1 Structure and Function Alan M. Rosenberg, Ross E. Petty Rheumatic diseases involve abnormalities of multiple organs and systems, but it is predominantly the musculoskeletal system that is affected. The musculoskeletal system consists of bones, joints, carti- lage, tendons, ligaments, muscles, and associated connective tissues. This chapter is an overview of normal musculoskeletal biology. In other sections, pathology of the musculoskeletal system and other sys- tems pertinent to rheumatic diseases, including immune, vascular, and neurological systems, are discussed in relevant contexts. THE SKELETON The musculoskeletal system is derived from embryonic mesoderm, a layer that first appears at approximately the fourth week of gestation. As the embryonic neural crest closes to create the notochord, cells from the dorsal margins migrate to form the teeth and some facial bones, cartilage, and connective tissue of the anterior skull. 1 Mesodermal cells arising from the prechordal region (located rostral to the notochord) form cartilage and bone for the posterior skull, paraxial mesodermal cells (somites) generate the axial skeleton, and lateral plate mesoder- mal cells form the appendicular skeleton. 1 As with all large organisms, the human endoskeleton provides a rigid framework, allowing movement to occur with power and pre- cision. The axial skeleton comprises the skull, mandible, auditory ossicles, hyoid bone, vertebral column, sternum, and ribs, and the appendicular skeleton includes the limbs and limb girdles. Ossification of the fibrous membranes and hyaline cartilage that constitute the primordial skeletal scaffold begins at approximately the seventh gesta- tional week, and by birth it is almost, but not entirely, complete. The skeleton is the framework that maintains body shape; sup- ports muscles and connective tissues, neurovascular networks, and skin; protects the body’s vital organs; and makes movement possible through articulations and muscle and tendon attachments. The skel- eton is the repository for hematopoietic and mesenchymal progeni- tor cells, a source of energy derived from yellow marrow adipocytes, a mediator of the immune response, and functions as an endocrine organ contributing to calcium, phosphate, and glucose homeosta- sis. 2-7 Further, the musculoskeletal system aids other bodily functions, including digestion by mastication, respiration by moistening inspired air passing through bone sinuses, and communication by way of audi- tory ossicles for hearing and the larynx for vocalization. The skeleton is a dynamic organ both anatomically and physio- logically. It is modified structurally during growth and maturation, in response to fluctuating mechanical loads, and as a normal physiolog- ical process throughout life. It is continuously generating new blood cells and contributing to the maintenance of the body’s biochemical and energy balance. Bone Bone is a composite of organic and inorganic materials. The hardness of bone is a consequence of the interaction of organic constituents such as collagen, proteoglycans, and matrix proteins, which confer ten- sile strength and permit flexibility to withstand stress, and inorganic materials such as hydroxylapatite and other calcium and phosphate salts, which account for bone’s compressive strength. Classification of Bones Bones are categorized based on either their respective shapes and attendant functions or the process by which they become ossified. Bone classification based on shape and function. All bones share similar ultrastructural characteristics, but gross anatomical shapes differ, reflecting adaptations to their respective functions. Long bones, with a greater length than width, have a thick wall of cortical bone to withstand and adapt to fluctuating stresses of weight-bearing and to allow for rotation and leverage. Short, cuboid-shaped bones, such as carpal bones, support and facilitate movement. Broad flat bones, such as cranial bones, sternum, and ribs, are thin and curvilinear; they serve as attachment sites for musculature and they shield vital organs. Irregular- shaped bones protect the face and spinal cord. Sesamoid bones develop as smooth, round ossifications to counteract compressive forces within tendons; patellae are the only sesamoid bones consistently present in humans, although the hallux sesamoid bone of the foot and the pisiform of the wrist are usually present as well. Bone classification based on ossification process. Bone is formed by either intramembranous or endochondral ossification. Intramembranous ossification occurs in facial, cranial, and clavicular bones as mesenchymal cells within the unossified skeleton coalesce and then differentiate into either capillaries or osteoblasts. The osteoblasts secrete unmineralized osteoid matrix into which mineral salts (notably calcium and phosphorus) are then deposited. When ossified, osteoid produced around capillaries becomes honeycomb-like trabecular bone (also referred to as cancellous or spongy bone), which encroaches on the vasculature, compacting it into red marrow. Endochondral ossification occurs in most appendicular and axial skeleton bones. Endochondral ossification involves the progressive 1 SECTION I Basic Concepts
Transcript
1
Structure and FunctionAlan M. Rosenberg, Ross E. Petty
Rheumatic diseases involve abnormalities of multiple organs and systems, but it is predominantly the musculoskeletal system that is affected. The musculoskeletal system consists of bones, joints, carti-lage, tendons, ligaments, muscles, and associated connective tissues. This chapter is an overview of normal musculoskeletal biology. In other sections, pathology of the musculoskeletal system and other sys-tems pertinent to rheumatic diseases, including immune, vascular, and neurological systems, are discussed in relevant contexts.
THE SKELETONThe musculoskeletal system is derived from embryonic mesoderm, a layer that first appears at approximately the fourth week of gestation. As the embryonic neural crest closes to create the notochord, cells from the dorsal margins migrate to form the teeth and some facial bones, cartilage, and connective tissue of the anterior skull.1 Mesodermal cells arising from the prechordal region (located rostral to the notochord) form cartilage and bone for the posterior skull, paraxial mesodermal cells (somites) generate the axial skeleton, and lateral plate mesoder-mal cells form the appendicular skeleton.1
As with all large organisms, the human endoskeleton provides a rigid framework, allowing movement to occur with power and pre-cision. The axial skeleton comprises the skull, mandible, auditory ossicles, hyoid bone, vertebral column, sternum, and ribs, and the appendicular skeleton includes the limbs and limb girdles. Ossification of the fibrous membranes and hyaline cartilage that constitute the primordial skeletal scaffold begins at approximately the seventh gesta-tional week, and by birth it is almost, but not entirely, complete.
The skeleton is the framework that maintains body shape; sup-ports muscles and connective tissues, neurovascular networks, and skin; protects the body’s vital organs; and makes movement possible through articulations and muscle and tendon attachments. The skel-eton is the repository for hematopoietic and mesenchymal progeni-tor cells, a source of energy derived from yellow marrow adipocytes, a mediator of the immune response, and functions as an endocrine organ contributing to calcium, phosphate, and glucose homeosta-sis.2-7 Further, the musculoskeletal system aids other bodily functions, including digestion by mastication, respiration by moistening inspired air passing through bone sinuses, and communication by way of audi-tory ossicles for hearing and the larynx for vocalization.
The skeleton is a dynamic organ both anatomically and physio-logically. It is modified structurally during growth and maturation, in
response to fluctuating mechanical loads, and as a normal physiolog-ical process throughout life. It is continuously generating new blood cells and contributing to the maintenance of the body’s biochemical and energy balance.
BoneBone is a composite of organic and inorganic materials. The hardness of bone is a consequence of the interaction of organic constituents such as collagen, proteoglycans, and matrix proteins, which confer ten-sile strength and permit flexibility to withstand stress, and inorganic materials such as hydroxylapatite and other calcium and phosphate salts, which account for bone’s compressive strength.
Classification of BonesBones are categorized based on either their respective shapes and attendant functions or the process by which they become ossified.
Bone classification based on shape and function. All bones share similar ultrastructural characteristics, but gross anatomical shapes differ, reflecting adaptations to their respective functions. Long bones, with a greater length than width, have a thick wall of cortical bone to withstand and adapt to fluctuating stresses of weight-bearing and to allow for rotation and leverage. Short, cuboid-shaped bones, such as carpal bones, support and facilitate movement. Broad flat bones, such as cranial bones, sternum, and ribs, are thin and curvilinear; they serve as attachment sites for musculature and they shield vital organs. Irregular-shaped bones protect the face and spinal cord. Sesamoid bones develop as smooth, round ossifications to counteract compressive forces within tendons; patellae are the only sesamoid bones consistently present in humans, although the hallux sesamoid bone of the foot and the pisiform of the wrist are usually present as well.
Bone classification based on ossification process. Bone is formed by either intramembranous or endochondral ossification. Intramembranous ossification occurs in facial, cranial, and clavicular bones as mesenchymal cells within the unossified skeleton coalesce and then differentiate into either capillaries or osteoblasts. The osteoblasts secrete unmineralized osteoid matrix into which mineral salts (notably calcium and phosphorus) are then deposited. When ossified, osteoid produced around capillaries becomes honeycomb-like trabecular bone (also referred to as cancellous or spongy bone), which encroaches on the vasculature, compacting it into red marrow.
Endochondral ossification occurs in most appendicular and axial skeleton bones. Endochondral ossification involves the progressive
1
SECTION I Basic Concepts
2 SECTION I Basic Concepts
replacement of the primordial cartilaginous scaffold with bone through a sequence in which chondrocytes proliferate, mature, and enlarge. By 8 weeks gestational age some mesenchymal cells have differentiated into cartilage-producing chondrocytes that create the cartilaginous skeletal framework. Building the skeletal scaffold begins as mesen-chymal stem cells form condensations at the site of future bone. Cells within the condensations do not differentiate directly into bone-pro-ducing osteoblasts; instead, they become proliferating chondrocytes, which synthesize the extracellular matrix comprising predominantly type II collagen, some type IX and XI collagens, and proteoglycans. Then, as mature, hypertrophied chondrocytes, they secrete a matrix rich in type X collagen, which provides the supporting framework for endochondral ossification.
As the cartilage matrix ossifies, blood flow to the chondrocytes is impeded, the cells die, and adjacent cartilage breaks down, leaving a medullary cavity into which new vessels invade to deliver bone-pro-ducing osteoblasts. While ossification continues from the primary ossification center in the diaphysis, cartilage continues to be produced at the ends of bones as the epiphyses. At birth, cartilage remains at the articular surface and the growth plate (the physis) situated between the epiphysis and the diaphysis. A mature long bone includes the epiphysis at the end of the bone, the diaphysis (the shaft of the bone), and the interposing metaphysis, which represents the region of the previous cartilaginous growth plate. Apophyses, such as the tibial tuberosity, posterior calcaneus, greater and lesser trochanters, and iliac crests, like epiphyses, are the sites of new bone formation but do not contribute to bone length; instead, they generate new bone in response to tendinous or ligamentous traction.
The Morphology and Physiology of BoneBone includes an outer layer of thick, compact, cortical bone and an inner layer of interlacing ossified struts (trabeculae). Alignment of the trabecular network with the long axis of the bone confers maximal tensile strength and compressive tolerance so that less bone mass is required. Bones subjected to predominantly tensile and compressive forces, such as vertebrae, have a thin cortex of compact bone; they exert their support by their shapes and trabecular architecture. Bones that are required to tolerate torsion, bending, and shearing forces, such
as the bones of the leg, tend to be cylindrical with a thick collar of cor-tical bone and a marrow cavity.
Type I collagen constitutes 90% of bone matrix. The triple heli-cal strands of collagen, when densely packed into parallel alignments (lamellae), confer optimal tensile strength to accommodate mechani-cal loading. Other organic constituents of bone contribute to regulat-ing mineralization, growth, and energy metabolism.
The circular osteon is the fundamental physiological unit of most mammalian compact bone (Fig. 1.1). The microstructure of the osteon is characterized by 5 to 20 concentric layers of compact (cortical) bone that encircle a central haversian canal (first identified by 17th-century English physician Clopton Havers), through which neurovascular bun-dles traverse to supply the adjacent osteocytes (Fig. 1.1). Osteons, mea-suring a few millimeters in length and less than 0.25 mm in diameter, tend to orient to the long axis of the bone. Osteons are characteristically present in mature bone that has undergone new bone growth, remodel-ing, or repair. Osteons and their associated intra-haversian blood vessels communicate by interconnecting transverse Volkmann canals (named for 19th-century German physiologist Alfred Volkmann).
Deeper layers of bone are made of spongy bone, the normal physio-logical consequence of bone breakdown induced by osteoclasts.
Bone CellsBone contains four cell types: mesenchymal stem cells, osteoblasts, osteocytes, and osteoclasts. Mesenchymal stem cells, located in the intertrabecular loose connective tissue adjacent to vascular channels and in the periosteum, have the potential to differentiate into cells of bone (osteoblasts, osteocytes, and osteoclasts), cartilage (chondro-cytes), muscle (myocytes), or fat (adipocytes).
The differentiation of mesenchymal stem cells into osteoclasts is regulated by a wide array of cytokines, growth factors, and hormones that, respectively, promote or inhibit osteoclastogenesis (Fig. 1.2).8-11 Groups of osteoblasts, which assemble into clusters to form the osteon, produce the organic constituents of bone, mainly collagen and to a lesser extent osteocalcin and osteopontin. Osteocalcin is a hormone that acts on pancreatic beta cells to promote insulin production in response to leptin and can activate bone-based adipocytes to regulate insulin responsiveness.12 Thus, there is an interaction between bone
Osteocytes
Concentric lamellae of osteon
Circumferential lamellae
Osteon
Central (haversian) canal
Neurovascular bundle
Perforating (Volkmann’s) canal
Periosteum Compact bone Spongy bone
Fig. 1.1 A depiction of the ultrastructural characteristics of bone. For clarity, 2 or 3 concentric lamellae are shown to represent the osteon; normally, the osteon is composed of 5 to 20 concentric lamellae. (Drawing by Alexandru Margarit and labeling by Alan Rosenberg. Printed with permission. All rights reserved.)
3CHAPTER 1 Structure and Function
development, fat cell function, and insulin production. Osteopontin participates in bone remodeling by securing osteoclasts to the inor-ganic (mineral) matrix of bone.13
After becoming entrapped in fully formed bone, osteoblasts become osteocytes within intramatrix cavities (lacunae). The mechanosensory osteocytes begin mineralizing hydroxylapatite, the bone’s organic matrix. Osteocytes are interconnected by cytoplasmic tendrils that traverse a net-work of canaliculi through which nutrients and metabolites are delivered to ensure osteocyte viability. Osteocytes, the most abundant bone cells, maintain the physiological balance between bone formation by osteo-blasts and bone resorption by osteoclasts, thus preserving bone integrity and mediating bone remodeling in response to mechanical stresses.
Osteoclasts mediate resorption at the inner surfaces of bone, such as the marrow cavity and cancellous bone trabeculae, and the outer
surfaces of bone, such as the epiphyses. Osteoclasts also participate in the conversion of cancellous bone into lamellar (compact) bone by creating spaces in which osteons can develop. Osteoclasts degrade bone by secreting lysosomal enzymes, resulting in calcium release to maintain mineral homeostasis.
Bone GrowthAfter birth, longitudinal bone growth occurs from the epiphyseal plate, which has four zones: reserve (resting), young proliferating car-tilage, maturing cartilage with hypertrophic chondrocytes, and calci-fying matrix (Fig. 1.3). The epiphyseal side of the plate is retained as articular cartilage, whereas on the diaphyseal side, the epiphysis ossifies to increase bone length. The dense cortex of long bones is enveloped by an outer layer of periosteum and an inner layer of endosteum.
Fig. 1.2 A simplified illustration of activation of molecular signaling cascades by soluble factors in the bone marrow that regulate mesenchymal stem cell differentiation into bone-producing cells. Cytokines, growth factors, and hormones within the bone marrow microenvironment (BMME) bind to their respective cognate receptors on the mesenchymal stem cell plasma membrane, initiating activation of signaling pathways that promote gene expression for differentiation of mesenchymal stem cells into cells that regulate osteoclas-togenesis. More detailed discussion of soluble factors in the bone marrow microenvironment that regu-late osteoclastogenesis is provided by Amarasekara et al.8 and Sobacchi et al.9 cAMP, Cyclic adenosine monophosphate; ERK, extracellular signal-regulated kinase; FAK, focal adhesion kinase; GH, growth hor-mone; IGF-1, insulin-like growth factor 1; IL, interleukin; JAK2, Janus kinase 2; MAPK, mitogen-activated protein kinase; MSC, mesenchymal stem cell; mTOR, mammalian target of rapamycin; NF-κB, nuclear factor κB; PI3K, phosphoinositide 3-kinase; PKA, protein kinase A; PKB, protein kinase B; PKCα, protein kinase C alpha; PLCγ, phospholipase C gamma; PTH, parathyroid hormone; p38, protein; RANKL, receptor activator of NF-κB ligand; SMAD, small mothers against decapentaplegic; STAT3, signal transducer and activator of transcription 3; TGF-β, transforming growth factor-beta; TNF, tumor necrosis factor; VEGF, vascular endothe-lial growth factor.
4 SECTION I Basic Concepts
The stages of skeletal maturation are reflected by the radiographic appearance of secondary ossification centers in long bones and physeal closure. The Tanner-Whitehouse14,15 and Greulich-Pyle16 protocols remain the conventional methods for assessing skeletal maturation (bone age) using left hand and wrist radiographs, although newer assessment methods have been considered.17 Bone growth at the phy-sis is governed by thyroid, growth, and sex hormones. Peak bone mass development during adolescence is determined by a variety of factors, including diet, physical activity, and synergistic effects of growth hor-mone and insulin-like growth factor. During the most rapid phases of adolescent growth, testosterone promotes cell division resulting in widening of the physis, and estrogen promotes calcification and clo-sure of the physis.18,19 Completion of ossification of the iliac apophyses is temporally related to cessation of the appendicular skeleton growth, although vertebral height may continue to increase and contribute to increasing height until the third decade of life.
The endochondral ossification that leads to longitudinal bone growth during childhood and adolescence requires the continuous availability of chondrocytes at the growth plate. During fetal and early neonatal periods, chondroprogenitor cells are progressively depleted. In contrast, as secondary ossification centers develop later in the bone growth process, chondrocytes in the physis begin to acquire charac-teristics of stem cells and, under the influence of two signaling path-ways (hedgehog and mammalian target of rapamycin complex 1),
differentiate into chondroprogenitor cells.20 In this way, a continuous supply of chondrocytes is available during growth.
Bone growth is accompanied by coincident angiogenesis, a process regulated by bone cells.21,22 Osteocytes, which have dendritic processes connected to blood vessels, induce angiogenesis by the vascular endo-thelial growth factor and mitogen-activated phosphorylated kinase signaling pathways.21
Growth factors are delivered to the physis through three routes of arterial supply: nutrient arteries, a metaphyseal-epiphyseal network, and a periosteal system (Fig. 1.4). In the resting state, bones receive approximately 10% to 15% of the cardiac output.23 Pathological states, such as joint inflammation, are associated with increased blood flow and acceleration of bone growth.
The nutrient blood vessels (together with nerves) penetrate the cor-tex through nutrient foramina, enter the medullary cavity, and then branch into ascending and descending arterioles that travel through the haversian system to supply blood at high-pressure flow to the inner two-thirds of mature bone (Fig. 1.4). The metaphyseal-epiphyseal vas-cular supply is derived from arteries arising from the periarticular vas-cular plexus that include metaphyseal, perichondral, and epiphyseal arteries. The periosteal vascular flow is a low-pressure system supply-ing the outer one-third of bone through the haversian and Volkmann canals. Evidence indicates that an elaborate and dense transcortical capillary network is the terminal pathway for blood delivery to bone.24
Fig. 1.3 Histology of the epiphysis. Cells at the top, the epiphyseal side, undergo rapid mitosis to increase the thickness of the epiphyseal plate resulting in increased bone length. In the hypertrophic zone, matrix is deposited and, as matrix accumulates, cells in the vicinity die. (The histological image, to which radiograph and labeling have been added, is attributed to the copyright holder, Springer Publishing, and appeared in Horvai A. Anatomy and histology of cartilage In TM Links, Editor, Cartilage Imaging, Springer, 2011, Figure 1-8. Reprinted with permission.)
5CHAPTER 1 Structure and Function
Circumferential growth, an increase in bone width, results from appositional subperiosteal membranous ossification on the bone surface.
Bone RemodelingMaintaining the integrity of mature bone, repairing damaged bone, and releasing calcium from bone are mediated by a shared process called bone remodeling. Bone remodeling, which occurs continuously throughout life, is more rapid during childhood and adolescence. The regulation of bone modeling during development and subsequent bone remodeling that occurs after birth requires direct interactions between osteoblasts and osteoclasts, endocrine and neurological con-trols, and innate immune and inflammatory regulators.
The metabolic events that initiate bone remodeling are unclear, but the death of osteocytes is likely to be an early event in the repair/remod-eling cascade. After osteocytes die, osteoclasts degrade bone matrix, forming lacunae into which mesenchymal cells migrate. Intralacunae mesenchymal cells differentiate into osteoblasts resulting in new bone matrix formation and subsequent mineralization. Depending on bone size and patient age, this remodeling process might take up to 6 months. Bone remodeling occurs continually even in adults; it is estimated that the entire healthy adult skeleton is remodeled several-fold during a lifetime.
The controlled balance between bone-forming osteoblasts and bone-resorbing osteoclasts is fundamental to sustaining bone health. Intercellular signaling by the wingless-related integrated site (Wnt) family of glycoproteins regulates the transcription cofactor β-cat-enin, which is required for signaling the expression of genes involved in development.25,26 The Wnt/β-catenin signaling pathway (referred to as the canonical Wnt signaling pathway) is crucial for growth and development of a wide array of tissues, including bone. Wnt signaling is integral for skeletal formation and embryonic limb development, chondrogenesis, production of bone matrix by osteoblasts, and the
differentiation and activity of osteoclasts. Wnt/β-catenin signaling prods mesenchymal stem cells to differentiate into osteoblasts, chon-drocytes, or osteocytes. The Wnt/β-catenin pathway enhances bone mass by promoting stem cell renewal and osteoblastogenesis, suppress-ing osteoblast and osteocyte apoptosis, and sensing mechanical load-ing. Bone remodeling also occurs in response to endocrine influences such as thyroid, growth, and sex hormones.
Bone and the Immune and Inflammatory SystemsNew knowledge about the important connections between the skeletal and immune systems has fostered osteoimmunology as a research and clinical field.3,27-30 Bone and immune and inflammatory systems are integrally related in both physiological and pathological circumstances.
Immune and inflammatory cells and mediators arise from pro-genitor cells in bone marrow. As an example, the receptor activator of the nuclear factor (NF)-κB ligand (RANKL), a member of the tumor necrosis factor (TNF) superfamily derived from osteocytes, primor-dial osteoblasts, and T cells, is a cytokine that links bone with immune response.31 RANKL mediates osteoclast differentiation by transmitting its signal through its cognate receptor, RANK. RANK and RANKL gene knockout mice, having no osteoclasts, have a phenotype characterized by severe osteopetrosis and immune deficiency associated with absent lymph nodes.32-35 RANKL is expressed by osteocytes and hypertrophic chondrocytes in response to factors that promote bone production, including vitamin D, parathyroid hormone, and prostaglandins. The binding of a RANK receptor on osteocytes to its ligand RANKL on osteoblasts is a key inducer of bone resorption. Osteoprotegerin, pro-duced by osteoblasts, inhibits osteoclastogenesis by serving as a decoy receptor for RANKL and thus inhibits bone resorption. In addition to its role in bone metabolism, RANKL/RANK also participate in immune activation, immune tolerance, and development of thymus and lymph node tissues.33,36
Fig. 1.4 Distribution of blood supply in growing bone. (Image design concept inspired by GJ Tortora and B Derrickson, Principles of Anatomy & Physiology, 14th Edition, John Wiley and Sons, page 175, Figure 6-4. Image drawn by Alexandru Margarit and labeled by Alan Rosenberg and reprinted with permission. All rights reserved.)
6 SECTION I Basic Concepts
The processes of bone turnover and inflammation are regulated by shared mediators, including cytokines and transcription factors.3,37 Bone marrow–derived T lymphocytes are integral to regulating bone remodeling and the response of bone cells to parathyroid hormone.37 Osteoclastic-mediated bone resorption is generated by T cell–derived cytokines.8 T cells also promote bone formation by direct interaction with bone cells in part by activating Wnt signaling in the osteoblast cell lineage. T helper 17 (Th17) cells stimulate bone resorption by promoting osteoclast differentiation.38 Th17 cells promote osteoclas-togenesis in rheumatic diseases by elaborating TNF, interleukin (IL)-1, IL-6, IL-17, and RANKL. IL-17 stimulates the release of RANKL by osteoblasts and osteocytes and upregulates RANK expression on osteo-cytes. By producing antiinflammatory cytokines IL-4, IL-10, and trans-forming growth factor-beta (TGF-β), T regulatory cells (Tregs) inhibit osteoclastogenesis and thus promote bone formation.
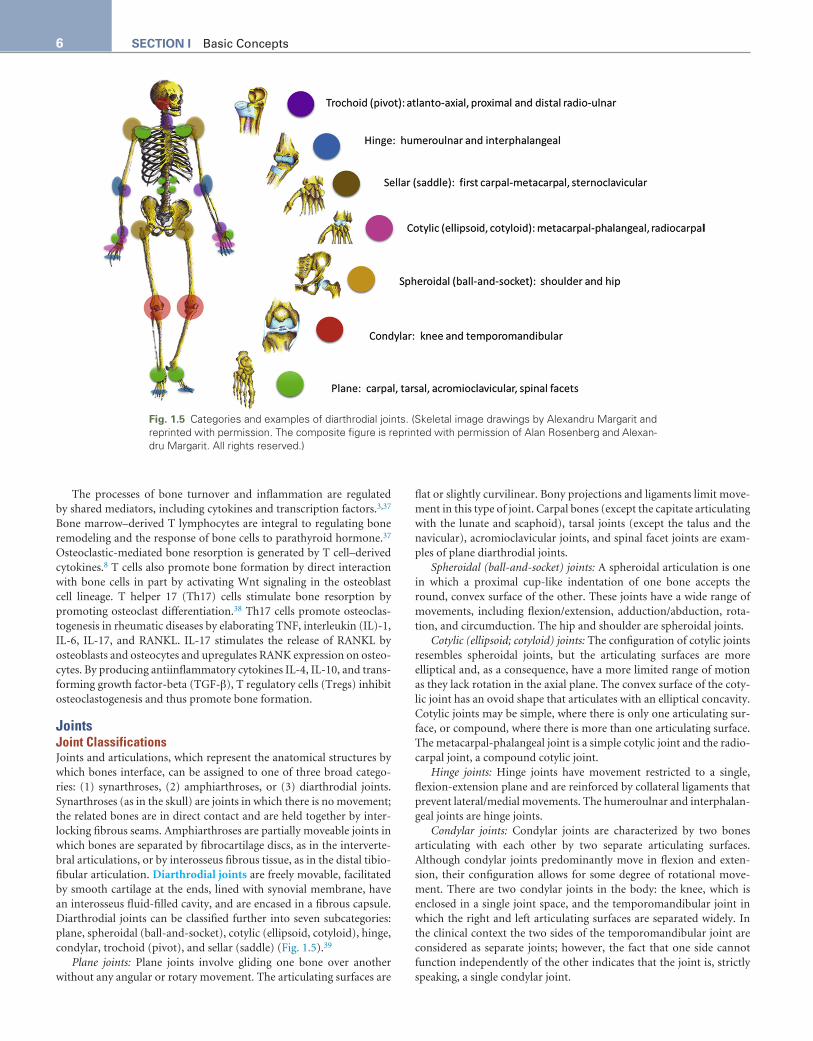
JointsJoint ClassificationsJoints and articulations, which represent the anatomical structures by which bones interface, can be assigned to one of three broad catego-ries: (1) synarthroses, (2) amphiarthroses, or (3) diarthrodial joints. Synarthroses (as in the skull) are joints in which there is no movement; the related bones are in direct contact and are held together by inter-locking fibrous seams. Amphiarthroses are partially moveable joints in which bones are separated by fibrocartilage discs, as in the interverte-bral articulations, or by interosseus fibrous tissue, as in the distal tibio-fibular articulation. Diarthrodial joints are freely movable, facilitated by smooth cartilage at the ends, lined with synovial membrane, have an interosseus fluid-filled cavity, and are encased in a fibrous capsule. Diarthrodial joints can be classified further into seven subcategories: plane, spheroidal (ball-and-socket), cotylic (ellipsoid, cotyloid), hinge, condylar, trochoid (pivot), and sellar (saddle) (Fig. 1.5).39
Plane joints: Plane joints involve gliding one bone over another without any angular or rotary movement. The articulating surfaces are
flat or slightly curvilinear. Bony projections and ligaments limit move-ment in this type of joint. Carpal bones (except the capitate articulating with the lunate and scaphoid), tarsal joints (except the talus and the navicular), acromioclavicular joints, and spinal facet joints are exam-ples of plane diarthrodial joints.
Spheroidal (ball-and-socket) joints: A spheroidal articulation is one in which a proximal cup-like indentation of one bone accepts the round, convex surface of the other. These joints have a wide range of movements, including flexion/extension, adduction/abduction, rota-tion, and circumduction. The hip and shoulder are spheroidal joints.
Cotylic (ellipsoid; cotyloid) joints: The configuration of cotylic joints resembles spheroidal joints, but the articulating surfaces are more elliptical and, as a consequence, have a more limited range of motion as they lack rotation in the axial plane. The convex surface of the coty-lic joint has an ovoid shape that articulates with an elliptical concavity. Cotylic joints may be simple, where there is only one articulating sur-face, or compound, where there is more than one articulating surface. The metacarpal-phalangeal joint is a simple cotylic joint and the radio-carpal joint, a compound cotylic joint.
Hinge joints: Hinge joints have movement restricted to a single, flexion-extension plane and are reinforced by collateral ligaments that prevent lateral/medial movements. The humeroulnar and interphalan-geal joints are hinge joints.
Condylar joints: Condylar joints are characterized by two bones articulating with each other by two separate articulating surfaces. Although condylar joints predominantly move in flexion and exten-sion, their configuration allows for some degree of rotational move-ment. There are two condylar joints in the body: the knee, which is enclosed in a single joint space, and the temporomandibular joint in which the right and left articulating surfaces are separated widely. In the clinical context the two sides of the temporomandibular joint are considered as separate joints; however, the fact that one side cannot function independently of the other indicates that the joint is, strictly speaking, a single condylar joint.
Fig. 1.5 Categories and examples of diarthrodial joints. (Skeletal image drawings by Alexandru Margarit and reprinted with permission. The composite figure is reprinted with permission of Alan Rosenberg and Alexan-dru Margarit. All rights reserved.)
7CHAPTER 1 Structure and Function
Trochoid (pivot) joints: Trochoid joints have movement limited to rotation around one axis. The mobile bone rotates within a ring formed in the adjacent bone by a concavity and associated ligaments. The atlantoaxial and proximal and distal radioulnar joints are exam-ples of this type of articulation.
Sellar (saddle) joints: Sellar joints are distinguished by each bone of the articulation having convex and concave surfaces that recipro-cally interlock. Like cotylic joints, they have a wide array of movements except for axial rotation. The first carpometacarpal joint (the articula-tion between the first metacarpal of the thumb and the trapezium) and the sternoclavicular joints are sellar joints.
Diarthrodial Joint StructuresDevelopment of diarthrodial joints. Embryonic development of
synovial joints is regulated by genes and signaling networks that promote cartilage condensation (growth differentiation factor-5), cavitation (WNT9A), and tissue differentiation.40 The diarthrodial joint anlage comprises a three-layered interzone. The dense chondrogenic middle zone is the site of cavitation, the future joint space, which occurs at 8 weeks gestational age.41 The various joint components, including joint capsule, synovial membrane, menisci, and intraarticular ligaments and tendons, are derived from interzone mesenchymal cells.
Anatomy of diarthrodial joints. Diarthrodial joints are enveloped by a capsule of dense fibrous tissue that can be reinforced by adjacent ligaments and tendons. The capsule, which attaches to bones of diarthrodial joints by fibrocartilaginous insertions, provides stability to the joint, limits joint mobility, and contains the intraarticular fluid.
The synovium attaches to the cartilage-bone junction. In healthy joints the synovium does not impinge on the articular surface; when inflamed, the synovial membrane may proliferate and encroach on the articular surface as inflammatory pannus.
Synovium. The synovial membrane (synovium), a tissue of ectodermal origin, lines the inner surface of the diarthrodial joint capsule and intraarticular ligaments, tendons, and fat pads but not articular cartilage or menisci. The synovial membrane is configured with pouches and folds which, together with its inherent deformability and property of nonadherence to adjacent tissues, allows unhindered movement. It is the source of synovial fluid that nourishes and lubricates the joint cartilage.
There are two synovial membrane layers: the inner intima and the outer subintima. The intima, composed of two to three layers of a mixture of macrophage and fibroblast cells, is interposed between the joint cavity and the subintima to which it is loosely attached. There is no basement membrane delineating the thin intima from the subintima; rather, the intima rests loosely on intermingling col-lagen fibrils, vascular and lymphatic vessels, and nerves of the sub-intima with a film of hyaluronic acid interposed between the two layers. This loose network accounts for the synovial lining’s physi-ological permeability. Normal synovial lining is also replete with a wide variety of adhesion molecules, vasculature, and cells. Adhesion molecules help ensure that crucial components are not lost during compression and shearing movements. Adhesion molecules also are involved in the recruitment of inflammatory cells under patholog-ical states.
Synovial lining cells within the synovial intimal layer can be type A macrophage-like synoviocytes or type B fibroblast-like synoviocytes. Synovial membrane macrophages scavenge intraarticular debris through phagocytosis and participate in antigen-mediated immune responses. Synovial membrane fibroblasts produce the main joint lubricants, lubricin and hyaluronic acid,42 as well as cytokines, pros-taglandins, collagen, proteoglycans, and metalloproteinases and their cognate inhibitors.
Vasculogenesis within the synovium begins during embryonic development. Normal synovial blood flow is governed locally by autocrine and paracrine influences and extrinsically by neuronal and humoral factors. With inflammation, there is accelerated angiogenesis and increased blood flow to the synovium. The inflamed synovial tis-sue is proportionally more proliferative than the vasculature, resulting in a relatively smaller vascular network and consequent synovial mem-brane hypoxia. Intraarticular hypoxia results in a switch in synovial cells from a resting metabolic state to a markedly increased metabolic state to ensure cell survival. As a result, metabolic intermediates accu-mulate and promote hypoxia-induced, proinflammatory signaling pathways.43
The synovial membrane microvasculature is linked to the vascu-lature of the periarticular bone and periosteum. The microvascular network includes arterioles and capillaries in the intimal region and venules and lymphatics in the subintima. The dense neural network follows blood vessels into the synovial lining. These nerve fibers tend to be slow conducting fibers resulting in sensations of aching and burning in inflammatory states.
Synovial fluid. Synovial fluid, ordinarily clear, transparent, pale yellow, and viscous, is a plasma transudate derived from the subintimal capillary network and serves to nourish mature avascular cartilage and lubricate, protect, and regulate temperature within joints. Hyaluronic acid in the fluid accounts for normal synovial fluid’s high viscosity. Hyaluronic acid appears to work synergistically with lubricin, a synovial fluid glycoprotein, to lubricate the articulating surfaces.44,45 The synovial fluid also serves as a shock-absorbing cushion interposed between articulating bones. Some synovial fluid coats the cartilage surface and some is absorbed into cartilage; when the cartilage is compressed, fluid is expressed to reduce friction. The small amount of synovial fluid in a healthy joint reduces compressive shock by dispersing pressure evenly across the articular surface.
The attributes of synovial fluid quantity, color, clarity, and viscosity can be easily assessed at the bedside to help distinguish normal from inflammatory, septic, and hemorrhagic synovial fluid. The volume of synovial fluid in a healthy joint is small (<3.5 mL in an adult knee). The fluid is clear, colorless, or pale yellow; it has a high viscosity, a neutral pH (7.4), a protein concentration of 1.7 to 2.1 g/dL, and a glu-cose level that is within approximately 10 mg/dL of the serum level.46-50 There are no red blood cells and fewer than 200/mm3 white blood cells. Ingress of larger molecules, such as complement, is limited. Synovial fluid is devoid of elements of the coagulation pathway, and, conse-quently, normal synovial fluid does not clot.51
Articular cartilage. Normal cartilage on the articular surface of diarthrodial joints has a glassy, translucent, pearl-white appearance, giving rise to the term hyaline cartilage (from the Latin hyalinus and Greek hyalinos meaning of glass or crystal). In addition to providing structural form, hyaline cartilage minimizes friction within articulations and, because of its inherent compressibility, protects from mechanical loads. The cartilage margins blend with the synovial membrane and with metaphyseal periosteum. Hyaline cartilage includes chondrocytes, which become progressively less abundant with advancing age, within an extracellular matrix that is predominantly type II collagen. The cell volume of adult cartilage is less than 2%.52 During cartilage development and fetal and postnatal growth, cartilage has abundant metaphyseal and epiphyseal vascularity. In children, the epiphyseal cartilage is vascular until approximately 18 months of age after which transphyseal vessels regress, leaving remnant epiphyseal vascular canals. These canals can increase in size with advancing age and can become more conspicuous in the presence of inflammation.53,54 Articular cartilage is aneural.
8 SECTION I Basic Concepts
Healthy articular cartilage is mostly water (70%). Type II collagen and aggrecan, an aggregate of proteoglycans, account for most of the dry weight of cartilage; other collagen types and unaggregated proteo-glycans contribute minimally to cartilage volume.
Macroscopically, hyaline cartilage appears as a homogenous tissue. Microscopically, cartilage has four discrete layers (Fig. 1.6). The most superficial region (the gliding or tangential zone) is characterized by flattened chondrocytes, fine collagen fibrils arranged in a tangential array, a high concentration of small molecular weight proteoglycans (decorin),55 and a low concentration of aggrecan. The middle (tran-sitional) zone, accounting for half the cartilage mass, includes round chondrocytes surrounded by thick strands of collagen fibers arranged radially. In the deep zone, chondrocytes are aligned in columns. The number of chondrocytes gradually decreases from the superficial to the deeper layers, but the volume of each cell progressively increases through to the deeper zones. Water content progressively decreases from the superficial to deeper zones. Collagen is the most abundant and proteoglycan content is the least abundant in the superficial zone; proteoglycans become proportionally more abundant in the deeper zones accounting for 50% of dry weight. As bone growth finishes, a calcified zone, formed by endochondral ossification, persists to pro-vide a protective buffer between the soft uncalcified articular cartilage and the underlying subchondral bone.
At some sites, such as the pubic symphysis, the annulus fibrosus of the intervertebral discs, the triangular fibrocartilage of the wrist, and the temporomandibular joint, fibrocartilage rather than hyaline carti-lage is interposed between bones of the respective joints.
Extracellular matrix.Collagen. Collagen (from the Greek kόlla for glue and gen for
producing), the dominant constituent of connective tissue, is the most abundant and ubiquitously distributed extracellular structural protein in the body, accounting for approximately one-third of the human body’s total protein content.
After transcription and translation of the procollagen α chains, four distinct stages occur for the assembly of collagen fibrils. The first stage is the transportation of the α chains into the rough endoplasmic retic-ulum where they are modified to form triple-helical procollagen. The
second stage is the modification of procollagen in the Golgi apparatus and its packaging into secretory vesicles. The third stage is the forma-tion of the collagen molecule in the extracellular space by cleavage of the procollagen. The final step is the cross-linking between the collagen molecules to stabilize the supramolecular collagen structure.
Collagen is distinguished from other proteins by its high content of proline (Pro), hydroxyproline (Hyp), and glycine (Gly), which occu-pies every third amino acid position. Most of the proline in collagen is present in the sequence Gly-Pro-Xaa, where Xaa (an unspecified amino acid) is frequently alanine or hydroxyproline. The stereochem-ical constraints imposed by proline and hydroxyproline result in the repeating tripeptide sequences to form the triple-helical configuration. Glycine is essential for ensuring the stability of the triple-helical con-formation. Tropocollagen, the basic unit of collagen, is a triple-heli-cal structure. The three polypeptide chains (except for the amino and carboxy termini) form a left-handed helix, while the tropocollagen superhelix coils to the right. Tropocollagen assembles into collagen fibrils, which are then bundled (as in tendon and bone), stacked (as in cornea), or loosely connected (as in interstitial tissue) to form the structural fibrous tissue within respective organs.
Although there are an estimated 28 different collagen types,56 type I collagen accounts for more than 90% of all collagen. Type II collagen has the most restricted tissue distribution, being confined to hyaline cartilage, intervertebral discs, and the vitreous of the eye.
Collagen and cartilage proteases: Proteinases, endopeptidase enzymes that cleave by hydrolyzing peptide bonds in the middle of the molecule, are integral to ensuring bone and cartilage homeostasis.57 Endopeptidases are classified based on the chemical group that is targeted for hydrolysis. Endopeptidases that act extracellularly in a neutral pH are serine and metalloproteinases, and those acting intracellularly in an acid pH are aspartate, cysteine, and threonine proteinases. Extracellular homeostasis is regulated partly by an array of proteolytic enzymes that account for continual physiological bone remodeling or degradation in pathological states such as inflammation. Matrix metalloproteinases (and their respec-tive inhibitors, tissue inhibitors of matrix metalloproteinases) are clas-sified into five broad categories (collagenases, gelatinases, stromelysins, matrilysins, and membrane type) that relate to the substrate they target, their structure, and their location within the cell.58
Proteoglycans: Proteoglycans are glycosylated proteins to which a multitude of anionic glycosaminoglycans chains (e.g., chondroitin sulfate, keratan sulfate, and heparan sulfate) are bound. Proteoglycans may link to each other to form aggrecan, or to hyaluronic acid or other matrix proteins such as collagen. Hydrodynamics is the predominant physiological function of proteoglycans as cationic water molecules bind to the negatively charged glycosaminoglycan sulfates. Aggregates of proteoglycans, which provide resistance to compression, are distrib-uted among the networks of collagen fibers and confer tensile strength to the articular cartilage. The water molecules bound to the hydrophilic proteoglycans within cartilage provide resistance to deformation on compression. As cartilage is compressed, water is extruded but quickly reabsorbed, allowing the cartilage to regain is precompressive form.
An essential biological role of heparan sulfate proteoglycan in the extracellular matrix is to bind an array of cytokines to regulate their availability and mobility. The relative abundance of various proteo-glycans varies with age; with advancing age, proteoglycan aggregates increase, keratin sulfate increases, and chondroitin sulfate decreases.
TENDONSTendons provide the anatomical connection between bone and muscle. The tendon allows a large mass of muscle to focus its force on a small area of bone, or if the tendon is split, to distribute the force of the
Fig. 1.6 Histological section of normal articular cartilage stained with hematoxylin and eosin. Healthy articular cartilage includes three zones: superficial, intermediate, and deep zones. The tidemark demarcates the deep zone from the calcified zone and the underlying subchondral bone. (The histological image, to which labeling has been added, is reproduced with permission of Dr. Andrew Horvai.)
9CHAPTER 1 Structure and Function
muscle to multiple bones. The tendon comprises predominantly type I collagen (85% of dry weight) and small amounts of type III collagen. Tendon fibers are orientated longitudinally along the lines of stress within a proteoglycan, elastin, neurovascular, and fibroblast matrix. Decorin, the most abundant proteoglycan in the tendon, establishes cross-linking between collagen fibers to facilitate the transfer of loads among fibers. The hierarchical tendon structure includes microfibrils, subfibrils, fibrils, fascicles, and the main tendon unit. The movement of many tendons, particularly large ones, is facilitated by gliding through tubes comprising an inner synovial sheath and an outer fibrous sheath; mesenchymal cells from the synovial sheath produce hyaluronic acid that promotes lubrication to facilitate movement. Fibroblasts within the tendons produce collagens, proteoglycans, and matrix metallopro-teinases and their inhibitors that mediate tendon damage and repair.
LIGAMENTS AND FASCIALigaments and fascia provide connections between bones, conferring stability and constraining movement. Although resembling tendons structurally, ligaments have more elastin and type I collagen fibers. Ligaments are longitudinally orientated, as in tendons, but tend to be crimped, allowing for the ability to stretch and recoil.
ENTHESESThe attachment of tendons and ligaments to bone occurs at the enthe-sis (from the Greek insertion), a distinct, specialized interface where tendon and ligament fibers gradually blend into bone tissue.59,60 The entheseal connection can take the form of either a dense fibrous con-nective tissue or, more commonly, fibrocartilage. Fibrous entheses are characterized by mineralized collagen fibers inserting directly into bone or periosteal tissue, entering into a broad section of the diaphysis (e.g., the deltoid insertion into the humerus). Fibrocartilage entheses are characterized by a four-zone gradient, transitioning from uncalci-fied tendon to bone (Fig. 1.7). The tendon/ligament, comprising types I and III collagen fibers, elastin, proteoglycans, and fibroblasts, transi-tions to zone two, an uncalcified fibrocartilaginous zone comprising types I, II, and III collagens, aggrecan, and chondrocytes. This second
zone appears as a tidemark demarcating the uncalcified from calci-fied fibrocartilage; the tidemark zone serves as a mechanical interface between soft and hard tissue. The third zone, calcified fibrocartilage, comprises mostly type II collagen with some aggrecan, types I and X collagen, and chondrocytes. The fourth zone is bone.
A unique T cell lineage within entheseal sites contributes to the patho-genesis of enthesitis. IL-23, produced in response to Th17 cells, is an essential cytokine in the development of inflammation at entheses, acting on the resident T cells to promote IL-17–mediated and IL-22–mediated inflammation and bone remodeling at the enthesopathic site.61,62
BURSAEThin-walled bursal sacs, which either appear during embryonic devel-opment contemporaneously with synovial membranes or develop postnatally, have ultrastructure resembling synovial tissue but are less vascular. By cushioning and facilitating gliding between points of pressure, bursae prevent damage that would otherwise be incurred by the rubbing of one tissue against another. Most bursae in the human body are in the vicinity of large joints such as the shoulder, hip, and knee. The location of bursae can be subfascial, subligamentous, sub-tendinous, or submuscular. Bursae may increase in size, or new bursal sacs (adventitious bursa) may develop in response to inflammation or trauma. In the presence of intraarticular inflammation, communica-tion between deep bursae and the joint space may develop. As exam-ples, the lateral subtendinous bursa of the gastrocnemius muscle can communicate with the knee joint, the subacromial bursa with the gle-nohumeral joint, and the iliopsoas bursa with the hip joint. Superficial bursae, such as the olecranon bursa and the prepatellar bursa, are less likely to form a conduit to the respective joint spaces.
SKELETAL MUSCLEAs strong and functional as the skeleton is, its operations would be impossible without the support and activities of muscles. The more than 600 human muscles account for approximately 40% of the body’s mass. During the third gestational week, the paraxial mesoderm under-goes somitogenesis, yielding spherical somites adjacent to each side of the neuronal groove. Skeletal muscle forms from these somites and mononucleated myoblasts differentiate into multinucleated myotubes.
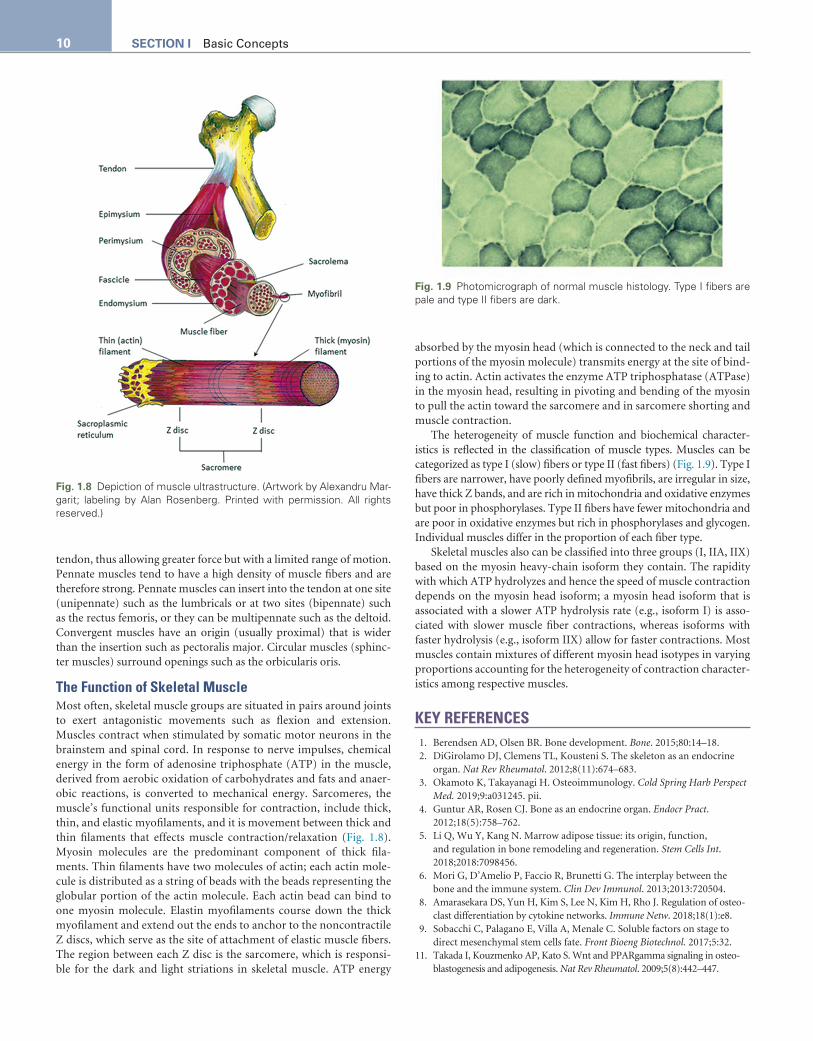
Entire muscles are surrounded and secured by the epimysium, a sheath of connective tissue (Fig. 1.8). Within each muscle are bun-dles of muscle fibers (fascicles) that function in concert. Fascicles are cloaked with a connective sheath, the perimysium. Within each fasci-cle, individual muscle fibers are each surrounded by a loose connec-tive tissue, the endomysium, through which terminal nerve fibers and capillaries pass. The perimysium and endomysium blend together as the muscle fibers merge with tendons, thus helping to reinforce the attachment of muscle to tendon. Muscle fibers each contain a nucleus, a plasma membrane (the sarcolemma), and cytoplasm (sarcoplasm). Inside each muscle cell are myofibrils, protein bundles comprising myofilamentous fibers (sarcomeres).
Classification of Skeletal Muscles Based on ShapeThe variety of shapes and configurations of skeletal muscles reflects their respective mechanical actions. Fusiform muscles, such as the biceps brachii, are centrally thick with tapered ends and have muscle fibers aligned in parallel along the long axis. Parallel muscles, such as the sartorius and sternocleidomastoid, also have muscle fibers aligned in parallel. They are longer and less strong than fusiform muscles but can endure relatively longer loading. Pennate (feather-like) mus-cles have fascicles that are obliquely aligned to the orientation of the
Fig. 1.7 Histological section, stained with Masson trichrome, showing a normal enthesis. The red vertical lines on the left are collagen tendon fibers. The blue color in the middle of the image is fibrocartilage. The red band on the right is the transition zone (tidemark) from fibrocarti-lage at the attachment site to bone. The white on the far right is fat within the bone cavity. (Image reproduced from http://www.enthesis.info/anatomy/enthesis_insertion.html with permission from Dr. Dennis G. McGonagle.)
tendon, thus allowing greater force but with a limited range of motion. Pennate muscles tend to have a high density of muscle fibers and are therefore strong. Pennate muscles can insert into the tendon at one site (unipennate) such as the lumbricals or at two sites (bipennate) such as the rectus femoris, or they can be multipennate such as the deltoid. Convergent muscles have an origin (usually proximal) that is wider than the insertion such as pectoralis major. Circular muscles (sphinc-ter muscles) surround openings such as the orbicularis oris.
The Function of Skeletal MuscleMost often, skeletal muscle groups are situated in pairs around joints to exert antagonistic movements such as flexion and extension. Muscles contract when stimulated by somatic motor neurons in the brainstem and spinal cord. In response to nerve impulses, chemical energy in the form of adenosine triphosphate (ATP) in the muscle, derived from aerobic oxidation of carbohydrates and fats and anaer-obic reactions, is converted to mechanical energy. Sarcomeres, the muscle’s functional units responsible for contraction, include thick, thin, and elastic myofilaments, and it is movement between thick and thin filaments that effects muscle contraction/relaxation (Fig. 1.8). Myosin molecules are the predominant component of thick fila-ments. Thin filaments have two molecules of actin; each actin mole-cule is distributed as a string of beads with the beads representing the globular portion of the actin molecule. Each actin bead can bind to one myosin molecule. Elastin myofilaments course down the thick myofilament and extend out the ends to anchor to the noncontractile Z discs, which serve as the site of attachment of elastic muscle fibers. The region between each Z disc is the sarcomere, which is responsi-ble for the dark and light striations in skeletal muscle. ATP energy
absorbed by the myosin head (which is connected to the neck and tail portions of the myosin molecule) transmits energy at the site of bind-ing to actin. Actin activates the enzyme ATP triphosphatase (ATPase) in the myosin head, resulting in pivoting and bending of the myosin to pull the actin toward the sarcomere and in sarcomere shorting and muscle contraction.
The heterogeneity of muscle function and biochemical character-istics is reflected in the classification of muscle types. Muscles can be categorized as type I (slow) fibers or type II (fast fibers) (Fig. 1.9). Type I fibers are narrower, have poorly defined myofibrils, are irregular in size, have thick Z bands, and are rich in mitochondria and oxidative enzymes but poor in phosphorylases. Type II fibers have fewer mitochondria and are poor in oxidative enzymes but rich in phosphorylases and glycogen. Individual muscles differ in the proportion of each fiber type.
Skeletal muscles also can be classified into three groups (I, IIA, IIX) based on the myosin heavy-chain isoform they contain. The rapidity with which ATP hydrolyzes and hence the speed of muscle contraction depends on the myosin head isoform; a myosin head isoform that is associated with a slower ATP hydrolysis rate (e.g., isoform I) is asso-ciated with slower muscle fiber contractions, whereas isoforms with faster hydrolysis (e.g., isoform IIX) allow for faster contractions. Most muscles contain mixtures of different myosin head isotypes in varying proportions accounting for the heterogeneity of contraction character-istics among respective muscles.
KEY REFERENCES 1. Berendsen AD, Olsen BR. Bone development. Bone. 2015;80:14–18. 2. DiGirolamo DJ, Clemens TL, Kousteni S. The skeleton as an endocrine
organ. Nat Rev Rheumatol. 2012;8(11):674–683. 3. Okamoto K, Takayanagi H. Osteoimmunology. Cold Spring Harb Perspect
Med. 2019;9:a031245. pii. 4. Guntur AR, Rosen CJ. Bone as an endocrine organ. Endocr Pract.
2012;18(5):758–762. 5. Li Q, Wu Y, Kang N. Marrow adipose tissue: its origin, function,
and regulation in bone remodeling and regeneration. Stem Cells Int. 2018;2018:7098456.
6. Mori G, D’Amelio P, Faccio R, Brunetti G. The interplay between the bone and the immune system. Clin Dev Immunol. 2013;2013:720504.
8. Amarasekara DS, Yun H, Kim S, Lee N, Kim H, Rho J. Regulation of osteo-clast differentiation by cytokine networks. Immune Netw. 2018;18(1):e8.
9. Sobacchi C, Palagano E, Villa A, Menale C. Soluble factors on stage to direct mesenchymal stem cells fate. Front Bioeng Biotechnol. 2017;5:32.
11. Takada I, Kouzmenko AP, Kato S. Wnt and PPARgamma signaling in osteo-blastogenesis and adipogenesis. Nat Rev Rheumatol. 2009;5(8):442–447.
Fig. 1.8 Depiction of muscle ultrastructure. (Artwork by Alexandru Mar-garit; labeling by Alan Rosenberg. Printed with permission. All rights reserved.)
Fig. 1.9 Photomicrograph of normal muscle histology. Type I fibers are pale and type II fibers are dark.
11CHAPTER 1 Structure and Function
20. Newton PT, Li L, Zhou B, et al. A radical switch in clonality reveals a stem cell niche in the epiphyseal growth plate. Nature. 2019;567(7747):234–238.
22. Sivaraj KK, Adams RH. Blood vessel formation and function in bone. Development. 2016;143(15):2706–2715.
23. Tomlinson RE, Silva MJ. Skeletal blood flow in bone repair and mainte-nance. Bone Res. 2013;1(4):311–322.
24. Grüneboom A, Hawwari I, Weidner D, et al. A network of trans-cortical capillaries as mainstay for blood circulation in long bones. Nat Metab. 2019;1(2):236–250.
26. Karner CM, Long F. Wnt signaling and cellular metabolism in osteoblasts. Cell Mol Life Sci. 2017;74(9):1649–1657.
27. Ginaldi L, De Martinis M. Osteoimmunology and beyond. Curr Med Chem. 2016;23(33):3754–3774.
37. Pacifici R. T cells, osteoblasts, and osteocytes: interacting lineages key for the bone anabolic and catabolic activities of parathyroid hormone. Ann N Y Acad Sci. 2016;1364(1):11–24.
39. Polley H, Hunder G. Rheumatologic Interviewing and Physical Examination of the Joints. 2nd ed. Toronto: W.B. Saunders Company; 1978.
40. Salva JE, Merrill AE. Signaling networks in joint development. Dev Dyn. 2017;246(4):262–274.
41. Decker RS. Articular cartilage and joint development from embryogenesis to adulthood. Semin Cell Dev Biol. 2017;62:50–56.
42. Hui AY, McCarty WJ, Masuda K, Firestein GS, Sah RL. A systems biology approach to synovial joint lubrication in health, injury, and disease. Wiley Interdiscip Rev Syst Biol Med. 2012;4(1):15–37.
43. Fearon U, Hanlon MM, Wade SM, Fletcher JM. Altered metabolic path-ways regulate synovial inflammation in rheumatoid arthritis. Clin Exp Immunol. 2019;197(2):170–180.
55. Zhang W, Ge Y, Cheng Q, et al. Decorin is a pivotal effector in the extracellular matrix and tumour microenvironment. Oncotarget. 2018;9(4):5480–5491.
59. Waghray N, Jyothi A, Imran M, Yaseen S, Chaudhary U. Enthesis: a brief review. Apollo Med. 2015;12:32–38.
60. Apostolakos J, Durant TJ, Dwyer CR, et al. The enthesis: a review of the tendon-to-bone insertion. Muscles Ligaments Tendons J. 2014;4(3):333–342.
61. Lories RJ, McInnes IB. Primed for inflammation: enthesis-resident T cells,. Nat Med. 2012;18(7):1018–1019.
Complete reference lists are available in www.expertconsult.com
ONLINE REFERENCES 1. Berendsen AD, Olsen BR. Bone development. 2015;80:14–18. 2. DiGirolamo DJ, Clemens TL, Kousteni S. The skeleton as an endocrine
organ. Nat Rev Rheumatol. 2012;8:674–683. 3. Okamoto K, Takayanagi H. Osteoimmunology. Cold Spring Harb Perspect
Med. 2019;9:a031245. pii. 4. Guntur AR, Rosen CJ. Bone as an endocrine organ. Endocr Pract.
2012;18(5):758–762. 5. Li Q, Wu Y, Kang N. Marrow adipose tissue: its origin, function,
and regulation in bone remodeling and regeneration. Stem Cells Int. 2018;2018:7098456.
6. Mori G, D’Amelio P, Faccio R, Brunetti G. The interplay between the bone and the immune system. Clin Dev Immunol. 2013;2013:720504.
7. Pino AM, Rodríguez JP. Is fatty acid composition of human bone marrow significant to bone health? Bone. 2019;118:53–61.
8. Amarasekara DS, Yun H, Kim S, et al. Regulation of osteoclast differentia-tion by cytokine networks. Immune Netw. 2018(1);18:e8.
9. Sobacchi C, Palagano E, Villa A, Menale C. Soluble factors on stage to direct mesenchymal stem cells fate. Front Bioeng Biotechnol. 2017;5:32.
10. Karsenty G, Kronenberg HM, Settembre C. Genetic control of bone for-mation. Annu Rev Cell Dev Biol. 2009;25:629–648.
11. Takada I, Kouzmenko AP, Kato S. Wnt and PPARgamma signaling in os-teoblastogenesis and adipogenesis. Nat Rev Rheumatol. 2009;5(8):442–447.
12. Kanazawa I. Osteocalcin as a hormone regulating glucose metabolism. World J Diabetes. 2015;6(18):1345–1354.
13. Reinholt FP, Hultenby K, Oldberg A, Heinegård D. Osteopontin-a possible anchor of osteoclasts to bone. Proc Natl Acad Sci USA. 1990;87(12): 4473–4475.
14. Tanner J, Whitehouse R, Cameron N, Marshall W, Healy M, Goldstein H. Assessment of Skeletal Maturity and Prediction of Adult Height (TW2). 2nd ed. London, UK: Academic Press; 1983.
15. Tanner G, Healy M, Goldstein H, Cameron N. Assessment of Skeletal Maturity and Prediction of Adult Height (TW3) Method. 3rd ed. London, UK: Saunders Ltd.; 2001.
16. Greulich W, Pyle S. Radiograph Atlas of Skeletal Development of the Hand and Wrist. 2nd ed. California: Stanford University Press; 1959.
17. Satoh M. Bone age: assessment methods and clinical applications. Clin Pediatr Endocrinol. 2015;24(4):143–152.
18. Chagin AS, Sävendahl L. Estrogens and growth: review. Pediatr Endocrinol Rev. 2007;4(4):329–334.
19. Chagin AS, Sävendahl L. Oestrogen receptors and linear bone growth. Acta Paediatr. 2007;96(9):1275–1279.
20. Newton PT, Li L, Zhou B, et al. A radical switch in clonality reveals a stem cell niche in the epiphyseal growth plate. Nature. 2019;567(7747):234–238.
21. Prasadam I, Zhou Y, Du Z, Chen J, Crawford R, Xiao Y. Osteocyte- induced angiogenesis via VEGF-MAPK-dependent pathways in endothelial cells. Mol Cell Biochem. 2014;386(1–2):15–25.
22. Sivaraj KK, Adams RH. Blood vessel formation and function in bone. Development. 2016;143(15):2706–2715.
23. Tomlinson RE, Silva MJ. Skeletal blood flow in bone repair and mainte-nance. Bone Res. 2013;1(4):311–322.
24. Grüneboom A, Hawwari I, Weidner D, et al. A network of trans-cortical capillar-ies as mainstay for blood circulation in long bones. Nat Metab. 2019;1(2):236–250.
25. MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17(1):9–26.
26. Karner CM, Long F. Wnt signaling and cellular metabolism in osteoblasts. Cell Mol Life Sci. 2017;74(9):1649–1657.
27. Ginaldi L, De Martinis M. Osteoimmunology and beyond. Curr Med Chem. 2016;23(33):3754–3774.
28. David JP. Osteoimmunology: a view from the bone. Adv Immunol. 2007;95:149–165.
29. Lorenzo J, Horowitz M, Choi Y. Osteoimmunology: interactions of the bone and immune system. Endocr Rev. 2008;29(4):403–440.
30. Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk be-tween the immune and bone systems. Nat Rev Immunol. 2007;7(4):292–304.
31. Nakashima T, Hayashi M, Fukunaga T, et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat Med. 2011;17(10):1231–1234.
32. Dougall WC, Glaccum M, Charrier K, et al. RANK is essential for os-teoclast and lymph node development. Genes Dev. 1999;13(18):2412–2424.
33. Kim D, Mebius RE, MacMicking JD, et al. Regulation of peripheral lymph node genesis by the tumor necrosis factor family member TRANCE. J Exp Med. 2000;192(10):1467–1478.
34. Kim N, Odgren PR, Kim DK, , et al. . Diverse roles of the tumor necrosis factor family member TRANCE in skeletal physiology revealed by TRANCE deficiency and partial rescue by a lymphocyte- expressed TRANCE transgene. PNAS. 2000;97(20):10905–10910.
35. Kong YY, Yoshida H, Sarosi I, et al. OPGL is a key regulator of osteo-clastogenesis, lymphocyte development and lymph-node organogenesis. Nature. 1999;397(6717):315–323.
36. Akiyama T, Shinzawa M, Akiyama N. RANKL-RANK interaction in immune regulatory systems. World J Orthop. 2012;3(9):142–150.
37. Pacifici R. T cells, osteoblasts, and osteocytes: interacting lineages key for the bone anabolic and catabolic activities of parathyroid hormone. Ann N Y Acad Sci. 2016;1364(1):11–24.
38. Sato K, Suematsu A, Okamoto K, et al. Th17 functions as an osteoclasto-genic helper T cell subset that links T cell activation and bone destruction. J Exp Med. 2006;203(12):2673–2682.
39. Polley H, Hunder G. Rheumatologic Interviewing and Physical Examination of the Joints. 2nd ed. Toronto: W.B. Saunders Company; 1978.
40. Salva JE, Merrill AE. Signaling networks in joint development. Dev Dyn. 2017;246(4):262–274.
41. Decker RS. Articular cartilage and joint development from embryogenesis to adulthood. Semin Cell Dev Biol. 2017;62:50–56.
42. Hui AY, McCarty WJ, Masuda K, Firestein GS, Sah RL. A systems biology approach to synovial joint lubrication in health, injury, and disease. Wiley Interdiscip Rev Syst Biol Med. 2012;4(1):15–37.
43. Fearon U, Hanlon MM, Wade SM, Fletcher JM. Altered metabolic path-ways regulate synovial inflammation in rheumatoid arthritis. Clin Exp Immunol. 2019;197(2):170–180.
44. Das S, Banquy X, Zappone B, et al. Synergistic interactions between graft-ed hyaluronic acid and lubricin provide enhanced wear protection and lubrication. Biomacromolecules. 2013;14(5):1669–1677.
45. Jay GD, Lane BP, Sokoloff L. Characterization of a bovine synovial fluid lubricating factor. III. The interaction with hyaluronic acid. Connect Tissue Res. 1992;28(4):245–255.
46. Cummings NA, Nordby GL. Measurement of synovial fluid pH in normal and arthritic knees. Arthritis Rheum. 1966;9(1):47–56.
47. Hamerman D, Schuster H. Hyaluronate in normal human synovial fluid. J Clin Invest. 1958;37(1):57–64.
48. Johnston JP. The viscosity of normal and pathological human synovial fluids. Biochem J. 1955;59(4):633–637.
49. Ropes MW, Muller AF, Bauer W. The entrance of glucose and other sugars into joints. Arthritis Rheum. 1960;3:496–514.
50. Ropes MW, Rossmeisl EC, Bauer W. The origin and nature of normal human synovial fluid. J Clin Invest. 1940;19(6):795–799.
51. Cho MH, Neuhaus OW. Absence of blood clotting substances from syno-vial fluid. Thromb Diath Haemorrh. 1960;5:108–111.
52. Poole AR, Kojima T, Yasuda T, Mwale F, Kobayashi M, Laverty S. Com-position and structure of articular cartilage: a template for tissue repair. Clin Orthop Relat Res. 2001:(391 Supp)S26–S33.
53. Gylys-Morin VM, Graham TB, Blebea JS, et al. Knee in early juvenile rheuma-toid arthritis: MR imaging findings. Radiology. 2001;220(3):696–706.
54. Laor T, Jaramillo D. MR imaging insights into skeletal maturation: what is normal? Radiology. 2009;250(1):28–38.
55. Zhang W, Ge Y, Cheng Q, et al. Decorin is a pivotal effector in the extracellular matrix and tumour microenvironment. Oncotarget. 2018;9(4):5480–5491.
56. Petty R. Structure and function. In: Petty R, Laxer R, Lindsley C, Wedderburn L, eds. Textbook of Pediatric Rheumatology. Philadelphia: Elsevier; 2016:1–13.
11.e2 References
57. Rengel Y, Ospelt C, Gay S. Proteinases in the joint: clinical relevance of proteinases in joint destruction. Arthritis Res Ther. 2007;9(5):221.
58. Verma RP, Hansch C. Matrix metalloproteinases (MMPs): chemical- biological functions and (Q)SARs. Bioorg Med Chem. 2007;15(6):2223–2268.
59. Waghray N, Jyothi A, Imran M, et al. Enthesis: a brief review. Apollo Med. 2015;12:32–38.
60. Apostolakos J, Durant TJ, Dwyer CR, et al. The enthesis: a review of the tendon-to-bone insertion. Muscles Ligaments Tendons J. 2014;4(3):333–342.
61. Lories RJ, McInnes IB. Primed for inflammation: enthesis-resident T cells,. Nat Med. 2012;18(7):1018–1019.
62. Sherlock JP, Joyce-Shaikh B, Turner SP, et al. IL-23 induces spon-dyloarthropathy by acting on ROR-γt+ CD3+CD4-CD8- entheseal resident T cells. Nat Med. 2012;18:1069–1076.