108
Ahmed Group Lecture 13 Molecular Signaling Lecgture 14
| Date post: | 27-Dec-2015 |
| Category: |
Documents |
| Upload: | arron-little |
| View: | 213 times |
| Download: | 0 times |

Ahmed GroupLecture 13
Molecular Signaling
Lecgture 14

Ahmed GroupLecture 13
Signal Transduction Pathways
Pathways of molecular interactions that provide communication between the
cell membrane and intracellular endpoints, leading to some
change in the cell

Ahmed GroupLecture 13
Goal of signaling
pathways is to ensure an appropriate
reaction to the type and
strength of an extracellular
stimulus
Principles of information transmission
processingprocessing
Environmental inputEnvironmental input
reactionreaction
growthgrowth
proliferationproliferation
death death
necrosis necrosis
apoptosis apoptosis
movement movement EFFECTOREFFECTORFUNCTIONSFUNCTIONS
Generate new informationGenerate new information•neurotransmissionneurotransmission•messengers messengers

Ahmed GroupLecture 13
Why do we need biosignaling pathways ?
functional integration of distant organs, tissues and cells requires communication;
Signaling is perhaps a primal requirement to respond to our environment;
The foundation of any complex response pathway lies with cellular biochemicals.

Ahmed GroupLecture 13

Ahmed GroupLecture 13
Major themes in ST
• The “internal complexity” of each interaction
• The combinatorial nature of each component molecule (may receive and send multiple signals)
• The integration of pathways and networks

Ahmed GroupLecture 13
1. Post-translational 1. Post-translational modifications (PTM)modifications (PTM)
•Reversible addition of a small Reversible addition of a small chemical group causes change chemical group causes change in activity or location of a signaling in activity or location of a signaling proteinprotein
•PTMs require the action of both PTMs require the action of both modifying and un-modifying modifying and un-modifying enzymes (allowing the signal to be enzymes (allowing the signal to be given given andand terminated) terminated)
Cellular tools for information transmission
MODIFICATIONMODIFICATION TARGETTARGET ADDITIONADDITION REMOVALREMOVAL ROLESROLES
PHOSPHORYLATIONPHOSPHORYLATIONPOPO44
--
TyrTyrSerSerThrThr
kinasekinase phosphatasephosphatase Activity switchActivity switchTargetingTargeting
SS-NITROSYLATION-NITROSYLATIONNONO
TyrTyrCysCys
SS-nitrosylase-nitrosylase Nitro-Nitro-hydrolasehydrolaseReductionReduction
Activity switchActivity switchTargeting?Targeting?
GLYCOSYLATIONGLYCOSYLATION N- or O-N- or O-eg. Asneg. Asn manymany manymany
ProcessingProcessingTargetingTargeting
LIPIDLIPIDeg. Farnesyleg. Farnesyl PrenylPrenyl Geranyl Geranyl MyristoylMyristoyl etcetc
variousvarious manymany manymanyProcessingProcessingTargetingTargetingActivityActivity
ACETYLATIONACETYLATION lysinelysineAcetylationAcetylationcomplexescomplexes
De-acetylationDe-acetylationcomplexescomplexes
Large scale Large scale conformational conformational changeschanges

Ahmed GroupLecture 13
Regulating proteins
How much protein is created?
Transcription,
splicing, degradation
, translation
Change in conformation by ligand binding. Only bound protein can bind DNA
Change in conformation by protein
phosphorylation. Only phospho-
protein can bind DNA
Only dimer complex of two proteins can bind DNA
Binding site is revealed only after
removal of an inhibitor
In order to bind DNA, the protein must first be
translocated to the nucleus

Ahmed GroupLecture 13
Cellular tools for information transmission
Signaling networks are composed of
highlyspecialized
proteins with devolved
functions
•Receptors•Transducers•Adapters•Scaffolds•Effectors

Ahmed GroupLecture 13
Cellular tools for information transmission
•ReceptorsReceptors receive information in form of a ligandreceive information in form of a ligandtransmembrane signalingtransmembrane signaling
•TransducersTransducerspass informationpass informationenzymatically activeenzymatically activemay be signal integratorsmay be signal integrators
•AdaptersAdapters no catalytic activityno catalytic activitymodulate proximity of transducersmodulate proximity of transducers
•ScaffoldsScaffoldsprovide architectureprovide architectureallow energetically unfavorable eventsallow energetically unfavorable events
•EffectorsEffectorsperform an end functionperform an end function
Signaling events are ordered both spatially and temporally
Signaling events are ordered both spatially and temporally

Ahmed GroupLecture 13
Integration of Signals
The signals from several different sources may be integrated though a single shared protein (A) or
protein complex (B)

Ahmed GroupLecture 13
Insulation by complex formation
• The same signaling molecule may participate in more than one pathway
• In such cases, it is sometimes insulated from some of its potential inputs and outputs and sequestered (with specific up- and downstream counterparts) by a specific scaffold molecule

Ahmed GroupLecture 13
Amplification
1 receptor activates multiple G proteins
Each enzyme Y produces many
second messangers, each
messanger activates 1 enzyme Y
1ligand-receptor
500 G-protein
500enzymes
105
(2nd messanger)
250(ion channels)
105-107
(ions)

Ahmed GroupLecture 13
Intracellular target
• Determining the “end” of a signaling pathway is often difficult
• For example, after transcription, a phosphatase may be synthesized that dephosphorylates one of the enzymes in the pathway
• One approach is to consider an event that is “biochemically different” (e.g. transcription, metabolism) as the intracellular target

Ahmed GroupLecture 13
Intracellular Endpoint
• Three major molecular targets– Regulation of gene expression (e.g. activate a transcription factor and translocate it to the nucleus)
– Changes in the cytoskeleton (e.g. induce movement or reorganization of cell structure)
– Affect metabolic pathways
• Many critical processes can occur in response to external signals, without any new synthesis of RNA or proteins. The most well known one is “cell suicide”, termed apoptosis

Ahmed GroupLecture 13
Change in the cell
• An animal cell depends on multiple extracellular signals
• Multiple signals are required to survive, additional to divide and still others to differentiate
• When deprived of appropriate signals most cells undergo apoptosis
DIFFERENTIATE
F G

Ahmed GroupLecture 13
Change in the cell
• The same signal molecule can induce different responses in different target cells, which express different receptors or signaling molecules
• For example, the neurotransmitter acetylcholine induces contraction in skeletal muscle cells, relaxation in heart muscle cells and secretion in salivary gland cells

Ahmed GroupLecture 13
Two Views of Signaling
• The biochemical view: What are the specific biochemical events that mediate signals?
• The logical view: Is a signal activatory or inhibitory?

Ahmed GroupLecture 13
Molecular SignalingReceptor/Ligand InteractionPhosphorylation/dephosphorylation reactionTranscriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
The LIGAND is the signaling molecule (e.g., hormone, pheromone, ion, neurotransmitter, drug).
The ligand binds to or “fits,” a site on a RECEPTOR molecule on the target cell.

Ahmed GroupLecture 13
Requirement of BiosignalingRequirement of Biosignaling
requires a receptor to detect signals;
the receptor must link to or generate an intracellular response;
Such linking molecules are known as “second messengers”;
This transduction system must meet four specific criteria.

Ahmed GroupLecture 13
Criterion 1: specificityCriterion 1: specificity
High specificity only the target cell is influenced; Receptor binding site ligand (signal molecule) complementary and non-covalent interaction follows the law of mass action

Ahmed GroupLecture 13
Criterion 2: amplificationCriterion 2: amplification
often short-lived& low concentration
A single receptor binding event may elicit responses inA single receptor binding event may elicit responses in multiple enzymemultiple enzyme

Ahmed GroupLecture 13
Criterion 3: DesensitizationCriterion 3: Desensitization
feedbackcontrol
the aim of biosignaling is to produce a rapid and majorthe aim of biosignaling is to produce a rapid and major cellular response to a transient signal.cellular response to a transient signal.

Ahmed GroupLecture 13
Criterion 4: IntegrationCriterion 4: Integration
cells frequently receive multiple signals; there are manycells frequently receive multiple signals; there are many reciprocal pathways within cells.reciprocal pathways within cells.

Ahmed GroupLecture 13
Signal source• A signaling cell produces a particular
particular type of signal molecule• This is detected in another target cell, by
means of a receptor protein, which recognizes and responds specifically to its ligand
• We distinguish between Endocrine, paracrine and autocrine signaling. The latter often occurs in a population of homogenous cells.
• Each cell responds to a limited set of signals, and in a specific way

Ahmed GroupLecture 13
Signaling Molecule
• The signal molecule is often secreted from the signaling cell to the extracellular space
• In some cases the signaling molecule is bound to the cell surface of the signaling cell. Sometimes, a signal in both cells will be initiated by such an event.

Ahmed GroupLecture 13
Receptors
• Cell surface receptors detect hydrophilic ligands that do not enter the cell
• Alternatively, a small hydrophobic ligand (e.g. steroids) may cross the membrane, and bind to an intracellular receptor
• Cells may also be linked through a gap junction, sharing small intracellular signaling molecules
GAP JUNCTIONS

Ahmed GroupLecture 13
Cell Surface Receptors• Ion channel linked:
Binding of ligand causes channel to open or close
• G-protein linked:Binding of ligand activates a G-protein which will activate a separate enzyme or ion channel
• Enzyme linked receptor: Binding of ligand activates an enzyme domain on the receptor itself or on an associated molecule

Ahmed GroupLecture 13
Receptor-Ligand Binding
• A dimeric ligand protein is formed by di-sulfide bonds between two identical protein monomers
• The ligand has two identical receptor binding sites and can cross link two adjacent receptors upon their binding
• This initiates the intracellular signaling process• We assume that ligand-receptor binding is irreversible
Ligand Receptor-Ligand complex

Ahmed GroupLecture 13
Receptor Activation
• The cytoplasmic domain of the receptor has intrinsic kinase activity
• Upon dimerization each receptor cross phosphorylates a specific tyrosine residue on its counterpart, which fully activates its kinase
• Then, each kinase autophosphorylates additional tyrosine residues on it own cytoplasmic part

Ahmed GroupLecture 13
Receptor Activation
• The cytoplasmic domain of the receptor has intrinsic kinase activity
• Upon dimerization each receptor cross phosphorylates a specific tyrosine residue on its counterpart, which fully activates its kinase
• Then, each kinase autophosphorylates additional tyrosine residues on it own cytoplasmic part

Ahmed GroupLecture 13
The activated receptor
• The phosphorylated tyrosines can be specifically identified by SH2 and SH3 domains on other proteins, including adapter proteins
• The activated receptor can then phosphorylate these bound proteins

Ahmed GroupLecture 13
Ion Channel Linked cell surface receptor:Ion Channel Linked cell surface receptor:Three stages of acetylcholine receptorThree stages of acetylcholine receptor

Ahmed GroupLecture 13
Receptor/G-proteinReceptor/G-proteinSystems of cell surface receptorsSystems of cell surface receptors
• a complex system: A receptor linked to trimeric GTP binding protein;
• Binding of the ligand produces a conformational change that causes the G-protein to leave the receptor and “dock” with a membrane bound enzyme.
• Activity of the enzyme initiates a cascade of events• Example: adenylate cyclase
phopholipase C

Ahmed GroupLecture 13
The Nobel Prize in Physiology and Medicine 1994
Alfred G. Gilman Martin Rodbell USA USA
1941- 1925-1998
"for their discovery of G-proteins and the role of these proteins in signal transduction in cells"

Ahmed GroupLecture 13
An example ofG-protein linked
Cell surfaceReceptor.

Ahmed GroupLecture 13
Enzyme Linked cell Enzyme Linked cell surface receptorssurface receptors
(a) tyrosine kinases
insulin receptor: prototype for this signaling pathway.
the receptor complex:extracellular ligand binding site cytosolic catalytic domain;
The enzyme is a tyrosine kinase, which phosphorylate tyrosine residues in specific target proteins.

Ahmed GroupLecture 13
IRS: insulin receptor substrate
protein kinase : Raf-1, MEK, MAPK (phosphorylate Ser or Tyr residue) MAKP (ERK, extracellular regulated kinase): mitogen - activated protein kinase; MEK: mitogen-activated, ERK-activating kinase

Ahmed GroupLecture 13
Activation of glycogen synthase by insulin

Ahmed GroupLecture 13
(b) guanylyl cyclases
produces cGMP (guanosine 3’5’-cyclic monophosphate ) from GTP, which serve as a second messenger;
most cGMP effects are mediated via a cGMP-dependentprotein kinase;
cGMP levels are restored to normal by a phosphodiesterase that produces GMP.

Ahmed GroupLecture 13
Intracellular receptors• Small hydrophobic signaling
molecules, such as steroids, can cross the cell membrane (e.g. estrogen, vitamin D, thyroid hormone, retinoic acid) and bind to intracellular receptors
• The hormone-receptor complex has an exposed DNA binding site and can activate transcription directly (or, more typically as a homo- or hetero-dimer)
• This usually initiates a cascade of transcription events
PRIMARY RESPONSE
SECONDARY RESPONSE
Shut off primary
response genes
Turn on secondary
response genes

Ahmed GroupLecture 13
Regulation of transcription by steroid hormones
steroid steroid receptorsreceptors

Ahmed GroupLecture 13
•Steroid, retinoic acid and thyroid hormones use a signalling process (by-passes the plasma membrane);• the hormones bind to soluble receptors (i.e. not membrane bound) with high affinity/specificity; •the binding energy induces conformational change that result in homo- or heterdimerisation with other receptors;•the oligomerized and liganded receptors bind to regulatory regions of DNA known as hormone response elements (HREs);•receptor interaction at HREs causes altered rates of gene transcription and subsequently protein levels and cellular effects.

Ahmed GroupLecture 13
Tamoxifen antagonist of estrogen, to treat hormone-dependent breast cancer
competes with estrogen for binding to the estrogen receptor;
tamoxifen-receptor complex has no effect on gene expression.
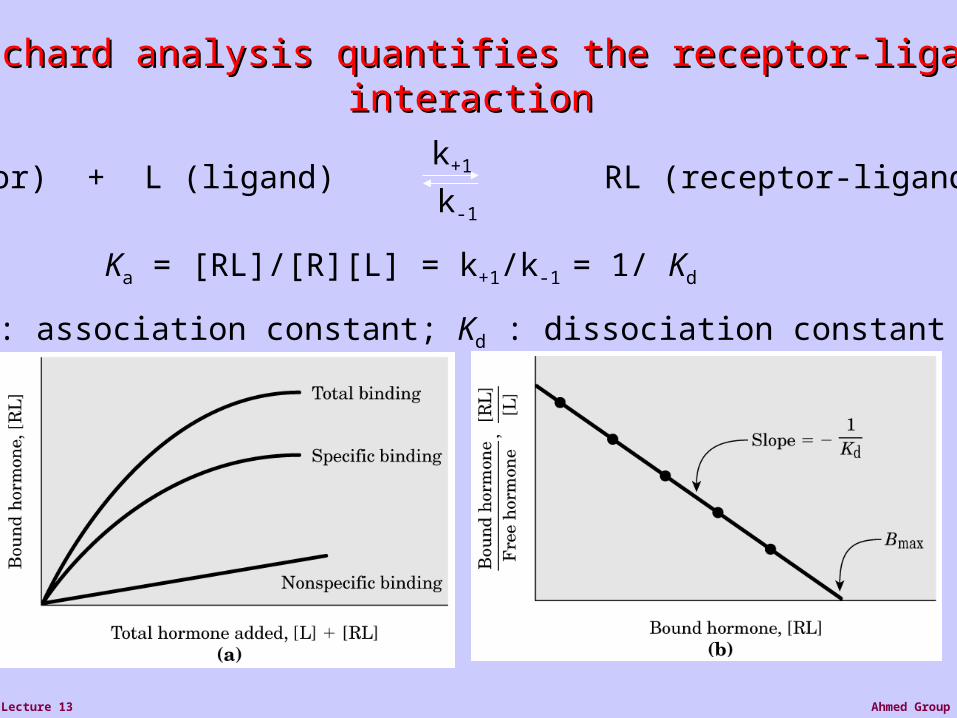
Ahmed GroupLecture 13
Scatchard analysis quantifies the receptor-ligand Scatchard analysis quantifies the receptor-ligand interactioninteraction
R (receptor) + L (ligand) RL (receptor-ligand complex)k+1
k-1
Ka = [RL]/[R][L] = k+1/k-1 = 1/ Kd
Ka : association constant; Kd : dissociation constant

Ahmed GroupLecture 13
EGF-signaling network
StatStat
PI3KPLC
UV, ECM, polarization
Raf1
MEK1,2
ERK1,2
EGF
EGFR
EGFR
EGF
Shc
SosRas
Grb-2Grb-2
Fos Jun
MycElk1
proEGF
EGF
GPCRthrombin
GHRGH
JAK2
Src
Nck
PAK1
MP1ERK1
MEK1

Ahmed GroupLecture 13
Molecular Signaling
Receptor/Ligand InteractionPhosphorylation/dephosphorylation reactionTranscriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
Protein P-Protein
ATP ADP
Protein kinase
Protein phosphatase
P
Protein phosphorylation/dephosphorylation provides a major mechanism for signal
transduction

Ahmed GroupLecture 13
Kinases (and phosphatases)Kinases (and phosphatases)1.1. Bind and orient ATPBind and orient ATP2.2. Bind and orient substrateBind and orient substrate3.3. Catalyze phosphate transferCatalyze phosphate transfer
Why is phosphate such a good information carrier?Why is phosphate such a good information carrier?
1.1. High bond energiesHigh bond energies2.2. Labile if unattachedLabile if unattached3.3. Linked to metabolic statusLinked to metabolic status200kDa
97kDa
68kDa
43kDa
restactive
pTyrproteins
pTyrSignaling complexes

Ahmed GroupLecture 13
A total of 518 protein kinases including: • 478 conventional protein kinases (ePKs) (16 have tandem catalytic domains) • 388 protein-serine/threonine kinases • 90 protein-tyrosine kinases
58 receptor protein-tyrosine kinases 32 non-receptor protein-tyrosine kinases
(~50 may lack catalytic activity; ~106 pseudogenes) • 40 atypical protein kinases (e.g. EF2K/alpha kinases)
Analysis by G. Manning, D. Whyte, R. Martinez, S. Sudarsanam (Sugen) (http://www.kinase.com)
The human kinome (protein kinases)

Ahmed GroupLecture 13
A total of ~140 protein phosphatases including:
• 38 protein-tyrosine phosphatases
• 38 serine/threonine phosphatases (18 PP1/2A; 20 PP2C)
• 62 Dual Specificity Phosphatases (e.g. MKPs, PTEN)
Analysis by G. Manning, D. Whyte, R. Martinez, S. Sudarsanam (Sugen) (http://www.kinase.com)
~2.5% genes directlydevoted to proteinphosphorylation anddephosphorylation
The human phosphatome (protein phosphatases)

Ahmed GroupLecture 13
PP1 PP2A
PP2B
Receptor-like
Ser or Thr
Tyr
Non-receptor
PPP PTP
PPM
Classification of Protein Phosphatases
Novel
PP2C
PHP
His
FCP
DSP
Dual specificity

Ahmed GroupLecture 13
OldInhibitor Activator
New
Type Name Type Name
1
2
2
2
PP1
PP2A
PP2B
PP2C
inhibitor-1&-2
okadaic acid
trifluoperazine
EDTA
-
-
Ca-calmodulin
PPP
PPP
PPP
PPM
PPP1
PPP2
PPP3
PPM2+, Mn2+ Mg
Classical Ser/Thr Protein Phosphatases

Ahmed GroupLecture 13
Seven Subfamilies of Tyrosine Kinase Receptors

Ahmed GroupLecture 13
Ras Activation
• By these protein-protein interactions, the SOS protein is brought close to the membrane, where is can activate Ras, that is attached to the membrane
• SOS activates Ras by exchanging Ras’s GDP with GTP.
• GAP inactivates it by the reverse reaction
SOS

Ahmed GroupLecture 13
Activation of the MAPK cascade
• Active Ras interacts with the first kinase in the MAPK cancade, Raf.
• It localizes Raf to the membrane, where it is activated by an unknown mechanism
• This starts the cascade

Ahmed GroupLecture 13
Activation of the MAPK cascade
• Each kinase in the cascade is activated by phosphorylation in a regulatory site, called the t-loop
• When T-loop is phosphorylated, a conformation change occurs and the catalytic cleft is “opened” and active
• Each kinase is bound by modifying enzymes (incoming signals) on its Nt lobe. It binds its substrate through its Ct lobe.
• The three kinases may be tethered together in one complex with the MP1 scaffold protein

Ahmed GroupLecture 13
MAPK (ERK1)
Binding MP1 molecules
Kinase site: Phosphorylate Ser/Thr
residues (PXT/SP motifs)
Regulatory T-loop: Change conformation
ATP binding site: Bind ATP, and use it for
phsophorylation
Binding to
substrates
Structure Process
COOH
Nt lobe
Catalytic coreCt lobe
NH2
p-Y
p-T

Ahmed GroupLecture 13
MAPK targets
• The MAPK phosphorylates and activates many different targets
• For example, after phosphorylation it may translocate to the nucleus and activate transcription factors
• It also phosphorylates the receptor kinase and other enzymes in the pathway in an inhibitory fashion (negative feedback)

Ahmed GroupLecture 13
The RTK-MAPK pathway
This is only one path in mammalian mitogenic signaling initiated from an RTK. In fact, additional signals are intiated at the RTK. Similar pathways were found in eukaryotic organisms as diverse as
yeast, drosophila, mouse and humans
RTK receptor
Adaptor proteins
Ras Activati
on
MAPK cascade
ERK1RAF
GRB2
RTK
RTK
SHC
SOS
RAS
GAP
PP2A
MKK1
GF GF
MP1
MKP1
IEG
IEP
IEP
J F

Ahmed GroupLecture 13
Molecular Signaling
Receptor/Ligand InteractionPhosphorylation/dephosphorylation reactionRadiation-induced Transcriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
Examples of Redox Regulated Mammalian Transcription Factors
• AP-1– Ref-1 & Thioredoxin
• Egr1– Zinc fingers, most common motif in the human proteome
• HIF-1/ ARNT– O2
– Fe+2
-ketoglutarate – Ascorbate
• PAS (Per/Arnt/Sim) Domain Proteins (NADPH & NADH sensitive)• NFκB

Ahmed GroupLecture 13
AP-1 (activator protein-1) activity is controlled by reversible cysteine oxidation
Evans, AR, et al., Mutat. Res. 461, 83-108, 2000

Ahmed GroupLecture 13
Zinc Fingers are a common redox sensitive DNA binding motif
Alberts et al., Molecular Biology of the Cell, 4th Edition

Ahmed GroupLecture 13
HIF-1 is Post-Translationally Regulated
O2, Fe+2, -KG, Asc

Ahmed GroupLecture 13
Wang GL, et al., Proc Natl Acad Sci 92(12): 5510, 1995
HIF-1 is O2 sensitive

Ahmed GroupLecture 13
OH• OH•
SpeciesReactive Oxygen
PTEN
Ionizing Radiation
p53
MDM2
p53
Anti-apoptosis
p53
MDM2
RbHYPO-PO 4
MDM2
RbHYPO-PO4
MDM2
RbHYPO-PO4
p53p53
transactivation
Bax
Apoptosis
EGR-1

Ahmed GroupLecture 13
Pro-survival NF-B Pathway isInduced by ROS

Ahmed GroupLecture 13
Molecular Signaling
Receptor/Ligand InteractionPhosphorylation/dephosphorylation reactionRadiation-induced Transcriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
Yellow: equally expressedRed: more expressed in tumor tissue (“over expressed”)Green: more expression in normal tissue (“under expressed”)
Tools for Gene Expression Analysis1. Northern blotting. 2. RT-PCR 3. Microarray technology

Ahmed GroupLecture 13
299213
2Gy191
0.1 Gy
Total gene set contains nearly 10,000 genes
703 Genes with Significant F-ratio
DIFFERENCES IN TRANSCRIPTION PROFILESBETWEEN LOW AND HIGH DOSE IRRADIATION IN
MURINE BRAIN CELLS
Up-regulated at 2Gy 245
Down-regulated at 2Gy 135
Up-regulated at 0.1Gy 182 Down-regulated at 0.1Gy 187
Yin 2003
Numbers of Genes
in HLB Cells 4 hr after IRDifferentially Regulated

Ahmed GroupLecture 13
Radiation-induced changes in gene
expression
Dose (cGy)
0 10 100 1000
Low Dose Genes
High Dose Genes
Wyrobek

Ahmed GroupLecture 13
UT IR IR + α-TGFβ
Hierarchical Clustering

Ahmed GroupLecture 13
What is proteomics?

Ahmed GroupLecture 13
Metabolic labeling and chemical proteomics strategy for the post-translational O-GlcNAc modification of nucleocytoplasmic proteins. Cells are treated with Ac4GlcNAz, which diffuses into the cytosol where it is deactylated to yield GlcNAz. GlcNAz is an analogue of the naturally occurring saccharide GlcNAc, differing only in the presence of the azide moiety on the acetamido group. This analogue is tolerated by the hexosamine biosynthetic pathway with the result that cellular proteins become modified with GlcNAz. The cell lysate is then subjected to the highly chemoselective Staudinger ligation and the bioorthogonal azide moiety reacts with a triarylphosphine probe to provide a stable amide linkage. Subsequent detection and identification of O-GlcNAz modified proteins can then be carried out using mass spectrometry or Western blot analysis.

Ahmed GroupLecture 13
High-thoroughput analysis of radiation-induced apoptosis through quantitative proteomic profiling using comparative amino acid-coded mass tagging
Investigators: Mansoor M. Ahmed, Weis Center for Research, Geisinger ClinicAlan Pollack, Fox Chase Cancer Center
ObjectiveTo identify the differential regulation of protein expression in normal prostate versus prostate tumor after ionizing radiation treatment using mass spectrometry coupled with Amino Acid-Coded mass tagging (AACT).
ApproachThe normal prostate cell line RWPE-1 and prostate tumor cell line LNCaP will be used in this study. Both cell line will be maintained in normal media as well as in AACT-nutrient media containing the deuterated form of various amino acids such as leucine, lysine, methionine and tyrosine. This culture approach will result in proteins synthesized having a slightly higher mass as opposed to proteins that are expressed in normal media. The cells in the AACT–media will then be exposed to IR dose of 5 Gy. After 24 hours of incubation, the cells will be lysed to obtain cytosolic, membrane and nuclear fraction of the proteins from each group.Each of the protein fraction of the irradiated and untreated group of the same cell line will then be mixed in a ratio of 1:1, and further separated by SDS-PAGE. The stained protein bands will be excised continuously with a 1-2 mm step from 10kDa to the loading well. The excised bands will be subjugated to trypsin digestion and peptide extraction followed by MLC connected directly to electrospray ionization source on a mass spectrometer. Since the elution time of the labeled and unlabelled proteins is essentially identical in the C18 reverse phase column, both isotopic forms of peptide would elute together at corresponding retention time but would differ in their mass-charge (m/z) ratios. The relative intensities of the two isotopic peaks for labeled and unlabeled peptides hence obtained would correspond to differential expression of protein synthesized in the normal and treated population. The identity of each protein will be detected by MS/MS experiment for each peptide, that will be concurrently performed to yield peptide sequence information for protein identification by MASCOT.Several low-abundance peptides can be identified by the single-peptide-based AACT-MALDI approach but not LC-MS/MS approach. This is due to the difference in the ionization mechanism, hence MALDI is a powerful alternative and complementary approach. We propose to study the protein profiling by both the methods. The proteins showing differential expression would then be verified using conventional Western and Immuno-blot analysis.

Ahmed GroupLecture 13
Molecular Signaling
Receptor/Ligand InteractionPhosphorylation/dephosphorylation reactionRadiation-induced Transcriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
Examples of DNA lesions induced by Radiation

Ahmed GroupLecture 13
Base changeBase change
U
Examples of DNA lesions induced by Radiation

Ahmed GroupLecture 13
Dimer formationDimer
formation
Examples of DNA lesions induced by Radiation

Ahmed GroupLecture 13
Interstrand cross link
Examples of DNA lesions induced by Radiation

Ahmed GroupLecture 13
Most severe type of damage
Double strand break
Examples of DNA lesions induced by Radiation

Ahmed GroupLecture 13
Radiation induced DNA lesions cause cell death &
transformation
Decreased cell survivalDecreased cell survival
Surviving cells transformed and become cancer cellsSurviving cells transformed and become cancer cells

Ahmed GroupLecture 13
Possible Biomarkers and Biological Dosimeters for Human Radiation
Exposure
• TRAIL receptor 2• FHL2• cyclin G• cyclin protein gene• -H2AX

Ahmed GroupLecture 13
Antibody-labeled histone -H2AX phosphorylated foci in human cells following 0.6Gy irradiation
EP Rogakou et al, J Cell Biol, 146:905-915, 1999

Ahmed GroupLecture 13
Antibody-labeled histone -H2AX phosphorylated foci in human mitoticchromosomesfollowing 0.6Gyirradiation
EP Rogakou et al, J Cell Biol, 146:905-915, 1999

Ahmed GroupLecture 13
Untargeted Effects of Exposure Untargeted Effects of Exposure to Ionizing Radiationto Ionizing Radiation
Effects in unexposed cells and their progenyi.e. in cells not directly hit.
Genomic instabilityGenomic instability Bystander EffectsBystander Effects

Ahmed GroupLecture 13
Radiation-induced Genomic Radiation-induced Genomic InstabilityInstability
micronucleusmicronucleuschromosomechromosome aberrationaberration
cell deathcell death gene gene mutationmutation
mitotic failureaneuploidy

Ahmed GroupLecture 13
Bystander Effects of Ionizing Bystander Effects of Ionizing RadiationRadiation
Signals via gap junctions
Signals via medium/plasma
N.B. 1950s and 60s Reports of clastogenic factors in blood of exposed individuals
1990s: Effects in more cells than
irradiated by -particles Cytotoxic factor(s) after low dose
low LET exposure

Ahmed GroupLecture 13
Checkpoints integrate repair of chromosome damage with events of cell cycle
• G1-S checkpoint– p53 – transcription factor that induces expression of DNA repair genes and CDK inhibitor p21
– p53 pathway activated by ionizing radiation or UV light (causing DNA damage) during G1 phase delays entry into S phase
– DNA is repaired before cell cycle continues
– If DNA is badly damaged cells commit suicide (programmed cell death or apoptosis)

Ahmed GroupLecture 13
Molecular Signaling
Receptor/Ligand InteractionPhosphorylation/dephosphorylation reactionRadiation-induced Transcriptional activationRadiation-induced gene expression: Gene expression profiling andProteomicsRadiation-induced signals: DNA damage response and non-DNADamage responsesCell survival and death pathways

Ahmed GroupLecture 13
G1-S phase transition

Ahmed GroupLecture 13
TGF-Smad SignalingPathway is induced By radiation
TGFR: receptor serine/ threonine kinaseSmad: transcription factorTFE: transcription factor

Ahmed GroupLecture 13
Pro-survival NF-B Pathway isInduced by radiation

Ahmed GroupLecture 13
Cell survival and proliferation are highly regulated.
Cell division modulated by cell cycle
Apoptosis eliminates damaged cells
Mutations in cancer cells allow cells to escape apoptosis and proliferation controls

Ahmed GroupLecture 13

Ahmed GroupLecture 13
ATM ATM
ATMP
ATM
CHK2
PATM Nbs1 P
ATMPBrca1
P
P
ATMP
p53P
Bax Cell Death
p21 waf1/cip1G1 Arrest
DNA Repair
Ionizing radiation Chromatin
changes
ReactiveOxygenSpecies
EGR-1
Ras AKT/PI3-K
NFB
Bcl-2
TNF-
MDR1
Chemo-Resistance
SurvivalProliferation
Caspaseactivation
Autophosphorylation
Substratephosphorylation
PFocus Formation
Radiation induced signaling response: wild type p53 background

Ahmed GroupLecture 13
ATM ATM
ATMP
ATMP
Bax
p21 waf1/cip1
DNA Repair
Ionizing radiation Chromatin
changes
ReactiveOxygenSpecies
Mutant p53
Autophosphorylation
Substratephosphorylation
Ras AKT/PI3-K
NFB
Bcl-2
TNF-
MDR1
Chemo-Resistance
SurvivalProliferation
Induced RadiationResistance
Focus Formation
Radiation induced signaling response: mutant p53 background
ATM Nbs1 P
ATMPBrca1
P
P

Ahmed GroupLecture 13
Grb2
SOSRas
Raf
MEK1/2
ERK1/2
ERK1/2
ERK1/2
ERK1/2
p90rsk
p90rsk
mTOR
AKT/pKB
CREB
c-FosSRF
c-Jun
STAT1/3ELK-1 Ets
PI3K
4E-BP1 p70S6K
PKC
TPAPlasma Membrane
Cytoplasm
Nucleus
Growth factor
Receptor (ex.: EGFR)Growth Factor
(ex.: EGF)
Growthdifferentiation
Survival
MAPK
pp
NFB
NFB
Other cell survival pathways that may affect radiation sensitivity
Ionizing
radiation

Ahmed GroupLecture 13
Death Signals
DNA damage (ionizing radiation)stress
heat shockoxidative stress (hypoxia, NO)
nutrient deprivationInterferon
protein synthesis inhibitorsTNF
CD-95 (Fas or Apo-1) Apo3 ligand (Apo3L, TWEAK)
Apo2L (TRAIL)
NGF

Ahmed GroupLecture 13
Death receptor ligands CD95 ligand (FasL)TNF and lymphotoxin Apo3 ligand (Apo3L, TWEAK)
Apo2L (TRAIL)
NGF
Death Receptors:CD95 (Fas, Apo1)TNFR1Death receptor3 (DR3, Apo3, WSL-1, TRAMP, LARD)DR4 DR5 (Apo2, TRAIL-R2, TRICK 2, KILLER)P75 nerve growth factor receptor

Ahmed GroupLecture 13
CD95 (Fas, Apo1)TNFR1Death receptor3 (DR3, Apo3, WSL-1, TRAMP, LARD)DR4 DR5 (Apo2, TRAIL-R2, TRICK 2, KILLER)P75 nerve growth factor receptor
CAR1 (avian death receptor)

Ahmed GroupLecture 13
Cysteine proteinases, cys at active site.
Activated by cleavage at aspartic acid.
Cleave target proteins at aspartic acid residues

Ahmed GroupLecture 13

Ahmed GroupLecture 13
Adams and Cory, 1998. Science 281:1322-1326. Structure of Bcl-XL with a BH3 peptide bound.

Ahmed GroupLecture 13
From:“The biochemistryof apoptosis”Hengartner, 2000 Nature407:770-776.

Ahmed GroupLecture 13
Turning-off Mechanisms of Signaling
• Receptor sequestration
• Receptor downregulation
• Receptor inactivation by its modification
• Inactivation of signaling proteins
• Production of inhibitory proteins including decoy proteins
• Cross-inhibition of different signaling pathways

















