Altered vitamin D status in liver tissue and blood plasma from Greenland sledge dogs (Canis familiaris) dietary exposed to organohalogen contaminated minke whale (Balaenoptera acuterostrata) blubber Christian Sonne a,n , Maja Kirkegaard a , Jette Jakobsen b , Bjørn Munro Jenssen c , Robert J. Letcher d , Rune Dietz a a Aarhus University, Faculty of Science and Technology, Department of Bioscience, Arctic Research Centre, Roskilde, Denmark b Technical University of Denmark, National Food Institute, Søborg, Denmark c Department of Biology, Norwegian University of Science and Technology, Trondheim, Norway d Wildlife and Landscape Science Directorate, Science and Technology Branch, Environment Canada, National Wildlife Research Centre, Carleton University, Ottawa, Canada article info Article history: Received 7 October 2013 Received in revised form 12 November 2013 Accepted 18 November 2013 Available online 13 April 2014 Keywords: 25-Hydroxyvitamin D 3 Sledge dogs Organohalogen contaminants PCBs Polychlorinated biphenyls Vitamin D 3 abstract This study compared vitamin D 3 (vitD 3 ) and 25-OH vitamin D 3 (25OHD 3 ) status in Greenland sledge dogs (Canis familiaris) given either minke whale (Balaenoptera acuterostrata) blubber high in organohalogen contaminants (OHCs) or clean porcine (Suis scrofa) fat for up to 636 days. A group of six exposed and six control sister bitches (maternal generation) and their three exposed and four control pups, respectively, were daily fed 112g whale blubber (193 mg ∑PCB/day) or porcine fat (0.17 mg ∑PCB/day). Mean level of ∑PCB in adipose tissue of exposed bitches and their pups was 3106 and 2670 ng/g lw, respectively, which was significantly higher than the mean concentration of 53 ng/g lw for all controls (p o0.001). The vitamin analyses showed that 25OHD 3 in liver of maternal exposed bitches were significantly lower than in controls (p ¼0.004) while vitD 3 was significantly highest in liver of exposed pups (p o0.003). Regarding blood plasma concentrations, exposed F generation pups had significantly higher concentra- tions of 25OHD 3 than controls (p ¼0.009). Correlation analyses showed that blood 25OHD 3 decreased significantly with increased adipose tissue concentrations of ∑PCB in exposed dogs (R 2 ¼0.64, p ¼0.005) and a similar trend was found for liver 25OHD 3 (R 2 ¼0.32, p ¼0.08). The results indicate that the homeostasis and metabolism of vitamin D compounds may respond differently to the dietary composi- tion of fatty acids and OHC exposure. It is unknown if the lower level of 25OHD 3 in the liver of exposed dogs would have any negative effects on immunity and reproduction and more focus should be conducted on this compound in Arctic wildlife. & 2014 Elsevier Inc. All rights reserved. 1. Introduction Vitamin D (vitD) is categorized as a fat soluble seco-steroid hormone rather than a vitamin and it has several important functions in the organism, including regulation of mineral and calcium metabolism hereunder bone mineralization. Other func- tions of the hormone include regulation of blood pressure, immune functions, cell proliferation and differentiation, gonadal functions, apoptosis and cancer protection (Kinuta et al., 2000; Li et al., 2004; Lou et al., 2004; Uitterlinden et al., 2004; Tuohimaa et al., 2005; Norman, 2008). VitD 3 exists in two main forms: vitamin D 2 (VitD 2 , ergocalciferol) and vitamin D 3 (VitD 3 , cholecal- ciferol) (Feldman et al., 2005; Norman, 2008). VitD 3 is absorbed in the intestine or produced in skin from 7-dehydrocholesterol during UV-light exposure. It is metabolized first in the liver to 25-OH vitamin D 3 (25OHD 3 ) and then further to 1,25-dihydrox- yvitamin D 3 (1,25OHD 3 ) mainly in the kidney but also in many other tissues. The latter is the active hormonal form of vitD 3 in tissues and binds to the nuclear vitamin D receptor (VDR) (Feldman et al., 2005). The liver hydroxylation of vitD 3 to 25OHD 3 is, as opposed to hydroxylation of 25OHD 3 to 1,25OHD 3 , unregulated, and 25OHD 3 is accepted as a good indicator for vitD 3 status (Hollis, 2005). 25OHD 3 and 1,25OHD 3 circulate in the blood stream as complexes with Vitamin D-binding protein (DBP), albumin and lipoproteins (Kalueff et al., 2004). There is, however, Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/ecoenv Ecotoxicology and Environmental Safety 0147-6513/$ - see front matter & 2014 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ecoenv.2013.11.017 n Correspondence to: Aarhus University, Faculty of Science and Technology, Department of Bioscience, Frederiksborgvej 399, PO Box 358, DK-4000 Roskilde, Denmark. Fax: þ45 87 15 50 15. E-mail address: [email protected](C. Sonne). Ecotoxicology and Environmental Safety 104 (2014) 403–408
Transcript
Altered vitamin D status in liver tissue and blood plasmafrom Greenland sledge dogs (Canis familiaris) dietary exposedto organohalogen contaminated minke whale (Balaenopteraacuterostrata) blubber
Christian Sonne a,n, Maja Kirkegaard a, Jette Jakobsen b, Bjørn Munro Jenssen c,Robert J. Letcher d, Rune Dietz a
a Aarhus University, Faculty of Science and Technology, Department of Bioscience, Arctic Research Centre, Roskilde, Denmarkb Technical University of Denmark, National Food Institute, Søborg, Denmarkc Department of Biology, Norwegian University of Science and Technology, Trondheim, Norwayd Wildlife and Landscape Science Directorate, Science and Technology Branch, Environment Canada, National Wildlife Research Centre, Carleton University,Ottawa, Canada
a r t i c l e i n f o
Article history:Received 7 October 2013Received in revised form12 November 2013Accepted 18 November 2013Available online 13 April 2014
This study compared vitamin D3 (vitD3) and 25-OH vitamin D3 (25OHD3) status in Greenland sledge dogs(Canis familiaris) given either minke whale (Balaenoptera acuterostrata) blubber high in organohalogencontaminants (OHCs) or clean porcine (Suis scrofa) fat for up to 636 days. A group of six exposed and sixcontrol sister bitches (maternal generation) and their three exposed and four control pups, respectively,were daily fed 112 g whale blubber (193 mg ∑PCB/day) or porcine fat (0.17 mg ∑PCB/day). Mean level of∑PCB in adipose tissue of exposed bitches and their pups was 3106 and 2670 ng/g lw, respectively, whichwas significantly higher than the mean concentration of 53 ng/g lw for all controls (po0.001). Thevitamin analyses showed that 25OHD3 in liver of maternal exposed bitches were significantly lower thanin controls (p¼0.004) while vitD3 was significantly highest in liver of exposed pups (po0.003).Regarding blood plasma concentrations, exposed F generation pups had significantly higher concentra-tions of 25OHD3 than controls (p¼0.009). Correlation analyses showed that blood 25OHD3 decreasedsignificantly with increased adipose tissue concentrations of ∑PCB in exposed dogs (R2¼0.64, p¼0.005)and a similar trend was found for liver 25OHD3 (R2¼0.32, p¼0.08). The results indicate that thehomeostasis and metabolism of vitamin D compounds may respond differently to the dietary composi-tion of fatty acids and OHC exposure. It is unknown if the lower level of 25OHD3 in the liver of exposeddogs would have any negative effects on immunity and reproduction and more focus should beconducted on this compound in Arctic wildlife.
& 2014 Elsevier Inc. All rights reserved.
1. Introduction
Vitamin D (vitD) is categorized as a fat soluble seco-steroidhormone rather than a vitamin and it has several importantfunctions in the organism, including regulation of mineral andcalcium metabolism hereunder bone mineralization. Other func-tions of the hormone include regulation of blood pressure,immune functions, cell proliferation and differentiation, gonadalfunctions, apoptosis and cancer protection (Kinuta et al., 2000;Li et al., 2004; Lou et al., 2004; Uitterlinden et al., 2004; Tuohimaa
et al., 2005; Norman, 2008). VitD3 exists in two main forms:vitamin D2 (VitD2, ergocalciferol) and vitamin D3 (VitD3, cholecal-ciferol) (Feldman et al., 2005; Norman, 2008). VitD3 is absorbed inthe intestine or produced in skin from 7-dehydrocholesterolduring UV-light exposure. It is metabolized first in the liver to25-OH vitamin D3 (25OHD3) and then further to 1,25-dihydrox-yvitamin D3 (1,25OHD3) mainly in the kidney but also in manyother tissues. The latter is the active hormonal form of vitD3 intissues and binds to the nuclear vitamin D receptor (VDR)(Feldman et al., 2005). The liver hydroxylation of vitD3 to25OHD3 is, as opposed to hydroxylation of 25OHD3 to 1,25OHD3,unregulated, and 25OHD3 is accepted as a good indicator for vitD3
status (Hollis, 2005). 25OHD3 and 1,25OHD3 circulate in the bloodstream as complexes with Vitamin D-binding protein (DBP),albumin and lipoproteins (Kalueff et al., 2004). There is, however,
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/ecoenv
Ecotoxicology and Environmental Safety
0147-6513/$ - see front matter & 2014 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.ecoenv.2013.11.017
n Correspondence to: Aarhus University, Faculty of Science and Technology,Department of Bioscience, Frederiksborgvej 399, PO Box 358, DK-4000 Roskilde,Denmark. Fax: þ45 87 15 50 15.
species difference and for example domestic cats (Felis catus) anddogs (Canis familiaris) are not able to synthesize vitD3 adequatelyin the skin following sunlight exposure why vitD3 is an essentialdietary vitamin for these two species (How et al., 1994; Kennyet al., 2004). Additionally, studies have also linked alterations inthe vitD-VDR system to behavioral disorders in mice such asanxiety, motor behavior, maze performance and exploration,suggesting that vitD3 and VDR are important factors in the centralnervous system and imbalance may significantly affect emotionalbehavior (Kalueff et al., 2004).
Only few studies have investigated the effects of organohalogencontaminant (OHC) on vitD3 status in mammals. Lilienthal (2000)studied PCB exposed rat (Rattus rattus) dams (5–40 mg ∑PCB/kg inthe diet), and reported decreased levels of both 25OHD3 and1,25OHD3 using OHC reconstituted congener patterns similar tothose found in human breast milk, even for the lowest dose.In addition to this, Routti et al. (2008) studied wild gray (Halio-cerus grypus) and ringed (Pusa hispida) seals from the Baltic regionrevealing a relationship between circulating 1,25OHD3, calcium,phosphate and thyroid hormone levels and hepatic PCB and DDTconcentrations.
In the present study, Greenland sledge dog bitches were fedminke whale (Balaenoptera acuterostrata) blubber containing highOHC concentrations or clean porcine (Suis scrofa) fat low in OHCfor 656 days. This composition and daily oral intake mimic theeffects from chronic OHC exposure of Arctic mammals. We reporton OHC exposure and subsequent tissue residue levels and thepotential effects on concentrations of vitD3 (liver) and 25OHD3
(liver and plasma).
2. Materials and methods
2.1. Experimental design
The maternal sledge dog generation (P) was composed of six exposed and sixcontrol sister bitches from Aasiaat in Disco Bay, West Greenland (N 681420 ,W 521510 , Fig. 1) obtained at two months of age. One bitch from each sister-pairwas assigned randomly to either the exposed or control group in order to minimizeage and genetic differences between the two groups (Table 1). The bitches weremated with the same unexposed male which resulted in the pup generation (F) thatwas composed of three exposed and four control pups all siblings within the group(Table 2). After weaning at 6–8 weeks of age, they were fed the same diet as theirmother. The exposed dogs were fed whale blubber without epidermis from a WestGreenland minke whale, rich in OHCs, polyunsaturated lipids and vitaminsobtained by native subsistence hunting while control dogs were fed porcine fat(lard) low in OHCs and polyunsaturated fatty acids (Sonne et al., 2006, 2007a,2007b, 2008a, 2008b, 2008c). All dogs were fed 50–200 g/day of either blubber orporcine fat which was equivalent to twenty percent of the daily total energy intake.All dogs were also fed 50–200 g/day of standardized Royal Canine Energy 4300/4800 dry dog pellets to cover basic nutrients, vitamins and microelements (www.RoyalCanin.com). The energy intake was balanced in order to achieve comparableweights among siblings in the two groups. Concentrations of vitamins A, E and Dwere higher in the whale blubber given the exposed dogs (Table 3). Description anddetailed discussion of concentrations of pollutants, lipids and nutrients in the diethas been published previously (Sonne et al., 2008b, 2008c). The dogs weresubjected to various treatments during the study as described elsewhere (Sonneet al., 2006, 2007a, 2007b, 2008a, 2008b, 2008c) and were euthanatized uponexperiment completion at adulthood for the maternal generation at an age of1.570.1 years (range 1.5–2 years) while offspring were all one year old. The animalexperiment was performed on a license granted by the Home Rule GovernmentChief Veterinarian in Greenland and conducted in accordance with national andinstitutional guidelines for the protection of animal welfare.
2.2. Blood and tissue sampling
Blood was sampled from the cava vein using a Vacutainer™ blood collectionsystem with an eighteen gauge needle and sterile Lithium-Heparin Greiner 9 mLVacuette and centrifuged at 3600 rpm at 1450g for 10 min. Subsequently 4 mLblood plasma from each dog was sampled in 10 mL sterile Nunc 348224 cryo tubesand frozen to �18 1C immediately. A total of 50 exposed and 50 control bloodplasma samples were taken from the six sister pairs of the P generation between
three and 21 months of age. From the seven (three exposed and four controlsiblings) F generation pups, nineteen blood plasma samples were taken betweenthree and twelve months of age. Liver and adipose tissue was sampled from allbitches and pups immediately after euthanasia and transported and stored at�18 1C until chemical analysis in the respective laboratories as described below.
2.3. Analysis of tissue vitamin D3 and 25OHD3
The vitamin D analyses were performed at the Technical University ofDenmark, National Food Institute, Denmark. The analytical method and theequipment used to determine vitD3 and 25OHD3 in whale blubber, porcine lardand dog livers has been previously described in detail (Jakobsen et al., 2004, 2007).Briefly, the internal standards of vitD2 and 25 OHD2 were added to the samples andsaponified with ethanolic potassium hydroxide, and the unsaponifiable matter wasextracted with diethylether–petroleum ether (1:1). This solution was purified on asilica solid-phase extraction column. Subsequently, clean-up was performed bypreparative HPLC-procedure using a silica column combined with an aminocolumn. Finally, the separation, detection and quantitation were performed on ananalytical HPLC-system with a reversed phase column and diode array detector(DAD) and UV-detector. The limit of detection (LOD) and limit of quantification(LOQ) were for both D3 and 25OHD3 0.02 mg/100 g and 0.1 mg/100 g, respectively.The LOD, LOQ and analytical method including the equipment used to determine25OHD3 in plasma has been previously described by Jakobsen et al. (2004, 2007,2009). A minor modification was introduced by using 25OHD2 (Sigma-Aldrich,Buchs, Schwitzerland) as internal standard. Briefly, plasma proteins were precipi-tated with ethanol and the supernatant was cleaned by a MFC18 solid-phaseextraction. Finally the separation, detection and quantitation of the 25OHD3
compounds were performed on an analytical HPLC-system using a cyano columnand DAD- and a UV-detector. Quantification limits for 25OHD3 was 6 nmol/l.All samples from each sledge dog was analyzed in the same run, while the inter-assay variation was 12.4 percent (n¼62), and the recovery efficiency was 94.176.7percent (n¼34). The analyses were performed in a laboratory accredited according toISO 17025.
2.4. Analyses of dietary vitamins, nutrients and fatty acids
In the dietary fat supplements, analyses for vitamin D (vitD3 and 25OHD3),vitamin A (all-trans retinol) and vitamin E (α-tocopherol) were performed atTechnical University of Denmark, National Food Institute, Søborg, Denmark. Themethod used for vitamin D is described above, and for vitamin A and E the methodsapplied were according to the EN12823-1 (2000) and EN12822 (2000),
Fig. 1. Location of Aasiaat (Egedesminde; 681420N. 521500W) in West Greenlandwhere the sledge dog study was conducted during year 2004–2005. Modified fromwww.google.com.
C. Sonne et al. / Ecotoxicology and Environmental Safety 104 (2014) 403–408404
respectively. Total vitamin C (ascorbic acid and dehydroascorbic acid) wereperformed at Eurofins Lab (Kolding, Denmark) and metals and fatty acids at AarhusUniversity, Department of Bioscience as described previously by Sonne et al.(2008c). Vitamin content in Royal Canin dog food pellets were described accordingto formula content by Royal Canin (www.royalcanin.com). All analyses for metaland vitamins were performed accredited according to ISO 17025 or ISO 2000.
2.5. Analysis of organohalogens and elements
Organohalogen contaminant (organochlorines and brominated flame retardants)analysis in blood plasma and fat samples were conducted at the National WildlifeResearch Center, Carleton University, Ottawa, Canada, according to methodsdescribed in detail elsewhere (Verreault et al., 2009a, 2009b; Dietz et al., 2004,2007, 2013a; Gebbink et al., 2008; Muir et al., 2006). Furthermore, OHC data forblood and fat has been published elsewhere as well (Verreault et al., 2008, 2009a,2009b; Dietz et al., 2013b). The following contaminants were analyzed: PCBs; 61congeners. OCs; 1,2,4,5-tetrachlorobenzene (TeClBz), pentachlorobenzene (PnClBz),hexachlorobenzene (HCB), α-hexachlorocyclohexane (HCH), β-HCH, γ-HCH, octa-chlorostyrene (OCS), the chlordanes and metabolites heptachlor epoxide, oxychlor-dane, trans-chlordane, cis-chlordane, trans-nonachlor, cis-nonachlor, p,p0-DDE, p,p0-DDD, p,p0-DDT, dieldrin, photomirex, mirex and tris(4-chlorophenyl)methane(TCPM). Also in fat, 35 PBDE congeners were monitored for (BDE-17, -25, -28, -54,-47, -49, -66, -77, -100, -119, -99, -116, -85, -155, -154, -153, -139, -138, -140, -180,-183, -184, -191, -187, -190, -171, -202, -201, -197, -203, -196, -207, -208, -206 and
-209), as well as several other BFRs including 2,2,0 ,4,40 ,5-pentabromobiphenyl (BB-101), 2,20 ,4,40 ,5,50-hexaBB (BB-153), pentabromotoluene (PBT), hexabromobenzene(HBB), total-(α)-hexabromocyclododecane (HBCDD), bis(2,4,6-tribromophenoxy)ethane (BTBPE) and pentabromoethylbenzene (PBEB). For plasma samples, thirteenPBDE congener (BDE-17, -258, -49, -47, -66, -100, -99, -85, -154, -153, -138, -183 and-209) were monitored for, but the only other BFRs monitored for were BB-101 andtotal-(α)-HBCDD. The reason lesser number of PBDE congeners and BFRs weremonitored in plasma is because based on fat analysis all other PBDEs and BFRs listedfor fat were below detection (see below).
As originally reported elsewhere (Verreault et al., 2008, 2009a, 2009b), theanalyte-specific method limit of quantification (MLOQ) was conservatively set (tominimize analytical response variation that occurs close to the detection limit) as asignal being ten times the standard deviation of the noise, The method limit ofdetection (MLOD) was based on the signal being three times the standard deviationof the noise. For the fat samples, where the mean lipid content was 8478 percent,the MLOD for all OHCs under study was generally 0.005 ng/g lipid weight (lw) andthe MLOQ was generally 0.01 ng/g lw. For the plasma samples, where the meanlipid content was 0.7270.14 percent, the MLOD for all OHCs under study wasgenerally 0.5 ng/g lipid weight (lw) and the MLOQ was generally 1.0 ng/g lw.
Elements analyses were performed at the accredited (DANAK, DSyEN ISOyIEC17025, No. 435) NERI-DAE Laboratory (Roskilde, Denmark; www.bios.au.dk) byFlow Injection Mercury System (mercury), Atomic Absorption Spectrometry (zincand iron) and Flow Injection Analyze System (selenium) as described by Asmundet al. (2004) and Dietz et al. (2006).
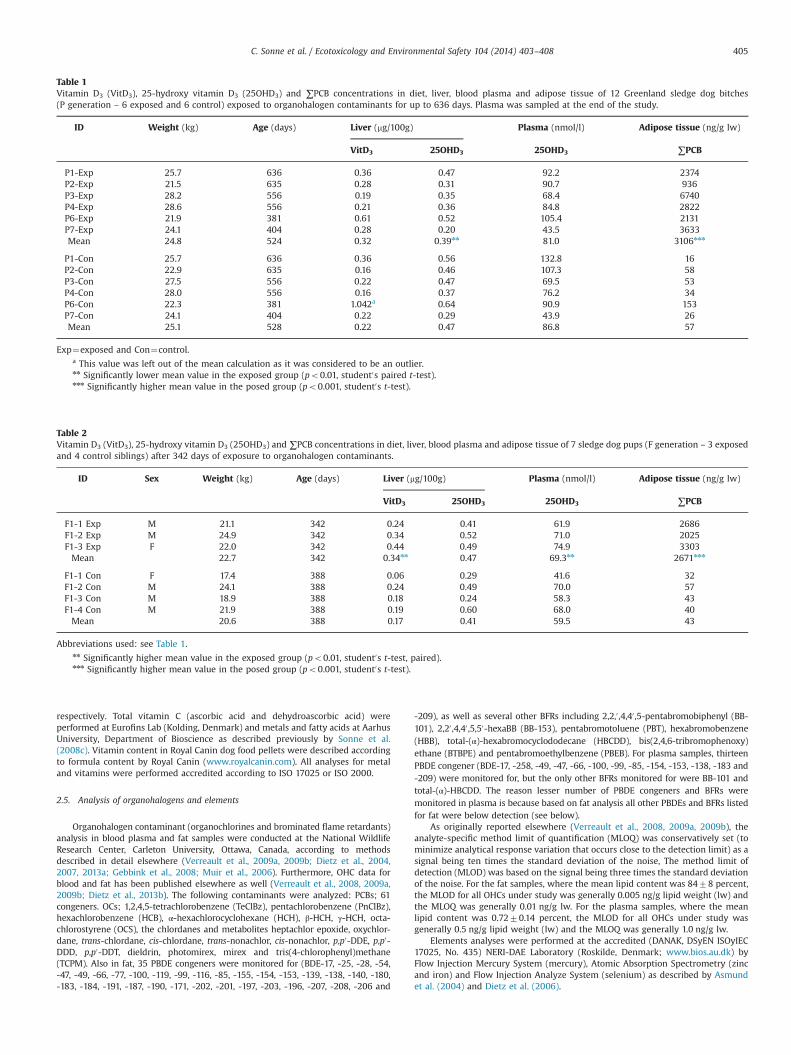
Table 1Vitamin D3 (VitD3), 25-hydroxy vitamin D3 (25OHD3) and ∑PCB concentrations in diet, liver, blood plasma and adipose tissue of 12 Greenland sledge dog bitches(P generation – 6 exposed and 6 control) exposed to organohalogen contaminants for up to 636 days. Plasma was sampled at the end of the study.
ID Weight (kg) Age (days) Liver (mg/100g) Plasma (nmol/l) Adipose tissue (ng/g lw)
Exp¼exposed and Con¼control.a This value was left out of the mean calculation as it was considered to be an outlier.nn Significantly lower mean value in the exposed group (po0.01, student0s paired t-test).nnn Significantly higher mean value in the posed group (po0.001, student0s t-test).
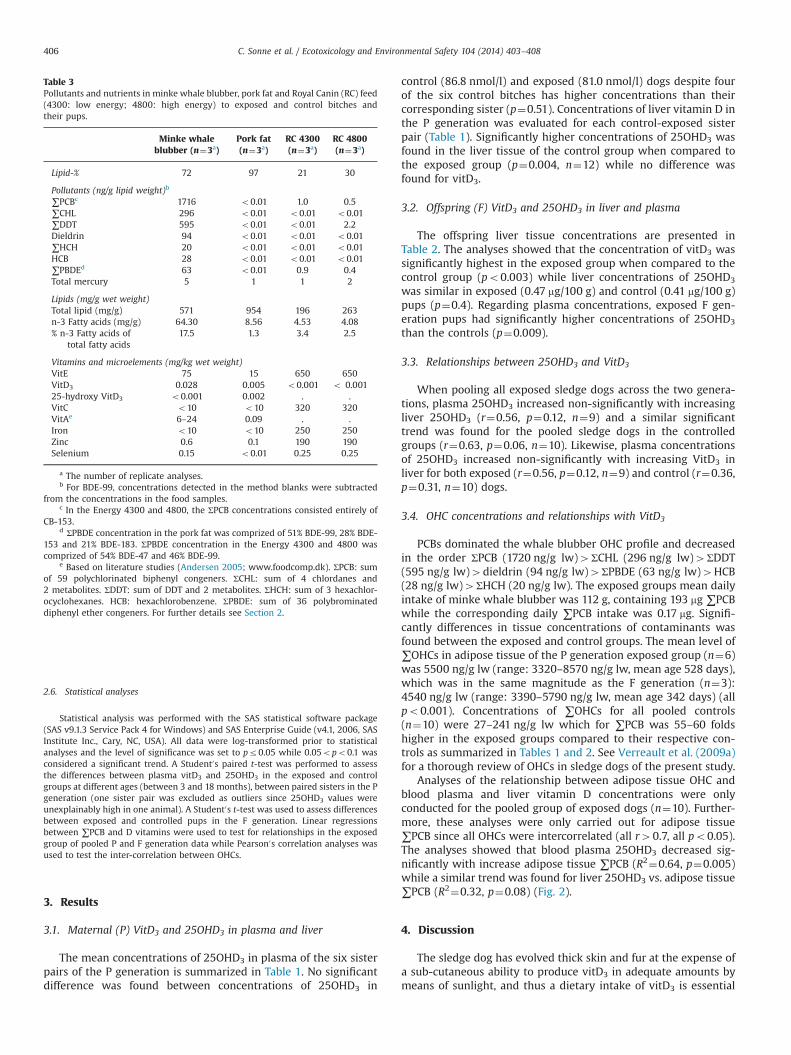
Table 2Vitamin D3 (VitD3), 25-hydroxy vitamin D3 (25OHD3) and ∑PCB concentrations in diet, liver, blood plasma and adipose tissue of 7 sledge dog pups (F generation – 3 exposedand 4 control siblings) after 342 days of exposure to organohalogen contaminants.
ID Sex Weight (kg) Age (days) Liver (mg/100g) Plasma (nmol/l) Adipose tissue (ng/g lw)
F1-1 Con F 17.4 388 0.06 0.29 41.6 32F1-2 Con M 24.1 388 0.24 0.49 70.0 57F1-3 Con M 18.9 388 0.18 0.24 58.3 43F1-4 Con M 21.9 388 0.19 0.60 68.0 40Mean 20.6 388 0.17 0.41 59.5 43
Abbreviations used: see Table 1.nn Significantly higher mean value in the exposed group (po0.01, student0s t-test, paired).nnn Significantly higher mean value in the posed group (po0.001, student0s t-test).
C. Sonne et al. / Ecotoxicology and Environmental Safety 104 (2014) 403–408 405
Statistical analysis was performed with the SAS statistical software package(SAS v9.1.3 Service Pack 4 for Windows) and SAS Enterprise Guide (v4.1, 2006, SASInstitute Inc., Cary, NC, USA). All data were log-transformed prior to statisticalanalyses and the level of significance was set to pr0.05 while 0.05opo0.1 wasconsidered a significant trend. A Student0s paired t-test was performed to assessthe differences between plasma vitD3 and 25OHD3 in the exposed and controlgroups at different ages (between 3 and 18 months), between paired sisters in the Pgeneration (one sister pair was excluded as outliers since 25OHD3 values wereunexplainably high in one animal). A Student0s t-test was used to assess differencesbetween exposed and controlled pups in the F generation. Linear regressionsbetween ∑PCB and D vitamins were used to test for relationships in the exposedgroup of pooled P and F generation data while Pearson0s correlation analyses wasused to test the inter-correlation between OHCs.
3. Results
3.1. Maternal (P) VitD3 and 25OHD3 in plasma and liver
The mean concentrations of 25OHD3 in plasma of the six sisterpairs of the P generation is summarized in Table 1. No significantdifference was found between concentrations of 25OHD3 in
control (86.8 nmol/l) and exposed (81.0 nmol/l) dogs despite fourof the six control bitches has higher concentrations than theircorresponding sister (p¼0.51). Concentrations of liver vitamin D inthe P generation was evaluated for each control-exposed sisterpair (Table 1). Significantly higher concentrations of 25OHD3 wasfound in the liver tissue of the control group when compared tothe exposed group (p¼0.004, n¼12) while no difference wasfound for vitD3.
3.2. Offspring (F) VitD3 and 25OHD3 in liver and plasma
The offspring liver tissue concentrations are presented inTable 2. The analyses showed that the concentration of vitD3 wassignificantly highest in the exposed group when compared to thecontrol group (po0.003) while liver concentrations of 25OHD3
was similar in exposed (0.47 mg/100 g) and control (0.41 mg/100 g)pups (p¼0.4). Regarding plasma concentrations, exposed F gen-eration pups had significantly higher concentrations of 25OHD3
than the controls (p¼0.009).
3.3. Relationships between 25OHD3 and VitD3
When pooling all exposed sledge dogs across the two genera-tions, plasma 25OHD3 increased non-significantly with increasingliver 25OHD3 (r¼0.56, p¼0.12, n¼9) and a similar significanttrend was found for the pooled sledge dogs in the controlledgroups (r¼0.63, p¼0.06, n¼10). Likewise, plasma concentrationsof 25OHD3 increased non-significantly with increasing VitD3 inliver for both exposed (r¼0.56, p¼0.12, n¼9) and control (r¼0.36,p¼0.31, n¼10) dogs.
3.4. OHC concentrations and relationships with VitD3
PCBs dominated the whale blubber OHC profile and decreasedin the order ΣPCB (1720 ng/g lw)4ΣCHL (296 ng/g lw)4ΣDDT(595 ng/g lw)4dieldrin (94 ng/g lw)4ΣPBDE (63 ng/g lw)4HCB(28 ng/g lw)4ΣHCH (20 ng/g lw). The exposed groups mean dailyintake of minke whale blubber was 112 g, containing 193 mg ∑PCBwhile the corresponding daily ∑PCB intake was 0.17 mg. Signifi-cantly differences in tissue concentrations of contaminants wasfound between the exposed and control groups. The mean level of∑OHCs in adipose tissue of the P generation exposed group (n¼6)was 5500 ng/g lw (range: 3320–8570 ng/g lw, mean age 528 days),which was in the same magnitude as the F generation (n¼3):4540 ng/g lw (range: 3390–5790 ng/g lw, mean age 342 days) (allpo0.001). Concentrations of ∑OHCs for all pooled controls(n¼10) were 27–241 ng/g lw which for ∑PCB was 55–60 foldshigher in the exposed groups compared to their respective con-trols as summarized in Tables 1 and 2. See Verreault et al. (2009a)for a thorough review of OHCs in sledge dogs of the present study.
Analyses of the relationship between adipose tissue OHC andblood plasma and liver vitamin D concentrations were onlyconducted for the pooled group of exposed dogs (n¼10). Further-more, these analyses were only carried out for adipose tissue∑PCB since all OHCs were intercorrelated (all r40.7, all po0.05).The analyses showed that blood plasma 25OHD3 decreased sig-nificantly with increase adipose tissue ∑PCB (R2¼0.64, p¼0.005)while a similar trend was found for liver 25OHD3 vs. adipose tissue∑PCB (R2¼0.32, p¼0.08) (Fig. 2).
4. Discussion
The sledge dog has evolved thick skin and fur at the expense ofa sub-cutaneous ability to produce vitD3 in adequate amounts bymeans of sunlight, and thus a dietary intake of vitD3 is essential
Table 3Pollutants and nutrients in minke whale blubber, pork fat and Royal Canin (RC) feed(4300: low energy; 4800: high energy) to exposed and control bitches andtheir pups.
a The number of replicate analyses.b For BDE-99, concentrations detected in the method blanks were subtracted
from the concentrations in the food samples.c In the Energy 4300 and 4800, the ΣPCB concentrations consisted entirely of
CB-153.d ΣPBDE concentration in the pork fat was comprized of 51% BDE-99, 28% BDE-
153 and 21% BDE-183. ΣPBDE concentration in the Energy 4300 and 4800 wascomprized of 54% BDE-47 and 46% BDE-99.
e Based on literature studies (Andersen 2005; www.foodcomp.dk). ΣPCB: sumof 59 polychlorinated biphenyl congeners. ΣCHL: sum of 4 chlordanes and2 metabolites. ΣDDT: sum of DDT and 2 metabolites. ΣHCH: sum of 3 hexachlor-ocyclohexanes. HCB: hexachlorobenzene. ΣPBDE: sum of 36 polybrominateddiphenyl ether congeners. For further details see Section 2.
C. Sonne et al. / Ecotoxicology and Environmental Safety 104 (2014) 403–408406
and therefore this species is depending on a sufficient dietaryintake (How et al., 1994; Kenny et al., 2004). Based on the dailyfood intake, the exposed sledge dogs of both P and F generationreceived app. 33 percent more dietary vitD3 than the controls for aperiod of 25 days prior to sampling. Despite this, the concentra-tions of 25OHD3 in liver were significantly lower in the exposed Pgeneration. The reason for the lower 25OHD3 in liver tissue of theexposed P generation dogs despite their higher intake comparedto controls remains speculative. One suggestion could be thattransport proteins, e.g. albumin and vitamin D binding protein(DBP), are affected by OHCs. Thus, vitD3 may not be adequatelytransported to the liver, consequently resulting in a lesser hepaticbiotransformation of vitD3 to plasma 25OHD3. In humans, DBP,albumin and lipoproteins are the principle carriers of vitD3, andDBP and albumin are both members of the albumin gene familyand synthesized in the liver (Birn and Christensen, 2006). Themechanism of an albumin and vitD3 correlation is not clearlyunderstood as yet (Wolf et al., 2007) but in humans, the affinity ofalbumin for 25OHD3 is substantially lower than that of the DBP.
The vitD3 concentration in liver showed a declining non-significant trend with increasing ∑PCB concentrations in the liverof exposed dogs. This is partly in accordance with a study onringed seals that showed that persistent organic pollutants likePCBs disrupt vitD3 metabolism (1,25OHD3) (Routti et al., 2008).In the same study, Routti et al. (2008) proposed that bone lesionsin Baltic gray seals may be associated with contaminant mediatedvitD3 and thyroid disruption, as a clear relationship betweencirculating 1,25OHD3, calcium, phosphate and thyroid and hepaticPCB and DDT load was observed. 1,25OHD3 was not measured inthe present study and the female seals studied by Routti et al.(2008) exhibited uterine occlusions and infertility, presumablybecause of higher concentrations of accumulated OHCs (∑PCB inliver: Baltic ringed seals, mean 34,000722,000 ng/g lw and Balticgray seals, mean 59,000740,000 ng/g lw and for the present dogstudy ΣPCB in liver: exposed P mean 43,111 ng/g lw and exposed F
mean 20,065 ng/g lw). Furthermore, low sample size or speciesspecific vitD3 dynamics could suggest the difference.
Another suggestion for the difference in vitamin 25OHD3
concentrations between the groups of P generation dogs despitedifferent dietary levels could be drug–drug interaction. VitD3 ishydroxylated to 25OHD3 in the microsomes and mitochondria inliver by the CYP-25-hydroxylation enzymes. One suggested CYPenzyme mediating the hydroxylation of vitD3 in microsomes isCYP2R1, which is preferentially expressed in the liver, and is notsexually dimorphic. Furthermore, CYP2R1 is highly conservedamong species ranging from fish to human (Cheng et al., 2003,2004). The ability of a xenobiotic substrate that up-regulateshepatic CYPs such as OHC exposure by affecting the activity of anatural substrate (e.g. vitD3 in liver) is a drug–drug interaction(Nebert and Russell, 2002). VitD3 is a seco-steroid hormone, andVerreault et al. (2009b) have shown that testosterone hydroxyla-tion activity resulting in the formation of various hydroxylatedtestosterone metabolites (and thus mediated by various CYPs) wasgreater in dogs from the present exposed generations relative tothe controls. This suggests that the induction of various CYPs andperhaps CYP2R1 as well, may explain the increased vitD3 to25OHD3 metabolism, which subsequently resulted in lower livervitD3 levels of the exposed P generation. Furthermore, there maybe increased metabolism of 25OHD3 in plasma to 1,25OHD3 intissues and kidney, as 25OHD3 in liver was lower in the exposedgroup (relative to controls). This indicated a greater need for1,25OHD3 in tissues of OHC exposed bitches.
Another aspect of vitD3 transport disruption could be fattyacids in the diet. One study concluded that n-3 polyunsaturated (inour case whale blubber) fatty acids, opposed to saturated (in ourcase porcine) fatty acids, are capable of decreasing the binding of25OHD3 to DBP in humans (Buillon et al., 1992). Another studyconclude that DBP possesses relatively weaker binding to fattyacids than albumin, so when albumin is depressed, DBP does nottake over it in terms of fatty acid scavenging and transportation(Swamy and Ray, 2008). Whether this possible inhibitory potencyis the same for canine DBP and the n-3 polyunsaturated fatty acidsfrom whale blubber given to the exposed group of dogs in thisstudy is unknown.
5. Conclusions
We compared vitamin D status in OHC exposed sledge dogswith controls. The study showed that 25OHD3 in liver tissue ofexposed bitches was significantly lower than in controls, whilevitD3 in liver tissue and 25OHD3 in plasma of exposed offspringwas highest. This means that the homeostasis and metabolismof vitamin D may respond differently to the dietary compositionof fatty acids and OHC exposure. It is unknown if the lower level of25OHD3 in the liver of exposed dogs would have any negativeeffects on immunity and reproduction why more focus should beconducted on this compound in Arctic wildlife.
Acknowledgments
The Danish Cooperation for Environment in the Arctic (DANCEA),Natural Science and Engineering Research Council (NSERC) Canada,Royal Canin, Kruuse Aps, EU Endoclima Marie Curie Training Site(NTNU) are acknowledged for financial support. T.D. Rasmussen, M.J.Rasmussen and S. Shahmiri (Organic Contaminants Research Labora-tory/Letcher Group, NWRC, Environment Canada) are thanked forcaring for the sledge dogs and chemical analyses of the sledge dogtissues, respectively.
Fig. 2. Blood and liver 25-hydroxy vitamin D3 (25OHD3) vs. adipose tissue ∑PCB insix OHC exposed sledge dog bitches and their three pups.
C. Sonne et al. / Ecotoxicology and Environmental Safety 104 (2014) 403–408 407
References
Andersen, SM, Vitamins and minerals in the traditional Greenland diet. NationalEnvironmental Research Institute, Denmark 2005. NERI Technical Report no.528, http://technical-reports.dmu.dk, 44pp.
Asmund, G., Vorkamp, K., Backus, S., Comba, M., 2004. An update on analyticalmethods, quality assurance and quality control used in the Greenland AMAPprogramme 1999 to 2002. Sci. Total Environ. 331, 233–245.
Birn, H., Christensen, E.I., 2006. Renal albumin absorption in physiology andpathology. Kidney Int. 69, 440–449.
Buillon, R., Xiano, D.Z., Corrs, R., Bafelen, H.V., 1992. Polyunsaturated fatty acidsdecrease the apparent affinity of vitamin D metabolites for human vitaminD-binding protein. J. Steroid Biochem. Mol. Biol. 42, 855–861.
Cheng, J.B., Levine, M.A., Bell, N.H., Mangelsdorf, D.J., Russell, D.W., 2004. Geneticevidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase.PNAS 101, 7711–7715.
Dietz, R., Riget, F.F., Born, E.W., Sonne, C., Grandjean, P., Kirkegaard, M., Olsen, M.T.,Asmund, G., Renzoni, A., Baagøe, H., Andreasen, C., 2006. Trends in mercury inhair of Greenlandic polar bears (Ursus maritimus) during 1892–2001. Environ.Sci. Technol. 2006 (40), 1120–1125.
Dietz, R., Riget, F.F., Sonne, C., Letcher, R.J., Backus, S., Born, E.W., Kirkegaard, M.,Muir, D.C.G., 2007. Age and seasonal variability of polybrominated diphenylethers in free-ranging East Greenland polar bears (Ursus maritimus). Environ.Pollut. 146, 166–173.
Dietz, R., Riget, F.F., Sonne, C., Letcher, R.J., Born, E.W., Muir, D.C.G., 2004. Seasonaland temporal trends in polychlorinated biphenyls and organochlorine pesti-cides in East Greenland polar bears (Ursus maritimus), 1990–2001. Sci. TotalEnviron. 331, 107–124.
Dietz, R., Rigét, F.F., Sonne, C., Born, E.W., Bechshøft, T., McKinney, M.A., Letcher, R.J.,2013a. Part 1: Three decades (1984–2010) of legacy contaminant trends in EastGreenland polar bears (Ursus maritimus). Environ. Int. 59, 485–493.
Dietz, R., Rigét, F.F., Sonne, C., Born, E.W., Bechshøft, T., McKinney, M.A., Drimmei, R.,Muir, D.C.G., Letcher, R.J., 2013b. Part 2: Three decades (1984–2010) of flameretardant trends in East Greenland polar bears (Ursus maritimus). Environ. Int.59, 494–500.
EN12822, 2000. Foodstuffs – Determination of Vitamin E by High PerformanceLiquid Chromatography. Measurement of α-, β-, λ-, δ-tocopherol. EuropeanCommittee for Standardization, Rue de Stassart, 36, B-1050 Bruxelles.
EN12823-1, 2000. Foodstuffs – Determination of Vitamin A by High PerformanceLiquid Chromatography. Measurement of all-trans and 13-cis Retinol. EuropeanCommittee for Standardization, Rue de Stassart, 36, B-1050 Bruxelles.
Feldman, D., Pike, J.W., Glorieux, F.H. (Eds.), 2005. Vitamin D, 2nd ed. ElsevierAcademic, San Diego, CA, USA.
Gebbink, W.A., Sonne, C., Dietz, R., Kirkegaard, M., Born, E.W., Muir, D.C.G., Letcher,R.J., 2008. Target tissue selectivity and burdens of diverse classes of brominatedand chlorinated contaminants in polar bears (Ursus maritimus) from EastGreenland. Environ. Sci. Technol. 42, 752–759.
Hollis, B.W., 2005. Circulating 25-hydroxyvitamin d levels indicative of vitamin Dsufficiency: implications for establishing a new effective dietary intake recom-mendation for vitamin D. J. Nutr. 135, 317–322.
How, K.L., Hazewinkel, H.A.W., Mol, J.A., 1994. Daily Vitamin D dependence of catand dog due to inadequate cutaneous synthesis of vitamin D. Gen. Comp.Endocrinol. 96, 12–18.
Jakobsen, J., Clausen, I., Leth, T., Ovesen, L., 2004. A new method for thedetermination of vitamin D3 and 25-hydroxyvitamin D3 in meat. J. FoodCompos. Anal. 17, 777–787.
Jakobsen, J., Maribo, H., Bysted, A., Sommer, H.M., Hels, O., 2007. 25-hydroxyvitamin D3 affects vitamin D status similar to vitamin D3 in pigs –
but the meat produced has a lower content of vitamin D. Br. J. Nutr. 98,908–913.
Jakobsen, J., Bysted, A., Andersen, R., Bennett, T., Brot, C., Brugel, S., Chashman, K.D.,Denk, E., Harrington, M., Teucher, B., Walczyk, T., Ovesen, L., 2009. Vitamin Dstatus assessed by a validated HPLC method: within and between variation insubjects supplemented with vitamin D3. Scand. J. Clin. Lab Invest. 69 (2),190–197.
Kalueff, A.V., Lou, Y.R., Laaksi, I., Tuohimaa, P., 2004. Increased anxiety in micelacking vitamin D receptor gene. Neuroreport 15, 1271–1274.
23, 33–43. (References and further reading may be available for this article. Toview references and further reading you must purchase this article).
Kinuta, K., Tanaka, H., Moriwake, T., Aya, K., Kato, S., Seino, Y., 2000. Vitamin D is animportant factor in estrogen biosynthesis of both female and male gonads.Endocrinology 141, 1317–1324.
Li, Y.C., Guilin, Qiao, Milan, Uskokovic, Wei, Xiang, Wei, Zheng, Juan, Kong, 2004.Vitamin D: a negative endocrine regulator of the renin–angiotensin system andblood pressure. J. Steroid Biochem. 89-90, 387–392.
Lilienthal, H., 2000. Reduced levels of 1,25-dihydroxyvitamin D(3) in rat dams and520 offspring after exposure to a reconstituted PCB mixture. Toxicol. Sci. 57,292–301.
Lou, Y.-L., Laaksi, I., Syvala, H., Bluer, M., Tammela, T.L.J., Ylikomi, T., Tuohimaa, P.,2004. 25-Hydroxyvitamin D3 is an active hormone in human primary prostaticstromal cells. FASEB 18, 332–334.
Muir, D.C.G., Backus, S., Derocher, A.E., Dietz, R., Evans, T.J., Gabrielsen, G.W., Nagy,J., Nordstrom, R.J., Sonne, S., Stirling, I., Taylor, M.K., Letcher, R.J., 2006.Brominated flame retardants in polar bears (Ursus maritimus) from Alaska,the Canadian Arctic, Greenland and Svalbard. Environ. Sci. Technol. 40,449–455.
Nebert, D.W., Russell, D.W., 2002. Clinical importance of the cytochromes P450.Lancet 360, 1155–1162.
Norman, A.W., 2008. From vitamin D to hormone D: fundamentals of the vitamin Dendocrine system essential for good health. Am. J. Clin. Nutr. 88, 491S–499S.
Routti, H., Nyman, M., Jenssen, B.M., Backman, C., Koistinen, J., Gabrielsen, G.W.,2008. Bone-related effects of contaminants in seals may be associated withvitamin D and thyroid hormones. Environ. Toxicol. Chem. 27, 873–880.
Sonne, C., Dietz, R., Kirkegaard, M., Letcher, R.J., Shahmiri, S., Andersen, S., Møller, P.,Olsen, A.K., Jensen, A.L., 2008a. Effects of organohalogen pollutants on haema-tological and urine clinical-chemical parameters in Greenland sledge dogs(Canis familiaris). Ecotoxicol. Environ. Saf. 69, 381–390.
Sonne, C., Dietz, R., Larsen, H.J.S., Loft, K.E., Kirkegaard, M., Letcher, R.J., Shahmiri, S.,Møller, P., 2006. Impairment of cellular immunity in West Greenland sledgedogs (Canis familiaris) dietary exposed to polluted minke whale (Balaenopteraacutorostrata) blubber. Environ. Sci. Technol. 40, 2056–2062.
Sonne, C., Fonfara, S., Dietz, R., Kirkegaard, M., Letcher, R.J., Shahmiri, S., Andersen,S., Møller, P., 2007a. Multiple cytokine and acute phase protein gene transcrip-tion in West Greenland sledge dogs (Canis familiaris) dietary exposed to organicenvironmental pollutants. Arch. Environ. Contam. Toxicol. 53, 110–118.
Sonne, C., Leifsson, P.S., Dietz, R., Kirkegaard, M., Jensen, A.L., Shahmiri, S., Letcher,J.L., 2008b. Greenland sled dogs (Canis familiaris) develop liver lesions whenexposed to a chronic and dietary low dose of an environmental organohalogencocktail. Environ. Res. 106, 72–80.
Sonne, C., Leifsson, P.S., Dietz, R., Kirkegaard, M., Møller, P., Jensen, A.L., Letcher, R.J.,Shahmiri, S., 2007b. Renal lesions in Greenland sledge dogs (Canis familiaris)exposed to a natural dietary cocktail of persistent organic pollutants. Toxicol.Environ. Chem. 89, 563–576.
Sonne, C., Rigét, F.F., Jensen, J.-E.B., Hyldstrup, L., Teilmann, J., Dietz, R., Kirkegaard,M., Andersen, S., Letcher, R.J., Jakobsen, J., 2008c. Does the nutrition profile ofvitamins, fatty acids and microelements counteract the negative impact fromorganohalogen pollutants on bone mineral density in Greenland sledge dogs(Canis familiaris)? Environ. Int. 34, 811–820.
Swamy, N., Ray, R., 2008. Fatty acid-binding site environments of serum vitaminD-binding protein and albumin are different. Bioorg. Chem. 36, 165–168.
Tuohimaa, P., Golovko, O., Kalueff, A., Nazarova, N., Qiao, S., Syvala, H., Talonpoika, R.,Lou, Y.-R., 2005. Calcidiol and prostate cancer. J. Steroid Biochem. 93, 183–190.
Uitterlinden, A.G., Fang, Y., Van Meurs, J.B., Pols, H.A., Van Leeuwen, J.P., 2004.Genetics and biology of vitamin D receptor polymorphisms. Gene 338,143–156.
Verreault, J., Dietz, R., Sonne, C., Gebbink, W.A., Shahmiri, S., Letcher, R.J., 2008.Comparative fate of organohalogen contaminants in two top carnivores inGreenland: captive sledge dogs and free-ranging polar bears. Comp. Biochem.Physiol. Part C 147, 306–315.
Verreault, J., Letcher, R.J., Christian, S., Dietz, R., 2009a. Dietary, age and trans-generational effects on the fate of organohalogen contaminants in captivesledge dogs in Greenland. Environ. Int. 35, 56–62.
Verreault, J., Maisonneuve, F., Dietz, R., Sonne, C., Letcher, R.J., 2009b. Comparativehepatic activity of xenobiotic-metabolizing enzymes and concentrations oforganohalogens and their hydroxylated analogues in captive Greenland sledgedogs. Environ. Toxicol. Chem. 28, 162–172.
Wolf, M., Shah, A., Gutierrez, O., Ankers, E., Monroy, M., Tamez, H., Steele, D., Chang,Y., Camargo Jr., C.A., Tonelli, M., Thadhani, R., 2007. Vitamin D levels and earlymortality among incident hemodialysis patients. Kidney Int. 72, 1004–1013.
C. Sonne et al. / Ecotoxicology and Environmental Safety 104 (2014) 403–408408