An evaluation of the genus Amphidinium (Dinophyceae) combiningevidence from morphology, phylogenetics, and toxin production, withthe introduction of six novel species
Sarah Karafasa,*, Sing Tung Tengb, Chui Pin Leawc, Catharina Alves-de-Souzad
aAlgal Resources Collection, University of North Carolina Wilmington, Marine Biotechnology Facility, 5600 Marvin K. Moss Ln., Wilmington, NC 28403, USAb Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, MalaysiacBachok Marine Research Station, Institute of Ocean and Earth Sciences, University of Malaya, Bachok, 16310 Kelantan, MalaysiadAlgal Resources Collection, University of North Carolina Wilmington, Marine Biotechnology Facility, 5600 Marvin K. Moss Ln., Wilmington, NC 28403, USA
A R T I C L E I N F O
Article history:Received 4 May 2017Received in revised form 1 August 2017Accepted 1 August 2017Available online xxx
Keywords:Secondary structureCompensatory base changeCytotoxicityScales28SITS
A B S T R A C T
The genus Amphidinium is an important group of athecated dinoflagellates because of its high abundancein marine habitats, its member’s ability to live in a variety of environmental conditions and ability toproduce toxins. Furthermore, the genus is of particular interest in the biotechnology field for its potentialin the pharmaceutical arena. Taxonomically the there is a history of complication and confusion over theproper identities and placements of Amphidinium species due to high genetic variability coupled withhigh morphological conservation. Thirteen years has passed since the most recent review of the group,and while many issues were resolved, some remain. The present study used microscopy, phylogenetics ofthe 28S region of rDNA, secondary structure of the ITS2 region of rDNA, compensatory base change data,and cytotoxicity data from Amphidinium strains collected world-wide to elucidate remaining confusion.This holistic approach using multiple lines of evidence resulted in a more comprehensive understandingof the morphological, ecological, and genetic characteristics that are attributed to organisms belonging toAmphidinium, including six novel species: A. fijiensis, A. magnum, A. paucianulatum, A. pseudomassartii, A.theodori, and A. tomasii.
journal home page : www.elsevier .com/ locat e/hal
1. Introduction
The genus Amphidinium is a group of athecated dinoflagellatesthat are incredibly diverse in that, while being highly conservedmorphologically, they thrive world-wide in a wide variety ofhabitats (Dodge,1982; Dolapsakis and Economou-Amilli, 2009; FløJørgensen et al., 2004a, 2004b; Larsen, 1985; Larsen and Patterson,1990), temperatures (Murray and Patterson, 2002), and trophicmodes (Flø Jørgensen et al., 2004; Murray et al., 2004; Murray andPatterson, 2002). They also produce toxins and bioactive com-pounds that can have both harmful effects. Reports indicate thatAmphidinium blooms cause fish kills and that the toxins theyproduce may increase the effects of Ciguatera Fish Poisoning asthey are often found in association with Gamberdiscus in affectedareas (Baig et al., 2006; Rhodes et al., 2010; Tindall and Morton,
1998). They also have beneficial effects, producing compounds thatexibit antifungal or antimicrobial properties (Echigoya et al., 2005;Kobayashi and Kubota, 2007; Kobayashi et al., 1991; Kobayashi,2008; Meng et al., 2010; Nuzzo et al., 2014; Satake et al., 1991;Washida, 2006). They grow easily in culture, and can be scaled upto mass culturing volumes appropriate for chemical analyses.
Presently the genus includes both heterotrophic and autotro-phic forms possessing a characteristically minute epicone that isdeflected towards the left. The autotrophic Amphidinium consist oftwo clades (Flø Jørgensen et al., 2004a, 2004b) that are sister to oneanother. The first will be referred to as the Herdmanii Clade andincludes: A. steinii Lemmerman, A. mootonorum Murray and D. J.Patterson, A. herdmanii Kofoid and Swezy, and A. cupulatisquama M.Tamura and T. Horiguchi. The second is the Operculatum Clade andincludes: A. carterae Hulburt, A. massartii Biecheler, A. gibbossum (L.Maranda and Y. Shimizu) Flø Jørgensen and Murray, A. trullaShauna Murray, Rhodes, and Flø Jørgensen, A. operculatumClaparède & Lachmann, and, most recently, A. thermaeumDolapsakis and Economou-Amilli. The organisms in the HerdmaniiClade are distinguishable by a high degree of genetic diversity.
Table 1Strain ID numbers, Genbank accession numbers, and collection locations of all Amphidinium species used in this study. Bolded strain IDs indicate strains that are maintained atthe Algal Resources Collection. Boxed strains have been examined using SEM.
Species Strain LSU Accession ITS Accession Collection Location
Amphidinium carteraeAmca1004-2 KY070353 St. John’s Island, USVI
KY070351 KY697954 Korotoga, FijiAmca1412-1 KY070349 Bahia Cofresí, Dominican RepublicAmca0812-1 KY070355 KY697939 La Parguera, Puerto RicoAmca1506-2 KY070350 KY697940 Korotoga, Fiji
KY070352 KY697941 Korotoga, FijiAmca0406-1 KY070360 KY697969 Little San Salvador Island, BahamasAmca0406-2 KY070362 KY697964 Little San Salvador Island, BahamasAmca0406-3 KY070359 KY697963 Little San Salvador Island, BahamasAmca0406-4,TAC010
EU046327 KY697977 Little San Salvador Island, Bahamas
Amca0406-5 KY070361 KY697953 Little San Salvador Island, BahamasAmca0406-6 KY697972 Little San Salvador Island, BahamasCCMP1748 AY4605862 KY697967 Main Lagoon, Carrie Bow Key, BelizeJL 3 AF2603801 NASM 11 AY4605802 Niteroi, Rio de Janeiro, BrazilCCMP 121 AY4605852 Symbiont of CassiopeiaCS-383 JQ3948063 Bicheno, TAS, AustraliaCS-212 JQ3948053 Bay of Naples, ItalyCS-21 JQ3948073 Halifax, CanadaUTEX LB1946 FJ8235324 NACCMP 124 AY4605842 EU927575 Puerto Penasco, Sonora, MexicoCCMP 2100 KY697981 FJ823530 4 Eilat, IsraelCS-740 AY4605782 JQ6474243 Port Botany, NSW, AustraliaCCMP 2980 KY697968 Florida Keys National Marine Sanctuary, Marquesas Keys, Florida, USACCMP 1684 KY697966 Muscat, Oman
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 129
Table 1 (Continued)
Species Strain LSU Accession ITS Accession Collection Location
Amphidinium magnumAmma0206-1 KY070341 KY697948 Grand Bahama Beach, BahamasAmma0206-2 KY697946 Grand Bahama Beach, BahamasAmma0206-3 KY070347 KY697949 Grand Bahama Beach, BahamasAmma0206-4 KY070342 KY697947 Grand Bahama Beach, Bahamas
KY070343 Grand Bahama Beach, BahamasAmma0206-6 KY070344 Grand Bahama Beach, Bahamas
Amphidinium massartiiEU046333 KY697952 Off shore North Carolina, USA (Gulf Stream)
TM16 AB8189528 Shirahama Beach, JapanHG 115 AB4773475 Ikei Beach, Ikei Is., Okinawa Prefecture, JapanCCMP 1821 AY4556707 Succotash Marsh, South Kingston, RI, USA
Amphidinium trulla AY4605942 Rangaunu, New ZealandK-0657 AY4556717 Jægerspris, Isefjorden, Denmark
Amphidinium tomasii
130 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 131
Likewise, within the Operculatum Clade, A. operculatum is highlydivergent from the remaining species. There is much less geneticdivergence, however, among the remaining members of theOperculatum Clade, i.e, A. thermaeum, A. carterae, A. massartii, A.gibbosum, and A. trulla.
In addition to size and shape, a wide variety of morphologicalfeatures were historically used to identify these species, includingthe presence, location, and number of nuclei, pusules, pyrenoids,chloroplasts, eyespots, scales, and life cycle stages. There is,however, such a high degree of overlap in these characters, bothwithin and between species, that finding distinct morphologicalcharacters to define taxonomic groups by has been challenging atbest. Flø Jørgensen et al., 2004a and Murray et al. (2004) madenotable strides redefining morphological characters in the typespecies of the genus and eliminating confusion among the strains.The issue became even clearer with the reassignment of organismsthat were erroneously considered Amphidinium into their propergenera (e.g. Hoppenrath et al., 2012; Horiguchi et al., 2012; Onumaet al., 2015; Takano et al., 2014). But, as more strains ofAmphidinium are established and more molecular informationbecomes available, it is clear that many features once consideredindicative of a species are no longer valid. For example, the definingcharacteristics of A. steinii were reported to be its large size,metabolic movement, and cyst formation (Kofoid and Swezy, 1921;Murray et al., 2004). Metabolic movement and cyst formation werealso later reported in A. thermaeum (Dolapsakis and Economou-Amilli, 2009), leaving only size to differentiate the two species, andeven then overlap of about 10 mm occurs despite the fact that thesetwo species are highly evolutionarily divergent. There remainsconfusion in Amphidinium taxonomy also because its nomencla-ture is still littered with the descriptions of organisms that areactually synonyms of other species.
The Algal Resources Collection (ARC) maintains over 750 strainsof harmful algae representing diatoms, phytoflagellumtes, anddinoflagellates. Included in the collection are approximately 58Amphidinium strains from locations world-wide. An effort toensure proper classification to species level for each strain led tothe discovery of morphologically and/or genetically unique strainsof Amphidinium in the Operculatum Clade, specifically. Thepurpose of this study was to use multiple lines of evidence tojustify the formation or maintenance of species level taxon in thegenus Amphidinium. Considering multiple lines of evidence willhelp develop a more accurate narrative of the evolution of
1 Daugbjerg et al. (2000).2 Murray et al. (2004).3 Murray et al. (2012).4 Stern et al. (2012).5 Tamura et al. (2009).6 Hikosaka-Katayama et al. (2012).7 Flø Jørgensen et al., 2004a.8 Owari et al. (2014).9 Lee et al. (2013).10 Rhodes et al. (2014).11 Hoppenrath and Leander (2010).12 Murray and Patterson (2002).13 Cyronak (2007).14 Dolapsakis and Economou-Amilli (2009).
Amphidinium. Through a combination of morphometric, genetic,secondary structure, compensatory base changes (CBC), andtoxicity analyses this study aims to offer new insights into thetaxonomy, systematics, and toxicity of the genus with anintroduction of six novel species.
2. Methods
2.1. Collection and isolation
The samples of Amphidinium maintained at ARC and used in thisstudy were collected by a variety of researchers and collaboratorsover a number of years and locations (Table 1). Completeinformation can be accessed on the ARC website (algalresources-collection.com) or through contact with ARC staff. In most cases,individual cells were isolated from raw water samples with sterilepipettes and placed into separate wells of a 96-well plate. Theisolated cells were grown in culture and stepped up serially to250 ml Erlenmeyer flasks using appropriate media. In other casesstrains were acquired from other collections as monocultures.
2.2. Microscopy
Strains were observed live using a Zeiss Axio Imager Z.1Microscope equipped with a 100 � oil objective and AxioVisionsoftware. Fluorescent images were captured for the autofluor-escent plastids using the Alexa Fluor 430 filter. Cells were alsoprepared for scanning electron microscopy as follows: 250 ml to1 ml of cultured cells was gently filtered through either 5 mm or8 mm Nucleopore filters (Whatman) and treated with a 2% osmiumtetroxide in a 40% solution of the culture salinity, in this case asalinity of 36 � 39, for 20 min. The samples were then rinsed withthe 40% salinity solution three times and then dehydrated with tworinses each of 10, 30, 50, 75, 95, and 100% ethanol over a period oftwo days. The filters containing cells were processed with a criticalpoint dryer, placed on stubs, and coated with 12 nm of platinum/palladium. Samples were viewed on a Philips XLS-FEG scanningelectron microscope, using the secondary electron or through thelens detectors where necessary. Ideally two strains representingeach major phylogenetic clade, including the type specimen fornovel species, were prepared for SEM (Table 1).
ion Collection Location
Raoul Island, Kermadec Islands Bahia Cofresí, Dominican Republic
Table 2Pooled of morphological features of Amphidinium species compiled from the results of the present study and from literature sources. Species are roughly arranged by size.
A. carteraec,d
A. massartii d,e A. cf. massartiic
A.thermaeuma,e
A. cf.thermaeum
A.pseudomassartii
A.theodori
A. fijiensis A. trulla d A.tomasii A.paucianulatum
straight straight straight NA short, straight short,straight
short, straight short,straight
straight short, straight longcurved
short, straight
Pyrenoid Observed yes yes yes varies yes yes yes yes yes varies yes yes, 2–4 yesPyrenoid Location central central central central central lateral central central central central central leteral centralPusules Observed no no no varies no no no cysts no no no yes yesScales observed no yes yes no no no yes no no no yes no noCell division motile motile motile cysts cysts motile cysts cysts motile motile motile mobile motileMetabolic movementobserved
no no no yes NA no yes yes no no no no no
Toxic yes no yes yes NA yes yes no yes yes yes yes yes
aDolapsakis and Economou-Amilli (2009).bFlø Jørgensen et al. (2004a, 2004b).cLee et al. (2013).dMurray et al. (2004).eMurray et al. (2012).
132
S. Karafas
et al.
/ Harm
ful Algae
68 (2017)
128–151
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 133
2.3. DNA extraction and sequencing
DNA was extracted using the Chelex method (Richlen andBarber, 2005), and the LSU rDNA D1 � D6 regions using the primersD1R (Scholin et al.,1994) and 28–1483 (Daugbjerg et al., 2000). The
Fig. 1. Amphidinium fijiensis. Amfi0508-1. Scale bars are 10 mm unless otherwise noted.plastids, (E) cell division within temporary cyst, (F–H) were taken at the same magnifi
universal primers ITSL (Hsiao et al., 1995) and D2C (Scholin et al.,1994) were used to amplify the ITS1-5.8S-ITS2-partial LSU regionsof the rDNA. For both primer pairs, GoTaq Green Mastermix wasused according to the manufacturers protocols (Promega, Madison,Wisconsin USA), and thermocycler conditions were as follows:
Py = pyrenoid. (A) Apical view, (B) ventral view, (C) lateral view, (D) epifluorescentcation of a single cell displaying metabolic movement.
134 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
initial denaturing at 94 �C for 2 min followed by 35 cycles of 94 �Cfor 30 s, 50 �C for 40 s, 72 �C for 1 min 45 s with a final extensionperiod at 72 �C for 5 min. Amplification products were viewed on a1% agarose gel and successful amplicons were cleaned usingExosap-IT (Affymetrix, Santa Clara, California USA) and used astemplates in Big Dye (v.3.1, Applied Biosystems, Foster City, CA,USA) sequencing reactions. Sequencing reactions were run on anABI 3100 Genetic Analyzer (DNA Analysis Core Facility, CMS) andedited and assembled using Sequencher (Gene Codes Corp., AnnArbor, MI, USA). Sequences obtained in this study were depositedin Genbank with the accession numbers presented in Table 1.
For the LSU sequence analysis, 78 Amphidinium sequences wereeither obtained from strains housed at ARC or retrieved fromGenbank (Table 1) and were aligned using Mega 5.2 (Tamura et al.,2011). Strains of Karlodinium (DQ114467) and Heterocapsa(AY571372) were used as outgroups. Total sequence length was1268 bp, with every effort made to only include sequences thatspanned the entire sequence length. When this was not the case,the phylogeny was tested with and without the shorter sequences,
Fig. 2. Amphidinium magnum. Amma0206-5. Unless otherwise noted scales bars are 10 m(D) ventral view SEM � asterisks indicate the beginning and end of the ventral ridge,
and only when there was no difference in topologies or significanteffect to the bootstrap values were the sequences maintained.
The GTR + G substitution model, determined in Mega 5.2(Tamura et al., 2011) as the best-fit model according to theBayesian Information Criterion scores, was used to perform allphylogenetic analyses. Between-group uncorrected pairwise dis-tances and maximum likelihood (ML) analysis with 1000 bootstrapreplicates were performed with Mega 5.2 and RAxML-HPC v.8 inthe CIPRES Portal, respectively. Additionally, Bayesian analysis wasperformed in Mr. Bayes 3.2 using three independent runs of1,000,000 generations and a 25% burn-in. Only support valuesgreater than 50% were reported in the phylogeny.
2.4. ITS2 secondary structure and CBC analyses
The secondary structure of the ITS2 region was examined for 59strains of Amphidinium, using a strain of Karlodinium (EF036540)and Heterocapsa (KF240777) as outgroups. The exact ITS2 termini
m. Py = pyrenoid, Pu = pusule, VR = ventral ridge. (A & B) ventral view, (C) dorsal view,(E) epifluorescence, (F) cell surface at high magnification.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 135
were determined with the construction of the 5.8S and 28Sproximal stem. Preliminary ITS2 secondary structure transcriptswere predicted by using RNAstructure ver. 5.8.1 (Bellaousov et al.,2013). The structure was optimized based on the common motifsof ITS2 as described in Koetschan et al. (2010).
Multiple sequence-structure alignment of the ITS2 dataset wasgenerated using 4SALE v1.7 (Seibel et al., 2006, 2008; Wolf et al.,2005, 2014), with a 12 � 12 scoring matrix (Wolf et al., 2014). Thealignment with sequence-structure information was used toconstruct a ML phylogeny using Phargorn (Schliep, 2011) asdetailed in Teng et al. (2016). Maximum likelihood parameterswere set to 1000 bootstrap replicates with NNI optimization. TheCBC matrix was generated by using 4SALE. The hemi-CBCs werehighlighted through the structure comparing in 4SALE.
Fig. 3. Amphidinium paucianulatum. Ampa0606-1. Scale bars are 5 mm unless otherwiseridge. (A & B) ventral views in DIC and SEM, respectively, (C) dorsal view, (D) close up o
2.5. Brine shrimp toxicity assay
Ten newly hatched Artemia salina were isolated into each well ofa 24-well plate containing 2 ml of seawater at a salinity of 36. Arepresentative strain from each clade identified in the phylogenieswas grown in culture in 1500 ml of the appropriate salinity for twoweeks. Fourteen hundred milliliters of cells in exponential growthphase were harvested with a Hermle Labnet Z383 K centrifuge intoa pellet and frozen at �80 �C until later chemical extraction couldbe performed. Pellets were then brought to room temperature andextracted 2 x with 80:20 MeOH:H2O with sonication. Thecombined filtered extracts were dried in vacuo. The extracts werethen re-suspended in 10 ml 95:5H2O:MeOH and 20 mg of eachextract was loaded onto a separate 200 mg SPE cartridge (WatersOasis Prime HLB). Columns were first eluted with 1 column volume
noted. Py = pyrenoid, S = scale, Asterisks mark the beginning and end of the ventralf flagellar insertion, (E) cell surface at high magnification, (F) epifluorescent view.
136 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
of 95:5 water/methanol. The columns were then eluted with 1CVof 90:10 MeCN:MeOH. This organic fraction was dried and 100 mgprepared for bioassay. The final concentration of each extract was1.6 mg�ml�1 in DMSO, 25 ml of which was applied to each well withbrine shrimp for a final concentration of 0.021 mg�ml�1. Sixreplicates were used for a total sample size of n = 60, both DMSOand seawater were used as controls. Toxicity was assessed every24 h until the health of the control shrimp began to deplete(usually by 72 h).
3. Results
3.1. Morphology � species descriptions
Morphological analysis was performed on any Amphidiniumstrain that was either genetically distinct or gave ARC investigatorspause during routine microscopic examinations. Six novel species,presented in Table 1, resulted from that analysis, the morphologiesof which are described below. Many other strains maintained atARC identified phylogenetically with previously established taxa,such as A. carterae, A. gibbosum, A. massartii, A. thermaeum, or A.trulla. Strains that fell into those species were examined withscanning electron microscopy (circled in Table 1) to verify thattheir morphology was consistent with the original descriptions foreach respective species, and were not examined further. Importantmorphological features identified in this study for the novelspecies introduced herein, as well as features from speciesestablished or analyzed in other studies, were consolidated andare presented in Table 2, which can serve as a quick reference of themost important characters in Amphidinium taxonomy. In mostcases there was not one specific trait that was unique for anyAmphidinium species, instead it was the specific combination ofcharacters that was important, and Table 2 allows for thesefeatures to easily be compared and contrasted between species.The descriptions provided herein provide more detail thanavailable in Table 2. Attempts were made to provide a uniformpattern of description for each species presented, however, theability to do so was dependent largely on cell orientation and
preparation condition in the case of SEM and cellular movementpatterns in live cells. Character information was only providedwhen it could be done with certainty and without ambiguities.
Amphidinium fijiensis Karafas and Tomas sp. nov.Description: Cell shape varied greatly from round to oval,
elliptical, or pear-shaped (Fig. 1). Typically round cells werestationary, resting at the bottom of the slide while oval, elliptical orpear-shaped cells were swimming freely in the water column. Celllength was 9.8 � 22.7 mm (12.7 � 2.0 mm, n = 127) and width was5.7 � 14.9 mm (11.3 � 1.6 mm, n = 127). The length to width ratiowas 0.8 � 2.2 (1.2 � 0.4, n = 127). The epicone (Fig. 1A) was minuteand left-deflecting with a rounded tip. In five cells the epiconelength averaged 7.73 mm � 1.1 mm, slightly more than half theaverage cell width. It protruded only minimally on the ventral side.The cingulum was displaced (Fig. 1B) and the proximal end nearlymade contact with the ventral ridge and sulcus so as to form a v-shape. The ventral ridge was short and straight (Fig. 1B), averaging5.29 mm � 0.8 mm (n = 6), and connected the two flagella insertionpoints. The length of the ventral ridge did not seem to change,though few cells were observable for this character. When the cellswere in the elongated state, however, the angle of the ventral ridgeto the longitudinal midline of the cell was very small (Fig. 1B). Thehypocone was widest at the cell’s midpoint when in elliptical andround forms, and in the posterior when pear-shaped. The anteriorshoulders of the hypocone were symmetrical in most cases, but notin all. Cells possessed rounded antapices The longitudinalflagellum was inserted in the middle 1/3 of the cell just belowthe proximal end of the cingulum at the beginning of the sulcus,which was shallow and did not reach the antapex of the cell(Fig. 1B). A central pyrenoid (Fig. 1C � “Py”) was visible in lightmicroscopy from which radiated a single plastid that formed aperipherally located lobe with many perforations (Fig. 1D). Nopusules could be observed in light microscopy. The nucleus wasposterior. Reproduction within temporary haline cysts (Fig. 1E).Metabolic movement was directly observed in samples ofAmfi0508-1 (Figs. 1F–H), and nearly all of the cells displayed theround morph after a few minutes on the slide. Plates and scalescould not be observed. Non-toxic.
l view, (C) close up of cell surface, (D-F) Lugol’s preserved. FI = flagellar insertion,
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 137
Type Locality: Korotoga FijiHolotype: Amfi0508-1. Voucher material will be deposited in
the New York Botanical Garden. Living material maintained at theAlgal Resources Collection (ARC), Wilmington NC
Etymology: Amphidinium fijiensis is named for its presumed typelocality.
Distribution: To date Amphidinium fijiensis has only beencollected from a fish tank containing “live rock” imported fromKorotoga Fiji.
Amphidinium magnum Karafas and Tomas sp. nov.Description: The oval cells (Fig. 2) were 25.9–47.3 mm long
(33.4 � 4.6 mm, n = 44) and 13.9–36.2 mm wide (23.9 � 4.1 mm,n = 44). The left deflecting epicone was flat on its anterior (Fig. 2Aand B), about half as wide as the cell width (11.7 � 1.8 mm, n = 29).Its right side was either 90� or sloped slightly left (Fig. 2A and B) inlight microscopy. The epicone overlapped the anterior portion ofthe hypocone just slightly (Fig. 2C), sometimes resulting in a softlycurved dip in the dorsal anterior portion of the hypocone. Thehypocone itself was symmetrical with the left and right sides equalin height and the widest point was typically at the cell center(Fig. 2D). Typically both sides of the cell were convex, though onrare occasions the left side was straight. The antapex was eitherrounded or came to a soft, round point. The cingulum was wide andhoused a thin transverse flagellum. The sulcus began just posteriorto the cell center (Fig. 2D) and the longitudinal flagellum wasinserted just inside the sulcus (�0.6 � 0.07 mm, n = 7). The sulcusdid not reach the antapex of the cell and the sulcus and cingulumdid not make contact. The two insertion points of the flagellainsertion were connected by a narrow, long ventral ridge (VR) inFig. 2D that in a small sample size (n = 4) averaged 17.34 � 0.8 mm.The ventral ridge nearly touched the proximal end of the cingulumat the transverse middle of the cell. At this point the ventral ridge,which was travelling at about a 30�45� angle from the insertion ofthe transverse flagellum, curved to make a more vertical path theremaining distance to the sulcal groove. The severity of the curve
Fig. 5. Amphidinium theodori. Amth0702-1. (A) apical view, (B-D) ventral views, (E) encys� an example of a scale is circled, (G) dividing encysted cells, (H) cluster of dividing e
varied. Two pusules were present (Fig. 2B), but a central pyrenoidwas lacking. Instead 2–4 organelles (Fig. 2A–C), possibly pyrenoids,were visible toward the periphery of the cells in light microscopy.The plastid lobes (Fig. 2E) were filamentous and sparse in thecenter of the cell and over the nucleus. They appeared concentrat-ed on the sides and appeared to be radiating from laterally locatedpyrenoids. The cell surface (Fig. 2F) was covered in a hexagonalplate structure, but scales were not observed. Metabolic movementand cysts were not observed. Toxic.
Type locality: Bahamas Beach, Bahamas; 26.716002 N,78.107986 W
Holotype: Amma0206-5, stubs prepared for SEM will bedeposited in New York Botanical Gardens. Living material ismaintained at the Algal Resources Collection, Wilmington NC.
Etymology: magnum refers to the large size of the speciesDistribution: BahamasAmphidinium paucianulatum Karafas and Tomas sp. nov.Description: Cells (Fig. 3) were elliptical or pear shaped and
16.5–40.6 mm long (27.1 �6.1 mm, n = 47) and 12.3–34.8 mm wide(19.6 � 4.5 mm, n = 44). The right and left side of the cells wereasymmetrical, with the left side being slightly longer and usuallystraight or concave (Fig. 3A). The right side was convex androunded (Fig. 3A). The cell was widest at the cells midpoint orslightly anterior in most cells (Fig. 3A–C), though a few wereobserved to be widest posterior to the cell’s midpoint. The epiconewas flattened anteriorly (Fig. 3A), and averaged 10.92 mm � 2.3 mm(n = 9) wide. When viewed from the ventral side the right side ofthe epicone appeared to slope towards the right of the cell beforeturning towards the left. The antapex of the cell appeared flattenedin light microscopy (Figs. 3A and C) and slightly rounded inelectron microscopy (Fig. 3B). The descending cingulum (Fig. 3B)began 0.2 � 0.03 cell lengths from the apex of the cell. Dorsally thecingulum path curved slightly to create a shallow, u-shapedindentation in the anterior dorsal portion of the hypocone (Fig. 3C).The proximal end of the cingulum appeared to connect completely
ted cell, Pyrenoid labeled to the right with a “Py” (F) close up of scales on cell surfacencysted cells.
138 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
with the sulcus in most cases (Fig. 3B and D), which did not reachthe antapex of the cell. The insertion of the longitudinal flagellumwas typically at the middle 1/3 of the cell (0.49 � 0.7 cell lengthsfrom apex) and appeared to be to the right of the beginning of thesulcus and immediately below the ventral ridge (Fig. 3B and D),
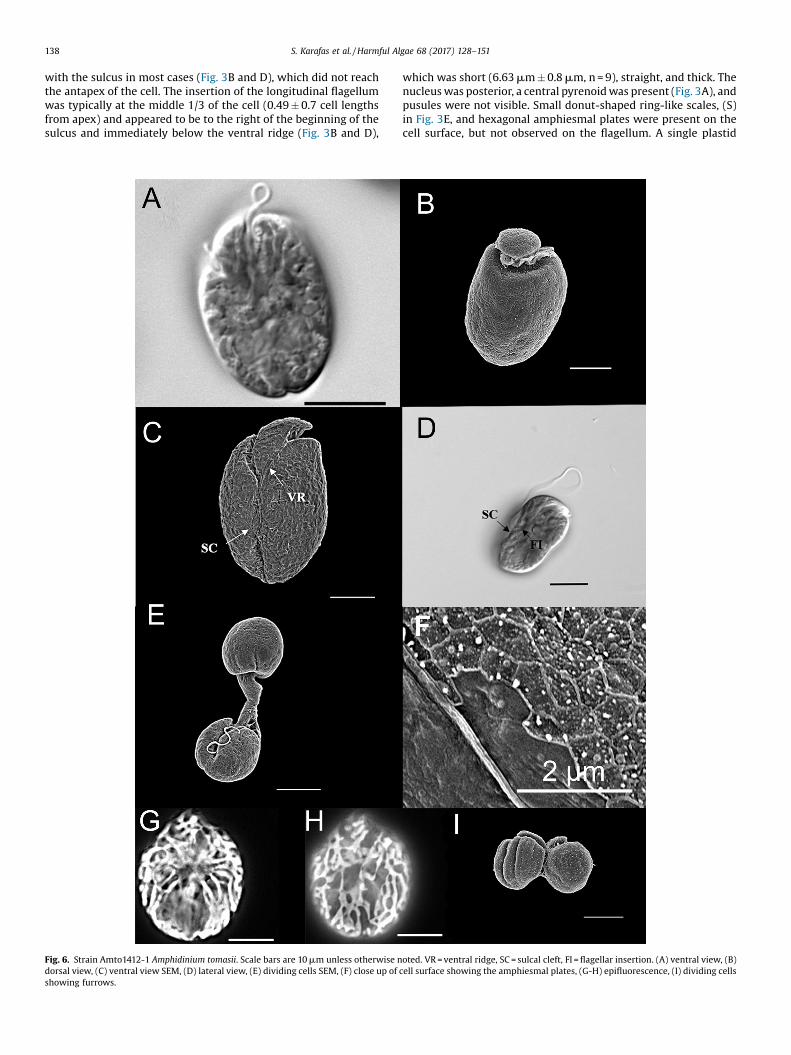
Fig. 6. Strain Amto1412-1 Amphidinium tomasii. Scale bars are 10 mm unless otherwise ndorsal view, (C) ventral view SEM, (D) lateral view, (E) dividing cells SEM, (F) close up of
showing furrows.
which was short (6.63 mm � 0.8 mm, n = 9), straight, and thick. Thenucleus was posterior, a central pyrenoid was present (Fig. 3A), andpusules were not visible. Small donut-shaped ring-like scales, (S)in Fig. 3E, and hexagonal amphiesmal plates were present on thecell surface, but not observed on the flagellum. A single plastid
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 139
radiated from the pyrenoid (Fig. 3F). The plastid lobes were thick,relatively few in number, and extended over the nucleus. Nometabolic movement or cysts were observed. Toxic.
Type locality: Palma Sola, Clearwater, Florida; 27.628854 N,82.655468 W
Holotype: Ampa0606-1, stubs prepared for SEM will bedeposited in New York Botanical Gardens. Living strains aremaintained at the Algal Resources Collection.
Etymology: paucianuli means “few rings” in Latin, referring tothe sparsely distributed ring-like scales on the cell surface.
Distribution: Clearwater Florida (type); Korotoga FijiAmphidinium pseudomassartii Karafas and Tomas sp. nov.Description: Round to elliptical cells (Fig. 4) are 12.8 � 21.2 mm
(16.7 � 2.1 mm, n = 66) in length and 7.7 � 18.7 mm (13.9 � 2.4 mm,n = 64) wide with a length to width ratio of 1.0-1.8 (1.2 � 0.2, n = 53).The left deflecting epicone is crescent shaped and has a rounded tip(Figs. 4A and B). There was considerable apical rise in the epiconesof cells observed and the right sides of those epicones were slopedtoward the left of the cell (Fig. 4B). The cingulum is displaced andmeets the short (4.88 mm � 0.5 mm, n = 6) straight ventral ridge toform a ventral v-shape when viewed in light microscopy. Thelongitudinal flagellum is inserted in the middle 1/3 of the cell at thebeginning of the sulcal groove directly below the proximal end ofthe cingulum (Fig. 4B). The sulcus is shallow and does not reach theantapex of the cell (Fig. 4B). No scales were observed on the cellsurface (Fig. 4C). The hypocone is asymmetrical with the left sidelonger than the right, even in rounder cells (Figs. 4B & D). Theantapex of the hypocone is rounded. The nucleus is posteriorlypositioned (Fig. 4E). The pyrenoid is visible and appears to belocated laterally (Fig. 4F) but pusules are not visible. Cell divisionoccurs in motile cells. Haline cysts were not observed. Toxic
Type locality: Tasmania, AustraliaHolotype: Strain AKLV01, SEM stubs will be deposited at the
New York Botanical Garden.
Fig. 7. Cells representing Amphidinium strain Amth0000600. Scale bars are 10 mm. Photoencysted cell dorsal view, (D) ventral view of elongated cell, (E) dorsal view, and (F) la
Etymology: pseudomassartii means “like massartii” in LatinDistribution: AustraliaRemarks: The type strain AKLV01 (Murray et al., 2004) is
represented in the ARC database as Amsi0006-1. Presently A.pseudomassartii is represented by only two strains isolated byImojen Pearce from Tasmania, Australia and was referred to asAmphidinium cf. massartii in Murray et al. (2004). In the originalpublication the authors noted that the strains were geneticallydistinct from other A. massartii, but refrained from assigning aunique taxonomic designation for lack of distinct morphologicalcharacters. Herein the strains are assigned to the novel speciesAmphidinium pseudomassartii on the basis of both morphology andgenetic analyses. Cells of A. pseudomassartii seem to lack the rounddoughnut-like scales that are present on the surface of A. massartiistrains, have wider epicones than A. massartii, and are toxic.Furthermore, the number of taxa included in the present study hasprovided even more insight into the genetic distinction of thisspecies from A. massartii and strengthens the case supportingtaxonomic distinction. The epithet “pseudomassartii” was chosento reflect the historic thinking throughout the literature that theorganism was closely related to A. massartii though the presentphylogenetics show that it is more closely related to other speciesof Amphidinium.
Amphidinium theodori Tomas and Karafas sp. novDescription: The oval to elliptical cells of Amphidinium theodori
(Fig. 5) are 13.0–20.9 mm (16.6 � 2.7 mm, n = 20) long and 9.8–17.0 mm (12.6 � 2.2 mm, n = 20) wide. The left deflecting epicone(Fig. 5A) is relatively flat, is 5.79 � 0.9 mm (n = 6) and has a rightside that slopes to the right before angling to the left (Fig. 5B–D).The insertion points of the flagella are connected by a short ventralridge (Fig. 5B), averaging 4.00 � 0.4 mm (n = 6). The longitudinalinsertion point is in the anterior 1/3 of the cell (Figs. 5B, 5C, and5D). The sulcus is not well defined. A central pyrenoid wasobserved (Fig. 5E) but pusules were not. The nucleus is posterior.
s courtesy of Tyler Cyronak. (A) ventral view, (B) epifluorescent/DIC combination, (C)teral view.
140 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
The antapex of the hypocone is round (Fig. 5B and D). Ring-likescales are present on the cell surface (Fig. 5F). Cell division occursin cysts of either a single division (Fig. 5G) or multiple divisionswithin the same cysts (Fig. 5H). Few plastid lobes appear to radiatefrom the central pyrenoid in a trunk-like fashion and spread acrossthe periphery of the cell. Metabolic movement was observed.Toxic.
Type Locality: Korotoga FijiHolotype: Amth0702-1, stubs will be deposited in the New York
Botanical Garden and living material is maintained at the AlgalResources Collection
Etymology: Amphidinium theodori is named in honor of Dr.Theodore Smayda for a life-time of contributions to the field ofphytoplankton ecology.
Distribution: Fiji, U.S. Virgin IslandsRemarks: The metabolic movement observed in this species
made characterizing shapes difficult. As observed in Figs. 5B–D,there are a variety of shapes the hypocone can take. The shouldersof the hypocone can be either symmetrical or not, the antapex canbe slightly pointed, and at times the hypocone even appearedtwisted. In some cells the ventral ridge appeared to completelymeet the proximal end of the cingulum.
Amphidinium tomasii Karafas sp. nov.Description: Cells were oval to elliptical in shape and slightly
flattened dorsoventrally (Fig. 6A). On rare occasion rounder cellswere observed. Cells were 18.6-42.2 mm in length (25.5 � 4.9 mm;n = 28) and 12.9-24.5 mm in width (18.4 � 3.4 mm; n = 25), andwhen oval, were typically widest at the center or anterior of thecell. The, very slightly, left-deflecting epicone (Fig. 6B, C) wascurved and rounded and measured 8.75 �1.8 mm (n = 17). Theapical rise of the epicone varied from no rise to extreme rise, but
Fig. 8. Maximum likelihood tree of the ribosomal large subunit of the Amphidinium. Valueposterior probabilities. The dark triangle represents the Operculatum Clade. The width odiversity.
most commonly it remained close to the cell and slightly overhungthe dorsal, anterior side of the hypocone. In SEM the right side ofthe epicone sloped to the right before angling to the left. Theanterior portion of the hypocone varied from symmetrical toasymmetrical, with the left side longer than the right. The left edgeof the hypocone varied from being straight to convex. The rightside of the hypocone was nearly always convex. The dorsal, apicalside of the hypocone occasionally possessed an anterior longitu-dinal cleave. The transverse flagellum was in a well-definedcingulum (Fig. 6B). The distal end of the cingulum terminated atthe transverse midpoint of the cell meeting or nearly meeting thethin ventral ridge. The narrow sulcus began around the ventralmidpoint of the cell and appeared to turn to the right briefly shiftleft again and widen posteriorly (Fig. 6C), giving the antapex of thecell a subtle or bi-lobed appearance when viewed in certainpositions (Fig. 6A). The curved appearance of the sulcal pathwayseemed to be due to a sulcal cleft (Fig. 6C–E). The longitudinalflagellum was inserted in the middle 1/3 of the cell (Fig. 6D). Thepronounced sulcal cleft and a very narrow distal portion of thesulcal groove (Fig. 6C) sometimes obscured the insertion point ofthe longitudinal flagellum so that it was not always visible andappeared to have posterior position. A ventral ridge measuring8.93 � 2.8 mm (n = 18) connected the two points of flagellainsertion. Pusules were not visible. A hexagonal plate structurewas seen on the cell surface, but scales were not observed (Fig. 6F).The plastid appeared to be single with many thin stranded lobesradiating from the cell center and running over the perimeter like amaze (Fig. 6G and H). On rare occasions a central structure,presumably a pyrenoid, was observed. The nucleus was located inthe posterior of the cell (Fig. 6A and D). Asexual division occurredin motile cells (Fig. 6E) and furrows were present during the
s above the line are bootstrap values and those below the line are Bayesian Inferencef the triangle represents robustness of taxa and the height of the triangle represents
Fig. 9. Maximum likelihood phylogeny of the ribosomal large subunit of the autotrophic Amphidinium strains belonging to the Operculatum Clade. These species arerepresented in Fig. 8 by the black triangle. Values above the line represent bootstrap values and below the line represent posterior probabilities from Bayesian inference.Strains are indicated by the collection identification numbers. Roman numerals indicate subclades referred to in the text.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 141
Fig. 10. Secondary structure phylogeny generated from the ITS2 region of theribosomal DNA of autotrophic Amphidinium strains from the Operculatum Clade.Numbers on nodes represent bootstrap values. Roman numerals indicate sublcadesreferred to in the text.
142 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
mitotic process (Fig. 6I). No immobile or encysted cells wereobserved. Toxic.
Type Locality: Bahia Cofresi, Dominican Republic; 19.820175 N,70.726158 W
Holotype: Amto1412-1, Fig. 1, stub prepared for SEM will bedeposited in New York Botanical Gardens, living material main-tained at the Algal Resources Collection
Etymology: Named after Dr. Carmelo Tomas in honor of his manyyears of service and expertise in phytoplankton systematics.
Distribution: Dominican Republic, Florida, Puerto RicoAmphidinium sp. ARC strain Amth0000600Description: Cells were usually round but also showed a second
morph of elliptical cell (Fig. 7). Measurements taken by Cyronak
(2007) indicated that cells averaged 17.5 �1.9 mm (14–23 mm,n = 51) long and 16.5 �1.7 mm (14–21 mm, n = 51) wide. When cellswere round (Fig. 7A) the hypocone sides were symetrical in lengthand convexity. When cells were elliptical (Fig. 7B and C) the leftside was just slightly longer than the right and the cell appearedwidest in the middle (Fig. 7D). The left-deflecting epicone hungover the anterior of the hypocone on the dorsal side (Fig. 7E). Thelongitudinal flagellum (Fig. 7A) had a central insertion, and thenucleus was posterior but not always very visible. A centralpyrenoid was apparent in most micrographs (Fig. 7F), but pusuleswere not. Few plastid lobes radiated from the pyrenoid to theperiphery of the cell where they appeared as one superficial lobewith perforations (Fig. 7B). Metabolic movement was not recordedin the cells, but Cyronak (2007) noted that the cells producedtemporary cysts (Fig. 7C) within which they could be observedrotating. Also noted was that cells had the ability to exit the cystsand move freely in the culture medium. The cells were largely non-motile.
3.2. LSU rDNA phylogeny
The Amphidinium genus forms a strongly supported cladeaccording to maximum likelihood booststrap (BS) values (100) andBayesian Inference posterior probability (PP) values (1). Bothanalyses resulted in the same topologies, but only the maximumlikelihood phylogeny is displayed in Figs. 8 and 9. While allAmphidinium sensu stricto grouped together strongly, two majorclades were resolved within autotrophic Amphidinium represent-ing the Herdmanii Clade and the Operculatum Clade. Within theHerdmanii Clade, A. steinii strains were sister to a fully supportedclade including A. cupulatisquama, A. herdmanii, and A. mootono-rum. Strains of A. herdmanii and A. mootonorum were resolvedtogether in another fully supported clade, but there was no supportfor the relationships among the strains.
Species within the Operculatum Clade that were establishedusing traditional morphometrics grouped together phylogeneti-cally as well. Bootstrap values and posterior probabilities of theclades describing each of these unique taxonomic units were highin most cases, 74–100% and 1 respectively. Relationships betweenthese taxonomic units, however, were not always stronglysupported. The Amphidinium operculatum (Fig. 8) strains weresister to a strongly supported clade (93/1) containing theremaining species in the Operculatum Clade. Long branch lengthsuggests that the divergence of this species from the others wasgreat. The placement of Amphidinium trulla, A. gibbosum, and A.paucianulatum as sister taxa to any of the other species from theOperculatum Clade was not strongly supported with BS values.
All Amphidinium carterae strains grouped together in a stronglysupported clade (99/1) that was sister to a clade containing A.pseudomassartii, A. tomasii, A. theodori, A. magnum, A. thermaeum, A.fijiensis, and A. massartii (Figs. 9 and 10). Strains of Amphidiniummassartii and A. cf. massartii were sister taxa with one another andthe two formed a clade that was sister to a clade containing A.pseudomassartii, A. tomasii, A. theodori, A. magnum, A. thermaeum,and A. fijiensis, though neither showed very strong bootstrapsupport. Strong support (92/1) grouped A. pseudomassartii, A.tomasii, A. theodori, A. magnum, and A. thermaeum into a clade tothe exclusion of A. fijiensis. Likewise A. pseudomassartii wasstrongly excluded from the remaining taxa (98/1). The Amphidi-nium tomasii strains grouped together in a clade that was sister toone containing A. theodori and A. magnum as sister clades, as wellas A. thermaeum and A. cf. thermaeum as sister clades.
Table 3 represents the uncorrected p-distances within andbetween each taxonomic group described. The greatest intraspe-cific genetic diversity exists within the A. operculatum, A. carterae,and the A. massartii clades with values of 0.05, 0.05 and 0.04,
Table
3Unco
rrec
tedpairw
isege
netic
distance
sof
thelargeribo
somal
subu
nitsequ
ence
dataforsp
eciesof
Amph
idinium.B
olded
values
onthediago
nal
indicateintraspec
ificdistance
san
dthos
ebe
low
thediago
nal
indicateinterspec
ific
distance
s.
Spec
ies
carterae
cupu
latisqua
nafijie
nsis
gibb
osum
herdman
iimoo
tono
rum
massartii
pseu
domassartii
cf.
massartii
mag
num
thermae
umpa
ucianu
latum
tomasii
theo
dori
cf.
thermae
umop
ercu
latum
steinii
trulla
outgroup
incoloratum
carterae
0.05
cupu
latisqua
na0.35
NA
fijie
nsis
0.12
0.33
0gibb
osum
0.20
0.34
0.18
0he
rdman
ii0.35
0.24
0.33
0.39
0.02
moo
tono
rum
0.35
0.24
0.33
0.39
0.01
NA
massartii
0.12
0.34
0.08
0.18
0.34
0.33
0.04
pseu
domassartii
0.13
0.36
0.08
0.20
0.36
0.35
0.09
0cf.m
assartii
0.13
0.34
0.08
0.18
0.34
0.34
0.06
0.07
0.01
mag
num
0.19
0.37
0.16
0.25
0.39
0.39
0.16
0.15
0.16
0thermae
um0.17
0.38
0.15
0.24
0.37
0.38
0.17
0.14
0.15
0.18
0pa
ucianu
latum
0.18
0.35
0.16
0.20
0.35
0.35
0.15
0.16
0.14
0.22
0.23
0tomasii
0.14
0.34
0.08
0.18
0.35
0.35
0.08
0.06
0.07
0.12
0.14
0.16
0theo
dori
0.14
0.34
0.12
0.21
0.37
0.36
0.15
0.11
0.14
0.11
0.12
0.19
0.10
0.01
cf.the
rmae
um0.15
0.38
0.15
0.21
0.36
0.36
0.14
0.12
0.14
0.14
0.07
0.19
0.12
0.10
NA
opercu
latum
0.41
0.45
0.41
0.44
0.44
0.43
0.41
0.39
0.41
0.42
0.44
0.41
0.42
0.42
0.43
0.05
steinii
0.29
0.28
0.27
0.32
0.30
0.30
0.28
0.26
0.27
0.32
0.30
0.32
0.27
0.29
0.31
0.42
0.02
trulla
0.15
0.35
0.16
0.20
0.37
0.37
0.15
0.18
0.16
0.19
0.22
0.18
0.15
0.17
0.20
0.41
0.30
0ou
tgroup
0.38
0.34
0.35
0.38
0.32
0.33
0.36
0.38
0.37
0.39
0.39
0.39
0.37
0.39
0.37
0.44
0.28
0.38
0.09
incoloratum
0.33
0.37
0.32
0.34
0.37
0.37
0.32
0.33
0.34
0.38
0.33
0.34
0.33
0.34
0.33
0.39
0.30
0.34
0.30
NA
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 143
respectively, and the remaining species of Amphidinium resulted invalues ranging from 0 to 0.02. As would be expected by branchlengths in the phylogeny, the largest interspecific distances werebetween A. operculatum and the other species of Amphidinium(0.40-0.45). Those species in the Herdmanii Clade ranged from0.27–0.39 genetic distance from any Amphidinium species in theOperculatum Clade. The remaining distances between the speciesof the Operculatum Clade (minus A. operculatum itself) rangedbetween 0.06-0.23.
3.3. ITS2 secondary structure phylogeny and CBC analysis
The phylogeny displayed in Fig. 10 is based on the secondarystructure of the ITS2 region of the ribosomal DNA of 59 strains fromthe Amphidinium carterae species complex, as well as Karlodiniumand Heterocapsa as outgroups. The ITS2 secondary structures ofAmphidinium comprised of four common helices, with the longesthelix III, a universal motif of 5.8S and 28S proximal stem, U–Umismatches at helix II, and AAA motif and UGGU at apex of thehelix III.
Strong monophyletic support for the Amphidinium speciespreviously established and those presented in this study rangedfrom 90 to 100%. Only A. thermaeum, A. theodori, and A.pseudomassartii did not show monophyletic support due to smallsample sizes. Two clades were formed in the phylogeny: the firstclade clustered A. tomasii, A. magnum, A. cf. thermaeum, A.psuedomassartii, A. massartii, and A. gibbosum together, andAmphidinium carterae, A. paucianulatum, and A. fijiensis comprisedthe second.
The first clade grouped together with weak support, but withinthat Amphidinium cf. thermaeum, A. theodori, and A. pseudomas-sartii did group together with support of 94%. Strains that aremorphologically referred to as Amphidinium massartii were placedinto two clades with one subclade including two strains fromFlorida, one strain from Palmyra Atoll in the Pacific Ocean, and onestrain collected from the Gulf Stream off the coast of North Carolinathat form a monophyly with 100% bootstrap support. The othersubclade of A. massartii included a strain from Fiji, a strain fromAustralia, three strains from Korea, and one strain from Oman (99%BS) and is sister taxa (98% BS) to three A. gibbosum strains.
In the second major clade within the A. carterae speciescomplex, A. carterae emerged as sister to A. paucianulatum and A.fijiensis, which were supported as sister taxa to one another with93% bootstrap value. Well-supported diversity was found within A.carterae, with four subclades emerging, each with bootstrapsupport values of 100% and unique ITS2 transcripts (Fig. 11)between which 1–3 CBCs existed. All of the subclades groupedtogether monophyletically with a bootstrap of 100%.
Tabular CBC and Hemi-CBC data for all taxa is available uponrequest. Figs. 12–14 shows the ITS2 transcripts of A. tomasii vs. A.magnum, A. theodori vs. A. psuedomassartii, and A. paucianulatumvs. A. fijiensis, respectively, with their CBC/Hemi-CBC comparisons.A comparison of Amphidinium tomasii and A. magnum (five and fourstrains, respectively) revealed an ITS2 sequence lengths of 217 and189 bp, respectively, with a CBC and a Hemi-CBC present in helix Iand two CBCs present in helix IV. A comparison of the ITS2transcript from A. theodori and A. pseudomassartii revealed asequence lengths of 192 and 224 bp, respectively, with eight CBCs(located at helices I, II, and III, respectively) and seven Hemi-CBCs(located at helices I, II, and III, respectively. Comparing A.puacianulatum and A. fijiensis (179 and 190 bp, respectively)revealed one CBC in helix II and three in helix III as well as threeHemi-CBCs in helix III and one in helix IV.
Conflicting CBC data resulted between strains of A. pseudo-massartii and the clade containing A. cf. massartii and A. gibbosum.Zero to one CBC was present between A. massartii and A. cf.
Fig. 11. Representative ITS2 transcripts from each of the four subclades of Amphidinium carterae. A = subclade I, B = subclade II, C = subclade III, and D = subclade IV.
144 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
massartii as well as A. massartii and A. gibbosum and three Hemi-CBCs were present between A. massartii and A. gibbosum,suggesting, if only considering these data, that some strains
Fig. 12. ITS2 transcript of Amphidinium tomasii compared to Amphidinium magnum. Thstructure are deviations of the sequence found in A. magnum. CBCs and Hemi-CBCs are
between these clades were the same species while others were not.Three to four CBCs existed between A. cf. massartii and the clade ofA. gibbosum (Fig. 15), though the secondary structure tree suggests
e folded figure represents A. tomasii and the nucleotides placed outside the folded indicated with solid and dotted boxes, respectively.
Fig. 13. ITS2 transcript of Amphidinium theodori compared to Amphidinium pseudomassartii. The folded figure represents A. theodori and the nucleotides placed outside thefolded structure are deviations of the sequence found in A. pseudomassartii. CBCs and Hemi-CBCs are indicated with solid and dotted boxes, respectively.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 145
these are closely related taxa. The clade of A. massartii had variableCBC/Hemi-CBC results that, if only considering CBC data, suggestadditional taxonomic structure.
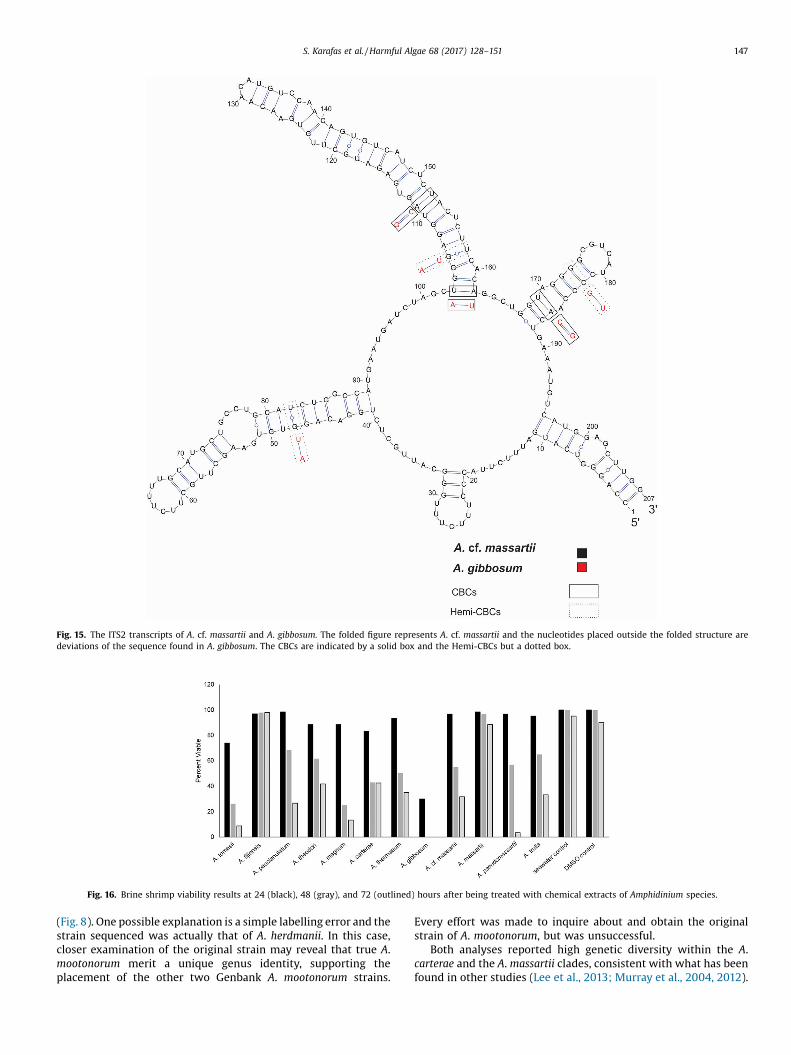
3.4. Toxicity
Data from the brine shrimp toxicity study was normalized andis shown in Fig. 16 and in tabular form in Table 2. The data areshown as pooled, though for logistical reasons not all tests wereperformed simultaneously. All species with the exception of A.fijiensis and A. massartii showed toxicity (Table 2). The mosttoxicity resulted from extractions of A. gibbosum where there wassubstantial cell death after 24 h and complete cell death after 48 h.Strains of Amphidinium tomasii, A. theodori, A. magnum, and A.carterae displayed the most initial cell death, aside from A.gibbosum, and each had less than 90% cell viability after 24 h, withA. tomasii showing the most initial toxicity among these with only74% cell viability at 24 h. The largest change in brine shrimpviability between 24 and 48 h resulted from brine shrimp treatedwith extracts of A. magnum with a 63% decrease. The remainingspecies showed a 27% � 48% decrease in brine shrimp viabilitybetween 24 and 48 h. From 48 � 72 h the largest decrease in brineshrimp viability was seen in those tests treated with extractions ofA. paucianulatum and A. pseudomassartii (42% and 53%, respective-ly). The extract from Amphidinium carterae only showed a decreaseof 1% between these time points, and the remaining speciesdisplayed a brine shrimp viability decrease of 12–23% between 48
and 72 h. The largest overall decreases in brine shrimp viabilitycould be found in those tests treated with A. magnum, A. tomasii,and A. pseudomassartii, where after 72 h only 13%, 9%, and 3% ofbrine shrimp were viable. The remaining species had 27–43% ofbrine shrimp still viable after 72 h.
4. Discussion
The results from the morphometrics, phylogenetics, secondarystructure and CBC data, and toxicity data agreed sufficiently tosupport the taxonomic distinctions described. The few cases,which will be discussed further, where the four lines of evidenceare not in complete agreement could be due, in part, to sample sizediscrepancies. For example, despite multiple attempts, the ITS2region of Amphidinium trulla did not successfully amplify andcomplete sequences for that region are limited on publicdatabases, possibly affecting bootstrap support values and thetopology of the secondary structure phylogeny. Instances such asthose highlight the importance of using multiple lines of evidencein supporting the introduction of novel taxa designations. In thatway general patterns begin to emerge even in the midst oftopological disagreements. For example, though the large subunitsequence data and ITS2 secondary structure data do not agree onthe terminal relationships between A. tomasii, A. magnum, A.thermaeum, A. theodori, and A. pseudomassartii, these five speciesalways cluster together.
Fig. 14. ITS2 transcript of Amphidinium paucianulatum compared to Amphidinium fijiensis. The folded figure represents A. paucianulatum and the nucleotides placed outsidethe folded structure are deviations of the sequence found in A. fijiensis. CBCs and Hemi-CBCs are indicated with solid and dotted boxes, respectively.
146 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
4.1. Scales
To date, body scales were reported in three clades ofAmphidinium, A. cupulatisquama and both clades of A. massartii(A. massartii and A. cf. massartii) (Lee et al., 2013; Murray et al.,2012; Sekida et al., 2003; Tamura et al., 2009). In A. cupulatisquamathe scales are cup-shaped with distinct three-dimensionality andin A. massartii and A. cf. massartii they are doughnut-shaped andslightly concave rings. In both studies the scales were present onthe cell surface outside the plasma membrane and amphiesmalplates. The present study further confirmed the existence of scalesin both A. massartii and A. cf. massartii, and additionally introducesthe presence of scales in two newly identified species A.paucianulatum and A. theodori (Table 2). Viewed on scanningelectron micrographs, the scales in both species appeared roughlythe size and shape as those originally described for A. massartii(AB477347) by Sekida et al. (2003). Due to their small size at themagnification limits of the scope, however, exact measurementscould not be inferred, nor could an unequivocal judgement bemade on their 3-dimensionality. Furthermore, SEM is a useful toolfor determining that scales are present, but cannot be used toconfirm that scales are not present if they are not observed, due tothe positioning of the scales on top of the plasma membrane andamphiesmal plates (Sekida et al., 2003). It is not always obvious in apreparation that the plates are fixed in place, and if the plates arestripped away then the scales are too. Even when it is clear thatplates are intact on the cells, one cannot be guaranteed that theplasma membrane has been fixed properly, despite the assumptionthat a good osmium preparation immediately fixes the plasmamembrane. Transmission electron microscopy is currently the bestway to deny the presence of scales on cells with certainty, which
went beyond the scope of the present study. Instead, research iscurrently underway to describe the TEM results for eachAmphidinium clade as part of a future publication. Therefore, withrespect to those strains where scales did not appear to be presentthe best that can be stated in this study is that scales were notobserved.
4.2. Species comparisons
It is odd that Amphidinium herdmanii and Amphidiniummootonorum are reported to be so morphologically distinct butlack any significant molecular distinction (p-dist = 0.004). Mor-phologically these two species are distinguishable by size, shape,and subcellular features such as nucleus position. The speciesAmphidinium mootonorum was described based on morphology(Murray and Patterson, 2002) and later its genetic sequence wasincluded in the revision of Amphidinium by Flø Jørgensen et al.,2004a, and likely the two studies used the same strain. In everyother case, taxonomic units of Amphidinium were well distin-guished molecularly, especially when unique morphologicalcharacters are present. The peculiarity of these two speciesclustering together was noted by Flø Jørgensen et al., 2004a, butnot expanded on in any way. More recently two other A.mootonorum strains have been deposited on Genbank (KT371435and GU295205) that do not even cluster within the Amphidiniumgenus, but instead as sister taxa to Ankistrodinium. These twostrains were not included in the presently reported phylogeny in aneffort to not confuse the study focus. This study is centered on theresolution of species identities within the Amphidinium opercula-tum species complex, specifically. It is important to briefly mentionthe issue because the strains are included in the larger phylogeny
Fig. 15. The ITS2 transcripts of A. cf. massartii and A. gibbosum. The folded figure represents A. cf. massartii and the nucleotides placed outside the folded structure aredeviations of the sequence found in A. gibbosum. The CBCs are indicated by a solid box and the Hemi-CBCs but a dotted box.
Fig. 16. Brine shrimp viability results at 24 (black), 48 (gray), and 72 (outlined) hours after being treated with chemical extracts of Amphidinium species.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 147
(Fig. 8). One possible explanation is a simple labelling error and thestrain sequenced was actually that of A. herdmanii. In this case,closer examination of the original strain may reveal that true A.mootonorum merit a unique genus identity, supporting theplacement of the other two Genbank A. mootonorum strains.
Every effort was made to inquire about and obtain the originalstrain of A. mootonorum, but was unsuccessful.
Both analyses reported high genetic diversity within the A.carterae and the A. massartii clades, consistent with what has beenfound in other studies (Lee et al., 2013; Murray et al., 2004, 2012).
148 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
In the case of Amphidinium carterae, those studies describedmultiple clades, and the present study reports at least four thatform a monophyly with strong support in both phylogenies.Initially it appeared that five subclades were present in these data,and in part that is true, however, the secondary structure dataoffered more resolution, though subclade III of the secondarystructure phylogeny cannot be directly compared to the sequencephylogeny. There is only one secondary structure transcript thatrepresents both sister clades of subclade I in Fig. 10, see Fig. 11,indicating that it is appropriate to consider those two sister cladestogether. The CBC data further indicates that these subclades aredistinct. One compensatory base change is considered sufficient toidentify species level diversity and has been used successfully to dojust that in many species including phytoplankton species andother benthic harmful algal species (Ahvenniemi et al., 2009;Amato et al., 2007; Coleman, 2000, 2009; Coleman and Vacquier,2002; Leaw et al., 2010, 2016; Lim et al., 2013; Müller et al., 2007;Ruhl et al., 2010; Schill et al., 2010; Sorhannus et al., 2010; Tenget al., 2016; Wolf et al., 2013). Morphological characters unique tothese subclades, if any, remain elusive and their discovery wasbeyond the scope of this study. The toxicity data offered for thisspecies represents only subclade I, strain Amca0406-1, whichshowed initial toxicity after 24 h, but a plateau in effects between48 and 72 h. Each individual subclade was not tested. Theindividual tested in this study falls into the same secondarystructure subclade as strain CS-740 which was reported by Murrayet al. (2012) to contain partial ketosynthase sequences. Rhodeset al. (2010) also report a strain of A. carterae that was fatally toxicin mice, CAWD 57. The strain fell out in a subclade unique to thosealready reported for the species (Clades 1, 2, & 3) but genetic datawas not made publicly available and cannot be compared. Thesetwo studies are examples of a changing trend to identify thesubclade from which A. carterae samples are being tested fortoxicity. This is important because toxicity, both presence anddegree, is variable between the subclades (Baig et al., 2006; Rhodeset al., 2010; Murray et al., 2012). Taken as a whole, this studysuggests that there is enough evidence to warrant furtherexamination of the intraspecific variation of A. carterae and thateach of the four subclades identified by the ITS2 secondarystructure transcripts may represent a unique species, however atthis time elevation of any is premature and all should be referred asa single species. Therefore, within the context of this study, theintraspecific variation found in A. carterae is the largest that can beassigned to one taxon and was used as a standard to delineateacceptable genetic diversity within a single species (Table 3).
In the past Amphidinium pseudomassartii was either classified asAmphidinium massartii or referred to as A. cf. massartii, acknowl-edging the distinct molecular differentiation of the group fromtraditional A. massartii (Lee et al., 2013; Murray et al., 2004, 2012).Through the addition of more Amphidinium strains, this studydemonstrated with both ribosomal sequence data and ITS2secondary structure data that strains of A. pseudomassartii areeven more genetically distinct from A. massartii than originallyproposed, and that they likely evolved somewhere in theAmphidinium thermaeum species complex. Therefore, to refer tothem as A. cf. massartii is erroneous, but until now researchers havebeen hesitant to identify strains of A. pseudomassartii as their owntaxonomic unit because of the lack of defining morphologicalcharacters. While strains of A. massartii examined with SEM readilyshowed the presence of ring-like surface scales (Lee et al., 2013;Murray et al., 2012; Sekida et al., 2003), scales were not observedon the strain of A. pseudomassartii examined. It is very likely thatthe lack of observable scales on seemingly well prepared scanningelectron micrographs may be the defining morphological characterthat has been lacking. As noted earlier, however, this charactercannot be verified unequivocally until the TEM work is completed.
Despite this, the strengthened phylogenetic support in theribosomal sequence data and the additional support offered inthe secondary structure phylogeny justify the elevation of thesestrains to a unique taxonomic species. Furthermore, chemicalextractions from cells of A. pseudomassartii were cytotoxic toreplicates of brine shrimp resulting in 97% cell death after 72 hcompared to 12% in A. massartii.
In the case of strains that were traditionally identified asAmphidinium massartii, the results reported in this study mirrorthose presented by Lee et al. (2013). Both studies reported twowell-supported subclades that possessed additional structurewithin. While the genetic structure is similar, the present resultsshowed no support for the monophyly of the species, as there is nosignificant bootstrap value in the sequence data and the secondarystructure phylogeny includes strains of A. gibbosum nestedbetween the two subclades of A. massartii. The genetic diversitybetween the two clades of A. massartii (if considered together) isalso higher than the standard set in this study by A. carterae (0.05).This likely implies that what was to date regarded as Amphidiniummassartii was, in fact, two species. Strains were available forcomparison that represented each subclade but no uniquemorphological traits could be observed that would supportseparating the species. Scales with presumably the samemorphology were definitively observed in this study and others(Lee et al., 2013; Murray et al., 2012) in strains representing bothclades. A t-test showed a significant difference between thetoxicity of Clade 1 and Clade 2 (p = 0.002) after 48 h. Extracts of A.massartii Clade 1 were not found to be especially cytotoxic to brineshrimp, killing only 12% after 72 h whereas Clade 2 was toxic,demonstrating 68% Artemis mortality after 72 h. Clade 1 resultswere consistent with what has been observed previously for A.massartii (Lee et al., 2013; Murray et al., 2012), but the results fromClade 2 suggest that there are ecological differences that existsbetween the two groups. Examination into the morphologicalfeatures of Clade 1 and 2 continues, but at this point it is prematureto identify either group as an entirely unique taxa. Instead, it isrecommended to refer to Clade 1 as Amphidinium massartii andClade 2 as Amphidinium cf. massartii. The original strain collectedfrom Biecheler from Sete, France in 1952 (Biecheler, 1952) nolonger exists in culture. Neither does any other verifiable A.massartii (Farmer and Roberts, 1989) from the same location. In theabsence of a substitute from the same location, the oldest verifiablestrain, CCMP 1342 collected in 1991 from Knight Key, Florida, USA,was chosen to represent as A. massartii proper. This strain evolvedin Clade 1, leaving Clade 2 representing A. cf. massartii, which willherein be referred to as such.
The present study agrees with that of Murray et al. (2004)determining that A. carterae can be distinguished from A. massartii,A. cf. massartii, and A. pseudomassartii by absence of observablescales, reported in ultrastructure studies of A. carterae (Farmer andRoberts, 1989; Roberts et al., 1988), the presence of cytotoxicity, asingle plastid superficially located, and a narrow epicone. Whilethe latter trait is qualitatively evident to the practiced eye, it can bedifficult to quantify because it requires that substantial cells arecorrectly oriented in electron microscopy. This study made verypreliminary comparisons of the epicone widths of A. carterae, A.massartii, A. cf. massartii, and A. pseudomassartii. Sample sizes wereextremely limited (5, 9, 2, and 6, respectively), but are interestingand suggest this may be a character in which more examination ismerited. The four species mentioned had means and standarddeviations of 1.5 � 0.5 mm, 2.1 �0.3 mm, 2.5 � 0 mm, and4.0 � 0.8 mm, respectively. While this trait exhibited much overlap,particularly between A. carterae, A. massartii, and A. cf. massartii,epicone width may turn out to be a useful character to differentiateA. pseudomassartii from the others, particularly if other informa-tion is lacking.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 149
The correct position of Amphidinium gibbosum is inconclusivefrom this study. Morphologically they are quite distinct from otherAmphidinium in size and shape, but genetically the LSU sequencedata and the secondary structure disagree. Both molecularanalyses suggested the species is in fact taxonomically uniquebut the tests do not agree as to its position. The sequence data didnot show any support for the position in which A. gibbosum wasdisplayed. On the other hand, the secondary structure phylogenyoffered support, but its position contradicts morphologicalobservations. As an exception, some pairwise comparisons withinthe CBC data conflicted as well. The data indicated that CBCs werenot present between any A. gibbosum strains and CCMP 1342,Amma0607-1, or Amma1506-1 but there was 1 CBC presentbetween A. gibbosum and the remaining strains of A. massartii andA. cf. massartii. These inconsistencies may be due to the highdiversity of A. massartii ITS2 region. That being said, it cannot beruled out completely that A. gibbosum is a legitimate sister taxa toA. massartii, thus implying that A. cf. massartii should represent itsown unique species.
There is little morphological evidence distinguishing strainAmth0000600 from Amphidinium thermaeum (Table 2), and theevolutionary divergence within A. thermaeum, with Amth0000600included, is lower than within A. carterae (Table 3). Cells from thepresent study were mostly round, whereas the original descrip-tions suggests that the majority of cells were oval or elliptical. Itwas suggested, however, that in the case of A. thermaeum cell shapeis a product of culture age and that spherical cells are products ofearly exponential growth. The present study did not observe anypusules in cells of Amth0000600 under light microscopy, whereasDolapsakis and Economou-Amilli (2009) reported two. Dolapsakisand Economou-Amilli (2009) reported variations within theirclone of A. thermaeum where the pusules were not always visible.Finally, the present study did not observe any metabolicmovement, but this characteristic is one that may only occurduring certain growth stages or be dependent on other variablessuch as geography. Murray et al. (2012) also report not observingmetabolic movement in a strain of A. thermaeum examined. StrainAmth0000600 is a clone that was established and housed at theAlgal Resources Collection and originally investigated using lightand epifluorescent microscopy by a former graduate student(Cyronak, 2007). In subsequent years genetic material wascollected and is still being maintained, but the strain itself,unfortunately, was unable to be maintained as part of the livingcollection beyond 2013. Though little evidence exists to considerthe strain of Amth0000600 to be something other than A.thermaeum, it is preferable to refer to strain Amth0000600 as A.cf. thermaeum because strain material no longer exists for moredetailed morphological examinations. Therefore until more taxathat identify with this strain genetically are available forexamination, the conservative approach is most appropriate.
The database Algaebase (Guiry and Guiry, 1996) has worked toresolve many issues of synonymy within the Amphidinium genus,but one of the largest sources of confusion, Amphidinium klebsiiKofoid and Swezy is still considered taxonomically valid. Thisorganism was originally described by Klebs (1884) and laterelevated to a species based on that description by Kofoid andSwezy (1921). In short, this species displayed the typicalAmphidinium morphology and had a slightly dorsoventrallyflattened hypocone with a broad, round antapex and asymmetricanterior shoulders, the left one being narrower than the right. Theexact location of the longitudinal flagellum is not indicated clearly,but it is implied to be between the middle and anterior of the cell.Multiple chromatophores are described. The original description ofthis organism is quite generic given what is now known about thegenus, not introducing any defining characters except one � thepresence of furrows (Klebs, 1884) which can be difficult to see and
was not verified in any other organism until now. As a result, itbecame a sort of catch-all for Amphidinium, with the species, at onetime or another, being synonymous with strains from A.operculatum, A. gibbosum, A. massartii, and A. carterae (Murrayet al., 2004). Even now, the genetic data for the LSU region of two A.klebsii that are on Genbank Database actually represent A. massartiiand A. gibbosum. It remains unclear if Klebs (1884) had in hispossession a valid new species that simply has not since beencollected, or if he was examining one of the other strains withwhich A. klebsii is often confused. The presence of furrows wasobserved in two different strains of Amphidinium tomasii(Amto1412-1, Amto1412-2). In both cases the cells were in theprocess of cell division, and no furrows were observed in cells post-division. A similar phenomena was observed in cells of Togulabritannica, a species formerly considered an Amphidinium, where asingle groove formed on the ventral side of motile cells only priorto cell division (Jørgensen et al., 2004). As far as can be determined,this is the first time that furrows of any kind have been noted in theAmphidinium sensu stricto since the description of A. klebsii. At thispoint A. tomasii and A. klebsii are not considered synonyms. The cellsizes differ greatly, averaging twice as large in A. klebsii as in A.tomasii for length and width. Furthermore, the chromatophoresreportedly radiate from the posterior of the cell in A. klebsii, whereas a single plastid centrally radiates in A. tomasii. Because thefurrows were not observed in any un-dividing cell it is likely thattheir formation is a temporary artifact of the division process. Alsolikely is that the artifact could be present in other Amphidiniumspecies but they must be observed at a specific point in the lifecycle, which has not yet been done. Further study into this issue iswarranted. Given the lack of defining characters unique to A. klebsiiand the erroneously positioned, according to Kofoid and Swezy(1921), transverse flagellum in the original description, Klebs(1884), it is recommended to formally eliminate A. klebsii as anaccepted taxonomic unit.
5. Conclusion
Multiple lines of evidence including LSU sequence data, ITS2secondary structure, CBC data, and toxicity provided sufficientevidence to justify establishing six new species within theAmphidinium operculatum species complex: A. tomasii, A. magnum,A. theodori, A. pseudomassartii, A. paucianulatum, and A. fijiensis.The same lines of evidence were used to split A. massartii into A.massartii and A. cf. massartii as well as assign one strain to A. cf.thermaeum. Two of these species were found to not have cytotoxiceffects, A. massartii and A. fijiensis, while the remaining speciesshowed varying degrees of toxicity, though the composition,mechanism, and contribution to CFP of the toxins remains unclear.
Acknowledgements
Thanks go to Tyler Cyronak who prompted interest in thisproject with his thesis work many years ago. Thank you to themany researchers who collected seawater samples and contribut-ed to building such a vast Amphidinium collection at ARC, inparticular Dr. A. N’Yeurt from the University of the South Pacific,Fiji. Thanks go to the scientists in the Wright Lab at the MarbioNCbuilding who performed chemical extractions for the cytotoxicitystudies, and also to Dr. Taylor and the EM lab for assistance with theSEM. A special thank you goes to the staff at the National Center forMarine Algae and Microbiota who donated 13 Amphidinium strainsto the ARC particularly for this project. This research did not receiveany specific grant from funding agencies in the public, commercial,or not-for-profit sectors. Most importantly, exceptional acknowl-edgment and gratitude is extended to Dr. Carmelo Tomas who, as
150 S. Karafas et al. / Harmful Algae 68 (2017) 128–151
former Director of the Algal Resources Collection, was monumen-tal in the completion of this project. He not only supported thisproject fiscally, but also served as a mentor, a reference, and asounding board.[SS]
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.hal.2017.08.001.
References
Ahvenniemi, P., Wolf, M., Lehtonen, M.J., Wilson, P., German-Kinnari, M., 2009.Evolutionary diversification indicated by compensatory base changes in ITS2secondary structures in a complex fungal species, Rhizoctonia solani. J. Mol. Evol.69, 150–163.
Amato, A., Kooistra, W.H.C.F., Ghiron, J.H.L., Mann, D.G., Pröschold, T., Montresor, M.,2007. Reproductive isolation among sympatric cryptic species in marinediatoms. Protist 158 (2), 193–207.
Baig, H.S., Saifullah, S.M., Dar, A., 2006. Occurrence and toxicity of Amphidiniumcarterae Hulburt in the North Arabian Sea. Harmful Algae 5 (2), 133–140.
Biecheler, B., 1952. Recherches sur les peridiniens. Bull. Biol. Fr. Belg. 36, 1–49.Coleman, A.W., Vacquier, V.D., 2002. Exploring the phylogenetic utility of ITS
sequences for animals: a test case for abalone (Haliotis). J. Mol. Evol. 54, 246–257.
Coleman, A.W., 2000. The significance of a coincidence between evolutionarylandmarks found in mating affinity and a DNA sequence. Protist 151, 1–9.
Coleman, A.W., 2009. Is there a molecular key to the level of biological species ineukaryotes? A DNA guide. Mol. Phylogen. Evol. 50, 197–203.
Cyronak, T.J., 2007. A Comparative Morphological, Phylogenetic, and AutecologicalStudy of New Amphidinium Species of Dinoflagellates, Biology and MarineBiology Master’s of Science Thesis. University of North Carolina, Wilmington,pp. 56.
Daugbjerg, N., Hansen, G., Larsen, J., Moestrup, Ø., 2000. Phylogeny of some of themajor genera of dinoflagellates based on ultrastructure and partial LSU rDNAsequence data, including the erection of three new genera of unarmoureddinoflagellates. Phycologia 39, 302–317.
Dodge, J.D., 1982. Marine Dinoflagellates of the British Isles. Her Majesty'sStationery Office, London, U.K.
Dolapsakis, N.P., Economou-Amilli, A., 2009. A new marine species of Amphidinium(Dinophyceae) from Thermaikos Gulf, Greece. Acta Protozool. 48, 153–170.
Echigoya, R., Rhodes, L.L., Oshima, Y., Satake, M., 2005. The structures of five newantifungal and hemolytic amphidinol analogs from Amphidinium carteraecollected in New Zealand. Harmful Algae 4 (2), 383–389.
Farmer, M.A., Roberts, K.R.,1989. Comparative analyses of the dinoflagellate flagellarapparatus. 3. Freeze substitutuion of Amphidinium rhynchocephalum. J. Phycol.25, 280–292.
Flø Jørgensen, M., Murray, S., Daugbjerg, N., 2004a. Amphidinium revisited I:Redefinition of Amphidinium (Dinophyceae) based on cladistic and molecularphylogenetic analysis. J. Phycol. 40, 351–365.
Flø Jørgensen, M., Murray, S., Daugbjerg, N., 2004b. A new genus of athecateinterstitial dinoflagellates, Togula gen. nov., previously encompassed withinAmphidinium sensu lato: inferred from light and electron microscopy andphylogenetic analyses of partial large subunit ribosomal DNA sequences. Phycol.Res. 52 (3), 284–299.
Guiry, M.D., Guiry, G.M., 1996. AlgaeBase. World-wide electronic publication,National University of Ireland, Galway.
Hikosaka-Katayama, T., Koike, K., Yamashita, H., Hikosaka, A., Kazuhiko, K., 2012.Mechanisms of maternal inheritance of dinoflagellate symbionts in acoelo-morph worm Waminoa litus. Zool. Sci. 29 (9), 559–567.
Hoppenrath, M., Leander, B.S., 2010. Dinoflagellate phylogeny as inferred from heatshock protein 90 and ribosomal gene sequences. PLoS One 5 (10), 1–12.
Hoppenrath, M., Murray, S., Sparmann, S.F., Leander, B.S., 2012. Morphology andmolecular phylogeny of Ankistrodinium gen. nov. (Dinophyceae): a new genus ofmarine sand-dwelling dinoflagellates formerly classified within Amphidinium. J.Phycol. 48, 1143–1152.
Horiguchi, T., Tamura, M., Katsumata, K., Yamaguchi, A., 2012. Testudodinium gen.nov. (Dinophyceae), a new genus of sand-dwelling dinoflagellates formerlyclassified in the genus Amphidinium. Phycol. Res. 60 (2), 137–149.
Hsiao, C., Chatterton, N.J., Asay, K.H., Jensen, K.B., 1995. Phylogenetic relationships ofthe monogenomic species of the wheat tribe, Triticeae (Poaceae) inferred fromnuclear rDNA (internal transcribed spacer) sequences. Genome 38, 211–223.
Klebs, G., 1884. Ein kleiner Beitrag zur kenntnis der Peridineen. BotanischeZeitschrift 42, 737–752.
Kobayashi, J., Kubota, T., 2007. Bioactive macrolides and polyketides from marinedinoflagellates of the genus Amphidinium. J. Nat. Prod. 70, 451–460.
Kobayashi, J., Shigemori, H., Ishibashi, M., Yamasu, T., Hirota, H., Sasaki, T., 1991.Amphidinolides G and H: new potent cytotoxic macrolides from the culturedsymbiotic dinoflagellate Amphidinium sp. J. Org. Chem. 56, 5221–5224.
Kobayashi, J.i., 2008. Amphidinolides and its related macrolides from marinedinoflagellates. J. Antibiot. 61 (5), 271–284.
Koetschan, C., Förster, F., Keller, A., Schleicher, T., Ruderisch, B., Schwarz, R., Müller,T., Wolf, M., Schultz, J., 2010. The ITS2 database III—sequences and structures forphylogeny. Nucleic Acids Res. 38 (suppl 1), D275–D279.
Kofoid, C.A., Swezy, O.,1921. The Free-living Unarmored Dinoflagellumta. Universityof California Press Berkeley, California.
Larsen, J., Patterson, D.J., 1990. Some flagellates (Protista) from tropical marinesediments. J. Nat. Hist. 24, 801–937.
Larsen, J., 1985. Algal studies of the Danish Wadden Sea II: A taxonomic study ofpsammobious dinoflagellates. Opera Bot. 79, 14–37.
Leaw, C.P., Lim, P.T., Cheng, K.-W., Ng, B.-K., Usup, G., 2010. Morphology andmolecular characterization of a new species of thecate benthic dinoflagellate,Coolia malayensis sp. nov. (Dinophyceae). J. Phycol. 46 (1), 162–171.
Leaw, C.P., Tan, T.H., Lim, H.C., Teng, S.T., Yong, H.L., Smith, K.F., Rhodes, L.L., Wolf, M.,Holland, W.C., Vandersea, M.W., Litaker, R.W., Tester, P.A., Gu, H., Usup, G., Lim, P.T., 2016. New scenario for speciation in the benthic dinoflagellate genus Coolia(Dinophyceae). Harmful Algae 55, 137–149.
Lee, K.H., Jeong, H.J., Park, K., Kang, N.S., Yoo, Y.D., Lee, M.J., Lee, J., Lee, S., Kim, T., Kim,H.S., Noh, J.H., 2013. Morphology and molecular characterization of theepiphytic dinoflagellate Amphidinium massartii isolated from the temperatewaters off Jeju Island, Korea. Algae 28 (3), 213–231.
Lim, H.C., Teng, S.T., Leaw, C.P., Lim, P.T., 2013. Three novel species in the Pseudo-nitzschia pseudodelicatissima complex: P. batesiana sp. nov., P. lundholmiae sp.nov., and P. fukuyoi sp. nov. (Bacillariophyceae) from the Strait of Malacca,Malaysia. J. Phycol. 49 (5), 902–916.
Meng, Y., Van Wagoner, R.M., Misner, I., Tomas, C., Wright, J.L.C., 2010. Structure andbiosynthesis of amphidinol 17, a hemolytic compound from Amphidiniumcarterae. J. Nat. Prod. 73, 409–415.
Murray, S., Patterson, D.J., 2002. The benthic dinoflagellate genus Amphidinium insouth-eastern Australian waters, including three new species. Eur. J. Phycol. 37,279–298.
Murray, S., Flø Jørgensen, M., Daugbjerg, N., 2004. Amphidinium revisited IIResolving species boundaries in the Amphidinium operculatum species complex(Dinophyceae), including the descriptions of Amphidinium trullasp. nov. andAmphidinium gibbosum comb. nov. J. Phycol. 40, 366–382.
Murray, S.A., Garby, T., Hoppenrath, M., Neilan, B.A., 2012. Genetic diversity,morphological uniformity and polyketide production in dinoflagellates (Amphidinium, Dinoflagellumta). PLoS One 7 (6), e38253.
Nuzzo, G., Cutignano, A., Sardo, A., Fontanta, A., 2014. Antifundal amphidinol 18 andits 7-sulfate derivative from the marine dinoflagellate Amphidinium carterae.Natural Products 77 (6), 1524–1527.
Onuma, R., Watanabe, K., Horiguchi, T., 2015. Pellucidodinium psammophilum gen. &sp. nov. and Nusuttodinium desymbiontumsp. nov. (Dinophyceae), two novelheterotrophs closely related to kleptochloroplastidic dinoflagellates. Phycologia54 (2), 192–209.
Owari, S., Hayashi, A., Ishida, K.-i., 2014. Subcellular localization of minicircle DNA inthe dinoflagellate Amphidinium massartii. Phycol. Res. 62, 1–8.
Rhodes, L.L., Smith, K., Harwood, T., Bedford, C., 2014. Novel and toxin-producingepiphytic dinoflagellates isolated from sub-tropical Raoul Island, KermadecIslands group. N. Z. J. Mar. Freshwater Res. 48 (4), 594–599.
Richlen, M.L., Barber, P.H., 2005. A technique for the rapid extraction of microalgalDNA from single live and preserved cells. Mol. Ecol. Notes 5, 688–691.
Roberts, K.R., Farmer, M.A., Schneider, R.M., Lemoine, J.E., 1988. The microtubularcytoskeleton of Amphidinium rhynchocephalum (Dinophyceae). J. Phycol. 24(544-553) .
Ruhl, M., Wolf, M., Jenkins, T.M., 2010. Compensatory base changes illuminatetaxonomically difficult taxonomy. Mol. Phylogen. Evol. 54 (2), 664–669.
Satake, M., Murata, M., Yasumoto, T., Fujita, T., Naoki, H., 1991. Amphidinol, apolyhydroxypolyene antifungal angent with an unprecedented structure, froma marine dinoflagellate, Amphidinium klebsii. J. Am. Chem. Soc. 113, 9859–9861.
Schill, R.O., Förster, F., Dandekar, T., Wolf, M., 2010. Using compensatory base changeanalysis of internal transcribed spacer 2 secondary structures to identify threenew species in Paramacrobiotus (Tardigrada). Organ. Divers. Evol. 10 (4), 287–296.
Schliep, K.P., 2011. Phangorn: phylogenetic analysis in R. Bioinformatics 27 (4), 592–593.
Scholin, C.A., Herzog, M., Sogin, M., Anderson, D.M., 1994. Identification of group-and strain-specific genetic markers for globally distributed Alexandrium(Dinophyceae): II. Sequence analysis of a fragment of the LSU rRNA gene. J.Phycol. 30, 999–1011.
Seibel, P.N., Müller, T., Dandekar, T., Schultz, J., Wolf, M., 2006. 4SALE–a tool forsynchronous RNA sequence and secondary structure alignment and editing.BMC Bioinf. 7 (1), 498.
Seibel, P.N., Müller, T., Dandekar, T., Wolf, M., 2008. Synchronous visual analysis andediting of RNA sequence and secondary structure alignments using 4SALE. BMCRes. Notes 1 (1), 91.
Sekida, S., Okuda, K., Katsumata, K., Horiguchi, T., 2003. A novel type of body scalefound in two strains of Amphidinium species (Dinophyceae). Phycologica 42 (6),661–666.
S. Karafas et al. / Harmful Algae 68 (2017) 128–151 151
Sorhannus, U., Ortiz, J.D., Wolf, M., Fox, M.G., 2010. Microevolution and speciation inThalassiosira weissflogii (Bacillariophyta). Protist 161 (2), 237–249.
Stern, R.F., Andersen, R.A., Jameson, I., Küpper, F.C., Coffroth, M.-A., Vaulot, D., LeGall, F., Véron, B., Brand, J.J., Skelton, H., Kasai, F., Lily, E.L., Keeling, P.J., 2012.Evaluating the ribosomal internal transcribed spacer (ITS) as a candidatedinoflagellate barcode marker. PLoS One 7, e42780.
Takano, Y., Yamaguchi, H., Inouye, I., Moestrup, Ø., Horiguchi, T., 2014. Phylogeny offive species of Nusuttodinium gen nov. (Dinophyceae), a genus of unarmouredkleptoplastidic dinoflagellates. Protist 165, 759–778.
Tamura, M., Yoshihito, T., Horiguchi, T., 2009. Discovery of a novel type of body scalein the marine dinoflagellate Amphidinium cupulatisquama sp. nov (Dinophy-ceae). Phycol. Res. 57, 304–312.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5Molecular evolutionary genetics analysis using maximum likelihood, evolu-tionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739.
Teng, S.T., Tan, S.N., Lim, H.C., Dao, V.H., Bates, S.S., Leaw, C.P., 2016. High diversity ofPseudo-nitzschia along the northern coast of Sarawak (Malaysian Borneo), with
descriptions of P. bipertita sp. nov. and P. limii sp. nov. (Bacillariophyceae). J.Phycol. 52 (6), 973–989.
Tindall, D.R., Morton, S.L., 1998. Community dynamics and physiology of epiphytic/benthic dinoflagellates associated with ciguatera. In: Cembella, A.D., Halle-graeff, G.M. (Eds.), Physiological Ecology of Harmful Algal Blooms. Proceedingsof the NATO-ASI. Springer, Verlag Heidelberg.
Washida, K., 2006. Karatungiols A and B two novel antimicrobial polyol compounds,from the symbiotic marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 47,2521–2525.
Wolf, M., Achtziger, M., Schultz, J., Dandekar, T., Müller, T., 2005. Homologymodeling revealed more than 20,000 rRNA internal transcribed spacer 2 (ITS2)secondary structures. RNA 11 (11), 1616–1623.
Wolf, M., Chen, S., Song, J., Ankenbrand, M., Müller, T., 2013. Compensatory basechanges in ITS2 secondary structures correlate with the biological speciesconcept despite intragenomic variability in ITS2 sequences � a proof of concept.PLoS One 8 (6), e66726.
Wolf, M., Koetschan, C., Müller, T., 2014. ITS2, 18S, 16S or any other RNA—simplyaligning sequences and their individual secondary structures simultaneously byan automatic approach. Gene 546 (2), 145–149.