Signal transduction of somatostatin inhuman B lymphoblasts
DIETER ROSSKOPF, MARKUS SCHURKS, IRIS MANTHEY,MARKUS JOISTEN, STEFAN BUSCH, AND WINFRIED SIFFERTInstitut fur Pharmakologie, Universitatsklinikum Essen, D-45122 Essen, GermanySubmitted 28 March 2001; accepted in final form 11 September 2002
Rosskopf, Dieter, Markus Schurks, Iris Manthey,Markus Joisten, Stefan Busch, and Winfried Siffert.Signal transduction of somatostatin in human B lympho-blasts. Am J Physiol Cell Physiol 284: C179–C190, 2003.First published September 18, 2002; 10.1152/ajpcell.00160.2001.—Somatostatin (SST) and somatostatin receptors(SSTR) are widely distributed in lymphoid tissues. Here, wereport on the stimulatory effects of SST in Epstein-Barrvirus-immortalized B lymphoblasts. By RT-PCR, we demon-strated the exclusive expression of the somatostatin receptorisoform 2A (SSTR2A) in B lymphoblasts. Addition of SSTrapidly increased the cytosolic free calcium concentration[Ca2�]i maximally by about 200 nM, with an EC50 of 1.3 nM,and stimulated the formation of inositol phosphates. Fur-thermore, SST increased binding of guanosine 5�-O-(3-thio-triphosphate) by 50% above basal. These effects were partlyinhibited by pertussis toxin (PTX), which indicates the in-volvement of PTX-sensitive G proteins. We provide furtherevidence that G�16, a PTX-insensitive G protein confined tolymphohematopoietic cells, is involved in the otherwise un-usual coupling of SSTR2A to phospholipase C activation. Inaddition, SST activated extracellular regulated kinases andinduced a 3.5-fold stimulation of DNA synthesis and a 4.4-fold stimulation of B lymphoblast proliferation, which wasaccompanied by an enhanced immunoglobulin formation.Thus SST exerts a growth factor-like activity on human Blymphoblasts.
G protein; immunoglobulin formation; MAP kinase; pertussistoxin; phospholipase C
B LYMPHOCYTES ARE THE PRINCIPAL mediators of adaptivehumoral immunity. The differentiation and selectionprocesses from quiescent B lymphocytes to antibody-secreting plasma cells and memory cells are governedby specific antigen receptors and a complex network ofmodulating signals in a specialized lymphoid environ-ment (10). These modulating signals are generated byselective cytokines, B cell coreceptors, and cell adhe-sion molecules confined to the lymphoid system. Inaddition, hormones and neurotransmitters that regu-late many different physiological systems are also in-volved in B cell control (9). Such agonists include plate-let-activating factor (PAF; Ref. 28), lysophosphatidicacid (LPA; Ref. 44), vasoactive intestinal polypeptide
(VIP; Refs. 17 and 18), and catecholamines (26), toname but a few.
Somatostatin (SST), a cyclic tetradecapeptide, wasfirst described as a potent inhibitor of growth hormonesecretion (39). Subsequent studies have shown thatSST has widespread physiological functions in hor-mone release, regulation of exocrine secretion, modu-lation of neural activity, and the inhibition of tumorgrowth (reviewed in Refs. 34, 39, and 46).
SST binding sites were first detected on circulatinghuman blood lymphocytes (5) but also on many lym-phoid tissues (42, 43). Somatostatin-receptor (SSTR)-based imaging has been widely used for the diagnosisof malignant lymphomas and hyperplastic or granulo-matous nonmalignant lymph nodes (31, 42, 43).
Despite this widespread distribution in lymphoidtissues, only limited information on the physiologicalroles of SST and SSTRs in the immune system exists(reviewed in Refs. 53 and 54). Several groups of inves-tigators have used lymphocytes to study SST effects onintracellular effector systems, including adenylyl cy-clase and the Na�/H� exchanger (23, 34). Both anti-proliferative and growth-promoting activities of SSThave been reported (27, 36, 37). More recent reportshave shown that SST regulates T cell interferon-� andinterleukin (IL)-2 release (7, 11). However, for the Bcell system, such functional data on the role of SST andits receptors are lacking.
Five different human SST receptor subtypes(SSTR1–SSTR5) have been cloned (16, 33, 34, 46, 58),and splice variants of SSTR2 exist (35). All SSTRsbelong to the family of G protein-coupled or heptaheli-cal receptors but differ with respect to their tissue-specific distribution and pharmacological properties(16, 33).
The predominantly inhibitory cellular actions of SSTare mediated by multiple effector pathways (23, 33,34), including the inhibition of adenylyl cyclase (19)and voltage-dependent Ca2� channels (25), a reducedmobilization of intracellular Ca2� (19), and the atten-uation of Na�/H� exchange activity. Furthermore, SSTstimulates voltage-dependent K� channels (59) andprotein tyrosine phosphatases (6), the activation of
Address for reprint requests and other correspondence: D. Rosskopf,Institut fur Pharmakologie, Universitatsklinikum Essen, Hufelandstr.55, D-45122 Essen, Germany (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Cell Physiol 284: C179–C190, 2003.First published September 18, 2002; 10.1152/ajpcell.00160.2001.
which coincides with an antiproliferative effect viaproapoptotic pathways (34). SST has also been re-ported to activate phospholipase A2 and the MAP ki-nase cascade (34). In some cell lines, predominantly onoverexpression of SSTRs, SST stimulates phospho-lipase C (PLC) activity (2, 8, 29, 47, 51).
Most effects of SST are transmitted via pertussistoxin (PTX-)-sensitive G proteins, although partiallyPTX-insensitive effects including activation of PLChave been reported (8, 48, 51). B lymphoblasts expressthe PTX-sensitive G proteins G�i2 and G�i3 but notG�i1 and G�o (44). G proteins of the G�q family arepredominantly involved in activation of PLC� isoforms(40), and G�q and G�11 are expressed in B lympho-blasts. Highly restricted to lymphohematopoietic cells,an additional G�q class protein exists, G�16 (and itsmurine homolog G�15; Refs. 3 and 57), which is able tolink “promiscuously” numerous heptahelical receptorsto PLC activation, a peculiarity that is not observedwith other G�q class proteins (32, 57).
Here, we investigated the effects of SST on prolifer-ation and immunoglobulin formation of human B lym-phoblasts, and we characterized the early signal trans-duction of SST in these cells. We report the novelobservation that SST stimulates PLC activity and in-creases [Ca2�]i in B lymphoblasts, events that mostlikely involve the coupling of SSTR2A to G�16 andPLC�2. These stimulatory effects of SST on early sig-nal transduction are accompanied by an increased cellproliferation and immunoglobulin formation.
MATERIALS AND METHODS
Materials. Fura 2-AM was purchased from MolecularProbes (Eugene, OR). [3H]-methylthymidine was boughtfrom Hartmann Analytics (Braunschweig, Germany). [35S]-guanosine 5�-O-(3-thiotriphosphate ([35S]GTP�S) (specificactivity 1,200–1,400 Ci/mmol) was from DuPont-NEN (BadHomburg, Germany), and myo-[3H]-inositol was from Amer-ican Radiolabed Chemicals (St. Louis, MO). Somatostatin(SST-14) and PAF were purchased from Calbiochem (BadSoden, Germany), and 12-O-tetradecanoylphorbol 13-acetate(TPA) was from Sigma (Deisenhofen, Germany). Primary andsecondary antibodies for immunoglobulin ELISA were pur-chased from Tago Immunologicals (Burlingame, CA). Unla-beled nucleotides were from Boehringer Mannheim, and PTXwas from List Biological Laboratories (Campbell, CA). Thephosphospecific p42/44 MAP kinase antibody was from NewEngland Biolabs (Schwalbach, Germany), and the polyclonalanti-ERK1/ERK2 antiserum was from Santa Cruz Biotech-nology (Heidelberg, Germany). Reverse transcriptase (RT;Superscript) was from Life Technologies (Eggenstein, Ger-many). Taq polymerase, RNasin, RNase-free DNase, andrestriction enzymes were purchased from Fermentas (St.Leon-Rot, Germany), and Pfu-Taq polymerase was from Pro-mega (Heidelberg, Germany).
B lymphoblast cell lines and cell culture. Human B lym-phoblast cell lines were derived from peripheral blood lym-phocytes and immortalized with Epstein-Barr virus as de-scribed (44, 48). They were cultured in RPMI 1640 mediumcontaining 2 mM L-glutamine, which was supplemented with100 U/ml penicillin, 100 �g/ml streptomycin, and 10% FCS(all from Life Technologies). If not indicated otherwise, cellswere routinely subcultured into fresh RPMI 1640 medium
with 10% FCS at a density of 106 cells/ml 1 day before theexperiment.
Detection of SSTRs by reverse transcription-polymerasechain reaction (RT-PCR). Lymphoblast RNA was preparedand reverse transcribed using Superscript reverse transcrip-tase as described (44). Before reverse transcription, RNA wastreated with RNase-free DNase according to the manufactur-er’s instructions. PCR primer sequences were taken from thereport by Kubota et al. (22). According to the publishedsequence for human SSTR2 (58), the reverse primer sequencewas modified to 5�-TCACCATGATCTGTCTTTGC-3�. PCRamplicons flanked by these primers contained at least onesingle restriction site, and the specificity of the synthesizedPCR products was confirmed by restriction analysis. Todiscriminate the splice variant SSTR2A from SSTR2B, RT-PCRs were repeated using the oligonucleotide primers5�-CTTCCGTCTCCATGGCCATCAGC-3� and 5�-GGTAAT-GCCTATACAGAGAATAAATA GG-3�. The specific ampli-cons generated with these primers comprise 686 base pair(bp) for SSTR2A transcripts and for genomic DNA. InSSTR2B, a fragment of 341 bp is removed by alternativesplicing, resulting in an amplicon of only 345 bp flanked bythese oligonucleotide primers (38). PCRs were carried out inreaction volumes of 50 �l containing 200 �M of each dNTP,10 pmol of each oligonucleotide primer, cDNA correspondingto 100 ng RNA, and 2.5 U Taq polymerase in reaction buffersupplied by the manufacturer. The amplification profile in-volved denaturation at 94°C (30 s), annealing at 55°C (45 s),and extension at 72°C (90 s) for 35 cycles. A negative controlcontaining 100 ng of RNA instead of cDNA was included ineach experiment. Because all SSTRs lack introns, genomicDNA was used as positive control. PCR products were ana-lyzed by electrophoresis on 1.5% agarose gels and stainedwith ethidium bromide. Primer sequences were derivedfrom the following GenBank entries: M81829 (SSTR1),M81830 (SSTR2A), L13033 (SSTR2B), AF184174 (SSTR2B),XM_009963 (SSTR3), NT_011387 (SSTR4), and NT_010552(SSTR5).
Measurement of cytoplasmic free Ca2�. Changes in intra-cellular free Ca2� concentration ([Ca2�]i) were measured, asdescribed in detail (44, 48), using the calcium-sensitive fluo-rescent dye fura 2. One day before the experiment, B lym-phoblasts were subcultured at a density of 2–5 � 105 cells/mlin RPMI 1640 medium containing antibiotics and 0.5% FCS.Dye loading, measurements of fluorescence, and calibrationof emission ratios (� 495 nm) after rapid alternatingexcitation wavelengths from � 340 to 380 nm were per-formed, exactly as described, in a Perkin-Elmer LS5B spec-trofluorimeter (44, 48). For each experiment, 1 � 107 dye-loaded cells were measured at 37°C in HEPES-buffercontaining 1 mM CaCl2 (44, 48). For experiments in Ca2�-free medium, EGTA (5 mM final) was added 5 s beforeaddition of agonist.
Analysis of inositol phosphate formation. Formation ofinositol phosphate (IP) was quantified in lymphoblasts grownat a density of 2 � 106/ml and labeled with 5 �Ci/ml myo-[3H]inositol in serum- and inositol-free RPMI 1640 mediumas described (44, 48). Lymphoblasts were stimulated, andwater-soluble IP were fractionated by anion-exchange chro-matography on AG 1-X8 formate resin (Bio-Rad, Munich,Germany) as detailed previously (44).
GTP�S binding assay. Agonist-induced activation of Gproteins was determined by measurement of [35S]GTP�Sbinding to digitonin-permeabilized lymphoblasts as de-scribed (44, 48, 56). Single measurements were conducted on1 � 106 lymphoblasts for 10 min at 30°C. Nonspecific bindingwas defined as the fraction of bound [35S]GTP�S not com-
C180 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
peted for by 10 �M unlabeled GTP�S. Measurements werecarried out in triplicate.
Cloning, sequencing, and expression of G� subunits andSSTR2 in COS-7 cells. The cDNAs encompassing the codingsequences of the human G�q, G�11, G�16, and SSTR2 werecloned by RT-PCR from B lymphoblast RNA using the oligo-nucleotide primers 5�-CCACCGCACCATGGCCCGCTCG-3�and 5�-TGGGGCCTGGGTCACAGCAGG-3 for the amplifica-tion of G�16 (accession no. NM_002068), 5�-GGAAGAAT-GACTCTGGAGTCCATCATGGCG-3� and 5�-CAGGCACA-ATTAGACCAGATTGTACTCC-3� for G�q (accession no.NM_002072), 5�- GGCCGGGACGATGACTCTGG-3� and 5�-GCGAAGTGTACGGAGGGAGAGATG-3� for G�11 (accessionno. AF011497), 5�-GTCTTTTCTTTCCACACCCCTGTG-3�and 5�-GAAGCACTTGCAAATAAAACAAGG AG-3� for G�14
(accession no. XM_005478), and the oligonucleotides 5�-AAAGCAGCCATGGACATGGCGG-3� and 5�-CCCCAAGCA-GTTCAGATACTGG-3� for the amplification of SSTR2 (acces-sion no. M81830), respectively. Reverse transcription wasperformed as described above. For PCR amplification, Pfu-Taq (Promega) was used according to the manufacturer’sinstructions. PCR products were cloned into pGEM-T easyvector (Promega), sequenced, and further subcloned into themammalian expression vector pcDNA3.1� (Invitrogen). Hu-man brain RNA was used for the amplification of G�14. ThePLC�2 expression vector was a kind gift of Dr. M. Simon(Pasadena, CA).
COS-7 cells were maintained in Dulbecco’s modified Ea-gle’s medium (DMEM) supplemented with 100 U/ml penicil-lin, 100 �g/ml streptomycin, and 10% FCS at 37°C. Fortransient transfection, 3 � 104 COS-7 cells were platedonto each well of a 24-well dish 1 day before the transfection.Cells were transfected using the transfection reagent Effect-ene (Qiagen, Hilden, Germany) according to the manufactur-er’s instructions. For each well, a total of 0.2 �g of DNA wasmixed with 1.6 �l of the enhancer reagent and 5 �l of thetransfection reagent in 60 �l of transfection buffer (all sup-plied by the manufacturer). Each component in the cotrans-fection experiment was represented by 0.06 �g of DNA, andthe total amount of DNA was supplemented to 0.2 �g/wellwith empty pcDNA3.1� vector DNA if necessary. This proce-dure resulted in a transfection efficacy of 70%, as estimatedby cotransfection assays using an enhanced green fluores-cence protein vector. After transfection (24 h), cell culturemedium was replaced with inositol-free DMEM supple-mented with 7 �Ci/ml myo-[3H]inositol and the cells wereincubated for another 24 h. The label medium was replacedwith inositol-free DMEM containing 20 mM LiCl, and cellswere stimulated by addition of 100 nM SST for 1 h. Cell lysisand separation of total IP were performed as described (32).
Measurement of MAP kinase activity. B lymphoblasts(1 � 107) were incubated in the presence or absence of SST(100 nM) for 2 min at 37°C, rapidly spun down, and lysedexactly as described (44). MAP kinase activity was deter-mined by immunoblotting with an antibody that recognizesthe phosphorylated, and thereby activated, MAP kinase iso-forms p44/42, i.e., ERK1/ERK2, as described (44). Aliquots ofthese lysates were analyzed in a second control Western blotfor equal expression and handling of ERK1/ERK2 proteinsduring preparation with an anti-ERK1/ERK2 antiserum.
Determination of cell proliferation, thymidine incorpora-tion, and immunoglobulin synthesis. Cells were seeded at aninitial density of 2 � 105/ml in serum-free RPMI 1640 me-dium, stimulated with SST, and propagated for 4 days (44).Cells were counted daily using a CASY cell analyzer system(Scharfe, Reutlingen, Germany). For determination of DNAsynthesis, B lymphoblasts were subcultured in 24-well dishes
under identical experimental conditions as described abovefor analysis of cell proliferation. They were propagated fortwo days and prepulsed with 1 �Ci of [3H]methylthymidinefor 18 h. The incorporation of radioactivity was determinedexactly as described (44). Supernatants of SST-stimulatedand control cells from the proliferation assay described abovewere harvested, and immunoglobulin concentrations werequantified by ELISA as described (44).
Statistical analysis and presentation of data. Data wereanalyzed using two-tailed Student’s t-tests and regardedsignificantly different at P � 0.05. If not indicated otherwise,all experiments were performed in triplicate using at leasttwo different cell lines. Data represent means � SE if notindicated otherwise.
RESULTS
Expression of SSTR type 2A transcripts in B lympho-blasts. In the first series of experiments, we examinedthe expression of the various SSTR isoforms in humanB lymphoblasts by RT-PCR using established oligonu-cleotide primers (22), which do not flank intron se-quences. By using human genomic DNA as control, allfive SSTRs were amplified (Fig. 1A), and the specificityof the PCR fragments was confirmed by restrictionanalysis (not shown). However, in RT-PCR studies, weonly detected mRNA transcripts encoding for the iso-form SSTR2 (Fig. 1A). To exclude fortuitous amplifica-tion of potentially contaminating genomic DNA in RT-PCRs, we included controls of the RNA without reversetranscription. Furthermore, all RNA specimens weretreated with RNase-free DNase before reverse tran-scription. No PCR amplicons were detected in thesesamples.
SSTR2-specific transcripts were identified in allRNA samples from six different B lymphoblast celllines (Fig. 2B). However, when we examined RNA fromquiescent peripheral blood lymphocytes of differentindividuals, a faint SSTR2-specific band was only ob-served in one of five samples (Fig. 2B). This observa-tion confirms previous reports (52) and suggests thatSSTR2 expression increases on lymphoblastic differen-tiation (52, 53).
For SSTR2, two splice variants have been identified(38), with SSTR2A being the wild-type receptor. InSSTR2B, a cryptic intron of 341 bp, located at the3�-end of the open reading frame, is removed by alter-native splicing. Thus the COOH-terminal sequence ofSSTR2B varies from SSTR2A (38). To distinguish be-tween these variants, we performed RT-PCR studieswith oligonucleotide primers that flank the crypticintron. In RNA samples from B lymphoblasts and pe-ripheral blood lymphocytes, we detected only long tran-scripts encoding SSTR2A (Fig. 2C). Thus we concludethat SSTR2A is the only known SSTR present onhuman B lymphoblasts.
Effects of SST on [Ca2�]i, IP formation, and bindingof GTP�S. Having demonstrated the expression ofSSTR2A-specific transcripts in B lymphoblasts, we ex-amined the effects of agonist stimulation on differenteffector systems. In the presence of 1 mM extracellularCa2�, basal [Ca2�]i amounted to 111 � 13 nM (n 31).On addition of 100 nM SST, [Ca2�]i increased by 191 �
C181EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
26 nM above baseline values (Fig. 2A). SST-evokedCa2� signals consisted of an initial peak followed by asustained plateau (Fig. 3A). When extracellular Ca2�
was chelated by addition of EGTA (5 mM) before stim-ulation with the agonist, SST-induced Ca2� increaseswere markedly reduced and amounted to 45 � 12 nMabove basal (Fig. 3B). Thus SST evoked both Ca2�
influx and mobilization from intracellular stores. Theincreases in [Ca2�]i were concentration dependent inthe range of 10 10 to 10 6 M SST (Fig. 3C), and theEC50 for SST-induced Ca2� signals was 1.3 � 0.9 nM.Maximum changes in [Ca2�]i were observed at 10 7 MSST. Because SSTRs couple to PTX-sensitive Gi and Goproteins, we examined the effects of PTX (50 ng/ml;16 h) on SST-induced [Ca2�]i transients. PTX treat-ment resulted in a distinct inhibition of SST-stimu-lated Ca2� signals by 71 � 12%. (Figs. 2A and 3A),which indicates that transmembrane signaling ofSSTR2A in human B lymphoblasts involves PTX-sen-sitive G proteins.
Ca2� mobilization from intracellular stores is fre-quently caused by the generation of inositol 1,4,5-trisphosphate (IP3). Hence, we analyzed the effect ofSST on IP levels in B lymphoblasts. Addition of 100 nMSST to B lymphoblasts resulted in an increased forma-tion of IP3 (Fig. 2B), as well as inositol 4-monophos-phate plus inositol 4,5-bisphosphate (IP2 � IP1). At 1min after stimulation, IP3 and (IP2 � IP1) levelsamounted to 162 � 16% (Fig. 2B) and 133 � 10% (notshown), respectively. Treatment of B lymphoblastswith 50 ng/ml PTX for 16 h resulted in a strong inhi-bition of SST-evoked IP formation, and markedly re-duced increases in IP3 (Fig. 2B) and (IP1 � IP2) levels(not shown) were observed.
To further demonstrate the involvement of G pro-teins in SST actions, we analyzed the SST-inducedbinding of the stable GTP analog [35S]GTP�S to per-meabilized lymphoblasts. SST (100 nM) increased[35S]GTP�S binding by 42 � 12%, and this effect wascompletely blunted in cells pretreated with PTX (Fig.
Fig. 1. Expression of somatostatin(SST) receptor (SSTR) isoforms in hu-man B lymphoblasts. A: transcriptionof specific mRNA for SST receptor iso-forms 1–5 was examined by RT-PCR.Lanes 1-5 show positive controls for theSSTR1–SSTR5 obtained with genomicDNA (left). Only SSTR2-specific tran-scripts were detected in RNA fromhuman B lymphoblasts by RT-PCR(right). The sizes of the HaeIII-di-gested marker FX174 are indicated atleft in base pairs (bp). The predictedsizes for the specific PCR products areSSTR1, 233 bp; SSTR2, 284 bp;SSTR3, 222 bp; SSTR4, 321 bp; andSSTR5, 223 bp. Ethidium bromide-stained 1.5% agarose gel. B: occurrenceof SSTR2-specific transcripts in RNApreparations from 6 different B lym-phoblasts (LB1–LB6) and from 5 dif-ferent peripheral blood lymphocytepreparations (LY1–LY5). Arrow indi-cates position of SSTR2-specific ampli-cons (284 bp). Numbers at left referto marker positions of AluI-digestedpBR322 DNA. C: discrimination be-tween alternatively spliced SSTR2-specific transcripts. PCR ampliconsshown left were generated with oligo-nucleotide primers used for the exper-iment in A, which do not differentiatebetween splice variants SSTR2A andSSTR2B (open arrow). The oligonucle-otide primers used at right encompassthe sequence fragment of SSTR2,which is deleted in SSTR2B by alter-native splicing. Predicted PCR ampli-cons for SSTR2A and SSTR2B are 686and 345 bp, respectively (indicated bybold arrows). Numbers at left referto marker positions of AluI-digestedpBR322 DNA. N, negative control; RNA,use of RNA which was not reverse tran-scribed; DNA, genomic DNA used forpositive controls. In LB2 and LY3, RNApreparations are the same as in B.
C182 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
4). Again, these experiments suggest that SSTR2Arelays signals via PTX-sensitive G proteins. However,it is important to note that the complete inhibition ofSST-induced [35S]GTP�S binding by PTX does not pre-clude an activation of PTX-insensitive G proteins bySST (56). PTX-sensitive G proteins are the mostabundant class of G� subunits. Thus binding of[35S]GTP�S to other G� proteins may escape detec-tion in this assay (56) and most likely explains theonly partial PTX-sensitivity of the SST-induced Ca2�
signals described above.Expression of transcripts encoding G� subunits of the
G�q family and reconstitution of SSTR2A-mediated IPgeneration. Our data presented so far suggest that bothPTX-sensitive and PTX-insensitive G proteins are in-volved in SST-stimulated IP generation, which is inaccordance with results of other groups (2, 8, 47). ThePTX-sensitive fraction of SST-stimulated IP formationis attributable most likely to the well-known activationof G�i proteins by SSTRs (23, 34). This results in theliberation of G�� subunits, which in turn activatephospholipase C � (PLC�) isoforms (40). However, be-cause G�i proteins are expressed ubiquitously, addi-tion of SST should result in IP generation in all cells
expressing SSTR2A, which is obviously not the case.Therefore, we wondered which components of the sig-naling cascade in lymphoblasts contribute to their pe-culiar behavior. Interestingly, the expression patternsof G�16 and PLC�2, two proteins involved in IP gener-ation, are restricted to lymphohematopoietic cells (3,
Fig. 2. Effect of SST on free calcium concentration ([Ca2�]i) andformation of inositol phosphates (IP) in human B lymphoblasts andeffect of pertussis toxin (PTX). A: B lymphoblasts were stimulatedwith SST (100 nM) and maximum [Ca2�]i were determined with thefluorescent dye fura 2 (filled bars). Control cells remained nonstimu-lated (hatched bars). At right, lymphoblasts were pretreated withPTX (50 ng/ml; 16 h). Data are means � SE of n 31 (for PTX, n 19) measurements on 4 different cell lines. B: B lymphoblasts werelabeled with [3H]-myo-inositol for 24 h. Inositol trisphosphate (IP3)formation was determined 1 min after stimulation with SST (100nM; filled bars). Control cells remained nonstimulated (hatchedbars). At right, cells were pretreated with PTX (50 ng/ml; 16 h).Results are means � SE from 1 representative experiment of a totalof 4 independent experiments conducted on a different cell line each.*Significant difference from control cells at P � 0.05.
Fig. 3. Ca2� signals in SST-stimulated human B lymphoblasts weremeasured with the calcium-sensitive dye fura 2. A: control B lym-phoblasts (left) and B lymphoblasts pretreated with PTX (50 ng/ml,16 h; right) were stimulated with 100 nM SST in the presence of 1mM extracellular Ca2�. B: extracellular Ca2� was chelated by addi-tion of EGTA (5 mM) before stimulation with 100 nM SST. Removalof extracellular Ca2� resulted in reduced basal as well as peak[Ca2�]i on stimulation with SST. Representative traces from exper-iments conducted on 4 different lymphoblast cell lines with measure-ments performed in triplicates each. C: fura 2-loaded B lymphoblastswere stimulated with SST at the indicated concentrations in thepresence of 1 mM extracellular Ca2� and the ensuing peak increasesin [Ca2�]i were measured. Results are from experiments performedon 3 different cell lines. Data points represent means � SE and aregiven as a percentage of maximum [Ca2�]i increase.
C183EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
40). Studies with other G protein-coupled receptorsexpressed in blood cells suggested that a basal activa-tion of a G�q family protein, most likely G�16, is re-quired for activation of PLC�2, which is then furtherpotentiated by free G�� subunits (4). Furthermore,G�16 is able to couple nonspecifically to a wide varietyof different heptahelical receptors (32). By RT-PCR, wedetected G�16-specific transcripts together with tran-scripts encoding the widely expressed G�q family mem-bers G�q and G�11 in human B lymphoblasts (Fig. 5,A–C). The identities of these transcripts were verifiedby cloning and sequencing. Transcripts of the fourthmember of this G�q family, G�14, were not detectablein B lymphoblasts but were present in RNA fromhuman brain (Fig. 5D).
To test the hypothesis that PLC�2 and G�16 contrib-ute to SST-mediated IP generation in B lymphoblasts,we reconstituted these components transiently inCOS-7 cells. Native COS-7 cells express neither SSTR2nor G�16, as revealed by RT-PCR (data not shown). Inaccord, addition of SST (100 nM) to nontransfectedCOS-7 cells did not result in an increase in IPs (Fig.6A). Sole expression of SSTR2A resulted in a signifi-cant but modest stimulation of IP formation on addi-tion of SST to 130 � 4.5% (P � 0.05; n 3). However,expression of SSTR2A together with G�16 led to dis-tinct SST-stimulated increases in IP formation to193 � 12%. Coexpression of PLC�2 together withSSTR2A resulted in higher basal IP levels but did notstimulate IP generation (135 � 7.4%) above the valuesobserved in cells expressing SSTR2A only (Fig. 6A).Maximum SST-stimulated increases in IP formationwere observed in COS-7 cells that expressed SSTR2Atogether with G�16 and PLC�2 (234 � 2% of nonstimu-lated controls; P � 0.05; n 3). These results suggestthat G�16 and PLC�2 are efficient mediators of SST-stimulated IP formation. In a final series of experi-ments, we tested whether other members of the G�qfamily can substitute for G�16 in this pathway. Thewidely expressed G� subunits G�q and G�11 werecloned and sequenced from human B lymphoblasts.G�14, which was not detectable in B lymphoblasts, wascloned from human brain. Again, expression of G�16together with PLC�2 mediated marked SST-stimu-lated increases in IP levels (290.3 � 20.3% of non-stimulated controls; n 3 independent experiments;P � 0.01, Fig. 6B). However, in the presence of G�q andG�11, SST stimulation resulted in only modest in-creases in IP levels (129.2 � 0.6% and 138.5 � 2.4% for
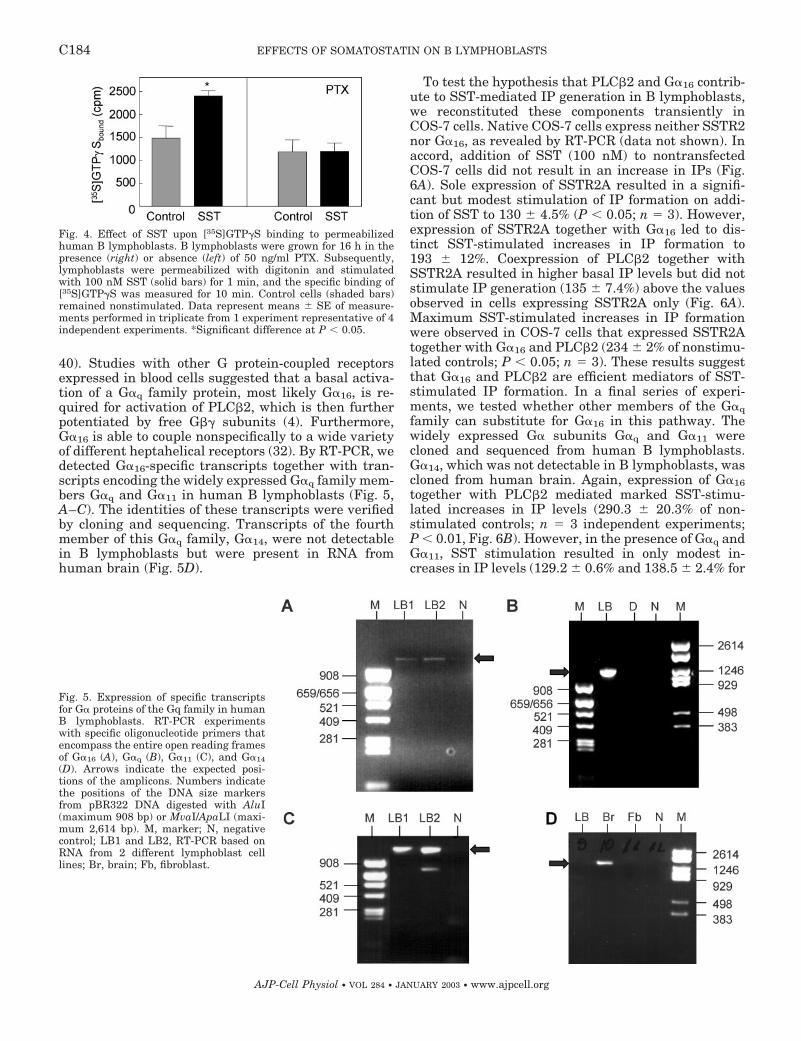
Fig. 4. Effect of SST upon [35S]GTP�S binding to permeabilizedhuman B lymphoblasts. B lymphoblasts were grown for 16 h in thepresence (right) or absence (left) of 50 ng/ml PTX. Subsequently,lymphoblasts were permeabilized with digitonin and stimulatedwith 100 nM SST (solid bars) for 1 min, and the specific binding of[35S]GTP�S was measured for 10 min. Control cells (shaded bars)remained nonstimulated. Data represent means � SE of measure-ments performed in triplicate from 1 experiment representative of 4independent experiments. *Significant difference at P � 0.05.
Fig. 5. Expression of specific transcriptsfor G� proteins of the Gq family in humanB lymphoblasts. RT-PCR experimentswith specific oligonucleotide primers thatencompass the entire open reading framesof G�16 (A), G�q (B), G�11 (C), and G�14
(D). Arrows indicate the expected posi-tions of the amplicons. Numbers indicatethe positions of the DNA size markersfrom pBR322 DNA digested with AluI(maximum 908 bp) or MvaI/ApaLI (maxi-mum 2,614 bp). M, marker; N, negativecontrol; LB1 and LB2, RT-PCR based onRNA from 2 different lymphoblast celllines; Br, brain; Fb, fibroblast.
C184 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
G�q and G�11, respectively). Interestingly, SST-medi-ated PLC�2 stimulation was also very efficient in thepresence of G�14 (478.0 � 24.2%; n 3, Fig. 6B), anobservation without obvious relevance for B lympho-blasts. Taken together, these results suggest, but donot prove, that G�16 and PLC�2 are responsible for theability of SSTR2A to stimulate increases in IP levels inB lymphoblasts, a property not observed in other celltypes.
MAP kinase activation, DNA synthesis, cell prolifer-ation, and synthesis of immunoglobulins. The experi-ments presented so far provided evidence for stimula-tory actions of SST in B lymphoblasts, as shown for IPformation and G protein activation. Next, we studiedwhether SST affects typical cellular functions of Blymphoblasts, i.e., proliferation and immunoglobulinsynthesis. Furthermore, we investigated whether SSTstimulates additional signaling pathways in these cells
and examined the effects of SST on MAP kinase acti-vation. Available evidence suggests that MAP kinaseactivation by heptahelical receptors that couple toPTX-sensitive G proteins involves a complex array ofadaptor proteins, GTPases, and kinases (for review,see Ref. 45). Liberated G�� subunits from activatedPTX-sensitive G proteins are thought to initiate thispathway (45). As shown in Fig. 7, SST (100 nM; 2 min)induced an increase in phospho-ERK1/2, which pre-dominantly affected p42ERK2. This activation wasstronger than that induced by PAF (100 nM; 2 min),another well-characterized B lymphocyte activator(28). SST-induced MAP kinase activation varied be-tween three- and sevenfold compared with the non-stimulated controls, as determined by image analysis.Treatment of B lymphoblasts with PTX (50 ng/ml; 16 h)completely blocked SST-induced MAP kinase activa-tion (not shown).
For the analysis of B lymphoblast proliferation, cellswere seeded at a density of 2 � 105 cells/ml inserum-free medium. After 24–36 h, we observed amarked increase in cell number of SST-stimulated cellsabove that of nonstimulated controls (Fig. 8A). Thisdifference reached its maximum on day 4, and cellnumbers of SST-stimulated cells were 4.4 � 1.5-foldhigher (n 6 independent experiments with 3 differ-ent cell lines) compared with nonstimulated controls.There was a considerable variation in this growth-promoting effect, ranging from 1.2-fold to 11.4-foldincreases depending on cell line and experiment. Inter-estingly, PTX inhibited the SST-stimulated B lympho-blast proliferation only modestly by 22 � 5% ( n 4;not shown).
In accordance with the growth-promoting action ofSST, [3H]thymidine incorporation into SST-stimulatedB lymphoblasts was 3.5 � 1.2-fold higher (n 6 inde-pendent experiments with 3 different cell lines) thanthat of nonstimulated controls (Fig. 8B).
In parallel to these proliferation assays, superna-tants were harvested and used for the quantification ofimmunoglobulin concentrations by ELISA. Immuno-globulin formation increased significantly 4.8 � 1.2-fold (n 6 independent experiments with 3 differentcell lines) above nonstimulated controls. Again, weobserved a considerable variation in the range of 2.4- to9.2-fold depending on cell line and experiment.
DISCUSSION
SST is a widely expressed neuropeptide that affectsnumerous organ systems and cell types in a prevail-ingly inhibitory fashion. Increasing evidence, predom-inantly from binding, nuclear imaging, and expressionstudies, suggests that SST also plays a role in theimmune system (53, 54). On the basis of observationsthat SST can evoke Ca2� signals in human B lympho-blasts (48), we investigated this signaling pathway inmore detail. Specifically, we tried to unravel whichcomponents of the B lymphoblast signaling networkare responsible for this unusual effector activation bySST. A second and independent focus of this work was
Fig. 6. SST-mediated IP formation in the presence of G� subunits ofthe G�q family and PLC�2. A: COS-7 cells were transiently trans-fected with expression vectors for the indicated proteins and forma-tion of total IPs was analyzed on stimulation with SST (100 nM).Shown are means � SE of measurements performed in quadrupli-cates from 1 experiment representative of 3 independent experi-ments. B: COS-7 cells were transiently transfected with vectorsencoding SSTR2, PLC�2, and the indicated G� protein. The forma-tion of total IPs on stimulation with 100 nM SST was measured as inA. Data represent means � SE from 3 independent experiments withmeasurements performed as triplicates. *Significant difference atP � 0.05. **Significant difference at P � 0.01.
C185EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
to elucidate the significance of SST for B lymphoblastfunction, and we examined two typical B cell re-sponses, proliferation and immunoglobulin formation.Our results suggest that SST acts as a B cell growthfactor. Both issues, early effector activation and cellproliferation, deserve a more detailed discussion. Fi-nally, we have to address the question of potentialphysiological roles for SST in B cell regulation, an issuewhich will remain mostly speculative.
SSTR expression and early signaling. The only SSTRisoform we detected in B lymphoblasts is SSTR2A,which is the predominant SSTR in nonneural tissues.Although this notion is drawn from RT-PCR studies, itis in accord with other expression, binding, and imag-ing studies (52, 53). SSTR2A is a typical heptahelicalreceptor that couples to heterotrimeric G proteins. Thisis also true for B lymphoblasts, as shown here by anincreased GTP�S binding on SST stimulation. Directbiochemical interaction studies provided evidence forcoupling of G�i3 and G�o to SSTR2 (24). Because Blymphoblasts lack G�o (44), which is predominantlyexpressed in neural tissues, SSTR2 most likely couplesto G�i subunits in this cell type. This notion is furthercorroborated by the inhibitory effects of PTX on SST-stimulated GTP�S binding, IP formation, increases in[Ca2�]i, and MAP kinase activation. Of note, theseeffects of PTX varied considerably between differenteffector systems. Whereas SST-stimulated MAP ki-nase activation and GTP�S binding were totallyblocked and Ca2� signals were inhibited by 70%, PTXdiminished SST-induced lymphoblast proliferation byonly 20%. Because PTX-sensitive G proteins are mostabundantly expressed in mammalian cells, and be-cause G�i proteins have a considerable rate of sponta-neous GDP/GTP exchanges, a concomitant activation
Fig. 7. Effect of SST on activation ofmitogen-activated (MAP) kinase. Blymphoblasts were stimulated for 2min with 100 nM SST, 100 nM plate-let-activating factor (PAF), or for 15min with 250 nM of the phorbol esterTPA or remained nonstimulated (con-trol). A: equal amounts of cell lysateswere analyzed by immunoblotting, andactivated MAP kinases were detectedwith a phosphospecific ERK1/ERK2 ki-nase antibody that recognizes p42 andp44 isoforms. B: control Western blotswith aliquots from lysates used in A.Immunostaining was performed withan antibody that recognizes both ERKisoforms. Arrows indicate positions ofmolecular weight standards. Experi-ments are representative of 3 others.
Fig. 8. Effect of SST on B lymphoblast proliferation, DNA synthesis,and immunoglobulin formation. A: B cells were seeded at 2–3 � 105
cells/ml on day 0. On day 1, either SST (F) or buffer (E) was added(arrow). Aliquots were counted daily. Data represent means � SDfrom 1 representative experiment, replicated with 4 different celllines. B: lymphoblasts were seeded on 24-well culture dishes asdescribed for A. On day 2, cells were prepulsed with 2 �Ci of[3H]methylthymidine for 16 h. Values are means � SE from 1 repre-sentative experiment with measurements performed in triplicate. C:supernatants from experiments in A were harvested on day 4 and IgMlevels were quantified. Values are means � SE from 1 representativeexperiment with measurements performed in triplicate. *Significantdifference from controls at P � 0.05.
C186 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
of PTX-insensitive G proteins most likely escapes grossbinding studies as performed here by GTP�S bindingto whole cells, a method which has even been optimizedfor the analysis of PTX-sensitive G proteins (56). Thusour results from GTP�S binding analyses do not ex-clude a simultaneous activation of, for example, G�qclass G proteins, which is in accord with the partialPTX-insensitivity of SST-stimulated Ca2� signals ob-served here. At this point, we have to consider furtherthat PTX is a highly sensitive but not an entirelyspecific tool for the analysis of G�i-dependent signalingpathways, especially if immune cells are concerned.There is evidence that long-term treatment with PTXstimulates proliferation of T and B lymphocytes (20),an effect that has been attributed to the B oligomer ofthe toxin and does not involve ADP ribosylation (20).Although we have verified in previous studies that ourPTX treatment conditions result in a complete modifi-cation of G�i proteins in B lymphoblasts (48), we alsoobserved that long-term treatment with the toxin in-consistently evoked minor increases in cell number(44), which could also explain the limited effect of PTXon SST-stimulated B lymphoblast proliferation shownhere.
SSTR2A-mediated Ca2� signals in B lymphoblasts.Although there is evidence from one tumor cell linethat SSTR2 can activate Ca2� channels (50), in mostcell types SSTR2 stimulation is associated with de-creased Ca2� signals caused by an inhibition of Ca2�
channels or an activation of K� channels (25, 33, 34,59). The original tracings shown in Fig. 3 suggest thecontribution of both Ca2� influx and Ca2� liberationfrom intracellular stores to the generation of Ca2�
signals in SST-stimulated B cells. Concomitantly, weobserved an activation of PLC by SST. To our knowl-edge, the human B lymphoblast is the only nontrans-fected cell type for which a direct activation of PLC bySSTR2 stimulation has been observed so far. Hence, wewondered which mechanisms confined to B lympho-blasts contribute to this unusual PLC activation. Lym-phohematopoietic cells specifically express PLC�2 andG�16, a G�q class G protein (3, 40). G�16 can couplepromiscuously to a wide variety of heptahelical recep-tors, including the 5-HT1A receptor, the formyl peptidereceptor, the �-opiod receptor, the �2-adrenergic recep-tor, the M2-muscarinic acetylcholine receptor, theLTB4 receptor, and the P2U purinoreceptor (4, 14, 32).Baltensperger and Porzig (4) provided evidence thatheptahelical receptors that couple to PTX-sensitive Gproteins in lymphohematopoietic cells require a basalactivation of a G�q class G protein, e.g., G�16, forefficient activation of PLC. Free G�� subunits arisingfrom concomitant G�i activation then further potenti-ate PLC activity in a PTX-sensitive manner (4). How-ever, sole free G�� dimers were unable to mediate PLCactivation in the absence of activatable G�16 in thissystem (4). Interestingly, G�16 activation has also beenrelated to cell growth stimulation of hematopoieticcells by heptahelical receptors (15).
To test whether the above described dual activationmodel explains SSTR2A-mediated PLC activation in
lymphoblasts, we conducted a series of reconstitutionexperiments. RT-PCR studies indicated that B lympho-blasts express transcripts for the G�q class proteinsG�16, G�q and G�11 but not G�14 (Fig. 5). Transfectionof SSTR2A into COS-7 cells, which lack endogenousSSTRs, resulted in a modest increase in IP levels onstimulation with SST. This 30% increase in IP levelsis in perfect accordance with the small effects observedon expression and stimulation of SSTR2 in COS-7 cells(2, 51), CCL39 cells (47), and F4C1 pituitary cells (8).However, coexpression of SSTR2A together with G�16and PLC�2 resulted in pronounced increases in IPlevels on stimulation with SST. A second series ofexperiments revealed that the widely expressed G�qclass proteins G�11 and G�q cannot efficiently substi-tute for G�16 to mediate SST-stimulated PLC�2 acti-vation. Interestingly, G�14, a G�q class protein withrestricted expression (e.g., kidney and brain; 57)shared the ability of G�16 to relay SSTR2A activationto PLC�2 stimulation. This feature of G�14 deservesfurther investigation but does not contribute to the Blymphoblast signal transduction discussed here.
Taken together, the data presented here support, butdo not ultimately prove, a scenario that the dual acti-vation of G�16 and G�i proteins by SST in B lympho-blasts causes PLC�2 activation and Ca2� transients.Such a dual activation mechanism for PLC stimulationappears to occur frequently in B lymphoblasts, as canbe inferred from the only partial PTX sensitivities ofLPA and PAF in other studies (44, 48).
In contrast to SSTR2A, SSTR3 can activate PLC�isoforms in nonlymphohematopoietic cells, as has beenshown for PLC�3 activation in intestinal smooth mus-cle cells (29), which indicates subtype-specific effectoractivation.
Somatostatin-induced B lymphoblast proliferation.The second novel observation of this report is an aug-mented B lymphoblast proliferation on stimulationwith SST, which was further accompanied by an in-crease in immunoglobulin secretion.
In most cell types, mainly antiproliferative effects ofSST have been observed (6, 33, 39). For the immunesystem, early studies reported growth-inhibiting ef-fects of SST in activated T lymphocytes (27, 37). In Blymphocytes, biphasic effects of SST have been de-scribed: SST at 10 9 to 10 8 M exhibited a modestinhibitory effect on lymphocyte DNA synthesis; how-ever, at 10 7 M SST-induced a twofold increase in DNAsynthesis (36). Other reports demonstrate that SSTpotentiates the proliferation of mitogen-activated Jur-kat and peripheral blood mononuclear cells (7, 21). Inthe case of Jurkat cells, this is caused by an increasedIL-2 secretion mediated by SSTR3 activation (7).
In our in vitro B cell model, SST stimulates prolifer-ation and immunoglobulin synthesis and may be re-garded as a B cell growth factor. The observed effectsresemble those of PAF and LPA, both in magnitudeand time course of the signals (38, 44, 48). Further-more, our data and results from others indicate thatthe expression of SSTR2 on lymphocytes is develop-mentally regulated (52, 53). Quiescent peripheral lym-
C187EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
phocytes lack SSTR2 or express transcripts only at lowlevels, whereas all B lymphoblast lines investigatedhere expressed these mRNA transcripts. A recent re-port describes a similar developmental regulation inthymocytes and provides evidence that SSTR2 activa-tion is involved in thymic maturation and thymocyteproliferation (49). This study also examined the effectsof SST on splenocyte proliferation. Whereas SST in-creased the proliferation of total splenocytes, it inhib-ited growth of purified splenic T cells and T lympho-cytes (49). Thus effects of SST on different lymphocytepopulations depend on the cellular context and may bemediated by the secretion of additional auto- and para-crine factors.
At present, we do not know the exact sequence ofevents coupling SSTR2A activation in B lymphoblaststo enhanced proliferation and immunoglobulin secre-tion. Current evidence suggests that sole activation ofPLC�2 is not sufficient for stimulation of proliferation(30), and additional effector systems are probable can-didates for this effect. Free G�� subunits, liberatedfrom activated G�i proteins, are frequently involved inthe initiation of growth signals, e.g., by activating theMAP kinase cascade (45). For lymphohematopoieticcells, evidence exists that G�16 also contributes togrowth regulation (15). Hence, inactivation of G�16 inerythroleukemia cells is accompanied by a decreasedproliferation rate (15). Both signaling pathways mayalso contribute to SSTR2A-induced B cell proliferation.
Given the biphasic and context-dependent effects ofSST on lymphocyte proliferation in the literature (7,27, 36, 37, 49), it is also conceivable that SST stimu-lates the secretion of autocrine growth factors in Blymphoblasts, which in turn stimulate lymphoblastproliferation. For T lymphocytes, an increased IL-2secretion on SSTR3 activation has been reported,which subsequently stimulates T cell proliferation (7).Furthermore, a SST-triggered release of autocrine Bcell growth factors could alternatively explain the dif-ferent inhibitory potencies of PTX on SST-stimulated“early signals” and proliferation observed here. Futureexperiments are needed to elucidate the mutual con-tribution of these signaling pathways for SST-stimu-lated B cell proliferation.
Somatostatin and the immune system. ImmortalizedB lymphoblasts are a frequently employed model sys-tem for differentiated antibody-secreting B cells (28,44). However, some caution should be taken whenextrapolating data derived from such cell lines. Basedprimarily on these in vitro studies, it is only possible tospeculate on the precise physiological roles of SST for Bcells in the immune system.
SSTRs are widely expressed in lymphoid tissues (31,42, 43), and both peptidergic nerves and autocrine SSTsecretion are potential sources for SST in the immunesystem (1, 12, 13, 49). High levels of SST binding sitesare found in germinal centers of Peyer’s patches, ton-sils, appendix, colonic lymphoid tissue, and on thymo-cytes (41, 49). In quiescent blood lymphocytes, mRNAencoding for SSTRs is found at low levels only (52). On
lymphoblastic differentiation and proliferation, begin-ning at early differentiation stages, B and T cells startto express SSTRs, with SSTR2 being the predominantSSTR isoform (52, 53), which is also confirmed in ourstudy for B lymphoblasts. SSTR2 transcripts are alsofound in plasma cells (53). Messenger RNAs for SSTR3,SSTR4, and SSTR5 have been detected in a few lym-phocyte cell lines, whereas SSTR1 mRNA was unde-tectable (7, 52, 53). There is evidence for autocrine SSTsecretion in lymphoid tissues, including thymus andspleen (1, 12, 13, 49). Lipopolysaccharide, interferon-�(IFN-�), and IL-10 stimulate SST expression in thespleen (55). On the other hand, SST inhibits IFN-�release by granuloma cells, splenocytes, and T cells(11), which suggests complex immunoregulatory cir-cuits.
SST has been reported to stimulate IL-2 secretionand T cell proliferation in Jurkat cells via SSTR3 (7),which resembles the findings of this study. Conversely,other studies show antiproliferative effects of SST inthe immune system (27, 37). With respect to B cellphysiology, SST has been reported to be ineffective instimulating immunoglobulin secretion of plasma cells,whereas vasoactive intestinal peptide markedly in-creased immunoglobulin production (17, 18).
Emerging evidence suggests that these varying andcontradicting effects of SST in the immune systemdepend on the context and developmental stage of thecells investigated. Results from a recent report supportthat notion. Hence, SST stimulates fetal thymocytegrowth and maturation under organ culture conditions(49). Similarly, total splenocyte proliferation increasesin the presence of SST (49). However, repeating theseexperiments with purified thymocytes or splenic T lym-phocytes revealed an antiproliferative effect of SST(49).
B lymphocyte proliferation and immunoglobulin for-mation are regulated in complex network of antigenactivation and accessory stimuli, e.g., cytokines, adhe-sion molecules, hormones, and neurotransmitters (9,10, 12), that govern the fate of the respective B lym-phoblast population to proliferation, immunoglobulinsynthesis, or apoptosis. SST obviously contributes tothis network. With the availability of more isotype-specific SSTR agonists and antagonists, we now havethe tools to challenge the in vivo significance of these invitro results.
We thank Dr. K. H. Jakobs for valuable comments on the manu-script.
This study was supported by the Deutsche Forschungsgemein-schaft.
REFERENCES
1. Aguila MC, Dees WL, Haensly WE, and McCann SM. Evi-dence that somatostatin is localized and synthesized in lymphoidorgans. Proc Natl Acad Sci USA 88: 11485–11489, 1991.
2. Akbar M, Okajima F, Tomura H, Majid MA, Yamada Y,Seino S, and Kondo Y. Phospholipase C activation and Ca2�
mobilization by cloned human somatostatin receptor subtypes1–5 in transfected COS-7 cells. FEBS Lett 348: 192–196, 1994.
C188 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
3. Amatruda TTIII, Steele DA, Slepak VZ, and Simon MI.G�16, a G protein �-subunit specifically expressed in hematopoi-etic cells. Proc Natl Acad Sci USA 88: 5587–5591, 1991.
4. Baltensperger K and Porzig H. The P2U purinoreceptor oblig-atory engages the heterotrimeric G protein G16 to mobilize in-tracellular Ca2� in human erythroleukemia cells. J Biol Chem272: 10151–10159, 1997.
5. Bhatena SJ and Recant L. Somatostatin receptors on circu-lating human blood cells. Horm Metab Res 12: 277–278, 1980.
6. Buscail L, Delesque N, Esteve JP, Saint-Laurent N, PratsH, Clerc P, Robberecht P, Bell GI, Liebow C, and SchallyAV. Stimulation of tyrosine phosphatase and inhibition of cellproliferation by somatostatin analogues: mediation by humansomatostatin receptor subtypes SSTR1 and SSTR2. Proc NatlAcad Sci USA 91: 2315–2319, 1994.
7. Cardoso A, El Ghamrawy C, Gautron JP, Horvat B, Gau-tier N, Enjalbert A, and Krantic S. Somatostatin increasesmitogen-induced IL-2 secretion and proliferation of human Jur-kat T cells via sst3 receptor isotype. J Cell Biochem 68: 62–73,1998.
8. Chen L, Fitzpatrick VD, Vandlen RL, and Tashjian AH Jr.Both overlapping and distinct signaling pathways for somatosta-tin receptor subtypes SSTR1 and SSTR2 in pituitary cells. J BiolChem 272: 18666–18672, 1997.
9. Cushley W and Harnett MM. Cellular signaling mechanismsin B lymphocytes. Biochem J 292: 313–332, 1993.
10. Delves PJ and Roitt IV. The immune system. N Engl J Med343: 37–49 and 108–117, 2000.
11. Elliott DE, Li J, Blum AM, Metwali A, Patel YC, andWeinstock JV. SSTR2A is the dominant somatostatin receptorsubtype expressed by inflammatory cells, is widely expressedand directly regulates T cell IFN-� release. Eur J Immunol 29:2454–2463, 1999.
12. Felten DL, Felten SY, Carlson SL, Olschowka JA, andLivnat S. Noradrenergic and peptidergic innervation of lym-phoid tissue. J Immunol 135: 755–765, 1985.
13. Fuller PJ and Verity K. Somatostatin gene expression in thethymus. J Immunol 143: 1015–1017, 1989.
14. Gaudreau R, Le Gouill C, Metaoui S, Lemire S, StankovaJ, and Rola-Pleszczynski M. Signalling through the leukotri-ene B4 receptor involves both �i and �16, but not �q or �11G-protein subunits. Biochem J 335: 15–8, 1998.
15. Ghose S, Porzig H, and Baltensperger K. Induction of ery-throid differentiation by altered G�16 activity as detected by areporter gene assay in MB-02 cells. J Biol Chem 274: 12848–12854, 1999.
16. Hoyer D, Bell GI, Berelowitz M, Epelbaum J, Feniuk W,Humphrey PPA, O’Carroll AM, Patel YC, Schonbrunn A,Taylor JE, and Reisine T. Classification and nomenclature ofsomatostatin receptors. Trends Pharmacol Sci 16: 86–88, 1995.
17. Kimata H and Fujimoto M. Induction of IgA1 and IgA2 pro-duction in immature human fetal B cells and pre-B cells byvasoactive intestinal peptide. Blood 85: 2098–2104, 1995.
18. Kimata H, Yoshida A, Ishioka C, Fujimoto M, and FurushoK. Vasoactive intestinal peptide enhances immunoglobulin pro-duction and growth in human plasma cells via mechanisms thatmay involve protein kinase C. J Clin Endocrinol Metab 81:3024–3032, 1996.
19. Koch BD, Dorflinger LJ, and Schonbrunn A. Pertussis toxinblocks both cyclic AMP-mediated and cyclic AMP-independentactions of somatostatin. J Biol Chem 260: 13138–13145, 1985.
20. Kolb JP, Genot E, Petit-Kosaks E, Paul-Eugene N, andDugas B. Effect of bacterial toxinson human B cell activation. I.Mitogenic activity of pertussis toxin. Eur J Immunol 20: 969–976, 1990.
21. Krantic S, Enjalbert A, and Rabourdin-Combe C. Measlevirus modulates human T-cell somatostatin receptors and theircoupling to adenylyl cyclase. Virology 71: 7470–7477, 1997.
22. Kubota A, Yamada Y, Kagimoto S, Shimatsu A, ImamuraM, Tsuda K, Imura H, Seino S, and Seino S. Identification ofsomatostatin receptor subtypes and an implication for the effi-cacy of somatostatin analogue SMS 201–995 in treatment ofhuman endocrine tumors. J Clin Invest 93: 1321–1325, 1994.
23. Law SF, Woulfe D, and Reisine T. Somatostatin receptoractivation of cellular effector systems. Cell Signal 7: 1–8, 1995.
24. Law SF, Yasuda K, Bell GI, and Reisine T. G�i3 and G�o
selectively associate with the cloned somatostatin receptor sub-type SSTR2. J Biol Chem 268: 10721–10727, 1993.
25. Lewis DL, Weight FF, and Luini A. Guanine nucleotide-binding protein mediates the inhibition of voltage-dependentcalcium current by somatostatin in a pituitary cell line. ProcNatl Acad Sci USA 83: 9035–9039, 1986.
26. Madden KS, Sanders VM, and Felten DL. Catecholamineinfluences and sympathetic neural modulation of immune re-sponsiveness. Annu Rev Pharmacol Toxicol 35: 417–448, 1995.
27. Mascardo RN, Barton RW, and Sherline P. Somatostatinhas an antiproliferative effect on concanavalin-activated ratthymocytes. Clin Immunol Immunopathol 33: 131–138, 1984.
28. Mazer B, Clay KL, Renz H, and Gelfand EW. Platelet-activating factor enhances Ig production in B lymphoblastoid celllines. J Immunol 145: 2602–2607, 1990.
29. Murthy KS, Coy DH, and Makhlouf GM. Somatostatin recep-tor-mediated signaling in smooth muscle. Activation of phospho-lipase C-�3 by G�� and inhibition of adenylyl cyclase by G�i1 andG�o. J Biol Chem 271: 23458–23463, 1996.
30. Noh DY, Shin SH, and Rhee SG. Phosphoinositide-specificphospholipase C and mitogenic signaling. Biochim Biophys Acta1242: 99–113, 1995.
31. O’Brien JP. Somatostatin receptor-based imaging in malignantlymphomas. J Nucl Med 36: 19–20, 1995.
32. Offermanns S and Simon MI. G�15 and G�16 couple a widevariety of receptors to phospholipase C. J Biol Chem 270: 15175–15180, 1995.
34. Patel YC. Somatostatin and its receptor family. Front Neuroen-docrinol 20: 157–198, 1999.
35. Patel YC, Greenwood M, Kent G, Panetta R, and SrikantCB. Multiple gene transcripts of the somatostatin receptorSSTR2: tissue selective distribution and cAMP regulation. Bio-chem Biophys Res Commun 192: 288–294, 1993.
36. Pawlikowski M, Stepien H, Kunert-Radek J, ZelakowskiP, and Schally AV. Immunomodulatory action of somatostatin.Ann NY Acad Sci 496: 233–239, 1987.
37. Payan DG, Hess CA, and Goetzl EJ. Inhibition by somatosta-tin of the proliferation of T-lymphocytes and Molt-4 lympho-blasts. Cell Immunol 84: 433–438, 1984.
38. Petersenn S, Rasch AC, Presch S, Beil FU, and SchulteHM. Genomic structure and transcriptional regulation of thehuman somatostatin receptor 2. Mol Cell Endocrinol 157: 75–85,1999.
39. Reichlin S. Somatostatin (parts 1 and 2). N Engl J Med 309:1495–1501 and 1556–1561, 1983.
40. Rhee SG. Regulation of phosphoinositide-specific phospholipaseC. Annu Rev Biochem 70: 281–312, 2001.
41. Reubi JC, Horisberger U, Waser B, Gebbers JE, and Lais-sue J. Preferential location of somatostatin receptors in germi-nal centres of human gut lymphoid tissue. Gastroenterology 103:1207–1214, 1992.
42. Reubi JC, Waser B, and Hanhagan M. In vitro and in vivodetection of somatostatin receptors in human malignant lympho-mas. Int J Cancer 50: 895–900, 1992.
43. Reubi JC, Waser B, and Horisberger U. In vitro autoradio-graphic and in vivo scintigraphic localization of somatostatinreceptors in human lymphatic tissue. Blood 82: 2143–2148,1993.
44. Rosskopf D, Daelman W, Busch S, Schurks M, Hartung K,Kribben A, Michel MC, and Siffert W. Growth-factor likeaction of lysophosphatidic acid on human B lymphoblasts. Am JPhysiol Cell Physiol 274: C1573–C1582, 1998.
45. Schwindinger WF and Robishaw JD. Heterotrimeric G-pro-tein ��-dimers in growth and differentiation. Oncogene 20:1653–1660, 2001.
46. Selmer I, Schindler M, Allen JP, Humphrey PP, and Em-son PC. Advances in understanding neuronal somatostatin re-ceptors. Regul Pept 90: 1–18, 2000.
C189EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org
47. Siehler S and Hoyer D. Characterisation of human recombinantsomatostatin receptors. 4. Modulation of phospholipase C activity.Naunyn Schmiedebergs Arch Pharmacol 360: 522–532, 1999.
48. Siffert W, Rosskopf D, Moritz A, Wieland T, Kaldenberg-Stasch S, Kettler N, Hartung K, Beckmann S, and JakobsKH. Enhanced G protein activation in immortalized lympho-blasts from patients with essential hypertension. J Clin Invest96: 759–766, 1995.
49. Solomou K, Ritter MA, and Palmer PD. Somatostatin isexpressed in the murine thymus and enhances thymocyte devel-opment. Eur J Immunol 32: 1550–1559, 2002.
50. Taylor JE. Somatostatin (SSTR2) receptors mediate phospho-lipase C-independent Ca2� mobilization in rat AR42J pancreascells. Biochem Biophys Res Commun 214: 81–85, 1995.
51. Tomura H, Okajima F, Akbar M, Abdul-Majid M, Sho K,and Kondo Y. Transfected human somatostatin receptor type 2,SSTR2, not only inhibits adenylate cyclase but also stimulatesphospholipase C and Ca2� mobilization. Biochem Biophys ResCommun 200: 986–992, 1994.
52. Tsutsumi A, Takano H, Ichikawa K, Kobayashi S, andKoike T. Expression of somatostatin receptor subtype 2 mRNAin human lymphoid cells. Cell Immunol 181: 44–49, 1997.
53. Van Hagen PM, Hofland LJ, ten Bokum AMC, Lichtenaur-Kaligis EGR, Kwekkeboom DJ, Ferone D, and LambertsSWJ. Neuropeptides and their receptors in the immune system.Ann Med 31: Suppl. 2., 15–22, 1999.
54. Van Hagen PM, Krenning EP, Kwekkeboom DJ, Reubi JC,vd Anker-Lugtenburg PJ, Lowenberg B, and LambertsSWJ. Somatostatin and the immune and haematopoetic system:a review. Eur J Clin Invest 24: 91–99, 1994.
55. Weinstock JV and Elliot D. The substance P and somatostatininterferon-� immunoregulatory circuit. Ann NY Acad Sci 840:532–539, 1998.
56. Wieland T, Liedel K, Kaldenberg-Stasch S, Meyer zu Her-ingdorf D., Schmidt M., and Jakobs KH. Analysis of recep-tor-G protein interactions in permeabilized cells. NaunynSchmiedebergs Arch Pharmacol 351: 329–336, 1995.
57. Wilkie TM, Scherle PA, Strathmann MP, Slepak VZ, andSimon I. Characterization of G-protein � subunits in the Gq
class: expression in murine tissues and in stromal and hemato-poietic cell lines. Proc Natl Acad Sci USA 88: 10049–10053,1991.
58. Yamada Y, Post SR, Wang K, Tager HS, Bell GI, and SeinoS. Cloning and functional characterization of a family of humanand mouse somatostatin receptors expressed in brain, gastroin-testinal tract, and kidney. Proc Natl Acad Sci USA 89: 251–255,1992.
59. Yamashita N, Shibuya N, and Ogata E. Hyperpolarization ofthe membrane potential caused by somatostatin in dissociatedhuman pituitary adenoma cells that secrete growth hormone.Proc Natl Acad Sci USA 83: 6198–6202, 1986.
C190 EFFECTS OF SOMATOSTATIN ON B LYMPHOBLASTS
AJP-Cell Physiol • VOL 284 • JANUARY 2003 • www.ajpcell.org