year
average air
temperature
2013 average air
temperature
30 year
average total
precipitation
2013 total
precipitation
14° C 19.8° C 118.62 cm 141.8 cm Source: University of Kentucky Agricultural Weather Center (2014)
Page 56
45
Table 2.3 Monthly growing season averages for air temperature and total precipitation
in Upton, Kentucky for 2013
Month Average Daily Temperature (C)
Monthly Sum of Daily Precipitation (cm)
January 2013 3.9 14.2
February 2013 3.9 5.0
March 2013 5 14.0
April 2013 13.9 13.0
May 2013 18.9 12.4
June 2013 23.3 12.0
July 2013 23.3 18.5
August 2013 23.9 10.2
September 2013 21.1 9.7
October 2013 14.4 9.4
November 2013 6.7 8.9
December 2013 3.9 14.5 Source: University of Kentucky Agricultural Weather Center (2014)
Page 57
46
All germination tests were conducted at the prescribed temperature and light
conditions as outlined by the AOSA, Rules of Testing Seed (2007) (Table 2.1). For the
control treatment, two replicates of 50 seeds were placed on 9-cm petri dishes prepared with
number 4 filter paper obtained from Whatman International Ltd., Maidstone, Kent ME14
2LE, U.K. moistened with 4 mL of deionized (DI) water added by pipette. Weather and
environmental data for the 2013 growing season and 30-year average climate data from
Clayton, NC was obtained from the University of Kentucky Agricultural Weather Center
(Table 2.2; Table 2.3).
Seeds were exposed to the following treatments:
1. Control (placed directly in germination plates and into germinator set at respective
species ideal temperature);
2. Rumen stage: in vivo rumen for 48 hours (Blackshaw and Rode 1991);
3. Rumen stage plus small intestinal phase: in vivo rumen for 48 hours and small intestinal
phase (SIP) for 24 hours (Calsamiglia and Stern 1995; Unruh Snyder et al. 2011);
For each treatment, there were a total of six replicates of 50 seeds for each species.
The seeds were counted, weighed and sealed into white polyester monofilament nitrogen free
10cm x 20cm Forage Bag with a 53 micron (+/- 10) porosity from Bar Diamond, Inc. in
Parma, Idaho. The sealed bags were then secured in a porous #3T PVC Sample Tube from
Bar Diamond, Inc. for 48 hours in the rumen of a cannulated steer. After 48 hours in the
rumen, the treatment groups were removed and transported to the lab in triple-sealed plastic
Page 58
47
bags to preserve temperature and pH conditions and rumen fluid sample was tested for
temperature and pH in the laboratory at North Carolina State University. Rumen fluid
temperature was 23 ° C and pH was 6.76. The bags were sorted into either the small
intestinal phase (SIP) (Calsamiglia and Stern 1995) or no SIP treatment (placed directly into
the germination phase), with three replicates per species represented in both treatments. The
no SIP bags were then rinsed with DI water, blotted dry and seeds were removed from bags
and placed in an oven for 5 minutes at 38° C. Upon removal from the drying oven, the seeds
were weighed, recorded, and seeds were placed on respective labeled germination plates and
into germinators set to species’ ideal germination conditions (AOSA 2007). For SIP
treatment reps, bags were immediately placed in a daisy jar containing 2 L of pepsin solution
(1.9 pH) for 1 hour followed by a prepared small intestine pancreatin solution (Calsamiglia
and Stern 1995). After 24 hours, the bags were removed and rinsed with DI water, blotted
dry, seeds were removed from bags and placed in the drying oven for 5 minutes at 38° C and
seeds were then weighed, recorded, placed on labeled respective germination plates and into
germinators set to species’ ideal germination conditions (AOSA 2007).
Germination occurred in a controlled environmental chamber set with ideal
alternating night/day temperatures with 12 hours of darkness and 12 hours of light conditions
for each species and settings were utilized for both control and experimental treatment.
Switchgrass treatments were placed in 20° C and big bluestem in alternating night/day
temperatures of 20/30° C. Germination plates were kept in sealed plastic bags to maintain
humidity and were grouped by treatment. All seed treatments were subjected to the same
Page 59
48
temperature and light settings as their respective control treatment, determined by species
(AOSA 2007).
Germination counts occurred daily over seven days where all treatments were read for
germinating percentages. Germination was defined and counted when the radical protruded
through the seed coat (Bewley 1997). Germinated seeds were recorded daily and removed
from the germination plate (to discourage mold growth). On day 7, a random sample of ten
hard seeds per germination plate were tested for viability according to correct protocol for
each species as outlined by the Tetrazolium Testing Committee of the Association of Official
Seed Analysts (AOSA 2000). Tetrazolium derivative used was 2,3,5-triphenyl tetrazolium
chloride (TTC) at a concentration of 0.25%. Seeds were placed in “rag-doll” saturated paper
towels overnight at room temperature to allow water imbibition. Seeds were then bisected
according to species protocol (AOSA 2000) with a razor blade near the caryopsis above the
embryo. Bisected seeds were placed in petri dishes and soaked in TTC solution of designated
time based on species (AOSA 2000) at approximately 38° C in a dark oven. Seed viability
was evaluated immediately after soaking was completed according to AOSA (2000) protocol.
Baseline tetrazolium testing for viability and germination testing on 100 seeds per seed lot
were performed by North Carolina Department of Agriculture and Consumer Services Plant
Industry Division in Raleigh, NC. Tetrazolium testing was performed on each seed lot
according to correct protocol for each species as outlined by the Tetrazolium Testing
Committee of the Association of Official Seed Analysts (AOSA 2000) to provide a baseline
estimate of seed viability.
Page 60
49
Figure 2.1 Diagram of longitudinally bisected seed and embryo exposed to tetrazolium
testing
Table 2.4 Switchgrass and big bluestem seed viability, hard seed, dormant seed and
control germination mean
Plant Species Total
Viability
Total Germination Hard
Seed
Dormant
Seed
Control
Germination
Mean %
Switchgrass 48.5% 60% 0% 1% 50.16%
Big bluestem 27% 18% 0% 0% 26.16% Source: North Carolina Department of Agriculture and Consumer Services Plant Industry Division Lab Results
Page 61
50
Statistical Analysis
The experimental design for this study was a randomized block with 3 replications for
Stage I and Stage II treatments and 2 replications for the control treatment. Data were
analyzed in SAS Statistical Software, Version 9.3, Cary, NC. ANOVA was then conducted
using PROC GLM and means were separated using the Tukey-Kramer Procedure (P≤0.05).
Statistical analyses were done utilizing number of seeds germinated, but data were presented
as mean percent germinated. Mean separation was determined through the least significant
difference (LSD). The analysis of variance table is as follows (species including big bluestem
and switchgrass):
Table 2.5 Native warm-season grass species ANOVA table
Source DF
Type III
Sum of
Squares
Mean
Square
F-
Value Pr>F
Germination Percentage Data
Model 7 3331.2 476.0 30.72 <.0001
Corrected Total 15 3455.2
Replicate 2 87.0 43.51 2.81 0.1191
Species 1 418.9 418.9 27.04 0.0008
Treatment 2 2360.4 1180.2 76.18 <.0001
Species*Treatment 2 327.0 163.5 10.55 0.0057
Error 8 123.9 15.5
Hard Seed Percentage Data
Model 7 19791.08 2827.30 484.68 <.0001
Corrected Total 15 19837.75
Replicate 2 14.67 7.34 1.26 0.3352
Species 1 15147.43 15147.43 2596.70 <.0001
Treatment 2 1259.67 629.83 107.97 <.0001
Species*Treatment 2 1027.08 513.54 88.04 <.0001
Error 8 46.67 5.83
Page 62
51
Results
A significant species by treatment interaction for germination was observed with a
LSD of 4.5. Germination of the switchgrass control treatment was 50.16% (Figure 2.2). This
was significantly greater than the rumen stage and the rumen stage plus SIP at 15.34% and
6.67% respectively (P≤0.05). Germination from the rumen stage was not significantly
different than germination observed for the rumen stage plus SIP (P≤0.05). Germination
declined by 69.42% with exposure to in situ rumen digestion (rumen stage). Germination
declined by 86.70% with exposure to in vitro small intestine digestion (rumen stage plus
SIP). Difference in germination with exposure to in sacco rumen digestion versus in sacco
rumen digestion and in vitro small intestine digestion was a reduction of 56.52%. Mean
germination percentage across all treatments for species main effect was 13.63% for big
bluestem and 24.05% for switchgrass. There was a significant species effect for mean percent
hard seed (P≤0.05). Mean percent hard seed across treatments for switchgrass was 65%.
Mean percent hard seed across treatments for big bluestem was 2.34%.
A significant species by treatment interaction for hard seed after treatment was
observed with a LSD of 5.5. Percent hard seed of switchgrass following control treatment
was 88.49% (Figure 2.3). This was not significantly different than the rumen stage or the
rumen stage plus SIP at 95.3% and 83.3% respectively (P≤0.05). Percent hard seed following
the rumen stage was not significantly different than percent hard seed observed following the
rumen stage plus SIP (P≤0.05).
Page 63
52
Germination of the big bluestem control treatment was 26.16%. This was
significantly greater than the rumen stage and the rumen stage plus SIP at 9.40% and 5.34%
respectively (P≤0.05) Germination from the rumen stage was not significantly different than
germination observed for the rumen stage plus SIP (P≤0.05). Germination declined by
64.07% with exposure to in situ rumen digestion (rumen stage). Germination declined by
79.59% with exposure to in vitro small intestine digestion (rumen stage plus SIP). Difference
in germination with exposure to in sacco rumen digestion versus in sacco rumen digestion
and in vitro small intestine digestion was a reduction of 43.19%.
Percent hard seed of big bluestem following control treatment was 26.49% (Figure
2.3). This was not significantly different than the rumen stage but was significantly different
than the rumen stage plus SIP at 14.07% and 7.33% respectively (P≤0.05). Percent hard seed
following the rumen stage was not significantly different than percent hard seed observed
following the rumen stage plus SIP (P≤0.05).
Treatment Main Effect
Mean germination percentage across NWSG species for treatment main effect was
38.16% for control treatment, 12.37% for rumen stage and 6% for rumen stage plus SIP
(Table 2.4). Control treatment mean germination percentage was significantly different than
rumen stage and rumen stage plus SIP (P≤0.05). Rumen stage mean germination percentage
was significant different from control and rumen stage plus SIP (P≤0.05). Rumen stage plus
SIP mean germination percentage was significantly different from rumen stage and control.
Mean germination for all species was reduced by 67.58% with exposure to in sacco rumen
digestion. Mean germination for all species was reduced by 84.28% with exposure to in vitro
Page 64
53
small intestine digestion. Difference in mean germination across species with exposure to in
sacco rumen digestion versus in sacco rumen digestion and in vitro small intestine digestion
was a reduction of 51.50%.
There was a significant treatment effect for mean percent hard seed (P≤0.05). Mean
percent hard seed following treatment across NWSG species was 57.49% for control
treatment, 54.70% for rumen stage and 45.33% for rumen stage plus SIP (Figure 2.3). Hard
seed percent following control treatment was not significantly different than rumen stage but
was significantly different than hard seed percent following rumen stage plus SIP (P≤0.05).
Rumen stage mean germination percentage was significant different from rumen stage plus
SIP (P≤0.05). Rumen stage plus SIP mean germination percentage was significantly different
from control (P≤0.05).
Page 65
54
Table 2.6 Percent mean germination of native warm-season grass seeds exposed to
varying stages of ruminant digestion including control treatment, rumen stage exposure
or rumen stage plus small intestinal exposure
1Percent germination obtained using three mean observations per species for rumen stage and rumen stage plus
SIP; two mean observations per species for control. 2Standard error (n=3 for rumen stage and rumen stage plus SIP; n=2 for control).
3Pecent germination within rows differ (P≤0.05) when followed by differing superscript letters; means within a
column followed by the same letter are not significantly different according to the Tukey-Kramer Procedure at
P≤0.05.
Treatment
Control Rumen Stage Rumen Stage plus
SIP
Plant
Species
%
Germination1
SE2
%
Germination SE
%
Germination SE
Switchgrass 50.16 a 3 3.00 15.34 b 0.67 b 6.67 c 0.67
Big
bluestem 26.16 a 8.00 9.40 c 0.70 c 5.34 c 0.67
Page 66
55
Figure 2.2 Percent mean germination of native warm-season grass seeds exposed to
control treatment, rumen stage exposure or rumen stage plus small intestinal exposure
1Percent germination obtained using three mean observations per species for rumen stage and rumen stage plus
SIP; two mean observations per species for control. 2Standard error (n=3 for rumen stage and rumen stage plus SIP; n=2 for control).
3Pecent germination differ (P≤0.05) when followed by differing superscript letters; means followed by the same
letter are not significantly different according to the Tukey-Kramer Procedure at P≤0.05.
0
10
20
30
40
50
60
Control Rumen Stage Rumen Stage plue Small Intestinal Phase
Ger
min
ati
on
Treatment
Big bluestem
Switchgrass
SE: 8
b
SE: 3
a
SE: 0.7
c
SE:
0.67
b
SE:
0.67
c
SE:
0.67
c
Page 67
56
Figure 2.3 Percent mean hard seed of native warm-season grass seeds exposed to
control treatment, rumen stage exposure or rumen stage plus small intestinal exposure
1Percent hard seed obtained using three mean observations per species for rumen stage and rumen stage plus
SIP; two mean observations per species for control. 2Standard error (n=3 for rumen stage and rumen stage plus SIP; n=2 for control).
3Pecent hard seed differ (P≤0.05) when followed by differing superscript letters; means followed by the same
letter are not significantly different according to the Tukey-Kramer Procedure at P≤0.05.
0
10
20
30
40
50
60
70
80
90
Control Rumen Stage Rumen Stage plue Small Intestinal Phase
Ha
rd S
eed
Treatment
Big bluestem
Switchgrass
SE: 0
b
SE:
1.42
b
SE:
1.97
b
SE:
0.94
b
SE:
0.94
b
SE: 0.54
b
Page 68
57
Discussion
Germination percentages for each treatment across species were significantly
different (P≤0.05). Seed species differ in their success in survivability when exposed to the
ruminant digestive tract (Gardener et al. 1993). Rumen digestion exposure alone reduced
germination of both species, however to a lesser extent than when exposed to the small
intestine. Increased seed mortality after exposure to the rumen may be explained by length of
exposure time (48 hours), ruminal microbe activity, high temperatures (39° C), rapid water
imbibition or anaerobic conditions damage may have occurred with rapid water uptake
(Bewley 1997). Seeds exposed to both rumen and small intestine digestion resulted in the
highest reduction in seed germination for both NWSG species. Potential factors contributing
to these results could be seed coat degradation post-rumen exposure and subsequent exposure
to acid in the abomasum and small intestine (Simao Neto et al. 1987; Gardener et al. 1993).
Additionally, degraded and damaged seed coat, caused by rumen exposure, may have been
penetrated by acidified pepsin in the small intestine treatment (1.9 pH) and, in turn caused
the seed embryo to lose viability. Past studies which utilized the small intestine simulation
procedure suggest similar results with similar grass species. Lowry (1997) reported reduction
in germination of Sherman big bluegrass (Poa secunda L.) of 69%, 86% reduction for
Bluebunch wheatgrass (Pseudoroegneria spicata L.) and a 44% reduction for Whitetop
(Lepidium draba L.) after exposure to rumen and SIP. Ruminant diet also affects retention
time, microbe activity and populations (Van Soest 1994). With a diet of grain and pasture
forage, steers used in this study may have had makeup of cellulolytic and saccharolytic
Page 69
58
microbes specially adapted to digest cellulose in the seed coat, exposing the seed embryo to
digestive tract processes and cause declines in viability (Gardener et al. 1993; Lowry 1996).
Seeds in this study were placed directly into the cow rumen and were not exposed to
mastication or complete rumination processes through manual ingestion. Seeds are impacted
by mastication and rumination (Gardener at al. 1993) and so study germination values may
have been impacted, resulting in either greater or lesser germination depending on seed
physiology of species, due to this lack of exposure. However, lack of exposure to these
digestive processes should produce small effects, as according to Simao Neto et al. (1987),
the greatest damage to digested seeds takes place in the rumen.
Blackshaw and Rode (1991) and Gardener et al. (1993) suggested the implications of
hard-seededness on the sustained viability of seeds after ruminant digestion. Seeds
characterized by hard seed coats decline in germination and viability more slowly than soft
seed coat species, including many NWSG (Archer and Pyke 1991; Blackshaw and Rode
1991). For percent hard seed, there was a significant species by treatment interaction
(P≤0.05). The percent hard seed following treatment was calculated by counting the number
of hard seeds remaining in petri dishes following respective treatment. Replicates which
experienced higher percentages of germination inherently contain fewer total seeds following
treatment as germinated seeds were removed. This caused an upward trend in mean hard seed
percent following treatment when comparing control, rumen stage and rumen stage plus SIP,
respectively. For big bluestem control treatment, there was 0% hard seed found following
treatment and 26.16% germination. This suggests that seeds that were able to germinate, did
successfully germinate leaving no viable seed remaining in the petri dish. Similarly,
Page 70
59
switchgrass control experienced the highest mean percent germination and lowest mean
percent hard seed remaining post-treatment, which suggests that the control treatment
hindered germination least of all available potential treatments. A higher mean percent hard
seed observed following switchgrass control treatment versus big bluestem control treatment
suggests that there is another factor influencing hard seeds to not germinate, potentially seed
dormancy, which is a common characteristic of switchgrass.
According to Gardener et al. (1993), dormancy of seeds is not necessarily vital for
survival in rumen and intestine digestion. Native warm-season grasses, including switchgrass
and big bluestem are frequently characterized by seed dormancy. Switchgrass and big
bluestem seed germination experienced a higher reduction percentage than species not
characterized by dormancy, including Palmer amaranth and Texas panicum. Big bluestem
germination was reduced by 79.59% and switchgrass germination was reduced by 86.70%
after exposure to ruminant digestion which suggests dormancy may not protect seed or
enhance germination when exposed to digestion. Seed, regardless of being characterized by
inherent dormancy, function as dormant within the digestive tract as it imbibes water but
germination is disallowed due to lack of oxygen. The seed coat allows for water
permeability, however is presumed in some cases to act as a barrier to acids, enzymes and
microbes present in the digestive system. When the seed is not characterized by inherent
dormancy, seeds resume germination processes when excreted with feces and exposed to
aerobic conditions (Gardener et al. 1993).
Growing climate and environmental factors affecting parent plants also may play a
role in the seed viability and germinability. Precipitation was notably greater for the the 2013
Page 71
60
growing season compared to the 30 year average in Upton, Kentucky, which may affect seed
viability, maturity and other physiological factors or parent plant factors. For the growing
season from April to October 2013, the 30 year average air temperature was 14° C, average
relative humidity maximum was 89% and relative humidity minimum was 47%. Thirty year
average maximum soil temperature when covered with grass was 17° C and minimum of 15°
C. Thirty year average total precipitation was 118.62 centimeters. Environmental conditions
for the 2013 growing season in Upton, Kentucky from April to October average air
temperature was 19.8° C, average relative humidity maximum was 86% and relative
humidity minimum was 47%. 2013 maximum soil temperature when covered with grass was
23° C and minimum of 21° C. 2013 total precipitation was 141.8 centimeters (University of
Kentucky Agricultural Weather Center, 2014).
Management Implications
Ungulates ingest and disperse seeds in sync with the ingestion of leaf foliage (Janzen
1984), however, there are relatively few attempts by land managers to utilize livestock as a
resource of dissemination for desirable plant establishment and community expansion
(Archer and Pyke 1991). By harnessing the natural processes and movements of livestock,
animals could potentially be fed seeds of desirable plant species or strategically grazed in
plant communities where desirable plant species are established and subsequently be moved
to targeted areas. If this method proves viable, advantages for management include biological
and economic. Broadcast seeding is the most widely used alternative and conventionally used
methods of seed application and is characterized by low establishment success rates.
Management techniques for seed application which ensure good seed to soil contact are
Page 72
61
frequently monetarily expensive and can be unfeasible in areas of uneven or infertile terrain.
If livestock were properly managed to disseminate seed, seeds excreted would be essentially
planted in a moist and nutrient-rich medium, potentially providing a high quality germination
substrate. Closely regulated management of grazing could allow for distribution over large
areas sustainably and at low economic costs. Conceivable issues with this method of
management occur with time span in which seeds emerge, most specifically with hard or
inherently dormant seeds and proper grazing. One potential use of this method could be the
establishment of communities with high densities of desirable species to be used as a source
of future seed for spreading plant communities.
Objective one of this study was to support validity of feeding grazing ruminant
livestock desirable forage and vegetation as a method of plant community expansion on
degraded grasslands or in areas in which conventional seeding technology is unsuitable. With
the results of this suggesting low grass seed germination rates post-digestive tract exposure, it
may be impractical to utilize grazing livestock as a management tool for propagating NWSG.
However, results show that all species studies have remaining viability post-ruminant
digestion, suggesting that further information of the capability of livestock to spread viable
seed could have applications. For switchgrass and big bluestem, rumen stage was
significantly different from the control treatment and was not significantly different rumen
stage plus SIP, suggesting the most damage to seed germination occurred in the rumen.
Exposure of grass seed to the rumen stage as compared to the control demonstrated the
greatest reduction in mean germination percentage. For every species, there were replicate
which were reduced in total final seed weight. This suggests that seed mass was either lost
Page 73
62
through digestion of cellulose in the seed coat by rumen microbes or the seed coat was
impacted by the digestive process (Gardener et al. 1993).
The NWSG species experienced diminished germination, but not elimination by
either treatment. Overall germination reduction of big bluestem when exposed to rumen stage
was 79.59% as compared to switchgrass, which was reduced by 86.70%. The chaffy
physiology of big bluestem seed may have allowed the seed to only lose the non-vital lemma
and palea and still maintain viability after digestive tract exposure (Simao Neto and Jones,
1986). Ocumpaugh et al. (1991) suggested linear reduction of grass seed germination with
respect to increased exposure to the ruminal conditions. Seed retention time within the rumen
and rate of passage may additionally impact viability through depending on degree of
imbibition with subsequent exposure to ruminal conditions with respect to differing seed coat
hardness (Blackshaw and Rode 1991; Gardener et al. 1993). According to Janzen (1984),
there may be a relationship between grass and grass seed morphology and seed survival when
subjected to digestion. Tall native warm-season bunch grasses potentially produce seeds
which are have lower survivability of digestion because seeds are evolutionarily less likely to
be ingested with plant foliage. Tall bunch grass seeds are often characterized by lignified
glumes, protective chaff, presence of awns and general poor nutritional makeup (CP) and low
palatability. Gardener et al. (1993) found that ruminants have the ability to reject seed heads
in favor of plant leaf material due to spatial distance allowed for by tall stems of bunch grass.
More research needs to be completed to determine quantities in which tall bunch grass seed
are ingested.
Page 74
63
Literature Cited
Adler, P.R., M.A. Sanderson, A.A. Boateng, P.J. Weimer and H.G. Jung. 2006. Biomass
yield and biofuel quality of switchgrass harvested in fall or spring. Agronomy Journal
98: 1518-1525.
Archer, S. and D. A. Pyke. 1991. Plant-animal interactions affecting plant establishment and
persistence on revegetated rangeland. J. Range Manage. 44:558-565.
Association of Official Seed Analysts. 2000. Tetrazolium testing handbook. Contribution No.
29.
Association of Official Seed Analysts. 2007. Rules for testing seeds. 54 (2).
Barrow, J.R and K.M. Havstad. 1992. Recovery and germination of gelatin-encapsulated
seeds fed to cattle. J. Arid Environ. 22395-399.
Beckman, J.J., L.E. Moser, K. Kubik and S.S. Waller. 1993. Big bluestem and switchgrass
establishment as influenced by seed priming. Agronomy Journal 85:199-202.
Bewley, D.J. 1997. Seed germination and dormancy. Plant Cell 9: 1055-1066.
Blackshaw, R. E. and L. M. Rode. 1991. Effect of ensiling and ruminant digestion by cattle
on weed seed viability. Weed Sci. 39:104-108.
Burns J.C., K.R. Pond, D.S. Fisher, and J.M. Luginbuhl. 1997. Changes in forage quality,
ingestive mastication, and digesta kinetics resulting from switchgrass maturity. J
Anim Sci 75:1368-1379.
Calsamiglia, S., and M. D. Stern. 1995. A three-step in vitro procedure for estimating
intestinal digestion of protein in ruminants. J. Anim. Sci. 73:1459-1465.
Gardener, C. J., J. C. Mclvor, and A. Jansen. 1993. Passage of legume and grass seeds
through the digestive tract of cattle and their survival in feces. J. Appl. Ecol. 30:63-
74.
Gould, F.W. and R.B. Shaw. 1983. Grass systematics. 2nd edition. Texas A&M University
Press, College Station, Texas.
Grubb, P. J. 1977. The maintenance of species-richness in plant communities: the importance
of the regeneration niche. Biol. Rev. 52:107-145.
Page 75
64
Harper, C.A., G.E. Bates, M.P. Hansbrough, M.J. Gudlin, J.P. Gruchy and P.D. Keyser.
2007. Native warm-season grasses: identification, establishment and management for
wildlife and forage production in the mid-south. University of Tennessee Extension.
Institute of Agriculture. Knoxville, Tennessee.
Henning, J.C. 1993. Big bluestem, indiangrass and switchgrass. University of Missouri.
Department of Agronomy.
Hsu, F.H., C.J. Nelson, and A.G. Matches. 1985. Temperature effects on seedling
development of perennial warm-season forage grasses. Crop Sci. 25: 249-255.
Janzen D. H. 1983. Dispersal of seeds by vertebrate guts. p. 232-262. In: D. J. Futuyma and
M. Slatkin (eds.), Coevolution. Sinauer Associates, Sunderland, Mass.
Janzen, D. H. 1984. Dispersal of small seeds by big herbivores: foliage is the fruit. Amer.
Natur. 123:338-353.
Jacobson, Erling T.; Taliaferro, Charles M.; Dewald, Chester L. 1985. New and old world
bluestems. In: Carlson, Jack R.; Mcarthur, E. Durant, chairmen. Range plant
improvement in western North America: Proceedings of a symposium at the annual
meeting of the Society for Range Management; 1985 February 14; Salt Lake City,
UT. Denver, Co: Society for Range Management: 40-50.
Jolaosho, A.O., J.A. Olanite, B.O. Oduguwa and E.A.O. Adekunle. 2005. Comparative
effects of three seasons on seed population and viability in cattle, sheep and goat
dung in southwest Nigeria. Nigerian Journal of Animal Production 32(1): 94-101.
Published by Nigerian Society for Animal Production.
Lowry, A. 1996. Influence of Ruminant Digestive Processes on Germination of Ingested
Seeds. Master's Thesis, Oregon State University.
Maze, J., Scagel, R. K. 1982. Morphogenesis of the spikelets and inflorescence of
Andropogon gerardii. Canadian Journal of Botany. 60(6): 806-817.
McIntosh, D.W. 2013. Forage and Biomass Dual-Purpose Harvest System Using Native
Warm-Season Grasses. Master's Thesis, University of Tennessee.
Mitchell, R., J. Fritz, K. Moore, L. Moser, K. Vogel, D. Redfearn, and D. Wester. 2001.
Predicting forage quality in switchgrass and big bluestem. J. Agron. 93 (1):118-124.
Mosali, J., J. T. Biermacher, B. Cook, and J. Blanton. 2013. Bioenergy for cattle and cars: A
switchgrass production system that engages cattle producers. J. Agron. 105 (4):960-
966.
Page 76
65
Mulkey, V. R., V. N. Owens, and D. K. Lee. 2008. Management of warm-season grass
mixtures for biomass production in South Dakota USA. Bioresour. Technol. 99
(3):609-617.
Newman, P.R. and L.E. Moser. 1988. Seedling root development and morphology of cool-
season and warm-season forage grasses. Crop Sci. 28: 148-151.
Norsworthy, J. K., K. L. Smith, L. E. Steckel, and C. H. Koger. 2009. Weed seed
contamination of cotton gin trash. Weed Technol. 23:574–580.
Ocumpaugh, W.R., J.W. Stnth, and S. Archer. 1991. Using cattle to distribute pasture seeds.
p. F40-F49. In: Texas A&M Univ. Agr. Res. Sta. at Beeville. Information Report 91-
1.
Ocumpaugh, W.R., S. Archer, and J.W. Stuth. 1996. Switch-grass recruitment from
broadcast seed fed to cattle. Journal of Range Management, 49:368–371
Owsley, C.M. Review of big bluestem. USDA-NRCS. Jimmy Carter Plant Materials Center.
Americus, Georgia.
Pitman, W.D. 2000. Adaptation of tall-grass prairie cultivars to west Louisiana. Journal of
Range Management 53: 47-51.
Sanderson, M. A., R. L. Reed, S. B. McLaughlin, S. D. Wullschleger, B. V. Conger, D. J.
Parrish, D. D. Wolf, C. Taliaferro, A. A. Hopkins, and W. R. Ocumpaugh. 1996.
Switchgrass as a sustainable bioenergy crop. Bioresour. Technol.. 56 (1):83-93
Stubbendieck, J.; Nichols, James T.; Roberts, Kelly K. 1985. Nebraska range and pasture
grasses (including grass-like plants). E.C. 85-170. Lincoln, NE: University of
Nebraska, Department of Agriculture, Cooperative Extension Service.
Unruh Snyder, L.J., K. Burger, J. Santini, N. Baird, and S. Lake. 2011. The effect of rumen
digestion on viability of common Indiana weed seeds. American Forage & Grassland
Council Proceedings, June 12-15th
French Lick, IN. pp. 5.
USDA. 2013. The PLANTS database. Natural Resources Conservation Service. National
Plant Data Center, Baton Rouge, LA 70874-4490 USA. http://plants.usda.gov
Simao Neto, M. and R. M. Jones. 1986. The effect of storage in cattle dung on viability of
tropical pasture seeds. Trop. Grassi. 20:180-183.
Page 77
66
Simao Neto, M. , R. M. Jones, and D. Ratcliff. 1987. Recovery of pasture seed ingested by
ruminants. 1. Seed of six pasture species fed to cattle, sheep, and goats. Aust. J. Exp.
Agr. 27:239-246.
Smoliak, S., R.L. Ditterline, J.D. Scheetz, L.K. Holzworth, J.R. Sims, L.E. Wiesner, D.E.
Baldridge, and G.L. Tibke. Big bluestem. Montana Interagency Plant Materials
Handbook.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. Cornell Univ. Press, Ithaca, New
York.
Vogel, K.P., Mitchell, R., Sarath, G. 2008. Development of Switchgrass Into a Biomass
Energy Crop. p. 64. In: R.S. Zalesny, Jr., R. Mitchell, and J. Richardson (eds.).
Proceedings of the Short Rotation Crops International Conference, Bloomington,
Minnesota, August 19-21.
Wolf, D., and D. Fiske. 1995. Planting and managing switchgrass for forage, wildlife, and
conservation. Virginia Cooperative Extension Publication 418-013.
Page 78
67
IMPACT OF RUMINANT DIGESTION ON GERMINATION OF INGESTED SEEDS
OF AGRONOMIC WEED SPECIES IN THE SOUTHEAST UNITED STATES
Erin Kiley, Dr. Lori Unruh-Snyder, Dr. Wesley Everman and Dr. Randy Wells
---Formatted for Weed Science---
Dissemination of seeds by livestock has been discussed as a means of spreading
problematic weed species to agronomic land through various means (Grubb 1977). Seeds
which retain viability subsequent to ruminant digestive tract exposure can be vital to the
process of introducing new weed species to previously uncontaminated land and greatly
spreading existing plant communities (Blackshaw and Rode 1991; Lowry 1996). Further
research is needed in this area to determine and to understand how grazing animals, including
cattle, can impact the dissemination of seeds and establishment of plant populations. The
scope of this study was to examine the effects of rumen and intestinal digestion on seed
germination of particularly problematic agronomic weed species in the Southeast U.S.,
including Palmer amaranth (Amaranthus palmeri L.) and Texas panicum (Panicum texanum
L.) with the purpose of addressing cattle as a means of transport and ruminal influence on
germination of negatively impactful species on crops in the Southeast U.S.
___________________________________________________________________________
*First, second, third, and fourth authors: Graduate Research Assistant, Assistant Professor,
Assistant Professor and Professor. Crop Science Department, P.O. Box 7620, North Carolina
State University, Raleigh, NC 27695-7620.
Corresponding authors email: [email protected]
Page 79
68
Two experiments were conducted to determine how ruminant digestion influences seed
viability and germination on problematic weeds in agronomic settings. Experiment one was
to determine the impact of rumen exposure and digestion on seed viability and germination.
Seeds were treated to in situ (live animal) rumen digestion in a cannulated steer for 48 hours.
Experiment two was to examine the effect of small intestine exposure and digestion to
determine impact on seed viability and germination and potential points of digestion which
cause or impact seed mortality. Seeds were treated to in vitro small intestine digestion for 24
hours. Seed germination varied by plant species and response to respective treatment. The
combination of rumen phase and small intestine procedure exposure (abomasal-simulated
digestion) or SIP resulted in highest seed mortality. Troublesome agronomic weeds Palmer
amaranth (Amaranthus palmeri L.) and Texas panicum (Panicum texanum L.) retained
viability but germination declined with digestion exposure. Palmer amaranth mean
germination percentage was reduced by 53.77% and Texas panicum mean germination
percentage was reduced by 56.83% with exposure to rumen phase plus SIP, which was the
highest rate of germination. Herbivory is a key process in range and grassland ecosystems
and it is imperative to consider the role of herbivores as part of a holistic system and utilize
this understanding to improve management approaches. Further research is needed in this
area to determine and to understand how grazing animals, including cattle, can impact the
dissemination of seeds and establishment of plant populations.
Page 80
69
Introduction
Problematic weed species can be spread through means of livestock manure
amendments and supplemental feed, such as cotton (Gossypium hirsutum L.) gin trash or
corn (Zea mays L.) and soybean (Glycine max L.) products (Norsworthy, et al. 2009),
negatively impacting yield of agronomic crops grown in subsequently contaminated areas
(Byrd et al. 2004). The National Cotton Council of America estimates cotton production to
account for approximately 11.3 million acres in the U.S. for 2014, which is an 8.2% increase
from 2013 (USDA 2013). By-product and waste remaining after the cotton ginning process,
including leaf and stem fragments, lint, dust, and seeds from cotton and weeds, is known as
gin trash (Mittal et al. 1979). Disposal of gin trash can be a challenging problem for gin
operations; one method is to feed gin trash to livestock in pelletized form or as raw material
(Bader et. al 1998; Kennedy and Rankins 2008; Norsworthy et al. 2009).
Dairy and beef cows are frequently fed cotton seed hulls, cotton gin trash or other by-
product related feed stocks derived from agronomic crops exposed to Palmer amaranth as
supplement (Legleiter 2013). Ingestion, digestion, and excretion by ruminants are all
processes which can influence seed viability and germination and successful dispersal of
plant communities (Norsworthy, et al. 2009). The digestive tract is an anaerobic environment
containing microbes which secrete proteolytic and cellulolytic enzymes and attach to the seed
coat in the rumen and improve fiber digestion (Gardener et al. 1993). Seeds are exposed to an
acidic environment (pH 2.5) in the abomasum and small intestine (Gardener et al. 1993).
Some seeds ingested by grazing ruminants will remain viable, or germinable, post-ingestion
Page 81
70
and digestion, however survival and success can vary widely across species (Blackshaw and
Rode 1991).
Due to its height potential and aggressive and rapid growth patterns, Palmer amaranth
is especially competitive to agronomic crops (Rowland et al. 1999). By 1995, Palmer
amaranth was the most troublesome weed species for cotton production North Carolina and
South Carolina, however, was not in the top 10 troublesome weeds in an additional states
(Webster and Coble 1997) and by 2009, Palmer amaranth was listed as the number one most
troublesome weed for cotton production in the southern U.S. (Webster and Nichols 2012).
Palmer amaranth reached this top position in large part due to the development of multiple
herbicide resistance to the ALS-inhibiting and glycine herbicide families (Norsworthy 2008).
Competition with Palmer amaranth can reduce yield in corn by up to 91% (Massinga et al.
2001). An issue of note is the potential for glyphosate-resistant Palmer amaranth to be
spread through gin trash disposal; if a field characterized by the presence of glyphosate-
resistant Palmer amaranth, seeds within residual cotton lint have the potential of being spread
to new fields and perpetuating resistance (Culpepper et al. 2006; Norsworthy et al. 2008;
Norsworthy et al. 2009). Texas panicum has been identified as a noxious weed species and
such, is difficult to control with many conventional weed control methods (USDA 2013).
According to Webster and Nichols (2012), Texas panicum is among the most troublesome
weeds for agronomic crop systems in the Southeast U.S., including corn and cotton
production. In corn production, grasses can be difficult to manage due to their similar
structure and herbicide tolerances; grasses account for one-third of the ranked weed species
of note in corn (Webster and Nichols 2012).
Page 82
71
In previous research, another Amaranthus species, redroot pigweed (Amaranthus
retroflexus L.) seed viability was reduced by only 52% through the process of rumen
exposure, which suggests that movement of cattle and other ruminant livestock allowed to
ingest weeds and weed seeds likely influence establishment of new populations (Norsworthy
et al. 2009; Unruh Snyder et al. 2011). Blackshaw and Rode (1991) examined twelve
commonly occurring problematic weed species in Alberta, Canada including redroot pigweed
and other hard-seeded species, in which they exposed the seeds to rumen digestion for 24
hours. The seeds were not, however, subjected to the intestinal digestion phase, which was
conducted as a component of this study (Unruh Snyder et al. 2011). Unruh Snyder et al.
(2011) found that redroot pigweed germination was enhanced by 20 to 25% through
exposure to the rumen plus intestinal phase of digestion. These results suggest that ruminants
may provide a means of weed dispersal with increased chance of germination. Survival,
viability and success of seeds when exposed to ruminant digestive tracts varies with animal
and plant species and both weed and forage species dispersed by livestock should be
considered by land managers (Janzen 1983 and 1984; Gardener et al. 1993).
Therefore, the objective of this study was to conduct two experiments to determine
how ruminant digestion influences seed viability and germination on native warm-season
grasses for potential forage, biomass and restoration use and on problematic weeds in
agronomic settings. The first experiment was to determine the impact of rumen exposure and
digestion on seed viability and germination. Seeds were treated to in situ (live animal) rumen
digestion in a cannulated steer for 48 hours and compared to a control treatment (Blackshaw
and Rode 1991). The second experiment was to examine the effect of small intestine
Page 83
72
exposure and digestion to determine impact on seed viability and germination and potential
points of digestion which cause or impact seed mortality. Seeds were treated to in vitro small
intestine digestion for 24 hours (Calsamiglia and Stern 1995; Unruh Snyder et al. 2011). It
was hypothesized that exposure to the ruminant digestive system would decrease germination
rates of native-warm season grass species, big bluestem and switchgrass.
Page 84
73
Materials and Methods
Research was conducted in 2013 and 2014 at North Carolina State University in
Raleigh, NC. Seed for Texas panicum and Palmer amaranth were collected from the Central
Crops Research Station near Clayton, NC (35ᵒ40’6.04”N, 78ᵒ30’7.00”W) in fall 2013. Soil at
Clayton was Dothan loamy sand (fineloamy, kaolinitic, thermic, Plinthic Kandiudults). The
seed lots were hand-harvested, and stored in paper bags located in a temperature controlled
laboratory at North Carolina State University at room temperature (approximately 21° C)
until processing to remove debris and residue. For Texas panicum and Palmer amaranth,
whole seed heads were clipped and placed in paper bags to be processed. Seed was cleaned
and segregated into heavy density fractions using an air screen seed cleaner. Following
cleaning, seed was stored in paper bags in a temperature controlled laboratory
(approximately 21° C) at North Carolina State University until experiment start date.
Table 3.1 Agronomic weed species germination temperature and seed size
Scientific Name Common Name Germination Temperature Seed Size
Weed species
Panicum texanum Texas panicum 20-30° C 4 to 6 mm
Amaranthus palmeri Palmer amaranth 25-35° C 2 to 4 mm
Source: Association of Official Seed Analysts (AOSA). 2007. Rules for testing seeds.
Table 3.2 Thirty year averages and 2013 growing season averages for air temperature
and total precipitation in Clayton, North Carolina
30 year average air
temperature
2013 average air
temperature
30 year average total
precipitation
2013 total
precipitation
19.7° C 19.06° C 69.40 cm 153.6cm Source: State Climate Office of North Carolina (2014). Raleigh, NC
Page 85
74
Table 3.3 Monthly growing season averages for air temperature and total precipitation
in Clayton, North Carolina for 2013
Month Average Daily Temperature (C)
Monthly Sum of Daily Precipitation (cm)
January 2013 6.9 7.8
February 2013 5.8 10.5
March 2013 7.7 4.9
April 2013 16 9.8
May 2013 19.2 15
June 2013 23.8 41.5
July 2013 25.2 36.7
August 2013 23.8 7.7
September 2013 20.6 9
October 2013 16.1 0.9
November 2013 9.2 1.8
December 2013 7.8 8 Source: State Climate Office of North Carolina (2014). Raleigh, NC
Page 86
75
All germination tests were conducted at the prescribed temperature and light
conditions as outlined by the AOSA, Rules of Testing Seed (2007) (Table 3.1). For the
control treatment, two replicates of 50 seeds were placed on 9-cm petri dishes prepared with
number 4 filter paper obtained from Whatman International Ltd., Maidstone, Kent ME14
2LE, U.K. moistened with 4 mL of deionized (DI) water added by pipette. Weather and
environmental data for the 2013 growing season and 30 year average climate data from
Clayton, NC was obtained from State Climate Office of North Carolina (Table 3.2; Table
3.3).
Seeds were exposed to the following treatments:
1. Control (placed directly in germination plates and into germinator set at respective
species ideal temperature);
2. Rumen stage: in vivo rumen for 48 hours (Blackshaw and Rode 1991);
3. Rumen stage plus small intestinal phase: in vivo rumen for 48 hours and small
intestinal phase (SIP) for 24 hours (Calsamiglia and Stern 1995; Unruh Snyder et al.
2011);
Six replicates of 50 seeds for each species were counted, weighed and sealed into
white polyester monofilament nitrogen free 10cm x 20cm Forage Bag with a 53 micron (+/-
10) porosity from Bar Diamond, Inc. in Parma, Idaho. The sealed bags were then secured in a
porous #3T PVC Sample Tube from Bar Diamond, Inc. for 48 hours in the rumen of a
cannulated steer. After 48 hours in the rumen, the treatment groups were removed and
Page 87
76
transported to the lab in triple-sealed plastic bags to preserve temperature and pH conditions
and rumen fluid sample was tested for temperature and pH in the laboratory at North
Carolina State University. Rumen fluid temperature was 23 ° C and pH was 6.98. The bags
were sorted into either the small intestinal phase (SIP) (Calsamiglia and Stern 1995) or no
SIP treatment (placed directly into the germination phase), with three replicates per species
represented in both treatments. The no SIP bags were then rinsed with DI water, blotted dry
and seeds were removed from bags and placed in an oven for 5 minutes at 38° C. Upon
removal from the drying oven, the seeds were weighed, recorded, and seeds were placed on
respective labeled germination plates and into germinators set to species’ ideal germination
conditions (AOSA 2007). For SIP treatment reps, bags were immediately placed in a daisy
jar containing 2 L of pepsin solution (1.9 pH) for 1 hour followed by a prepared small
intestine pancreatin solution (Calsamiglia and Stern 1995). After 24 hours, the bags were
removed and rinsed with DI water, blotted dry, seeds were removed from bags and placed in
the drying oven for 5 minutes at 38° C and seeds were then weighed, recorded, placed on
labeled respective germination plates and into germinators set to species’ ideal germination
conditions (AOSA 2007).
Germination occurred in a controlled environmental chamber set with ideal
alternating night/day temperatures with 12 hours of darkness and 12 hours of light conditions
for each species and settings were utilized for both control and experimental treatment. Texas
panicum treatments were placed in alternating night/day temperatures of 20/30° C and
Palmer amaranth in alternating night/day temperatures of 25/35° C. Germination plates were
kept in sealed plastic bags to maintain humidity and were grouped by treatment. All seed
Page 88
77
treatments were subjected to the same temperature and light settings as their respective
control treatment, determined by species (AOSA 2007).
Germination counts occurred daily over seven days where all treatments were read for
germinating percentages. Germination was defined and counted when the radical protruded
through the seed coat (Bewley 1997). Germinated seeds were recorded daily and removed
from the germination plate (to discourage mold growth). On day 7, seeds a random sample of
ten hard seeds per germination plate were tested for viability with tetrazolium testing
according to correct protocol for each species as outlined by the Tetrazolium Testing
Committee of the Association of Official Seed Analysts (AOSA 2000). Tetrazolium
derivative used was 2,3,5-triphenyl tetrazolium chloride (TTC) at a concentration of 0.25%.
Seeds were placed in “rag-doll” saturated paper towels overnight at room temperature to
allow water imbibition. Seeds were then bisected according to species protocol (AOSA 2000)
with a razor blade near the caryopsis above the embryo. Bisected seeds were placed in petri
dishes and soaked in TTC solution of designated time based on species (AOSA 2000) at
approximately 38° C in a dark oven. Seed viability was evaluated immediately after soaking
was completed according to AOSA (2000) protocol. Baseline tetrazolium testing for viability
and germination testing on 100 seeds per seed lot were performed by North Carolina
Department of Agriculture and Consumer Services Plant Industry Division in Raleigh, NC.
Tetrazolium testing was performed on each seed lot according to correct protocol for each
species as outlined by the Tetrazolium Testing Committee of the Association of Official Seed
Analysts (AOSA 2000) to provide a baseline estimate of seed viability.
Page 89
78
Table 3.4 Texas panicum and Palmer amaranth seed viability, hard seed, dormant seed
and control germination mean
Plant
Species
Total
Viability
Total
Germination
Hard
Seed
Dormant
Seed
Control
Germination
Mean %
Texas
panicum 61% 60% 0% 54% 24.97%
Palmer
amaranth 98% 75% 0% 0% 31.97%
Source: North Carolina Department of Agriculture and Consumer Services Plant Industry Division
Page 90
79
Statistical Analysis
The experimental design for this study was a randomized block with 3 replications for
Stage I and Stage II treatments and 2 replications for control treatment. Data were analyzed
in SAS Statistical Software, Version 9.3, Cary, NC. ANOVA was then conducted using
PROC GLM and means were separated using Tukey-Kramer Procedure (P≤0.05). Statistical
analyses were done utilizing number of seeds germinated, but data were presented as mean
percent germinated. Mean separation was determined through the least significant difference
(LSD). The analysis of variance table is as follows (species including Texas panicum and
Palmer amaranth):
Table 3.5 Agronomic weed species ANOVA table
Source DF
Type III
Sum of
Squares
Mean
Square
F-
Value Pr>F
Germination Percentage Data
Model 7 810.23 115.75 3.99 0.0354
Corrected Total 15 1042.22
Replicate 2 43.98 21.99 0.76 0.4994
Species 1 32.56 32.56 1.12 0.3203
Treatment 2 540.48 270.24 9.32 0.0081
Species*Treatment 2 57.91 28.95 1.00 0.4101
Error 8 231.99 29.00
Hard Seed Percentage Data
Model 7 15647.05 2235.29 129.90 <.0001
Corrected Total 15 15784.72
Replicate 2 98.35 49.17 2.86 0.1158
Species 1 14006.86 14006.86 813.98 <.0001
Treatment 2 236.01 118.01 6.86 0.0184
Species*Treatment 2 204.68 102.34 5.95 0.0261
Error 8 137.66 17.21
Page 91
80
Results
A significant treatment main effect was observed with no difference in germination
between Palmer amaranth and Texas panicum. This indicates that these species are similarly
affected by the rumen and small intestine. Germination of the control treatment was 29.5%
which was significantly greater than rumen only stage (17.53%) and the rumen stage plus
SIP (12.78%) (Figure 3.1). Although numerically greater, the germination observed
following the rumen only stage was not significantly different from the rumen stage plus SIP
(Table 3.6). Overall germination was reduced by 48.77% with exposure to in sacco rumen
exposure while Germination was reduced by 53.77% when exposed to in vitro small intestine
phase. A 10% difference in germination was observed with exposure to in sacco rumen
digestion compared to in sacco rumen digestion and in vitro small intestine digestion.
A significant species by treatment interaction for hard seed after treatment was
observed with a LSD of 5.97. Percent hard seed of Palmer amaranth following control
treatment was 63.70% (Figure 3.2). This was not significantly different than the rumen stage
or rumen stage plus SIP at 72.03% and 75.86% respectively. Percent hard seed following the
rumen stage was not significantly different than percent hard seed observed following the
rumen stage plus SIP. Percent hard seed of Texas panicum following control treatment was
12.70% (Figure 3.2). This was not significantly different than the rumen stage or rumen stage
plus SIP at 2.67% and 15.44% respectively. Percent hard seed following the rumen stage was
significantly different than percent hard seed observed following the rumen stage plus SIP
(P≤0.05).
Page 92
81
There was a significant treatment effect for mean percent hard seed (P≤0.05). Mean
percent hard seed following treatment across agronomic weed species was 38.20% for
control treatment, 37.35% for rumen stage and 45.65% for rumen stage plus SIP (Figure 3.2).
Hard seed percent following control treatment was not significantly different than rumen
stage but was significantly different than hard seed percent following rumen stage plus SIP
(P≤0.05). There was a significant species effect for mean percent hard seed (P≤0.05). Mean
percent hard seed across treatments was for Palmer amaranth was 70.53%. Mean percent
hard seed across treatments for Texas panicum was 10.30%.
Table 3.6 Percent mean germination of agronomic weed seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure
1Percent germination obtained using three mean observations per species for Rumen Stage and Rumen Stage
plus SIP; two mean observations per species for control. 2Standard error (n=3 for Rumen Stage and Rumen Stage plus SIP; n=2 for control).
3Pecent germination within rows differ (P≤0.05) when followed by differing superscript letters; means within a
row followed by the same letter are not significantly different according to the Tukey-Kramer Procedure at
P≤0.05.
Treatment
Control Rumen Stage Rumen Stage plus
SIP
Plant Species %
Germination1
SE2
%
Germination SE
%
Germination SE
Palmer
amaranth 32.56
a 5.00 16.48
b 2.22 14.78
b 1.39
Texas panicum 25.56a 4.14 18.67
b 4.06 10.78
b 3.00
Page 93
82
Figure 3.1 Percent mean germination of agronomic weed seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure
1Percent germination obtained using three mean observations per species for Rumen Stage and Rumen Stage
plus SIP; two mean observations per species for control. 2Standard error (n=3 for Rumen Stage and Rumen Stage plus SIP; n=2 for control).
3Pecent germination differ (P≤0.05) when followed by differing superscript letters; means followed by the same
letter are not significantly different according to the Tukey-Kramer Procedure at P≤0.05.
0
5
10
15
20
25
30
35
40
Control Rumen Stage Rumen Stage plue
Small Intestinal Phase
Ger
min
ati
on
Treatment
Palmer amaranth
Texas panicum
SE: 5
a
SE: 4.14
a
SE: 2.2
b
SE:
4.06
b SE:
14.78
b
SE: 3
b
Page 94
83
Figure 3.2 Percent mean hard seed of agronomic weed seeds exposed to control
treatment, rumen stage exposure or rumen stage plus small intestinal exposure
1Percent hard seed obtained using three mean observations per species for rumen stage and rumen stage plus
SIP; two mean observations per species for control. 2Standard error (n=3 for rumen stage and rumen stage plus SIP; n=2 for control).
3Pecent hard seed differ (P≤0.05) when followed by differing superscript letters; means followed by the same
letter are not significantly different according to the Tukey-Kramer Procedure at P≤0.05.
0
10
20
30
40
50
60
70
80
90
Control Rumen Stage Rumen Stage plue Small
Intestinal Phase
Ha
rd S
eed
Treatment
Palmer amaranth
Texas panicum
SE:
4.96
b
SE:
1.42
b
SE:
2.45
b
SE:
0.66
b
SE:
2.37
b
SE:
1.52
b
Page 95
84
Discussion
Seed species differ in their success in survivability when exposed to the ruminant
digestive tract (Gardener et al. 1993). In this study, the treatment combining rumen and small
intestine digestion resulted in the highest reduction in seed germination for both species
investigated. Potential factors contributing to these results could be seed coat degradation
post-rumen exposure and subsequent exposure to acid in the abomasum and small intestine
(Simao Neto and Jones 1986; Gardener et al. 1993). Rumen exposure reduced germination of
all species. Increased seed mortality after exposure to the rumen may be explained by length
of exposure time (48 hours), ruminal microbe activity, high temperatures (39° C), rapid water
imbibition or anaerobic conditions damage may have occurred with rapid water uptake
(Bewley 1997). Small intestine exposure increased mortality of all seed species. This may be
due to the degraded and damaged seed coat, caused by rumen exposure, being penetrated by
acidified pepsin in the small intestine treatment (1.9 pH) and causing the seed embryo to lose
viability. Past studies which utilized the small intestine simulation procedure suggest similar
results with like species. Lowry (1997) reported reduction in germination of Sherman big
bluegrass (Poa secunda L.) of 69%, 86% reduction for Bluebunch wheatgrass
(Pseudoroegneria spicata L.) and a 44% reduction for Whitetop (Lepidium draba L.) after
exposure to rumen and SIP. Ruminant diet also affects retention time, microbe activity and
populations (Van Soest 1994). With a diet of grain and pasture forage, steers used in this
study may have had makeup of cellulolytic and saccharolytic microbes specially adapted to
digest cellulose in the seed coat, exposing the seed embryo to digestive tract processes and
Page 96
85
cause declines in viability (Gardener et al. 1993; Lowry 1996). Seeds in this study were
placed directly into the cow rumen and were not exposed to mastication or complete
rumination processes through manual ingestion. Seeds are impacted by mastication and
rumination (Gardener at al. 1993) and so study germination values may have been impacted,
resulting in either greater or lesser germination depending on seed physiology of species,
based on this lack of exposure. However, lack of exposure to these digestive processes
should produce small effects, as according to Simao Neto and Jones (1986), the greatest
damage to digested seeds takes place in the rumen.
Growing climate and environmental factors affecting parent plants also may play a
role in the seed viability and germinability. Precipitation was notably greater for the 2013
growing season compared to the 30 year average in Clayton, North Carolina which may have
affected seed viability, maturity and other physiological factors or parent plant factors. For
the growing season from March to October, the 30 year average air temperature was 19.7° C.
Thirty year average total precipitation was 69.4 centimeters. Environmental conditions for
the 2013 growing season in Clayton, North Carolina from March to October average air
temperature was 19.06° C and total precipitation was 153.6 centimeters (State Climate Office
of North Carolina, 2014).
Management Implications
One objective of this study was to support validity of grazing ruminant livestock as a
source of agronomic weed plant community dispersal and spread into uncontaminated areas.
Most notably, the dispersal of noxious and invasive agronomic weed seed by livestock is a
valid concern for land managers as demonstrated by the results of this study (Archer and
Page 97
86
Pyke 1991; Lowry 1996). More research is necessary to develop applicable management
strategies to mitigate, diminish and contain the successful spread of weeds species to
agronomic land. For every species, there were replicates which were reduced in total final
seed weight. This suggests that seed mass was either lost through digestion of cellulose in the
seed coat by rumen microbes or the seed coat was impacted by the digestive process.
According to results from Janzen (1983), most small seeds are present in the digestive
tract of cows for approximately 2-3 days and are capable of moving up to 14 km per day-1
,
suggesting seeds have the ability to be spread over large areas of land. This information, in
combination with the results of this and other studies, showing maintained viability of weed
seed post-ruminant digestive tract exposure, suggests that seed of certain species has the
capability to be spread to new and favorable growth agronomic crop sites by livestock. The
results of this study suggest that weed species have the most potential success of being
disseminated by cattle when compared to favorable forage species (Kiley et al. 2014). With
respect to this, species well adapted to dry, infertile substrate conditions which produce a
high volume of small seeds and are readily eaten by ruminants in combination with leaf
matter, such as Texas panicum, or Palmer amaranth fed directly to cows in cotton gin trash,
have a high likelihood of colonizing new areas of pasture or agronomic fields amended with
contaminated manure by cattle (Gardener et al. 1993).
Blackshaw and Rode (1991) and Gardener et al. (1993) suggested the implications of
hard-seededness on the sustained viability of seeds after ruminant digestion. The hard seed
coat of Palmer amaranth may be a factor in higher survivability and germination post-small
intestine treatment. Seeds characterized by hard seed coats, like Palmer amaranth and redroot
Page 98
87
pig weed, decline in germination and viability more slowly than soft seed coat species,
including many NWSG (Archer and Pyke 1991; Blackshaw and Rode 1991). For percent
hard seed, there was a significant species by treatment interaction (P≤0.05). The percent hard
seed following treatment was calculated by counting the number of hard seeds remaining in
petri dishes following respective treatment. Replicates which experienced higher percentages
of germination inherently contain fewer total seeds following treatment as germinated seeds
were removed. This caused an upward trend in mean hard seed percent following treatment
when comparing control, rumen stage and rumen stage plus SIP, respectively, which was an
inverse relationship with germination percentages. This suggests that seeds that were able to
germinate, did germinate leaving no viable seed remaining in the petri dish. Palmer amaranth
control experienced the highest mean percent germination and lowest mean percent hard seed
remaining post-treatment, which suggests that the control treatment hindered germination
least of all available potential treatments. A higher mean percent hard seed observed
following Palmer amaranth control treatment versus Texas panicum control treatment
suggests that there is another factor influencing hard seeds to not germinate, potentially seed
dormancy or failure to experience environmental germination triggers.
Agronomic crop weed management in the southeastern region of the U.S. has been
fundamentally changed by recent introductions and dominance of glyphosate-resistant and
other herbicide-resistant cultivars of corn, cotton, and soybean, in combination with improper
rotation of herbicide mechanisms. In cotton and soybean production, multiple herbicide
applications within a single season are typically used due to the lack of robust crop growth
and canopy closure (Nandula et al. 2009; Webster and Nichols 2012). Cotton has limited
Page 99
88
tolerance to a select number of herbicide mechanisms, as compared to other agronomic
crops. Weed communities and flora has changed over the past several decades, likely due to
broad use of glyphosate as a management tool. Seven of the top 15 most troublesome weeds
species of cotton and soybean crops, both crops which rely heavily on the greatest use of
glyphosate, are new species of concern in 2009 as compared to 1995 (Webster and Nichols
2012). These changes demonstrate the capacity of weed species to adapt show to recurrent
selection from herbicides. Weeds which increased in importance most between 1994 and
2008, according to Webster and Nichols (2012), include Palmer amaranth, increasing in
relative rank to 7 from 25 for corn and increasing to relative rank to 1 from 10 for cotton.
Texas millet ranks as second most troublesome weed for corn and number 8 for cotton.
Palmer amaranth did not appear in the 1974 or 1994 surveys, however, pigweeds did indicate
the presence of Amaranthus species. The increase in prevalence of Palmer amaranth as a
troublesome agronomic weed species is likely due to herbicide resistance (Webster 2004;
Webster and Nichols 2012). Future research should seek methods to hinder the rapid spread
of herbicide-tolerant and evolution of herbicide-resistant weed species which, according to
this study, have the capability of being widely spread by ruminant livestock.
Managers may have some control over the process of extending seed retention time
within the ruminant tract by increasing fiber in the diet to slow rate of passage, and thus
decrease seed viability and contamination (Lowry 1996). In light of this study’s results, a
technique to be considered by managers is to contain or dry-lot cattle after being exposed to
or grazed in an area contaminated with Palmer amaranth or Texas panicum or fed
contaminated feedstocks to reduce spread of these problematic species. Another option for
Page 100
89
land and livestock managers to consider is to graze weed-contaminated areas prior to seed
maturation to prevent cattle consuming the weed seeds and mitigate plant community spread.
Page 101
90
Literature Cited
Archer, S. and D. A. Pyke. 1991. Plant-animal interactions affecting plant establishment and
persistence on revegetated rangeland. J. Range Manage. 44:558-565.
Association of Official Seed Analysts. 2000. Tetrazolium testing handbook. Contribution No.
29.
Association of Official Seed Analysts. 2007. Rules for testing seeds. 54 (2).
Bader, M. J., R. K. Bramwell, R. L. Stewart, and G. M. Hill. 1998. Gin trash studies
conducted in Georgia. Pages 1698-1699 in P. Dugger and D. Richter, eds.
Proceedings of the Beltwide Cotton Conference. Memphis, TN: National Cotton
Council of America.
Beckie, H. J. 2011. Herbicide-resistant weed management: focus on glyphosate. Pest Manag.
Sci. 67:1037–1048.
Bensch, C. N., M. J. Horak, and D. Peterson. 2003. Interference of redroot pigweed
(Amaranthus retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A.
rudis) in soybean. Weed Sci. 51:37–43.
Bewley, D.J. 1997. Seed germination and dormancy. Plant Cell 9: 1055-1066.
Blackshaw, R. E. and L. M. Rode. 1991. Effect of ensiling and rumen digestion by cattle on
weed seed viability. Weed Sci. 39:104-108.
Byrd, J. D., C. T. Bryson, and R. G. Westbrooks. 2004. Tropical soda apple (Solatium viarum
Dunal) identification and control. Published by the Mississippi Department of
Agriculture and Commerce.
Calsamiglia, S., and M. D. Stern. 1995. A three-step in vitro procedure for estimating
intestinal digestion of protein in ruminants. J. Anim. Sci. 73:1459-1465.
Chandler, J. M. and P. W. Santelmann. 1969. Growth characteristics and herbicide
susceptibility of Texas panicum. Weed Sci. 17:91–93.
Culpepper, A. S., T. L. Grey, W. K. Vencill, J. M. Kichler, T. M. Webster, S. M. Brown, A.
C. York, J. W. Davis, and W. W. Hanna. 2006. Glyphosate-resistant Palmer amaranth
(Amaranthus palmen) confirmed in Georgia. Weed Sci. 54:620-626.
Dastgheib, F. 1989. Relative importance of crop seed, manure and irrigation water as sources
of weed infestation. Weed Res. 29:113-116.
Page 102
91
Egley, G. H. and J. M. Chandler. 1983. Longevity of weed seeds after 5.5 years in the
Stoneville 50-year buried seed study. Weed Sci. 31:264–270.
Gardener, C. J., J. C. Mclvor, and A. Jansen. 1993. Passage of legume and grass seeds
through the digestive tract of cattle and their survival in feces. J. Appl. Ecol. 30:63-
74.
Grubb, P. J. 1977. The maintenance of species-richness in plant communities: the importance
of the regeneration niche. Biol. Rev. 52:107-145.
Holt, G.A., G.L. Barker, R.V. Baker and A. Brashears. 2000. Characterization of cotton gin
byproducts produced by various machinery groups used in the ginning operation.
Transactions of the ASAE 43:1392-1400.
Janzen D. H. 1983. Dispersal of seeds by vertebrate guts. p. 232-262. In: D. J. Futuyma and
M. Slatkin (eds.), Coevolution. Sinauer Associates, Sunderland, Mass.
Janzen, D. H. 1984. Dispersal of small seeds by big herbivores: foliage is the fruit. Amer.
Natur. 123:338-353.
Jha, P., J. K. Norsworthy, W. Bridges, and M. B. Riley. 2008a. Influence of glyphosate
timing and row width on Palmer amaranth (Amaranthus palmeri) and pusley
(Richardia spp.) demographics in glyphosate-resistant soybean. Weed Sci. 56:408–
415.
Johnson, W.C., Mullinix, Jr., B. 2005. Texas panicum (Panicum texanum) interference in
peanut (Arachis hypogaea) and implications for treatment decisions. Peanut Science.
32:68-72.
Keeley, P. E., C. H. Carter, and R. J. Thullen. 1987. Influence of planting date on growth of
Palmer amaranth (Amaranthus palmeri). Weed Sci. 35:199–204.
Kennedy, J. B. and D. L. Rankins, Jr. 2008. Comparison of cotton gin trash and peanut hulls
as low-cost roughage sources for growing beef cattle. Prof. Anim. Sci. 24:40-46.
Legleiter, T., Johnson, B. 2013. Palmer Amaranth Biology, Identification, and Management.
Purdue Weed Science.
Lowry, A. 1996. Influence of Ruminant Digestive Processes on Germination of Ingested
Seeds. Master's Thesis, Oregon State University.
Massinga, R. A., R. S. Currie, M. J. Horak, and J. Boyer, Jr. 2001. Interference of Palmer
amaranth in corn. Weed Science 49: 202–208.
Page 103
92
Menges, R. M. 1987. Allelopathic effects of Palmer amaranth (Amaranthus palmeri) and
other plant residues in soil. Weed Science 35: 339–347.
Mittal, D. K., R. E. Fornes, R. D. Gilbert, and P. E. Sasser. 1979. Chemical composition of
cotton dusts part I: analysis of cotton plant parts, gin trash, and weed samples. Textile
Res. J. 49:364-368.
Nandula, V.K., D.H. Poston, K.N. Reddy, K. Whiting. 2009. Response of Soybean to
Halosulfuron Herbicide. International Journal of Agronomy. 2009(754510):1-7.
Norsworthy, J. K., G. M. Griffith, R. C. Scott, K. L. Smith, and L. R. Oliver. 2008.
Confirmation and control of glyphosate-resistant Palmer amaranth (Amaranthus
palmeri) in Arkansas. Weed Technol. 22:108-113.
Norsworthy, J. K., K. L. Smith, L. E. Steckel, and C. H. Koger. 2009. Weed seed
contamination of cotton gin trash. Weed Technol. 23:574–580.
Ocumpaugh, W.R., S. Archer, and J.W. Stuth. 1996. Switch-grass recruitment from
broadcast seed fed to cattle. Journal of Range Management, 49:368–371.
Poore, M H. 2008. Alternative feeds for beef cattle during periods of low forage availability.
North Carolina State University, North Carolina Cooperative Extension Service,
Department of Animal Science.
Prostko, E. P., T. L. Grey, and J. W. Davis. 2006. Texas panicum (Panicum texanum) control
in irrigated field corn (Zea mays) with foramsulfuron, glyphosate, nicosulfuron, and
pendimethalin. Weed Technol. 20:961–964.
Ribeiro, D. N., Z. Pan, F. E. Dayan, S. O. Duke, V. K. Nadula, D. R. Shaw, and B. S.
Baldwin. 2012. Apomixis involvement in inheritance of glyphosate resistance in
Amaranthus palmeri from Mississippi. Abstracts of the Weed Science Society of
America 2012 Annual Meeting.
Rogers, G.M., M.H. Poore and J.C. Paschal. 2002. Feeding cotton products to cattle. Chapter
5 In: G.M. Rogers and M.H. Poore (eds) Alternative feeds for beef cattle. Vet. Clin.
Food Anim. 18:267-294.
Rowland, M. W., D. S. Murray, and L. M. Verhalen. 1999. Full-season Palmer amaranth
(Amaranthus palmeri) interference with cotton (Gossypium hirsutum). Weed Sci.
47:305–309.
Sauer, J. D. 1955. Revision of the dioecious amaranths. Madrono 13:5–46.
Page 104
93
Sauer, J. D. 1957. Recent migration and evolution of the dioecious amaranths. Evolution
11:11–31.
Schroeder, J., C.C. Dowler, and J.R. Stansell. 1990. Texas panicum (Panicum texanum)
growth as affected by irrigation management and planting date. Weed Sci. 38:374-
378.
Unruh Snyder, L.J., K. Burger, J. Santini, N. Baird, and S. Lake. 2011. The effect of rumen
digestion on viability of common Indiana weed seeds. American Forage & Grassland
Council Proceedings, June 12-15th
French Lick, IN. pp. 5.
USDA. 2013. The PLANTS database. Natural Resources Conservation Service. National
Plant Data Center, Baton Rouge, LA 70874-4490 USA. http://plants.usda.gov
Simao Neto, M. and R. M. Jones. 1986. The effect of storage in cattle dung on viability of
tropical pasture seeds. Trop. Grassi. 20:180-183.
Steckel, L. E., C. L. Main, A. T. Ellis, and T. C. Mueller. 2008. Palmer amaranth
(Amaranthus palmen) in Tennessee has low level glyphosate resistance. Weed
Technol. 22:119-123.
Trucco, F., D. Zheng, A. J. Woodyard, J. R. Walter, T. C. Tatum, A. L. Rayburn, and P. J.
Tranel. 2007. Nonhybrid progeny from crosses of dioecious Amaranths: implications
for gene-flow research. Weed Sci. 55:119–122.
Vangessel, M.; Q. Johnson. Texas Panicum Control in Field Crops and Commercial
Vegetables. Newark, DE: University of Delaware, Department of Agriculture,
Cooperative Extension Service.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. Cornell Univ. Press, Ithaca, New
York.
Ward, S.M., Webster, T.M., Steckel, L.E. 2013. Palmer amaranth: A review. Weed
Technology. 27:12-27.
Webster, T. M. and H. D. Coble. 1997. Changes in the weed species composition of the
southern United States: 1974 to 1995. Weed Technol. 11:308–317.
Webster, T. M. 2004. Weed survey— southern states. Proc. South. Weed Sci. Soc. 57:404–
426.
Page 105
94
Webster, T. M. and R. L. Nichols. 2012. Changes in the prevalence of weed species in the
major agronomic crops of the Southern United States: 1994/1995 to 2008/2009. Weed
Sci. 60:145–157.
Wise, A. M., T. L. Grey, E. P. Prostko, W. K. Vencill, and T. M. Webster. 2009. Establishing
the geographical distribution and level of acetolactate synthase resistance of Palmer
amaranth (Amaranthus palmeri) accessions in Georgia. Weed Technol. 23:214–220.
Wright, S. R., H. D. Coble, C. D. Raper, Jr, and T. W. Rufty, Jr. 1999. Comparative
responses of soybean (Glycine max), sicklepod (Senna obtusifolia), and Palmer
amaranth (Amaranthus palmeri) to root zone and aerial temperatures. Weed Sci.
47:167–174.
York, A.C., and H.D. Coble. 1977. Fall panicum interference in peanuts. Weed Sci. 25:43-
47.
Page 107
96
Appendix 1- Trial 1 Germination Raw Data
Species
Initial
seed wt. g
Final
seed
count
Final total
wt.
Difference
in seed
wt. g
Viability
Baseline Temperature
Germ
Day
1
Germ
Day
2
Germ
Day
3
Germ
Day
4
Germ
Day
5
Germ
Day
6
Germ
Day
7
Total #
Germimation
Percent
Germination
Mean %
Germ
Rumen
Stage
Big
bluestem 0.1011 50 0.1608 0.0597 27% 20-30 0 0 0 1 2 1 0 4 8.00%
9.40% Big
bluestem 0.1090 49 0.1821 0.0731 27% 20-30 0 1 1 0 1 0 2 5 10.20%
Big
bluestem 0.0923 50 0.1201 0.0278 27% 20-30 0 0 2 1 2 0 0 5 10.00%
Rumen
Stage
plus
SIP
Big
bluestem 0.1011 50 0.1059 0.0048 27% 20-30 0 0 0 0 1 2 0 3 6.00%
5.33% Big
bluestem 0.1185 50 0.1092
-
0.0093 27% 20-30 0 0 0 0 0 1 1 2 4.00%
Big
bluestem 0.1008 50 0.0978
-
0.0030 27% 20-30 0 0 0 0 1 0 2 3 6.00%
Control Big
bluestem 0.0852 50 n/a n/a 27% 20-30 0 3 3 2 0 1 0 9 18.00%
26.00%
Control Big
bluestem 0.0852 50 n/a n/a 27% 20-30 0 4 7 3 1 1 1 17 34.00%
Rumen
Stage
Switchgrass 0.0929 50 0.1123 0.0194 48.5% 20.0 0 0 0 0 2 2 3 7 14.00%
15.33% Switchgrass 0.0966 50 0.0876 -
0.0090 48.5% 20.0 0 0 0 0 3 3 2 8 16.00%
Switchgrass 0.0986 50 0.1234 0.0248 48.5% 20.0 0 0 0 0 1 3 4 8 16.00%
Rumen
Stage
plus
SIP
Switchgrass 0.0960 50 0.0580 -
0.0380 48.5% 20.0 0 0 0 0 0 2 1 3 6.00%
6.67%
Switchgrass 0.0976 50 0.0612 -
0.0364 48.5% 20.0 0 0 0 0 1 2 1 4 8.00%
Page 108
97
Switchgrass 0.0982 50 0.0572 -
0.0410 48.5% 20.0 0 0 0 0 0 2 1 3 6.00%
Control Switchgrass 0.0966 50 n/a n/a 48.5% 20.0 0 0 8 4 5 3 2 22 44.00%
50.00%
Control Switchgrass
0.0966 50 n/a n/a 48.5% 20.0 0 0 7 6 6 9 0 28 56.00%
Rumen
Stage
Texas
panicum 0.2445 50 0.2712 0.0267 61% 20-30 0 3 3 2 1 0 0 9 18.00%
18.67% Texas
panicum 0.2508 50 0.2643 0.0135 61% 20-30 0 5 5 3 0 0 0 13 26.00%
Texas
panicum 0.2098 50 0.2431 0.0333 61% 20-30 0 4 1 1 0 0 0 6 12.00%
Rumen
Stage
plus
SIP
Texas
panicum 0.2440 50 0.2241
-
0.0199 61% 20-30 0 2 1 1 0 0 1 5 10.00%
10.78% Texas
panicum 0.2539 49 0.2187
-
0.0352 61% 20-30 0 3 1 3 1 0 0 8 16.33%
Texas
panicum 0.2262 50 0.2262 0.0000 61% 20-30 0 1 0 0 1 0 1 3 6.00%
Control Texas
panicum
0.2384 50 n/a n/a 61% 20-30 0 2 3 4 3 3 0 15 30.00% 26.00%
Control Texas
panicum
0.2384 50 n/a n/a 61% 20-30 0 0 5 2 3 1 0 11 22.00%
Rumen
Stage
Palmer
amaranth 0.0181 47 0.0214 0.0033 98% 25-35 0 0 2 2 1 2 2 9 19.15%
16.38% Palmer
amaranth 0.0165 50 0.0411 0.0246 98% 25-35 0 0 2 1 2 0 1 6 12.00%
Palmer
amaranth 0.0163 50 0.0312 0.0149 98% 25-35 0 1 4 3 0 0 1 9 18.00%
Page 109
98
Rumen
Stage
plus
SIP
Palmer
amaranth 0.0155 50 0.0179 0.0024 98% 25-35 0 1 1 4 0 1 1 8 16.00%
14.78% Palmer
amaranth 0.0173 50 0.0121
-
0.0052 98% 25-35 0 2 1 3 0 0 0 6 12.00%
Palmer
amaranth 0.0161 49 0.0204 0.0043 98% 25-35 0 1 2 5 0 0 0 8 16.33%
Control Palmer
amaranth
0.0149 50 n/a n/a 98% 25-35 3 5 5 0 2 2 2 19 38.00% 33.00%
Control Palmer
amaranth
0.0149 50 n/a n/a 98% 25-35 3 1 6 1 0 0 3 14 28.00%
Page 110
99
Appendix 2- Trial 2 Germination Raw Data
Species
Initial
seed wt.
g
Final
seed
count
Final
total wt.
Difference
in seed
wt. g
Viability
Baseline Temperature
Germ
Day 1
Germ
Day 2
Germ
Day 3
Germ
Day 4
Germ
Day 5
Germ
Day 6
Germ
Day 7
Total #
Germimation
Percent
Germination
Mean %
Germ
Rumen
Stage
Little
bluestem 0.0786 50 0.1211 0.0425 28% 20-30 0 0 0 1 0 0 0 1 2.00%
8.05% Little
bluestem 0.0825 49 0.1324 0.0499 28% 20-30 0 1 1 1 0 1 0 4 8.16%
Little
bluestem 0.0971 50 0.0960
-
0.0011 28% 20-30 0 1 2 1 1 1 1 7 14.00%
Rumen
Stage
plus
SIP
Little
bluestem 0.0835 49 0.1112 0.0277 28% 20-30 0 0 0 0 0 1 0 1 2.04%
2.68% Little
bluestem 0.0147 50 0.1115 0.0969 28% 20-30 0 0 0 0 1 1 0 2 4.00%
Little
bluestem 0.0914 50 0.1178 0.0264 28% 20-30 0 0 0 0 0 0 1 1 2.00%
Control Little
bluestem
0.0069 50 n/a n/a 28% 20-30 0 0 0 0 1 1 0 2 4.00% 5.00%
Control Little
bluestem
0.0797 50 n/a n/a 28% 20-30 0 0 0 0 1 1 1 3 6.00%
Rumen
Stage
Indiangrass 0.1065 50 0.1194 0.0129 29% 20-30 0 0 1 1 1 0 0 3 6.00%
4.67% Indiangrass 0.0917 50 0.1054 0.0137 29% 20-30 0 0 0 1 1 0 1 3 6.00%
Indiangrass 0.0897 50 0.1191 0.0294 29% 20-30 0 0 0 1 0 0 0 1 2.00%
Rumen
Stage
plus
SIP
Indiangrass 0.0953 50 0.1256 0.0303 29% 20-30 0 0 0 0 0 0 0 0 0.00%
0.00%
Indiangrass 0.1120 50 0.1279 0.0159 29% 20-30 0 0 0 0 0 0 0 0 0.00%
Page 111
100
Indiangrass 0.0913 50 0.1141 0.0228 29% 20-30 0 0 0 0 0 0 0 0 0.00%
Control Indiangrass
0.1030 50 n/a n/a 29% 20-30 0 0 0 1 1 1 0 3 6.00% 5.00%
Control Indiangrass
0.0884 50 n/a n/a 29% 20-30 0 0 0 1 1 0 0 2 4.00%
Rumen
Stage
Eastern
gama 4.0911 50 7.6532 3.5621 55% 20.0 0 0 0 0 1 1 1 3 6.00%
2.67% Eastern
gama 4.9752 50 8.7324 3.7572 55% 20.0 0 0 0 0 0 1 0 1 2.00%
Eastern
gama 3.6984 50 7.5836 3.8852 55% 20.0 0 0 0 0 0 0 0 0 0.00%
Rumen
Stage
plus
SIP
Eastern
gama 4.2334 50 8.4819 4.2485 55% 20.0 0 0 0 0 0 0 0 0 0.00%
0.00% Eastern
gama 5.0633 50 9.1641 4.1008 55% 20.0 0 0 0 0 0 0 0 0 0.00%
Eastern
gama 4.8480 50 9.0173 4.1693 55% 20.0 0 0 0 0 0 0 0 0 0.00%
Control Eastern
gama
5.6516 50 n/a n/a 55% 20.0 0 0 0 0 0 0 0 0 0.00% 2.00%
Control Eastern
gama
5.0123 50 n/a n/a 55% 20.0 0 0 0 0 0 1 1 2 4.00%