An improved protocol for rapid extraction of membrane enzymes from Gram positive bacteria Pooja Attri, Drukshakshi Jodha, Jasbir Singh and Suman Dhanda * Received 24th December 2011, Accepted 3rd May 2012 DOI: 10.1039/c2ay05931b A simple and reproducible method for extraction of membrane enzymes from Gram positive bacteria in active form has been developed. The method employs the incubation of cells with lysozyme and sonication in the presence of salt and detergent followed by repeated freezing and thawing to disrupt the lipid–protein interactions. Although developed for Dipeptidylpeptidase-III as a demonstration, this protocol has been used for the successful extraction of some synthetic substrate hydrolyzing activities from three different Gram positive bacteria with yield of more than 90% for most studied activities. This method can be routinely used for a wide range of applications and it can also be used to extract and solubilize other enzymes/proteins only by manipulating detergent, salt and their concentration. It is simple, straightforward, reproducible, cost effective and a less time consuming protocol for extracting and solubilizing membrane enzymes. Introduction Membrane associated proteins of microorganisms often present a challenge as well as opportunity, in the quest for better thera- peutic and prophylactic interactions against infectious diseases, as membrane associated proteins are important for pathogenicity of bacteria. Pediococcus acidilactici is an important microor- ganism with wide applications in the food industry. It is generally regarded as safe (GRAS) but its vancomycin resistant strains may cause blood septicemia. 1 Dipeptidylpeptidase-III (DPP-III) removes dipeptide moieties from amino-terminus of oligopep- tides. 2,3 It has been purified from different species but its func- tions are not well studied. 4 As the DPP-III is thought to be involved in enkephalin degradation with micromolar affinity 3,5 it has emerged as an important target for developing inhibitors which could act as analgesics for patients of neuropathic and cancer pain. 6 DPP-III is unexplored in microbes so, in order to understand the functional significance of DPP-III, studies were planned for purification and characterization of DPP-III from Pediococcus acidilactici, a microbe in the dairy industry. More- over it is easier to develop the knock out in microbes which help to know the particular function. Although DPP-III is a cytosolic enzyme in mammals, 7 studies in our laboratory revealed that it is located in membranes in Pediococcus acidilactici. Study of membrane proteins is one of major difficulty because of prob- lems encountered in extraction while maintaining solubility and activity. To date, several methods for extracting membrane proteins have been described. 8–13 All these have their own inherent advantages and disadvantages and none benefit from the existence of internationally recognized standards for universally applicable extraction of membrane proteins. In the present technical note, a simple, straightforward, reproducible and less time consuming protocol is reported for extracting and solubilizing membrane enzymes. As a test, this protocol has also been successfully used for extraction of aminopeptidase B, DPP-II, cathepsin C, proline iminopeptidase, Ala-Ala-bNA, Leu-Ala-bNA, Glu-Ala-bNA, Ile-bNA, Ser-Met- bNA, Phe-bNA, Trp-bNA, Val-bNA, Tyr-bNA and Ser-bNA hydrolyzing activities in three different Gram positive lactic acid bacteria (LAB) viz. Pediococcus acidilactici, Lactobaccillus plantarum, Lactobacillus brevis. Experimental Bacterial strains All the bacterial strains (Pediococcus acidilactici NCDC 252, Lactobaccillus plantarum NCDC 020 and Lactobacillus brevis NCDC 001) were purchased from National Dairy Research Institute (NDRI), Karnal, India. Chemicals and reagents All the bacteria were grown at their respective temperatures in Man Rogosa Sharpe (MRS) purchased from Himedia chemical laboratory. All the chemicals used for the experiments were of the highest possible quality. Lysozyme, sodium phosphate, SDS, Tween-20, Tween-80 and TritonX-100, ethanol, butanol, CoCl 2 , CaCl 2 , EDTA, NaCl was purchased from Himedia chemical Laboratory, Bombay. Fast Garnet GBC, Arg-4mbNA from Sigma Aldrich. Department of Biochemistry, Kurukshetra University, Kurukshetra, India. E-mail: [email protected]; Fax: +91-1744-238277; Tel: +91- 1744-238196 ext. 2751 2574 | Anal. Methods, 2012, 4, 2574–2577 This journal is ª The Royal Society of Chemistry 2012 Dynamic Article Links C < Analytical Methods Cite this: Anal. Methods, 2012, 4, 2574 www.rsc.org/methods TECHNICAL NOTE Downloaded by Beijing University on 02/05/2013 08:26:52. Published on 13 June 2012 on http://pubs.rsc.org | doi:10.1039/C2AY05931B View Article Online / Journal Homepage / Table of Contents for this issue
Transcript
Dynamic Article LinksC<AnalyticalMethods
Cite this: Anal. Methods, 2012, 4, 2574
www.rsc.org/methods TECHNICAL NOTE
Dow
nloa
ded
by B
eijin
g U
nive
rsity
on
02/0
5/20
13 0
8:26
:52.
Pu
blis
hed
on 1
3 Ju
ne 2
012
on h
ttp://
pubs
.rsc
.org
| do
i:10.
1039
/C2A
Y05
931B
View Article Online / Journal Homepage / Table of Contents for this issue
An improved protocol for rapid extraction of membrane enzymes from Grampositive bacteria
Pooja Attri, Drukshakshi Jodha, Jasbir Singh and Suman Dhanda*
Received 24th December 2011, Accepted 3rd May 2012
DOI: 10.1039/c2ay05931b
A simple and reproducible method for extraction of membrane enzymes fromGram positive bacteria in
active form has been developed. The method employs the incubation of cells with lysozyme and
sonication in the presence of salt and detergent followed by repeated freezing and thawing to disrupt the
lipid–protein interactions. Although developed for Dipeptidylpeptidase-III as a demonstration, this
protocol has been used for the successful extraction of some synthetic substrate hydrolyzing activities
from three different Gram positive bacteria with yield of more than 90% for most studied activities.
This method can be routinely used for a wide range of applications and it can also be used to extract and
solubilize other enzymes/proteins only by manipulating detergent, salt and their concentration. It is
simple, straightforward, reproducible, cost effective and a less time consuming protocol for extracting
and solubilizing membrane enzymes.
Introduction
Membrane associated proteins of microorganisms often present
a challenge as well as opportunity, in the quest for better thera-
peutic and prophylactic interactions against infectious diseases,
as membrane associated proteins are important for pathogenicity
of bacteria. Pediococcus acidilactici is an important microor-
ganism with wide applications in the food industry. It is generally
regarded as safe (GRAS) but its vancomycin resistant strains
may cause blood septicemia.1 Dipeptidylpeptidase-III (DPP-III)
removes dipeptide moieties from amino-terminus of oligopep-
tides.2,3 It has been purified from different species but its func-
tions are not well studied.4 As the DPP-III is thought to be
involved in enkephalin degradation with micromolar affinity3,5 it
has emerged as an important target for developing inhibitors
which could act as analgesics for patients of neuropathic and
cancer pain.6 DPP-III is unexplored in microbes so, in order to
understand the functional significance of DPP-III, studies were
planned for purification and characterization of DPP-III from
Pediococcus acidilactici, a microbe in the dairy industry. More-
over it is easier to develop the knock out in microbes which help
to know the particular function. Although DPP-III is a cytosolic
enzyme in mammals,7 studies in our laboratory revealed that it is
located in membranes in Pediococcus acidilactici. Study of
membrane proteins is one of major difficulty because of prob-
lems encountered in extraction while maintaining solubility and
activity. To date, several methods for extracting membrane
proteins have been described.8–13 All these have their own
Department of Biochemistry, Kurukshetra University, Kurukshetra, India.E-mail: [email protected]; Fax: +91-1744-238277; Tel: +91-1744-238196 ext. 2751
2574 | Anal. Methods, 2012, 4, 2574–2577
inherent advantages and disadvantages and none benefit from
the existence of internationally recognized standards for
universally applicable extraction of membrane proteins.
In the present technical note, a simple, straightforward,
reproducible and less time consuming protocol is reported for
extracting and solubilizing membrane enzymes. As a test, this
protocol has also been successfully used for extraction of
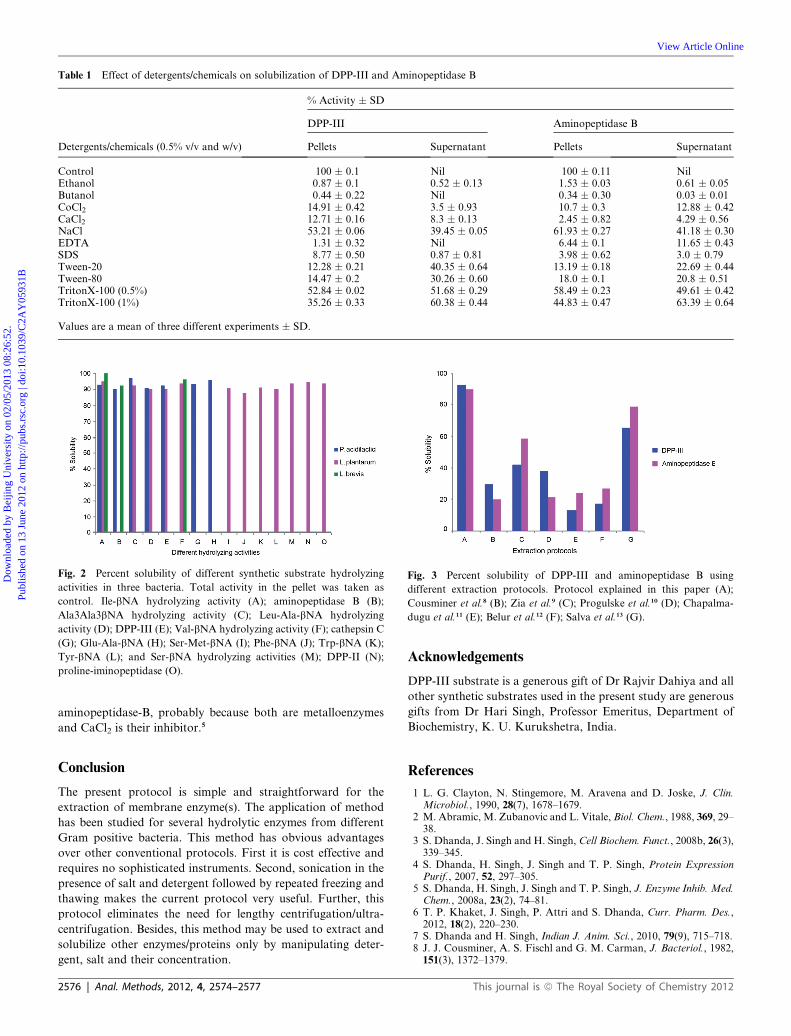
Tyr-bNA (L); and Ser-bNA hydrolyzing activities (M); DPP-II (N);
proline-iminopeptidase (O).
Fig. 3 Percent solubility of DPP-III and aminopeptidase B using
different extraction protocols. Protocol explained in this paper (A);
Cousminer et al.8 (B); Zia et al.9 (C); Progulske et al.10 (D); Chapalma-
dugu et al.11 (E); Belur et al.12 (F); Salva et al.13 (G).
Dow
nloa
ded
by B
eijin
g U
nive
rsity
on
02/0
5/20
13 0
8:26
:52.
Pu
blis
hed
on 1
3 Ju
ne 2
012
on h
ttp://
pubs
.rsc
.org
| do
i:10.
1039
/C2A
Y05
931B
View Article Online
aminopeptidase-B, probably because both are metalloenzymes
and CaCl2 is their inhibitor.5
Conclusion
The present protocol is simple and straightforward for the
extraction of membrane enzyme(s). The application of method
has been studied for several hydrolytic enzymes from different
Gram positive bacteria. This method has obvious advantages
over other conventional protocols. First it is cost effective and
requires no sophisticated instruments. Second, sonication in the
presence of salt and detergent followed by repeated freezing and
thawing makes the current protocol very useful. Further, this
protocol eliminates the need for lengthy centrifugation/ultra-
centrifugation. Besides, this method may be used to extract and
solubilize other enzymes/proteins only by manipulating deter-
gent, salt and their concentration.
2576 | Anal. Methods, 2012, 4, 2574–2577
Acknowledgements
DPP-III substrate is a generous gift of Dr Rajvir Dahiya and all
other synthetic substrates used in the present study are generous
gifts from Dr Hari Singh, Professor Emeritus, Department of
Biochemistry, K. U. Kurukshetra, India.
References
1 L. G. Clayton, N. Stingemore, M. Aravena and D. Joske, J. Clin.Microbiol., 1990, 28(7), 1678–1679.
2 M. Abramic, M. Zubanovic and L. Vitale, Biol. Chem., 1988, 369, 29–38.
3 S. Dhanda, J. Singh and H. Singh, Cell Biochem. Funct., 2008b, 26(3),339–345.
4 S. Dhanda, H. Singh, J. Singh and T. P. Singh, Protein ExpressionPurif., 2007, 52, 297–305.
5 S. Dhanda, H. Singh, J. Singh and T. P. Singh, J. Enzyme Inhib. Med.Chem., 2008a, 23(2), 74–81.
6 T. P. Khaket, J. Singh, P. Attri and S. Dhanda, Curr. Pharm. Des.,2012, 18(2), 220–230.
7 S. Dhanda and H. Singh, Indian J. Anim. Sci., 2010, 79(9), 715–718.8 J. J. Cousminer, A. S. Fischl and G. M. Carman, J. Bacteriol., 1982,151(3), 1372–1379.
This journal is ª The Royal Society of Chemistry 2012