ALEMAYA UNIVERSITY OE AGRICULTURE SO HO Oil, OF GRAIDUWTE STUDIES RECEIVED Date Month Year — ALEMAVA UNIVERSITY OF AGRICULTURE LIBRARY YIELD AND YIELD COMPONENTS OF ETHIOPIAN MUSTARD (BRASSICA CARINATA A. BRAUN) 7 * AND RAPESEED (BRASSICA NAPUS L.) AS AFFECTED BY SOME AGRONOMIC PRACTICES NIGUSSIE ALEMAYEHU JULY, 1990
Transcript
ALEMAYA UNIVERSITY OE AGRICULTURE
SO HO Oil, OF GRAIDUWTE STUDIES
R E C E I V E D
Date Month Year —
ALEM AVA UNIVERSITY OF
AG RICULTURE LIBRARY
YIELD A N D YIELD COMPONENTS OF ETHIOPIAN M U S T A R D (BRASSICA CARINATA A. BRAUN)
7 * A N DRAPESEED (BRASSICA NAPUS L.)
AS AFFECTED B Y SOME AGRONOMIC PRACTICES
NIGUSSIE ALEMAYEHU
JULY, 1990
YIELD AND YIELD COMPONENTS OF ETHIOPIAN
MUSTARD (BRASSICA C AR IN ATA A. BRAUN)#
AND
RAPESEED (BRASSICA NAPUS L.)AS AFFECTED BY SOME AGRONOMIC
PRACTICES
H AR AM AYA UNIVERSITY LIBRARY
300028030
A Thesis
Presented to the School of Graduate Studies of Alemaya University of Agriculture
In Partial Fulfillment of the Requirements for the Degree of Master of Science in Agriculture (Agronomy)
by
Nigussie Alemayehu July 1990
ALEMAYA UNIVERSITY OF ACRICULTURETHE HRADUATE SCHOOL
Title of Thesis:Yield and Yield Components of Ethiopian mustard (Brassica carinata B.)
The author was born in Oct. 1962 from his mother Weizero Mulunesh Woldegiorgis and father Ato Alemayehu Asfaw at Koffele, 275 km south of Addis Ababa, in Arsi Administrative Region.
When he was 8, he began his elementary school at Koffele Elementary and Junior High School in 1970 and was schooled there until he completed8th grade in 1976. Hethen shifted his residence toAssela, the capital of Arsi, topursue his high school education at Assela Comprehensive Secondary School. During his four-years stay at this school, he was educated under the stream of
Agriculture which probably induced a keen interest in him to* proceed with agriculture later in his university life.
After completing high schoolwith a Diploma inAgriculture and passing the Ethiopian School Leaving Certificate Examination <ESLCE ) in 1980, he joined thethen Alemaya College of Agriculture under the auspice of Addis Ababa University.
In July 1984,he graduated with a B.Sc. in Agriculture (Plant Sciences) and was recruited by the Institute of Agricultural Research (IAR), During the next two and half-years (1984 - May 1987) of his career, he served as anassistant Research Officer in the Highland Oil Crops Improvement Project under Field Crops Division at Holetta AgriculturalResearch Center (HRC). Thenin May 1987, he wastransferred to the Agronomy/Phvsiology Divisionat HRC with the same position to handle the agronomic and physiologic research in highland Oil Crops (rapeseed, mustard, linseed and
niger seed), and worked there until September of the same year.
In September 1987, he joined the Graduate School of Alemaya University of Agriculture under the sponsorship of the International Development Research Center (IDRC) of Canada. He graduated in July 1990 with an M.Sc. in Agronomy. Heis fervent to go back to IAR and assume research in Oilseeds.
ACKNOWLEDGEMENTS
I embarked on the thesis research project under the advisor-
ship of the late Dr. Hiruy Belayneh, Institute of Agricultural
Research (IAR). Dr. Hiruy was involved right from the initiation and preparation of the thesis proposal, and provided unreserved assistance all the way through up until the analyses of the data. Thus, his participation in the execution and follow-up of the experiment was invaluable. Unfortunately, however, Dr. Hiruy did not live long enough to witness the write-up of this thesis. His untimely death has touched me most, but to him I extend to highest tribute. As a token of such an appreciate, I dedicate the thesis to his memory.
$ *
Despite other mutually exclusive commitments, my co-advisor, Dr. Mesfin Abebe of Alemaya University of Agriculture (AUA) assumed full responsibility when my major advisor passed away. As a consequence he took a pain to critically reviewed and correct the draft thesis to give it the present shape. I am indebted to Dr. Mesfin's indispensable assistance and advice.
My M.Sc. training program was supported by the International Development Research Center (IDRC) of Canada, to whom I express my deep gratitude. Egually, the academic leave of absence accorded by the IAR, the assistance rendered by the Holetta Agricultural
i v
i acknowledge with heart-feit thanks the enthusiastic support and encouragement I received from: Ato Getinet Aiemaw, Ato \itbarek
Semeane, Ato Amsal Tarekegn, Ato Adefris Teklewold, Ato Tadesse Deme, Ato Mengistu Negi and Ato Tadesse Debele of the IAR.
I am highly grateful to Dr. Abbas O.Omran, Oil Seeds Network Coordinator of IDRC for his valuable comments and suggestions pertaining to statistical analyses; and his staff who provided me with unreserved technical assistance in the preparation phase of
the manuscript.
My thanks are also due to W/t Elizabeth Yohannes and W/t
Elizabeth Baslyos for the final preparation of the manuscript, without whose untiring and cheerful assistance, this thesis wouldn’t have seen the light of day at the time it did.
The continuous encouragement and moral support of my familyand friends during my Master’s program at AUA is indeed cherishable and will be remembered with appreciation.
TABLE OF CONTENTS
BIOGRAPHICAL SKETCH ii
ACKNOWLEDGEMENTS ivDEDICATION vi
LIST OF TABLES X
LIST OF FIGURES xiLIST OF MAPS xv
ABSTRACT xvi ,I . INTRODUCTION 1II. REVIEW OF LITERATURE - 8
2.1 Effects of the factors considered onAgronomic characters 8
2.1.1 Species 8 *2.1.2 Date of sowing y ?2.1.3 Method of sowing 1 1
2.1.4 Fertilizer - 122.1.5 Weed control 1 3
2.1.6 Interactions of factors 13
2.2 Effects of the factors considered on • Seed yield, Oil content and Oil yield 14
III MATERIALS AND METHODS 23t \3.1 Experimental Sites 233.2 Experimental Design and Treatments 24
3.3 Parameters Collected and Methods of Collection 26IV. RESULT AND DISCUSSION 3 0
4.1 Effects of the Factors Considered on
TABLE OF CONTENT CONTD. Page
Agronomic Characters 304.1.1 Species 304.1.2 Date of sowing 364.1.3 Method of sowing ft . 414.1.4 Fertilizer 444.1.5 Weed Control 474.1.6 Interactions of factors Effects of the Factors considered on Seed
50
yield , Oil content and Oil yield 754.2.1 Species 754.2.2 Date of sowing 774.2.3 Method of sowing 794.2.4 Fertilizer 804.2.5 Weed control 814.2.6 Interactions of factors '8 3
viii
TABLE OF CONTENT CONTD. : Fage4.3 Correlation of characters 96
V. SUMMARY AND CONCLUSION 103VI. RECOMMENDATION 110
i
VII. BIBLIOGRAPHY 112VIII. APPENDIX 129
ix
LIST OF TABLES IN TEXT
Table 1
Table 2
Table 3
Table 4
Agronomic characters as affected by- species, sowing date, sowing method, fertilizer and weed control' at Holetta
and Kulumsa.
Seed yield, oil content and oil yield as
affected by species, sowing date, sowing method, fertilizer and weed control at Holetta and Kulumsa.
Simple correlation coefficients between yield components of oil seed brassica at Holetta and Kulumsa.
Path-coefficient analyses showing the direct and indirect effects of four characters on seed yield at Holetta and Kulumsa. *
Page
31
76 -
*
97
101
x
LIST OF FIGURES IN TEXT
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Figure 6
Figure 7
Effect of the interactions between species andsowing date on plant height at Holetta and Kulumsa. 51
Effect of the interactions between species, sowing date and fertilizer on number of
secondary branches per plant at Kulumsa. 51
Effect of the interactions between sowing dateand fertilizer on stand at Holetta and Kulumsa. 54
Effect of the interaction between sowing dateand fertilizer on plant height at Holetta. 54 f*
Effect of the interaction between species,sowing date and method of sowing on seedlingdensity at Holetta and Kulumsa. 54
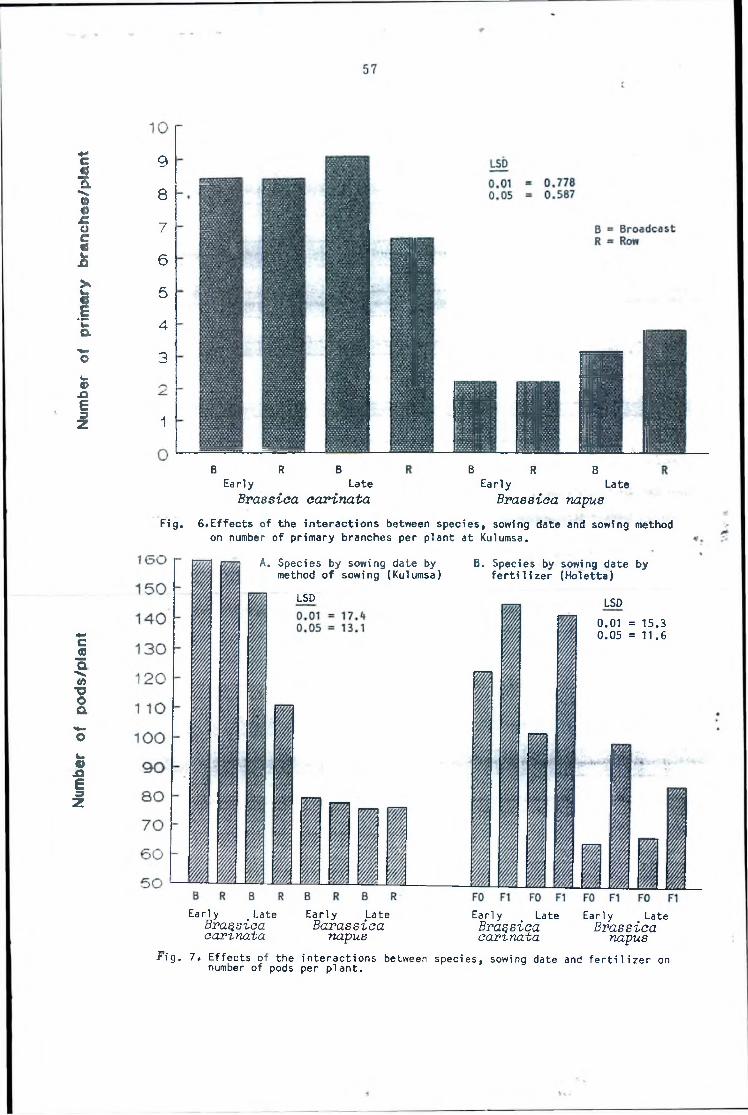
Effect of the interactions between species,sowing date and method of sowing on number ofprimary branches per plant at Kulumsa. 57Effect of the interactions between species,sowing date and fertilizer on number of podsper plant at Holetta and Kulumsa. 57
xi
Figure
Figure
Figure
Figure
Figure
Figure
LIST 0? FIGURES IN TEXT (Contrd)gage
8 Effects of the interactions between species, sowing date, sowing method and fertilizer on 1000-seed seed weight at Holetta. 60
9 Effect of the interaction between species, sowing date and fertilizer on number ofprimary branches per plant at Holetta and Kulumsa. 60
10 Effects of the interaction between species, sowing date and fertilizer on
days to flowering at Holetta and Kulumsa. 64
11 Effects of the interactions sowing date, fertilizer and days to maturity at Holetta
between species,weed control onand Kulumsa. 64
12 Effects of the interactions between species, sowing date and fertilizer on number ofseeds per plant at Holetta and Kulumsa. 67
13 Effects of the interactions betweensowing date, fertilizer and weed controlon number of pods per plant at Kulumsa. 67
xii
LIST OF FIGURES IN TEXT (Cant’d)
Figure 14 Effects of the interactions between
sowing date, fertilizer and weed controlon number of seeds per plant at Holetta andKulumsa. 73
Figure 15 Effects of the interactions between
species, sowing date and fertilizeron oil content at Holetta and Kulumsa. 84
Figure 16 Effects of the interactions betweenspecies and fertilizer on oil yield at Holettaand Kulumsa. 8 4
Figure 17 Effect of the interactions between
species, sowing date, fertilizer andweed control on seed yield at Kulumsa. 88
Figure 18 Effects of the interactions between
species method of sowing, fertilizer and weed control on seed yield at Holetta and Kulumsa. 88
Figure 19 Effects of the interactions betweensowing date, fertilizer and weed controlon seed yield at Holetta and Kulumsa. 94
xiii
LIST OF HAPS
PageMap 1. Rapeseed/mustard growing areas in Ethiopia
xv
LIST OP FIGURES IN TEXT (Contfd)
Figure 20 Effect of the interactions between
sowing date, fertilizer and wefed control
on oil yield at Holetta and Kulumsa.
Figures 21 A (Holetta) and B (Kulumsa) Path-coefficientdiagrams.
Page
94
100
xi v
YIELD AND YIELD COMPONENTS OF ETHIOPIAN MUSTARD (BRASSICA
CARINATA A. BRAUN) AND RAPESEED (BRASSICA NAPUS L.) AS AFFECTEDBY SOME AGRONOMIC PRACTICES
ByNigussie Alemayehu, B.Sc (Alemaya College of Agriculture)Major Advisor - Hiruy Belayneh, Ph.D. (Washington State
Since antiquity, Ethiopian mustard (B. carinata A. Braun) has
been cultivated for array purposes, mainly in the Centeral, South- Eastern and North-Western highlands of Ethiopia. While Argentine rape seed (B. napus L), is a recent introduction, mostly in State
Farms of Arsi and Bale regions.Currently Ethiopian mustard and the Argentine rape seed
together constitute the third most important oleiferous crops in
Ethiopia both interms of area and production, next only to niger (Guizotia abyssinica Cass.) and linseed (Linum usitatisimum L.).
Yet, the national average yield remained much below the world average record leading to an acute shortage of edible oil at the face of ever increasing demand. This low productivity is attributed by and large to miss-management of the crop and lack of appropriate packages of agronomical technology.
Although piecemeal research of the past decade has revealed
xv i
that considerable amount of loss could occur due to lack of
optimum: date of sowing, method of sowing, fertilizer application and weed control, the relative importance and the interactive
effects of these factors on the yield and yield components of oleiferous brassica is virtually absent in Ethiopia. Therefore, the paucity of technical information on such a vital crop has instigated this experiment.
The trial was conceived with 32 combinations of five
management factors each at two levels namely, species, sowing
dates, sowing methods, fertilizers and weed control, laid out in an incomplete block design in which the highest order interaction was completely confounded. The experiment was conducted in fourfold replication at Holetta; (38° E. Long, and 9° N. Lat.), and Kulumsa; (39° E. Long, and 8° N. Lat. ) in the main cropping season of 1988/89,
At Holetta, fertilizer > sowing date > species > weed control > method of sowing were found to be sequentially important while at Kulumsa, the sequential significance of the factors were: species> sowing date > fertilizer > weed control > method of
sowing. The correlation and path-coefficient analyses revealed that plant height and number of pods per plant are the yield components that exert the greatest influence for seed and oil yields in oleiferous brassica*
The main and interactive effects of the various management
factors on the different agronomic characters, seed yield, oil / content and oil yield; and the relationships between the component
xv ii
parts and seed yield of oil seed brassica indicate that in most cases, the introduced B. napus cultivar, (Tower sel3) „as more
responsive to a better level of a given management factor when
grown under conditions of optimum level of the other factors while the indigenous B. carinata cultivar (S-67) responded tremendously with only slight intervention of one or two management factors.
X V I 1 1
I INTRODUCTION
The genus Brassica of the Brassicaceae family as a whole is1
believed to have originated around the Mediterranean, Eastern
Afghanistan and the adjoining portion of Pakistan and North-Eastern Africa (Hemingway, 1976, McNaughton, 1976 a,b). The genus isprobably one of the most widespread plant genera containing species that have evolved through natural interspecific hybridization and the establishment of the hybrids as full-fledged independent
yspecies of both cultivated and possibly wild forms. Many
investigators, including U (1935), Nwankiti (1970), Appelqvist, eta l . . ( 1972), McNaughton ( 1973 ), Prakash ( 1973, 1974), Gowers
i(1974), Hemingway (1976 ), Annand, et a l . , (1980), Prakash, e t .a l « . ( 1983) and Fan, et a l . , (1985) have recognized that there arethree basic diploid species; B. nigra Koch. (black mustard), B.
oleracea L. (cabbages) and B. campestris L. (turnip rape) with chromosome (n) numbers of 8, 9 and 10, respectively.
Chromosome doubling susbsquent to hybridization gave rise to three amphidiploid species namely. B. napus L. (rape seed) which is the result of a natural cross between B. oleracea and B.
campestris; B . carinata A. Braun (Ethiopian mustard) a cross between B. oleracea and B. nigra and B. Juncea (L.) Czern and Coss (brown or Indian mustard) a cross between B. campestris and B.
nigra.
The three amphidiploid and B. campestris are the leading oleiferous Brassica spp. Downey ( 1983 ) and Sing, et aJL., (1984)
indicated that B. napus and B. campestris predominate in the
temperate regions while in the semi-tropics of Asia B. campestris
and B. juncea are of major significance.
AccFoding to Holmes (1980), Stringham, et a l ., (1980) and
Prakash, et al . , (1983), oil has been extracted from rape (B .napus) and other oilseed brassicas in Europe at least since the
fifteenth century and for a much longer (perhaps about two millenia B.C.) in India and probably China. -?Downey (1971), Appelqvit, et al ., (1972) and Holmes (1980) indicated that oil is now producedfrom rapes and mustards in every continent. Such worldwide proliferation has led to substantial hectarage being devoted to
oilseed brassica. As a case in point, the 1986 FAO sensus estimated a total acreage of well over 145 million hectars with a production of about 20 million metric tons. Over half the area is * in Asia wherein China and India, which stand first and second globally, constitute more than three quarter of the acreage. Canada, Bangladesh, Pakistan and Northern Europe (France, Poland, West Germany, Norwey and Sweden) in aggregate stand third.
The fruit of rape seed or mustard is a silique which contains 10-45 seeds. The oil present in the embryo represents about 38- 45% of the seed dry weight and is a good cooking or salad oil. This is particularly true for cultivars with low level of the long Chain 22-carbon monoenoic acid-erucic acid.
Subsequent to oil extraction, the meal which is protein rich (30-45%) can be used either as a high-protein feed supplement or as an organic fertilizer. Industrial uses include: leathertanning, the manufacture of varnishes, soap, and lamps (Vaughan,
2
*r
1970; Downey, 1971; Appelqvist, et al., 1972; Bhan, 1979; Downey,
1983 ) . Bettis et a 1 • » ( 1982 ) have even gone further to suggest the immense potential of rape oil as a substitute for diesel feul.
Ethiopian mustard (B. carinata) and rape seed (S. napus) are
the only oleiferous brassicas currently under production in%
Ethiopia. Mustard and rape seed, in Ethiopia, occupy well over 50
thousand hectares which renders them the third important oilseed crops following niger ( Guizotia abyssinica Cass.) and linseed (Linum usitatisimum L.) (Central Statistics Authority, 1987). According to Seegeler (1983) and Hiruy (1983), Ethiopian mustard and rape seed have been cultivated primarily in the 500-1200 mm
annual rainfall belt in mid-and high- altitudes ranging between 1700 and 2800 m a.s.l.
~Since antiquity, mustard has been exclusively grown by farmers with small holding mainly in what used to be Gojam, Arsi, Sidamo, Western Welega and Centeral and Southern Shoa (Map 1). It occupies some 43 thousand hectares and is cultivated either as a
* 1sole crop or intercropped with cereals notably with maize and sorghum.’ Rape seed, on the other hand, is a recent introduction and is grown mainly on State Farms at Arsi (Lole, Gofar and Dixis) and Bale (Serofta, Sinana and Herero). The hectareage of rape seed, on State Farms has increased from 207 ha in 1982 to 7299 ha in 1988, an increase of over 3000% (Hiruy, 1989, unpublished).
i Ethiopian mustard (B. carinata) is often grown on well drained and organic matter-rich soils close to homesteads. The crop is cultivated primarily for its culinary value. Boiled mustard leaves
3
are utilized as "cabbage", "gomen wot"- a kind of stew; crushed seeds are often used to grease the clay baking pan ( "mitad" ) beforeIthe local bread ("injera” ) is baked. Some of its preparations are traditionally used as component of spices and condiments as well as a cure to minor ailments.
Much of the produce from the State Farms (rape) contributes
to the limited agricultural export commodities. The industrial value of the crop is indeed immense and not fully explored.
Mustard and rape seed are often tolerant to the major diseases and pests that attack cereals. Thus, they can be included in the crop rotation scheme to break disease or pest cycles in the production of cereals.
According to Hiruy ( 1983 ) and FAO (1986), the average nationa.1 ?
yield is very low, only four to five q/ha as compared to the world
average of 13-14 q/ha. The reasons for this predicament will be elucidated further but the fact remains that there is an accute shortage of edible oil at the face of ever increasing demand; because the average oil consumption in Ethiopia, as Hiruy (1983) reported it, is in the order of about one liter per person per year. Of this the share from brassica, assuming the average oil content to be 40% and specific gravity of the oil 0.91 (Appelqvist, et al., 1972) and taking in to account the average national yield and total acreage in the country, is between 0.20 and 0.24 1/personper year.
(Research results of the past decade (Mid 70’s-Mid 80Js), on
the other hand, reveal that the potential of the crop if cultivated
4
under proper sowing date, sowing method, fertilizer application and weed control is well over 30q/ha (Hiruy, gjt al. , 1987) which then
increases the above share to 1.46 1/person per year,i.e 5 times more than the maximum possible under the current situation. Ethiopian farmers, however, seldom weed their brassica fields, nor do they sow the crops at the optimum time. This is partly due to the fact that by the time farmers ought to carry out such agronomic practices for brassica, the cereals which fetch better market prices, are ready for similar operations. Such mutualexclusiveness interms of operation calendar and the priorities farmers give to cereals leads to competition for labour and has made the oil seeds victims to the fates of delayed sowing and weed
competition. Equally, farmers very rarely apply fertilizers to oil seed brassicas and for that matter to other oilseed crops. Although piecemeal research of the past ten years (Hiruy, et a l .« 1986 ) has revealed that considerable amount of loss could occur due to the lack of observance of optimum sowing date, sowing method,
judicious fertilizer application and effective weed control, the relative importance and the interactive effects of these factors on the performance of Ethiopian mustard is scanty in the world at large and virtually absent in the specific case of Ethiopia. Therefore, the paucity of technical information on such a vital crop in the socio-economic life of the Ethiopian people has instigated the need for research to address itself to this information gap and hence this experiment was conceived with the following specific objectives as to:
5
quantify the relative importance of sowing date, sowing method, fertilizer and weed control on the seed and oil
yields of Ethiopian mustard (Brassica carinata) and rape seed (B . napus);
determine the interactive effects of these major
agronomic practices on the yield and yield components of the crop and thereby fine-tune the agronomic package of brassica as a whole;
elucidate the relationships between the different yield components of the crops which offer a basis to explain the difference between genotypes and management practices in line with improvement programs; and
finally formulate a set of recommendation^) for possiblea
compatible interventions from the interaction of the factors that will enhance productivity and thus narrow the current gap between the yields obtained by farmers and researchers.
7
II. REVIEW OF LITERATURE
2.1. Effects of the considered Factory
on Agronomic characters The factors under consideration include: species, date of
sowing, method of sowing, fertilizer application and weed control practices and hence the review of the relevant literature on each specific factor is inhere provided as a prelude to the elucidation of the subsequent experimental findings.
2.1.1. SpeciesIt is recognized that the mustard ( B. carinata) cultivars have
longer growth duration than rape seed cultivars (£.' napus)
(Mizushima, et al. , 1967; Chauhan, et al.. , 1979, 1984 ). Thevariation, between species or genotypes of the same species of oilseed brassica, in plant height, number of primary and secondary branches, days to maturity and number of seeds per pod are indeed tremendous (Kaul, et aJL., 1986 ). The authors also noted that
number of pods per plant and days to flowering also vary greatly with species of oil seed brassicas. Paul, et al., (1976) found similar results including 1000-seed weight but with pronounced variations among the genotypes of B. juncea, under Ethiopian conditions, Hiruy, et slL*> ( 1986 ) and Hiruy (1987 ) found that cultivars of B . napus are earlier in flowering and maturity, shorter in height, more compact with reduced branching characteristics than those of B. carinata.
8
9
2.1.2. Date of sowing
There is a.wealth of information, some as yet not resolved controversies, on the ranges of effects that sowing dates have pertaining to both the pre-and post so called "optimum” date of sowing established for specific localities. Many of the workers found different manifestations in plant characters emanating from differences in dates of sowing.
Fulkerson (1971) and Mendham, et al., (1981 a), reported that early dates of sowing produced significantly taller plants. Auld, et al.,(1984), however, noted that plant height was not sensitive
to sowing date. Similar results have also been reported by Degnehardt, et ajL . , (1981 a). with early sowing, Mendham, et a l . ,
* * >( 1975 ) found very late plants of rape seed with an enormous number >of flowers and a dense canopy containing almost twice as many sitesfor pod development as existed in the late sown ones. Early sowingproduced consistently more seeds per plant with a far greater rangethan late sowings (Mendham, et M * . 1981 a). Further, early sowingproduced fewer seeds per pod in B . napus and they were supportedby the works of Thurling ( 19 74 b) and Mendham, et a l . , ( 1975 ).Degenhardt, et a l ., (1981 a), on the other hand, found that sowingdate had no significant effect on number of seeds per plant. They
/reported little change in seed per pod in B, napus with successive delays in seeding.
It appears that there are numerous conflicting findings vhich by virtue of controversies need further in-depth research to
resolve the contradictions and bring about reproducible and
definitive concepts on the effect of early sowing on the character/of oil seed brassicas.
According to Degnehardt, et jlL* , (1981 a), delayed seedingresulted in a significant decrease in seedling density, because" earlier sowing dates provided warmer soil conditions, better moisture regimes and, therefore, resulted in better germination and more number of seedlings. " This is supported by the work of
Pretty, et, aX* > ( 1976 ) who also noted that stress, caused bydelayed sowing during germination generally results in reduced seedling density. Yet, Christensen, et al., (1985), found that later sowing resulted in fewer seedling density reduction of B.
campestris and no visible trend with B. napus concluding that
seedling density did not appear to be related to date of sowing.A highly significant effect of sowing date on the flowering
and maturity dates of rape seed was reported by Degnehardt, et a l . , (1981 a). They found that a delay of 28 days resulted in five-day reduction in the time required to reach maturity of first pod and a seven-day reduction in the time required to reach last flower.
The authors, however, pointed out that delayed sowing resulted in no significant difference in the seed formation period indicating that compensation for delayed sowing occured through a reduction in time to growth stages upto the end of flowering of first pod. Trends of decline in days to maturity of rape seed (B . napus) as a result of advancement of sowing date beyond the optimum were
reported by Kondra ( 1976 ) and Christensen, et al. , (1985).
10
11 1
According to the latter, however, further excessive delays increased days to maturity.
In B. napusf Thurling (1974 b) found that number of primary and secondary branches decreased with each successive delay in
sowing contrary to the finding of Degnehardt, et al. , (1981 a)indicating the need for further research to resolve this controversy.
Successive delays in sowing rape seed are reported to have a marked reduction in the number of pods per plant (Thurling 1974 b;Holmes 1980; Degnehardt, et a l . , 1981 a). Conversely, Mendham, et
\
a l . , (1981 a) found excessive number of pods with early sowings,while delayed sowing restricted the number of pods a crop could carry to final harvest.
Thurling (1974 b) and Ali~Khan (1976) found a decrease in* 1000-seed weight with l&te sowing dates. Degnehardt, et al., (1981
a) and Hiruy, ejt al., (1983), however, reported no significant effect of sowing date on 1000-seed weight similar to Mendham , et al. , (1981 a) who in different years, found inconsistent andconflicting results in seed weight with delayed sowing.
2.1.3. Method of sowing
Comparing the effects of methods of sowing in rape seed, Clarke, et al., (1978) obtained comparable plant stands for the broadcast and drilled plots. The broadcast-seeded had more branches and pods per plant than drill seeded material. Seeding
method, however, did not affect the number of seeds per pod and
1000-seed weight. However, broadcast sown rape seed tend to be taller than the row seeded«(Fulkerson, 1971). On niger (Guizotia
abyssinica Cass.), Patil (1979) found higher number of buds
(capitula) per plant with row than broadcast sowing.
2.1.4. Fertilizer
Tremendous responses to fertilizers have been reported by various workers. According to Christensen, et al., (1985) nitrogen and phosphorus resulted in delayed maturity of rape seed but were not considered agronomically important. Mendham, et al. , (1981 b) noted slightly advanced time of flower initiation with the application of nitrogen and the height of individual plants was approximately doubled similar to the work of Sheppard, et- al. ,(1980 ) .
According to Olsson (1960) and Forbes (1985 b), fertilizer application resulted in increased growth and number of primary branches per plant. The latter, further noted an increased number of secondary branches, in agreement with Joarder (1983),
Higher levels of nitrogen led to the development of increased number of pods per plant and seeds per pod (Olsson, I960; Allen, et al. , 1971). While Olsson (1960) and Mendham, ejt aj,. , (1981 b) found higher number of seeds per plant and heavier seeds.
12
13
2.1.5. Weed controlt
A reduction in available nutrients, light, and moisture duei
to weed competition would be expected to restrict the growth of pod bearing branches in rape seed with a resultant effect of less pods per plant (Forbes, 1985 b). Jain, et a l ., (1979), Kaul, et
a l ., (1986) and Kumar, et a l ., (1989) emphasized that the vigor of rape seed and mustard, including height and pod bearing branches can considerably be reduced from competition with weeds with an ultimate effect of delayed maturity.
2.1.6. Interaction of factors*
Bhan (1979) signified the interactive effects of sowing date and fertilizer application on mustard. Improved growth of plants as measured by plant height, number of primary and secondary branches and yield attributes namely, pods per plant, seeds per pod and 1000-seed weight may be held responsible for increased yield of mustard due to proper time of sowing and fertilizer application. The positive effects reflected in the overall vigor of rape seed and mustard, which are expected from early sowing could be off-set by lack of provision of timely weed control (Jain, et al., 1979). Inferences can also be drawn from the work of Staniforth (1958) who found a significant interactive effect between date of sowing and weed control on the growth of soybean (Glycin max (L.) Merril),
Auld, et a l ., (1984) emphasized the importance of the interaction
of sowing date and cultivar in the general growth of rape seed.Pretty, et a l ., (1976) have also found a considerable amount oft
interactive effects between date of sowing and fertilizer on seedling density, stand and yield attributes of rape seed.
2.2 Effects of the Considered Factors on Seed Yield. Oil Content and Oil Yield
2.2 .1. Species
Interms of seed yield and oil content, it is recognized that there are tremendous variations among and within the species of Brassica. Mizushima, et al, , ( 1967) have noted considerably higher yield in B. carinata than in B, napus. Hiruy, et al.. , (1986) and"* %
Hiruy (1987) reported higher seed yield, lower oil content and higher oil yield in B. carinata than in B. napus under Ethiopian conditions. Paul, et al., (1976), Uppstrom (1977), Astahana, et
a l . , (1979) and Clarke (1979) working with different species ofoilseed brassica found that the variation in seed, oil yields, and oil content, between genotypes of the same and different species were immense.
2.2.2. Date of sowing
Moss, et al. , (1971), Pretty, et al. , ( 197 6) , Synder, et al, , (1978) and Tayo, et a/L. , (1979) underlined the importance of sowing
14
date because optimum date of sowing takes advantage of light and
temperature regimes that best fit the optimum photosynthetic period of the crop such that the period between germination and maturity
is fully utilized. Otherwise, delayed seeding is recognized to
result in a significant decrease in seed yield of oil seed brassicas. Therefore, seed yield would be greatest in the earlier seeding where the period from sowing to the first flower is much longer than in subsequent sowings (Ali-Khan, 1976: Kondra, 1976; Degnehardt, et a l . , 1981 a; Auld, et a l . , 1984; Christensen, et
al., 1985).In specific terms, Thurling ( 1974 a ), Mendham, et al. , (1975 ) ,
T a y o , et a l . , ( 1975 ) Degnehardt, et al . , (1981 b) and Mendham, et,al., (1981 b) found a highly significant and continuous decline in seed yield of B. napus with delayed sowing. The authors emphasized., leaf area as a cause whereby delayed sowing having smaller than apparent critical size plants at flower initiation were unable to produce the maximum leaf area and hence had lower oil and seed yield. Equally, Gross, et a l . , (1966), Kondra (1976) andChristensen, et al. , (1985 ) found a significant decrease in oilcontent and yield of B. napus with late sowing and a peak with the optimum sowing date. On the other hand, Mendham, et aJL. , (1981 a) and Auld, et. _al . , (1984) observed that oil content was notsignificantly affected by sowing date. Hiruy, et al. ( 1983), in Ethiopia, found that with late sowing yields of both B. napus and B. carinata were greatly reduced. In essence, piecemeal findings
15 1
have indicated that there are potential seed yield losses of 23%
from delayed sowing of brassica (Hiruy, et al_. , 1987).
2.2.3. Method of sowing
Fulkerson (1971) found that higher yields have been produced by B. napus under the row than the broadcast method of sowing. Kondra (1975), Clarke, et al., (1978), Jain, et aJ., (1979) and
Anonymous (1981, 1985) reported increased seed yield and nosignificant oil content in rape seed with sowing in the row. Conversely, Helps (1971) and Kaul, et al., (1986), found broadcast over row seedings of rape seed and mustard resulted in higher seed yield but showed no difference in oil content.
2.2.4. Fertilizer
A great deal of research on the fertilizer requirements of oilseed brassicas has made it clear that the response of these crops is indeed tremendous. The oilseed brassicas have a high requirement for N and P, more than is provided by most soils and generous use of these fertilizers is, therefore, necessary for optimum yield (Allen, et al ., 1971). Similarly, Ractz, et a l .,(1965 ) , Krogman, et a l . , (1975 ) , Sheppard, et al., (1980 ) , Aulakh, et ajL. , (1980), Joarder (1983) and Nuttal, et a_l. , (1987) foundthat nitrogen and phosphorus increased seed yield of rape seed.
On the importance of nitrogen and phosphorus to rape seed and
16
other oilseed brassicas, Holmes (1980) states "increasing the
nitrogen supply increases the amount of noted that protein formed and, therefore, the amount of protoplasm. ... the over all eff-ect is to increase crop growth dramatically. ... and to provide, in
oilseed brassica, a large frame on which more flowers and eventually more pods can develop. ... phosphorus is a constituent of nucleic acids and nucleoproteins, organic phosphorus compounds are involved in energy transfer reactions and in respiration. Thus, the importance of this element for seed production is far
from doubt."Different results have been reported by various workers, on
the effect of fertilizer on oil content of oilseed brassica. Dybing (1964), Krogman, et. al. , (1975 ). On the other hand, Henery, et a l . , ( 1978) and Joarder ( 1983 ) noted that nitrogen lowered the'1oil content of rape seed significantly while Aulakh, et al. , (1980) reported little effect on oil percentage of B. campestris and B . juncea but the total oil yield increased many fold with N- application due to increased seed yield. Conversely, a marked reduction in oil concentration in rape seed of upto 4-5% was found by other workers from N-application (Olsson, 1960; Nuttal, et. a l . , 1987; Sheppard, et al. , 1980). The same authors, however, notedthat phosphorus did not affect oil content. Under Ethiopian
\
conditions, Hiruy ( 1985 ) Taye (1986) and Hiruy, et ajL. , ( 1987 ).emphasized that oil seed brassica is generally influenced greatly
iby N and P. As a consequence, seed yield losses could reach as
high as 120% due to lack of N and P.
1 ?
2.2.5. Weed control
Plants show extreme plasticity in response to environmental
conditions. The presence of a neighbour plant’ constitutes one of
the most potent external factors that may limit plant size and ultimate yield (Zimdahl, 1980). This is substantiated by the work
of Forbes ( 1985 a) who found that seed yield of B. napus was severely depressed by competition from annual weeds and their onetime removal during the growing season greatly minimized the extent of yield loss. The author emphasized that growth analysis has demonstrated that leaf area index is more strongly affected than net assimilation rate by weed competition. This seems to suggest that weeds compete primarily for nutrients, since either
shading or water deficit might be expected to cause reduction in
both leaf area index and net assimilation rate. This is",corroborated by the works of Forbes (1985 b),
Jain, et al ., ( 1979 ) , Kaul, et al., (1986 ) and Kumar, et a l , , (1989) found that when weeds were allowed to grow with B. napus, they reduced the dry matter yield by more than 70% and seed and oil yields to a considerably higher extent in comparison with a weed- free crop indicating that single well-timed removal of weeds is adequate to prevent yield loss.
In Ethiopia, Badege, ejt jal. , ( 1986) noted that mustard andrape seed, at their vegetative stage, are sensitive to weed competition. In an attempt of quantification of the loss, “Hiruy, et al. , (1987 ) estimated seed yield loss of 84% due to lack of weed control.
18
2.2.6. Interactions of factors
19Dfctc Month Year
a l e m a v a u n i v e r s i t y o f| AGRlCULTURF LIBKARV
A significant effect of interaction between species and sowing dates was reported by Hiruy, et al, , (1983). As a consequence, the mustard cultivar produced higher seed yield with earlier sowing while the rape seed performed similarly both at earlier and later
sowing dates.On the interaction of sowing date and fertilizer on seed
yield, Christensen, et a l . , ( 1985) found a significant effect which led to an efficient utilization of N and P when rape was sown at the most optimum time.
Holmes (1980) and Forbes (1985 a) have summarized the interaction effects of sowing date, fertilizer and weed control on seed yield of rape seed and concluded that the response of the crop,
to fertilizer is greatly influenced by other agronomic practices including optimum sowing date and timely weed removal.
'2.3. Correlation of Characters
Correlation studies in oilseed brassicas offer valuable information on the nature and degree of interrelationship between different morphological characters of the crops. Selection based on any trait might change the other character(s ), directly or indirectly, regardless of the interest of the breeder. This is because of the fact that the attainment of characteristic form and function in a crop depends upon a chain of interrelated events
which are sequential in time and subject to the modifying influences of non-genetic variables. Moreover, these events do not occur haphazardly, but follow an integrated pattern. Thus,
component compensation is clearly a feature of relationship between components of seed yield (Adams, 1967; Thurling, 1979 a; Tatchel, 1983 ) .
In this regard, Tayo, et al.f (1975) and Thurling (1974 a) reported that seed yield was significantly correlated with the number of pods per plant and its components, number of pod bearing branches and number of pods per branch. A highly significant positive correlation between number of pods per plant and seed yield was also reported by Agarwal, et ajL. , ( 1973 ). Equally,Singh, et al* » ( 1969) and Singh, et aj,*> ( 1979) found seed yield
* * **to be significantly associated with plant height, number of primary'
and secondary branches, pods per plant, seeds per pod and 1000-seed weight. J
According to Olsson (I960), Zuberi, et al. , ( 1973 ) andThurling (1974 c), the correlation between seed yield and pods per plant and seeds per pod were all positive and significant. However, seed weight was found to be negatively correlated with number of pods per plant (Zuberi, et al. , 1973 ) and number of seeds per pod (Olsson, 1960). Nevertheless, the former authors found no significant correlation with number of seeds per pod. Basedon path coefficient analysis, Mendham, et a l . , (1975, 1981 a) found that the order of importance of yield components to seed yield include: number of pods, number of seeds per pod and seed weight.
20
Singh, et al., (1978, 1979), on the other hand, found plant height, number of pods per plant, and secondary branches to be the most important components of yield in oil seed brassica. Earlier,Singh, et a l ., (1969) found a highly significant positivecorrelation between seed yield and primary branches and pods per
plant. However, the path analysis revealed that both characters had rather negative influences on seed yield.
2.4. Environmental Influences 4
It is confirmed that oilseed rape is basically a temperate
crop and was initially restricted to such areas of the Northern and Southern Hemispheres (Weiss, 1983). As a result, the brassicas* *which thrive under relatively low temperature conditions are one ‘ of the very few edible oil sources that can be successfully
produced in the temperate regions (Downey, 1971; Appelqvist, et al. , 1972 ) .
Rape seed prefers moderate day temperatures of below 25°Cduring growth but is very sensitive to frost (Olsson, 1960;Acharya, et. a_l. , 1982; Weiss, 1983 ). A slight frost at floweringadversely affects pollen formation and viability which reducesthe number of flowers and pods produced. Conversely, it is ableto withstand a temperature of 40°C for limited periods during the
vegetative phase, but the effect of high temperature on flowering is variable.
21
v.\.
22 J
Somewhat conflicting evidence was forwarded, by different
authors regarding the relations between mean day temperatures during the grain-filling period and oil content. Olsson (1960) relates high soil moisture and high temperatures during seed
filling with high oil content. While Weiss (1983) attaches more importance to the inverse relationship that exists between mean
daily temperature during seed-filling and seed oil content.
Approximately 450-500 mm of rainfall, mainly in the vegetative and flowering period is desirable with a total of some 700 mm for maximum seed production (Hiruy, 1983; Seegeler, 1983; Weiss, 1983).
Brassicas, however, appear to be generally tolerant to severe
soil moisture deficits. They produce a good crop utilizing mainly residual soil moisture, provided they receive some rain at seeding £to ensure the germination of the small seeds, the advanced seedling f stage to promote seedling development and the main flowering stage for subsequent seed set. Soil moisture stress during flowering especially when accompanied by high temperature reduced seed weight and to a lesser extent oil content. Too much water, however, causes problems since they prefer well aerated soils and thus are liable to suffer from water logging. This provides a conducive environment that may favour fungal attacks on the roots and leads to decreased resistance to' low temperatures (Appelqvist, et al 19 72); Acharya, et al 1982),
Rape seed and other oilseed brassicas can tolerate a wide range of pH (5.0-8.5) and can also be grown on marginal soils (Weiss, 1983). Hiruy (1989 unpublished) found a pH range of 6.0
to 7.5 to be highly suitable, 5.0- 6.0/7.5-8.5 moderately suitable
and 4.5-5.0 marginal.
There is a paucity of information on species, dates of sowing, method of sowing, fertilizer and weed control practices experienced
both world-wide and under Ethiopian conditions. The information on the international scene is, at .times, conflicting as to warrant no reliable inferences that could directly be adopted to the local conditions but call for further research in line with the materials and methods elaborated hereunder.
I l l . MATERIALS AND METHODS
3.1. Experimental Sites
The experiment was conducted at Holetta and Kulumsa Agricultural Research Centers of the Institute of Agricultural Research in the 1988/89 main growing season.
Holetta in Shoa Administrative Region has the coordinates of: 38° E. and 9° N. and Kulumsa, in Arsi Administrative Region is: 390 E. and 8 0 N. They are situated at altitudes of 2400 m (Holetta) and 2200 m (Kulumsa) a.s.l.
The average annual rainfall of Holetta is 1274 mm and that of Kulumsa 74 3 mm. Of the above, Holetta receives 60% (764.4 mm) and Kulumsa 55% (408.7 mm) of the annual rainfall between June toSeptember. The 1988/89 annual rainfall was 1161 mm for Holetta and 875 mm for Kulumsa with 80% (928.8 mm) and 54% (472.5mm) of the amounts concentrated between June to September.
23
The mean maximum and minimum temperatures for the period were
20.0 0 C and 9.2 0 C for Holetta and 21.3 °C and 11.2°C forKulumsa. These are not drastically different from theannual averages of 22.4° C and 6.4 0 C of Holetta and 22.8°C and10.4°C of Kulumsa (Appendix 1}. Some of the relevant soilcharacteristics of the sites are:
24
P2O5 pH in Chro- Sand Silt ClayLocation N% DDm H2O Hue Value roa Color % % % ClsassHoletta, 0.25 13 5.20 SYR 4 3 reddish 33
brown 28 39 Clayloam
Kulumsa 0.22 50 6.10 5YR 3 1 very 23darkgray
29 48 Clay
3.2, Experimental Design and Treatments*
Thirty two combinations of five factors each at two levels ■ namely, 2 species x 2 sowing dates x 2 sowing methods x ‘2 fertilizer levels x 2 weeding practices were laid out in an incomplete block de s ign in which the highest order interact ion i s completely confounded.
.^The large number of treatments considered necessitated the use of this design which otherwise would introduce errors if a complete block design was employed. At each location, thetreatments were replicated four times thus giving a total of 128 plots per site. Each incomplete block contained 16 treatments which were identified using the modular arithmetic procedure.
The plot size used was 9m2 (1.8m x 5m) with six rows of 30cm each and 5m length for the row sowing as opposed to the 1.8m width and 5m length for the broadcasting.
The two recommended varieties representing two species of oil
seed brassica were "Tower sel3M (Brassica napus L.) and " S-67” (B.
carinata A. Braun).Tower sels is a variety developed by Ethiopian breeders
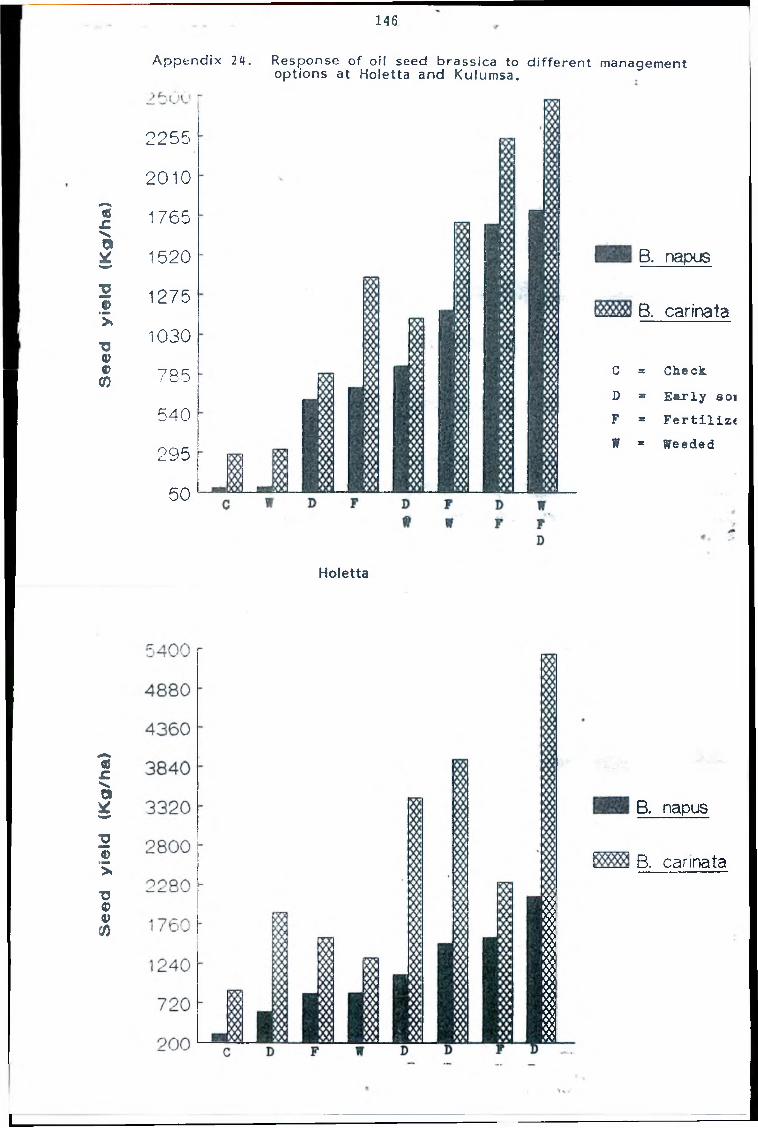
through selection from an introduced variety called Tower-a low (< 0.2%) erucic acid variety initially developed in Canada. Tower sel3 produces seed yields upto 3000 kg/ha and oil contents of 48%
under experimental conditions.S-67 is one of the few varieties of B. carinata developed and
released in Ethiopia. This variety has been 'reported to produce
seed yields of up to 4800 kg/ha and possesses oil contents up to 46% under experimental conditions. Erucic acid level in the oil
is high >30% (Hiruy, et. al., 1983; 1986).The sowing dates included; a) an optimum date i.e., 17th and
24th of June. The optimum ranges are between 10th and 20th of June for Holetta and between mid to end of June for Kulumsa (Hiruy, et
a l ., 1986) and;b) one delayed sowing which is a month after the first date
of sowing i.e. , 17 ** and 24^^ of July at Holetta and Kulumsa,
respect ively.The two sowing methods included; a) drilling the seeds in six
rows of 30 cm space and 5m length at a seed rate of 10 kg/ha. The seeds were manually drilled in each row at about 1 cm depth from
the surface and covered with soil and;b) broadcasting the seed over the plot of 1.8m width and 5m
length at the same seed rate i.e., 10 kg/ha. After the seeds were
25
approximately evenly distributed over the plot, they were covered
with soil using a rake.
The two fertilizer levels were: a) no fertilizer and b) arecommended level of 46-30 N-P kg/ha (Hiruy, et. al., 1983} as DAP
and urea combination applied at the time of sowing. The fertilizer was drilled or broadcasted depending on the method of seeding. With row sowing the fertilizer was drilled manually in each of the six rows and covered with a thin layer (2-3 cm) of soil .just before
seeding inorder to avoid a direct contact with the seed. Similarly with broadcast seeding the fertilizer was broadcasted and covered
with soil.The weed treatments were represented by a) no weeding and b)
hand weeding. The hand weeding was done a month after sowing at*
about the second true leaf stage. The weeding was dene once at the specified stage because earlier it was found that the advantage of
subsequent weed removals over the first timely done hand weeding was not economical (Badege, et. a.1., 1986; Hiruy, ejt a l . , 1987).
3.3. Parameters Collected and Methods of Collection
A. Seedling density (Stand establishment): This was calculated,for the whole plot based on the number of seedlings that have emerged two weeks after weeding (six weeks after sowing) and the average number of seeds sown in each plot.
The average number of seeds sown, for each species, was determined based on the count made on four samples each of nine
grams (weight of seeds required for a plot). The seeds from each
sample were counted using seed counter and the average over the
four samples was considered to be the number of seeds sown in each plot of 9mz . Using this procedure, 1923 seedlings were expected in each plot of S-67 (Brassica carinata) and 2026 seedlings of Tower sel3 (B. napus).
B) Days to flowering: This was assessed based on visual
observation and constitutes the number of days from sowing to about 50% blooming.
C) Days to Maturity : Based on visual observation, it is thenumber of days from sowing to the time when about75% of the pods change color from green to yellow-green.
D) Yield components: In order to determine the various facts,ten plants were randomly selected (Singh et al. , 1978;Ibrahim, 1983) from the center four rows of the row sown and the center 1.2 m by 5m of the broadcast sown plots and tagged at the time of maturity,
i) Plant height : The average height of the ten taggedplants measured using a height meter from the groundlevel up to the tip of the main stem,
i i ) Number of primary branches: Average of the ten taggedplants
i i i ) Number of secondary branches: Average of the ten taggedplants
27
iv ) Number of pods per plant: Average number of pods of the
ten tagged plantsv) Number of seeds per plant: Average of the ten tagged
plants.vi) Number of seeds per pod: Calculated by dividing the
number of seeds per plant by the number of pods per plantvii) Thousand-seed weight: The seeds of the identified ten
plants were bulked in to one lot from which 1000 seeds were counted and weighed
E) Stand percentage at maturity: This was calculatedfrom the number of plants harvested and expected from the center four rows (6m 2 ) of row sown and the center 1.2m by 5m ( 6m 2 ) of the broadcast plots. Based on the average number of, seeds initially sown in 6m2 , the expected number of plants
calculated to be 1282 for S-67 and 1350 for Tower sel3.
F) Seed yield: The weight of seeds from the center fourrows (6m2 ) of row sown and the center 1.2m by 5m (6m2 ) of broadcast sown plots plus the weight of seeds from the ten tagged plants used to determine the yield components all converted into yield/hectare.
28
G) Oil content: Twenty-two grams of seeds which are free frominert matter were taken from each plot and dried in an ovenat 135°C for two hours or 75°C for 12 hours. The dried seeds
were then subjected to the Nuclear Magnetic Resonance (N M R ) oil analyses (Madson, 1976) from which a direct reading of theoil percentage was made. >-
H) Oil yield: This was calculated from the ratio of seed yield
and oil content (based on 3.3 F and G).
The analysis of variance was performed in accordance with theprocedure outlined by Cochran, et. aJ. , ( 1957). In the path-coefficient analysis, a method similar to that used by Dewey, et al. , (1959), Singh, et. al*> (1978) and Ibrahim (1983) was employed to partition the correlation coefficients into direct and indirect effects. The path-coefficients were obtained by the simultaneous solution of the following equations. >**
-t- 2P35 r34 P45Where rij = The simple correlation coefficient between character
(i) and character (j ) ;Pij = The direct effect of i on j;
rij Pjk = The indirect effect of i on k via j; andThe x- variable consists of all residualfactors.
29
Since the experiment was conducted for one season :at each
location, combined analysis was considered unjustified and hence separate analysis was done for the two locations because it
doesn’t pay dividends interms of the effort that would be invested, yet some preliminary comparisons based on means are made.
Subsequent to the above approach, the following results and the discussion thereof are further elucidated here under.
IV. RESULT AND DISCUSSION
4.1 Effects of the Considered Factors on AgronomicCharacters
4.1.1. Species
Reference to Table 1 and Appendices 2 and 3 indicates that there were highly significant differences between S-67 (Brassica
carinata) and Tower sel3 {B. napus} in stand, nukber of primary and secondary branches per plant, number of seeds per plant, seeds per pod and 1000-seed weight-at both locations. In addition, there was highly significant difference, between the two species, in seedling density at Holetta.
At Holetta, Tower sel3 (B. napus) seems to be denser, at the seedling stage, than S-67 (B.. carinata). About 45% of the seedsof the former and 38% of the latter species have been established. At Holetta, 48% of the expected plants of S-67 reached maturity as compared to 42% of Tower sel3, while at Kulumsa 40% of Tower sel3
reached maturity as compared to 39% of S-67.
30
31
i { r : s o n c c h a r a c t e r s as a f f e c t e d a* s p e c i e s , $ 3 « : d j s i t e sd » : 3 ; ie: f e r t i l i z e r ind i e e i c o n t r o l it S o l e t t a a n d I u I u a s s .
Seediias deesity Vutber of fuiber of Days to Days to\ Stand Frisary • secondary floierinj satunty
\ branches branchesper plant per plant
Holetta lalaisi H o le t t a [ t l u s a lole t t a [ u h i s a loletta laloisa H o le t t a l a k i s a Hole t t a l a l m s airissiaarinti (S-67) 38 44 48 3? 5,57 8.08 8.2 12.3 85 78 176 154S.upas 'Toier
selj) 45 46 42 40 3.35 2.80 0.3 1 7 80 i L 157 138
Earij scfin? 45 59 48 43 4.71 55.28 4.5 5.9 83 •» 4 - •* 164 147late sciing 3T 31 Iu 3 ( 4.21 5.60 j f n IA 32 76 169 145
Broiiccs: , r 14' 1:5.? 131s **41 0 75 *: r 1 ^3*t. V / 3.60J3f 1 f 14? . j'j . ' j* .1 0<' >.41 11.92 4.13 3.52fithout fertilizer 107 140 3?.2 93.1 ■ 1 ».
• ‘t. 35* t * 1 *.* * z 4.02 3.54lith fertilizer H I 156 117.2 123.? 1133 1534 «« M H 11ii/.ji i L . n 4.20 3.56lith 10 reeding ll? 144 95.3 83.4 336 1035 9.23 12.17 4.07 3.59lith feeding 129 152 137.1 133.5 1037 1532 5.93 11.76 4.15 3.52Kean 124 143 97 4 1 *» 0*1 5.58 11.97 4.11 3.56
:.7.* 13 13. -6 I 1 •J IS P 12 3
L-SS»01 5.37 3.91 6.? ft i C. f $.93 M v.7 4 3 .55 1 1*1 0.134•35 4.51 5.72 5.3 5.5 535 ; 1 ^ £? J 7? 3.16? 3.101
At Holetta, S-67 (B. carinata) produced six primary branches
per plant ■ as compared to three of Tower sel3 ( B. napus). At t
Kulumsa, S-67 had eight primary branches per plant whereas Tower sel3 had about three. At both locations, species was the most important factor inducing the largest amount of variability observed in this character. Similar trends of increase and importance vis-a-vis species was observed in number of secondary
branches per plant.At Holetta, S-67 produced about eight secondary branches per
plant as compared to 0.2 of Tower sel3 . At Kulumsa, the values were 12 for S-67 and 2.2 for Tower Sel3 . The results support what
Hiruy, et a l ., (1986) and Hiruy (1987) found. They reported that cultivars of B. napus are more compact with reduced branching
— *» habit than cultivars of B. carinata* »
At both locations, the mustard (B. carinata) variety S-67 flowered a few days later than the rape seed (B. napus) variety Tower sel3. The statistically significant delay was longer at Kulumsa where it took 78 days for S-67 and 72 days for Tower sel3 to flower. At Holetta, however, the varieties flowered after 85
and 80 days, respectively. Under the prevailing conditions of both sites, S-67 had delayed maturity of about two weeks when Tower sel3 was ready for harvest. The average values for S-67 and Tower sel3 being 176 and 157 days at Holetta with corresponding values of 154
and 138 at Kulumsa.The largest difference in days to maturity was attributed to
varietal difference. Similar results, whereby it took longer for
3 3
mustard cultivars than rape seed cultivars to flower and mature,
have been reported by Mizushima, et. ajl. , (1967), Chauhan, et, al ., (1984), Hiruy, et al ., (1983, 1986) and Hiruy (1987).
The mustard (B. carinata) variety S-67 grew taller, by 20 and
28 cm, than Tower sel3 at Holetta and Kulumsa, respectively. At Holetta, varietal difference was the third (following fertilizer and date of sowing), important factor in plant height while at Kulumsa, it contributed the largest amount of variability observed in this character. The results agree with the findings of Hiruy, et a l ., (1983, 1986, 1987) who reported greater values of plantheight in B. carinata cultivars than in B. napus cultivars.
Transformation of the average differences between the two species to percentages reveals that S-67 (B . carinata) produced 62 and 86% more number of pods per plant than the average values of 78.7 and 77.6 pods per plant of Tower sel3 (B. napus) at Holetta and Kulumsa, respectively. With these effects, species took the lead in inducing most of the variabilities observed in the number of pods per plant at both locations. At both locations, the number of seeds per plant was also higher for S-67 than for Tower sel3.
e
Similar transformation to percentages reveals that S-67 (B.
carinata) produced 10% at Holetta and 56% at Kulumsa more number of seeds per plant than the respective averages of 926 and 1004 seeds per plant of Tower sel3 (B. napus). These differences rendered species to be the fourth,important factor for higher number of seeds per plant after fertilizer, sowing date and weed control at Holetta; and the second, next only to fertilizer at
34
Kulumsa. The results appeared reproducible in that Mizushima, e£
a l .. (1967) and Chauhan, et a l . , (1984) have also found increased'i
number of pods and seeds per plant in mustard than in rape seed.S-67 (B . carinata) being taller and possessing by far greater
number of pod bearing branches vas able to produce more number of pods and seeds per plant than Tower Sela (B.napus) which was shorter with less number of pod bearing branches and hence less number of pods and seeds per plant.
At both locations, Tower sel3 (B. napus) produced higher number of seeds per pod than S-67 (B. carinata). Transforma
tion of the average difference, between the two species to percentages indicated that Tower sela (B. napus) produced 49% (Holetta) and 23% (Kulumsa) more number of seeds per pod than the
respective mean values of 7.70 and 10.71 seeds per pod of S-67 (B.>
carinata).
Although the magnitude varied greatly, the difference in seeds per pod due to species difference, at both locations, was the largest compared to that of any other factor considered. It can be noted that S-67 (B. carinata) despite its tall, profusedbranches with enormous pods habit (Table 1) produced by far less number of seeds per pod than the short, compact branches with less number of pods from Tower sels (B. napus). This is probably because of the fact that pods per plant and seeds per pod are negatively associated in oilseed brassica (Ollsson, I960; Zuberi, et a l ., 1973; Thurling, 1974 c).
At both locations, S-67 (B. carinata) had heavier seeds than
35
Tower sel3 (B. napus). S-67 had 4.29 and 3.66g per 1000 seeds as compared to 3.93 and 3.46 of Tower sel3 at Holetta and Kulumsa, respectively. This amounts to an advantage of 0.36 and 0.20 g per 1000- seeds for Holetta and Kulumsa. Species difference, once again, was responsible in inducing the largest amount of variability in 1000-seed weight. This is in agreement with thereports of Hiruy, et. aj . , ( 1983 ) who also found that B. carinata
cultivars had heavier seeds than B. napus cultivars.
4.1.2. Date of sowing
Table 1 and Appendices 2 and 3 provide the effect andsignificance of the effect of sowing date on different agronomic
characters of oilseed brassica.At both locations, sowing date had highly significant effects
on seedling density, stand, days to flowering, plant height, number of pods per plant and number of seeds per plant*. Highlysignificant effects were also observed on number of primary and secondary branches per plant, days to maturity, number of seeds per pod and 1000-seed weight at Holetta. At Kulumsa, significant effects were noted on number of primary and secondary branches per
plant and days to maturity.Early sowing resulted in higher mean seedling density values
of 46 and 59% as compared to corresponding 37 and 31% of late sowing at Holetta and Kulumsa, respectively. At Holetta, with a difference between early and late sowings of about 9%, sowing date
3G
was the second, next only to species, most important factor to induce the variability observed in seedling density while at
«Kulumsa, with an average difference of 28%, sowing date had the greatest influence for higher degree of seedling establishment.
Both at Holetta and Kulumsa, stand percentage was higher under early than late sowing dates. Early sowing increased stand by 6 and 7% at the two locations. The results are in agreement with what Pretty, et aJL. , ( 1976 ) and Degnehardt, et al., (1981 a )
found.The higher number of seedlings and subsequently number of
plants that reached maturity observed with early sowing at both sites might be due to the relatively warmer and more favourable moisture regimes (Appendix 1) which provided better conditions for
* *higher rate of germination and early establishment of the seedlings which could successfully compete with weeds and escape the excessive soil moisture at their most susceptible stage and hence more number of plants at harvesting.
Early sowing increased number of primary and secondary branches per plant at Holetta but reduced both characters at Kulumsa. Transformation of the differences from the absolute values to percentages, between early and late sowing dates, reveals that early sowing has increased the number of primary and secondary branches per plant at Holetta by 12 and 30%, respectively, but decreased them by 6 and 9% at Kulumsa. At Holetta, sowing date was the third most important factor for higher number of primary and secondary branches, next to species and
37
fertilizer, and the second, next to species. But at Kulumsa, both
characters were least affected by sowing date*The results at Holetta are in agreement with the findings of
Thurling (1974 b) who reported decreased number of primary and secondary branches with delayed sowing while results from Kulumsa
agree with the findings of Degnehardt, et .al* ♦ (1981 a) whoreported increased primary and secondary branches with delayed
seeding. These dichotomous reports provide a clue to the
conflicting trends observed at the two locations in that environmental influences might play the greatest role in thedevelopment of primary and secondary branches in brassicas.
Although not of agronomic significance, early sowing had atendency to increase days to flowering at Holetta but reduced it
(
by two days at Kulumsa, At Holetta, late sowing delayed maturity by five days but at Kulumsa, a reverse trend of two days was noted although the magnitude was not agronomically important.
The resultant effect of early sowing was decreased grain- filling period (81 days) as compared to 87 days of late sowing at Holetta but increased period (73 days ) as opposed to 69 days at
Kulumsa.The results at Holetta are consistent with the findings of
Christensen, et jlL., (1985) who found delayed maturity resulted from excessive delay of sowing date and that of Kulumsa was in agreement with the results of Kondra (1976) who reported reduced days to maturity with advancement of sowing from the optimum sowing date. It can be perceived that growth generally was speeded up at
38 1
Kulumsa where the growth period was relatively warmer and the
temperature extremes were more mitigated than it was at Holetta
(Appendix 2). This might’ be the reason why late sown plants at
Kulumsa completed their cycle faster.At both locations, plant height was increased with early
sowing. At Holetta, plant height was increased by 24 cm over 112 cm of late sown plants; and at Kulumsa, it was increased by 28 cm
over the mean 134 cm of late sown plants. With these effects,
sowing date stood as the second, next to fertilizer, at Holetta and first at Kulumsa, most important factor as far as plant height is
concerned.A casual observation of plant height in Table 1 reveals that
plants were more vigorous at Kulumsa than at Holetta. The resultsy**«
are in support of the works of Fulkerson (1971) and Mendham, e£ al. , (1981 b) who, with early sowing, found taller rape seedplants. The association of tallness to early sowing again might
be due to the provision of conducive temperature and moisture conditions for early germination and establishment of sturdier and vigorous seedlings that can successfully out compete weeds and make efficient use of nutrients and virtually become taller plants.
Early sowing, invariably, increased number of pods and seeds per plant at both locations. Transformation of the average differences, between early and late sowing dates, of the mean values indicates that the number of pods per plant was increased by 9% at Holetta and by 16% at Kulumsa. Similarly, number of seeds per plant was invariably increased by early sowing. With delayed
3 9
sowing, number of seeds per plant has decreased by 44 and 26% at
Holetta and Kulumsa, respectively. With these attributes, sowing
date took the third, after species and fertilizer and the second, next to fertilizer, at Holetta; and the fourth, next to species,
weed control and fertilizer and the fourth, following fertilizer, species and weed control, at Kulumsa orders of importance as far as number of pods per plant and seeds per plant, respectively, are concerned.
The findings of Thurling (1974 b), Mendham, et al., (1975), Holmes ( 1980), Degnehardt, et al,, (1981 a) and Mendham, et al. ,
(1981 a) support the results in that they have also reported increased number of pods and seeds per plant with early sowing.
r
It is a pp a r e n t l y clear that early sowing with its favourable* * ^
conditions increased the sites, via increased number of pod bearing* branches and plant height, where more number of pods could develop and virtually more seeds were produced.
At Holetta, late sowing appeared to increase number of seeds per pod. Percentage transformation of the mean difference of the late and early sown plots shows an increase of 47% over the mean value of 7.76 seeds per pod of early sown plots. sowing date appeared the second, next to species, most important factor for higher number of seeds per pod. Thurling (1974 b) and Mendahm, et a l .. (1975, 1981 a) have also reported similar results where bydelayed sowing increased number of seeds per pod. The reason might possibly be due to the inverse relationship that exists between number of seeds per pod and pods per plant (Olsson, 1960; Zuberi,
40
et a l .. 1973) which was increased by early sowing.
At Holetta, delayed sowing reduced 1000-seed weight by 0.34g
below the average 4.28g of early sown plots. Thurling (1974 b) and Ali-khan (1976) have also reported results of decreased seed weight due to delayed sowing.
4.1.3. Method of sowing
Reference to Table 1 and Appendices 2 and 3 indicates that the effect and significance of sowing method on agronomic characters of brassicas are such that method of sowing had a highly
significant effect on seedling density, stand, number of secondary branches per plant and number of seeds per plant at both locations. It also manifested highly significant effects on number of primary branches per plant, days to flowering and number of pods per plant at kulumsa; while at Holetta, it significantly affected number of primary branches per plant and days to flowering.
Both at Holetta and Kulumsa, method of sowing had a great deal of importance in the number of seedlings that could be established. Row sowing has increased the density of the seedlings from 32 to 51% at Holetta and fro® 3 7 to 53% at iulumsa. Among the factorsconsidered, method of sowing seems to exert the highest influence on seedling establishment at Holetta and ranked second, next only to sowing date, at Kulumsa. At both locations, stand percentage was higher in row sown than broadcasted plots. The respective increases were 8% for Holetta and 3% for Kulumsa. Clarke, et al. ,
41
(1978) comparing the effects of method of sowing in rape seed found seedling establishment and later stand were comparable for
broadcasting and drilling*The higher values obtained for both characters with row sowing
might be due to the fact that row sowing ensured proper coverage of the seeds so that there would be minimum loss from birds particularly crow. On top of this, losses from mechanical damages during weeding, by the workers were almost nill in the row sown plots. However, with broadcasting where the seeds are haphazardly distributed, there is no guarantee of complete seed coverage which,
therefore, are prone to bird attack apart from the high damage during weeding. These phenomena have probably led to reduced number of seedlings and hence low stand percentage thereof.
Plants sown in rows produced less number of primary branches,
than broadcast sown plants which had 6 and 9% more primary branches than row sown plants at Holetta and Kulumsa, respectively. Similarly, broadcasting increased number of secondary branches per plant by 18% at Holetta and 20% at Kulumsa.
At both locations, method of sowing stood as the least important factor, among the factors considered, as far as primary branches are concerned. However, it was the fourth, following species, date of sowing and fertilizer, at Holetta; and the third, after species and weed control at Kulumsa, important factor for higher number of secondary branches per plant. These results are consistent with the findings of Clarke, e_t al. , (1978 ) who alsoreported increased number of primary and secondary branches with
42 *
broadcasting over those of row sowing.The explanation for the results can be attached to the
findings of Tayo, et al. , ( 1975) who explicitly stated thatcompensations, for characters, in oilseed brassica are common phenomena. Therefore, under broadcasting where less number of pants per unit area were found than under row sowing, there were more number of primary and secondary branches per the same unit
area to compensate for the reduced population density. Conversely, with row sowing where there were more plants per unit area, the
Anumber of primary and secondary branches were less.
Although significant, the difference in days to flowering of row and broadcast sown plants was not that impressive both at Holetta and Kulumsa, Yet, row sowing shortened days to flowering by one and two days at the two locations, respectively. From tfce preceding paragraph, it is vivid that broadcast sown plants were characterized by more profused branches. Such branchy plants of oilseed brassica are mostly late as compared to more compact types.
At Kulumsa, there was a trend of increased number of pods per plant with broadcasting. Transformation of the average difference to percentage reveals that this method of sowing increased the character by 9% over the mean value of 106 pods per plant of row sown plots. Broadcasting, once again, has increased number of seeds per plant over row sowing both at Holetta and Kulumsa. Broadcast sown plants produced 7 and 9% more seeds per plant at the two respective sites. Method of sowing, however, appeared the least, among the considered factors, important to induce
43
variability in the number of seeds a plant can produce. The results are in agreement with the works of Clarke, et (1978)
who also found increased number of pods and seeds per plant with broadcasting. The increase in both characters can be explained by taking into account the higher number of pod bearing branches produced by individual plants of broadcasting.
4.1.4,Fertilizer
The effects of fertilizer on different characters of oilseed
brassicas are shown in Table 1; while the significances of these effects are given in Appendices 2 and 3. Highly significant effects of fertilizer were observed on stand, number of primary and secondary branches per plant, days to flowering and maturity,plant t height, number of pods and seeds per plant and number of seeds per pod at both Holetta and Kulumsa. In addition, such effects were noticed on seedling density and 1000-seed weight at Holetta.
Fertilizer increased the average seedling density by 3% over the mean 40% of non-fertilized plots at Holetta. Concomitantly, fertilizer increased stand percentage by 12 and 5 at Holetta and Kulumsa, respectively. The results are consistent with the observation of Hiruy, et al., (1987) who found increased number of plants reaching maturity with fertilizer application.
The average number of primary branches per plant has been increased by 27 and 17% at Holetta and Kulumsa, respectively. Although the absolute average difference (<1) between fertilized
44
and non-ferti1ized plots at Holetta was not that marked, it
remained the second, next to species, most important factor at both
locations to indyce the greatest variability observed in number of
primary branches.Fertilizer application also increased the number of secondary
branches per plant by 30% at Holetta and 20% at Kulumsa, These/
rendered fertilizer to be the second, following species, at Holetta
and the third, following species and weed control, at Kulumsaimportant factor to affect number of secondary branches.
These results are consistent with the findings of Olsson (1960), Joarder (1983) and Forbes (1985 b) who also found increased number of primary and secondary branches per plant with fertilizerapplication indicating that plants under conditions of judicious
\
fertilizer levels were able to possess increased height an$
virtually increased number of branches.iFertilizer shortened days to flowering by about 10 days at
Holetta and by 6 days at Kulumsa ihdicating that fertilizer is the most important factor to affect days to flowering of oilseedbrassica. Fertilizer application has also shortened maturity time by two weeks at Holetta and by a week at Kulumsa. With these effects, fertilizer was the second, next to species, most important factor which was responsible for the variability observed in days
to maturity.These results are in agreement with the findings of Hiruy,
et al .» (1983, 1986, 1987) and Hiruy (1985) who in Ethiopia, found that fertilizer shortened both days to flowering and maturity very
45 i
appreciably. However, Christensen, e t a l ., (1985), in Canada,
found delayed flowering and subsequently days to maturity with the, i application of nitrogen and phosphorus. These conflicting trends
might arose from the multitudes of variations in environments wherethe experiments were conducted and the varieties used.
Fertilizer increased plant height by 34 and 16 cm at Holetta
and Kulumsa. The result is supported by the reports of Sheppard,
et a l . , (1980 ) , Mendham, et al., (1981 b ) and Hiruy, et al- > (1986, 1987). As would be expected, the plants utilized the additional nutrients made available to them in the form of fertilizer.
The number of pods per plant was increased by 31% at Holetta
and 38% at Kulumsa. It was the second, next to species, at Holetta; and the third following species and weed control, at
Kulumsa; most important factor to induce the variability in this character. Fertilizer also increased the number of seeds ofindividual plants by 60 and 63% at Holetta and Kulumsa,
respectively.At both locations, fertilizer was the first important factor
to induce the largest difference in number of seeds per plant. At both locations, fertilizer resulted in higher number of seeds per pod. At Holetta, it stood as the third, following species and sowing date, important factor because, it increased the number of seeds per pod by 22% over the average absolute value of 8.64 seeds per pod of non-fertilized plots. Similarly at Kulumsa, it was the second, next to species, inducing a 14% higher number of seeds per pod than the average 11.16 seeds per pod of non-fertilized plots.
At Holetta, plants under fertilized condition had 0.18g more weight per 1000-seed than the average value of 4.02g under the non
fertilized condition. The results on number of pods and seeds per
plant, seeds per pod and 1000-seed weight are in agreement with
those of Olsson (1960), Allen, et al., (1971) and Mendham, et al. ,
(1981 b) who have also found that fertilizer affected each of these characters favourably. Plants grown under better fertilized conditions grew much taller and hence had more sites for the development of more pods and therefore virtually more seeds.
4.1.5«Weed ControlTable 1 reveals the effect of weed control on agronomic
characters of S-67 (Brassica. carinata) and Tower sels (B, napus),*
The significances of the effect are given in Appendices 2 and 3.. Accordingly, weed control had highly significant effect on stand, number of primary branches per plant and number of seeds per plant
at both locations. It also had such effects on seedling density, plant height and number of seeds per pod at Holetta and number of secondary branches per plant, days to maturity and number of pods per plant at Kulumsa. Weed control also had significant effect on
number of pods per plant at Holetta and plant height at Kulumsa.At Holetta, the seedling density under less competitive
conditions exceeded the average 40% of the unweeded plots by 3%. Weedy conditions reduced number of harvestable plants by 4 and 3% below the average 47 and 41% of stand at Holetta and Kulumsa, respectively.
It is apparent that under conditions of intense competition, the young and relatively slow growing brassica seedlings are very
easily over come by the weed species which were fast in growth and early in establishment. This enabled them to compete successfully hence reducing the number of crop seedlings and eventually the number of plants reaching maturity.
Plants under less weed competition had generally more number
of primary branches. Transformation of the difference, between the mean values of weeded and unweeded plots, in to percentages indicates that weed control increased number of primary branches per plant by 9% at Holetta and 24% at Kulumsa.
With these magnitudes, weed control stood as the fourth, following species, fertilizer and sowing date, at Holetta and the third, next to species and fertilizer, at Kulumsa, important factor'" as far as this character is concerned. Weed control, at Kulumsa, had also increased number of secondary branches per plant by about 74% over the mean 5.3 of unweeded plots. This rendered weed control to be the second, next only to species, important factor affecting number of secondary branches.
The results of this experiment agree with the works of Jain, et al., (1979), Forbes ( 1985 b) and Kaul, et al« > (1986) whoemphasized that a reduction in the available nutrients, light, moisture and space due to weed competition would be expected to restrict the growth of primary and secondary branches.
At Kulumsa, there was a trend of increased days to maturity with conditions of less weed competition. Plants under less weedy
48
situations required four more days than required by those grown under competitive situations.
At both locations, weed control increased plant height. T ie respective increases were 10 and 8 cm over the corresponding average values of 119 and 144 cm of unweeded plots. Consistent to
this result, Jain, et al. , ( 1979 ), Kaul, et al. , (1986 ) and Kumar, et aj,. , (1989) also found increased plant height with lessconditions of weed competition. This might suffice to explain the longer maturity period observed at Kulumsa with less weedy condition since it is c o m m o n l y observed that taller plants with profused branching habits are late maturing.
Both at Holetta and Kulumsa, plants under less weedy conditions produced more number of pods per plant than those under weedy conditions. Due to lack of weed control, losses in the, number of pods per plant amounted to 8 and 45 at Holetta and Kulumsa, respectively. These values correspond to about 8 and 34% of the respective potential that could be accrued by single timely hand weeding. This is in agreement with the reports of Forbes (1985 b) who found increased number of pods per plant with timely removal of weeds. This is possibly because of increased number of
pod bearing primary and secondary branches which resulted from weed-free growing conditions (Table 1).
Number of seeds per plant was also higher under less competitive conditions at both locations. It increased by 17 and 48%' over the average values of 886 and 1036 seeds produced by individual plants grown under weedy conditions of Holetta and
49
Kulumsa, respectively.
With the indicated effects at both locations, weed control«
stood as the third, following fertilizer and sowing date at Holetta and fertilizer and species at Kulumsa, important factor to induce higher number of seeds per plant. This is because of the increased
number of pods and pod bearing branches that resulted from weed removal (Table 1).
At Holetta, number of seeds per pod due to weed control has been increased by 8% over the average 9.23 seeds per pod of
unweeded plots. This effect made weed control to stand as thefourth important factor for higher number of seeds per pod following species, date of sowing and fertilizer.
( * *i 4 &
4.1.6. Interactions of factors '
4.1.6.1. Species and sowing date (SD): Plant height was affected, significantly at Holetta, and highly significantly at Kulumsa, by the interaction of species and sowing date (Appendices 2 and 3).
At both locations, both S-67 (Brassica carinata) and Tower sel3 (B. napus) were much taller under earlier than late sowingdates (Figure 1 and Appendix 4). Further, the increase due toearly sowing was much higher in S-67 (B. carinata) than Tower sel3 {B. napus). Early sowing increased the height of the former species by 29 cm (Holetta) and 40 cm (Kulumsa) and that of the latter by 19 and 15 cm at the respective locations.
50
No.
of
se
con
dar
y b
ran
ches
/pla
nt
Plan
t he
ight
(c
m)
51
D1 D2 D1 D2
Brassica carinata Brassica napusFigf.l.Effects of the interactions between species and sowing date
on plant height.
LSDt Hoi etta Kulumsa
0.01 BA5 12.600.05 6.37 9.50
Holett
Kulum<D1 = Early
D2 = Late
F0 F1 F0 F1 F0 F1 F0 F1
Brassica Brassica Early Late
carinata napus
Fig. 2, Effects of the interactions between species, sowing date and fertilizer on
number of secondary branches per plant at Kulumsa.
Early sowing, by providing more favourable temperature and
moisture regimes during the seed germination and the subsequent growth period resulted ih more vigorous plants than late sowing.
The higher rate of increase in S-67 (B. carinata) might be due to
its longer growth cycle, than Tower sela (B , napusJ, which,therefore, by early sowing could fully utilize the favourable conditions to manifest its potential to a greater extent*
4.1.6.2. Species and fertilizer (SF): The species and fertilizer interaction had highly significant effect on the number of secondary branches per plant at Kulumsa (Appendix 3)*
At both levels of fertilizer, S-67 (B. carinata)
produced more number of secondary branches per plant than Toweri i
sel3 {B. napus). The highest difference between the mean values of the two species, transformed in to percentage was under fertilized (921%) than non fertilized (268%) conditions. The response of the two species, to fertilizer, was somewhat contradictory in that B. carinata (S-67) produced 39% more number of secondary branches due to fertilizer whereas B, napus (Tower sela) tended to produce less although the magnitude was not that - marked (Figure 2 and Appendix 5). This might be due to the indeterminate nature of B. carinata (Hiruy, et al. , 1983) whichupon fertilizer application grew more luxuriously as compared to the B. napus cultivar which is rather more determinate and less branchy. Hence, with additional nutrients this species tends to
52 *
grow taller with more of primay branches and pods than secondary branches.
4.1.6.3. Sowing date and fertilizer (D F ): At Holetta, theinteraction of sowing date and fertilizer has highly significantly and significantly affected stand and plant height, respectively. However, at Kulumsa, significant and highly significant effects were noted on stand and number of secondary branches per plant, respectively (Appendices 2 and 3).
At both locations, fertilizer increased number of harvestable plants from both early and late sowings (Figure 3 and Appendix 6). It increased stand of early sown plots by 6 and 3% and that of late sown ones by 16 and 7% at Holetta and Kulumsa, respectively.
Early sowing also increased this character under both levels o£ fertilizer and locations. The respective increases from nonfertilized plots were 11 and 10% and from fertilized plots 1 and 6%.
At both locations, the higher response to fertilizer was at
the later sowing dates and that to sowing date was observed at no- fertilizer conditions. These imply that the negative influences of lack of fertilizer or delayed sowing can be off-set by early sowing or judicious fertilizer application to some extent.
At Kulumsa, fertilizer increased the number of secondary branches per plant only under early sowing date. Transformation of the average differences to percentages shows that the increase due to fertilizer amounted 43%. With late sowing, however, there
53
o*>-O c f0 4-1(/)
£
o(J
70
60
<oc<t>T>9Cvo4>cn
50
40
30
Early Late \Fig. 3* Effects of interactions between
sowing date and fertilizer on stand. FO = No fertilizer
F1 = With fertilizer
Late
Fig. 4. Effects of the interactionsbetween sowing date and fertilizer on plant height at Holetta.
FO = No ferti1i zer F1 = With fertilizer
0.01 = 8. 50.05 = 6.37
LSD04
Hoietta Kulumsa
5.5 2.34.2 1.;
Holetta u £LBSBSa K u S u n s a
150 '’c' 1 1 130 -C 120
110
Holetta
H U i KulumsaB = Broadcast
R = Row
Early Late Early Late
Brassica carinata Brassica napusFig. 5»Effects of the interactions between species, sowing date and method of sowing on seeding
densi ty.
was rather a decrease of secondary branches per plant by 13%. On
the other hand, early sowing increased secondary branches by 23% with fertilizer application, but reduced it by 60% with no
fertilizer application (Figure 2 and Appendix 5).The results of Bhan (1979) support the current results in
that they have also found increased number of secondary branches due to fertilizer or early sowing only under better condition of the other. This indicates that it is worthwhile to consider both fertilizer and early sowing for higher number of secondary branches
than each factor alone.Both fertilizer and early sowing increased plant height at
Holetta (Figure 4 and Appendix 4). The increase, due to either factors, however, varied in magnitude. The effect of fertilizer was more pronounced under late sowing than under early sowingswhile that of early sowing was under non-fertilized condition.
/
Hence, fertilizer increased plant height by 26 cm with early sowing and by 44 cm with late sowing. Early sowing, on the other hand, increased plant height by 33 cm with no-f ert il izer and by 15cm with
fertilizer.It can be noted that plants under less favourable conditions
of sowing date grew at a higher rate upon fertilizer application than plants under more favorable sowing date conditions but with the same fertilizer level. Similarly, early sowing resulted in higher rate of growth, as measured by plant height, with nofertilizer than with fertilizer conditions.
55
4.1.6.4• Species, date of sowing and method of sowing (SDM):Significant interactive effects were noted on seedling density at
both Holetta and Kulumsa; and on 1000-seed weight at Holetta;
highly significant effects on number of primary branches and pods
per plant at Kulumsa (Appendices 2 and 3).Reference to Figure 5 and Appendix 7 reveals that at both
locations, both S-67 (Brassica carinata) and Tower sel3 (B. napus)
were established at higher densities under row than broadcast
sowings.At Holetta, row sowing of both species was more beneficial
for seedling establishment than sowing date. It was particularly more relevant to Tower sel3 (B . napus) than to S-67 (B. carinata)
because row sowing increased seedling density of early and lateX 4
sown Tower sela (B. napus) by 75 and 68%, respectively* Nevertheless, row sowing has also increased seedling density of early and late sown S-67 (B. carinata) by 66 and 26%.
At Kulumsa, both species responded more to sowing date than to method of sowing as opposed to that of Holetta. The magnitude
of the response, however, varied with method of sowing. Early sowing increased the density of broadcast sown S-67 ( B. carinata)
and Tower Sel3 (B. napus) seedlings by 96 and 67%, respectively, while that of row sown S-67 (B. carinata) and Tower Sels (B. napus) were increased by 81 and 105%.
The results in Figure 6 and Appendix 8 indicate that at Kulumsa, S-67 (B. carinata) had a tendency to produce higher number of primary branches with .broadcasting than with row sowing whereas
Num
ber
of
pods
/pla
nt
Num
ber
of
prim
ary
bra
nch
es/
pla
nt 9
8
7
6
5
4
3
1
Fig.
B R BEarly Late
Brassioa oarinata
B R BEarly Late
Brassioa napus
6«Effects of the interactions between species, sowing date and sowing method on number of primary branches per plant at Kulumsa.
A. Species by sowing date by method of sowing (Kulumsa)
LSD
B. Species by sowing date by fertilizer (Holetta)
L S D0.01 = 15.3 0.05 = 11.6
Early LateBra§sicacar^nata
Early LateBaras si aa napus
Early LateBrassioacarvnata
Early LateBrassioanapus
Fig. 7* Effects of the interactions between species, sowing date and fertilizer on number of pods per plant.
the reverse held true for Tower sel3 (B. napus). With delayed
sowing, the former species produced 38% more primary branches under
broadcasting than row sowing while the latter had 20% under row sowing. Similar trends were also observed with early sowing but the magnitudes were not that marked. Thus, broadcasting was
important for S-67 ( B . carinata) and row sowing for Tower sel3 (B .
napus) as far as number of primary branches are concerned.With broadcasting, both species produced more primary branches
under late than early sowing dates. The magnitude of the increase, however, was much pronounced for Tower sel3 (B. napus) (43%) than it was for S-67 (B. carinata) (7%). With row sowing, on the other
hand, the two species responded differently in that early sown S- 67 (B . carinata) produced 28% more number of primary branches than
the lkte sown ones, but Tower sel3 (B. napus) produced 71% less•f«number of primary branches. At lower population density of late,
sowing Tower sel3 (B. napus) produced more number of primary branches per plant to compensate for the sparse stand. There appeared a synergistic effect of broadcasting and compensation for low population density in S-67 (B. carinata). Row sown S-67 (B.
carinata) was rather weaken instead of compensating for sparsity resulted from late sowing.
Irrespective of method of sowing, both S-67 (B. carinata)and Tower sel3 (B. napus), on the overall, produced more number of pods per plant under early than late sowing dates at Kulumsa (Figure 7 and Appendix 9). With percentage transformations of the mean differences, it was observed that early sown plants of S-67
IB. carinata) produced 8% more pods under broadcasting and 43% under row sowing than those sown late. Tower sela (B. napus), on
the other hand, produced 5 and 2% more pods per plant due to early
sowing under broadcasting and row sowing, respectively.Both species had similar responses to methods of sowing under
earlier sowing date. The same response trend was noted for the late sown Tower sels (£. natpus) unlike S-67 which with late sowing produced 34% more number of pods per plant under broadcast than row
sowing.This implies that early sowing of both species resulted in
higher number of pods per plant regardless of the methods of sowing. It also emphasizes the significance of broadcasting for S-67 which, with this method of sowing, could produce more than one third over the amount produced under row sowing even with the
aunfavourable sowing date condition. In addition, the response of S-67 {B. carinata) to favourable conditions of sowing date (early)was by far higher under less favourable condition of sowing method (row) and similarly that of favourable method (broadcasting) was under less favorable sowing date, implying further, that this
species can pay off with less amount of input, in this case either early sowing or broadcasting, for better number of pods per plant.
From Figure 8 and Appendix 10 it can be discerned that with early sowing, at Holetta, the 1000-seed weight of S-67 (B.
carinata ) was higher under row than broadcast sowings (0.58g) while Tower sel3 KB. napus) had the reverse tendency (0.03g). With late sowing, on the other hand, both S-67 and Tower sel3 had seeds
5 9
Num
ber
of
prim
ary
bran
ches
/pla
nt
1000
se
ed
wei
ght
(g)
60
B. Species by sowing date by ferti1i zer
____ Early # Late Early # Late Early # Laterassica Brassica Brassica Brassicacarinata napus carinata napus2
Fig. 8« Effects of the interactions between species, sowing date, sowing method and fertilizer on 1000-seed weight at Holetta. B = Broadcast, R = RowFO = With out fertilizer, F1 = With fertilizer
A. S?ecfes by sowing date by method sowi ng
LSDO.Ot = 0.447
os * 0.237
5 _Kulumsa
4F0 = No ferti1i zer
^ FI = With ferti1i z
LSD
0.010.05
Holetta Kulumsa
0.591 0.7780.445 0.587
F0 F1 F0 F1 F0 F1 F0 F1Early Late Early Late
Brassica carinata Brassica napusFig. 9. Effects of the interactions between species, sowing date and
fertilizer on number of primary branchas per plant.
61 i
heavier by 0.31 and 0.07g under broadcast than row sowing.Under broadcast, late sown S-67 (B. carinata) exhibited seeds
heavier by 0.33g than that of early sown ones. Tower sela IB.tnapus), however, had seeds heavier by 0.54g due to early than late
sowings«Under row sowing, both S-67 and Tower sel3 exhibited seeds
heavier by 0.56g and 0.58g when early than late sown. It can be noted that Tower sels (B. carinata) was affected more by sowing date than by method of sowing. Early sowing generally resulted in heavier seeds of this species. With S-67 (B. carinata), however,
both date and method were important in that under early sowing date, row sowing was more important while under late sowing date, broadcasting resulted in heavier seeds.
4.1.6.5, Species, sowing date and fertilizer (SDF); At Holetta, the interactive effect of species, sowing date and fertilizer significantly affected number of primary branches, days to maturity, number of pods per plant and 1000-seed weight. Further, highly significant interactive effects were discerned on days to flowering and number of seeds per plant. At Kulumsa, highly significant effects occurred on number of primary branches per plant, days to flowering and number of seeds per plant (Appendices2 and 3 ) .
Results in Figure 9 and Appendix 11 indicate that at both locations, S-67 (Brassica carinata) produced more number ofprimary branches per plant under early than late sowing dates while
62 ;
the reverse was evident for Tower sel3 (B. napus). Generally,
early sowing induced more number of primas^ branches under both
fertilizer regimes at both lofations.The response to fertilizer, however, varied with species and
sowing dates. At both locations, S-67 (B. carinata) showed moreresponse under delayed sowings (18 and 35% at Holetta and Kulumsa) than under earlier sowings with respective values of 9 and 20%. Conversely, Tower sel3 (B. napus) was more responsive to fertilizer under earlier sowing dates with values of 45% at Holetta and 75% at Kulumsa than under delayed sowings with corresponding values of
8 and 3%.The higher values of S-67 (B. carinata) under early sowing
and that of Towre sel3 (B. napus) under delayed sowing might be *associated with length of growth cycle; because the former is bjr - far a late maturing, more branchy and indeterminate type. Therefore, as more favourable growth period is provided by early sowing, it can fully utilize the period to manifest its potential. Tower sel3 (B. napus), on the other hand, is an early maturing variety with compact branching habit and more determinate. It, therefore, can reach the stage where its branches would normally develop without being significantly affected by delayed sowing but rather took advantage of reduced population density that resulted from delayed sowing, to develop more number of primary branches as compensatory factor. It can also be noted that S-67 (B. carinata)
can respond more to improved conditions of fertilizer or sowing/
date under poor condition of sowing date or fertilizer,
respectively, Tower sel3 (B. napus), however, performs at better
conditions of each factor. This implies that S-67 (B. carinata)is less demanding in terms of these management factors while Tower sel3 (B. napus) is more demanding as far as primary branches are
concerned.Fertilizer generally shortened the time to flowering of both
species at both locations (Figure 10 and Appendix 12). It shortened that of early sown S-67 (B. carinata) by four days andof Tower sel3 ( B. napus) by 13 days at Holetta and with corresponding values of 8 and 11 days at Kulumsa.
Similarly, late sown plants of the two species flowered 9 and 15 days earlier at Holetta but 7 and 5 days earlier at Kulumsa.
Irrespective of sowing date, both species flowered about the same time at Holetta when grown under conditions of no fertilizer; but. ?.
with fertilizer, it was much shortened by early sowing for Tower sel3 (B. napus) than it was for S-67 (B. carinata), the formerbeing early by eight days. At Kulumsa, however, early sowing
without fertilizer shortened the time to flowering of Tower sel3 (B . napus) by nine days and with fertilizer, by seven days. It can be noted that at Holetta the reduction in days to flowering by fertilizer application was higher with late sowing while at Kulumsa the reduction was by far greater with early sowing. Further, it was possible, at Holetta, to shorten the time to flowering of Tower sel3 (B. napus) through fertilizer application to a greater extent than it was possible for S-67 (B. carinata).
63 i
Day
s to
mat
urity
D
ays
to fl
ow
erin
g
9 0 r
640.010.05
LSD Holetta Kulumsa
1 .‘69 1.41 3.362.54
E B B Holett
Hill KulumsF0 = Wi thout
ferti1i zer
F1 = With
ferti1i zer
120
Fig.
F0 F1 FO F1 Early Late
Brassioacarinata
11
F0 F1 FO F1
Early Late FO FI FO
Brassioa Early Late
napusEffects of the interactions between species, sowing date, fertilizer and weed control on days to maturity.
60FO F1 FO F1
Early Late
Brassioa carinataFig. 10*Effects of the interactions between
days to flowering.
FO F1 FO F1
Early Late
Brassioa napusspecies, sowing date and fertilizer on
180
1 1
162
FO = Without fertilizer LSD
F1 = With fertilizer 0.01 = 5.02
WO = With no weeding 0.05 = 3.78W1 = With weeing
r A. Sepecies by sowing date by fertilizer (Hoi
B. Sowing date by fertilizer by weed control (Kulumsa)
Fertilizer generally shortened days to maturity of Tower sel3
\(B. napus) under both sowing dates at Holetta. It reduced days to
maturity of this species by about 15 days for the early and one month for the late sowing dates. Delayed sowing increased days to maturity of Tower sela (B . napus) by 18 days under no-fertilizer condition but no effect of sowing date was noted under fertilized condition. Neither fertilizer nor sowing date imposed any noticeable change in the time to maturity of S-67 (B . carinata) , (Figure 11 and Appendix 13). Like days to flowering, the time to
maturity of Tower sel3 (B. napus) can be manipulated by fertilizer or adjustment of sowing date. Fertilizer appeared to play an appreciable role in reducing length of the growth period, particularly of late sown Tower sels (B. napus) under conditions where late sowing is the only option.
Both S-67 (B. carinata) and Tower sel3 (B. napus) produced\
greater number of pods per plant under fertilized and early sowing date conditions of Holetta (Figure 7 and Appendix 9). Furtherexamination of Figure 7 reveals tremendous variations in the magnitudes of the differences between the two sowing dates of the two species under the two fertilizer levels. Delayed sowing under no-fertilizer condition reduced the number of pods per plant of S- 67 (B. carinata) by 20% but increased that of and Tower sel3 (B.
napus) by 3%. With fertilizer, however, there was only a slightreduction of 2.7% in S-67 whereas in Tower sel3 there wasconsiderably higher reduction of 17%.
The response of the two species to fertilizer also varied with
6 5
/
66 ‘
sowing dates. S-67 (B. carinata) showed a marked response (38%)
under delayed sowing than under earlier sowing date (18%). The
response to fertilizer of early sown Tower sels (B . napus) was 52%
more pods per plant as compared to 25% under delayed sowing. The
result makes it clear that S-67 (B . carinata) responded to earlysowing at an appreciably higher magnitude under no fertilizer than
/with fertilizer conditions, while the reverse held true for Tower
sel3 (B. napus). Similarly, the response of the species to fertilizer was in a reverse fashion. S-67 had higher number ofpods with delayed sowing whereas Tower sel3 had with early sowing.
IOnce again, this underlines the fact that S-67 (B. carinata) canpay off substantially with less amount of input interms of fertilizer or early sowing. Tower sel3 (B . napus), on the other
hand, is more demanding. „
It is apparent from Figure 12 and Appendix 14 that the number of seeds produced by both species varied consistently in response to the different sowing dates and fertilizer levels.
At both locations, fertilizer increased seeds per plant of both species under both sowing dates. At Holetta, the responses of the species to fertiliser were much more higher (74% for S-67 and 99% for Tower sel3) under delayed sowing than under earlier sowing dates (27 and 66% for the two species, respectively). The reverse was true at Kulumsa with corresponding values for S-67 (B .
carinata) and Tower sel3 (B. napus) of 32 and 28% under delayed sowing; and 101 and 88% under earlier sowing dates. At Holetta, Tower sels (B. napus was more responsive to fertilizer than S-67
No.
of
po
ds/p
lan
t N
o.
of
seed
s/pl
ant
2500
2280
2060
1840
1620
LSD Hoietta Kulumsa
0 . 0 1 1 0 8 8 90 . 0 5 8 1 6 7
9 BI Holetta
F0 F1 F0 F1 FOEarly Late Early # Late
Brassvca carinata Brassica napusFig. Alt Effects of interactions between species, sowing date and
fertilizer on number of seeds per plant.
Bill Kuluns;
= With no ferti1i z<
= With fertilizer
50W 0 W 1 W O W 1 w o W 1 w o W1
F0 F1 FO F1Ear]y Late
Fig. 13,Effects of the interactions between sowing date, fertilizer and weed control on number of pods per plant at Kulumsa.
170 -
158 - 146 -
LSD
0.01 = 17.4 0.05 = 13.1
WO = With no weeding W1 = With weeding
FO = With no fertilizer
F1 = With fertilizer
under both sowing dates; but at Kulumsa, t,he reverse was true for
S-67 (B. carinata). This trend was also reflected for the two*
sowing dates. At Holetta, under condition of no-fertilizer, S-67 (B. carinata) produced 48% and Tower sel3 (B. napus) 107% more
seeds with early sowing than with late sowing. Under fertilized condition, there were corresponding values of (8% S-67) and 73% (Tower sel3 ) increases with early sowing.
At Kulumsa, S-67 (B. carinata) produced 61 and 6% more seeds due to early sowing under fertilized and non-ferti1ized conditions, respectively. Tower sle3 (B. napus), on the other hand, tended to produce more (14%) seeds per plant due to delayed sowing under no fertilizer condition, Under fertilized condition, however, there were 29% more seeds per plant due to early sowing.
At Holetta where growth is relatively slow and soil phosphorus* level is much lower (about 6 ppm) than at Kulumsa (about 22 ppm) the response to fertilizer was more prominent with delayed sowing. At Kulumsa, however, there appeared a synergistic effect of early
sowing and fertilizer application. At Holetta, there was more response to early sowing of Tower sel3 (B . napus) than S-67 (B.
carinata) with the higher response occurring under no-fertilizer condition.
At Kulumsa, the late maturing S-67 (B. carinata) enjoyed the long growing period provided by early sowing with ample nutrients from the fertilizer application and thus grew more luxuriously which resulted in more number of seeds per plant. Tower sel3 , on the other hand, under no-fertilizer condition had more seeds per
68
plant with delayed than early sowings. This might be due to compensation of seed number for population density which was
significantly lower under delayed than early sowing.However, there appeared a synergistic effect of fertilizer
and early sowing.At Holetta, fertilizer generally increased the weight of the
seeds of both S-67 (B. carinata) and Tower sels ( B. napus) so did early sowing date (Figure 8 and Appendix 10 )v The magnitude of the
species response to sowing date, however, varied with fertilizer,. S-67 (B. carinata) showed higher response to sowing date underfertilized than non-fertilized conditions.
Early sowing increased 1000-seed weight of S-67, when fertilized, by 0.18g and when not fertilized by 0.04g. On the other hand, Tower sel3 (B. napus) was more responsive to sowing,
date under non-fertilized condition, Early sowing increased 1000- seed weight of non-fertilized plots by 0.91g and that of fertilized plots by 0.22g. In similar manner, the two species responded to fertilizer in opposing fashion. S-67 (B. carinata) responded more with early sowing than with delayed sowing where fertilizer increased 1000-seed weight of this species by 0.17g when early sown and by 0.03g when late sown. As opposed to S-67, the response of Tower sel3 (B . napus) to fertilizer was 0.08g under earlier and O.llg under delayed sowing dates. The longer maturing species, S- 67 (B. carinata) needs both fertilizer and early sowing to produce heavier seeds. This is may be because early sowing, with
sufficient amount of nutrients to synthesize more protoplasm for
69 ‘
large seeds, creates favourable environment during grain filling period. The effect of fertilizer with delayed sowing in Tower sel3
(B. napus), apart from its direct effect on the size of the seed through the development of cells and constituents might be due to the shortening of the time it takes for flowering and thereby increasing the period for steady grain-filling (Table 1).
4.1.6.6. Sowing date, fertilizer and weed control (DFW): Highlysignificant effect of the interaction between date of sowing, fertilizer and weed control was observed on the number of seeds
per plant at Holetta; number of pods and seeds per plant at Kulumsa. In addition, days to maturity was significantly affected at Kulumsa (Appendices 2 and 3).
Figure 11 and Appendix 13 reveal that under delayed sowing at^ Kulumsa, fertilizer has shortened the time to maturity of both weeded and unweeded plots by 11 and 12 days, respectively. Under early sowing date, however, the non-fertilized unweeded plots matured two days earlier than the fertilized ones; whereas the weeded fertilized plots matured three days earlier. Nevertheless, the effect of fertilizer with early sowing, irrespective of weed
control, was agronomically unimportant. It was indeed of paramount significance with delayed sowing where oilseed brassicas were sown at the fall of the optimum dates.
Under conditions of no-fertilizer, early sown weeded and
unweeded plots matured three days earlier than the corresponding
late sown ones. Conversely, the late sown fertilized plots were
70 i
’ j-"-- r •••.1 ' •- ecdeu ' '• r’ ‘ r +’h .n the early sown ones.
Once a.; - ' . *\ ’ ”oc ®' f i ?v + impor tance of fertilizer to1 v ~ irity of brassicas. The effect of early
■-v'mg in reducing this time, although statistically significant, was agronomically less important.
Based on Figure 13 and Appendix 8, fertilizer on the overallincreased the number of pods per plant at all levels of sowing date
and weed control at Kulumsa. Further, with early sowing,fertilizer increased the number of pods per plant by 69 and 54%under conditions of no weeding and weeding, respectively. With
late sowing, the increase of the unweeded plots was 64%. while theless than one percent increase in weeded plots was notagronomically important. It can be noted that pod development was *
highly promoted by fertilizer under early sowing date conditions ‘ irrespective of the weed situation. But with delayed sowing, fertilizer is an indispensable input for higher number of pods per plant under no weeding condition.
With no weed control practice, early sowing increased pods per plant of fertilized plots by 36% and that of the non-f ert il ized plots by 32%. With weed control, however, early sowing was advantageous under fertilized condition because it increased the character by 23% whereas it reduced those of non-ferti1ized plots by the same magnitude.
The higher number of pods per plant due to early sowing under no weed control practice might be because of the fact that early sowing increased the crop stand right from the early stage
-1
(seedling density) so that the crop could suppress the weeds and
became more vigorous resulting in many pods. With weed control
but fertilizer, however, there appeared a decline of pod number due to early sowing. This might have been so because of increased population density which could lead to mutual shading and competition and hence reduced pod number.
Fertilizer generally increased the number of seeds per plant at all levels of sowing date, weed control and locations (Fig.14 and Appendix 15). With early sowing and no weeding it has increased number of seeds per plant by 63% at Holetta and by 110% at Kulumsa compared to the 33 and 87% respective increases with
weeding under the same sowing date. With delayed sowing and no weeding the values were 74% (Holetta) and 68% (Kulumsa) in contrast
to the values obtained under weeding (92% Holetta and 17% Kulumsa).In similar manner, weed control generalLy resulted in higher,
number of seeds per plant. With early sowing and no-fertilizer, weeded plots produced 34% at Holetta and 58% at Kulumsa, more seeds per plant than those of unweeded plots. With e a r l y sowing and fertilized condition, weeding increased number of seeds per plant by 9 and 40% at Holetta and Kulumsa, respectively. Si m i l a r l y ,
with late sowing but no fertilizer, weeding increased seeds per plant by 8% (Holetta) and 82% (Kulumsa), while with fertilizer the number of seeds per plant increased by 19 and 27% at Holetta and Kulumsa, respectively.
From the results one can note that, at Kulumsa under conditions of both sowing dates when the population density of the
No.
of se
eds/
plan
t
73
Hoietta Kulumsa
WO W1 WO W1FO F1
EarlyFO F1
H o le t t
Kulum: s a
FO = With
F1 = Wi thWO = With
W1 = With
Late
Fig. H * Effects of the interactions between sowing date, fertilizer and weed control on number of seeds per plant.
crop is expected to reduce due to lack of either fertilizer or weed
control, the number of seeds per plant increased at a higher rate
upon provision of the other input, weed control or fertilizer. The same held true at Holetta for the early sown plots. With late sown plots, however, the response was higher when both the inputs (fertilizer and weed control) were provided. This might be
associated with the vigor and earliness of seedling establishmentf
and compensation for less population density later in the growth cycle. Early sowing at Holetta coupled with relatively higher natural fertility level of the soil at Kulumsa (0.22% N and, 22 P- ppm) might have enabled the seedlings to establish at better
condition. Even with late sowing, the seedling establishment was greater at Kulumsa which again may be due to higher nutrient level of the soil and - better climatic conditions specifically, temperature and rainfall, (Appendix 1). At Holetta, however, delayed sowing resulted in delayed seedling establishment which were not strong enough to out compete weeds. Therefore, plants at Kulumsa and those early sown at Holetta took the advantage of fertilizer or weed control and developed more seeds per plant than the ones which received both fertilizer and weed control and hence were densely populated. The weak plants from delayed sowing at Holetta, however, demanded both fertilizer and weed control to produce greater number of seeds per plant.
It can also be noted that the response to weed control in all combinations of sowing date and fertilizer was higher at Kulumsa than it was at Holetta; so was the response to fertilizer under
early sowing irrespective of the weed situation. But the response to fertilizer under delayed sowing'was higher at Holetta than it
was at Kulumsa, regardless of the weed control. Further, at both
locations, under early sowings, fertilizer appeard more important than weed control for number of seeds per plant, while under delayed sowing, fertilizer remained to be important at Holetta but weed control took the lead at Kulumsa.
4.2. Effects of the Considered Factors on Seed yield. Oil content and Oil yield
4.2.1, Species
The average differences in seed yield, oil content and oil yield between Brassica carinata (S-67) and B. napus (Tower sel3 ) are given in Table 2. At both locations, the effect of species in all the three characters was highly significant (Appendices 2 and 3).
Brassica carinata (S-67) gave an average seed yield of about 13 q/ha at Holetta and 26 q/ha at Kulumsa as compared to thecorresponding 9 and 11 q/ha of B. napus (Tower sel3 ) . Brassica
napus (Tower sel3) produced 3.3% at Holetta and 3% at Kulumsa, moreoil than B. carinata (S-67). With respect to oil yield, however,B. carinata (S-67) produced 133 and 520 kg/ha more than B. napus
(Tower sel3) at the two respective locations.
7 5
Table 2. Seed yield, oil co&ieit and oil yield as affected by species, soviig date, sowing le’iod, fertiliser and seed control at Holetta m f a h i s a
76
Seed yield k.g/k& Oil content S Oil yield kg/ha
Holetta [al'iisa Holetta [tthisa Holetta. Jaluisa
Jrassica carinata fS-671 1252 2572 33.1 37,7 501 970hussies m\is ("oser s e b j 85; 1105 42.4 40.7 358 450
Early sowing 1438 m i 41.4 38.3 53: 348Late sowing 712 1203 40.0 40.1 m 482
With no weeding 958 1406 41.0 3S.4 337 553Witt weeding 1180 2271 40.4 39.0 478 885
Kean iQTs 1839 40.7 33.2 435 717
C . 7,1 15 g 2 3 1c 10
58 0.545 0.660 of
77
With these differences among the factors included, species
stood as the third, following fertilizer and date of sowing, important factor to induce variabilities in seed yield and oil yield at Holetta. At Kulumsa, it took the lead inducing the largest amount of variability observed in these characters.
The results were in agreement with the findings of Mizushima,
jet a l . , (196 7), Hiruy, et ai. , ( 1986 ) and Hiruy ( 1987 ) who reported higher levels of seed yield and oil yield but lower oil content in B. carinata than in B. napus cultivars.
The reason that S-67 ( B. carinata) gave by far greater seed
yield than Tower sel3 (B. napus) may be because of its enormous growth which was manifested through increased height, pod bearing branches, pods and seeds per plant and longer grain-filling period (Table 1). Although the oil percentage in S-67 appeared# significantly lower than that of Tower sel3, the oil yield from S- 67 was higher than Tower sel3 both at Holetta and Kulumsa because the exceedingly higher seed yield from the former species overcompensated the loss in oil content.
4.2.2. Date of sowing
At both Holetta and Kulumsa, highly significant effects of sowing date were observed on seed yield, oil content and oil yield (Appendices 2 and 3).
The results presented in Table 2 indicate that early sowing resulted in higher seed yield than late sowing which caused seed
yield losses of 102% (Holetta) and 106% (Kulumsa). This is below the respective average values of about 14 and 25 q/ha that could
have potentially been realized through the optimum time of sowing
at Holetta and Kulumsa. Early sowing also increased oil content by 1.4% at Holetta but reduced the same by 1.8% at Kulumsa. Yet, it did increase oil yield considerably at both locations; with increases of 310 and 466 kg/ha at Holetta and Kulumsa, respectively. With these impacts, sowing date appeared to be the second most important factor determining higher seed yield next to fertilizer (Holetta) and species (Kulumsa).
The effect of sowing date on seed yield and oil yield at both locations and oil content at Holetta resemble the findings of -Gross, et al* » (1966), Thurling (1974 a), Mendham, et a l . , (1975 ),Tayo, et a l . , ( 1975 ), Ali-Khan (1976 ), Kondra(1976 ) , Degnehardt,
et a l ., (1981 a , b ) , Auld, et. al . , (1984) and Christensen, et al . , 4(1985) in that they have reported increased amounts of seed yield, oil content and oil yield due to early sowing of oil seed brassicas.
By creating more favourable moisture and temperature conditions for the growth and development of the crops, early sowing promoted the production of seed and oil. The lower oil content observed at Kulumsa as a result of early sowing might be due to the higher (> 23°c ) temperatures that prevailed towards the maturity of the crop (late November) (Appendix 2). 0 i 1deposition is active at the later ten days and is sensitive to temperature in brassicas (Downey, 1971; Appelqvist, el al . . 1 972).
However, this decline in oil content at this site was off-set by trie exceedingly higher seed yield which made up the oil yield from
early sowing to be almost twice as much from late sowing and this
further signifies the importance and indispensability of early
sowing.
4.2.3* Method of sowing
The effects of sowing method on seed yield and oil yield were highly significant at both Holetta and Kulumsa (Appendices 2 and 3). Results in Table 2 indicate that at Holetta, row sowing conferred about 13% yield advantage over the average value of about10 q/ha of broadcasting. At Kulumsa, however, broadcasting was more advantageous than row sowing by the same magnitude with respective values of about 19 and 17 q/ha. At Holetta, row sowing-
conferred an advantage in oil yield of 51 kg/ha; while at Kulumsa, broadcasting was advantageous by 79 kg/ha.
Among the factors included at both locations, method of sowing was the least important factor as far as both seed yield and oil yield are concerned. The results from Holetta agree with the results of Fulkerson (1971) who reported increased seed yield and oil yield in Brassica napus with row sowing than broadcasting and with the results of Patil (1979) who found increased seed yield and oil yield in nigerseed (Guizotia abyssinica) due to row sowing. The results from Kulumsa are in support of the works of Kondra (197 5), Clarke e_t al . , (1978), Jain, et al . , (1979) and Anonymous
79
11981, 1985) where by increased values of seed yield and oil yield
were obtained with broadcasting than row sowing. This implies that the effect of method of sowing on seed and oil yields vary with
environment. The increases at Holetta might be mainly via plant height and 1000-seed weight which were also appreciably increased bv row sowing. At Kulumsa, on the other hand, it might be mainly through increased number of pod bearing branches, number of pods and seeds per plant which were also increased significantly by
broadcasting (Table 1).
4.2.4. Fertilizer
The effects of fertilizer on seed yield and oil yield were found to be highly significant both at Holetta and Kulumsa; while the effect on oil content was significant only at Holetta
(Appendices 2 and 3).It can be discerned from Table 2 that seed yield was increased
by 219% (Holetta) and 77% (Kulumsa) due to fertilizer. This is in contrast to the respective 16 and 13 q/ha obtained from nonfertilized plots at Holetta and Kulumsa. Fertilizer increased oil content by 1.3% and oil yield by 471 kg/ha at Holetta and 409 kg/ha at Kulumsa. As a consequence, fertilizer stood foremost in inducing the largest amount of variability in seed yield observed at Holetta, but the third important factor, after species and
sowing date at Kulumsa.
Higher seed and oil yields in brassicas due to fertilizer have been reported by several workers including Ractz, et al.., (1965),
Allen, et al • > (1971), Krogman, et a l . , (1975 ), Aulakh, a l . .
(1980), Holmes (1980), Sheppard, et a l ., (1980), Joarder (1983),
Hiruy et .al. , ( 1983, 1986, 1987) and Nuttal, ejt al. . (1987 )substantiating what the present study revealed. The reasons might be that plants under fertilized conditions possessed larger framework (height and pod bearing branches) on which more number of pods and virtually seeds could develop (Table 1).
The higher oil content noted from fertilized plots was notconsistent with the works of Dybing (1964), Krogman, et. a l . ,(1975 ), Henery, et aj,. , (1978) and Joarder (1983) where by theyreported decreased oil content from nitrogen fertilizer but no
*effect of phosphorus. The differences of the results mighty
therefore, be due to the use of both nitrogen and phosphorus in this study whereas the latter may have diluted the negative effect of nitrogen. The differences might also_ be due to an indirect effect of fertilizer on seed size,in that fertilizer increased the seed size (Table 1). Larger seeds of oil seed brassica, for thatmatter other oil seed also, have proportionally higher oil content(Appelqvist, et. a_l. , 1972 ).
4.2.5. Weed Control
Highly significant effects of weed control occurred on seed yield and oil yield at both locations, and on oil content at
8 1 .
✓Holetta only (Appendices 2 and 3). At both locations, weed control
appeared to be the fourth important factor following fertilizer and
sowing date at Holetta; and species and sowing date at Kulumsa', because there were potential seed yield losses of about 22 and 62% below the corresponding average values of 12 and 23 q/ha of weeded plots at Holetta and Kulumsa (Table 2). On the average, unweeded
plots at Holetta produced 0.6% more oil than weeded plots. At both locations, however, weed control increased oil yield by 81 and 333 kg/ha at Holetta and Kulumsa, respectively.
Lower seed yield and oil yield in oilseed brassicas due to weed competition were reported also by Forbes (1985 a), Jain, et al. , (1986 ), Kaul, et aJL. , (1979 ) and Kumar, et al.. , (1989). Itwas observed that when oil seed brassicas were grown under
conditions of no-weeding, the competition from the weed reduced- 2 significantly the number of seedlings, number of harvestable plants and the height of the crop. It also virtually reduced the number of primary branches, number of pods and number of seeds per plant (Table 1) which might have possibly contributed to the reduction of seed and oil yields observed at both locations.
The higher level of oil content obtained under no- weeding conditions of Holetta and even of Kulumsa, although not statistically significant (Table 2), is difficult to explain with the data at hand and, therefore, needs further in-depth invest igations♦
82 1
4.2.6. t Interaction of factors
4.2.6.1. Species and date of sowing (S D ): The analyses ofvariance (Appendices 2 and 3) show that at both locastions oil
content was affected highly significantly by the interaction of species and date of sowing. Both seed yield and oil yield at both
locations were not affected by the interaction at a significant level .
It can be seen from Figure 15 and Appendix 16 that early sowing resulted in higher oil content of both species at Holetta. The increase in Brassica carinata (S-67), however, was not that impressive (0.4%); while in B. napus (Tower sel3) the 2.4% increase
Iwas substantial. At Kulumsa, on the othr hand, late sowing seemed to induce more oil accumulation in both species, the increases being 3.2% for B. carinata (S-67) and 0.5% for B. napus (Tower sel3 ) . Further, in contrast to, what has been observed at Holetta, B. carinata (S-67) showed a greater vriation due to sowing date
differences.The increase in oil content of both species due to early
sowing at Holetta might be associated with the snowballing effect of seed size on oil content (Appelqvist, et. al . , 1972) wherebylarger seeds of oilseed brassicas proportionally possess higheroil contents, The seed size, as measured by 1000-seed weight, at Holetta was significantly higher with early than late sowings. AtKulumsa, oil content was higher with late sowing date. This is
\perhaps because of the relatively lower temperature (about 21°C)
8 3 i
Oil
Yiel
d (k
g/h
a)
Oil
con
ten
t {%
)
4 1
39
37
Fig. 15.
1500 r
1360
1220
1080
940
800
660
520
380
240
10 O
LSD
0.010.05
Hoietta
5139
Kulumsa
4735
Holetta
Kulums<
With no fertilizi With fertilizer
Fig. 16
D1 D2 D1 D2 FO F1 FO F1
Brassica Brassica Early Latecarinata napus
Effects of the interactions between species, sowing date and
fertilizer on oil content.
A. Species by sowing date B. Sowing by fertilizer
0.490 0.933
0.369 0.704
Holetta Kulumsa
iM H I Holetta
■ Kulums
D1 = Early
D2 = Late
= With no fertiliz
= Wi th ferti1i zer
FO F1 FO
Brassica carinata Brassica napus, Effects of the interactions between species and fertilizer on
oil yield.
conditions that prevailed towards the maturity of the late sown(crops as compared to the higher (about 23° C) of the early sown
crops (Appendix 1). This is because higher temperatures towards the last ten days of maturity reduce the accumulation of oil in oilseed brassicas (Downey, 1971).
4.2.6.2. Species and fertilizer (SF):At both locations, oil yield was highly and significantly affected by the interaction of species and fertilizer (Appendices 2 and 3). Figure 16 and Appendix 17 reveal that at both locations, Brassica carinta (S-67)
and B. napus (Tower sel3 ) produced more oil yield under fertilized than non-fertilized conditions. Fertilizer increased oil yield of S-67 and Tower sel3 by 522 and 411 kg/ha at Holtta; and by 484 and 328 kg/ha at Kulumsa. ^
It is apparent that at both locations, Tower sel3 (B. napus)
was more responsive to fertilizer than S-67 (B . carinata) implying that the B. napus cultivar requires more input, in this case fertilizer, than the B. carinata cultivar. The increase in oil yield as a result of fertilizer application at Holetta might be due to the increased seed yield and oil content (Table 2). At Kulumsa, although the oil contents of both fertilized and non-fertilized plots were comparable, (Appendix 4) the exceedingly higher seed yield from fertilized plots (Table 2) overcompensated for the loss resulting from lower oil content.
85 ‘
4.2.6.3. Sowing date and fertilizer (D F ): The interactive effectof sowing date and fertilizer on oil content was highly significant at Holetta and significant at Kulumsa (Appendices 2 and 3). Figure 15 and Appendix 16 show that at both locations, early sown plots had more or less the same amount of oil inresponse to fertilizer with only a slightly higher amounts (0.4%) from the non- fertilized plots at Kulumsa. With delayed sowing dates, fertilizer increased oil content by 2.7% at Holetta and 1.3% at Kulumsa.
At Holetta, early sowing increased oil content of non- fertilized plots by 2.7% and plants under fertilized conditions
were not responsive to sowing date as it pertains to oil content. At Kulumsa, however, plants under both levels of fertilizer produced more oil with delayed sowing, but higher concentration (2.6%) was produced under fertilized than unfertilized (0.9%f
conditions.It can be noted from the results that fertilizer was important
for higher oil content only under delayed sowing dates at both locations. The same was true for early sowing which increased oil content at Holetta under zero level of fertilizer while at Kulumsa both delayed sowing and fertilizer were important for higher oil
content.The negative effect of early sowing and that of fertilizer
under early sowing date observed at Kulumsa might be related to prevailing temperature condition. The temperature during the last few days of maturity of the early sown plants was relatively higher than that which prevailed during this time of the late sown ones
87 ;
(Appendix 1). Yet, it was found that higher temperatures (>20°G) during the last ten days of maturity decrease oil content (Downey,
1971; Appelqvist, et a l . , 1972). The effect of fertilizer, by1virtue of its effect in reducing the time to maturity, even exposed the early sown plants to higher temperature conditions during the
critical stage of oil accumulation.
4.2.6.4. Species, sowing date and fertilizer (SDF): The effect ofthe interaction between species, sowing date and fertilzier on seed yield was highly significant at Kulumsa only (Appendix 3).
It can be descerned from Figure 17 and Appendix 18 that both Brassica carinata (S-67) and B. napus (Tower sel3) had higher seed yields in response to fertilizer, even when late sown. The
magnitude of response for the two species, however, varied with
sowing date.With early sowing, B. carinata (S-67) had a response to ferti
lizer of 73% while B. napus (Tower sel3 ) had 114%. With late sowing, more seed yield response of 47% (B . carinata) and 110% ( B. napus) was possible than would be obtained from unfertilized
conditions.In a similar manner, both species responded positively to
early sowing under both fertilizer levels. Under unfertilized conditions,B. carinata (S-67) produced 117% and B. napus (Tower sel3 ) 47% more seed yield than when late sown. Under fertilizedcondition also both B. carinata (S-67) and B. napus (Tower sel3)
Seed
yi
eld
(kg/
ha)
S
eed
yiel
d (k
g/h
a)
oo
_ A. Species by sowing date by ! fertilizer
B. Species by sowing date by weed control
-I'd50 t"I4 100 r
3650
3200
2750
2300
1850
1400
950
500
LSD
0.01 = 136 0.05 = 103
JBBg.FO F1 FO F1 FO F1 FO F1Early Late Early Late
Brassioa Brassioacarinata napus
Fig. 17. Effects of the interactions between species, sowing date, fertilizer and
weed control on seed yield at Kulumsa.
4500
4070
3640
3210
2780
2,350
1920
1490
1060
630
200
r A. Species by method of sowing by
ferti1i zer
B. Species by method of sowing by
weed control
LSD Hoietta Kulumsa
Holette
Kulums
B = BroadcastR = Row
FO = With no fertiliF1 = With fertilizerWO = With no weedintW1 = With weedi ng
FO F1 FO FI FO F1 FO F1 B R B R
Brassioa Brassioacarinata napus
WO W1 wo W1 wo W1 wo wiB R B R
Brassica Brassicacarinata napus
Fig. 1 8 r Effects of the interactions between species, method of sowing,, fertilizer and weed control on seed yield.
WO W1 WO W1 WO W1 WO WI
Early Late Early Late Brassioa Brassioacarinata napus
FO = Without fertilizerF1 = Wi th ferti1i zer
WO = With no weeding W1 = With weeding
produced 115 and 49% more seeci yield, respectively, due to early
sowing.The result indicates that the response to fertilizer was by
far greater in B. napus (Tower sel3 ) than it was in B. carinata (S-
67), implying that under the prevailing conditions of Kulumsa, S-
67 is superior to Tower sel3 in a sense that it could produce substantial amount of seed yield even without fertilizer
applicat ion.The response to sowing date was exactly the opposite to what
was true for fertilizer in that B. carinata (S-67) produced by far greater seed yield with early sowing than with late sowing. B.
napus (Tower sel3 ), on the other hand, showed exceedingly less response to sowing date than B. carinata which implies that the yield sacrifice in B. napus due to delayed sowing is not as severe as it is in B. carinata. In general terms, one can say that B.
carinata (S-67) is more sensitive to sowing date than to fertilizer
while the reverse holds true for B. napus (Tower sel3).
4.2.6.5 . Species, method of sowing and fertilizer (SMF): By theinteraction of species, method of sowing and fertilizer, seed yield was affected significantly at Holetta and highly significantly at Kulumsa (Appendices 2 and 3).
The results presented in Figure 18 and Appendix 19 indicate that under both methods of sowing, fertilizer has increased seed yield of both species. Fertilizer increased seed yield of broadcast sown Brassica carinata (S-67) and B. napus (Tower sel3 )
by 197 and 193% at Holetta and by 53 and 126% at Kulumsa. When row
sown, it increased seed yield of S-67 and Tower sel3 by 243 and 269% at Holetta with respective increases of 80 and 98% at Kulumsa.
At Holetta, row sowing increased the seed yield of fertilized and non-fertilized B. napus (Tower sel3) by 37 and 9% and that of the fertilized B. carinata (S-67) by 5% but reduced that of non
fertilized B. carinata (S-67) by 4%.At Kulumsa, broadcasting under conditions of no-ferti1izer
increased the seed yield of B. carinata (S-67) by 27% while it had almost negligible (1%) reduction in B. napus (Towr sel3). When fertilized, broadcast sowing increased the seed yield of both B.
carinata (S-67) and B. napus (Tower sel3) by 8 and 13%,
respectively.The results indicate that the response to fertilizer of both
species was higher at Holetta than at Kulumsa which might be due to the low soil fertility of Holetta as compared to Kulumsa (Table P. 24). Further, the response of B. carinata (S-67) to fertilizer was lower than that of B. napus (Tower sel3 ) implying that S-67 can perform better than Tower sel3 even under no-fertilizer conditions.
At Holetta, row sowing was more advantageous with respect to seed yield of B. napus (Tower sel3 ) and fertilized B. carinata ( S- 67) but under unfertilized condition seed yield of S-67 was more with broadcasting. This might be because of the spares population under broadcasting which, however, resulted in increased number of pod bearing branches, pods per plant, seeds per plant and seeds per pod obtained under this sowing method (Table 1). At Kulumsa,
90
broadcasting of both species was more important than row sowing
with both levels of fertilizer-.
4. 2.6.6. Species, date of sowing and weed control (SDW); Seedyield, at Kulumsa, was affected highly significantly by the
interaction of species, date of sowing and weed control (Appendix3 ) .
Early sowing increased the seed yield of unweeded and weededBrassica carinata (S-67) by 165 and 125% and that of unweeded andweeded B. napus (Tower sela) by 80 and 33%. This indicates thatthe long maturing B. carinata is more sensitive to sowing datethan the early maturing B. napus (Tower sel3 ) (Table 1).
In a similar manner, weeding increased early and late sown B. carinata (S-67) by 50 and 76% and that of B. napus (Tower sel3 ) by 53 and 107%. Although the magnitude varied, both species responded to weed control more under late than early sowing. The response of B. carinata (S-67) was not that impressive as the response of B. napus (Tower sel3 ) . This indicates once again that B. carinata
(S-67) is more tolerant to weed competition than the introduced B. napus cultivar, Tower sel3 (Figure 17 and Appendix 18 ).
4.2.6.7, Species, method of sowing and weed control (SMW): Theeffect of the interaction of species, method of sowing and weed control, on seed yield was significant both at Holetta and Kulumsa (Appendices 2 and 3)»
91
Figure 18 and Appendix 20 reveal that at Holetta, row sowing
increased seed yield of weeded and unweeded Brassica napus (Tower
sel 3 ) with respective values of 24 and 38% and that of weeded B.
carinata (S-67) by 5% but there was no effect on the seed yield of
unweeded B. carinata (S-67). At Kulumsa, broadcasting B . carinata
(S-67) increased seed yield of unweeded by 23% and weeded by 10%
while that of unweeded B. napus (Tower sel3 ) by 3 7%; but it
reduced seed yield of weeded B. napus (Tower sel3) by 6%.
The results indicated that at Holetta sowing method had
magnificent effect on seed yield of B. napus (Tower sel3 5 but only
slight or none on seed yield of B. carinata (S-67) which signifies
the fact that B. napus is more responsive to management factors
hence is more demanding than B . carinata (S-67). At Kulumsa as
well the lower yield from weeded B. napus (Tower se l3) with
broadcasting underlines the fact that this species performs better
with row sowing while B. carinata (S-67) produces substantial yield
.just with weeding alone.Weed control increased seed yield of broadcasted B. carinata
(S-67) by 17% and B. napus (Tower sel3) by 32% at Holetta and by
49 and 45%, at Kulumsa respectively. With raw sowing, yield was
increased by 23% (S-67) and 19% (Tower sel3 ) at Holetta with
respective increases of 67 and 110% at Kulumsa, due to weed
cont r o l ,At Kulumsa, both species responded more to weed control under
row than broadcast sowing once again indicating that row sowing at
this location is rather less important particularly for B. carinata
(S-67) than broadcast sowing.
4 .2 .6 .8. Sowing d ate; fe rtilizer and weed control (DFW) :Appendices 2 and 3 indicate that highly significant interactions
of sowing date, fertilizer and weed control were noted on seed
yield and oil yield at Holetta, while a significant effect occured
at Kulumsa.At all levels of sowing date and weed control, fertilizer
increased seed yield at both locations (Figure 19 and Appendix 21).
With early sowing, fertilizer increased the average seed yield of
unweeded plots by 180% at Holetta and by 116% at Kulumsa; and that
of weeded plots by 118 and 64%, respectively. With late sowing,
it increased seed yield of unweeded and weeded plots by 470 and #
629% at Holetta and by 80 and 60% at Kulumsa, respectively. £
The results indicate that with early sowing, fertilizer was
more important under unweeded than weeded conditions of both
locations. The same held true with late sowing at Kulumsa but not
at Holetta where tremendously higher response to fertilizer was
observed under weeded condition implying that late sown crops were
not vigorous enough to make efficient use of the fertilizer applied
but when the weeds were removed the stress was some how minimized
to result in better nutrient uptake which resulted in improved
g r o w t h .Further, at both levels of sowing date and weed control,
the response to fertilizer was by far higher at Holetta than at
Kulumsa which underlines the fact that fertilizer is more important
9 3
Oil
yiel
d (k
g/h
a)
Seed
yi
eld
(kg/
ha)
94
Hoietta Kulumsa
Fig. 19.Effects of the interactions between sowing date, fertilizer
and weed control on seed yield.
1500 -LSD Hoietta Kulumsa
6650
Earl y Late
Fig. 20*Effects of the interactions between sowing date, fertilizer and weed control on oil yield.
hiolett;
Kulums
FO = With no fertilize
F1 = Wi th ferti1i zer
WO = With no weeding
W1 = With weeding
Holettc
Ku turns
FO = Wi th no ferti1i: F1 = With fertilizer WO = With no weeding W1 = With weeding
at Holetta where the natural soil fertility level is less than that
of Kulumsa (Table P 24).
Weed control increased seed yield at both locations and
under all levels of sowing date and fertilizer. With early sowing,
weed control increased the average seed yield of unfertilized plots
by 39 and 80% over the respective values of 7 and 12.5 q/ha of
unweeded plots at Holetta and Kulumsa, respectively. It also
increased the seed yield of fertilized plots by 8 and 37% over the
respective values of 19.6 and 27 q/ha of unweeded plots at the two
locat i o n s .With late sowing, weed control increased the average seed
yield of unfertilized plots by 9% (Holetta) and 101% (Kulumsa) and
that of fertilized plots by 39 and 79% at Holetta and Kulumsa,
respectively. ^
It can be noted that at both locations, weeding was more
advantageous for unfertilized conditions of early sown brassica
and for late sown ones at Kulumsa. At Holetta, late sown plots
were more responsive to weed control under fertilized conditions.
Further, in all cases, weed control was by far more important at
Kulumsa than it was at Holetta.
Figure 20 and Appendix 22 descern that early sowing,
fertilizer and weed control at both locations increased oil yield.
With early sowing, fertilizer inreased oil yield of unweeded plots
by 180% at Holetta and by 113% at Kulumsa; and that of weeded plots
by 118 and 63%, respectively. With late sowing, it again increased
oil yield of unweeded and weeded plots by 504 and 687% (Holetta)
Under conditions of no-fertilizer, weed control increased oil
yield of early sown plots by 38% (Holetta) and 81% (Kulumsa). With
late sowing, the same factor increased oil yield by 6% at Holetta
and by 101% at Kulumsa. Similarly, under conditions of fertilizer
application, weed control increased oil yield of early sown plots
by 7 and 39% at Holetta and Kulumsa, rspectively. The corres
ponding increases for the late sown plots were 38% (Holetta) and
76% (K u l u m s a ) .The results indicate that the crops were more responsive to
fertilizer than to weed control at Holetta while at Kulumsa early
sown plots were more responsive to fertilizer and late sown ones
were to weed control. This again emphasizes that at Holetta
fertilizer plays the major role in increasing the productivity of
oil seed brassica; while at Kulumsa the importance of fertilizer
was over ridden by weed control due to the shift of the sowing date
from the optimum.
9 G
and by 84 and 61% (Kulumsa).
4.3* Correlations of Characters
Simple correlation coefficients on the linear relationships
of the different characters to each other at Holetta and Kulumsa
are presented in Table 3.
At both locations, all the characters, save number of seeds
per pod, were positively related to each other. The number of seeds per pod was invar i ably negatively related to all other
characters. Its relation to any one of the characters, however,
K uL
umsa
Ta bl e 3. S i np l e c o r r e l a t i o n c o e f f i c i e n t s betweer y i e l d c o a p o n e n t s of oil seed brassica at H o le t t a (above diagonal) and K u lu s s a (below diagonal)
9 7
Plantheight
N u m b e r of of p r ia a r yb r a n c h e s p er pl an t
N u ab e r of of s e c o n d a r ybr an c h e s p er pl an t
N u a b e r of p o d sp er p l an t
N u a b e r of s e e d spe r pod
1000-seedw eight
Seedy ield
P l an t h e ig h t 0 . 6 4 7 9 0 . 5 2 3 4 A 77J0 -0.2064 0 . 58 7 5 0 . 91 0 C *N u s b e r of p r im a r y b r a n c h e s p er plant 0 . 6 7 8 0 0.8783. 0 .5471 h j , 4233. 0.3311 0 . 53 1 3 *Nuafaer of s e co n d a ry b r a n c h e s p er plant 0 . 52 1 ? 0.9063. 0 . 31 4 8 -0,4557 0 . 36 7 E 0 .4270
N u a b e r of p o d s pe r pl an t 0 . 77 5 9 0 . 8 5 5 7 0 . 8 9 4 8 -0.2707 0. 52 2 6 0.6854 t
N u ib e r of s e B d s p er pod - 0 . 2 7 6 0 - 0 . 4 4 0 5 - 0 . 5 2 0 3 -0.3 4 3 9 -0.2327 0 .32941000-seedw e ig h t 0,28 6 4 0 . 4 2 6 5 0. 36 9 4 0.2571 -0.3156 0 .5566 tS e e d yi el d 0 . B 7 9 6 * 0.686-3* 0 . 6 6 6 9 * 0 . 8 3 1 0 * - 0.0413 0.2471
* S i g n i f i c a n t at 0.01 level
Holetta
was not significant except for the number of secondary branches
per plant observed at Kulumsa.t
Plant height was positively correlated with number of primary
branches, secondary branches and pods per plant at both locations,
but with 1000-seed weight at Holetta alone.
Number of primary branches was positively and significantly
correlated with number of secondary branches and pods per plant at
both locations. Yet, the number of pods per plant was positively
and significantly correlated with 1000-seed weight only at Holetta.
Seed yield was positively and significantly correlated with
plant height, number of primary branches and pods per plant at both
locations. In addition, it was positively correlated wij^h 1000-
seed weight at Holetta and with number of secondary branches at
K u l u m s a .These results are consistent with the results of Olsson
(1960), Singh, et a l . , (1969, 1979), Agarwal, et ad., (1973).
Zuberi, .et al. , ( 1973 ), Thurling (1974 a,c,), and Tayo, e_t a l . ,
(1975) who found positive and significant correlation between seed
yield and number of primary and secondary branches, plant height,
number of pods and seeds per plant, seeds per pod and 1000-seed
w e i g h t .On the basis of correlation coefficients, plant height
followed by numbr of pods per plant, was considered as the
component exerting a major influence upon seed yield both at
Holetta and Kulumsa. Number of primary branches was the thirc
(Kulumsa) and fourth (Holetta) important component influencing see<
S 8
yield. The number of secondary branches was the fourth important
component at Kulumsa, while 1000-seed weight was the third at
H ol e t t a ,
A casual observation of the correlation coefficients in Table
3 fails to provide much insight into the precise overall
relationships that exist in the sequential series of
characteristics and thus tend to lead towards a misleading
conclusions to be drawn. Neverthless, this complicated and hidden
relationship is brought to light through the path- coefficeint
analysis presented in Figs. 21(A) and (B).
The path analyses indicate that the effect (the direct and/or
indirect effects imposed by other characters via it) of plant
height on seed yield was positive both at Holetta and Kulumsa."/*■* a
Equally positive was that of pods per plant; but the effects 'of
primary and secondary branches and 1000-seed weight on seed yield
were negative (Table 4).
Based on path- coefficient analyses Mendham, et a l ., (1975,
1981 a) and Singh, et al., (1978, 1979) concluded that plant height
and number of pods per plant are the most important components in
contributing seed yield in oilseed brassicas which substantiate the
present findings. Similar result whereby the effect of primary
branches on seed yield turned out to be negative despite the
positive significant correlation between seed yield and primary
braches was found by Singh, e_t jlL • > (1969).
99
A (Holetta)
B (Kulumsa)
Figs ?J.(A) and (B),path-coefficient diagrams.Note The double arrowed lines indicate mutual association as measured
by correlation coefficients rij and the single arrowed lines represent direct influences as measured by path coefficient pij.
At Holetta, plant height was by far the most important
component exerting the greatest influence on seed yield; while at
Kulumsa, both plant height and number of pods .were equally important in exerting direct and indirect influences. The negative direct effects of primary and secondary branches at Kulumsa and that of 1000-seed weight at Holetta were negligible (Table 4).
The path-analyses revealed rather different picture than did the simple correlation analyses. The correlations between seed yield and number of primary branches (r = 0.5313) and 1000-seed w t . (r = 0.556) at Holetta; and that of primary (r = 0.6863) and
secondary branches ( r = 0.6669) at Kulumsa impart misleadingimpressions that they are very important components for higher seed
yield. Conversely, the path-analyses exposed the fact that they were not at least of first order importance for seed yield. Nevertheless, both simple correlations and path-analyses confirmed that plant height and number of pods per plant were more important
for seed yield than any other character.
102
V. SUMMARY AND CONCLUSION
The indigenous Brassies carinata culivar, S-67 appeared more
vigorous having more number of yield components including number of primary and secondary branches, plant height, number of pods and seeds per plant and 1000-seed weight under both conditions of Holetta and Kulumsa. The indeterminate nature of this cultivar made it possible for it to possess more number of secondary than primary branches as compared to the more compact and short B. napus cultivar Tower s e b with characteristics of more or less determinate and synchronized flowering habit, with prospects to
flower and mature early thus produce more number of seeds per pod.Early sowing, fertilizer and weed control invariably provided
more suitable growing conditions as were reflected by the increased growth and vigor of the crops. There were tremendous advantages in terms of seedling establishment, stand, plant height and number of pods and seeds per plant at both locations, and further primary and secondary branches and 1000-seed weight at Holetta.
Though at Kulumsa the number of pod bearing branches were rather reduced by early sowing the time to flowering and maturity was affected differently by sowing date at the two locations in that early sowing tended to increase days to flowering with aresultant effect of some what reduced grain-filling period atHoletta. Conversely at Kulumsa , early sowing shortened days toflowering and also days to maturity but with a net effect ofi n C IT c 3 S Gu grain-filling period.
103
At both locations, high degree of compensation of characters were observed with the methods of sowing. Row sowing increased
the number of seedling, established and number of harvestable
plants. Broadcasting, on the other hand, induced plants to develop
more number of primary and secondary branches and number of pods and seeds per plant to make up the losses due to the sparse
population density,Despite the fact that growth was faster at Kulumsa, the crops
grew more luxuriously as this was manifested through increased
number of primary and secondary branches, plant height, number ofpods and seeds per plant and seeds per pod. The seeds fromKulumsa, however, were much lighter than the seeds from Holetta.
At both locations, the seedling establishment was generally higher with early and row sowing. At Holetta, the response to" sowing method was greater than to sowing date. At Kulumsa, on theother hand, sowing date was more important. The increase due to
/row sowing was higher in B. napus (Tower sel3) than B. carinata
(S-67). The increase due to early sowing of B. carinata (S-67), was higher under broadcasting while that of early sown B . napus (Tower
sel3 ) was under row sowing.At both locations, fertilizer and early sowing increased the
number of harvestable plants. The reponse to fertilzier was higher under delayed sowing and that of early sowing was under no fertilizer conditions. This implies that the negative eflect of the lack of either factor is off-set by providing the other.
104 :
Brassica carinata (S-67) produced more number of primary
branches under early sowing dates and broadcasting, whereas B.
napus (Tower sel3 ) did under delayed sowing dates and row sowing. Both species produced higher number of primary branches with fertilizer application. The increase due to fertilizer in B.
carinata (S-67) was higher under delayed sowing while in B. napus
(Tower sel3), it was under early sowing.At both levels of fertilizer B, carinata (S-67) produced more
number of secondary branches than B . napus (Tower sel3 ) . The highest difference between the two species occured under fertilized condition where B. napus (Tower sel3) rather tended to produce less number of secondary branches. At Kulumsa, both early sowing and
fertilizer increased number of secondary branches, the increase,
however, was higher at better condition of the other. ^Under both early and late sowing dates the time to flowering
of both species was shortend by fertilizer both at Holetta and Kulumsa, At Holetta, under no-fertilizer condition, both species flowered more or less at the same time. But with fertilizer B.
napus (Tower sel3) flowered much earlier than B. carinata (S-67) due to early sowing. The reduction in days to flowering due to fertilizer was higher with delayed sowing at Holetta and with early-
sowing at Kulumsa.At Holetta, fertilizer reduced the time to maturity of B.
napus (Tower sel3 ) but not of B. carinata (S-67). The reduction was by 15 days under early sowing and by about a month-with delayed sowing. Without fertilizer, delayed sowing retarded maturity of
B. napus (Tower sel3) by 18 days . There was no effect of sowing
date or fertilizer on maturity period of B. carinata (S-67) at this location. At Kulumsa, however, fertilizer shortened the time to maturity under delayed sowing.
At both locations, early sowTing increased the height of both species. The increase was higher in B. carinata (S-67). Inaddition, fertilizer increased plant height at Holetta. The increase was higher under delayed than early sowing dates; and that of early sowing under no-fe rti1izer condition.
Both species produced more number of pods per plant with fertilizer and early sowing at Holetta and Kulumsa. The reduction at Holetta due to delayed sowing under conditions of no fertilizer was higher in B. carinata (S-67) but under fertilizer condition, it was in B. napus (Tower sel3). The response to fertilizer of both species was higher under delayed sowing.
At Kulumsa, B. carinata (S-67) produced more number of pods
under broadcasting while B. napus (Tower sel3) did under row sowing. Pod development was promoted by fertilizer under early sowing; and by early sowing under no-fertilizer and no weeding.
At both locations, fertilizer and weed control increased number of seeds per plant. At Holetta, early sowing increased
seeds per plant under no fertilizer condition. The response to fertilizer was higher under delayed sowing. B. napus (Tower sel3 ) was more responsive to fertilizer than B. carinata (S-67). AtKulumsa, B. carinata (S-67) produced more number of seeds per plant with early sowing and fertilizer while B. napus (Tower sell) did
10? 1
under delayed sowing and no fertilizer conditions. The response to fertilizer was higher under delayed sowing. With early sowing,
fertilizer was more important than weed control while under delayed sowing the reverse was found to be evident.
Both at Holetta and Kulumsa the B. carianta cultivar S-67 produced appreciably higher seed and oil yields than the B. napus cultivar. Tower sel3 . The latter, however, exhibited higher oil content. At both locations, early sowing, fertilizer and weed control resulted in higher seed and oil yields. The effects of
these management factors on oil content were either inconsistent or obscure; because oil content tended to be reduced with early sowing at Kulumsa and with weed control at Holetta, and it was
indifferent to fertilizer at Kulumsa.At Holetta, row sowing conferred an appreciable magnitude both
in seed and oil yields; while at Kulumsa, broadcasting did. The increase at Holetta was thought to be mainly via plant height, and 1000-seed weight which were also increased with row sowing. At Kulumsa, on the other hand, the increases were attached to number
of pod bearing branches and pods and seeds per plant which also increased with the same method of sowing that increased seed and oil yields at this site.
The interactions of factors indicated that both B. carinata
(S-67) and B. napus (Tower sel3) produced higher seed and oil yields in response to early sowing, fertilizer and weed control. The response of Tower sel3 to fertilizer and weed control was on the overall higher than that of S-67 which was rather more
sensitive to sowing date than the former species. The effect of
one of these factors on seed and oil yield of B. carinata (S-67) was generally more pronounced at the adverse conditions of the rest whereas the response of B. napus (Tower sel3) to any one of the factors was higher at an optimum level of the rest of the factors (Appendices 23 and 24).
This signifies the fact that the long-maturing indigenous B.
carinata (S-67) has such outstanding agronomic qualities as suppression of weeds, better performance even under no fertilizer application than the introduced cultivar Tower sel3 (B. napus).
Further,S-67 on the overall, responded little or none to method of sowing at Holetta but broadcasting suited better for this species at Kulumsa, while row sowing was favourable for Tower sel3 which again emphasizes that the latter cultivar is more demanding and can pay off only under high-imput conditions.
Early sowing increased the oil content of both species at
Holetta but reduced it at Kulumsa. The increase at Holetta was higher in B. napus (Tower sel3) and at Kulumsa, in B. carinata (S- 67) due to delayed sowing. With delayed sowing at Holetta, the oil content could be increased by fertilizer application. The effect of fertilizer with delayed sowing at Kulumsa was additive.
Under conditions of all management factors, the seed and oil
yields from kulumsa were exceedingly higher than those from Holetta. The high yielding S-67 (B carinata), at Holetta, gave comparable yield to the low yielding Tower sel3 (B. napus ) at
Kulumsa. Similarly, higher yields at Holetta obtained from early
108
sowing, fertilizer and weed control were more or less comparable
to lower yields obtained at Kulumsa due to advese effects from the
lack of these management factors. Kulumsa, therefore, appeared to
be more suitable for the production of oil seed brassica.At both locations, plant hieght was positively correlated with
number of primary branches, secondary branches and pods per plant; with 1000-seed weight at Holetta. Number of primary bracnhes was also positively correlated with number of secondary branches and pods per plant at both locations; so also was number of pods per
plant with 1000-seed weight at Holetta.Seed yield was positively correlated with plant height, number
of primary branches and pods per plant at both locations and with 1000-seed weight at Holetta but with number of secondary branches at Kulumsa. However, plant height and numnber of pods per plant-
were found to be the most important components exerting the
greatest influence on seed yield
109
VI. RECOMMENDATION
It may be pre-mature to arrive at definitive recommendations, based on the present findings of one year but the possibility of drawing some tentative conclusions to enlighten the general trends of responses to be expected is not necessarily ruled out. Thus, the following, interalia, conclusions are made based on which some leading or suggestive recommendations are forwarded.
Though due attention should be given to choice of species, sowing date, method of sowing ,fertilizer and weed control to procure substantial seed and oil yields in oil seed brassicas, the relative importance of each factor between locations varies greatly. At Holetta, fertilizer > sowing date> species > weed * control > method of sowing were found to be sequentially important ^ while at Kulumsa, there was a reciprocal change in the first and the third important factors. Therefore, the sequential importance of the factors are species> sowing date > fertilzier > weed control > method of sowing. Accordingly, priorities should be given to the factors in order to attain higher levels of seed and oil yields.
The interactive effects have shown that,' in most cases, theintroduced B. napus cultivar, Tower sel3 was more responsive to< a better level of a given management factor!s) when grown und^rconditions of optimum level of the other factor (s) *M1-- ■ '
indigenous B. carinata cultivar, S-6” L ' ' - '
only slight interver.ti " ‘ — 1
H O
-9
Therefore, under circumstances where farmers cannot afford the complete package of sowing date method of sowing, fertilizer and weed control, they can obtain substantial amount of yield by cultivating B. carinata (S-67) with reasonably less amount ofi nput.
The correlation and path-coefficient analyses revealed that plant height and number of pods are the yield components that exert the greatest influence for higher seed yield in oil seed brassicas, Thus, the production of the tall but lodging, resistant indigenous cultivar S-67 should be promoted in the less mechanized smallholding sector. But in the long-run the improvement of this species, through breeding, with the view of improving its fatty acid composition to meet the maximum erucic acid allowable i*n edible oils and with prospects of its suitability to mechanized agriculture without seriously affecting its height and pod numbers, should be pursued at a greater momentum.
111
VII. BIBLIOGRAPHY
112
*
BIBLIOGRAPHY
Acharva, S.N., J. Deieck and R.H. Downey. 1982. Selection and heritability studies on canola/rapeseed for low temperature germination. Can. J. Plant Sci. 63: 377-384.
Adams M.W. 1967. Bases of yield component compensation in cropplants with specific reference to the field bean. Phaseolus
vulgaris. Crop Sci. 7: 404-410.
Agarwal , B.L. and B. Rai. 197 3. Component analysis of the factors influencing oil production in Toria. Ind. J. Genet,
and Plant Breed. 33: 142-146.
Ali-Khan, S.T. 1976. Seed yield, seed weight, percent protein and protein yield of field peas as affecte by seeding dates Can. J. Plant Sci. 57:17-20.
Allen, E.J., D.G. Morgan and W.J. Ridgman. 1971. Physiological analysis of the growth of oil seed rape. J. Agric. sci. 77:339-341.
Annand I.J. and R.K. Downey. 1980. A study of erucic acid alleles in digenomic rape seed (Brassica napus L.). Can. J.
Plant Sci. 61:199-203.
Anonymous. 1981. Progress report of rape seed-mustard research(September 1979-August 1981) and extension proposal. G.B.
Pant University of Agriculture and Technology. Pantnagar.
India, 20 pp.
_______________. 1985. Research progress report for the period April
1984 to March 1985. G.B. Pant University of Agriculture and Technology. Pantnagar. India, 25 pp,
Appelqvist, L.A. and R. Ohlson. 1972. Rape seed: Cultivartion,composition, processing and utilization. Elsevier publishing comp. Inc. Amesterdam, The Netherlands. 391 pp.
*
Astahana, A.N., S.D. Dube, P.U. Tiwari, P.N. Gambhir and T*S.£ Rajan. 1979. Variability studies and breeding for increased
oil content in rape seed and mustard. Exp. Agric. 15: 65- 71.
Aulakh, M.S., N.S. Pasricha and N.S. Sahota. 1980. Yield nutrient concentration and quality of mustard crops as influenced by
nitrogen and sulfur fertilizers. J. Agric. Sci. 94: 654-549.
Auld, D.L., B.L. Bettis and M.J. Dial. 1984. Planting date and cultivar effect on winter rape production. Agron. J. 76: 19 7— 220 .
114
Badege Gebre, Rezene Fissehaie and A. Stround. 1986. Preliminary weed research findings on two Ethiopian highland oil seed
crops, rape seed (•Brassica napus) and niger (Guizotia
abyssinica). In Proceedigns of the Third Oilcrops Network Workshop. Abbas Ororan (e d .) Addis Ababa, Ethiopia. IDRC MR-
153e Ottawa, Canada pp.58-65.
Bettis, B.L., C.L. Peterson, D.L. Auld., D.J. Driscoll and E.D. Peterson. 1982. Feul characteristics of vegetable oil from oil seed crops in the Pacific North West. Agron. J. 335-339.
Bhan, S. 1979. Effect of soil moisture and nitrogen onmustard under Gangetic alluvium of Uttar Pradesh. Ind. J.
Agron. 24:180-186.
Bishnoi, K.C. and K. Singh. 1979. Oil yield and qualityparameters of three raya varieties as affected by sowing time
and nitrogen levels. Ind. J. Agro. 24:255-263.
Central Statistics Authority. 1987. Time series dataon area, production and yield of major crops. Addis Ababa,
Ethiopia. 66 pp.
Chauhan, Y.S. and D. Singh. 1979. Genetic archetecture of some quantitative characters in Indian mustard. Ind. J. Genet, and Plant Breed. 39: 255-262.
Chauhan, Y.S. and S.C. Bhargava. 1984. Physiological analysis of growth and variation of rape seed and mustard. J. Agric. Sci.
103:249-252.
Christensen, J.V., W.G. Legge, R.M. Depauw, A.M.F. Hennig, J.S.McKenzie, B. Siemens and J.B. Thomas. 1985. Effect of seeding
date, nitrogen and phosphate fertilizers on growth, yield and quality of rape seed in North West Alberta. Can. J. Plant
Sci. 65:275-284.
Clarke, J.M. , F.R. clarke and G.M. Simpson. 1978. Effect of method and rate of seeding on yield of Brassica napus Can. J.
Plant Sci. 58: 549-550.
_____________. 1979. Intra-plant variation in number of seeds perpod seed weight in Brassica napus: "Tower". Can. J. Plant.
Sci. 59:959-962.
Cochran, W.G. and G.M. Cox. 1957, Experimental designs, 2nd ed. John Willey and Sons. Newyork, 661 pp.
Degnehardt, D.F. and Z.P. Kondra. 1981 a. The influence of seeding date and seeding rate on seed yield and yield components of five genotypes of Brassica napus. Can. J. Plant
Sci. 61:175-183.
116
Degnehardt, D.F. and Z.P. Kondra. 1981 b. The influence of seeding date and seeding rate on seed yield and growth characters of five genotypes of Brassica napus. Can. J. Plant Sci. 61:185-190.
117
Dewey, D.R. and K.H. L U . 1959. A correlation and path-coefficientanalysis of crested wheat grass seed production. Agron. J. 51:515-518.
Downey, R.K. 1971. Agricultural and genetic potentials of
4 -____________ . 1983. The origin and discription of the brassica
oil seed crops. In High and Low Erucic Acid Rapeseed Oils. Academic press, Canada, pp. 1-18.
Dybing. C.D. 1964. Influences of nitrogen level on flax growth and oil production in varied environments. Crop Sci. 4: 491- 494.
Fan, Z. and W. Tai. 1985. A cytogenetic study of breeding foroil quality in rape. Can. J. Plant Sci. 43:271-275.
Food and Agriculture Organization of the United Nations (FAO).1986. Production Yearbook. 40: P. 115.
Forbes. J.C. 1985 a. Weed-crop competition studies in swedes, I. The effect of time of weed removal on crop yield. Ann. Appl. Biol. 106 : 505-5111
118
Forbes. J.C. 1985 b. Weed-crop competition studies in swedes. II.
" The effects of weed competion on crop growth parameters. Ann. Appl. Biol. 106:413-423.
Fulkerson, R.S. 1971. Agronomic studies with fodder rape(Brassica
napus var. Biennis). Can. J. Plant Sci. 41:305-308.
Gowers, S. 1974. The production of FI hybrid swedes (Brassica
napus spp rapifera) by the utilization of self-incompatibility. Euphytica. 23:205-208.
Gross, a.T. and B.R. Stefansson. 1966. Effect of planting date on protein, oil, and fatty acid content of rape seed and turnip rape. Can. J. Plant Sic. 46-389-395.
Helps, M.B. 1971. Methods of sowing, seed rate and nitrogen level for oil seed rape. Exp. Husb. 20:69-72
Hemingway, J. 1976. Mustards Brassica spp. and Sinapis alba
(cruciferae ) > In Evolution fo Crop Plants. N.W. Simmonds (ed.) Longman. London 339 pp.
Appeadii 2. Ctd.
132
Soiree of nrUtioi Degree of Buber of Buber of luber of 1000-seed Seed Oil Oilfreedot pods seeds seeds leigkt . yield coiteit yield
per plot per piut per pod
Species (S) 1 73024 « 159338 « 456.93 tt 4.08 « 5275564 » 348.8 « 489139 ttDate of soniig (D) 1 4047 u 4501838 « 405.27 « 3.70 tt 16449414 tt 61.5 tt 2852504 ttletkod of sowiag (Hi 1 452 tt 281691 « 1.39 IS 0.06 IS 464889 tt 0.2 SS 82486 «Fertiliser if) 1 23288 tt 6282759 tt 89.68 « 2.42 tt 40962563 tt 54.0 w 6331207 ttSeed coitrol (1) 1 1486 t 737186 tt 27.14 tt 0.25 IS 1331304 tt 10.9 tt 205089 tt
SB 1 26 IS 961937 « 6.32 IS 1.63 « 2521 IS 32.5 tt 5442 ISSF 1 490 IS 362001 « 8.38 IS 1,00 t 1069453 tt 1.3 IS 93192 ttBF 1 133 IS 5920 IS 8.00 IS 0.60 IS 155683 * 55.8 tt 24877 IS
m i 219 IS 1218 IS 4.00 IS 1.441 140 IS 0.01 IS 191 ISSBF 1 127! t 297115 « 0.13 IS 1.41 « 270 IS 0.01 IS 3 ISSIF 1 591 IS 3944 IS 4.S3 IS 0.01 IS 107300 * 0.S0 8S 18826 ISS&K 1 308 IS . 3370 IS 4.08 IS 0.48 IS 10768 * 0.40 IS 1819 ISOF! 1 1 IS 17041 tt ll.SC IS 0.84 IS 435478 tt 0.60 IS 733S1 «
Error 20 270 13335 3.113 0.230 25794 0.551 6038
tt Eigkly significant* SigRificaatIS lot sigaificast
Appeidii 3. l e u squires of tke ckincters it Ciluisa
13
Soarce of uriatioi«
Degree Seedlitg of freedoi deuitj
Stud Suber of priiarj braackes per plut
Saiber of secoid&rj bruckes per plut
Dipto
fioieriii
Dipto
latiritj
Plutkeigkt
Species (i) 1 134 IS 128 tt 901.00 tt 3346.64 tt 1052 tt 3838 tt 24809 «Date of soiiigtD! 1 23240 tt 1800 tt 2. 82 t 16,75 tt 229 « 118 tt 24476 ttletkod of Bomgilj 1 8T91 « 276 tt 7. 80 tt 53.17 tt 193 « 80 IS 72 ISFertiliier (F) 1 12 IS 722 tt 51.26 tt 58.46 tt 1808 tt 1030 tt 71T0 ttfed control (1) 1 3 IS 238 tt 42.09 tt 446.63 tt 17 IS 375 « 1785 *
SD 1 867 « 25 SS 28.12 tt 3.72 IS 174 « 7 IS 4827 ttSF 1 8 IS 1 IS 12.25 tt 235.72 tt 2 IS 3 BS 358 IS
' OF 1 6 IS 98 t 0.07 SS 164.03 tt 100 « 635 tt 1 SS
SDK 1 308 * 12 IS 17.55 tt 0.06 IS 1 IS 9 IS 903 ISSBF 1 65 i s 52 IS 8.00 tt 3.28 IS 126 tt 1 IS 124 ISSftF 1 28 IS 16 IS 1.85 tt 0.83 IS 1 BS 24 IS 66 ISSOI 1 1 IS 18 IS 0.45 IS 5.74 IS 1 IS , 2 IS i ISs u 1 23 IS 7 IS 1.58 IS 4.69 IS 20 BS 9 IS 25 ISDFi 1 29 IS 6 IS 0.88 IS 5.74 IS 35 IS 126 t 306 IS
Error SO 64 12 0.696 2.656 13 29 365
t t a igk ly sigiificut * Sigiificut SS lot sigiificut
Species (S) 1 153389 « 10160508 « 224.38 « 1.34 U 68929943 tt 282.0 tt 8115745 ttDate of Bowiag (D) 1 9028 tt 2821678 tt 4.50 SS 0.17 IS 51713179 tt 106.0 « 6294558 ttKetAori of so¥ii4 (I) 1 67712 « 383885 « 0.88 IS 0.24 IS 1497531 tt 3.1 IS 182138 «Fertiliser (F) 1 <6642 « 12041901 « 68.23 « 0.01 IS 33666141 tt 6.5 SS 503(2(6 ttfeed control (S) 1 S7712 « 8022015 « 6.22 IS 0.17 IS 23907748 « 2.8 IS 346T0220 tt
SD 1 8036 « 1201250 tt 2.99 SS 0.01 SS 22640551 « 54.1 tt 233( isSF 1 215 es 959113 « 3.27 IS 0.22 IS 1712869 tt 0.1 IS 154211 ttOF 1 1501 « 3161418 tt 3.29 IS 0.81 IS 5696578 tt 632941 t t
m 1 4574 « 7468 IS 4.70 IS 0.01 IS 1219 IS 6.5 * 2253 ISSDF 1 23 ES 387816 tt 3.60 IS 0.01 IS 2149960 « 0.02 IS 2454 ISSIF 1 210 IS 2073 RS 2.14 IS 0.16 IS 347465 tt 3.5 IS 559 ISSD* 1 228 ES 1208 iS 15.08 IS 0.10 SS 900985 tt 0.1 gs 113 ISSli 1 698 IS 4106 IS 13.99 IS 0.30 IS 92503 tt 9.8 11 1343 ISDFi 1 3894 « 537581 « 0.01 IS 0.03 IS 121956 tt 0.2 IS 23537 t
Appeidii 4. Hut keight (C») is affected by tke interactions of species ud *o»ii| date (SO) and sowing date and fertiliser (DF( it Boeltti aid by SD it Uluu
D1 = Early sowing FO= Hitkout fertiliserD2 = Late sowiig FI = iitk fertiliser
Appendix 5. Rniber of secoodary branches per plait as affected by tke interaction of species and sowing date (SD) and date of sowing and fertilizer (BF) at hlusa
4ppeadii 8. luber of priury bruckei per plut u affected by tke iateractiois ofspecies, date of sotiaj aid tetkod of Eotiig (EDI) tuber of pods per plut by date of soiia*, fertiliser ud teed coitrol (BFI) at [alust
Snssici c ir iu tt_______ Snssict m u ______fittest fertiliser f it i fertilifer
01 = Early Botiag 10 = litk ao feedifigD2 = Late sotiig II = litk leedieg
Appendix 9. Saiber of pods per plant as affected by the iateractioas of species, date of sowing ud letkod of sowing (SDK) at [alnisa and species, date of soting and fertiliser (SDF) at Eoletta
D1 = Early soving FO = litkoat fertiliserB2 = Late sowing FI = litk fertiliser
1 3 8
Appeidii 10. 1000-seed leigkt (|) as affected by tke iiteractiois of species, soiiif date aad letkod of soiiig (SDK) ifid specie*, sosing date ud fertiliser (SDP) at Soletta
Brassica carimU Brassica tipis fcm ic i cariaala Brassica napis
D1 = lirly sowing FO = Sithoat fertiliserD2 = Late sowing FI = With fertilizer
Appendix 13. Days to eahrity as affected by the interactions of species, date of sowing and fertilizer (SDF) at loletta and by date of sowing, fertilizer and wed control m \ at Kulmsa,
Snssica c ir iuU Brassica aapas lith no weeding lith weeding
01 : Early sowing FO = lithoat fertilizerD2 = Late sowing FI = lith fertilizer
Henery. J.L. and K.B. McDonald. 1978. The effects of soil and fertilizer nitrogen and moisture stress on yield, oil and
protein content of rape. Can. J. Plant Sci. 58: 303-310.
Hiruy Belayneh. 1983. Highland oil crops production and research in Ethiopia. In Oil Crop Proceedings Cairo, Egypt, pp 62- 71. IDRC MR-93e. Ottawa, Canada.
________________ . 1985. Review of mustard/rapessed agronomy resarchin Ethiopia. Paper presented at Agronomy Committee Meeting, Cambridge, U.K., GCIRC Bulletin No. 2: 52-57*
__________________ , K.W. Riley, Nigatu Tadesse and Getinet Alemaw.* 41983. The response of three oilseed Brassica spp to
different planting dates and seed rates in the highlands of Ethiopia, Eth. J. Agri. Sci. 5: 22-31.
_______________ and Nigussie Alemayehu. 1986. Progress inrape seed/mustard reserch in Ethiopia. In Oil Crops: Niger and Rapeseed/Mustard. Abbas Omran ( e d . ) . Proceedings of the Third Oil Crops Network Workshop Addis Ababa, Ethiopia, p(p. 12-17. IDRC-MR 153e, Ottawa, Canada.
______________ and ________________ 1987. Comporative performance ofEthiopian mustard (Brassica carinata A. Braun) and Argentine rapeseed (Brassica napus L.) under improved and traditional
119
*
farming practices. Paper presented at the SeventhInterationai Rape Seed Congress, Poznan, Poland, 11-14 May.1987.
Hiruy Belayneh. 1987. Present status and future strategies of oil
seed brassica resarch in Ethiopia. In Oil Crops the Brassica Sub-net Work Proceeding Uppsala, Sweden, pp. 15-26. IDRC-MR168e, Ottawa, Canada. \
Holmes, M.R.J. 1980. Nutrition of the oil seed rape crop. Applied science publishers Ltd. London, 158 pp.
Ibrahim, A.E.S. 1983. Associations, path-analysis and multiple coefficients of determination of some yield components in ground nut (Arachis hypogea L.). Oleagineux 38: 323-327.
Jain, V.K. and G. Tripathi. 1979. Production technology of mustard. In Farmer and Parlimament. 14: 5-6. Delhi, India.
Joarder, O. 1983. Oil content, yield and morphological responseof rape (Brassica campestris L.) to irrigation and fertilizer treatment. J. Agric. Sci. 100: 253-255.
Kaul, A.K. and M.L. Das. 1986. Oil seeds in Bangladesh. BRAC Printers, Dhaka, Bangladesh. 323 pp.
120
Kondra, Z.P. 1975. Effects of row spacing and seeding rate on rape seed. Can. J. Plant Sci. 55: 339-341.
«f
___________ . 1976. Effect of planting date on rape seed.Can. J. Plant Sci. 57: 607-609.
Kondra , Z . P . , D . C . Carobe 11 and J.R, King. 1983. Te mpe ratureeffects on germination of rape seed (Brassica napus L. and B.
campestris L.). Can. J. Plant Sci. 63: 1063-1065.
Krogman, K.K. and E.H. Hobbs, 1975. yield and morphological
response of rape (Brassica campestris L. C v . Span) to irrigation and fertilizer treatments. Can. J. Plant Sci. 55:903- 909. **
Kumar, A. and R.P. Singh, 1989. Agronomic investigations on rape
seed and mustard at Pantnagar. Paper presented at the Brassica sub-net work meeting held at G.B. Plant University of Agriculture and Technology, Pantnagar, India, from Jan. 4- 6, 1989.
Madson, E. 1976. Nuclear Magnetic Resonance Spectrometry a method of determination of oil content in rape seed JAOCS 53: 467- 469.
synthesis, cytology, fertility and general considerations. Euphytica 22: 301-309.
_________________• 1976 a. Turnip and relatives. Brassica campestris
(Cruciferae). In Evolution of Crop Plants. N.W. Simmonds (ed.). Longman, London and New York. 339 pp.
McNaughton, I.H. 1973 b. Swedes and rapes , Brassica napus
(Cruciferae). In Evolution of Crop Plants. N.W. Simmonds (ed.). Longman, London and New York. 339 pp.
Mendham, N.J. and P.K. Scott. 1975. The limiting effect of planet size at inflorescence initiation on subsquent growth and yield of oilseed rape (Brassica napus). J. Agric. Sci. 84: 487-502.
___________ > ___________ and P.A. Shipway. 1981 a. The effects ofdelayed sowing and weather on growth, development and yield
, of winter oil-seed rape (Brassica napus). J. Agric. Sci. 96: 389-416.
----- ------------ j________________ and__________________ b. The effectsof seed size, autumn nitrogen and plant population density on the response to delayed sowing in winter oil-seed rape (Brassica napus). J. Agric. Sci. 96: 417-428.
122
Mizushima, U. amd S. Tsunoda. 1967. A plant exploration inBrassica and allied genera. Tohoku J. Agric. Res. 17: 249-
274.
Moss, D.N. and R.B. Musgrave. 1971. Photosynthesis and cropproduction. Advan, in Agron. 23: 317-336,
Nuttal, W.F., H. Ukrainetz. J. W. B. Stewart and D.T. Spurr. 1987. The effect of nitrogen, sulfur and boron on yield and quality of rape seed (Brassica napus L.) and Brassica compestris L.) Can J. Soil Sci. 67: 545-559.
(j
Nwankiti, D. 1970. Cytogenetic and breeding studies withBrassica. I. Cytogenetic experiments with Brassica napo
campestris. Hereditas 66: 109-126.
Olsson, G. 1960. Some relations between number of seeds per pod, seed size and oil content and the effects of selection for these characters in Brassica and Sinapis. Hereditas 46: 29- 70.
Patil, C.B. 1979. Yield and yield attributes of niger as affected
by different agrotechniques. Ind. J. Agric. Sci. 49: 633-636.Paul, N. K. , 0.1 . Joarder and A.M. Eunus. 1976. Genotypic and
Phenotypic variability and correlation studies in Brassica
Juncea L. Pflanzenzitchtz 77: 145-154.
Prakash, S. 1973. Haploidy in Brassica nigra Koch. Euphytica 22:
613-614 .
___________ . 1974. Probable basis of diplodization of Brassica
Juncea coss. Can. J. Genet. Cytol. 16: 232-234.
Prakash, S. and R.N. Raut. 1983. Artificial synthesis of Brassica napus and its prospects as an oil seed crop in India. Ind. J.
Genet, and Plant Breed. 43: 282-290.
Pretty, K.M. and K.B. McDonald. 1976. Reduce crop stress with right planting time. Better crops with plant food. 60: 4-10.
Ractz, G.J., M.D. Webber, R.J. Sober and R.A. Hedlin. 1965.Phosphorus and nitrogen utilization by rape, flax and wheat.
Agron. J. 57: 335-337.i
Seegeler. C.J.P. 1983. Oil Plants in Ethiopia* their taxonomy and agricultural significance. Center for Agricultural Publishing and Documentation, Pudoc. Wageningen. Neatherlands. 368 pp.
Sheppard, S.C. and T.E. Bates. 1980. Yield and Chemical composition of rape in response to nitrogen, phosphorus and
potassium. Can. J. Soil Sci. 60: 153-162.
124
Singh, B., J.N. Sachan, R. Bhajan, S.P. Singh and D. Singh. 1984. Current status of Indian rape seed and mustard breeding research at G.B. Pant University of Agriculture and Technology. In Oil Crops News Letter. A.O. Omran (ed.). No.
1. p p . 62-67.
Singh, Dharampol, D. Singh, S.P. Singh, A.B. Singh and S.N. Mishi 1969. Relationship among some important agronomic characters in Indian mustard (B. juncea (L.l. Czern and Coss. ). Ind.
J. Agric. Sci. 39: 362-365.
Singh, R.M., B.S. Talukdar, Y.S. Chauhan and A.K. Singh. 1978.*
Path-coefficient studies in Indian mustard (Brassica Juncea>* H(L.). Czern and coss.). In Proceedings of 5th International Rape seed Conference. Vol. 2. Malmo, Sweden, June 12-16, 1978. p p . 71-7 5.
Singh, S.P., A.N. Srivastava and R.P. Katiyar. 1979. Path analysis in Indian Colza. Ind. J. Genet, and Plant Breed. 39: 150-153*
Staniforth, D.W. 1958. Soybean-foxtail competition under varying soil moisture conditions. Agron. J. 50: 13-15,
Stringham. G.R. and D.I. Me Gregor. 1980. Inheritance and fatty acid composition of yellow embryo mutant in turnip rape, (Brassic campestris L.). Can. J. Plant Sci. 60: 97-102.
125
Synder, F.W. and G.C. Carlson. 1978. Photosynthate partitioning in sugarbeet. Crop Sci. 18: 647-661.
Tatchel, G.M. 1983. Compensation in spring-sown oil seed rape (Brassica napus L.) plants in response to injury to their flower buds and pods. J. Agric. Sci. 101: 565-573.
Taye Bekele. 1986. Response of oilseed brassica to fertilziers. In Oil Crops Niger and Rapeseed/Mustard. Abbas Omran (ed.).
Proceedings of the Third Oil Crops Network Workshop Addis Ababa, Ethiopia. 1986. IDRC MR-153e pp 40-45.
Tayo, T.O. and D.G. Morgan. 1975. Quantitative analysis of the- *.
growth, development and distribution of flowers and pods in oil seed rape (Brassica napus L.). J. Agric. Sci. 85: 103-1 1 0 .
____________ and _____________1979. Factors influencing flowerand pod development in oil seed rape (Brassi,ca napus L. ).J. Agric. Sci. 92: 363-373*
Thurling, N. 1974 a. Morphophysiological determinants of yield inrape seed (Brassica campestris) and B. napus). I. Growth and morphological characters. Aust. J. Agric. Res. 25: 697-710.
Thurling, N. 1974 b. Morphophysiological determinants of yield in
rape seed (Brassica campestris and B. napus). II. yield components. Aust. J. Agric. Res. 25: 711-721.
____________________ C. An evaluation of an index method of selectionfor high yield in turning rape (Brassica campestris L. spp. Oleifera metzg.). Euphytica 23:321-331.
U, N. 1935. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of
fertilization. Jap. J. Bot. 9: 389-452.
Uppstrom, B. 1977. Breeding method for improvement of brassicaoil crops. In the second FAO/SIDA seminar of field food crops in Africa and the Near East, Lahare, Pakistan 15 pp,
Vaughan, J.G. 1970. The stucture and utilization of oil seeds. Chapman and hall Ltd. 11 New Fetter Lane, London. England.2 79 p p .
Weiss, E.A. 1983. Oil crops. Tropical agriculture series. Longman Inc. New Youk. U.S.A. 660 p p ,
Zimdahl, R.L. 1980. . Weed-crop competition. A review.International plant protection center. Oregon. U.S.A. 195 pp.
127
Zuberi, M.I. and S.U. Ahmed. 1973. Genetic study of yield andsome of its components in Brassica campestris L. var. "Toria". Crop Sci. 13: 13-15.
128
VIII. APPENDIX
129
Appendix 1. Meteorological data for Holetta and Kulumsa.
o<uu3uojU<ueu6a>H
-5 -
Jun Jul Aug ISep "T"Oct "T~Nov
200
- 150
- 100
- 50
- 0 200
- 150
- 100
u 50
- 0
u-te•HS
DecKey:Ten days mean of maximum and minimum temperatures ( X?.. X = ten yearsmean at Holetta and five yearsr mean at Kulumsa,-O— —o-=» 1988) and rain-' fall (A-~— & * ten years mean at Holetta and five years mean at Kulumsa D--- Q = 1988 ).
ippeidii 2. Xeu tqmee of ike ckiricteri at BoletU
131
Soiree of uriitioi Degree of freedoi
Seedlitgdeuity
Staid Inker of of priiary kruckei per plut
Sober of secoidary brucket per plut
Daystofloieriig
*Dayito
■atirity
Plutkeigkt
Species (t) 1 2253 tt 877 tt 157.75 « 1985.29 t t 561 tt 11495 tt 11459 t tDate of loviif(D) 1 1760 « 1122 tt 7.95 « 29.36 tt 36 « 1041 tt 19135 «letkod of Boiiig(lt) 1 164 « 1631 tt 1.73 t 22.36 tt 15 t 6 IS 815 tFertiliser (F) 1 387 « 4198 tt 15.33 tt 27.66 tt 3528 » 4129 tt 38330 «feed coitrol if) 1 12072 tt 727 tt 4.24 tt 0.10 IS 3 IS 1 IS 2355 IS
SD 1 38 IS 3 IS 26.01 tt 1.50 IS 181 « 417 tt 492 tSF 1 16 IS 6 IS 0.83 IS 2.50 IS 450 tt 3839 tt 6 ISOF 1 30 fiS 897 t t 0.71 SS 0.87 IS 91 « 508 tt 897 tt
SDH 1 51* 1 IS 0.83 SS 2.62 IS 1 IS 2 IS 142 ISSDF 1 25 IS 2 SS 2.73 * 0.20 IS 18 « 617 t t 9 ISSHF 1 6 IS 169 IS 0.64 ES 9.52 IS IIS 1 IS 461 ISan 1 34 IS 35 IS 0.16 IS 2.62 IS 13 IS 16 IS 71 ISDR 1 2? IS 142 BS 0.08 IS 0.60 K 2 IS 2 IS 56 18
FO = litkoit fertiliser !0 = S itk 10 feedingFI - lith fertiliser SI = fitk seeding
/
ippeidii if. Oil coiteit (X) ti affected by tke iiterictiois of species ud date of soviif (SD) ud d&te of soiiif aid fertiliser (DPI tt Icletta ud lilim
141
Holetta Iilusa
Brassica B eusici Sms i a Brassicacariiata tapis c&riuU iapu(S-6T) (Tower sell) Reu FO FI l e u ( H T ) (tower sell) Keu FO FI l e u
Di = Early soiiig FO = Without fertiliserD2 = Late soiiag FI = Kith fertiliser
f
Appendii 18. Seed yield (kf/ka) u affected by tke iateractioiB of species, date of soiiigaid fertiliser (SBF) aid species, date of soiitj aid weed coitrol (SSI) at Eiluu
142
I r u i iu cuiuU(S~if) Btmict npts(Toier ieij) Brassica car iiata(S-fT) Brassica up is (Toier sell)
I p p e a d i i 2D. Seed y i e l d (kg/ha) is a f f e c t e d by th e i n t e r a c t i o n s r. s p e c i e s . l e t h o d of soriag and feed coatrol (SID it loletta jad !C<2luasa
loletta . . . «Inhisa
tnssici a r i n t i ( H V Inssici upisUore: seh) Brassica carinata{S-67) Brassica napns {Tofer seh)