An NADH-tetrazolium-coupled sensitive assayfor malate dehydrogenase in mitochondria
and crude tissue homogenates
Cheng Luo a, Xuemin Wang a,b, Jiangang Long a, Jiankang Liu a,c,d,⁎
a Institute for Nutritional Science, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences,Shanghai, 200031, PR China
b Department of Biochemistry and Molecular Biology, College of Basic Medical Sciences,Second Military Medical University, Shanghai, 200433, PR China
c Children's Hospital Oakland Research Institute, Oakland, CA 94609, United Statesd Institute for Brain Aging and Dementia, University of California, Irvine, CA 92697-4540, United States
Received 8 March 2006; received in revised form 14 April 2006; accepted 18 April 2006
Abstract
A sensitive spectrophotometric assay for determining mitochondrial malate dehydrogenaseactivity is described. The assay measures NADH production by coupling it to the reduction of2-(p-iodophenyl)-3(p-nitrophenyl)-5-phenyl tetrazolium chloride (INT). Via an intermediate electroncarrier, either phenazine methosulfate or lipoamide dehydrogenase, INT accepts electrons and isreduced to a red-colored formazan, which can be quantified by spectrophotometer at 500 nm.This assay uses only commercial reagents but gives a 2–5 fold (with lipoamide dehydrogenase)or 5–20 fold (with phenazine methosulfate) activity increase over currently available assays forpure enzyme in mitochondria isolated from human neuroblastoma cells, rat brain and liver, andcrude homogenates of rat brain and liver. The assay can be easily performed with 96-well plateand less than 2.5 μg protein of isolated mitochondria or crude tissue homogenate. These results
Abbreviations:MDH,malatedehydrogenase;PMS,phenazinemethosulfate; INT,2-(p-iodophenyl)-3(p-nitrophenyl)-5-phenyl tetrazolium chloride; TCA, tricarboxylic acid; BSA, bovine serum albumin; CMDH, conventional MDH assay;PIMDH, PMS-coupled INT MDH assay; LIMDH, lipoamide dehydrogenase-coupled INT MDH assay; Kpi buffer,potassium phosphate buffer.⁎ Corresponding author. Institute for Brain Aging and Dementia, University of California, 1261 Gillespie Neuroscience
Research facility, Irvine, CA 92697-4540, United States. Tel.: +1 949 824 9896; fax: +1 949 824 2071.E-mail address: [email protected] (J. Liu).
Malate dehydrogenases (MDHs, EC 1.1.1.37) are involved in various cellular processes.Eukaryotic cells contain both mitochondrial and cytoplasmic NAD-dependent MDHs.Mitochondrial MDH is involved in the tricarboxylic acid (TCA) cycle, catalyzing theinterconversion of oxaloacetate and malate linked to the oxidation/reduction of dinucleotidecoenzymes, while cytoplasmic MDH is responsible for shuttling oxaloacetate-malate throughthe mitochondrial membrane [1–4]. The currently available MDH assay is a spectrophoto-metric measurement of NADH production [5–7]. The conventional method is simple but lackssensitivity and stability due to limitations such as a colorless product from the reaction and ashort linear period of time.
Using a tetrazolium dye-coupled procedure, we have developed a more sensitive andcolorimetric-continuous method to assay MDH. The electron acceptor dye 2-p-iodophenyl-3-p-nitrophenyl-5-phenyltetrazolium cation (INT) measures NADH production from MDH in bothmitochondrial preparations and crude tissue homogenates. Studies of the redox reactioninvolving NADH and INT have shown that the transfer of reducing equivalents between thereactants is direct and quantitative and that the redox steps are confined to a microenvironmentas in the case of NAD(P)H-dependent enzymatic reactions [8–10]. The intermediate electroncarriers we have used are either phenazine methosulfate (PMS) [11,12] or lipoamidedehydrogenase (EC 1.6.4.3) [13,14]. The transfer of reducing equivalents from NADH to thedye under these conditions is rapid, effective, and irreversible. MDH activity is measured inthe presence of sufficient malate and appropriate NAD+, by measuring the formation of INT-formazan at 500 nm [13,15]. The optimum pH and temperature for the assay system havebeen established. The assay has been compared with the commonly used NADH assay andtested using commercially available pure MDH, MDH in mitochondria isolated from humanneuroblastoma cells and rat brain and liver, and MDH in crude homogenates of rat liver andbrain.
2. Materials and methods
2.1. Chemicals
Lipoamide dehydrogenase (Type III), INT, andMDH from porcine heart (EC 1.1.1.37), NAD+,and rotenone were purchased from Sigma Chemical Co. (St. Louis, MO); PMS from Wako PureChemical Industries, Ltd. BCA™ Protein Assay Kit from Pierce Biotechnology, Inc. (Pierce,Rockford, IL). Tris base was from Amersco Inc. (Palm Harbor, FL). Other chemicals were all A.P.reagents from local vendors.
103C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
2.2. MDH assays
We used a conventional spectrophotometric method as the standard MDH assay, whichmeasures the formation of NADH at 340 nm at 37 °C in the presence of appropriate substrates andcofactors [6]. The reaction mixture contained 5 mM magnesium chloride, 35 mM KH2PO4,2.5 μM rotenone, 2 mM NaN3, and 0.5 mM EDTA, 15 mM malate, pH 7.25. A base-line wasdetermined with sample and reference cuvettes both containing the entire reaction mixture exceptfor NAD+. To initiate the reaction, 0.5 mM NAD+ was added to the sample cuvette and an equalvolume of water was added to the reference sample. The reaction rate was monitored at 340 nm(SPECTRAmax 190, Molecular Devices Corporation, USA) for 2 min.
Two MDH assays have been developed with 2 electron carriers, i.e., lipoamide dehydrogenaseas a LIMDH assay and PMS as a PIMDH assay. The assays were performed using a 96-well plate.The INT-coupled assay reaction mixture, total 250 μl, contained 5 mM magnesium chloride,50 mM Kpi pH 7.8, 3 μM rotenone, and 0.1% BSA,0.5 mM NAD+, INT (0.6 mM), variousconcentrations of mitochondria (from 5 to 50 μg/ml protein), and either PMS (0.2 mM) orlipoamide dehydrogenase (0.1 mg/ml). The reaction was initiated by adding 15 mM malate.Absorption was measured at 500 nm by SPECTRAmax 190 (Molecular Devices Corporation,USA) for 2 min. Blanks were carried out in the absence of malate, and the obtained rate wassubtracted from the sample rates. Units of activity for these assays were determined using mMINT-formazan /minute/mg mitochondrial protein. Stock solutions of INT were prepared at aconcentration of 6 mM in ddH2O, stirred at room temperature until INT was completelydissolved. PMS stocks were prepared at a concentration of 100 mM. Both solutions were stable atlest for 1–3 months if stored in the dark at 4 °C. In our experiments, the two stock solutions wereprepared monthly and other solutions were all prepared freshly before use.
2.3. Crude tissue homogenate preparation
Male Sprague Dawley rats (180–200 g) were purchased from Shanghai SLAC LaboratoryAnimal Co. Ltd (Shanghai, China). The animals were euthanized by decapitation after anovernight fast and the brain and liver were removed for immediate homogenate preparation.Homogenates were prepared as described [16] with a slight modification. Briefly, tissues wererinsed with saline, weighed, and put into ice-cold isolation buffer containing 0.25 M sucrose,10 mM Tris, 0.5 mM EDTA, pH 7.4. Tissues were sheared carefully then minced, and rinsed toeliminate residual blood, and then homogenized in 2.5 vol of isolation buffer. The homogenatewas adjusted to 8 vol with isolation buffer and centrifuged at 1000 ×g for 10 min; the supernatantfraction was decanted and saved. All of the above procedures were carried out at 4 °C. The tissuehomogenate protein concentration was determined using a BCA™ Protein Assay kit (Pierce23225) and bovine serum albumin (BSA) as a standard. The tissue homogenates were stored at−80 °C until enzyme analysis.
2.4. Mitochondrial isolation from rat liver and brain
Similar to the preparation of crude homogenates described above, the supernatant fractions werecombined and centrifuged at 10,000 ×g for 10min. The mitochondrial pellet was washed twice withisolation buffer. All of the above procedures were carried out at 4 °C. The mitochondrial proteinconcentration was determined using the BCA™ Protein Assay kit (Pierce 23225) and bovine serumalbumin (BSA) as a standard. The mitochondria were stored at −80 °C until enzyme analysis.
104 C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
2.5. Mitochondrial isolation from cultured human neuroblastoma SK-N-MC cells
Mitochondria from human neuroblastoma cells were isolated as described [17] with a slightmodification. Briefly, SK-N-MC cells were digested with trypsinase, collected and pelleted bycentrifugation at 1000 ×g for 10 min. The cell pellet was resuspended in ice-cold RSB (ahypotonic buffer for swelling the tissue culture cells) containing 10 mM NaCl, 1.5 mMMgCl2, 10 mM Tris–HCl, pH 7.5, and transferred to a 15-ml Dounce homogenizer. The cellswere allowed to swell for 5–10 min and homogenized with a pestle (small-clearance) forseveral strokes. The homogenate was diluted with a stock MS buffer to give a finalconcentration of 1×MS buffer containing 210 mM mannitol, 70 mM sucrose, 5 mM Tris–HCl, 1 mM EDTA, pH 7.5. The homogenate was mixed by inverting the tube for a couple oftimes and centrifuged at 1000 ×g for 10 min. The supernatant fraction was collected andcentrifuged at 10,000 ×g for 10 min. The mitochondrial pellet was washed twice with isolationbuffer with centrifugation. All of the above procedures were carried out at 4 °C. Themitochondrial protein concentration was also determined using the BCA™ Protein Assay kit.The mitochondria were stored at −80 °C until enzyme assay.
3. Results
3.1. Cofactor requirements for PIMDH and LIMDH assays
The requirements of MDH cofactors are presented in Table 1. There are no significantdifferences in cofactor requirements between the PIMDH and LIMDH assays. There is a 100%requirement for malate and NAD+, a 10–20% requirement for BSA, but no requirement forMgCl2.
3.2. The effects of Kpi and Tris buffers on PIMDH and LIMDH assays
The effects of two buffers, i.e., Kpi and Tris, on PIMDH and LIMDH assays were tested. For theLIMDH assay, there is no significant difference between the use of Kpi buffer and Tris buffer (datanot shown). However, the MDH activity (ΔOD500, milli-units per minute, mean±S.D. of 6experiments) in Kpi buffer is 443.96±9.65, about 2 fold of that in Tris buffer (294.22±5.47) inPIMDH assay. In addition, MDH activity in Kpi buffer is also higher with the PIMDH assay thanwith the LIMDH assay. Therefore, a Kpi buffer was selected for both assays.
Assays were performed using PIMDH and LIMDH procedure as described under “Materials and methods”, using Trisbuffer and omitting single cofactors as listed in the table. Data are percent activity±S.D. (n=8).
0
20
40
60
80
100
7 8 9 10 11
pH
Rea
ctiv
e ra
te (
%)
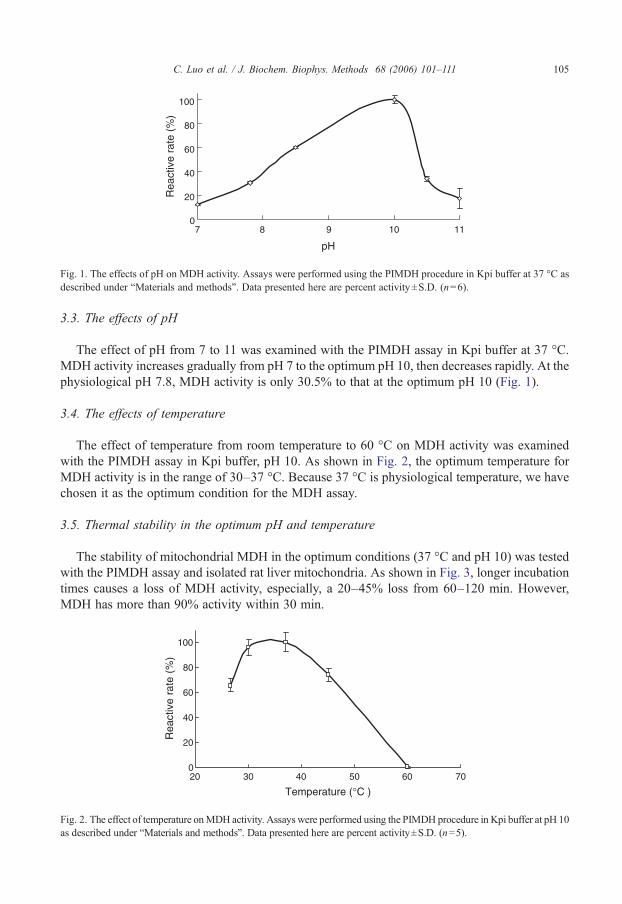
Fig. 1. The effects of pH on MDH activity. Assays were performed using the PIMDH procedure in Kpi buffer at 37 °C asdescribed under “Materials and methods”. Data presented here are percent activity±S.D. (n=6).
105C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
3.3. The effects of pH
The effect of pH from 7 to 11 was examined with the PIMDH assay in Kpi buffer at 37 °C.MDH activity increases gradually from pH 7 to the optimum pH 10, then decreases rapidly. At thephysiological pH 7.8, MDH activity is only 30.5% to that at the optimum pH 10 (Fig. 1).
3.4. The effects of temperature
The effect of temperature from room temperature to 60 °C on MDH activity was examinedwith the PIMDH assay in Kpi buffer, pH 10. As shown in Fig. 2, the optimum temperature forMDH activity is in the range of 30–37 °C. Because 37 °C is physiological temperature, we havechosen it as the optimum condition for the MDH assay.
3.5. Thermal stability in the optimum pH and temperature
The stability of mitochondrial MDH in the optimum conditions (37 °C and pH 10) was testedwith the PIMDH assay and isolated rat liver mitochondria. As shown in Fig. 3, longer incubationtimes causes a loss of MDH activity, especially, a 20–45% loss from 60–120 min. However,MDH has more than 90% activity within 30 min.
0
20
40
60
80
100
20 30 40 50 60 70
Temperature ( °C )
Rea
ctiv
e ra
te (
%)
Fig. 2. The effect of temperature onMDH activity. Assays were performed using the PIMDH procedure in Kpi buffer at pH 10as described under “Materials and methods”. Data presented here are percent activity±S.D. (n=5).
0
20
40
60
80
100
0 20 40 60 80 100 120
Time (minutes)
Rea
ctiv
e ra
te (
%)
Fig. 3. Thermal stability of MDH activity. Assays were performed using the PIMDH procedure in Kpi buffer at 37 °C, pH 10.Data presented here are percent activity±S.D. (n=6).
CMDH
0.19
0.2
0.21
0.22
0 30 60 90 120
0 30 60 90 120
0 30 60 90 120Reaction time (S)
Reaction time (S)
Reaction time (S)
LIMDH
0.1
0.2
0.3
0.4
PIMDH
0.4
0.6
0.8
1
1.2
ΔOD
500
ΔOD
500
ΔOD
340
a)
b)
c)
Fig. 4. A comparison of linearity and sensitivity of 3 assays (CMDH, LIMDH, and PIMDH). Assays were performed asdescribed under “Materials and methods” at 37 °C, pH 10. Data presented are absorption change in 2 min. The activity ofMDH was measured as formation of NADH at 340 nm in CMDH assay and as formation of INT-formazan at 500 nm inPIMDH and LIMDH assays.
106 C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
107C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
3.6. Linearity of the reactive velocity and sensitivity
The linearity of the reactive rate of the 3 assays (CMDH, PIMDH, and LIMDH) was comparedunder the same mitochondrial concentration and the same substrate concentrations (malate andNAD+). As shown in Fig. 4, the time of initial reactive velocity in CMDH is less than 40 s,however, the time of initial reactive velocity in PIMDH or LIMDH is longer than 120 s. In
a)
b)
c)
Liver
0
100
200
300
400
500
600
5 10 25 50Mitochondrial concentration (μg/ml)
5 10 25 50
Mitochondrial concentration (μg/ml)
5 10 25 50
Mitochondrial concentration (μg/ml)
ΔOD
ΔOD
ΔOD
CMDH
LIMDH
PIMDH
CMDH
LIMDH
PIMDH
CMDH
LIMDH
PIMDH
Brain
0
100
200
300
400
500
SK-N-MC cells
0
100
200
300
400
Fig. 5. A comparison of the PIMDH, LIMDH and the commonly used conventional MDH (CMDH) assays. a) Rat livermitochondria; b) rat brain mitochondria; c) human neuroblastoma SK-N-MC cell mitochondria. Assays were performed asdescribed under “Materials and methods” at 37 °C, pH 10. Data presented here are ΔOD: milli-units per minute±S.D.(n=5). The activity of MDH in the CMDH assay was measured as formation of NADH at 340 nm, and measured as theformation of INT-formazan at 500 nm in the PIMDH and LIMDH assays. The value on the y-axis is milli-units/min,calculated using the initial rate.
108 C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
addition, the absorbance at 340 nm at 120 s is about 0.215 in the CMDH assay, 0.34 at 500 nm inthe LIMDH assay, and 1.15 at 500 nm in the PIMDH assay, suggesting that both PIMDH andLIMDH assays are more sensitive than the CMDH assay, and PIMDH is most sensitive.
3.7. Comparison of the PIMDH and LIMDH assays with the conventional method inmitochondria
A comparison of the PIMDH assay, the LIMDH assays, and the commonly used conventionalMDH (CMDH) assay was carried out with mitochondria isolated from human neuroblastoma SK-
a)
b)
c)
Pure MDH
0
100
200
300
400
500
0
100
200
300
400
500
600
0.06 0.12 0.24 0.58
Pure malate dehydrogenase (units/ml)
CMDH
PIMDH
CMDH
PIMDH
CMDH
PIMDH
Liver
Homogenate concentration (μg proteins/ml)
Brain
0
200
400
600
800
10 20 50 100
10 20 50 100
Homogenate concentration (μg proteins/ml)
ΔOD
ΔOD
ΔOD
Fig. 6. A comparison of the PIMDH assay and the commonly usedMDH (CMDH) assay. a) Puremalate dehydrogenase; b) ratliver homogenate; c) rat brain homogenate. Assays were performed as described under “Materials and methods” at 37 °C, pH10.Data presented here areΔOD:milli-units perminute±S.D. (n=5). The activity ofMDH in the CMDHassaywasmeasuredas formation of NADH at 340 nm, and measured as formation of INT-formazan at 500 nm in the PIMDH assay. The value onthe y-axis is milli-units/min, calculated using the initial rate.
109C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
N-MC cells and rat liver and brain. As shown in Fig. 5, both PIMDH and LIMDH assays are moresensitive than the CMDH assay. The initial reactive velocity of LIMDH is about 2–5 fold andPIMDH about 5–10 fold of that of the CMDH assay in the three samples of mitochondria in therange of mitochondrial concentrations from 5–50 μg/ml protein (1.25–12.5 μg protein).
3.8. Comparison of the PIMDH assay with the conventional method using commercial pureMDH and crude homogenates of rat liver and brain
Since thePIMDHassay ismore sensitive than theLIMDHassay (Fig. 5) and alsoPMS is less expensiveandmore stable than lipoamide dehydrogenase,we recommend theuseof thePIMDHassay as a routine labmethod. We further compared its sensitivity with a commercially available MDH assay and found that thePIMDH assay is about 15 times more sensitive than the CMDH assay.
A test of the application of this assay to crude tissue homogenate for total MDH was carriedout with the PIMDH and CMDH assays. As shown in Fig. 6, the PIMDH assay can be used withcrude homogenates of rat liver and brain to assess the mitochondrial and cytoplasmic total MDHwith a 20 fold sensitivity as compared to the CMDH assay. The amount of crude homogenatecould be measured in less than 2.5 μg protein for liver and brain.
4. Discussion
An INT-coupled assay has been used for assaying other NADH-producing dehydrogenases,such as pyruvate dehydrogenase complex [13,18], succinate dehydrogenase [15], lactatedehydrogenase [19,20]. However, a similar procedure for measuring MDH has not beendescribed. In the present study, we developed an INT-coupled assay for measuring MDH activity.The assay requires cofactors malate and NAD+, electron carrier lipoamide dehydrogenase orPMS, and BSA, and is optimal at 37 °C and using a Kpi buffer, pH around 10. This assay isapplicable to either mitochondria isolated from cells and animal tissues such as rat liver and brainor from crude tissue homogenates. Compared to the commonly used MDH assay, this assay ismore sensitive.
As shown in Figs. 5 and 6, the PIMDH assay is about 2–3 fold more sensitive than the LIMDHassay suggesting that PMS is a better electron carrier than lipoamide dehydrogenase. PMS acts as anefficient intermediate transfer agent between NADH and tetrazolium salt, and thus the formazanproduced in the recommended reaction mixture serves as a measure of the dehydrogenase activity[20]. Therefore, the PIMDH assay is highly recommended for common use. In addition to itssensitivity, the lower cost of PMS, greater stability and relative ease of preparation and storage of thestock solution of PMS compared with lipoamide dehydrogenase are other advantages.
MDH activity is two-fold higher in Kpi buffer than in Tris–HCl buffer in the PIMDH assay. Itis known that phosphate is an obligatory component in the reaction mixture and in other enzymeassays. For example, Kpi buffers provide considerably higher activity than other buffers insuccinate dehydrogenase assays [21–23]. Though the mechanism of the higher sensitivity of Kpibuffer than that of Tris buffer needs to be studied further, it is reasonable to suggest the use of Kpibuffer in the present MDH assay for higher sensitivity.
The partial requirement for BSA in the LIMDH and PIMDH assays may be due to the fact thatPMS or lipoamide dehydrogenase alone is not efficient in transferring electrons to tetrazoliumsalts. However, in the presence of non-specific protein, PMS or lipoamide dehydrogenaseprovides a complex more closely resembling the natural protein structure so that electrons couldbe more efficiently transferred to the tetrazolium salt [20]. In addition, the presence of a simple
110 C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
protein in the reaction mixture might keep the formazan to be dispersed finely enough to permitdetermination of the color density in aqueous solution [21,24].
The optimum pH for the assay is approximately 10 for the mitochondrial MDH for theoxidation of malate in Kpi buffer. This is consistent with reports in conventional MDH assays byGrimm and Doherty [25]. McShan [26] emphasized that the optimal pH varies with the dye usedas the electron acceptor. To test whether a similar effect occurs in the MDH assay, we incubatedmitochondria in Kpi buffer, pH 10 at 37 °C. MDH reached 95% activity within 30 min and wasmaintained at more than 80% for an hour, suggesting MDH is stable in 50 mM Kpi, pH 10.
In conclusion, we describe a new, sensitive spectrophotometric assay for measuring MDHactivity. The assaymeasures NADHproduction by coupling to reduction of INT via the intermediateelectron carrier, phenazinemethosulfate. This assay uses only commercial reagents but gives amuchhigher MDH activity than from currently available assays in isolated mitochondria and also in crudetissue homogenates. The assay is simple, sensitive, stable and inexpensive, and therefore shouldhave wide application for those who have an interest in mitochondrial dehydrogenase assays.
5. Simplified description of the method and its applications
The reaction mixture of 250 μl in a 96-well plate contains 50 mMKpi pH 10.0, 3 μM rotenone,0.1%BSA, 0.5 mMNAD+, 0.6 mμ INT, 5–50 μg/ml protein mitochondria, and 0.2 mMPMS. Thereaction is initiated by adding 15 mMmalate. The absorption is observed at 500 nm for 2 min. Asshown in this report, this MDH assay can be used for measuring the activity of pure enzyme,mitochondria isolated from cells and animal issues, and also crude homogenates of animal tissues.Since mitochondrial dysfunction is considered an important contributor to aging and disease, thisassay may be widely used by researchers who have an interest in mitochondrial function in agingand disease.
Acknowledgements
We acknowledge the helpful comments and language editing of Dr. Elizabeth Head, Institute forBrain Aging and Dementia, University of California, Irvine. This study was supported by anOversea Scholars Award from the Chinese Academy of Sciences, the National Science Foundationof China (Grant 30171030, 30472175), Shanghai Municipal Government Award for DiabetesProject, and Shanghai Pujiang Talent Award.
References
[1] Thorne CJ, Kaplan NO. Physicochemical properties of pig and horse heart mitochondrial malate dehydrogenase. J BiolChem 1963;238:1861–8.
[2] Frieden C, Fernandez-Sousa J. Kinetic studies on pig heart cytoplasmic malate dehydrogenase. J Biol Chem1975;250:2106–13.
[3] Tomita T, Fushinobu S, Kuzuyama T, Nishiyama M. Crystal structure of NAD-dependent malate dehydrogenasecomplexed with NADP(H). Biochem Biophys Res Commun 2005;334:613–8.
[4] Goward CR, Nicholls DJ. Malate dehydrogenase: a model for structure, evolution, and catalysis. Protein Sci1994;3:1883–8.
[5] Kitto GB. Intra- and extramitochondrial malate dehydrogenase from chicken and tuna heart. Methods Enzymol1969;13:107–16.
[6] Humphries KM, Szweda LI. Selective inactivation of alpha-ketoglutarate dehydrogenase and pyruvatedehydrogenase: reaction of lipoic acid with 4-hydroxy-2-nonenal. Biochemistry 1998;37:15835–41.
111C. Luo et al. / J. Biochem. Biophys. Methods 68 (2006) 101–111
[7] Ochoa S. Malic dehydrogenase from pig heart. In: Colowick SP, Kaplan N0, editors. Methods in enzymology, vol. I;1955. p. 735–9.
[8] Uppu RM. Novel kinetics in a biomimetic redox reaction involving NADH and tetrazolium salts in aqueous micellarsolutions. J Inorg Biochem 1995;58:193–207.
[9] Rao UM. A micellar model for investigating the chemical nature of hydrogen transfer in NAD(P)H-dependentenzymatic reactions. Biochem Biophys Res Commun 1989;159:1330–6.
[10] Bergel A, Souppe J, Comtat M. Enzymatic amplification for spectrophotometric and electrochemical assays of NAD+and NADH. Anal Biochem 1989;179:382–8.
[11] Lim HH, Buttery JE. Determination of ethanol in serum by an enzymatic PMS-INT colorimetric method. Clin ChimActa 1977;75:9–12.
[12] Roos KJ. Rapid, sensitive and inexpensive method for estimating blood and urine alcohol concentration. Clin ChimActa 1971;31:285–7.
[13] Hinman LM, Blass JP. An NADH-linked spectrophotometric assay for pyruvate dehydrogenase complex in crudetissue homogenates. J Biol Chem 1981;256:6583–6.
[14] Switchenko AC, Miyada CG, Goodman TC, Walsh TJ, Wong B, Becker MJ, et al. An automated enzymatic methodfor measurement of D-arabinitol, a metabolite of pathogenic Candida species. J Clin Microbiol 1994;32:92–7.
[15] Munujos P, Coll-Canti J, Gonzalez-Sastre F, Gella FJ. Assay of succinate dehydrogenase activity by a colorimetric-continuous method using iodonitrotetrazolium chloride as electron acceptor. Anal Biochem 1993;212:506–9.
[16] Krahenbuhl S, Chang M, Brass EP, Hoppel CL. Decreased activities of ubiquinol:ferricytochrome c oxidoreductase(complex III) and ferrocytochrome c:oxygen oxidoreductase (complex IV) in liver mitochondria from rats withhydroxycobalamin[c-lactam]-induced methylmalonic aciduria. J Biol Chem 1991;266:20998–1003.
[17] Clayton DA. JND: isolation of mitochondria from tissue culture cells. Cells: A Laboratory Manual. New York: ColdSpring Harbour Press; 1998.
[18] Schwab MA, Kolker S, van den Heuvel LP, Sauer S, Wolf NI, Rating D, et al. Optimized spectrophotometric assayfor the completely activated pyruvate dehydrogenase complex in fibroblasts. Clin Chem 2005;51:151–60.
[19] Hochella NJ, Weinhouse S. Automated assay of lactate dehydrogenase in urine. Anal Biochem 1965;13:322–35.[20] Nachlas MM, Margulies SI, Goldberg JD, Seligman AM. The determination of lactic dehydrogenase with a
tetrazolium salt. Anal Biochem 1960;1:317–26.[21] Nachlas MM, Margulies SI, Seligman AM. A colorimetric method for the estimation of succinic dehydrogenase
activity. J Biol Chem 1960;235:499–503.[22] Kearney Edna B. Tpsanz: on the requirement of succinic dehydrogenase for inorganic phosphate. Archives of
Biochemistry and Biophysics 1955;55:579–81.[23] Kearney EB, Singer TP. Studies on succinic dehydrogenase. I. Preparation and assay of the soluble dehydrogenase. J Biol
Chem 1956;219:963–75.[24] Nachlas MM, Tsou KC, De Souza E, Cheng CS, Seligman AM. Cytochemical demonstration of succinic
dehydrogenase by the use of a new p-nitrophenyl substituted ditetrazole. J Histochem Cytochem 1957;5:420–36.[25] Grimm FC, Doherty DG. Properties of the two forms of malic dehydrogenase from beef heart. J Biol Chem