All-Trans Retinoic Acid Induces Chromatin Remodeling at the Promoter of the Mouse Liver, Bone, and Kidney Alkaline Phosphatase Gene in C3H10T 1/2 Cells Yang Wan • Songhai Yang • Fenyong Sun • Jiayi Wang • Qiongyu Chen • An Hong Received: 9 January 2011 / Accepted: 31 May 2011 / Published online: 22 January 2012 Ó Springer Science+Business Media, LLC 2012 Abstract The alkaline phosphatase (ALP) gene is an important marker of osteo- blast differentiation and bone formation. Although the molecular mechanisms of increased ALP expression in response to all-trans retinoic acid (ATRA) have been reported, the role of ATRA in chromatin structure changes remains unknown. Our results show that the expression of mouse liver, bone, and kidney ALP (mL/B/K- ALP) induced by ATRA in C3H10T 1/2 cells was related to the retinoic acid nuclear receptors, RARa and RARb, which are not involved in the MAPK pathway. DNase I hypersensitivity analysis revealed an inducible hypersensitive site in the mL/B/K- ALP promoter at *520 bp upstream of the transcription start site. Chromatin immunoprecipitation experiments showed a cascade of transcription cofactor recruitment events during ATRA-induced upregulation of mL/B/K-ALP. Together, our results provide a link between ATRA-induced mL/B/K-ALP gene transcription and chromatin remodeling. Keywords Osteoblast differentiation Bone formation Gene regulation Histone modifications MAPK pathway Yang Wan and Songhai Yang have contributed equally to this study. Y. Wan F. Sun Q. Chen A. Hong (&) Institute of Genetic Engineering, Key Lab for Genetic Medicine of Guangdong Province, National Engineering Research Center of Genetic Medicine, Jinan University, Guangzhou 510632, Guangdong, China e-mail: [email protected]S. Yang Shaoguan Tielu Hospital, Shaoguan 512023, Guangdong, China J. Wang Department of Laboratory Medicine, Ruijin Hospital, Shanghai Jiaotong University School of Medicine, Shanghai 200025, China 123 Biochem Genet (2012) 50:495–507 DOI 10.1007/s10528-011-9494-9
Transcript
All-Trans Retinoic Acid Induces ChromatinRemodeling at the Promoter of the Mouse Liver,Bone, and Kidney Alkaline Phosphatase Genein C3H10T 1/2 Cells
Yang Wan • Songhai Yang • Fenyong Sun •
Jiayi Wang • Qiongyu Chen • An Hong
Received: 9 January 2011 / Accepted: 31 May 2011 / Published online: 22 January 2012
� Springer Science+Business Media, LLC 2012
Abstract The alkaline phosphatase (ALP) gene is an important marker of osteo-
blast differentiation and bone formation. Although the molecular mechanisms of
increased ALP expression in response to all-trans retinoic acid (ATRA) have been
reported, the role of ATRA in chromatin structure changes remains unknown. Our
results show that the expression of mouse liver, bone, and kidney ALP (mL/B/K-
ALP) induced by ATRA in C3H10T 1/2 cells was related to the retinoic acid nuclear
receptors, RARa and RARb, which are not involved in the MAPK pathway. DNase
I hypersensitivity analysis revealed an inducible hypersensitive site in the mL/B/K-
ALP promoter at *520 bp upstream of the transcription start site. Chromatin
immunoprecipitation experiments showed a cascade of transcription cofactor
recruitment events during ATRA-induced upregulation of mL/B/K-ALP. Together,
our results provide a link between ATRA-induced mL/B/K-ALP gene transcription
(sc-11417), and HDAC4 (sc-11418) were purchased from Santa Cruz Biotechnol-
ogy (Santa Cruz, CA, USA). Antibodies to diacetyl-H3 (07-593) and tetra-acetyl-H4
(06-866) were obtained from Upstate (Charlottesville, VA, USA). The RAR
agonists Ch55, RARa (AM580), RARb2 (AC55649), and RARc (CD1530) were
496 Biochem Genet (2012) 50:495–507
123
purchased from Tocris Bioscience (Bristol, UK). Cycloheximide (CHX) and ATRA
were purchased from Sigma-Aldrich (St. Louis, MO, USA). The MAPK signaling
pathway inhibitors SB600125 (JNK inhibitor), SB203580 (p38 inhibitor), and
PD98059 (ERK inhibitor) were purchased from Calbiochem (San Diego, CA, USA).
C3H10T 1/2 cell lines were purchased from American Type Culture Collection
(Manassas, VA, USA) and cultured in Dulbecco’s modified Eagle’s medium
containing 10% fetal bovine serum, supplemented with L-glutamine (2 mM),
penicillin (100 U/ml), and streptomycin (100 mg/ml).
RNA Isolation and Quantitative Reverse Transcription PCR Analysis
Total RNA was isolated from cell cultures using Trizol reagent (Invitrogen,
Carlsbad, CA, USA). Total RNA (1 lg) was used for reverse transcription with the
MMLV reverse transcriptase (Toyobo Co., Osaka, Japan). PCR reactions were
performed in triplicate with primers for mL/B/K-ALP (F, 50 CTGACTGACCCTTC
GC TCTC 30, and R, 50 TGCTTGGCCTTACCCTCAT 30) and 18S RNA (F, 50
CCTGGATACCGCAGCTAGGA 30, and R, 50 GCGGCGCAATACGAATGCCCC
30) in separate wells, and monitored using Rotor-Gene 6000 (Corbett Life Science,
Sydney, Australia). Then quantitative reverse transcription PCR (qRT-PCR) was
performed to detect the mRNA expression level of the mL/B/K-ALP gene at
different time points as indicated. Results are presented as the amount of mL/B/K-
ALP mRNA present relative to that in untreated cells, and normalized to the level of
18S RNA.
DNase I Hypersensitivity Analysis
Cells were collected by trypsin digestion and lysed with buffers. Genomic DNA was
purified by phenol/chloroform and subsequent ethanol precipitation. The nuclei and
genomic DNA were digested with increasing concentrations of DNase I (0, 1, 3,
5 U/ml) (Takara Co., Tokyo, Japan) for 10 min at 37�C. Approximately 15 lg
DNase I-treated DNA was digested with Eco47III (Takara) and subjected to
electrophoresis on a 0.8% agarose gel using 19 TBE buffer. The DNA was
transferred to a nylon membrane. Southern blot assay probes were generated by
PCR using these primers: forward, 50 CACCCACCTCTACCTCATTG 30, and
reverse, 50 GGCTGTCCTGGAACTCACTC 30 (?2093 to ?2958). The genomic
DNA of the C3H10T 1/2 cells was used as template; PCR products were purified
and labeled using 32P.
Chromatin Immunoprecipitation Assays
Assays were performed according to the manufacturer’s instructions. A total of
2 9 106 cells was fixed with 1% formaldehyde, washed with cold phosphate-
buffered saline, and lysed in buffer. Nuclei were sonicated to shear DNA
(100–1,000 bp), and the lysates were pelleted and precleared. After immunopre-
cipitation with 3 lg antibody overnight, the protein–DNA-antibody complexes were
incubated with protein A beads (Invitrogen) and then washed with buffers. The
Biochem Genet (2012) 50:495–507 497
123
protein–DNA complexes were eluted in 1% SDS ? 0.1 M NaHCO3, and
crosslinking was reversed at 65�C. After protein digestion at 42�C, DNA was
purified using a DNA purification kit (Tiangen Biotech, Beijing, China). An equal
volume of ChIP DNA was subjected to real-time PCR using the primers F-50
GTAGGCAGAAAGACGCACAG 30 and R-50 TTCAGGGTGCAGCTGGCTTG 30,targeting the promoter region of -529 to -377, relative to the transcription start site
(TSS). Results are presented as the amount of antibody immunoprecipitation
relative to the untreated cells and normalized to the level of no antibody.
Results
ATRA Upregulation of mL/B/K-ALP Gene Expression
The upregulating effects of ATRA on the mL/B/K-ALP gene have been confirmed in
different cell lines by other studies (Heath et al. 1992). To detect ATRA-induced
mL/B/K-ALP gene expression in C3H10T 1/2 cells, we treated cells with 1 lM
ATRA for 0, 0.5, 1, 3, 6, 12, 24, 48, and 72 h, then RNA was extracted and qRT-
PCR was performed. The results show that the mL/B/K-ALP mRNA expression
level was upregulated in a time-dependent manner, which is similar to a previous
study by Orimo and Shimada (2005). Specifically, the changes in mL/B/K-ALPmRNA expression level were more obvious during 0–12 h than at other time points
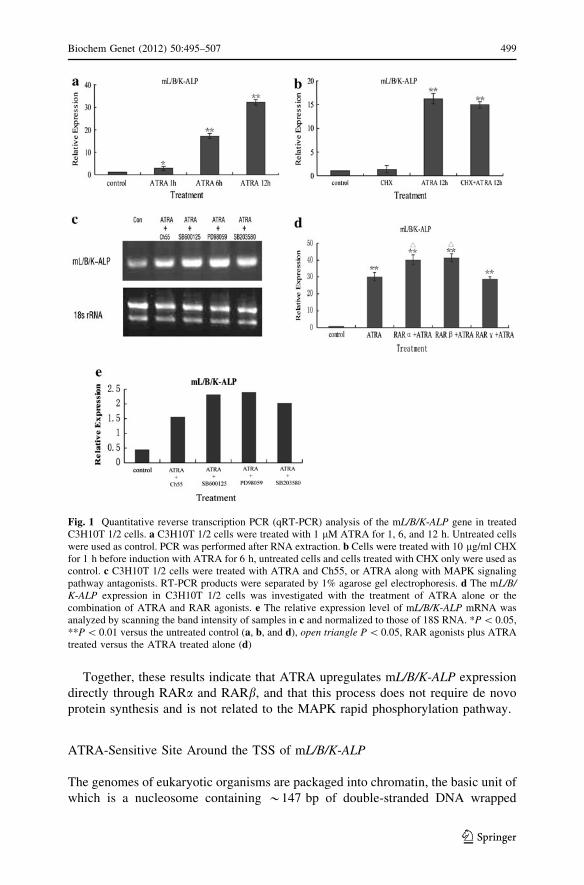
(Fig. 1). At 1 h after ATRA treatment, mL/B/K-ALP gene expression was
approximately double that in untreated cells. The expression level continued to
increase, to 16 times that in untreated cells at 6 h and to 32 times the untreated level
at 12 h. When the cells were treated with 10 lg/ml CHX for 1 h before the
induction (Fig. 1), CHX did not affect the upregulating effects of ATRA. This result
suggests that ATRA-mediated mL/B/K-ALP gene expression does not require de
novo protein synthesis, and that the promoter is likely to be the direct target of
ATRA.
The major signaling pathway of retinoic acids is through the classic genomic
pathway of nuclear receptors (RARs). Moreover, ATRA does not bind the retinoic
acid X receptor (RXR) (Chambon 1996; Leid et al. 1992). Thus, we treated C3H10T
1/2 cells with 1 lM ATRA and 50 lM Ch55, a general agonist of RAR, to elucidate
the mechanism of ATRA-mediated mL/B/K-ALP upregulation. Transcription of mL/B/K-ALP was upregulated significantly in the presence of the RAR agonist (Fig. 1).
Moreover, treatment with ATRA together with inhibitors of the MAPK pathway
10 lM) did not affect mL/B/K-ALP upregulation (Fig. 1). These results suggest
that mL/B/K-ALP upregulation by ATRA is mediated directly by RARs and does
not involve the MAPK rapid phosphorylation pathway.
Next, C3H10T 1/2 cells were treated with 1 lM ATRA and 50 lM of the
agonists to RARa (AM580), RARb (AC55649), or RARc (CD1530) for 12 h, then
qRT-PCR was performed. RARa and RARb agonists upregulated the inducing
effects of ATRA on mL/B/K-ALP expression. The RARc agonist, however, did not
increase the ATRA-induced mL/B/K-ALP mRNA expression (Fig. 1).
498 Biochem Genet (2012) 50:495–507
123
Together, these results indicate that ATRA upregulates mL/B/K-ALP expression
directly through RARa and RARb, and that this process does not require de novo
protein synthesis and is not related to the MAPK rapid phosphorylation pathway.
ATRA-Sensitive Site Around the TSS of mL/B/K-ALP
The genomes of eukaryotic organisms are packaged into chromatin, the basic unit of
which is a nucleosome containing *147 bp of double-stranded DNA wrapped
Fig. 1 Quantitative reverse transcription PCR (qRT-PCR) analysis of the mL/B/K-ALP gene in treatedC3H10T 1/2 cells. a C3H10T 1/2 cells were treated with 1 lM ATRA for 1, 6, and 12 h. Untreated cellswere used as control. PCR was performed after RNA extraction. b Cells were treated with 10 lg/ml CHXfor 1 h before induction with ATRA for 6 h, untreated cells and cells treated with CHX only were used ascontrol. c C3H10T 1/2 cells were treated with ATRA and Ch55, or ATRA along with MAPK signalingpathway antagonists. RT-PCR products were separated by 1% agarose gel electrophoresis. d The mL/B/K-ALP expression in C3H10T 1/2 cells was investigated with the treatment of ATRA alone or thecombination of ATRA and RAR agonists. e The relative expression level of mL/B/K-ALP mRNA wasanalyzed by scanning the band intensity of samples in c and normalized to those of 18S RNA. *P \ 0.05,**P \ 0.01 versus the untreated control (a, b, and d), open triangle P \ 0.05, RAR agonists plus ATRAtreated versus the ATRA treated alone (d)
Biochem Genet (2012) 50:495–507 499
123
around an octamer of histone proteins (Khorasanizadeh 2004). It is generally
accepted that local chromatin structure affects gene transcription. We used DNase I
hypersensitivity assays to reveal regions of open chromatin around the mL/B/K-ALPgene in C3H10T 1/2 cells. The genomic fragments between -18,763 and
?4,178 bp (relative to the TSS) were analyzed. The probe was located in the
vicinity of the downstream restriction site (?2,093 and ?2,958 bp). Two bands
were obtained in ATRA-treated C3H10T 1/2 cells on Southern blots (Fig. 2). The
band of 22.9 kb represents the original Eco47III–Eco47III restriction fragment of
mL/B/K-ALP, the signal of which weakened with the increase in DNase I enzyme.
The lower band showed an increase in signal with the increase in concentration of
DNase I. It represents a single DNase hypersensitive site that was estimated to be
*4.7 kb upstream of the Eco47III site at ?4,178 bp, adjacent to the TSS of the mL/B/K-ALP promoter located at -520 bp (Fig. 2). In control experiments, increasing
DNase I concentrations did not produce any specific bands at 4.7 kb, suggesting that
Fig. 2 DNase I hypersensitive site analysis of the mL/B/K-ALP promoter region. a C3H10T 1/2 cellswere treated with or without 1 lM ATRA for 12 h. The arrowhead shows bands (about 4.7 kb) produceddue to DNase I cleavage. As a control, purified genomic DNA from C3H10T 1/2 cells was digested invitro with increasing concentrations of DNase I. b Schematic representation of the probe used to map theDNase hypersensitive sites within the mL/B/K-ALP promoter by indirect end-labeling techniques
500 Biochem Genet (2012) 50:495–507
123
the 520 bp hypersensitive site is a result of chromatin remodeling rather than DNase
I recognition.
Together, these results indicate that a new DNase I hypersensitive site was
produced in C3H10T 1/2 cells after ATRA induction, and the appearance of this site
may be related to the mL/B/K-ALP gene transcription after induction. ATRA
induction may have caused a change in the conformation of the chromatin near the
TSS, thereby exposing the previously embedded DNase I hypersensitive site.
Changes in Transcriptional Activity of mL/B/K-ALP Promoter Related
to Chromatin Remodeling and Histone Modifications
Studies have shown that transcription activation or inhibition requires not only the
binding of transcription factors at specific sites in the promoter but also the
recruitment of other protein complexes for chromosome remodeling and modifi-
cations. Chromatin can be modified in ways that affect DNA function through the
action of two main types of multiprotein complexes. The first is the adjustment of
specific histone–DNA contacts by ATP-dependent nucleosome remodeling enzymes
(Lusser and Kadonaga 2003). The second is the post-translational covalent
modification of histones through the addition or removal of phosphoryl, methyl,
or acetyl groups, among other covalent modifications (Lo et al. 2004). To
understand ATRA-induced chromatin remodeling and histone modifications, we
examined the interactions between the mL/B/K-ALP promoter and 11 factors using
chromatin immunoprecipitation in noninduced cells and in cells induced with
ATRA for 1, 6, or 12 h. For this, we chose primers around the ATRA-sensitive site
(Fig. 3).
We first examined the binding of RNA polymerase II, the core component of the
gene transcription complex, onto the mL/B/K-ALP promoter. A large amount of
RNA polymerase II was recruited to the transcription region of the promoter at 1 h
after ATRA treatment (Fig. 3). Thereafter, the degree of enrichment gradually
declined with the increase of induction time.
In the transcription complex, histone acetylation and promoter transcription
activity are closely associated. Typically, histone acetylation is associated with
transcriptional activation, and histone deacetylation is associated with a decrease in
transcriptional activity (Sterner and Berger 2000; Fischle et al. 2003). The levels of
histone acetylation are regulated by histone acetyltransferases (HATs) and histone
deacetylases (HDACs). Gene transcription can be affected not only by modifications
in the composition of the nucleosome octamer but also by the methylation,
acetylation, and phosphorylation of specific amino acids in H3 and H4 histones.
Therefore, we examined the changes in H3 and H4 acetylation levels during
induction. The H3 acetylation level in induced cells was 10 times that of noninduced
cells at 1 h, 12 times at 6 h, and 7.8 times at 12 h after ATRA induction (Fig. 3). H4
acetylation was 3 times that of control cells at 1 h after ATRA induction, rose to a
maximum of 12 times the control at 6 h, and returned to the 1-h level at 12 h after
induction.
We then examined the recruitment and enrichment of the acetyltransferases, p300
and GCN5, in the transcriptional complex (Fig. 3). Similar to the changes observed
Biochem Genet (2012) 50:495–507 501
123
Fig. 3 Quantitative RT-PCR results using various antibodies with ChIP. a ChIP production location onthe mL/B/K-ALP promoter. b–i C3H10T 1/2 cells were treated with ATRA for 0, 1, 6, or 12 h, followedby ChIP using different antibodies: RNA polymerase II (b), histones H3 (c) and H4 (d), acetyltransferasesp300 (e) and GCN5 (f), histone deacetylases (g), and the ATP-dependent chromosome remodeling factorsBrg1 (h) and Brm (i). All results are means of three independent experiments ± standard deviation.*P \ 0.05, **P \ 0.01 versus the untreated control
502 Biochem Genet (2012) 50:495–507
123
in H3 and H4 acetylation levels, a substantial amount of p300 was recruited to the
vicinity of the TSS at 6 h after ATRA induction. At 12 h, the p300 level decreased
to the level observed at 1 h after induction. GCN5 levels were twice that in
noninduced cells at 6 h after induction. GCN5 levels increased dramatically, by 131
times, at 12 h post-induction. These results demonstrated that histone acetylation
and recruitment of acetylases increased near the TSS of the mL/B/K-ALP promoter
after ATRA treatment.
We examined the recruitment of HDAC1, 2, 3, and 4 into the chromosome during
induction, with effects the opposite of those of p300 and GCN5 (Fig. 3). HDAC1, 2,
and 3 recruitment was similar, with their levels increasing significantly at 1 h after
induction, followed by a decrease after 6 h. HDAC4 was not recruited until 6 h after
induction, and its level subsequently decreased.
Using the DNase I hypersensitive assay, we showed that a new open site was
produced near the TSS after ATRA induction; thus, chromatin remodeling exposed
a previously embedded DNase I hypersensitive site. We examined the expression of
the ATP-dependent chromosome remodeling factors, Brg1 and Brm of the SWI2/
SNF2 family, during the induction (Fig. 3). The greatest degree of enrichment of
both Brg1 and Brm was observed at 6 h after induction, with Brg1 and Brm levels at
3 and 9 times that in control cells. At 12 h after induction, Brg1 and Brm levels had
decreased to 1.5 and 4.4 times the control levels.
Discussion
Retinoic acid regulates multiple biological processes, including cell proliferation
and differentiation, by virtue of its ability to modulate the rate of transcription of
numerous target genes. Retinoic acid induces the expression of the L/B/K-ALP gene
in rat fibroblast cell lines (Gianni et al. 1993). Even though the molecular
mechanism of this activation has been extensively studied (Orimo and Shimada
2005), the relationship between chromatin structure and transcriptional activity
remains unclear.
The transcriptional activities of retinoic acid are mediated by two members of the
nuclear hormone receptor superfamily: the RXR, which is activated by the 9-cis
isomer of retinoic acid, and the RAR, which responds to both 9-cis and ATRA.
These retinoid receptors bind to specific response elements in the promoter regions
(retinoic acid response elements, RAREs) of target genes and function as ligand-
inducible transcription factors (Chambon 1996; Leid et al. 1992; Mangelsdorf et al.
1994). RXR can bind to DNA and activate transcription as a homodimer. In
contrast, tight binding to DNA and transcriptional activation by RAR usually occurs
through heterodimerization with RXR (Durand et al. 1992; Hallenbeck et al. 1992;
Leid et al. 1992; Yu et al. 1991). Upon ligand binding, RAR–RXR heterodimers
recruit multicomponent coactivator complexes that in turn remodel chromatin and
bridge to the general transcription machinery to modulate gene expression (Glass
and Rosenfeld 2000; Xu et al. 1999). Both of these receptors are composed of three
subtypes: a, b, and c (Durand et al. 1992). Because RARs are the initial response
receptor of the activation by ATRA, we treated cells with RARa, b, and c agonist.
Biochem Genet (2012) 50:495–507 503
123
The results revealed that ATRA regulated mL/B/K-ALP expression through RARaand RARb (Fig. 1).
Retinoic acid can also activate gene transcription through nongenetic mecha-
nisms, by activating signal pathways that lead to the phosphorylation of
transcription factors (Canon et al. 2004). Because ATRA rapidly upregulates mL/B/K-ALP gene expression, we speculated that this process involved the MAPK rapid
phosphorylation pathway. When ATRA-induced C3H10T 1/2 cells were treated
with JNK, ERK, or P38 antagonists, upregulation of the mL/B/K-ALP gene
expression was unaffected. Moreover, treatment of cells with a general RAR agonist
only resulted in the induction of mL/B/K-ALP gene expression.
These results show that the ATRA-mediated upregulation of mL/B/K-ALP
involves RAR and is not related to the MAPK rapid phosphorylation pathway.
Using the luciferase activity assay and EMSA to examine fragments of different
lengths within 4.5 kb of the hL/B/K-ALP promoter in SaOS-2 osteosarcoma cell
lines, Orimo and Shimada (2005) identified a RARE located between -1,012 and
-999 bp upstream of the TSS of hL/B/K-ALP. In our study, we conducted a large-
scale screening of 22.9 kb (-18,763 to ?4,178 bp) fragments around the TSS using
DNase I hypersensitivity analysis and found a new DNase I-sensitive site at
*520 bp upstream of the TSS of the mL/B/K-ALP promoter in ATRA-induced
C3H10T 1/2 cells. Chromosomal structure is an obstacle for the binding of
transcription factors and the formation of the preinitiation complex (Adelman and
Lis 2002). Thus, the activation of gene transcription may be accompanied by
chromosome remodeling, which results in the appearance of new DNase I-sensitive
sites due to the increase in the accessibility of the local chromatin (Hebbes et al.
1994). Therefore, we inferred that ATRA mediates the upregulation of mL/B/K-ALPexpression by regulating its promoter accessibility, and that this site contains a
RARE. We found that in the mL/B/K-ALP gene, the potential RARE is located at
approximately -520 bp, but in the human gene the RARE is located at
approximately -630 bp (Orimo and Shimada 2005). Accordingly, the putative
RARE is consistent with the previous report of the murine RARE by Escalante et al.
(1996). As the DR5 sequence is a/g g g/t tcannnnn a/g g g/t tca (Leblanc and
Stunnenberg 1995), we detected a sequence, ggttca cagacgcgca gggtca (-324 to
-302), similar to the DR5 sequence around our RARE. Although the free bases are 10
instead of 5, our results can provide a better understanding of the mL/B/K-ALP gene.
Gene expression is under strict control. A series of transcription factors,
transcriptional cofactors, and chromosome-modifying enzymes act within the
genome to activate or inhibit gene expression (Reid et al. 2009). Histone-modifying
enzymes, as well as ATP-dependent remodeling enzymes, play an important role in
the regulation of the open state of chromatin. The regulation of histone or
nonhistone acetylation levels by HATS and HDACs has an important impact on the
open state of the gene. HATS and HDACs do not have specific binding sites and
need to be guided by specific transcription factors to act at the specific position of
the target gene. Histone acetylation increases its affinity for the SWI/SNF complex
(Han et al. 2006). The combined actions of histone acetylation and remodeling
enzymes allow the transcription spacer of the promoter to open further and allow the
binding of the transcription factor, thereby providing a binding site for other
504 Biochem Genet (2012) 50:495–507
123
transcriptional coactivators. In addition, this modification may increase the stability
of transcription complexes (Zhao and Benveniste 2008).
In this study, we used chromatin immunoprecipitation to examine the enrichment
of various factors in the specific promoter region of the mL/B/K-ALP gene during
ATRA induction. For purposes of discussion, we propose a model for the induction
of mL/B/K-ALP gene expression in C3H10T 1/2 cells that summarizes some of the
findings described here (Fig. 4). In the untreated condition, mL/B/K-ALP is
expressed at a low basal level. The chromosome is closed, and the TSS is wrapped
within the nucleosomes and unable to bind to the transcription complex. When the
cells are treated with ATRA, ATRA interacts with RARs, and the ATRA–RAR
complex binds to the RARE located in the promoter of the mL/B/K-ALP gene. H3
and H4 become acetylated under the action of acetylases, such as p300, GCN5,
HDAC 1, 2, 3, and 4; also, Brg1 and Brm are recruited to the vicinity of the TSS,
and the regulatory region of the gene is opened, allowing the recruitment of RNA
polymerase II. The transcription complex begins to form, and mL/B/K-ALPtranscription is initiated.
It is noteworthy that the levels of RNA polymerase II are contrary to the levels of
mL/B/K-ALP gene expression (Figs. 1, 3). In general, HATs are involved in
transcriptional activation, and HDACs play an important role in transcription
inhibition. We found, however, that the levels of HATs and HDACs during ATRA
induction were similar (Fig. 3). These results suggest that there may be other
transcription factors involved in the inducing progress of ATRA on mL/B/K-ALP.
In summary, our study demonstrates that in C3H10T 1/2 cells, ATRA
upregulates mL/B/K-ALP expression through RARa and RARb, unrelated to the
MAPK pathway. ATRA induces mL/B/K-ALP expression by promoting changes in
the structure of chromatin through the sequential and combined action of HATs,
HDACs, Brg1, and Brm.
Acknowledgments This study was supported by the National Natural Science Foundation of China
(Nos. 30600328 and 30740030).
Fig. 4 Schematic representation of the possible mechanism underlying ATRA treatment at the mL/B/K-ALP promoter in C3H10T 1/2 cells
Biochem Genet (2012) 50:495–507 505
123
References
Adelman K, Lis JT (2002) How does Pol II overcome the nucleosome barrier? Mol Cell 9:451–452
Budhu AS, Noy N (2002) Direct channeling of retinoic acid between cellular retinoic acid-binding protein
ii and retinoic acid receptor sensitizes mammary carcinoma cells to retinoic acid-induced growth
arrest. Mol Cell Biol 22:2632–2641
Canon E, Cosgaya JM, Scsucova S, Aranda A (2004) Rapid effects of retinoic acid on CREB and ERK
phosphorylation in neuronal cells. Mol Biol Cell 15:5583–5592
Chambon P (1996) A decade of molecular biology of retinoic acid receptors. FASEB J 10:940–954
Durand B, Saunders M, Leroy P, Leid M, Chambon P (1992) All-trans and 9-cis retinoic acid induction of
CRABPII transcription is mediated by RAR–RXR heterodimers bound to DR1 and DR2 repeated
motifs. Cell 71:73–85
Escalante AD, Recillas TF, Hernandez GD, Castro OS, Terao M, Garattini E, Covarrubias L (1996)
Retinoic acid and methylation cis-regulatory elements control the mouse tissue non-specific alkaline
phosphatase gene expression. Mech Dev 57:21–32
Fischle W, Wang Y, Allis CD (2003) Binary switches and modification cassettes in histone biology and
beyond. Nature 425:475–479
Garen A, Levinthal C (1960) A fine-structure genetic and chemical study of the enzyme alkaline
phosphatase of E. coli, I: purification and characterization of alkaline phosphatase. Biochim Biophys
Acta 38:470–483
Gianni M, Terao M, Sozzani S, Garattini E (1993) Retinoic acid and cyclic AMP synergistically induce
the expression of liver/bone/kidney-type alkaline phosphatase gene in L929 fibroblastic cells.
Biochem J 296:67–77
Glass CK, Rosenfeld MG (2000) The coregulator exchange in transcriptional functions of nuclear
receptors. Genes Dev 14:121–141
Hallenbeck PL, Marks MS, Lippoldt RE, Ozato K, Nikodem VM (1992) Heterodimerization of thyroid
hormone (TH) receptor with H-2RIIBP (RXR beta) enhances DNA binding and TH-dependent
transcriptional activation. Proc Natl Acad Sci USA 89:5572–5576
Han S, Lu J, Zhang Y, Cheng C, Han L, Wang X, Li L, Liu C, Huang B (2006) Recruitment of histone