Contents lists available at SciVerse ScienceDirect
Respiratory Physiology & Neurobiology
j our nal ho me p age: www.elsev ier .com/ locate / resphys io l
ge and hypothyroidism affect dopamine modulation of breathing and D2
eceptor levels
ory Sykora, Mitch Amor, Evelyn Schlenker ∗
asic Biomedical Sciences, Sanford School of Medicine of the University of South Dakota, United States
r t i c l e i n f o
rticle history:ccepted 2 October 2012
eywords:ypothyroidismypoxiaopamine D2 receptor
a b s t r a c t
During and following hypoxic exposure young male hypothyroid hamsters treated with the dopamine D2
receptor agonist bromocriptine increased breathing, while ventilation was depressed in bromocriptine-treated euthyroid hamsters. Moreover, D2 receptor expression was increased in carotid bodies andstriatum, but not in the nucleus tractus solitaries (NTS) of hypothyroid relative to euthyroid hamsters.Here ventilation was determined in older male hypothyroid and euthyroid hamsters given vehicle orbromocriptine, and exposed to baseline air, hypoxia, and then air. Bromocriptine without hypoxia served
arotid bodytriatumTS
as a time control. Relative to vehicle, bromocriptine depressed ventilation in both groups exposed to airor to hypoxia, but hypothyroid bromocriptine-treated hamsters increased ventilatory responsiveness tohypoxia, while euthyroid hamsters decreased ventilatory responsiveness to hypoxia and exhibited post-hypoxic depression. Hypothyroidism had no effect on D2 receptor expression in carotid bodies or striatum,but increased it in the NTS. Thus, in hamsters bromocriptine modulates breathing and expression of D2
e len
receptor depending on th
. Introduction
Hypothyroidism is a common endocrine disorder that is classi-ed by lower than normal levels of circulating thyroid hormones.hysiologically thyroid hormones, thyroxine (T4) and triiodothyro-ine (T3) control the rate of metabolic reactions in the body andlay essential roles in the maturation and function of the centralervous system and other organs (Anderson, 2001; Dussault anduel, 1987; Schlenker, 2012). The prevalence of hypothyroidism inhe United States ranges from 4.6% to 9.5% in adults and is depend-nt on age and gender (Cannaris et al., 2000). Worldwide iodineeficiency is the most common cause of hypothyroidism, but eithern absence of the thyroid gland or a deficiency in stimulating hor-ones from the hypothalamus or pituitary gland can also cause
ypothyroidism (Rugge et al., 2011). Treatment of hypothyroidismncludes administration of synthetic and animal-derived forms of4 and T3; however, treatments with thyroid hormones do notlways normalize function (Jha et al., 2006).
The brain has long been known to be extremely sensitive to
he effects of thyroid hormones, influencing neural developmentnd function (Jedrusiak et al., 1995). Effects of hypothyroidismay cause cognitive dysfunction resulting from altered central
∗ Corresponding author at: Basic Biomedical Sciences, Sanford School of Medicinef the University of South Dakota, Vermillion, SD 57069, United States.el.: +1 605 677 6150; fax: +1 605 677 6381.
catecholamine expression, specifically dopamine (Oh-Nishi et al.,2005; Akaike et al., 1991). Dopamine is a major neurotransmitteralso affecting voluntary movement, mood, attention, and mem-ory (Beaulieu and Gainetdinov, 2011). Dopamine’s physiologicalactions are exerted by G protein-coupled receptors which aredivided into two major subtypes: D1-like (D1 and D5) and D2-like(D2, D3 and D4) (Beaulieu and Gainetdinov, 2011; Oh-Nishi et al.,2005). Amongst the dopaminergic receptors, the D2-like receptorsreduce adenylate cyclase activity and stimulate opening K+ chan-nels, which are responsible for the predominantly inhibitory effectsof D2 receptors (Missale et al., 1998).
Some of the highest levels of dopaminergic D2 receptors in thecentral nervous system are found in the striatum. An abundantamount of D2 receptors are also located within the nucleus tractussolitaries (NTS, a respiratory integratory region) and peripherallywithin the glomus cells of the carotid body (CB) (Kline et al., 2002;Beaulieu and Gainetdinov, 2011; Schlenker and Schultz, 2012).While the NTS and CB are well known to modulate breathing(McKay et al., 2003; Bairam and Carroll, 2005; Evans et al., 1999),especially responses to hypoxia, studies in humans also indicatedthat the striatum plays a role in regulation and control of breathing(Evans et al., 1999; McKay et al., 2003).
Exposure to hypoxia can induce changes in the dopaminergiccarotid body system by altering the chemoreceptor sensitivity to
oxygen (Bairam and Carroll, 2005; Simsek et al., 2004). In experi-ments with hypothyroid rats exposed to hypoxia, ventilation waslower in hypothyroid animals compared to euthyroid animals dueto alterations in the function of the carotid body (Olea et al., 2007).
everal other animal and human experiments also confirm theole of hypoxic exposure on D2 receptor modulation of breath-ng (Bascom et al., 1991; Huey et al., 2003; Pedersen et al., 1997;chlenker, 2007). Most studies indicate that stimulation of D2eceptors depresses ventilation, but some have reported an excit-tory effect of D2 receptors on ventilation during and followingxposure of animals to hypoxia (Schlenker and Schultz, 2012).
Moreover, hypothyroidism can affect the function and expres-ion of D2 receptors in several brain regions and the carotid bodySchlenker, 2007; Guner et al., 2002; Schlenker and Schultz, 2012).or example, Cameron and Crocker (1990) demonstrated that treat-ng adult rats with propylthiouracil (PTU) for 4 weeks did not affecttriatal D2 receptor levels when compared to those of euthyroidats, but PTU-induced hypothyroid rats displayed a larger behav-oral sensitivity to D2 receptor agonists. When rats were renderedypothyroid for 10 weeks, not only did striatal D2 receptor lev-ls increased relative to those of euthyroid rats, but behavioralesponses to apomorphine also were enhanced (Crocker et al.,986). Thus, longer term hypothyroidism has an effect on thexpression of D2 receptors and D2-receptor mediated behavioralhenotypes. In a more recent study, Schlenker and Schultz (2012)lso reported an increase in D2 receptor protein levels in the stri-tum and carotid body, but no difference not in the nucleus tractusolitaries (NTS) of male hamsters rendered hypothyroid for threeonths relative to euthyroid animals of the same age. What effect
onger term hypothyroidism would have on D2 receptors in maleamsters had not been investigated.
In addition, Schlenker and Schultz noted that bromocriptine, D2 receptor agonist, administered to hypothyroid and euthy-oid hamsters had no effect on ventilation during the exposuref animals to air, but bromocriptine stimulated ventilation inypothyroid hamsters during and after exposure of hypoxia rela-ive to effects of vehicle (Schlenker and Schultz, 2012). In contrast,uthyroid hamsters receiving bromocriptine decreased their ven-ilation during and following exposure to hypoxia compared toheir responses to vehicle treatments (Schlenker and Schultz,012). Thus, hypothyroidism affected ventilation in hypothyroidnd euthyroid hamsters in opposite ways during and followingxposure to hypoxia.
In the present study we evaluated the effects of the longer termTU treatment of male hamsters on ventilation treated with vehicler bromocriptine before, during, and after exposure of hypoxia. Inddition we investigated if longer term PTU treatment affected pro-ein levels of D2 receptors in the NTS, striatum and carotid bodies.
e hypothesized that the older hypothyroid hamsters would showlevated D2 receptors in the carotid bodies, NTS and striatum com-ared to euthyroid hamsters as reported by Schlenker and Schultz2012) in younger hamsters. In addition, treatment with the D2eceptor agonist bromocriptine should increase ventilation duringnd after exposure to hypoxia in hypothyroid hamsters relativeo respective vehicle. We extended our measurements to 10 minollowing the hypoxic exposure to determine if the post hypoxicesponses were longer lasting than what we had seen in previoustudies.
. Experimental procedures
.1. Animals
Two-month-old male golden Syrian hamsters were acquiredrom Harlan Sprague Dawley Inc. (Indianapolis, IN). Animals were
ept in groups of 3–4 in Ancare cages and exposed to 14 h of lightnd 10 h of darkness (to maintain normal thyroid hormone levelsn the euthyroid hamsters, Saita et al., 2005; Vaughan et al., 1982).he hamsters were divided into two groups. Eight hamsters were
Neurobiology 185 (2013) 257– 264
given tap water to drink and composed the euthyroid group. Theother nine hamsters received 0.04% propylthiouracil (PTU) in tapwater to induce hypothyroidism by preventing conversion of thy-roxine to triiodothyronine (Oppenheimer et al., 1972). Experimentswere conducted in both groups starting 5 months following theinitiation of treatments. Food (8604 rodent pellets obtained fromHarlan Sprague Dawley) and fluids (water or PTU) were availablead libitum. All procedures used in this study were approved by theUniversity of South Dakota Animal Care and Use Committee.
2.2. Respiratory chamber
Respiratory measurements were performed in hamsters placedinto a 20.2 cm × 7.9 cm Plexiglas cylindrical chamber using thebarometric method (Schlenker et al., 1994). One end of the cham-ber was closed but included ports for calibration (using a 1 ml glasssyringe), measuring chamber temperature (using a Fisher Scien-tific digital thermometer), determining pressure changes withinthe chamber associated with breathing (using a Statham low pres-sure transducer coupled to an Acknowledge system by BioPac dataacquisition system), and an input port for air or 10% oxygen in nitro-gen. Fractional contents of oxygen were measured with Vacu-MedModel 17515A gas analyzer. Barometric pressure was determinedusing a W.M. Welch (Chicago, IL) mercury barometer. Relativehumidity within the chamber averaged 50%. Inspiratory and expi-ratory times were quantified from 10 consistent breaths averagedfor each portion of the study and was used to calculate frequencyof breathing by adding inspiratory and expiratory times and divid-ing the sum into 60 (s/min). The minute ventilation was determinedby multiplying frequency by tidal volume. Tidal volume and minuteventilation were multiplied by 1000/body weight (BW).
2.3. Ventilatory studies
Three days prior to the commencement of experiments, animalswere acclimated to the chamber for 30 min each day. On the dayof an experiment hamsters were weighted and injected subcuta-neously with either vehicle (saline) or 1.0 mg/kg bromocriptine insaline (Tocris Bioscience, Ellisville, MO). This dose of bromocrip-tine was used a previous study (Schlenker and Schultz, 2012). Afterreceiving a subcutaneous injection, the hamster was placed intochamber for a 35-min acclimation period to room air. During thelast minute of exposure to air, the frequency, tidal volume, andminute ventilation were determined.
The hamster was then exposed to hypoxia (10% oxygen in nitro-gen) for 5 min. During the last minute of exposure to hypoxia, tidalvolume, frequency, and minute ventilation were determined. Afterthe hypoxic exposure, the chamber was washed out with room airfor 5 and 10 min and tidal volume, frequency, and minute ventila-tion were determined during the last minute of each time period.Following removal from the chamber, the hamster’s body temper-ature was measured using a Sensortek BAT-12 thermometer and aPhysiotemp thermocouple.
In a separate experiment in the same group of animals, hamsterswere given 1.0 mg/kg bromocriptine and underwent the same pro-tocol outlined above. Instead of being exposed to hypoxia for 5 min,the hamster was exposed to room air for 5 and 10 min in its placeas a time control study. Studies were randomized with at least 3days between treatments.
2.4. Immunohistochemistry
Following the ventilatory studies, hamsters were deeplyanesthetized and first perfused transcardially with phosphatebuffered saline (PBS) and then 4% paraformaldehyde in PBS. Thebrain, carotid bodies, and thyroid glands were quickly removed,
ogy &
psa
dwss
wdoafisD11V3l
ccfgoc
aaptoacficnto
2
ava(pcdwtpA
3
3w
c
C. Sykora et al. / Respiratory Physiol
ost-fixed for 5 h in 4% paraformaldehyde and then placed into 25%ucrose in PBS. Thyroid glands were weighed. The carotid bodiesnd brains were frozen at −80 ◦C.
Carotid bodies from hypothyroid and euthyroid hamsters wereehydrated and then embedded in paraffin. 7–10 �m thin slicesere cut and mounted on slides. Prior to immunohistochemical
tudies, sections were deparaffined, and placed in a sodium citrateolution heated in a vegetable steamer for antigen retrieval.
Brains were cut using a freezing microtome at 40 �m slices thatere then stored in cryoprotectant. Two areas of the striatum, theorsal and ventral regions (1.5–0.8 Bregma) and the medial nucleusf the NTS (−10.3 mm relative to Bregma) were investigated (Morinnd Wood, 2001). To evaluate D2 receptor protein levels, slicesrom the striatum, NTS, and carotid bodies were incubated in H2O2n 0.1 M phosphate buffered saline (PBS), blocked using 10% goaterum for 1 h, and then incubated in primary antibody (polyclonal2 receptor antibody produced in rabbit, Sigma–Aldrich, AB5084P,:1000). After an overnight incubation, tissues were incubated for
h in a biotinylated goat anti-rabbit secondary antibody (1:400,ector), placed in Avidin Biotin Complex, and then developed using,3′-diaminobenzidine and H2O2. Sections of the carotid body were
ightly counterstained with hemotoxylin.To quantify the relative amount of D2 receptors in the glomus
ells, NIH ImageJ software (rsb.info.nih.gov/ij) was used. Specifi-ally, the largest cross sections of carotid bodies were determinedrom each hamster and the cross sectional areas of all D2 positivelomus cells were measured in that carotid body. Then the ratiof the sum of glomus cells expression D2 receptors to carotid bodyross sectional area was calculated.
To evaluate D2 receptor expression in the two parts of the stri-tum and the NTS, at least three slices from each animal werenalyzed using NIH ImageJ software. Squares of a constant size werelaced on the same two locations in the ventral and dorsal striatumhe density of D2 receptor expression was determined as a relativeptical density (OD). In the NTS, the area of positive neurons in
square of a constant size in the same regions of the medial andentral nuclei of the NTS for all animals was determined at a magni-cation of 40×. For all immunohistochemical analysis, slides wereoded so that the analysis was conducted in a double-blinded man-er and slices from all the structures analyzed were run at the sameime to minimize differences of techniques during the processingf tissues.
.5. Statistical analysis
Statistical analysis of the ventilatory data consisted of two-waynalysis of variance (ANOVA) to test the effects of treatment (PTUersus tap water), drug administration (saline or bromocriptine),nd gas exposure (air or hypoxia). If the result was significantP < 0.05), post hoc Holm–Sidak test for paired and unpaired com-arisons were conducted. Body weight, thyroid gland weight, andarotid body, NTS, and striatum D2 receptor protein quantificationata were analyzed using unpaired Student’s t tests. If the dataere skewed, we used nonparametric test such as Mann–Whitney
est (for unpaired comparisons) or the Wilcoxon sign rank test (foraired comparisons). Data were analyzed using SPSS version 19.0.ll data are presented as means and standard deviation.
. Results
.1. Body weight (BW), body temperature and thyroid gland
eight
Body weights of the PTU-treated hamsters (154.9 ± 8.7 g) wereomparable to those of euthyroid hamsters (161.0 ± 13.8). Body
Neurobiology 185 (2013) 257– 264 259
temperatures were not different between the two groups andbromocriptine had no effect on this parameter (data not shown).Thyroid gland weights were significantly greater in hypothy-roid compared to euthyroid hamsters (214.9 ± 16.8 mg versus14.1 ± 2.1 mg, P < 0.001).
3.2. D2 receptor protein levels in carotid bodies, striatum, and NTS
D2 receptors were expressed in the carotid bodies of euthy-roid and hypothyroid animals. Carotid body D2 receptor proteinlevels relative to total carotid body areas were not significantlydifferent between the groups (hypothyroid = 0.23 ± 0.06 and euthy-roid = 0.25 ± 0.05).
In the striatum, D2 receptors protein levels (measured as opti-cal densities) were significantly lower in the ventral-striatum ofthe PTU-treated hamsters (0.18 ± 0.02) as compared to the dorsal-striatum of the PTU-treated hamsters (0.20 ± 0.02, P = 0.004).Likewise in the euthyroid hamster there was a lower level of D2receptor protein levels in the ventral-striatum (0.17 ± 0.02) versusthe dorsal-striatum (0.19 ± 0.02, P = 0.003). However, D2 receptorprotein levels in each region of the striatum were comparablebetween the two groups.
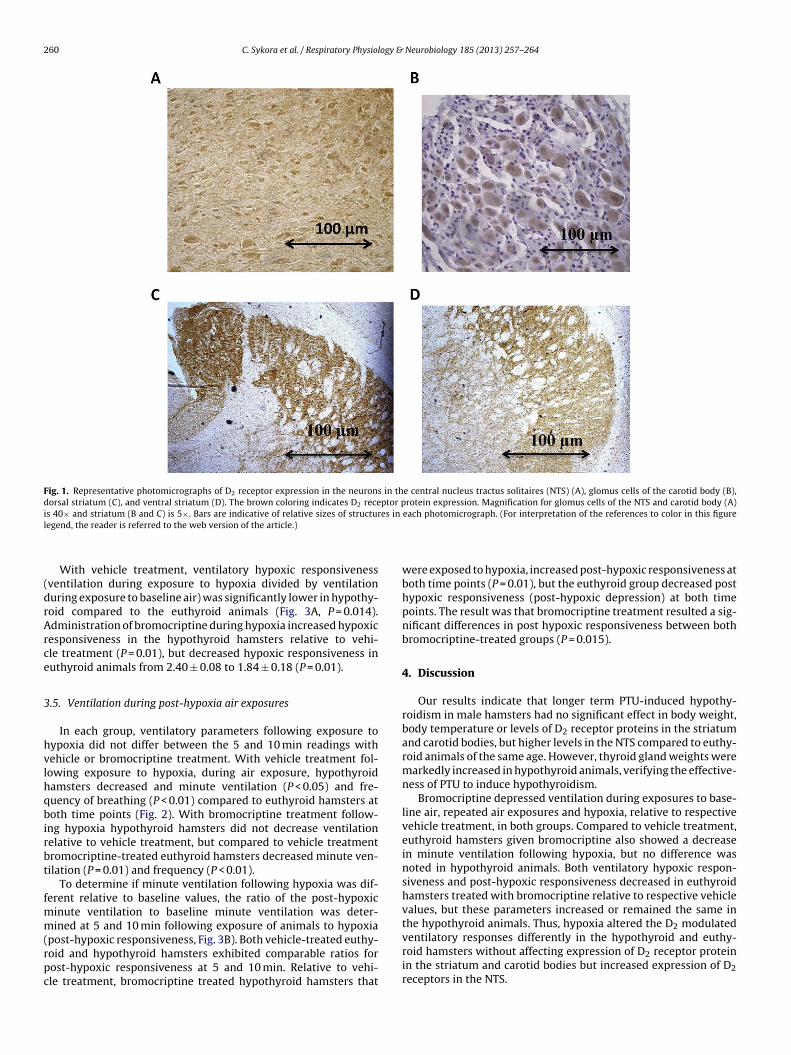
In the NTS, the relative optical density of D2 positive neuronswas significantly greater in the hypothyroid compared to the euthy-roid animals (80.3 ± 12.1 versus 66.7 ± 10.5, P < 0.05). These resultscontrast to those found in the striatum and carotid bodies. A rep-resentative photomicrograph of D2 receptor protein expression ofa euthyroid hamster in the 4 regions is shown in Fig. 1.
3.3. Ventilation during exposure to baseline air
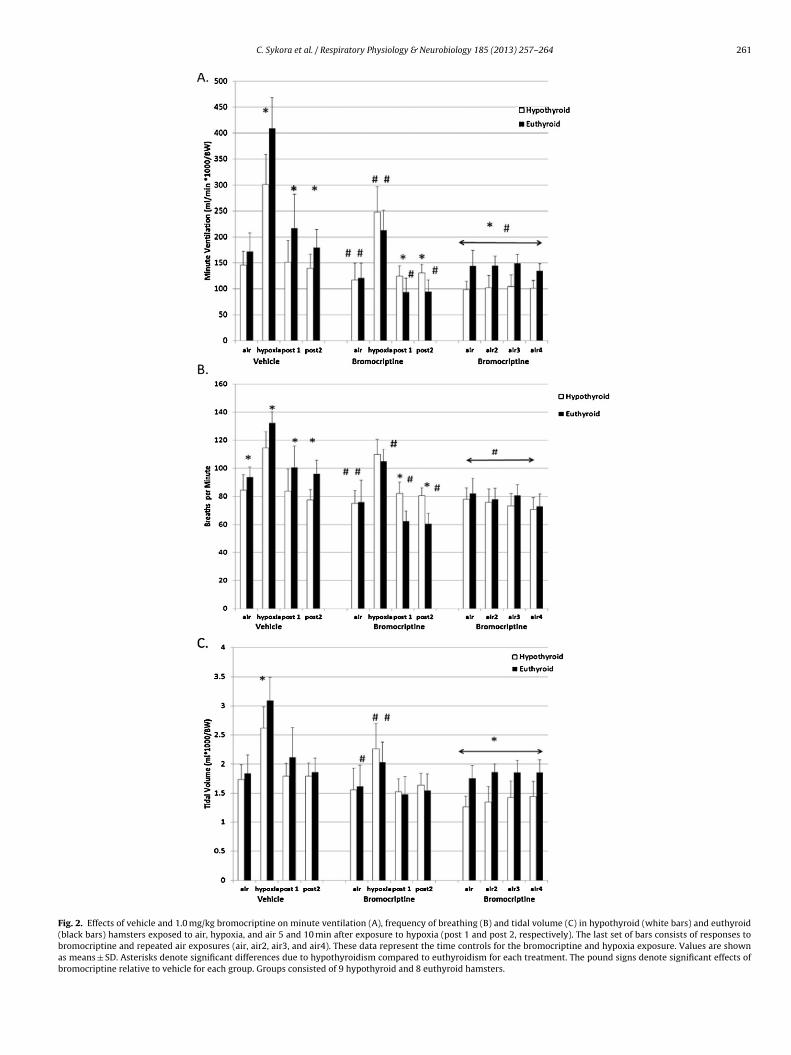
During room air baseline conditions, vehicle-treated hypothy-roid hamsters exhibited a decreased frequency of breathing (Fig. 2B,P < 0.05) relative to euthyroid animals. Treatment with bromocrip-tine during air exposure, prior to hypoxia decreased minuteventilation in both euthyroid (P = 0.02) and hypothyroid (P = 0.002)groups compared to their respective vehicle treatments, due toa significant decrease in tidal volume (P = 0.03) and frequency ofbreathing (P = 0.03) in the euthyroid hamsters. In the hypothyroidhamsters relative to vehicle treatment these two parameters werelower but not significantly so. Thus, relative to vehicle administra-tion of bromocriptine decreased minute ventilation in both groupsof hamsters.
Of interest is that during bromocriptine treatment followed byrepeated air exposures (last set of bars in Fig. 2), minute ven-tilation was lower than vehicle treated animals of both groups(P < 0.01), but also lower in the hypothyroid relative to the euthy-roid group (P < 0.05). This effect was due to a lower tidal volumein the hypothyroid animals, but not a decrease in frequency ofbreathing.
3.4. Ventilation during exposure to hypoxia
With vehicle treatment, euthyroid hamsters exposed to hypoxiashowed a higher minute ventilation than hypothyroid hamsters(Fig. 2, P < 0.002) due to a significantly greater tidal volume(P = 0.04) and frequency of breathing (P = 0.005). Administration ofbromocriptine decreased ventilation significantly in both groups(P < 0.01 for both), and eliminated the ventilatory differencesbetween the two groups. The decrease in minute ventilation inthe bromocriptine-treated animals during hypoxia was due to adecrease in tidal volume for both groups (euthyroid P = 0.003 and
hypothyroid P = 0.04) and a decrease in frequency (P = 0.005) in theeuthyroid hamsters. Thus, bromocriptine appears to have a greaterdepressive effect on ventilation during exposure to hypoxia in theeuthyroid group than on the hypothyroid group.
260 C. Sykora et al. / Respiratory Physiology & Neurobiology 185 (2013) 257– 264
Fig. 1. Representative photomicrographs of D2 receptor expression in the neurons in the central nucleus tractus solitaires (NTS) (A), glomus cells of the carotid body (B),dorsal striatum (C), and ventral striatum (D). The brown coloring indicates D2 receptor protein expression. Magnification for glomus cells of the NTS and carotid body (A)i es in
l
(drArce
3
hvlhqbirbt
fmm(rpc
s 40× and striatum (B and C) is 5×. Bars are indicative of relative sizes of structuregend, the reader is referred to the web version of the article.)
With vehicle treatment, ventilatory hypoxic responsivenessventilation during exposure to hypoxia divided by ventilationuring exposure to baseline air) was significantly lower in hypothy-oid compared to the euthyroid animals (Fig. 3A, P = 0.014).dministration of bromocriptine during hypoxia increased hypoxicesponsiveness in the hypothyroid hamsters relative to vehi-le treatment (P = 0.01), but decreased hypoxic responsiveness inuthyroid animals from 2.40 ± 0.08 to 1.84 ± 0.18 (P = 0.01).
.5. Ventilation during post-hypoxia air exposures
In each group, ventilatory parameters following exposure toypoxia did not differ between the 5 and 10 min readings withehicle or bromocriptine treatment. With vehicle treatment fol-owing exposure to hypoxia, during air exposure, hypothyroidamsters decreased and minute ventilation (P < 0.05) and fre-uency of breathing (P < 0.01) compared to euthyroid hamsters atoth time points (Fig. 2). With bromocriptine treatment follow-
ng hypoxia hypothyroid hamsters did not decrease ventilationelative to vehicle treatment, but compared to vehicle treatmentromocriptine-treated euthyroid hamsters decreased minute ven-ilation (P = 0.01) and frequency (P < 0.01).
To determine if minute ventilation following hypoxia was dif-erent relative to baseline values, the ratio of the post-hypoxic
inute ventilation to baseline minute ventilation was deter-ined at 5 and 10 min following exposure of animals to hypoxia
post-hypoxic responsiveness, Fig. 3B). Both vehicle-treated euthy-oid and hypothyroid hamsters exhibited comparable ratios forost-hypoxic responsiveness at 5 and 10 min. Relative to vehi-le treatment, bromocriptine treated hypothyroid hamsters that
each photomicrograph. (For interpretation of the references to color in this figure
were exposed to hypoxia, increased post-hypoxic responsiveness atboth time points (P = 0.01), but the euthyroid group decreased posthypoxic responsiveness (post-hypoxic depression) at both timepoints. The result was that bromocriptine treatment resulted a sig-nificant differences in post hypoxic responsiveness between bothbromocriptine-treated groups (P = 0.015).
4. Discussion
Our results indicate that longer term PTU-induced hypothy-roidism in male hamsters had no significant effect in body weight,body temperature or levels of D2 receptor proteins in the striatumand carotid bodies, but higher levels in the NTS compared to euthy-roid animals of the same age. However, thyroid gland weights weremarkedly increased in hypothyroid animals, verifying the effective-ness of PTU to induce hypothyroidism.
Bromocriptine depressed ventilation during exposures to base-line air, repeated air exposures and hypoxia, relative to respectivevehicle treatment, in both groups. Compared to vehicle treatment,euthyroid hamsters given bromocriptine also showed a decreasein minute ventilation following hypoxia, but no difference wasnoted in hypothyroid animals. Both ventilatory hypoxic respon-siveness and post-hypoxic responsiveness decreased in euthyroidhamsters treated with bromocriptine relative to respective vehiclevalues, but these parameters increased or remained the same inthe hypothyroid animals. Thus, hypoxia altered the D2 modulated
ventilatory responses differently in the hypothyroid and euthy-roid hamsters without affecting expression of D2 receptor proteinin the striatum and carotid bodies but increased expression of D2receptors in the NTS.
C. Sykora et al. / Respiratory Physiology & Neurobiology 185 (2013) 257– 264 261
Fig. 2. Effects of vehicle and 1.0 mg/kg bromocriptine on minute ventilation (A), frequency of breathing (B) and tidal volume (C) in hypothyroid (white bars) and euthyroid(black bars) hamsters exposed to air, hypoxia, and air 5 and 10 min after exposure to hypoxia (post 1 and post 2, respectively). The last set of bars consists of responses tobromocriptine and repeated air exposures (air, air2, air3, and air4). These data represent the time controls for the bromocriptine and hypoxia exposure. Values are shownas means ± SD. Asterisks denote significant differences due to hypothyroidism compared to euthyroidism for each treatment. The pound signs denote significant effects ofbromocriptine relative to vehicle for each group. Groups consisted of 9 hypothyroid and 8 euthyroid hamsters.
262 C. Sykora et al. / Respiratory Physiology &
Fig. 3. Effects of vehicle and 1.0 mg/kg bromocriptine on hypoxic (A) and post-hypoxic responsiveness (B) in hypothyroid (white bars) and euthyroid (black bars)hamsters. Values are shown as mean ± SD. Asterisks denote significant differencesdue to hypothyroidism compared to euthyroid in each group. The pound signsdr
4
hlrtfealsltarttarlNhobr
the plasma membrane (Tirotta et al., 2008; Genedani et al., 2010;
enote significant effects of bromocriptine (hypoxia) relative to vehicle for eachespective group.
.1. Effects of hypothyroidism on D2 receptor protein levels
The results of the current study showed that in male hamstersypothyroidism induced by PTU for five months do not affect the
evels of D2 receptor protein in the carotid bodies or striatum whenelative to the euthyroid hamsters, but increased D2 expression inhe NTS. These outcomes are in contrast to an earlier study thatound treatment of hamsters with PTU for 3 months resulted inlevated D2 receptor protein levels in the carotid bodies and stri-tum, but had no effect on D2 receptor levels in the NTS relative toevels in euthyroid hamsters (Schlenker and Schultz, 2012). Othertudies performed on hypothyroid rats have indicated that theength of hypothyroidism also affected levels of D2 receptor pro-eins. For example short term hypothyroidism (4 weeks) did notffect striatal D2 receptor expression, but longer term hypothy-oidism (12 weeks) results in increased the levels of D2 receptor inhe striatum relative to euthyroid animals and increased D2 recep-or modulated behaviors such as sniffing and locomotion (Cameronnd Crocker, 1990; Crocker et al., 1986). Why long term hypothy-oidism in older hamsters produces comparable D2 concentrationevels in the carotid bodies and striatum, but higher levels in theTS to those of euthyroid hamsters is not clear. It is not knownowever if expression of D2 receptors decrease in hypothyroid
r increase in euthyroid hamsters over time to become compara-le in older hamsters. To determine the course of change in D2eceptors in carotid bodies and brain regions, measurements of D2
Neurobiology 185 (2013) 257– 264
receptor expressions need to be conducted simultaneously in olderand younger hypothyroid hamsters.
4.2. Effects of hypothyroidism on ventilation
Although ventilation was comparable in euthyroid andhypothyroid hamsters during baseline air exposure, hypothyroidhamsters exposed to hypoxia and air following hypoxia exhib-ited lower ventilation relative to euthyroid animals. In previousstudies we noted that ventilation was lower for hypothyroid ham-sters compared to euthyroid for all three breathing periods: air,hypoxia and air following hypoxia (Schlenker and Schultz, 2012).Past experiments concluded that the decrease in ventilation ofhypothyroid animals was due to a lower body temperature andmetabolic rate as well as reduced carotid body hypoxic sensitiv-ity relative to euthyroid animals (Schlenker et al., 1994; Schlenkerand Schultz, 2012; Olea et al., 2007). In addition, the presentstudy hypoxic responsiveness was lower in hypothyroid relativeto euthyroid hamsters, but no difference was shown in the post-hypoxic responsiveness between the two groups. These results forhypoxic and post-hypoxic responsiveness are similar to the find-ings in a previous study where hamsters were hypothyroid for threemonths (Schlenker and Schultz, 2012).
4.3. Effects of D2 receptor modulation on ventilation
Earlier studies have indicated that treatment with D2 receptoragonists decreased ventilation of animals during exposure to airand hypoxia (Huey et al., 2003; Gonzalez et al., 1994). The presentresults agree with conclusions from these studies, but contrastswith findings by Schlenker and Schultz (2012) who demonstratedno effect of bromocriptine on minute ventilation of younger ham-sters when exposed to air, prior to hypoxia.
Euthyroid hamsters in both studies (Schlenker and Schultz,2012 and the present study) treated with bromocriptine relativeto vehicle during and following exposure to hypoxia exhibited adecrease in ventilation, hypoxic responsiveness, and post-hypoxicresponsiveness compared to respective vehicle due to the normalinhibitory effects of D2 receptors. In contrast, the hypoxic andpost-hypoxic responsiveness of hypothyroid hamsters treatedwith bromocriptine and exposed to hypoxia displayed no dif-ference when compared to vehicle treatment. These results areopposite to those reported by Schlenker and Schultz (2012)as younger hypothyroid hamsters increased in hypoxic andpost-hypoxic responsiveness relative to vehicle while euthyroidanimals showed no difference in hypoxic responsiveness and adecrease in post-hypoxic responsiveness compared to respectivevehicle.
When hamsters were exposed to hypoxia and treated withbromocriptine longer term hypothyroidism resulted in less of adepression of ventilation compared with that of euthyroid ham-sters. How and why the inhibitory action of bromocriptine observedin hypothyroid hamsters during exposure to air was attenuatedduring and following hypoxia needs to be determined. Severalpossible mechanisms are currently being explored. The first mech-anism may be that compared to euthyroidism, hypothyroidismcauses a greater internalization of D2 receptors during exposureto hypoxia thus leaving the increased release of dopamine to actof D1 receptors. It has been well established that dopamine stim-ulation causes D2 receptor desensitization by dopamine-inducedD2 receptor endocytosis, resulting in the endosomal recycling ofD2 receptors which determines the abundance of D2 receptors on
Li et al., 2012). Since the function of D2 receptors is determined byplasma membrane levels of receptors, coupling to G-proteins andsecond messenger systems, alterations in receptor internalization,
ogy &
rahl(gmcc2
rrriaadfgsrJCyohithwrtbeadtc
drtwsdgoiwrat
A
Eac
R
A
C. Sykora et al. / Respiratory Physiol
ecycling, and degradation of D2 receptors, differences in any orll of these possibilities between the hypothyroid and euthyroidamsters may influence the differences noted in ventilation fol-
owing stimulation by bromocriptine and exposure to hypoxiaXiao et al., 2009; Celver et al., 2010). Possible ways to investi-ate these possibilities is use of confocal microscopy, an ELISAethod, and differential centrifugation and determination of colo-
alization of D2 receptors in plasma membrane and/or cytosolicompartments (Thibault et al., 2011; Chen et al., 2012; Lane et al.,012).
Another manner that could cause the differential effect of D2eceptor stimulation on ventilation in the two groups may beelated to the presynaptic versus postsynaptic location of the D2eceptors. D2 receptors may be located pre- and post-synapticallyn the form of D2 short and long isoforms, respectively (Beaulieund Gainetdinov, 2011). The D2 short isoforms affect the productionnd release of dopamine while the D2 long forms transduce effectsopamine and D2 agonists at the post synaptic site. A number ofactors can affect the amounts of these receptor isoforms includingenetics (also differentially affecting behavioral phenotypes), sexteroid hormone levels, development, neuropeptides such as neu-otensin, and exposure to hypoxia for 6–24 h (Guivarc’h et al., 1995;oseph and Bairam, 2004; Thibault et al., 2011; Lane et al., 2012;olelli et al., 2010). The effects of hypothyroidism on each isoform iset unknown and could possibly explain the underlying differencesbserved on ventilation between the hypothyroid and euthyroidamsters following bromocriptine treatment and hypoxia. Prelim-
nary studies (Schlenker and Sykora) in our laboratory do suggesthat D2 long receptors are lower in the NTS of hypothyroid femaleamsters compared to euthyroid animals. Moreover, If DL receptorsere internalized to a greater extent than DS receptors in hypothy-
oid hamsters exposed to hypoxia, this may be an explanation forhe findings in the present study. However, these animals had noteen challenged with either bromocriptine and/or hypoxia so theffects of these treatments on expression of the isoforms need to belso evaluated. Moreover, it may be possible to selectively knock-own DL isoforms in specific brain regions (such as the NTS) andhis may be another way of determining the role of this isoform onontrol of breathing.
In summary, longer periods of hypothyroidism in male hamstersid not affect body temperatures or change the expression of D2eceptors in the striatum or carotid bodies, but increase D2 levels inhe NTS relative to euthyroid animals. Stimulation of D2 receptorsith bromocriptine depressed breathing in both groups of ham-
ters breathing air and in euthyroid hamsters depressed breathinguring and following exposure to hypoxia, whereas under the sameas exposures bromocriptine showed less of a depressive effectn breathing in hypothyroid hamsters. Moreover, bromocriptines now available for treatment of patients with type II diabetes,
ho may also exhibit hypothyroidism (DeFronzo, 2011). From theesults of previous studies and the present investigation age mayffect control of breathing in patients treated with bromocriptine;hat possibility needs to be investigated.
cknowledgements
This project was funded by NIH Center of Biomedical Researchxcellence (COBRE) pilot grant P20 RR-05567. I would like tocknowledge Abha Chalpe for her technical help in embeddingarotid bodies.
eferences
nderson, G.W., 2001. Thyroid hormones and the brain. Frontiers in Neuroen-docrinology 22, 1–17.
Neurobiology 185 (2013) 257– 264 263
Akaike, M., Kato, N., Ohno, H., Kobayashi, T., 1991. Hyperactivity and spatial mazelearning impairment of adult rats with temporary neonatal hypothyroidism.Neurotoxicology and Teratology 13, 317–322.
Bascom, D.A., Clement, I.D., Dorrington, K.L., Robbins, P.A., 1991. Effects of dopamineand domperidone on ventilation during isocapnic hypoxia in humans. Respira-tion Physiology 85, 319–328.
Bairam, A., Carroll, J.L., 2005. Neurotransmitters in carotid body development. Respi-ratory Physiology and Neurobiology 149, 217–232.
Beaulieu, J.-M., Gainetdinov, R., 2011. The physiology signaling, and pharmacologyof dopamine receptors. Pharmacological Research 63, 182–217.
Cameron, D.L., Crocker, A.D., 1990. The hypothyroid rat as a model of increased sensi-tivity to dopamine receptor agonists. Pharmacology Biochemistry and Behavior37, 627–632.
Cannaris, G.J., Manowitz, N.R., Mayor, G., Ridgway, E.C., 2000. Colorado thyroid dis-ease prevalence study. Archives of Internal Medicine 160, 526–534.
Celver, J., Sharma, M., Kovoor, A., 2010. RGS9-2 mediates specific inhibition ofagonist-induced internalization of D2-dopamine receptors. Journal of Neuro-chemistry 114, 739–749.
Chen, H.T., Ruan, N.Y., Chen, J.C., Lin, T.Y., 2012. Dopamine D2 receptor-mediatedAkt/PKB signaling: initiation by D2S receptor and role in quinpirole-inducedbehavioral activation. ASN Neuro 4 (6), e00098.
Colelli, V., Fiorenza, M.T., Conversi, D., Orsini, C., Cabib, S., 2010. Strain-specificproportion of the two isoforms of the dopamine D2 receptor in the mouse stri-atum: associated neural and behavioral phenotypes. Genes, Brain and Behavior9, 703–711.
Crocker, A.D., Overstreet, D., Crocker, J.M., 1986. Hypothyroism leads to increaseddopamine receptor sensitivity and concentration. Pharmacology Biochemistryand Behavior 24, 1593–1597.
DeFronzo, R.A., 2011. Bromocriptine: a sympatholytic, D2-dopamine agonist for thetreatment of Type 2 diabetes. Diabetes Care 34, 789–794.
Dussault, J.H., Ruel, J., 1987. Thyroid hormones and brain development. AnnualReview of Physiology 49, 321–334.
Evans, K.C., Shea, S.A., Saykin, A.J., 1999. Functional MRI localisation of central ner-vous system regions associated with volitional inspiration in humans. Journalof Physiology 520, 383–392.
Genedani, S., Carone, C., Guidolin, D., Filaferro, M., Marcellino, D., Fuxe, K., Agnati,L.F., 2010. Differential sensitiviy of A2A and especially D2 receptor trafficking tococaine compared with lipid rafts in cotrasfected CHO cell lines. Novel actions ofcocaine independent of the DA transporter. Journal of Molecular Neuroscience41, 347–357.
Gonzalez, C., Almaraz, L., Obeso, A., Rigual, R., 1994. Carotid body chemorecep-tors: from natural stimuli to sensory discharges. Physiological Reviews 74,829–898.
Guner, I., Yelmen, N., Sahin, G., Oruc, T., 2002. The effect of intracerebroventricu-lar dopamine administration on the respiratory response to hypoxia. TohokuJournal of Experimental Medicine 196, 219–230.
Guivarc’h, D., Vernier, P., Vincent, J.D., 1995. Sex steroid hormones change the dif-ferential distribution of the isoforms of the D2 dopamine receptor messengerRNA in the rat brain. Neuroscience 69, 159–166.
Jedrusiak, J., Brus, R., Kostrzewa, R.M., 1995. Dopaminergic neuronal systemsmodulate the central cardiovascular effects of TRH in rats. Polish Journal ofPharmacology 47, 43–52.
Jha, A., Sharma, S.K., Tandon, N., Lakshmy, R., Kadhiravan, T., Handa, K.K., Gupta, R.,Pandey, R.M., Chaturvedi, P.K., 2006. Thyroxine replacement therapy reversessleep-disordered breathing in patients with primary hypothyroidism. SleepMedicine 7, 55–61.
Joseph, V., Bairam, A., 2004. Differential regulation of short and long dopamine D2
receptor mRNA levels by hypoxia in the adrenals of 1-day-old and adult rabbits?Brain Research – Molecular Brain Research 130 (1–2), 115–123.
Kline, D.D., Takacs, K.N., Ficker, E., Kunze, D.L., 2002. Dopamine modulates synaptictransmission in the nucleus of the solitary tract. Journal of Neurophysiology 88,2736–2744.
Lane, D.A., Chan, J., Fitzgerald, M.L., Kearn, C.S., Mackie, K., Pickel, V.M., 2012. Quinpi-role elicits differential in vivo changes in the pre- and postsynaptic distributionsof dopamine D2 receptors in mouse striatum: relation to cannabinoid-1 (CB1)receptor targeting. Psychopharmacology (Berlin) 22, 1101–1113.
Li, Y., Roy, B.D., Wang, W., Zhang, L., Sampson, S.B., Yang, Y., Lin, D.-T., 2012. Identifi-cation of two functionally distinct endosomal recycling pathways for dopamineD2 receptor. Journal of Neuroscience 32, 7178–7190.
McKay, L.C., Evans, K.C., Frackowiak, R.S.J., Corfield, D.R., 2003. Neural corre-lates of voluntary breathing in humans. Journal of Applied Physiology 95,1170–1178.
Missale, C., Nash, S.R., Robinson, S.W., Jaber, M., Caron, M.G., 1998. Dopamine recep-tors: from structure to function. Physiological Reviews 78, 189–225.
Morin, L.P., Wood, R.I., 2001. A Stereotaxic Atlas of the Golden Hamster. AcademicPress, San Deigo.
Oh-Nishi, A., Saji, M., Furudate, S.I., Suzuki, N., 2005. Dopamine D2-like receptorfunction is converted from excitatory to inhibitory by thyroxine in the develop-
mental hippocampus. Journal of Neuroendocrinology 17, 836–845.
Olea, E., Gonzalez, C., Gallego, R., Geijo-Barrientos, E., Almaraz, L., 2007. PTU-induced hypothyroidism reduces hypoxic ventilatory drive by impairingcarotid body chemsensitivity in adult rats. Acta Physiologica 190 (Suppl. 655),P51.
2 logy &
O
P
R
S
S
S
S
64 C. Sykora et al. / Respiratory Physio
ppenheimer, J.H., Schwartz, H.L., Surks, M.I., 1972. Propylthiouracil inhibits theconversion of l-thyroxine to l-triiodothyronine. An explanation of the antithy-roxine effect of propylthiouracil and evidence supporting the concept thattriiodothyronine is the active thyroid hormone. Journal of Clinical Investigation51, 2493–2497.
edersen, M.E.F., Dorrington, K.L., Robbins, P.A., 1997. Effects of haloperidol on ven-tilation during isocapnic hypoxia in humans. Journal of Applied Physiology 83,1110–1115.
ugge, B., Balshem, H., Sehgal, R., Relevo, R., Gorman, P., Helfand, M., 2011. Screeningand Treatment of Subclinical Hypothyroidism or Hyperthyroidism. Agency forHealthcare Research and Quality (US) Report No. 11, Rockville (MD). Compara-tive. Effectiveness Reviews 24.
chlenker, E.H., 2007. In hamsters dopamine D2 receptors affect ventilation duringand following intermittent hypoxia. Comparative Biochemistry and PhysiologyPart A: Molecular and Integrative Physiology 148, 674–680.
chlenker, E.H., 2012. Effects of hypothyroidism on the respiratory system and con-trol of breathing human studies and animal models. Respiratory Physiology andNeurobiology 181, 123–131.
sumption but not ventilation in hypothyroid hamsters. Physiology and Behavior56, 635–638.
chlenker, E.H., Schultz, H.D., 2012. Hypothyroidism stimulates D2 receptor-mediated breathing in response to acute hypoxia and alters D2 receptors levels incarotid bodies and brain. Respiratory Physiology and Neurobiology 180, 69–78.
Neurobiology 185 (2013) 257– 264
Saita, E., Tohei, A., Jin, W.Z., Takahashi, S., Suzuli, A.K., Wantanabe, G., Taya, K.,2005. Effects of hypothyroidism on gonadal function after transition of shortday photoperido in amle golden hamsters (Mesocricetus, auratus). Journal ofReproduction and Development 51, 221–228.
Simsek, G., Karaturan, Y., Guner, I., Sahin, G., Oruc, T., Karter, Y., 2004. The role ofperipheral chemoreceptor activity on the respiratory responses to hypoxia andhypercapnia in anaesthetised rabbits with induced hypothyroidism. ChineseJournal of Physiology 47, 153–160.
Thibault, D., Albert, P.R., Pineyro, G., Trudeau, L.É., 2011. Neurotensin trigg-ers dopamine D2 receptor desensitization through a protein kinase C andbeta-arrestin1-dependent mechanism. Journal of Biological Chemistry 286,9174–9184.
Tirotta, E., Fontaine, V., Picetti, R., Lombardi, M., Samad, T.A., Oulad-Abdelghani,M., Edwards, R., Borrelli, E., 2008. Signaling by dopamine regulates D2 receptortrafficking at the membrane. Cell Cycle 7, 2241–2248.
Vaughan, M.K., Powanda, M.C., Richardson, B.A., King, T.S., Johnson Reiter, R.J., 1982.Chronic exposure to short photoperiod inhibits free thyroxine index and plasmalevels of TSH, T4 and triiodothyronine (T3) and cholesterol in female Syrian ham-sters. Comparative Biochemistry and Physiology Part B: Comparative Physiology
71, 615–618.
Xiao, M.-F., Xu, J.-C., Tereshchenko, Y., Novak, D., Schachner, M., Kleene, R., 2009.Neural cell adhesion molecule modulates dopaminergic signaling and behaviorby regulating dopamine D2 receptor internalization. Journal of Neuroscience 29,14752–14763.