V o l u m e 8 0 | I s s u e 5 | O c t 0 b e r 2 0 1 5 225
Maturation changes auxin profile during the process of adventitious rooting in PrunusG. Osterc and F. ŠtamparUniversity of Ljubljana, Biotechnical Faculty, Institute for Fruit Growing, Viticulture and Vegetable Growing, Ljubljana, Slovenia
IntroductionMany years have passed since the maturation process in
woody plants was described in all its complexity by differ-ent authors for the first time (Fortanier and Jonkers, 1976; Olesen, 1978). Different authors have studied different as-pects of the maturation process already before (Seeliger, 1924) but the interrelations among these segments has been demonstrated in the eighties (Osterc, 2009). The today’s understanding of this process is a result of differ-ent experiments carried out later, also explaining a very
German Society for Horticultural Science
wide involvement of the maturation process into the hor-ticulture production (Dodd and Power, 1988; Spethmann, 1997). A very important aspect in understanding the mat-uration process is the possibility of reverting the mature plants into juvenile ones. In vitro propagation methods of-fer an excellent possibility of creating physiologically juve-nile plant material which stays genetically identical to the original stock plant. Therefore, in vitro methods in woody plants should be mostly used as the procedure of creating juvenile stock plant material and not so much as a method of directly propagating the plant material for the market. Based on many previous studies it is well known that ad-ventitious root formation (ARF) is significantly easier in juvenile stock plant material than in a mature one (Morgan and Williams, 1976; Osterc et al., 2009; Osterc and Štam-par, 2011; Plietzsch and Heiliger, 1997; Spethmann, 1997). However, we still do not know why these juvenile plants react so extremely different compared to genetically iden-tical mature plants.
The knowledge of biochemical patterns of ARF has been strongly improved since the discovery of IAA as the main substance contributing to ARF (Went, 1934). Today we know that the presence of IAA is important immediately after removal of plant parts from the stock plant during the induction and initiation phase of ARF (De Klerk et al., 1995). The IAA-profile is very similar among species dur-ing this time. The induction phase starts with the sever-ance point and terminates with a significant IAA-peak and with lower peroxidase activity, mostly 24 h after severance from the stock plant (Nag et al., 2001; Osterc and Štam-par, 2011; Štefančič et al., 2007). After that, the initiation phase takes place and lasts until the decrease of IAA levels with the simultaneous increase of the peroxidase activity,
SummaryThe changes in indole-3-acetic acid (IAA) and in-
dole-3-acetic acid aspartate (IAA-Asp) levels in leafy cuttings of Prunus subhirtella ‘Autumnalis’ were studied in 2011. Leafy cuttings were severed from mature, semi-mature and juvenile stock plants on 20th of June when a half of them were treated with indole-3-butyric acid (IBA) powder (0.5% IBA + 10% Captan on talcum basis) while the rest remained untreated. Auxins (IAA, IAA-Asp) were measured at the time of severance and after 4 hours, 1 and 2 days. Cuttings from juvenile stock plants rooted signifi-cantly better (80%) than cuttings from semi-mature (50%) and mature stock plants (36.7%) and they de-veloped the longest roots (5.19 cm). The IAA-profile in juvenile cuttings reached its maximum (peak) 4 h after severance, whereby the maximum value 4.27 µg g-1 fresh weight (FW) was reached 4 h after sever-ance when the cuttings were pre-treated with IBA. The IAA-profile in semi-mature cuttings reached a peak of 1.48 µg g-1 FW on day 1 after severance but only when the cuttings had been pre-treated with IBA. The IAA-profile in mature cuttings did not reach any significant peak, however the values tended to increase on first day after severance in cuttings without IBA application. The IAA-Asp accumulation ranged between 3.20 and 9.11 µg g-1 FW, which was significantly the strongest in mature cuttings one day after severance without IBA treatment.
Significance of this studyWhat is already known on this subject?• It was very well known for a long period of time that
juvenile material develops roots much easier com-pared with mature material.
What are the new findings?• The IAA-profile in cutting bases differs in cuttings
regarding the physiological character of the origin of these cuttings.
What is the expected impact on horticulture?• Worse propagation is often not the result of an un-
suitable method but the consequence of unsuitable (mature) propagation material.
eJHS Magazine Vol 80-6.indd 225 12-10-15 15:19
226 E u r o p e a n J o u r n a l o f H o r t i c u l t u r a l S c i e n c e
Osterc and Štampar | Maturation changes auxin profile during the process of adventitious rooting in Prunus
reaching a peak, mostly 72 h after severance (Nag et al., 2001). A very quick degradation in this phase of ARF was also found for indole-3-butyric acid (IBA) in different plum cultivars (Sándor et al., 2010). The forms of IAA present in plants are different. IAA is rarely present in plants in free form but mostly in conjugate form, in dicots mainly in the form of amide conjugates (Woodward and Bartel, 2005). The type of IAA form in the plant at the time of severance might be very important regarding ARF. Certain IAA con-jugates (IAA-Alanine, IAA-Leucine) have auxin activity, whereas others (IAA-Asp) have only low auxin activity (Bartel et al., 2001; Ludwig-Müller, 2011). It is known from our previous results that IAA-profiles in cutting bases are similar during ARF among different physiological levels of stock plants. IAA-Asp accumulation is especially strong in mature cuttings which tend to lower rooting success (Os-terc and Štampar, 2011).
In this paper, we compare the duration of the induction and initiation phases among stock plants of different phys-iological levels based on the IAA-profile in leafy cuttings. Based on the IAA-peak we compare the transition from the induction to the initiation phase in cuttings derived from stock plants of different physiological levels. We also com-pare the aspartate profile during this time because the conjugate IAA-Asp is known as the IAA-inactivation com-pound. Additionally, we check the effectiveness of exoge-nously supplied auxin for the ARF induction and initiation phase, also in the light of cutting material of different phys-iological levels.
tumnalis’ leafy cuttings were harvested from different stock plants, distinguished by their physiological status. Three treatments were established: (1) mature plants, more than 40-year-old cherry plants; (2) semi-mature plants, 5-year-old plants propagated by leafy cuttings ob-tained from mature cherry plants; and (3) juvenile plants, 5-year-old in vitro derived plants (the rejuvenated above-mentioned mature cherry stock plants). All plants were located in the experimental field of the Biotechnical Fac-ulty in Ljubljana (Slovenia) to ensure equal environmental conditions. Mature stock plants were rarely pruned (only the dry branches), whereas semi-mature and juvenile stock plants were pruned every spring. Therefore, mature plants were up to 10 m high, semi-mature and juvenile plants were bushy, up to 1.5 m high. Leafy cuttings were harvested on 20th of June 2011. The propagation material was always ad-justed to 12 cm long leafy cuttings with apical meristems. On average, the cuttings had three to four fully developed leaves.
Cutting propagationThe cuttings were inserted in the substrate mixture of
peat and sand (1:1). Prior to insertion, the substrate mix-ture was fertilized with a slow-release fertilizer (2.0 g L-1 3-4 M Osmocote® Exact 16+11+11+3 Mg+Te; Scotts Interna-tional, Heerlen, The Netherlands), and the pH value was ad-justed to 4.0 with dolomitic lime. The first half of cuttings did not receive any hormone treatment before they were set into the propagation bed. Another half of the cuttings were treated with IBA powder (0.5% IBA on talcum basis with 10% Captan) on their bases before inserting into the
substrate. All experimental treatments (cuttings from ma-ture, semi-mature and juvenile stock plant material with and without IBA application) were replicated 3 times with 17 cuttings per plot (12 cuttings per plot for auxin analyses, 5 cuttings for rooting evaluation).
The experiment was set in an unheated plastic house equipped with a fogging system (Plantfog-Befeuchtungs- anlagen Nebelsysteme, Fishamend, Austria). The air tem-peratures in the plastic house reached up to 50°C dur-ing daytime and ranged between 18 and 20°C during the night. The substrate temperature (rooting zone) did not undergo such oscillations (between 20 and 24°C), mainly due to a quality fogging system. Fogging was regulated au-tomatically (the intervals were set manually dependent on weather conditions) to obtain a 90–95% relative humidity. Fogging intervals lasted approx. 30 s, with a 60 s pause and were switched off during the night (7.00 pm – 7.00 am).
The evaluation of propagation success was always per-formed four weeks after severance. The number of success-fully rooted leafy cuttings and the main roots were counted and the root bush length was measured.
Extraction and analyses of IAA and IAA-AspLeafy cuttings (three cuttings per treatment for each
sampling date) for auxin analysis were collected on the day of severance and immediately transferred to labora-tory facilities. All the leaves were cut off and discarded and only the basal parts (root emergence zone, lower 3 cm of the cuttings) were used for further analysis. The samples were washed and stored at -20°C until the analyses were performed.
Auxins were extracted following the methods of Gonçalves et al. (2008), Kovač et al. (2003) and Štefančič et al. (2007). Samples were ground to a fine powder us-ing a mortar and pestle with liquid nitrogen. Each sample was divided into two portions, 0.15 g each, and extracted separately with 1 mL BHT-MeOH solution [0.5 g of BHT (2,6-di-tert-butyl-4-methylphenol) per 1 L solution] and 4 mL 5 mM K-phosphate buffer, adjusted to pH 6.5. After 1 h at 4°C, the extract was filtered and 3 mL 5 mM K-phos-phate buffer (pH value 6.5) was added to each sample. For auxins purification, Strata C18-E columns (pore size 55 μm, retention capacity 500 mg, tube size 6 mL; Phenomenex, Torrance, CA, USA) were used. The complete sample ex-tracts were first run through the column pre-conditioned with K-phosphate buffer (pH value 6.5) and then washed with 4 mL of 5 mM K-phosphate buffer (pH value 6.5). The eluate was acidified to pH value 2.5 with 0.5 M H3PO4, then transferred to a second column pre-conditioned with the K-phosphate buffer (pH value 2.5). The column was rinsed with 2 mL twice-distilled water and eluted with 2 mL of 80% (v/v) methanol.
The concentrated eluate (1 mL) was separated by TSP (Thermo Separation Products) HPLC using a Chromsep (Varian, Palo Alto, CA, USA) column [SS 250 x 4.6 mm, Hy-persil 5 ODS (Octa Decyl Silica)] and analysed by a fluores-cence (Spectrasystem FL2000; SpectraPhysics, San Jose, CA, USA) and a UV-VIS detector (K-2500; Knauer, Berlin, Germany). The mobile phase consisted of solvent A (aceto-nitrile/glacial acetic acid/twice-distilled water, 10/2/88, v/v) and solvent B (100% acetonitrile). The gradient was isocratic from 84% of solvent A and 16% of solvent B for the first 20 min to 95% of solvent A and 5% of solvent B for the rest of the 30 min method, and the flow rate was 1 mL min-1. The excitation of the fluorescence detector
eJHS Magazine Vol 80-6.indd 226 12-10-15 15:19
V o l u m e 8 0 | I s s u e 5 | O c t 0 b e r 2 0 1 5 227
Osterc and Štampar | Maturation changes auxin profile during the process of adventitious rooting in Prunus
was set at 292 nm and emission at 360 nm. The UV-VIS detector was adjusted to measure absorption at 280 nm (Gonçalves et al., 2008). IAA and IAA-Asp were quantified by fluorimetry, comparing peak areas with those of corre-sponding standards. Losses were evaluated by standards that had passed through the extraction and purification processes. Auxins were identified using a mass spectro-meter (Thermo Scientific, LCQ Deca XP MAX) with an atmospheric-pressure chemical ionization (APCI) opera-ting in the positive ion mode. The analyses were carried out using full scan data-dependent MSn scanning from m/z 50 to 1,000. The injection volume was 10 μL and the flow rate was maintained at 1 mL min-1. The capillary temperature was 275°C, the sheath gas and auxiliary gas were 35 and 10 units respectively; and the source voltage was 16 kV. Spec-tral data were elaborated using the Excalibur software (Thermo Scientific). The identification of compounds was confirmed by comparing retention times and their spectra as well as by adding the standard solution to the sample and by fragmentation. The calculated auxin contents (IAA and IAA-Asp) of each sample (n=3) were the means of two replicate measurements.
Statistical analysesStatistical analyses were carried out with the Stat-
graphics Plus (version 4.0) programme, using analyses of variance (ANOVA). Statistically significant differences between treatments (physiological status and hormone treatment) were tested with the Duncan test at a 0.95 con-fidence level.
Results
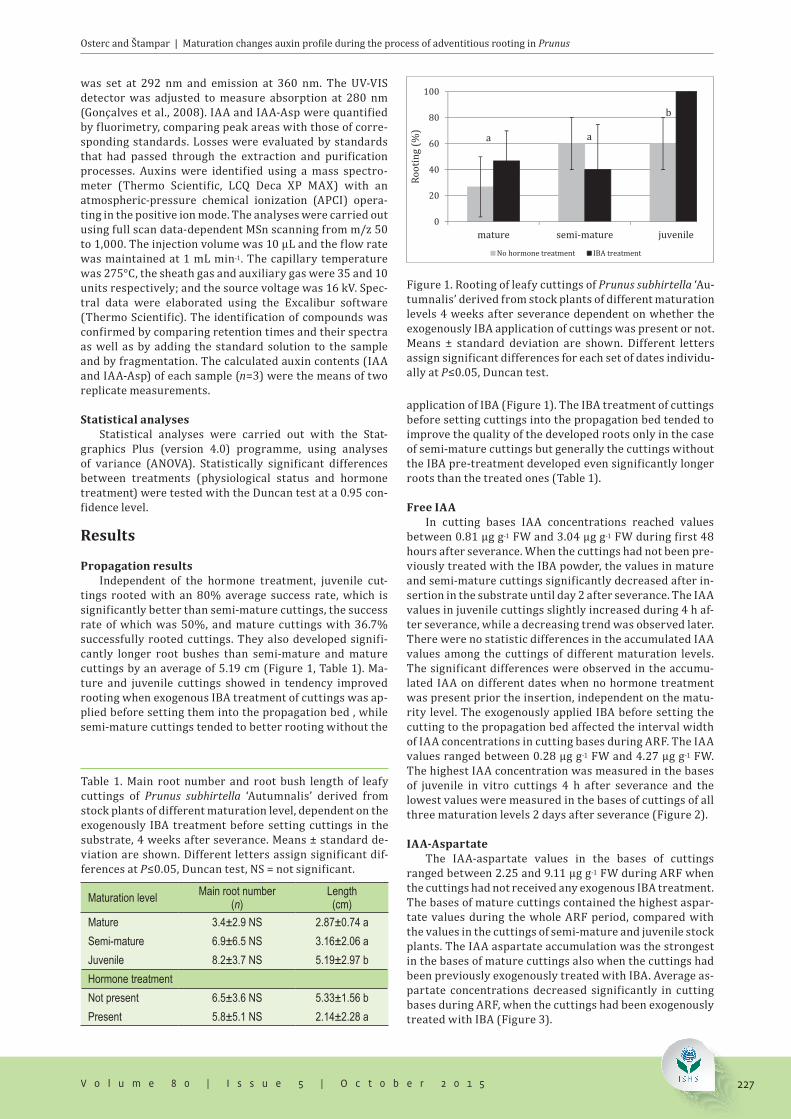
Propagation resultsIndependent of the hormone treatment, juvenile cut-
tings rooted with an 80% average success rate, which is significantly better than semi-mature cuttings, the success rate of which was 50%, and mature cuttings with 36.7% successfully rooted cuttings. They also developed signifi-cantly longer root bushes than semi-mature and mature cuttings by an average of 5.19 cm (Figure 1, Table 1). Ma-ture and juvenile cuttings showed in tendency improved rooting when exogenous IBA treatment of cuttings was ap-plied before setting them into the propagation bed , while semi-mature cuttings tended to better rooting without the
application of IBA (Figure 1). The IBA treatment of cuttings before setting cuttings into the propagation bed tended to improve the quality of the developed roots only in the case of semi-mature cuttings but generally the cuttings without the IBA pre-treatment developed even significantly longer roots than the treated ones (Table 1).
between 0.81 µg g-1 FW and 3.04 µg g-1 FW during first 48 hours after severance. When the cuttings had not been pre-viously treated with the IBA powder, the values in mature and semi-mature cuttings significantly decreased after in-sertion in the substrate until day 2 after severance. The IAA values in juvenile cuttings slightly increased during 4 h af-ter severance, while a decreasing trend was observed later. There were no statistic differences in the accumulated IAA values among the cuttings of different maturation levels. The significant differences were observed in the accumu-lated IAA on different dates when no hormone treatment was present prior the insertion, independent on the matu-rity level. The exogenously applied IBA before setting the cutting to the propagation bed affected the interval width of IAA concentrations in cutting bases during ARF. The IAA values ranged between 0.28 µg g-1 FW and 4.27 µg g-1 FW. The highest IAA concentration was measured in the bases of juvenile in vitro cuttings 4 h after severance and the lowest values were measured in the bases of cuttings of all three maturation levels 2 days after severance (Figure 2).
IAA-AspartateThe IAA-aspartate values in the bases of cuttings
ranged between 2.25 and 9.11 µg g-1 FW during ARF when the cuttings had not received any exogenous IBA treatment. The bases of mature cuttings contained the highest aspar-tate values during the whole ARF period, compared with the values in the cuttings of semi-mature and juvenile stock plants. The IAA aspartate accumulation was the strongest in the bases of mature cuttings also when the cuttings had been previously exogenously treated with IBA. Average as-partate concentrations decreased significantly in cutting bases during ARF, when the cuttings had been exogenously treated with IBA (Figure 3).
Table 1. Main root number and root bush length of leafy cuttings of Prunus subhirtella ‘Autumnalis’ derived from stock plants of different maturation level, dependent on the exogenously IBA treatment before setting cuttings in the substrate, 4 weeks after severance. Means ± standard de-viation are shown. Different letters assign significant dif-ferences at P≤0.05, Duncan test, NS = not significant.
Figure 1. Rooting of leafy cuttings of Prunus subhirtella ‘Au-tumnalis’ derived from stock plants of different maturation levels 4 weeks after severance dependent on whether the exogenously IBA application of cuttings was present or not. Means ± standard deviation are shown. Different letters assign significant differences for each set of dates individu-ally at P≤0.05, Duncan test.
9
Figure 1. Rooting of leafy cuttings of Prunus subhirtella ‘Autumnalis’ derived from stock plants of different maturation levels 4 weeks after severance dependent on whether the exogenously IBA application of cuttings was present or not. Means ± standard deviation are shown. Different letters assign significant differences for each set of dates individually at P<=0.05, Duncan test.
0
20
40
60
80
100
mature semi-mature juvenile
Rooti
ng (%
)
No hormone treatment IBA treatment
b
aa
eJHS Magazine Vol 80-6.indd 227 12-10-15 15:19
228 E u r o p e a n J o u r n a l o f H o r t i c u l t u r a l S c i e n c e
Osterc and Štampar | Maturation changes auxin profile during the process of adventitious rooting in Prunus
Figure 2. IAA concentrations in leafy cutting bases of Prunus subhirtella ‘Autumnalis’ at the time of severance and immediately after severance dependent on maturation level of cutting material and IBA treatment of cut-tings before their inserting in the substrate. Means ± standard deviation are shown. Dif-ferent letters assign significant differences for each set of dates individually at P≤0.05, Duncan test.
Figure 3. IAA-Asp concentrations in leafy cut-ting bases of Prunus subhirtella ‘Autumnalis’ at the time of severance and immediately af-ter severance dependent on maturation level of cutting material and IBA treatment of cut-tings before their inserting in the substrate. Means ± standard deviation are shown. Dif-ferent letters assign significant differences for each set of dates individually at P≤0.05, Duncan test.
11
Figure 3. IAA-Asp concentrations in leafy cutting bases of Prunussubhirtella ‘Autumnalis’ at the time of severance and immediately after severance dependent on maturation level of cutting material and IBA treatment of cuttings before their inserting in the substrate. Means ± standard deviation are shown. Different letters assign significant differences for each set of dates individually at P<=0.05, Duncan test.
0
2
4
6
8
10
12
14
Conc
entra
tion (
µg g-
1 FW
)
No IBA treatment beforeinserting in substrate
b
aa
02468
101214
severance date 4 h 1 day 2 daysadult semi-adult juvenile
IBA treatment beforeinserting in substrate
aababbbaaCo
ncen
tratio
n (µg
g-1 FW
) 10
Figure 2. IAA concentrations in leafy cutting bases of Prunussubhirtella ‘Autumnalis’ at the time of severance and immediately after severance dependent on maturation level of cut-ting material and IBA treatment of cuttings before their inserting in the substrate. Means ± standard deviation are shown. Different letters assign significant differences for each set of dates individually at P<=0.05, Duncan test.
Conc
entra
tion(
µg g-1
FW)
No IBA treatment beforeinserting in substrate
c bc ab a1
2
3
4
5
6
0
severance date 4 h 1 day 2 days
mature semi-mature juvenile
IBA treatment beforeinserting in substrate
aab
abb
b
bc
c
0
1
2
3
4
5
6
Conc
entra
tion (
µg g-1 F
W)
eJHS Magazine Vol 80-6.indd 228 12-10-15 15:19
V o l u m e 8 0 | I s s u e 5 | O c t 0 b e r 2 0 1 5 229
Osterc and Štampar | Maturation changes auxin profile during the process of adventitious rooting in Prunus
DiscussionFour hours after being inserted in the substrate, the IBA
treated cuttings showed higher IAA values in their bases in juvenile and mature cuttings than the cuttings with no IBA treatment before being transferred in the substrate. This result showed that IBA metabolised very quickly in IAA in the cherry species ‘Autumnalis’. This conversion occurred during the first 4 hours after severance. Our re-sults support some previous findings (Epstein et al., 1993; Ludwig-Müller, 2000) about IBA metabolisation after its absorption but all these findings reported that this conver-sion lasts longer, until day 2. We succeeded to show in some our previous experiments with Prunus subhirtella ‘Autum-nalis’ (Osterc and Štampar, 2011) that the conversion is probably faster, within day 1 after severance, as already mentioned for some other species (Nag et al., 2001). The results of the presented experiment move this level even further back, toward as little as 4 hours after severance. Surely, it has to be taken into account that this result was obtained in Prunus subhirtella, and that other species can react differently.
The conversion from IBA to IAA was especially signifi-cant in juvenile cuttings where already 4 hours after the insertion of cuttings a significant IAA-peak occurred as a result of the conversion. Later on, on day 1 and day 2 the IAA values were lower. The IAA-peak after 4 hours was noticed in juvenile cuttings also in the case when cuttings did not receive any hormone pre-treatment. Based on ex-periments of Nag et al. (2001), who linked the duration of the induction phase with an IAA-peak, the length of the in-duction phase in our juvenile cuttings was very short, only 4 hours. This early IAA increase should be optimal for the root induction process because juvenile cuttings showed a significantly better root formation compared with mature and semi-mature cuttings. Interestingly, there was no IAA increase after severance in not pre-treated semi-mature cuttings. In pre-treated cuttings the IAA-peak was noticed only on day 1. Maturation in plant material changed the length of the induction phase and had a negative effect on root formation as shown by lower rooting rates in semi-mature cuttings. Our results support the hypothesis that it could be more optimal for the root induction process when IAA peak occurs before day 1 as previously reported (Nag et al., 2001; Štefančič et al., 2007).
In our experiment the true juvenility could be ob-served only in the case of the originally in vitro derived stock plants. Regarding root formation cutting stock plants (semi-mature cuttings) reacted similarly as mature cut-tings, although the rooting success was higher than in mature cuttings but not statistically significant. A similar result has already been obtained also in our previous ex-periment where the semi-mature cuttings even showed the lowest rooting success of all (Osterc and Štampar, 2011). An exogenous application of IBA tended to improve rooting in mature and juvenile cuttings but the difference was not sig-nificant. This result clearly supported numerous findings (Osterc et al., 2009; Spethmann, 1997, 2007) that exoge-nous auxin treatment is not obligatory for the root induc-tion process itself. The bases of the cuttings always contain a specific endogenous hormone level derived from the api-cal parts of the shoots. If the stock plants grow well, the IAA synthesis in shoots of these plants is optimal as well. This naturally originating IAA is accumulated subsequently in shoot bases (as a result of polar auxin transport) and this
IAA level is normally sufficient for the root induction (Ban-durski et al., 1995; Ford et al., 2002). Exogenous hormone treatment can only slightly affect the endogenous hormone status due to the weaker hormone absorption from cut-tings, whose tissues are about lignifying at that time. On the other hand, we were able to show in some previous results that the absorbed auxin did not remain in cutting bases (root emergence zone) in its full concentration but that it was partly transported with the mass-flow to the upper cutting parts (Osterc and Spethmann, 2001; Osterc and Štampar, 2011). Therefore, this application should only affect the quality of the developed root system. In contrast, in our experiment the cuttings without hormone pre-treat-ment developed more and longer roots. The fact is that the quality of the root system is not affected only by endoge-nous factors (hormone level and exogenous hormone ap-plication which effect this level) but also exogenously, by substrate temperature and humidity, by the transpiration rate of cuttings etc. (Spethmann, 1997). Unsuitable exoge-nous factors (occasionally problems with fogging system) have also been the main reasons for slightly weaker results (root quality) in the case of hormone pre-treatment in our experiment.
The IAA-Asp concentrations were significantly higher in the bases of mature cuttings on all analysing dates re-gardless of the hormone pre-treatment of cuttings. This result also strongly supports our previous results with this cherry species (Osterc and Štampar, 2011). The IAA-Asp values in the bases of cuttings, which had been previously pre-treated with IBA, were at a similar level or even smaller than in the cuttings with no IBA pre-treatment. The natu-rally synthesized IAA metabolised into the IAA-Asp with the strongest rate in mature cuttings. This means that IAA was strongly inactivated especially in mature cuttings and it could be assumed that the lowest potential for root de-velopment in mature cuttings was in fact the result of this inactivation. The results with mature cuttings strongly support previous findings about the IAA inactivation in the form of IAA-Asp (Bartel et al., 2001; Osterc et al., 2009; Osterc and Štampar, 2011; Ludwig-Müller, 2011). The IAA-Asp can be therefore recognised as the main compound of blocking the root induction process due to the inactivation of IAA.
ConclusionsThe results of the present experiment clearly show that
different physiological levels of the propagating material changed the IAA-profile in cuttings during the root induc-tion and initiation phase. Regarding the propagation re-sults, the most successful in vitro derived material showed an extremely short induction phase, only 4 hours, while, on the other hand, semi-mature and mature cuttings showed a prolonged induction phase, until day 1 after severance. Less successful propagation results in mature cuttings had to be additionally ascribed to a strong conversion of IAA to IAA-Asp, a very well-known inactive form of IAA. The role of other possible IAA conjugates regarding rooting success in woody plants stays unclear. This is an open issue which needs to be tackled in the future.
AcknowledgmentsThis work is part of the Programme Horticulture No.
P4-0013-0481 financed by the Slovenian Research Agency.
eJHS Magazine Vol 80-6.indd 229 12-10-15 15:19
230 E u r o p e a n J o u r n a l o f H o r t i c u l t u r a l S c i e n c e
Osterc and Štampar | Maturation changes auxin profile during the process of adventitious rooting in Prunus
ReferencesBandurski, R.S., Cohen, J.D., Slovin, J.P., and Reinecke, D.M. (1995). Auxin biosynthesis and metabolism. In Plant Hormones: Physiology, Biochemistry and Molecular Biology, P. Davies, ed. (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 39–65. http://dx.doi.org/10.1007/978-94-011-0473-9_3.
Bartel, B., LeClere, S., Magidin, M., and Zolman, K.B. (2001). Inputs to the active indole-3-acetic acid pool: de novo synthesis, conjugate hydrolysis, and indole-3-butyric acid β-oxidation. J. Plant Growth Regul. 20, 198–216. http://dx.doi.org/10.1007/s003440010025.
De Klerk, G.-J., Keppel, M., Ter Brugge, J., and Meekes, H. (1995). Timing of the phases in adventitious root formation in apple microcuttings. J. Exp. Bot. 46, 965–972. http://dx.doi.org/10.1093/jxb/46.8.965.
Dodd, R.S., and Power, A.B. (1988). Clarification of the term topophysis. Silvae Genetica 37, 14–15.
Epstein, E., Zilkah, S., Faingersh, G., and Rotebaum, A. (1993). Transport and metabolism of indole-3-butyric acid in easy- and difficult-to-root cuttings of sweet cherry (Prunus avium L.). Acta Hortic. 329, 292–295. http://dx.doi.org/10.17660/actahortic.1993.329.69.
Ford, Y.-Y., Bonham, E.C., Cameron, R.W.F., Blake, P.S., Judd, H.L., and Harrison-Murray, R.S. (2002). Adventitious rooting: examining the role of auxin in an easy- and a difficult-to-root plant. Plant Growth Regul. 36(2), 149–159.
Fortanier, E.J., and Jonkers, H. (1976). Juvenility and maturity of plants as influenced by their ontogenetical and physiological ageing. Acta Hortic. 56, 37–44. http://dx.doi.org/10.17660/actahortic.1976.56.2.
Gonçalves, J.C., Diogo, G., Coelho, M.T., Vidal, N., and Amancio, S. (2008). Quantitation of endogenous levels of IAA, IAAsp and IBA in micro-propagated shoots of hybrid chestnut pre-treated with IBA. In Vitro Cell Dev. Biol.-Plant 44, 412–418. http://dx.doi.org/10.1007/s11627-008-9151-0.
Kovač, M., Piskernik, D., and Ravnikar, M. (2003). Jasmonic acid-induced morphological changes are reflected in auxin metabolism of beans grown in vitro. Biol. Plant. 47, 273–275. http://dx.doi.org/10.1023/B:BIOP.0000022264.47660.17
Ludwig-Müller, J. (2000). Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 32, 219–230. http://dx.doi.org/10.1023/A:1010746806891.
Ludwig-Müller, J. (2011). Auxin conjugates: their role for plant development and in the evolution of land plants. J. Exp. Bot. 62, 1757–1773. http://dx.doi.org/10.1093/jxb/erq412.
Morgan, D.L., and McWilliams, E.L. (1976). Juvenility as a factor in propagating Quercus virginiana Mill. Acta Hortic. 56, 263–268. http://dx.doi.org/10.17660/actahortic.1976.56.30.
Nag, S., Saha, K., and Choudhuri, M.A. (2001). Role of auxin and polyamines in adventitious root formation in relation to changes in compounds involved in rooting. J. Plant Growth Regul. 20, 182–194. http://dx.doi.org/10.1007/s003440010016.
Olesen, P.O. (1978). On cyclophysis and topophysis. Silvae Genetica 27, 173–216.
Osterc, G. (2009). A change in perspective: stockplant qualities that influence adventitious root formation of woody species. In Adventitious root formation of forest trees and horticultural plants – from genes to applications, K. Niemi and C. Scagel, eds. (Kerala, India: Research Signpost), pp. 175–185.
Osterc, G., and Spethmann, W. (2001). Studies on auxin uptake in Prunus and Malus green cuttings. Propagation of Ornamental Plants 1, 3–9.
Osterc, G., and Štampar, F. (2011). Differences in endo/exogenous auxin profile in cuttings of different physiological ages. J. Plant Physiol. 168, 2088–2092. http://dx.doi.org/10.1016/j.jplph.2011.06.016.
Osterc, G., Štefančič, M., and Štampar, F. (2009). Juvenile stock plant material enhances root development through higher endogenous auxin level. Acta Physiol. Plant. 31, 899–903. http://dx.doi.org/10.1007/s11738-009-0303-6.
Plietzsch, A., and Heiliger, P. (1997). In-vitro-vermehrte Mutterpflanzen den konventionellen überlegen. Deutsche Baumschule 2, 62–64.
Sándor, G., Bodor, P., Jócsák, I., Brunori, A., Tóth, M., and Végvári, Gy. (2010). Hardwood cuttings preparation timing and effect on the IBA uptake and metabolism in Prunus rootstocks. Propagation of Ornamental Plants 10, 75–80.
Seeliger, R. (1924). Topophysis und Zyklophysis pflanzlicher Organe und ihre Bedeutung für die Pflanzenkultur. Angewandte Botanik 6, 191–200.
Spethmann, W. (1997). Autovegetative Gehölzvermehrung. In Die Baumschule, G. Krüssmann, ed. (Berlin, Germany: Parey Verlag), pp. 382–449.
Spethmann, W. (2007). Increase of rooting success and further shoot growth by long cuttings of woody plants. Propagation of Ornamental Plants 7, 160–166.
Štefančič, M., Štampar, F., Veberič, R., and Osterc, G. (2007). The levels of IAA, IAAsp and some phenolics in cherry rootstock ‘GiSelA 5’ leafy cuttings pretreated with IAA and IBA. Sci. Hort. 112, 399–405. http://dx.doi.org/10.1016/j.scienta.2007.01.004.
Went, F.W. (1934). A test method for rhizocaline, the root forming substance. Proceedings of the Section of Sciences, Koninklijke Akademie van Wetenschappen te Amsterdam, Amsterdam 37, 445–455.
Woodward, A.W., and Bartel, B. (2005). Auxin: regulation, action, and interaction. Annals of Botany 95, 707–735. http://dx.doi.org/10.1093/aob/mci083.
Received: Jun. 2, 2014Accepted: Mar. 26, 2015
Addresses of authors: Gregor Osterc* and Franci ŠtamparUniversity of Ljubljana, Biotechnical Faculty, Institute for Fruit Growing, Viticulture and Vegetable Growing, Jamni-karjeva 101, 1000 Ljubljana, Slovenia* Corresponding author; E-mail: [email protected]