An unconventional antimicrobial protein histone from freshwater prawn Macrobrachium rosenbergii: Analysis of immune properties Q3 Jesu Arockiaraj a, * , Annie J. Gnanam b , Venkatesh Kumaresan a , Rajesh Palanisamy a , Prasanth Bhatt a , Muthukumaresan Kuppusamy Thirumalai c , Arpita Roy c , Mukesh Pasupuleti d , Marimuthu Kasi e a Division of Fisheries Biotechnology & Molecular Biology, Department of Biotechnology, Faculty of Science and Humanities, SRM University, Kattankulathur, Chennai 603 203, Tamil Nadu, India b Institute for Cellular and Molecular Biology, The University of Texas at Austin, 1 University Station A4800, Austin, TX 78712, USA c SRM Research Institute, SRM University, Kattankulathur, Chennai 603 203, India d Lab PCN 206, Microbiology Division, CSIR-Central Drug Research Institute, B.S.10/1, Sector 10, Jankipuram Extension, Sitapur Road, Lucknow 226031, Uttar Pradesh, India e Department of Biotechnology, Faculty of Applied Sciences, AIMST University, Semeling Bedong, 08100 Bedong, Kedah, Malaysia article info Article history: Received 19 June 2013 Received in revised form 6 August 2013 Accepted 20 August 2013 Available online xxx Keywords: Prawn Histone Bioinformatics Gene expression Pathogen Antimicrobial activity Q1 abstract In this study, we have reported the first histone characterized at molecular level from freshwater prawn Macrobrachium rosenbergii (MrHis). A full length cDNA of MrHis (751 base pairs) was identified from an established M. rosenbergii cDNA library using GS-FLXÔ technique. It encodes 137 amino acid residues with a calculated molecular mass of 15 kDa and an isoelectric point of 10.5. MrHis peptide contains a histone H2A signature between 21 and 27 amino acids. Homologous analysis showed that MrHis had a significant sequence identity (99%) with other known histone H2A groups especially from Penaeus monodon. Phylogenetic analysis of MrHis showed a strong relationship with other amino acid sequences from histone H2A arthropod groups. Further phylogenetic analysis showed that the MrHis belongs to histone H2A superfamily and H2A1A sub-family. Secondary structure of MrHis showed that the protein contains 50.36% a-helical region and 49.64% coils. The 3D model of MrHis was predicted by I-Tasser program and the model was evaluated for quality analysis including C-score analysis, Ramachandran plot analysis and RMSD analysis. The surface view analysis of MrHis showed the active domain at the N terminal. The antimicrobial property of MrHis protein was confirmed by the helical structure and the total hydrophobic surface along with its net charge. The MFE of the predicted RNA structure of MrHis is 128.62 kcal/mol, shows its mRNA stability. SchiffereEdmundson helical wheel analysis of the N-ter- minal of MrHis showed a perfect amphipathic nature of the peptide. Significantly (P < 0.05) highest gene expression was noticed in the hemocyte and is induced with viral (WSBV and MrNV) and bacteria (Anopheles hydrophila and Vibrio harveyi) infections. The coding sequence of recombinant MrHis protein was expressed in a pMAL vector and purified to study the antimicrobial properties. The recombinant product showed antimicrobial activity against both Gram negative and Gram positive bacteria. In this study, the recombinant MrHis protein displayed antimicrobial activity in its entirety. Hence, it is possible to suggest that the activity may be due to the direct defense role of histone or its N-terminal antimi- crobial property. However, this remains to be verified by detailed investigations. Ó 2013 Published by Elsevier Ltd. 1. Introduction Histones are alkaline proteins present in the nucleus that package the DNA into nucleosomes. They are accountable for supporting the size and shape of a nucleosome [1] and form a major protein component of chromatin. Moreover, it acts as a spool around which DNA winds and plays a major role in the regulation of gene expression [2]. A chromatin consists of atleast one histones per 100 base pairs of DNA molecules [3]. Histone can be categorized under two superfamily groups such as core histones and linker histones. Core histones include H2A, H2B, H3 and H4 and linker histones include H1 and H5. Moreover, these two superfamilies * Corresponding author. Tel.: þ91 44 27452270; fax: þ91 44 27453903. E-mail address: [email protected](J. Arockiaraj). Contents lists available at ScienceDirect Fish & Shellfish Immunology journal homepage: www.elsevier.com/locate/fsi 1050-4648/$ e see front matter Ó 2013 Published by Elsevier Ltd. http://dx.doi.org/10.1016/j.fsi.2013.08.018 Fish & Shellfish Immunology xxx (2013) 1e12 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 YFSIM2733_proof ■ 31 August 2013 ■ 1/12 Please cite this article inpress as: Arockiaraj J, et al., An unconventional antimicrobial protein histone from freshwater prawn Macrobrachium rosenbergii: Analysis of immune properties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
An unconventional antimicrobial protein histone from freshwaterprawn Macrobrachium rosenbergii: Analysis of immune properties
Jesu Arockiaraj a,*, Annie J. Gnanamb, Venkatesh Kumaresan a, Rajesh Palanisamy a,Prasanth Bhatt a, Muthukumaresan Kuppusamy Thirumalai c, Arpita Roy c,Mukesh Pasupuleti d, Marimuthu Kasi e
aDivision of Fisheries Biotechnology & Molecular Biology, Department of Biotechnology, Faculty of Science and Humanities, SRM University,Kattankulathur, Chennai 603 203, Tamil Nadu, Indiab Institute for Cellular and Molecular Biology, The University of Texas at Austin, 1 University Station A4800, Austin, TX 78712, USAc SRM Research Institute, SRM University, Kattankulathur, Chennai 603 203, Indiad Lab PCN 206, Microbiology Division, CSIR-Central Drug Research Institute, B.S. 10/1, Sector 10, Jankipuram Extension, Sitapur Road,Lucknow 226031, Uttar Pradesh, IndiaeDepartment of Biotechnology, Faculty of Applied Sciences, AIMST University, Semeling Bedong, 08100 Bedong, Kedah, Malaysia
7172
7374757677787980818283848586
a r t i c l e i n f o
Article history:Received 19 June 2013Received in revised form6 August 2013Accepted 20 August 2013Available online xxx
1050-4648/$ e see front matter � 2013 Published byhttp://dx.doi.org/10.1016/j.fsi.2013.08.018
87888990919293949596
Please cite this article in press as: Arockiararosenbergii: Analysis of immune properties,
a b s t r a c t
In this study, we have reported the first histone characterized at molecular level from freshwater prawnMacrobrachium rosenbergii (MrHis). A full length cDNA of MrHis (751 base pairs) was identified from anestablished M. rosenbergii cDNA library using GS-FLX� technique. It encodes 137 amino acid residueswith a calculated molecular mass of 15 kDa and an isoelectric point of 10.5. MrHis peptide contains ahistone H2A signature between 21 and 27 amino acids. Homologous analysis showed that MrHis had asignificant sequence identity (99%) with other known histone H2A groups especially from Penaeusmonodon. Phylogenetic analysis of MrHis showed a strong relationship with other amino acid sequencesfrom histone H2A arthropod groups. Further phylogenetic analysis showed that the MrHis belongs tohistone H2A superfamily and H2A1A sub-family. Secondary structure of MrHis showed that the proteincontains 50.36% a-helical region and 49.64% coils. The 3D model of MrHis was predicted by I-Tasserprogram and the model was evaluated for quality analysis including C-score analysis, Ramachandran plotanalysis and RMSD analysis. The surface view analysis of MrHis showed the active domain at the Nterminal. The antimicrobial property of MrHis protein was confirmed by the helical structure and thetotal hydrophobic surface along with its net charge. The MFE of the predicted RNA structure of MrHis is�128.62 kcal/mol, shows its mRNA stability. SchiffereEdmundson helical wheel analysis of the N-ter-minal of MrHis showed a perfect amphipathic nature of the peptide. Significantly (P < 0.05) highest geneexpression was noticed in the hemocyte and is induced with viral (WSBV and MrNV) and bacteria(Anopheles hydrophila and Vibrio harveyi) infections. The coding sequence of recombinant MrHis proteinwas expressed in a pMAL vector and purified to study the antimicrobial properties. The recombinantproduct showed antimicrobial activity against both Gram negative and Gram positive bacteria. In thisstudy, the recombinant MrHis protein displayed antimicrobial activity in its entirety. Hence, it is possibleto suggest that the activity may be due to the direct defense role of histone or its N-terminal antimi-crobial property. However, this remains to be verified by detailed investigations.
� 2013 Published by Elsevier Ltd.
9798 99
100101102103104
1. Introduction
Histones are alkaline proteins present in the nucleus thatpackage the DNA into nucleosomes. They are accountable for
: þ91 44 27453903.aj).
Elsevier Ltd.
105106107108
j J, et al., An unconventional aFish & Shellfish Immunology
supporting the size and shape of a nucleosome [1] and form amajorprotein component of chromatin. Moreover, it acts as a spoolaroundwhich DNAwinds and plays amajor role in the regulation ofgene expression [2]. A chromatin consists of atleast one histonesper 100 base pairs of DNAmolecules [3]. Histone can be categorizedunder two superfamily groups such as core histones and linkerhistones. Core histones include H2A, H2B, H3 and H4 and linkerhistones include H1 and H5. Moreover, these two superfamilies
109110
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
along with the subfamilies are involved in many cellular processessuch as transcriptional regulation, chromosome structure, DNArepair and heterochromatin formation [4e6].
Cho et al. [7] reported that histones are conserved eukaryoticcationic proteins present in the cells and are involved in the anti-microbial activities. Given the additional antimicrobial activity,these histones are placed in the group of antimicrobial peptides.Antimicrobial peptides (AMPs) are evolutionary conserved mole-cules with a net cationic charge of þ3 to þ6. AMPs are found inalmost all living forms ranging from bacteria to humans includingthe crustacean family. In line with other AMPs from various otheranimals, the crustacean antimicrobial peptides (AMPs) are alsogrouped under four different categories [8,9]. They are, single-domain linear a-helical AMPs and peptides enriched in certainamino acids (e.g., bac-like, callinectin, astacidin 2, armadillidin andhomarin), single-domain peptides containing cysteine residuesengaged in disulfide bonds (e.g., defensin, anti-LPS factor and scy-gonadin), multi-domain or chimeric AMPs (e.g., penaeidin, crustin,hyastatin, arasin and stylicin) and unconventional AMPs possessingmultifunctional proteins and protein fragments with antimicrobialactivities (e.g., histones, histone derived fragments, hemocyaninand hemocyanin derived peptides). Recently, a few researchershave reported the antimicrobial activity of core histones and his-tone derived proteins as well as in linker histones [10e18]. Notethat the antimicrobial activities of histones and their derivedproducts have been shown in many organisms including verte-brates such as human [19,20], amphibian [21,22] and fish[11,14,16,18,23,24] and invertebrates such as crustaceans [8,25] andmollusks [10,12,13,15] proving that this phenomenon is all alongthe phyla. Various researchers have reported the antimicrobialactivity of histones from the fish and amphibian’s skin and mucus[16,23,26,27]. The antimicrobial properties of histones are deter-mined based on the number of cationic residues and amphipathicsecondary structure. Further, this property of histones appears to beconnected to their role of damaging pathogenicmicrobes and not totheir capacity of arranging stable pores [23]. However, the processby which histones are procured to the sites outside the nucleus andthe control of histone production in response to pathogenic in-fections is far from being clear [8].
The freshwater prawn Macrobrachium rosenbergii, a potentialaquaculture candidate, is widely distributed in Asia. It is one of theeconomically important cultivable freshwater species in India [28]due to its delicious flesh. However, infectious diseases especially,viral and bacterial infections are causing a serious damage to theaqua industry resulting in heavy economic loss as well disturbancein the food supply chain [29]. It is well known that prawns lack anacquired defensemechanisms and their immune system depend oninnate immunity, which include a cell mediated humoral immunitythat activate AMPs to destroy the invading pathogen [13,30]. Hencethe following study was incited where we have shown that histonemolecules play a prominent role in the defense of M. rosenbergiifrom both viral as well as bacterial. Molecular characterization ofdefense genes would supply a thorough knowledge on the immunesystem of prawn and provide a new understanding on healthmanagement practices and disease control strategy in prawnculture.
2. Materials and methods
2.1. Construction of cDNA library and identification of MrHis
A full length cDNA ofMrHis was identified from the constructedM. rosenbergii cDNA library by the genome sequence FLX� tech-nology. Briefly, total RNA was isolated using Tri Reagent� (LifeTechnologies) from the tissue pool including muscle, gills,
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
haemocyte, hepatopancreas and brain of healthyM. rosenbergii. ThemRNA then isolated by using an mRNA isolation kit (MiltenyiBiotech). The first strand cDNA synthesis and normalization werecarried out with CloneMiner� cDNA library construction kit(Invitrogen) and Trimmer Direct Kit: cDNA Normalization Kit(BioCat GmbH). Thereafter, the GS-FLX� sequencing ofM. rosenbergii cDNA was sequenced according to the manufac-turer’s protocol (Roche). The raw data were processed with theRoche quality control pipeline using the default settings. Seqclean(http://compbio.dfci.harvard.edu/tgi/software/) software was usedto screen for and remove normalization adaptor sequences, ho-mopolymers and reads shorter than 40 bp prior to assembly.Further, the sequences were subjected to assembly usingMIRA (ver.3.2.0) software [31] into full length cDNAs. From the establishedcDNA library of M. rosenbergii sequence database, a histone genewas identified through BLAST annotation program on NCBI (http://www.blast2go.com/b2ghome).
2.2. MrHis sequence analysis using bioinformatics tools
The full length MrHis sequence was compared with other se-quences available in NCBI database and the similarities wereanalyzed. The open reading frame (ORF) and amino acid sequenceof MrHis were obtained by using DNAssist (ver. 2.2) [32]. Charac-teristic domains or motifs were identified using the PROSITE profiledatabase (http://prosite.expasy.org/scanprosite/) where as theidentity, similarity and gap percentages were calculated usingFASTA program [33]. The N-terminal transmembrane sequence wasdetermined by DAS transmembrane prediction program (http://www.sbc.su.se/wmiklos/DAS). Signal peptide analysis was doneby using the online software SignalP (http://www.cbs.dtu.dk). Pair-wise and multiple sequence alignment was constructed in ClustalOmega Program (http://www.ebi.ac.uk/Tools/msa/clustalo/) andedited using BioEdit (ver. 7.1.3.0). The phylogenetic tree was con-structed using Neighbor-Joining method on MEGA 5 by using thedefault settings. The secondary structure of MrHis was predictedusing Polyview (http://polyview.cchmc.org/).
Moreover, 3D structure of MrHis deduced amino acid sequencewas predicted by I-TASSER program (http://zhanglab.ccmb.med.umich.edu/I-TASSER). I-TASSER generates full length model ofproteins by excising continuous fragments from threading align-ments and then reassembling them using replica-exchangedMonteCarlo simulations [34,35]. The five models obtained by I-TASSERprogram was further evaluated for quality analysis including con-fidence score (C-score analysis), Ramachandran plot analysis androot mean square distance (RMSD) analysis. C-score is calculatedbased on the significance of threading template alignments and theconvergence parameters of the structure assembly simulations[36]. Ramachandran plot analysis was carried out to analyze thedispersion of the amino acid residues of the predicted structures ofMrHis. The phi psi plot of the five predicted structures was con-structed using the online tool (http://mordred.bioc.cam.ac.uk/wrapper/rampage.php) [37]. RMSD score is to find out the opti-mum structure of MrHis among the five predicted 3D proteinmodels by I-TASSER program. The models were validated using themodel evaluation server (http://modbase.compbio.ucsf.edu/modeval/) [38]. Based on the results of these model analyses thebest model was taken for further structural analysis on PyMolprogram (ver. 0.99). Further, the 3D structure of MrHis wascompared with the RCSB protein data bank to find out the struc-tural similarity with other experimentally identified structures(http://www.rcsb.org/pdb/home/home.do). The antimicrobialproperty of the MrHis protein was predicted using antimicrobialpeptide database (http://aps.unmc.edu/AP/main.php) [39]. More-over, to find out the stability of the MrHis mRNA, the cDNA
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
sequence of MrHis was converted into their corresponding RNAsequence using DNAssist (ver. 2.2.) to predict the RNA structure ofMrHis. The converted RNA sequence was submitted to RNA foldserver (http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi) and thestructure of MrHis RNA was predicted with minimum free energy(MFE) [40]. Further, SchieffereEdmundson helical wheel wereconstructed in DNASTAR program using a 40 amino acid residues(1e40) from the N-terminal region of MrHis to find out itsamphipathic nature. The amphipathic nature was calculated by thedistribution of polar and non-polar amino acid residues of MrHis.
2.3. Animal handling and immune challenge
Healthy prawns (average body weight of 15 g) were obtainedfrom Sri Sai Aqua Farm, Nellore, Andhra Pradesh, India. They weremaintained in 15 flat-bottomed fiber tanks (50 L) with aerated andfiltered de-chlorinated freshwater (water quality: dissolved oxy-gen, 5.8 � 0.2 mg/L; water temperature, 28 � 1 �C and pH,7.2 � 0.2). All prawns were acclimatized for one week before beinginjected to immune stimulants. During acclimatization period, theprawns were fed ad libitum two times daily at 09.00 and 16.00 hwith a commercially available prawn feed (pellet size: 2.2 mm,Cargill Animal Nutrition, Andhra Pradesh, India). A maximum of 10prawns per tank were maintained during the experiment.
To induce the mRNA expression level, the prawns were injectedwith WSBV and MrNV as reported in our earlier studies [41,42].Briefly, PCR confirmed WSBV and MrNV infected prawn tail tissuewas homogenized in sterile 2% NaCl solution and centrifuged in atabletop centrifuge at 5000 rpm for 5 min at 4 �C. The supernatantwas filtered through 0.45 mm filter and 150 ml per 15 g animal wasinjected into M. rosenbergii. Tissue homogenate prepared fromhealthy tail muscle served as control.
For bacterial infection, M. rosenbergii were injected intraperi-toneally with Anopheles hydrophila or Vibrio harveyi (5 � 106 CFU/ml) suspended in 1X phosphate buffer saline (50 ml per 15 g prawn)by using 22s syringe needle. Equal volume of PBS buffer injectedindividuals served as control. Samples were collected before (0 h),and after injection at regular intervals (3, 6, 12, 21 and 48 h) andwere immediately snap-frozen in liquid nitrogen and stored at�80 �C until total RNA was isolated. Using a sterilized syringe, thehaemolymph (0.3e0.5 ml per prawn) was collected from the prawnheart and immediately centrifuged at 5000 RPM for 10 min at 4 �C.Total RNA extraction was done from the sedimented haemocyte.
All samples were analyzed in three duplications and the resultsare expressed as relative fold of mean � standard deviation as re-ported by Livak and Schmittgenm [43].
2.4. Total RNA extraction and synthesis of first strand cDNA
Total RNA from the control and infected M. rosenbergii wereisolated using Tri Reagent� (Life Technologies), according to themanufacture’s protocol with slight modifications [44,45]. Using2.5 mg of RNA, first strand cDNA was synthesized by SuperScript�
VILO� cDNA Synthesis Kit (Life technologies). The resulting cDNAsolution was stored at �20 �C for further analysis.
2.5. Gene expression analysis by real time PCR
An expression level of MrHis in the haemocytes, pleopods,walking legs, eye stalks, gills, hepatopancreas, stomach, intestine,brain, heart and muscle were measured by quantitative real timepolymerase chain reaction (qRT-PCR). Real time analysis was car-ried out on a BIO-RAD CFX384 Touch Real Time PCR DetectionSystem. Reactions were performed in a 20 ml containing cDNA 4 ml(10 ng), 10 ml of Fast SYBR� Green Master Mix, 0.5 ml of each primer
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
(20 pmol/ml) and 5 ml dH2O. The qRT-PCR had following cycle: 1cycle of 95 �C for 10 s, followed by 35 cycles of 95 �C for 5 s, 58 �C for10 s and 72 �C for 20 s and finally 1 cycle of 95 �C for 15 s, 60 �C for30 s and 95 �C for 15 s. The same qRT-PCR cycle profile was used forthe internal control gene, b-actin. The internal control primers weredesigned from the b-actin of M. rosenbergii (GenBank AccessionNumber AY651918). The primers used in this study were presentedin Table 1. After the PCR program and electrophoresis, data wereanalyzed with BIO-RAD software. To maintain consistency, thebaseline was set automatically by the software. The comparative CTmethod (2�ddCT method) was used to analyze the expression levelof MrHis [43]. All samples were analyzed in three duplications andthe results are expressed as relative fold of mean � standarddeviation.
2.6. Cloning the coding sequence of MrHis into expression vector
All the cloning experiments were carried out according to ourearlier protocol [28,29,46]. MrHis gene specific primers (Table 1)were designed with the corresponding restriction enzyme sites forBamH1 and Pst1 in forward and reverse respectively. Using plasmidDNA of MrHis as a template along with Taq DNA polymerase (Invi-trogen BioServices India Pvt. Ltd, Bangalore, India), PCR was carriedout to amplify the coding sequence. The PCR product was purifiedusing the QIAquick Gel Extraction Kit (QIAGEN India Pvt. Ltd., NewDelhi, India). After PCR, the amplified product was purified by usingthe GeneJETTM (Thermo Scientific). Further, the cloned productswere sequenced using the ABI Prism-Bigdye Terminator CycleSequencing Ready Reaction kit by using the same primers used forthe amplification and analyzed using an ABI 3730 sequencer. Then,both insert and vector were digested with the respective restrictionenzymes. The ligated product was transformed into XL1-Blue cellsand the correct recombinant product was transformed into compe-tent Escherichia coli BL21 (DE3) cells for protein expression.
2.7. Overexpression and purification of recombinant MrHis protein
Transformed E. coli BL21 (DE3) cells were incubated in ampi-cillin (100 mg/mL) Luria broth (LB) overnight. This culture was thenused to inoculate 100 mL of LB broth in 0.2% glucose-rich mediumwith ampicillin at 37 �C until cell density reached 0.7 at OD600.E. coli BL21 (DE3) harboring pMAL-c2x-MrHis was induced for overexpression with 1 mM isopropyl-b-thiogalactopyranoside (IPTG)and incubated at 15 �C for 6 h. Cells were harvested by centrifu-gation (10 000 RPM for 20 min at 4 �C). Then the cells were re-suspended in column buffer (TriseHCl, pH 7.4, 200 mM NaCl) andfrozen at �20 �C overnight. After thawing on ice, cells were dis-rupted by sonication. The crude MrHis fusion protein fused withmaltose binding protein (MBP) was purified using pMAL� protein
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
fusion and purification system protocol (New England Biolabs UKLtd, United Kingdom). Further, DEAE-Sepharose� ion exchangechromatography method was used to purify the recombinantMrHis protein away from MBP and the protease, and we also stip-ulated an additional purification step for removing trace contami-nants according to the manufacture’s protocol (New EnglandBiolabs UK Ltd, United Kingdom). Then the purity of the expressedenzymewas verified by 12% SDS-PAGE and the molecular weight oftarget protein was evaluated using protein molecular weightstandards. Proteins were visualized by staining with 0.05% Coo-massie blue R-250. The concentrations of purified proteins weredetermined by the method of Bradford [47] using bovine serumalbumin (BSA) as the standard. The purified protein was kept at�80 �C until determination of molecular functional activities.
2.8. Antimicrobial activity of purified recombinant MrHis protein
The antimicrobial activity of purified recombinant MrHis proteinwas performed according to the methodology described by Li et al.[13] and Hikima et al. [48]. The assay was conducted using Gramnegative (A. hydrophila, E. coli, Edwardsiella tarda, Vibrio para-haemolyticus, Vibrio alginolyticus and V. harveyi) and Gram positive(Bacillus subtilis, Streptococcus iniae, Staphylococcus aureus, Entero-coccus faecium and Lactococcus lactis) bacteria. As suggested by DeZoysa et al. [10] 100 mg ampicillin and the same volume of DEPCtreated nuclease-free de-ionized water was used as positive andnegative controls respectively. The 1.5% broth agar containing testedbacteria (OD600 ¼ 0.1) was casted onto petriplates. The purified re-combinantMrHis protein sample at various concentrations (0, 25, 50and 100 mg), the positive control and the negative control weredropped into the individual wells in the agar plates and incubated at35 �C for 20 h. The diameter of the inhibition zone (millimeter) was
Fig. 1. The full length nucleotide and amino acid sequence of MrHis. The single letter aminoare italicized. The start (ATG) and stop codon (TAA) are bolded. The nucleotide and amino acishaded in grey. Histone H2A signature (21e27) is given in box. DNA binding site (28e78) isThe amino acid residues Asn38eTyr39eAla40eGlu41 forms homodimerization interface is hi
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
measured. The assays were performed in three replicates and thevalues were presented here as mean � standard deviation.
2.9. Statistical analysis
For comparison of MrHis relative gene expression and antimi-crobial activity, statistical analysis was performed using one-wayANOVA and mean comparisons were performed by Tukey’s Mul-tiple Range Test using SPSS 11.5 software at the 5% significant level.
3. Results
3.1. MrHis cDNA analysis
The obtained complete M. rosenbergii histone sequence wassubmitted to EMBL GenBank database under the accession numberHG001454. The full length sequence of MrHis was 751 base pairs(bp), with two uncoding region each at both ends i.e, 50 untrans-lated region (UTR) of 95 bp and a 245 bp 30 UTR (Fig. 1). The openreading frame (ORF) is 411 bp and encoded for 137 amino acidresidues with a calculated molecular mass of 15 kDa and an iso-electric point of 10.5.
3.2. MrHis amino acid analysis
MrHis amino acid (aa) sequence does not have a signal peptideregion and transmembrane region, as per the SignalP analysis andDAS transmembrane prediction program respectively. ScanPrositeanalysis revealed that the predicted MrHis protein contains a his-tone H2A signature at Ala21eGly22eLeu23eGln24ePhe25ePro26eVal27. Other than this gene specific motif, MrHis protein contains 6common motifs including 2 protein kinase C phosphorylation sites
acid code is shown below the corresponding nucleotide sequences. The 50 and 30 UTRd sequences are numbered in the left and right side respectively. Histone H2A domain isunderlined. The acetylation sites (K6 and K9) and ubiquitination site (K119) are rounded.ghlighted in dark grey.
at Ser2eGly3eArg4 and Ser18eSer19eArg20, 1 ATP/GTP-binding sitemotif A at Gly7eGly8eLys9eVal10eLys11eGly12eLys13eSer14 and 3N-myristoylation sites at Gly99eVal100eThr101eIle102eAla103eGln104, Gly106eVal107eLeu108ePro109eAsn110eIle111 and Gly124eAla125eLys126eGly127eSer128eLys129. Further motif search revealedthat MrHis belongs to the H2A super family which contains a DNAbinding (28e78), acetylation (K6 and K9) and ubiquitination sites(K119).
3.3. Homologous analysis of MrHis
The homologous analysis showed that MrHis had a significantsequence identity with other known histone H2A groups. Thehighest sequence identity (99%) was observed with histone H2Afrom Penaeus monodon (crustacean) and Anopheles gambiae andDrosophila melanogaster (insects), all these belong to Phylum:Arthropod (Table 2). Significant similarities were observed amongother arthropods which has high homology in the N terminal re-gion (data not shown). All the organisms taken for sequence sim-ilarity analysis showed not less than 60% similarity with MrHis.
3.4. Multiple sequence alignment
The deduced amino acid sequences of the MrHis were alignedwith the other histone H2A family members (including crustacean,fish, insect, bird andmammal) (Fig. 2). The results reveal that Salmosalar has the longest amino acid sequence (142 aa). Even though thelength of the amino acids varied from species to species, manyconserved residues were observed among the sequences. In the N-terminal region, the histone H2A signature remains conserved inthe sequences selected for analysis. As reported in the literature theN terminal region is more conserved than the C-terminal regionand also more number of gaps are present in the C-terminal regioncompared to the N-terminal region [49].
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
3.5. Phylogenetic tree
The phylogenetic tree formed two separate clusters of verte-brates (includes amphibian, fishes, bird and mammals) and in-vertebrates (mollusk, crustaceans and insects). Histone H2Asuperfamily FZ sub-family from Homo sapiens formed as an outgroup. The phylogenetic analysis of MrHis showed a strong rela-tionship with other amino acid sequences from histone H2Aarthropod groups (Fig. 3). Moreover, to find the evolutionary rela-tionship of MrHis, the sequence was compared with the other H2Asub-family classes including H2A1A, H2A1K, H2A1M, H2A1L,H2A1J, H2A1D, H2AC, H2AFJ and H2AFZ fromH. sapiens. The resultsshowed that MrHis is evolutionarily related to the H2A1A sub-family of H2A superfamily. Furthermore, to confirm the sub-family of MrHis, we constructed a separate phylogenetic tree us-ing other histones including H1, H2, H3 and H4. The results showedthatMrHis is clustered together with H2A group and showed a highdivergence from H1, H3 and H4 (data not shown). Hence, based onthese analyses, it is confirmed that the identified MrHis, belongs tohistone H2A superfamily and H2A1A sub-family.
3.6. Structural analysis of MrHis
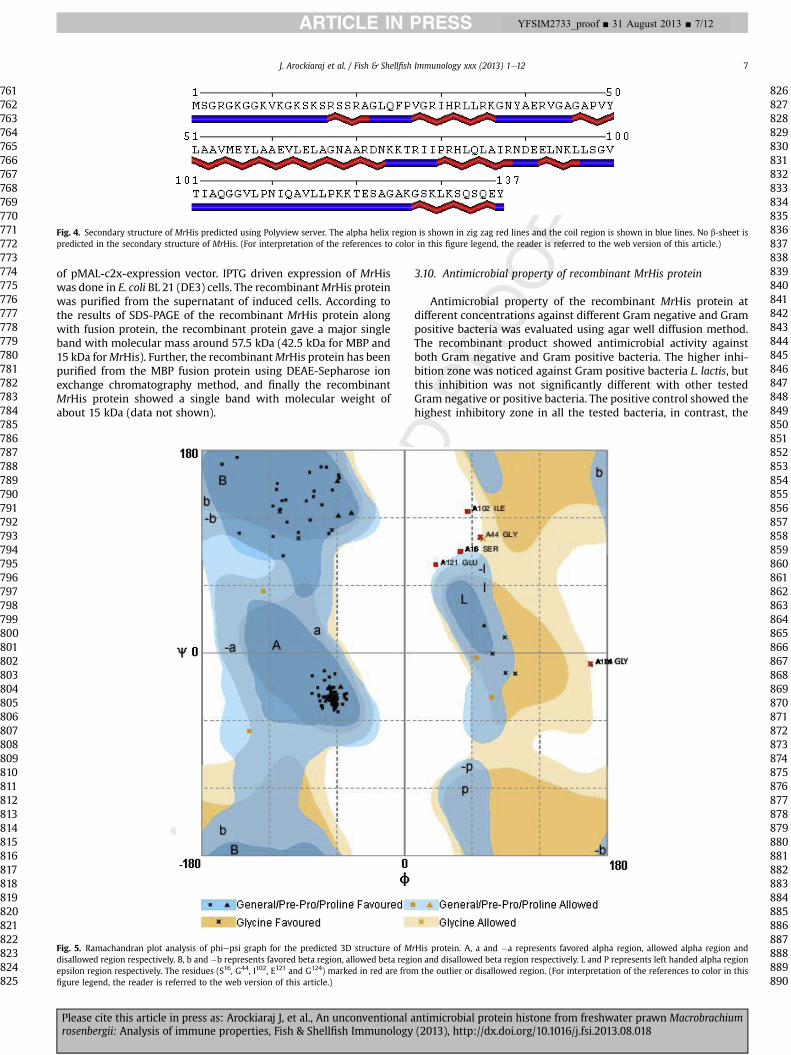
The secondary structural analysis of MrHis showed that theprotein contains 50.36% a-helical region (69 amino acid residues)and 49.64% coils (68 amino acid residues). No b-sheet region hasbeen noticed in the secondary structure of MrHis (Fig. 4).
I-TASSER program predicted five different models for MrHis pro-tein. The quality of the models was validated using Ramachandranplot analysis and the best representation of psi phi graph analysis isgiven in Fig. 5. The representation clearly indicates the proline andglycine were present in favored and allowed regions. Further, theRMSD value (Table 3) of the five models was calculated to obtain thebestmodel for furtherevaluation. Basedon these evaluations, thebestmodel (model-1, Table 2) was selected and is given in Fig. 6. Further,the model was viewed in PyMol surface view. This analysis showedthe active domain region of MrHis at the N terminal region. Further,the RCSB database analysis showed that theMrHis has themaximumstructural identity to histone H2A from Xenopus laevis, which isexperimentally, obtained using X-Ray diffraction method.
The antimicrobial property ofMrHis proteinwas confirmed by thehelical structural analysis and the total hydrophobic surface alongwith itsnet charge ofþ15. The results indicate thatMrHis hasa total of28 hydrophobic residues on the hydrophobic surface; hence, it has ahigh chance for interactionwith themembraneofmicrobes and act asan antimicrobial protein. MrHis has a total net charge of þ15 withprotein binding potential (Boman index) of 1.76 kcal/mol. The pre-dictedRNAfoldstructureofMrHiswithminimumfreeenergy (MFE) isgiven in Fig. 7. The MFE of the predicted RNA structure of MrHisis �128.62 kcal/mol. The predicted mRNA structure shows that theRNA is mostly paired and very few nucleotides are left unpaired.
SchiffereEdmundson helical wheel analysis of the N-terminalregion (40 residues) ofMrHis clearly showed a perfect amphipathicnature of the peptide (Fig. 8). Hydrophilic residues such as serine(S), lysine (K), histidine (H), arginine (R), asparagine (N), glutamine(Q) and hydrophobic residues such as valine (V), glycine (G), alanine(A), phenylalanine (F), tyrosine (Y), leucine (L), isoleucine (I), pro-line (P), methionine (M) face each other in the helical wheel.
3.7. Tissue distribution of MrHis
To investigate the tissue distribution of MrHis transcripts, totalRNA extraction followed by cDNA synthesis were synthesized fromvarious M. rosenbergii tissues and subjected to quantitative realtime PCR analysis. Fig. 9A shows that the MrHis transcript is
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
Fig. 2. Multiple sequence alignment ofMrHis. Analysis was performed by ClustalW, using representatives of histone H2A from tiger prawn Penaeus monodon (ABX84387), mosquitoAnopheles gambiae (XP_318365), Atlantic salmon Salmo salar (ACI66032), domestic fowl Gallus gallus (NP_001025924) and human Homo sapiens (Q96QV6). The identical sequencesare highlighted in black. The H2A signature is marked in double arrow. Deletions are indicated by dashes. The position of the sequences is numbered in the right margin.
J. Arockiaraj et al. / Fish & Shellfish Immunology xxx (2013) 1e126
expressed in all the tissues analyzed. Significantly (P < 0.05)highest expression was noticed in haemocyte followed by hepato-pancreas, muscle, stomach, heart, gill, brain, pleopod, eye stalk,intestine and walking leg.
3.8. mRNA transcription after viral and bacterial infection
Based on the results of tissue distribution analysis,MrHis mRNAexpression in M. rosenbergii is induced in haemocyte followingchallenge by virus (WSBV and MrNV) and bacteria (A. hydrophilaand V. harveyi). WSBV injected M. rosenbergii, MrHis mRNAexpression significantly (P < 0.05) increased (19-fold) at 12 h p.i.,when compared to the control group (Fig. 9B). MrNV induced
Fig. 3. The phylogenetic tree of MrHis constructed by Neighbor-Joining Method in MEGA 5.0an alignment corresponding to the full length amino acid sequences. The numbers shown anumber given in the parenthesis is GenBank accession numbers. The details of sequences
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
mRNA expressionwas significantly (P< 0.05) higher (7-fold) at 12 hpost-injection (p.i.) compared to the control (Fig. 9C). The relativeexpression pattern of MrHis in the haemocyte after A. hydrophilainjection is shown in Fig. 9D. MrHis mRNA transcription signifi-cantly (P < 0.05) increased (15 fold) at 12 h p.i., then subsequentlydecreased by 48 h p.i. In V. harveyi injected individuals, significantly(P < 0.05) higher expression was observed at 12 h p.i., and was 16-fold higher than the control (Fig. 9E).
3.9. Overexpression and purification of recombinant MrHis protein
The putative mature MrHis molecule was expressed in E. colicells after cloning the cDNA into the BamH1 and Pstl restriction sites
after aligning the amino acid sequences using ClustalW program. The tree is based ont the branches denote the bootstrap majority consensus values of 1000 replicates. Theused for constructing the phylogenetic tree are given in Table 1.
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
Fig. 4. Secondary structure of MrHis predicted using Polyview server. The alpha helix region is shown in zig zag red lines and the coil region is shown in blue lines. No b-sheet ispredicted in the secondary structure of MrHis. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
J. Arockiaraj et al. / Fish & Shellfish Immunology xxx (2013) 1e12 7
of pMAL-c2x-expression vector. IPTG driven expression of MrHiswas done in E. coli BL 21 (DE3) cells. The recombinantMrHis proteinwas purified from the supernatant of induced cells. According tothe results of SDS-PAGE of the recombinant MrHis protein alongwith fusion protein, the recombinant protein gave a major singleband with molecular mass around 57.5 kDa (42.5 kDa for MBP and15 kDa forMrHis). Further, the recombinantMrHis protein has beenpurified from the MBP fusion protein using DEAE-Sepharose ionexchange chromatography method, and finally the recombinantMrHis protein showed a single band with molecular weight ofabout 15 kDa (data not shown).
Fig. 5. Ramachandran plot analysis of phiepsi graph for the predicted 3D structure of Mdisallowed region respectively. B, b and �b represents favored beta region, allowed beta regiepsilon region respectively. The residues (S16, G44, I102, E121 and G124) marked in red are fromfigure legend, the reader is referred to the web version of this article.)
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
3.10. Antimicrobial property of recombinant MrHis protein
Antimicrobial property of the recombinant MrHis protein atdifferent concentrations against different Gram negative and Grampositive bacteria was evaluated using agar well diffusion method.The recombinant product showed antimicrobial activity againstboth Gram negative and Gram positive bacteria. The higher inhi-bition zone was noticed against Gram positive bacteria L. lactis, butthis inhibition was not significantly different with other testedGram negative or positive bacteria. The positive control showed thehighest inhibitory zone in all the tested bacteria, in contrast, the
rHis protein. A, a and �a represents favored alpha region, allowed alpha region andon and disallowed beta region respectively. L and P represents left handed alpha region
the outlier or disallowed region. (For interpretation of the references to color in this
negative control showed the lowest inhibitory zone in all the testedbacteria (Fig. 10). The result indicates that the antimicrobial activityof MrHis is concentration dependant manner i.e., the activity in-creases according to the concentration of the recombinant productincreases.
4. Discussion
In vertebrates and invertebrates, Histone H2A variant is re-ported to be involved in host immune response by acting as anti-microbial peptides (AMPs) [13]. The AMPs produced from HistoneH2A have not yet reported in freshwater prawn, to the best of ourknowledge this is the first report from freshwater prawn histoneH2A. In this study, molecular cloning, gene expression of histoneH2A from freshwater prawn M. rosenbergii and its recombinantproduct were over expressed and purified to study whether asimilar mechanism exists inM. rosenbergii. The characterizedMrHisfrom the constructed cDNA library of M. rosenbergii, has all thehistone H2A characteristic features including H2A signature motifand have 99% sequence identity with other homologous ofP. monodon, A. gambiae and D. melanogaster. Therefore, based on thehomologous, domain and motif as well as phylogenetic analysis, itis tempting to suggest that the characterized MrHis is histone H2Avariant.
The secondary structural analysis showed that MrHis containsmany coils and a-helices. Boman [50] and Papagianni [51] hasearlier stated that protein possessing antimicrobial property willhave more number a-helix regions. Moreover, the structural anal-ysis showed that the N-terminal region of MrHis is rich in cationicamino acids including lysine (10%) and arginine residues (8%),hence MrHis contains a net positive charge of þ15, which isimportant for an AMP as suggested by Boman [50] and Tagai [52]. Inaddition to this, due to the rich amount of lysine at the N-terminalsite of histone H2A it is also involved in DNA binding [53]. Theobtained 3D structure of MrHis protein by I-TASSER program was
Fig. 6. Predicted 3D structure ofM. rosenbergii histone with the suspected antimicrobial pepantimicrobial peptide region is highlighted in blue. (For interpretation of the references to
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
viewed through PyMol surface view and it contains a proline hingeregion, which is a major structural component of AMPs and isresponsible for cell penetration [21].
The predicted RNA structure of MrHis along with its MFE valueclearly indicated that the mRNA ofMrHis is highly stable. Accordingto Xiong and Waterman [54], the MFE value of mRNA depends onAU/GC pairing. The paired base pairs receive negative value andunpaired base pairs receive positive value. In this study, MrHismRNA received �128.62 kcal/mol, hence it is possible to suggestthat many nucleotide base pairs of MrHis are paired and a very fewnucleotides in the mRNA are left unpaired thus indicating stabilityof MrHis mRNA molecule.
Furthermore, we analyzed 40 amino acid residues ofMrHis fromthe N-terminal region using SchieffereEdmundson helical wheelprogram to study the amphipathic nature of the N-terminal peptidesequence. It is clearly indicated that MrHis is a linear a-helicalmolecule, having hydrophobic and hydrophilic residues onopposing sides similar to the earlier reports [7,10,13,15,16,18,19].Moreover, the AMP prediction program clearly shows that theMrHis protein can have antimicrobial property as the N-terminalregion ofMrHis has a significant similarity with the earlier reportedAMPs including buforin I [21], hipposin [24], EC-hepcidin1 [55],Oxyopinin 1 [56] and cyto-insectotoxin 1a [57]. All these AMPsshare several characteristics features including molecular mass, netpositive charge, hydrophobic andmembrane-active molecules withmultiple arginine and lysine residues [21,24,55e57]. Similarly,MrHis displays the physical properties such as molecular mass15 kDa, hydrophobic surface with 28 hydrophobic residues and thetotal net charge of þ15, thus confirming the antimicrobial propertyof MrHis. Interestingly, there is no cysteine residue noticed inMrHis, which clearly shows thatMrHis is an a-helical AMP withoutcysteine residue.
The gene expression analysis showed that MrHis is highlyinduced in haemocyte after viral and bacterial infection. Hence itpossible to suggest that MrHis may have an immune responseagainst microbial infections accumulating in M. rosenbergii bydirect defence role ofMrHis or histone derived peptides as reportedby De Zoysa et al. [10]. In contrast, the histone expression was notinduced in scallop hemocyte by pathogenic bacteria [13]. De Zoysaet al. [10] reported that histone H2A expressionwas highly inducedin disk abalone gill by bacterial infection. Due to these contrastingresults [10,13], it is still not clear how the lysis process occursin vivo, as reported elsewhere, whether by the direct defence role ofhistone or any other derived peptide of histone. Therefore, thisaspect needs more detailed investigation to find out the role ofhistone H2A in aquatic organisms.
tide region highlighted in yellow surface view. The proline hinge region in the predictedcolor in this figure legend, the reader is referred to the web version of this article.)
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
Fig. 7. The mRNA structure of MrHis predicted using RNA fold server. The different regions of the mRNA are marked appropriately.
Fig. 8. The SchiffereEdmundson helical wheel diagram of N terminal region (40 residues) of MrHis designed using DNASTAR-Protean software tool. The amino acid residues arenumbered from the amino terminal end.
J. Arockiaraj et al. / Fish & Shellfish Immunology xxx (2013) 1e12 9
Please cite this article in press as: Arockiaraj J, et al., An unconventional antimicrobial protein histone from freshwater prawn Macrobrachiumrosenbergii: Analysis of immune properties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
Fig. 9. Relative quantification of MrHis gene expression by real time PCR. A: Results of tissue distribution analysis of MrHis from various organs of M. rosenbergii. Data are given as aratio to MrHis mRNA expression in walking leg. B, C, D and E: The time course of MrHis mRNA expression in haemocyte at 0, 3, 6, 12, 24, and 48 h post-injection with virus (WSBVand MrNV) and bacteria (A. hydrophila and V. harveyi) respectively. All samples were analyzed in three duplications and the results are expressed as relative fold of one sample asmean � standard deviation.
J. Arockiaraj et al. / Fish & Shellfish Immunology xxx (2013) 1e1210
MrHis coding sequence was validated by using the pMAL-c2x-expression vector and expressed in E. coli as a fusion protein. Re-combinant MrHis protein was purified to homogeneity usingpMAL� protein fusion and purification system. Themolecular massof protein was approximately 15 kDa on 12% SDS-PAGE gel, in linewith previous reports [7,10,13,15,16,18,22,23].
Some researchers used a short peptide (40 amino acid resi-dues from N-terminal region of histone), either as a syntheticform or as a natural product to study the antimicrobial activity ofhistone [10,13]. Hence, an attempt was made in this study tounderstand the antimicrobial activity of purified recombinantMrHis protein, containing both N and C terminal regions. Inter-estingly, the recombinant MrHis protein showed antimicrobialactivity against all the tested Gram negative and Gram positivebacteria. But the results were significant at the higher concen-tration of recombinant MrHis protein (100 mg) and positive
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
control only. Thus, indicating MrHis antimicrobial activity isconcentration dependant. Our results regarding the antimicro-bial activity were different from that of the earlier reports by DeZoysa et al. [10] and Li et al. [13]. We believe that the differencein the activity could be due to the use of synthetic and naturalhistone products along with the intact histone by these re-searchers. In this study, the recombinant MrHis protein displayedantimicrobial activity in its full length form. However, previousreports [10,13] indicate the role of N-terminal region alone in theactivity. This probably suggests that the presence of C-terminalregion in MrHis protein does not affect the N-terminal region’santimicrobial activity. Hence, it is possible to suggest that theactivity may be due to the direct defence role of histone or its N-terminal antimicrobial property. However, this remains to beverified by detailed investigations on the role of MrHis in prawndefence mechanism, the importance of histone and/or histone
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018
Q2
Fig. 10. The antimicrobial activity of recombinant MrHis protein against the Gramnegative (A) and Gram positive bacteria (B). The diameters of the inhibition zoneshowing the expression of the recombinant MrHis protein at different concentrationsalong with its positive and negative controls.
J. Arockiaraj et al. / Fish & Shellfish Immunology xxx (2013) 1e12 11
derived peptides and its vigorous modification before and afterpathogenic infection.
Acknowledgments
This research is supported by SERB Young Scientist FellowshipScheme (SERC/LS-437/2011), Science and Engineering ResearchBoard (SERB), Department of Science and Technology, Ministry ofScience and Technology, Government of India, New Delhi.
References
[1] Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structureof the nucleosome core particle at 2.8 A resolution. Nature 1997;389:251e60.
[2] Alva V, Ammelburg M, Lupas AN. On the origin of the histone fold. BMC StructBiol 2007;7:17.
[3] Ward R, Bowman A, El-Mkami H, Owen-Hughes T, Norman DG. Long distancePELDOR measurements on the histone core particle. J Am Chem Soc 2009;131:1348e9.
[4] Redon C, Pilch D, Rogakou E, Sedelnikova O, Newrock K, Bonner W. HistoneH2A variants H2AX and H2AZ. Curr Opin Genet Dev 2002;12:162e9.
[5] Bhasin M, Reinherz EL, Reche PA. Recognition and classification of histonesusing support vector machine. J Comput Biol 2006;13:102e12.
[6] Wu N, Yue HM, Chen BO, Gui JF. Histone H2A has a novel variant in fish oo-cytes. Biol Reprod 2009;81:275e83.
[7] Cho JH, Sung BH, Kim SC. Buforins: histone H2A-derived antimicrobial pep-tides from toad stomach. Biochim Biophys Acta 2009;1788:1564e9.
[8] Rosa RD, Barracco MA. Antimicrobial peptides in crustaceans: review. ISJ2010;7:262e84.
[9] Pasupuleti M, Malmsten M, Schmidtchen A. Antimicrobial peptides: a keycomponents of innate immunity. Crit Rev Biotechnol 2011;32:143e71.
[10] De Zoysa M, Nikapitiya C, Whang I, Lee JS, Lee J. Abhisin: a potential anti-microbial peptide derived from histone H2A of disk abalone (Haliotis discusdiscus). Fish Shellfish Immunol 2009;27:639e46.
[11] Chaithanya ER, Philip R, Sathyan N, Kumar PRA. Molecular characterizationand phylogenetic analysis of a histone-derived antimicrobial peptide teleostin
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
from the marine teleost fishes, Tachysurus jella and Cynoglossus semifasciatus.ISRN Mol Biol 2013. Article ID 185807.
[12] Dorrington T, Villamil L, Gómez-chiarri M. Upregulation in response toinfection and antibacterial activity of oyster histone H4. Fish ShellfishImmunol 2011;30:94e101.
[13] Li C, Song L, Zhao J, Zhu L, Zou H, Zhang H, et al. Preliminary study on a po-tential antibacterial peptide derived from histone H2A in hemocytes of scallopChlamys farreri. Fish Shellfish Immunol 2007;22:663e72.
[14] Nama BH, Seo JK, Go HJ, Lee MJ, Kim YO, Kim DG, et al. Purification andcharacterization of an antimicrobial histone H1-like protein and its gene fromthe testes of olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol2012;33:92e8.
[15] Sathyan N, Philip R, Chaithanya ER, Kumar PRA. Identification and molecularcharacterization of mlluskin, a histone-H2A-derived antimicrobial peptidefrom molluscs. ISRN Mol Biol 2012. Article ID 219656.
[16] Park IY, Park CB, Kim MS, Kim SC. Parasin I, an antimicrobial peptide derivedfrom histone H2A in the catfish Parasilurus asotus. FEBS Lett 1998;437:258e62.
[17] Kashima M. H1 histones contribute to candidacidal activities of humanepidermal extract. J Dermatol 1991;18:695e706.
[18] Richards RC, O’Neil DB, Thibault P, Ewart KV. Histone H1: an antimicrobialprotein of Atlantic salmon (Salmon salar). Biochem Biophys Res Commun2001;284:549e55.
[19] Hirsch JG. Bactericidal action of histone. J Exp Med 1958;108:925e44.[20] Kim HS, Cho JH, Park HW, Yoon H, Kim MS, Kim SC. Endotoxin-neutralizing
antimicrobial proteins of the human placenta. J Immunol 2002;168:2356e64.[21] Park CB, Kim MS, Kim SC. A novel antimicrobial peptide from Bufo bufo gar-
garizans. Biochem Biophys Res Commun 1996;218:408e13.[22] Kim HS, Park CB, Kim MS, Kim SC. cDNA cloning and characterization of
buforin I, an antimicrobial peptide: a cleavage product of histone H2A. Bio-chem Biophys Res Commun 1996;229:381e7.
[23] Fernandes JMO, Kemp GD, Molle MG, Smith VJ. Anti-microbial properties ofhistone H2A from skin secretions of rainbow trout, Oncorhynchus mykiss.Biochem J 2002;368:611e20.
[24] Birkemo GA, Luders T, Andersen O, Nes IF, Nissen-Meyer J. Hipposin, a histonederived antimicrobial peptide in Atlantic halibut (Hippoglossus hippoglossus L).Biochim Biophys Acta 2003;1646:207e15.
[25] Patat SA, Carnegie RB, Kingsbury C, Gross PS, Chapman R, Schey KL. Antimi-crobial activity of histones from hemocytes of the pacific white shrimp. Eur JBiochem 2004;271:4825e33.
[26] Robinette D, Wada S, Arroll T, Levy MG, Miller WL, Noga EJ. Antimicrobialactivity in the skin of the channel catfish Ictalurus punctatus: characterizationof broad-spectrum histone-like antimicrobial proteins. Cell Mol Life Sci1998;54:467e75.
[27] Fernandes JM, Molle G, Kemp GD, Smith VJ. Isolation and characterization ofoncorhyncin II, a histone H1-derived antimicrobial peptide from skin secre-tions of rainbow trout, Oncorhynchus mykiss. Dev Comp Immunol 2004;28:127e38.
[28] Arockiaraj J, Sarasvathi E, Puganeshwaran V, Arun S, Rofina YO, Subha B.Molecular cloning, characterization and gene expression of an antioxidantenzyme catalase (MrCat) from Macrobrachium rosenbergii. Fish ShellfishImmunol 2012e;32:670e82.
[29] Arockiaraj J, Puganeshwaran V, Sarasvathi E, Arun S, Rofina YO, Subha B. Geneprofiling and characterization of arginine kinase-1 (MrAK-1) from freshwatergiant prawn (Macrobrachium rosenbergii). Fish Shellfish Immunol 2011;31:81e9.
[30] Roch P. Defense mechanisms and disease prevention in farmed marine in-vertebrates. Aquaculture 1999;172:125e45.
[31] Chevreux B, Pfisterer T, Drescher B, Driesel AJ, Muller WE, et al. Using themiraEST assembler for reliable and automated mRNA transcript assembly andSNP detection in sequenced ESTs. Genome Res 2004;14:1147e59.
[32] Patterton HG, Graves S. DNAssist: the integrated editing and analysis of mo-lecular biology sequences in windows. Bioinform Appl Note 2000;16:652e3.
[33] Pearson WR. Rapid and sensitive sequence comparison with FASTP andFASTA. Meth Enzymol 1990;183:63e98.
[34] Roy A, Kucukural A, Zhang Y. I-TASSER: a unified platform for automatedprotein structure and function prediction. Nat Prot 2010;5:725e38.
[35] Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform2008;9:40.
[36] Roy A, Yang J, Zhang Y. COFACTOR: an accurate comparative algorithm forstructure-based protein function annotation. Nucl Acids Res 2012;40:W471e7.
[37] Lovell SC, Davis IW, Arendall III WB, de Bakker PIW, Word JM, Prisant MG,et al. Structure validation by Calpha geometry: phi, psi and C-beta deviation.Proteins: Str Fun Gen 2002;50:437e50.
[38] Shen MY, Sali A. Statistical potential for assessment and prediction of proteinstructures. Protein Sci 2006;15:2507e24.
[39] Pace CN, Vajdos F, Fee L, Grimsley G, Gray T. How to measure and predict themolar absorption coefficient of a protein. Protein Sci 1995;4:2411e23.
characterization and gene expression in response to viral and bacterial in-fections. Microbiol Res. http://dx.doi.org/10.1016/j.micres.2013.04.007; 2013.
[43] Livak KJ, Schmittgenm TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(�Delta Delta C(T)) method. Methods2001;25:402e8.
[44] Arockiaraj J, Easwvaran S, Vanaraja P, Singh A, Othman RY, Bhassu S. Immu-nological role of thiol-dependent peroxiredoxin gene in Macrobrachiumrosenbergii. Fish Shellfish Immunol 2012;33:121e9.
[45] Arockiaraj J, Avin FA, Vanaraja P, Easwvaran S, Singh A, Othman RY, et al.Immune role of MrNFkBI-a, an IkB family member characterized in prawnM. rosenbergii. Fish Shellfish Immunol 2012;33:619e25.
[46] Arockiaraj J, Gnanam AJ, Muthukrishnan D, Pasupuleti M, Milton J, Singh A. Anupstream initiator caspase 10 of snakehead murrel Channa striatus, containingDED, p20 and p10 subunits: molecular cloning, gene expression and proteo-lytic activity. Fish Shellfish Immunol 2013;34:505e13.
[47] Bradford MM. A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of proteinedye binding. AnalBiochem 1976;72:248e54.
[48] Hikima J, Minagawa S, Hirono I, Aoki T. Molecular cloning, expression andevolution of the Japanese flounder goose-type lysozyme gene, and the lyticactivity of its recombinant protein. Biochim Biophys Acta 2001;1520:35e44.
[49] Morgan BA, Mittman BA, Smith MM. The highly conserved N-terminal do-mains of histones H3 and H4 are required for normal cell cycle progression.Mol Cell Biol 1991;11:4111e20.
Please cite this article in press as: Arockiaraj J, et al., An unconventional arosenbergii: Analysis of immune properties, Fish & Shellfish Immunology
[51] Papagianni M. Ribosomally synthesized peptides with antimicrobial proper-ties: biosynthesis, structure, function, and applications. Biotechnol Adv2003;21:465e99.
[52] Tagai C, Morita S, Shiraishi T, Miyaji K, Iwamuro S. Antimicrobial properties ofarginine- and lysine-rich histones and involvement of bacterial outer mem-brane protease T in their differential mode of actions. Peptides 2011;32:2003e9.
[53] Balicki D, Putnam CD, Scaria PV, Beutler E. Structure and function correlationin histoneH2A peptide-mediated gene transfer. Proc Natl Acad Sci USA2002;99:7467e71.
[54] Xiong M, Waterman MS. A phase transition for the minimum free energy ofsecondary structures of a random RNA. Adv Appl Math 1997;18:111e32.
[55] Zhou JG, Wei JG, Xu D, Cui HC, Yan Y, Ou-Yang ZL, et al. Molecular cloning andcharacterization of two novel hepcidins from orange-spotted grouper, Epi-nephelus coioides. Fish Shellfish Immunol 2011;30:559e68.
[56] Corzo G, Villegas E, Gómez-Lagunas F, Possani LD, Belokoneva OS, Nakajima T.Oxyopinins, large amphipathic peptides isolated from the venom of the wolfspider Oxyopes kitabensis with cytolytic properties and positive insecticidalcooperativity with spider neurotoxins. J Biol Chem 2002;277:23627e37.
[57] Vassilevski AA, Kozlov SA, Samsonova OV, Egorova NS, Karpunin DV,Pluzhnikov KA, et al. Cyto-insectotoxins, a novel class of cytolytic and insec-ticidal peptides from spider venom. Biochem J 2008;411:687e96.
1448
ntimicrobial protein histone from freshwater prawn Macrobrachium(2013), http://dx.doi.org/10.1016/j.fsi.2013.08.018