Page 1
ORIGINAL PAPER
Analysis of Binding Interaction Between AntibacterialCiprofloxacin and Human Serum Albumin by SpectroscopicTechniques
Ankita Varshney • Yunus Ansari • Nida Zaidi •
Ejaz Ahmad • Gamal Badr • Parvez Alam •
Rizwan Hasan Khan
� Springer Science+Business Media New York 2014
Abstract The binding of ciprofloxacin (CFX) to human
serum albumin (HSA) has been investigated by fluorescence
displacement and induced circular dichroism (ICD) mea-
surements. Displacement measurements were performed
with CFX in the absence and presence of marker ligands
(hemin for domain I, bilirubin for interspace of domain IA
and IIA, chloroform for domain II, and diazepam for domain
III) to establish CFX binding site in one of the three major
domains of HSA. The primary binding site of CFX is located
in site I of HSA (domain IIA) in close vicinity to the site
where chloroform (CHCl3) binds. It is depicted from the
decrease in quenching constant of HSA–CHCl3 system
(0.02 ± 0.06) 9 10-3 L mol-1 compared to HSA–CFX–
CHCl3 system (0.01 ± 0.06) 9 10-3 L mol-1 as obtained
by the fluorescence displacement spectroscopy. Further-
more, far-UV CD results show that the binding of CFX leads
to change in the helicity of HSA. The ICD results indicated
that the CFX binds to the domain IIA of HSA which is in
agreement with the fluorescence displacement results.
Keywords Ciprofloxacin (CFX) � Binding parameters �Circular dichroism � Human serum albumin
Introduction
Human serum albumin (HSA) is most abundant serum
protein in humans. It binds and transport a large variety of
ligands including hormones, fatty acids, drugs, etc. [1–5]. It
is a globular, multifunctional protein composed of three
structurally similar domains each containing two subdo-
mains and having molecular weight of 67 kD stabilized by
17 disulfide bonds [6–8]. Apart from ligands binding and
transport, it involves in maintaining pH and osmotic
pressure, preventing photodegradation of folic acid, and is
also a marker of inflammatory state [9, 10].
The group of fluoroquinolones is one of the most suc-
cessful classes of antibacterial drugs. These compounds are
of exceeding interest because their clinical role has greatly
expanded since they were introduced in the 1980s. One of
these quinolones, ciprofloxacin (CFX), has in vitro activity
against a wide range of Gram-negative and Gram-positive
microorganisms. The mechanism of action of quinolones,
including CFX, is different from that of other antimicrobial
agents such as beta-lactams, macrolides, tetracyclines, and
aminoglycosides; therefore, organisms resistant to these
drugs may be susceptible to CFX. There is no known cross-
resistance between CFX and other classes of antimicrobials.
Notably, the drug has 100 times higher affinity for bacterial
DNA gyrase than for mammalian. This fluoroquinolone has
been applied in the empirical treatment of a variety of
infections, particularly those of genitourinary, gastrointes-
tinal, and respiratory tracts [11]. Chemically, CFX is a
1-cyclopropyl-6-fluoro-4-oxo-7-piperazin-1-yl-quinolone-
3-carboxylic acid (Fig. 1). Since it has an extended aromatic
part and functional groups suitable for hydrogen bonding, it
can be expected that this phenolic type molecule is able to
interact strongly with biomacromolecules and that these non-
covalent interactions may play a decisive role in its
A. Varshney � Y. Ansari � N. Zaidi � E. Ahmad � P. Alam �R. H. Khan (&)
Interdisciplinary Biotechnology Unit, Aligarh Muslim
University, Aligarh 202002, U.P., India
e-mail: [email protected] ; [email protected]
G. Badr
Laboratory of Immunology & Molecular Biology, Zoology
Department, Faculty of Science, Assiut University,
Assiut 71516, Egypt
123
Cell Biochem Biophys
DOI 10.1007/s12013-014-9863-1
Page 2
mechanism of action. Because of its pharmacological
activity, the investigation of the interactions between this
compound and serum albumins is very important [12]. Fur-
thermore, binding of drugs to albumin alters the pattern and
volume of distribution, lowers the rate of clearance, and
increases the plasma half-life of the drug [12–15]. Since CFX
is practically insoluble in water (*1.1 mg L-1) at neutral
pH and rapidly decomposes in alkaline solution, its binding
to serum albumin is very important to exert the beneficial
therapeutic activities.
Protein binding has long been considered one of the
most important physicochemical characteristics of drugs,
playing a potential role in distribution, excretion, and
therapeutic effectiveness [16]. The multiplicity of binding
sites on HSA for endogenous and exogenous small mole-
cules makes it difficult to assess interactions, whether
competitive or cooperative, between different ligands
bound to the protein. The flexible structural organization
allows the protein structure to adapt to a variety of ligands.
However, it is important to address this issue in order to
obtain a fuller description of the ligand-binding properties
of HSA [8, 17, 18]. As conformational adaptability of HSA
extends well beyond the immediate vicinity of the binding
site(s), cooperativity and allosteric modulation arise among
binding sites; this makes HSA a multimeric protein.
In this study to explore the binding of CFX to HSA,
quenching of tryptophan fluorescence was carried out.
Furthermore far-UV CD spectroscopy was employed to
confirm the secondary structural changes upon CFX bind-
ing to HSA. The probable binding site of CFX on HSA is
also predicted from marker displacement experiment.
Materials and Methods
Materials
HSA (A1887; [96 %), CFX (17850; [98.0 %), warfarin
(A2250; [98 %), hemin (Hem) (H5533; [80 %), diaze-
pam (DIA) (D0406; [98 %), and chloroform (C2432;
>99.5 %) purchased from Sigma-Aldrich. Bilirubin (BR)
was purchased from Hemedia. The number in the paren-
thesis corresponds to the purity of the compounds. All of
the other reagents were of analytical grade.
Methods
Protein Concentration Determination
Protein concentration was determined spectrophotometri-
cally using E1cm of 5.30 at 280 nm [19] on a Hitachi
spectrophotometer, model U-1500 or alternately by the
method of Lowry et al. [20].
Sample Preparation
HSA and drug solutions were prepared in 20 mM sodium
phosphate buffer (pH 7.4). HSA was passed through Seph-
acryl-S200 gel filtration column and dialyzed. Site markers
for HSA were also prepared in 20 mM phosphate buffer and
ethanol [ethanol concentration did not exceed 5 % (v/v)] and
their concentration was calculated appropriately. All the
solutions were prepared by weight/volume (w/v).
Binding Displacement Measurement Using Site
Markers
Different site markers, Hem for site in subdomain IA [21],
DIA for site II (subdomain III A) [22, 23], chloroform
(CHCl3) for site in subdomain 1IA [24], and BR for site in
interspace of subdomain IA and IIA [25], were used for
performing displacement experiments. The titration of CFX
was carried out to the solution having protein and site marker
in the ratio of 1:1. The fluorescence emission spectra were
recorded in the 300–400 nm range after exciting at 295 nm.
The binding constant values of drug–protein–marker were
evaluated using Stern–Volmer equation.
Fluorescence Quenching Measurement of HSA
Fluorescence measurements were performed on a Shima-
dzu spectrofluorimeter, model RF-5301 PC. The fluores-
cence spectra were measured at 25 ± 0.1 �C with a 1 cm
path length cell. Both excitation and emission slits were set
at 3 nm. Intrinsic fluorescence was measured by exciting
the protein solution at 295 nm and emission spectra were
recorded in the range of 300–400 nm.
Calculations/Data Analysis
The quenching equation is presented by
F0=F ¼ 1 þ kqs0 Q½ � ¼ 1 þ Ksv Q½ �; ð1Þ
Fig. 1 Chemical structure of ciprofloxacin
Cell Biochem Biophys
123
Page 3
where F and F0 are the fluorescence intensities with and
without quencher, respectively, kq is the quenching rate
constant of the biomolecule, Ksv is the Stern–Volmer
quenching constant, s0 is the average lifetime of the bio-
molecule without ligand, and [Q] is the concentration of
the quencher used. Fluorescence quenching data of HSA
complexed with markers in the absence and presence of
CFX were analyzed to obtain various binding parameters.
The binding constant (Kb) and binding affinity were
calculated according to the given equation
Log ½ðF0 � FÞ=F� ¼ log Kb þ n log Q½ �; ð2Þ
where F0 and F are the fluorescence intensities with and
without the ligand, respectively.
A plot of log [(Fo - F)/F] versus log [Q] gave a straight
line using least-squares analysis whose slope was equal to n
(binding affinity) and the intercept on Y-axis to logK
(K = binding constant). The binding constant (K) thus
obtained was used to calculate the standard free energy change
DG0binding of the ligand binding to HSA from the relationship
DG0binding ¼ �2:303 RT ln Kb: ð3Þ
Circular Dichroism Spectroscopy
Circular dichroism (CD) was performed on a Jasco J-715
spectropolarimeter at 25 ± 0.2 �C, in a rectangular cell with
1.0 cm path length equipped with magnetic stirring. Each
spectrum was signal-averaged at least three times with a
bandwidth of 1.0 nm and a resolution of 0.5 nm at a scan
speed of 100 nm min-1. Induced CD (ICD) spectra resulting
from the interaction of the drug with HSA were obtained by
subtracting the CD spectrum of the protein from that of the
complex.
Results and Discussions
Binding of Hemin in the Absence and Presence
of Ciprofloxacin
X-ray crystal structure of HSA–Hem complex has shown a
single binding site for Hem on domain I [18, 19]. Hem is a
large planar molecule and can be used as a probe for moni-
toring the effect of drug on the binding properties of domain
I. The fluorescence quenching spectra of HSA at various
concentrations of Hem in the absence and presence of CFX
are shown in Fig. 2a, b and the data are summarized in
Tables 1 and 2. Equilibration of HSA with Hem caused
concentration-dependent quenching in the intrinsic fluores-
cence intensity, which suggests the binding of Hem to HSA.
A little decrease in association constant and almost no
change in the binding sites n were observed in the presence of
CFX. On the other hand, stability of HSA–Hem complex
(Kb = 3.01 9 108 M-1, DG0 = -48.30 kJ mol-1) decreases
compared to the HSA–CFX–Hem complex (Kb = 29.37 9
108 M-1, DG0 = -53.93 kJ mol-1). These results suggest
that the presence of CFX did not affect the binding of Hem to
domain IB.
Binding of Bilirubin in the Absence and Presence
of Ciprofloxacin
The binding of BR, a toxic metabolite of heme, to HSA
[18, 26] has been studied extensively for many years. A
number of studies that measured the affinity of proteolytic
fragments of HSA for BR showed that the high-affinity
BR-binding site was located near subdomain IIA. There is,
therefore, great clinical interest in understanding the
binding of BR to albumin and the effects of drugs and other
competitors on this binding. Aliquots of BR to the protein
solution were added in the absence and presence of CFX,
and decrease in protein fluorescence were measured after
each addition of BR. The fluorescence intensity of HSA
decreased regularly, and slight blue shift was observed for
the emission wavelength with increasing BR concentration
up to BR/albumin molar ratio of 0–10, indicating that the
presence of BR could quench the fluorescence of HSA–
CFX complex. Furthermore, determining the various
binding parameters (Fig. 3a, b; Tables 1, 2) depicts clearly
that there was no competitive binding between CFX and
BR since we observed very less change in the binding
pattern when two ligands were allowed to bind. We
observed decreased stability of HSA–BR complex
(Kb = 0.1088 9 108 M-1, DG0 = -40.08 kJ mol-1)
when compared to HSA–CFX–BR complex (Kb = 20.73 9
108 M-1, DG0 = -53.07 kJ mol-1).
Binding of Chloroform in the Absence and Presence
of Ciprofloxacin
The site of action of the volatile general anesthetics
remains controversial, but evidence in favor of its binding
to subdomain IIA of HSA is accumulating. In this study, in
the absence and presence of CFX, binding to chloroform to
HSA is monitored by fluorescence quenching measure-
ments. Chloroform causes a decrease in the fluorescence
emission quantum yield as shown in Fig. 4a, b. A slight
blue shift of 2 nm in the emission wavelength maximum
was observed, suggesting that the binding of chloroform is
associated with the changes in the dielectric environment
of the indole ring in HSA, because electron transfer from
the excited indole ring to chloroform might be responsible
for the observed fluorescence quenching. Figure 4 shows
the Stern–Volmer plot from the slope of which
Cell Biochem Biophys
123
Page 4
Ksv = 0.02 ± 0.08 M-1 was calculated. It is observed that
Ksv = 0.02 ± 0.08 M-1 was reduced to almost half
(0.01 ± 0.08 M-1) in the presence of CFX which is sug-
gestive of static interaction between fluorophore and CFX.
Furthermore a little increase in the binding site of HSA n in
the absence and presence of CFX depict that an allosteric
binding exists on domain II for both the CFX and chloro-
form (Fig. 4; Tables 1, 2).
The approach used allows direct monitoring of anes-
thetic binding to the protein and, in addition, provides
information about the location of the anesthetic in the
protein matrix. The results indicate that chloroform occu-
pies the same binding site on this model mammalian pro-
tein, HSA, as that occupied by CFX.
0
50
100
150
200
250
Wavelength (nm)F
luor
esce
nce
Inte
nsit
y
0
50
100
150
200
250
Wavelength (nm)
Flu
ores
cenc
e In
tens
ity
Fo/
F
0
2
4
6
8
Hemin [ M]
C
-1.2
-0.8
-0.4
0
0.4
0.8
1.2
300 350 400 300 350 400
0 1 2 3 4 -7 -6.5 -6 -5.5 -5
Log [Hemin]
D
Log
[(F
o-F
)/F
]
BA
µ
Fig. 2 a Fluorescence spectra
of hemin quenching in the
absence of CFX. b Fluorescence
spectra of hemin quenching in
the presence of CFX. c Stern–
Volmer plots of hemin
quenching in the absence (open
circle) and presence (filled
circle) of CFX. d Log [(F0 -
F)/F] versus Log [Q] plots of
hemin quenching in the absence
(open circle) and presence
(filled circle) of CFX for
determining the binding
constant and binding sites
Table 1 Effects of domain-specific ligands on binding constants of HSA and HSA–CFX systems
Ligands Systems Ksv 9 106 [L mol-1]a Kq 9 1014 [L mol-1 s-1]a R2
Hemin HSA 1.77 1.77 0.9917
HSA–CFX 1.40 1.40 0.9895
Bilirubin HSA 0.4 0.4 0.9902
HSA–CFX 0.2 0.2 0.9572
Chloroform HSA 0.02 9 10-3 0.02 9 10-3 0.9992
HSA–CFX 0.01 9 10-3 0.01 9 10-3 0.9951
Diazepam HSA 0.09 0.092 0.9962
HSA–CFX 0.06 0.06 0.9969
a The mean value of 4 individual experiments with standard deviation ±0.06–±0.11 %
Table 2 Thermodynamic and binding parameters of HSA and HSA–
CFX system
Ligand Systems Kb [M-1]a n DG0 [kJ mol-1]
Hemin HSA 3.01 9 108 1.40 -48.30
HSA–CFX 29.37 9 108 1.60 -53.93
Bilirubin HSA 0.1088 9 108 1.24 -40.08
HSA–CFX 20.73 9 108 1.68 -53.07
Chloroform HSA 18.13 1.03 -7.16
HSA–CFX 11.98 1.08 -6.14
Diazepam HSA 3.48 9 104 0.93 -25.87
HSA–CFX 3.09 9 104 0.94 -25.57
a The mean value of 4 individual experiments with standard deviation
±0.06–±0.11 %
Cell Biochem Biophys
123
Page 5
0
80
160
240
320
300 350 4000
50
100
150
200
250
0.8
1.2
1.6
2
C
-2
-1.6
-1.2
-0.8
-0.4
0
300 350 400
0 0.5 1 1.5 2 2.5 -6.6 -6.4 -6.2 -6 -5.8 -5.6
Log
[(F
o-F
)/F
]
D
Flu
ores
cenc
e In
tens
ity
Flu
ores
cenc
e In
tens
ity
Wavelength [nm] Wavelength [nm]
a
j
a
j
Fo/
F
Bilirubin [ M] Log [Bilirubin]
A B
µ
Fig. 3 a Fluorescence spectra
of bilirubin quenching in the
absence of CFX. b Fluorescence
spectra of bilirubin quenching in
the presence of CFX. c Stern–
Volmer plots of bilirubin
quenching in the absence (open
circle) and presence (filled
circle) of CFX. d Log [(F0 -
F)/F] versus Log [Q] plots of
bilirubin quenching in the
absence (open circle) and
presence (filled circle) of CFX
for determining the binding
constant and binding sites
0
20
40
60
80
100
-1.6
-1.2
-0.8
-0.4
0
0.4
0
20
40
60
80
100
300 350 400
Wavelength [nm] Wavelength [nm]
Flu
ores
cenc
e In
tens
ity
Flu
ores
cenc
e In
tens
ity
a
j
a
j
0.8
1.2
1.6
2
2.4
300 350 400
-2.2 -1.8 -1.4 -10 20 40 60 80 100
CHCl3 [mM] Log [CHCl3]
Fo/
F
Log
[(F
o-F
)/F
]
BA
DC
Fig. 4 a Fluorescence spectra
of chloroform quenching in the
absence of CFX. b Fluorescence
spectra of chloroform
quenching in the presence of
CFX. c Stern–Volmer plots of
chloroform quenching in the
absence (open circle) and
presence (filled circle) of CFX.
d Log [(F0 - F)/F] versus Log
[Q] plots of chloroform
quenching in the absence (open
circle) and presence (filled
circle) of CFX for determining
the binding constant and
binding sites
Cell Biochem Biophys
123
Page 6
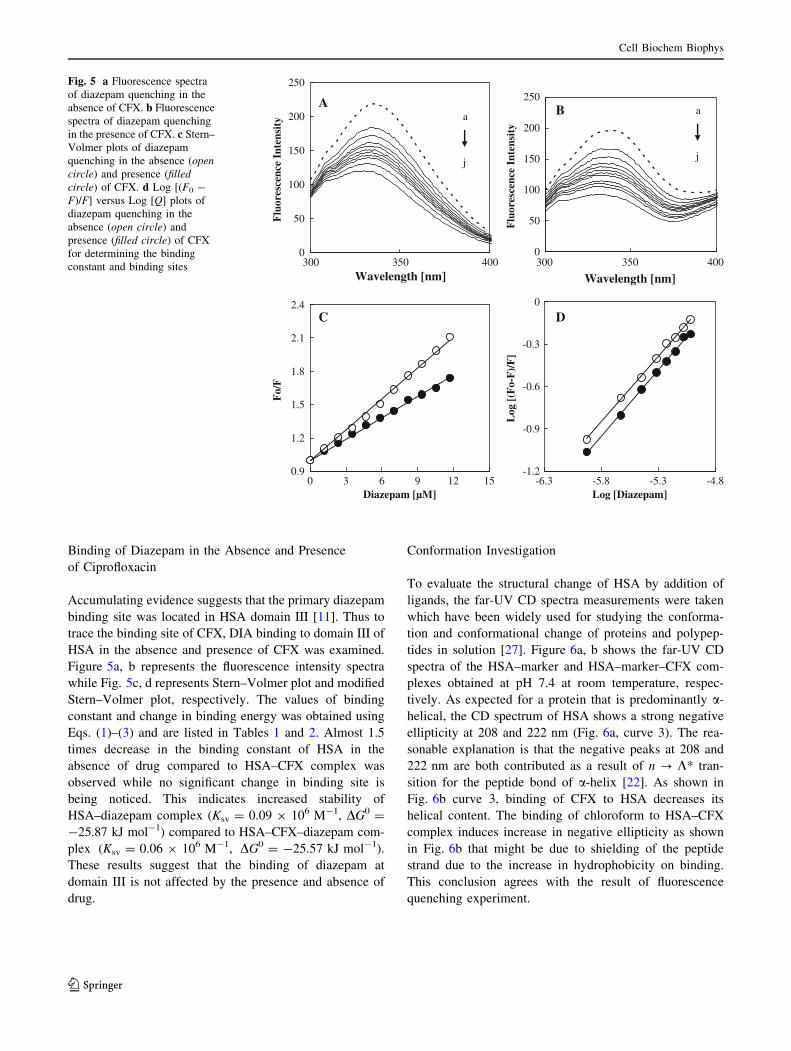
Binding of Diazepam in the Absence and Presence
of Ciprofloxacin
Accumulating evidence suggests that the primary diazepam
binding site was located in HSA domain III [11]. Thus to
trace the binding site of CFX, DIA binding to domain III of
HSA in the absence and presence of CFX was examined.
Figure 5a, b represents the fluorescence intensity spectra
while Fig. 5c, d represents Stern–Volmer plot and modified
Stern–Volmer plot, respectively. The values of binding
constant and change in binding energy was obtained using
Eqs. (1)–(3) and are listed in Tables 1 and 2. Almost 1.5
times decrease in the binding constant of HSA in the
absence of drug compared to HSA–CFX complex was
observed while no significant change in binding site is
being noticed. This indicates increased stability of
HSA–diazepam complex (Ksv = 0.09 9 106 M-1, DG0 =
-25.87 kJ mol-1) compared to HSA–CFX–diazepam com-
plex (Ksv = 0.06 9 106 M-1, DG0 = -25.57 kJ mol-1).
These results suggest that the binding of diazepam at
domain III is not affected by the presence and absence of
drug.
Conformation Investigation
To evaluate the structural change of HSA by addition of
ligands, the far-UV CD spectra measurements were taken
which have been widely used for studying the conforma-
tion and conformational change of proteins and polypep-
tides in solution [27]. Figure 6a, b shows the far-UV CD
spectra of the HSA–marker and HSA–marker–CFX com-
plexes obtained at pH 7.4 at room temperature, respec-
tively. As expected for a protein that is predominantly a-
helical, the CD spectrum of HSA shows a strong negative
ellipticity at 208 and 222 nm (Fig. 6a, curve 3). The rea-
sonable explanation is that the negative peaks at 208 and
222 nm are both contributed as a result of n ? K* tran-
sition for the peptide bond of a-helix [22]. As shown in
Fig. 6b curve 3, binding of CFX to HSA decreases its
helical content. The binding of chloroform to HSA–CFX
complex induces increase in negative ellipticity as shown
in Fig. 6b that might be due to shielding of the peptide
strand due to the increase in hydrophobicity on binding.
This conclusion agrees with the result of fluorescence
quenching experiment.
0
50
100
150
200
250
0
50
100
150
200
250
300 350 400
-1.2
-0.9
-0.6
-0.3
0
Wavelength [nm]Wavelength [nm]F
luor
esce
nce
Inte
nsit
y
Flu
ores
cenc
e In
tens
ity
a
j
a
j
0.9
1.2
1.5
1.8
2.1
2.4
300 350 400
-6.3 -5.8 -5.3 -4.80 3 6 9 12 15
Fo/
F
Log
[(F
o-F
)/F
] Diazepam [µM] Log [Diazepam]
A B
C D
Fig. 5 a Fluorescence spectra
of diazepam quenching in the
absence of CFX. b Fluorescence
spectra of diazepam quenching
in the presence of CFX. c Stern–
Volmer plots of diazepam
quenching in the absence (open
circle) and presence (filled
circle) of CFX. d Log [(F0 -
F)/F] versus Log [Q] plots of
diazepam quenching in the
absence (open circle) and
presence (filled circle) of CFX
for determining the binding
constant and binding sites
Cell Biochem Biophys
123
Page 7
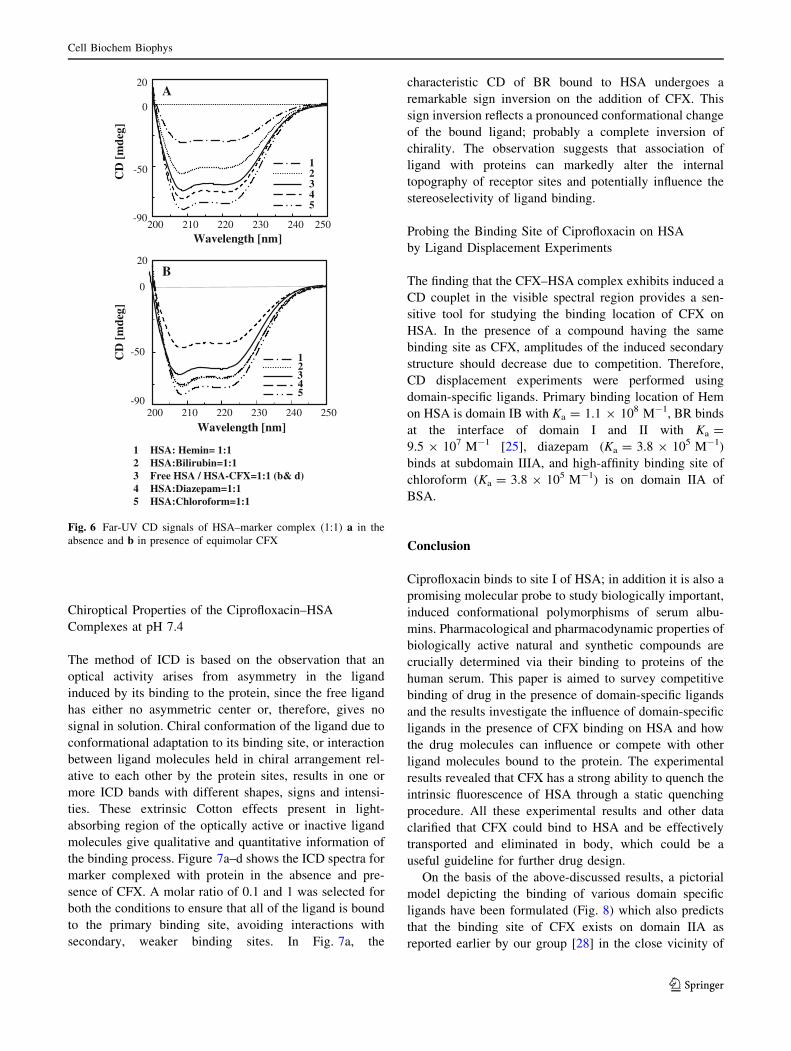
Chiroptical Properties of the Ciprofloxacin–HSA
Complexes at pH 7.4
The method of ICD is based on the observation that an
optical activity arises from asymmetry in the ligand
induced by its binding to the protein, since the free ligand
has either no asymmetric center or, therefore, gives no
signal in solution. Chiral conformation of the ligand due to
conformational adaptation to its binding site, or interaction
between ligand molecules held in chiral arrangement rel-
ative to each other by the protein sites, results in one or
more ICD bands with different shapes, signs and intensi-
ties. These extrinsic Cotton effects present in light-
absorbing region of the optically active or inactive ligand
molecules give qualitative and quantitative information of
the binding process. Figure 7a–d shows the ICD spectra for
marker complexed with protein in the absence and pre-
sence of CFX. A molar ratio of 0.1 and 1 was selected for
both the conditions to ensure that all of the ligand is bound
to the primary binding site, avoiding interactions with
secondary, weaker binding sites. In Fig. 7a, the
characteristic CD of BR bound to HSA undergoes a
remarkable sign inversion on the addition of CFX. This
sign inversion reflects a pronounced conformational change
of the bound ligand; probably a complete inversion of
chirality. The observation suggests that association of
ligand with proteins can markedly alter the internal
topography of receptor sites and potentially influence the
stereoselectivity of ligand binding.
Probing the Binding Site of Ciprofloxacin on HSA
by Ligand Displacement Experiments
The finding that the CFX–HSA complex exhibits induced a
CD couplet in the visible spectral region provides a sen-
sitive tool for studying the binding location of CFX on
HSA. In the presence of a compound having the same
binding site as CFX, amplitudes of the induced secondary
structure should decrease due to competition. Therefore,
CD displacement experiments were performed using
domain-specific ligands. Primary binding location of Hem
on HSA is domain IB with Ka = 1.1 9 108 M-1, BR binds
at the interface of domain I and II with Ka =
9.5 9 107 M-1 [25], diazepam (Ka = 3.8 9 105 M-1)
binds at subdomain IIIA, and high-affinity binding site of
chloroform (Ka = 3.8 9 105 M-1) is on domain IIA of
BSA.
Conclusion
Ciprofloxacin binds to site I of HSA; in addition it is also a
promising molecular probe to study biologically important,
induced conformational polymorphisms of serum albu-
mins. Pharmacological and pharmacodynamic properties of
biologically active natural and synthetic compounds are
crucially determined via their binding to proteins of the
human serum. This paper is aimed to survey competitive
binding of drug in the presence of domain-specific ligands
and the results investigate the influence of domain-specific
ligands in the presence of CFX binding on HSA and how
the drug molecules can influence or compete with other
ligand molecules bound to the protein. The experimental
results revealed that CFX has a strong ability to quench the
intrinsic fluorescence of HSA through a static quenching
procedure. All these experimental results and other data
clarified that CFX could bind to HSA and be effectively
transported and eliminated in body, which could be a
useful guideline for further drug design.
On the basis of the above-discussed results, a pictorial
model depicting the binding of various domain specific
ligands have been formulated (Fig. 8) which also predicts
that the binding site of CFX exists on domain IIA as
reported earlier by our group [28] in the close vicinity of
200 250 210 220 230 240 -90
-50
0
20
21
345
B
0
200 210 220 230 240-90
-50
20
12345
250
A
CD
[m
deg]
1 HSA: Hemin= 1:1 2 HSA:Bilirubin=1:1 3 Free HSA / HSA-CFX=1:1 (b& d) 4 HSA:Diazepam=1:1 5 HSA:Chloroform=1:1
CD
[m
deg]
Wavelength [nm]
Wavelength [nm]
Fig. 6 Far-UV CD signals of HSA–marker complex (1:1) a in the
absence and b in presence of equimolar CFX
Cell Biochem Biophys
123
Page 8
the site where chloroform binds. Our work not only pro-
vides the multiplicity of binding sites on HSA and dem-
onstrates the conformational plasticity of HSA on drug
binding, but it may also provide structural information for
the modulation of HSA–drug interaction by various spec-
troscopic approaches based on HSA–drug interaction.
The binding of small molecules to proteins and protein–
protein interactions are key processes in cell biochemistry.
-13
12
-10
0
320250 260 280 300
DI: HSA=0.1:1 DI: CFX: HSA=0.1:1:1
DI: CFX: HSA=1:1:1 DI: HSA=1:1
C
-6
-4
-2
0
2
4
350
A
HE: HSA=0.1:1 HE: CFX: HSA=0.1:1:1
HE: CFX: HSA=0.1:1:1 HE: HSA=1:1
CD
[m
deg]
Wavelength [nm]
CD
[m
deg]
Wavelength [nm]
-40
-20
0
20
40
CD
[m
deg]
BR: HSA=0.1:1 BR: CFX: HSA=0.1:1:1 BR: HSA=1:1 BR: CFX: HSA=1:1:1
B
Wavelength [nm]
-6
-3
0
3
6
9
12
400 450 500 350 400 450 500 550
300 350 400 450 500
D
CHCl3: HSA=0.1:1 CHCl3: CFX: HSA= 0.1:1:1
CHCl3: CFX: HSA= 1:1:1 CHCl3: HSA= 1:1
Wavelength [nm]
CD
[m
deg]
Fig. 7 ICD (expressed in
mdeg) of marker ligands: hemin
(a), bilirubin (b), diazepam (c),
and chloroform (d) bound to
HSA and HSA–CFX at a molar
ratio of 0.1 and 1. Line symbols
are shown in inset. Protein
concentration was 10 lM in
0.06 M sodium phosphate
buffer, pH 7.4
Fig. 8 A pictorial model
indicating the binding of
domain-specific ligands and
predicting the binding site of
ciprofloxacin on human serum
albumin
Cell Biochem Biophys
123
Page 9
The usual paradigm is that ligand binding induces a change
in the conformation of the target protein which, in turn,
produces a given response and fundamental importance.
Using the binding of various domain-specific ligands to
HSA, we have concluded that CFX interacts with domain II
of HSA in the close vicinity to chloroform.
Recent advances of gene cloning, together with com-
plete understanding of albumin structure and function,
provide for a greater abundance of future applications.
Accordingly, our information on the relative stability of
HSA and its domains should provide a basis for drug
design, as they possess binding sites for a variety of
exogenous and endogenous ligands.
Acknowledgments Facilities provided by A.M.U are gratefully
acknowledged. Ankita Varshney, Ejaz Ahmad, Nida Zaidi, and Par-
vez Alam thank the Council of Scientific and Industrial Research,
New Delhi and Yunus Ansari thanks the department of Biotechnol-
ogy, Govt. of India for financial assistance.
References
1. Bhattacharya, A. A., Grune, T., & Curry, S. (2000). Crystallo-
graphic analysis reveals common modes of binding of medium
and long-chain fatty acids to human serum albumin. Journal of
Molecular Biology, 303, 721–732.
2. Varshney, A., Sen, P., Ahmed, E., Rehan, M., & Subbarao, N.
(2010). Ligand binding strategies of human serum albumin: how
can the cargo be utilized. Chirality, 22, 77–87.
3. Gulam, J., Zunszain, P. A., Petipas, I., Bhattacharaya, A. A.,
Otagiri, M., & Curry, S. (2005). Structural basis of the drug-
binding specificity of human serum albumin. Journal of Molec-
ular Biology, 353, 38–52.
4. Zaidi, N., Ahmad, E., Rehan, M., Rabbani, G., Ajmal, M. R.,
Zaidi, Y., et al. (2013). Biophysical insight into furosemide
binding to human serum albumin: a study to unveil its impaired
albumin binding in uremia. Journal of Physical Chemistry B, 117,
2595–2604.
5. Zaidi, N., Ajmal, M. R., Rabbani, G., Ahmad, E., & Khan, R. H.
(2013). A comprehensive insight into binding of hippuric acid to
human serum albumin: a study to uncover its impaired elimina-
tion through hemodialysis. PLoS One, 8, e71422.
6. Peters, T. (1996). All about albumin. Biochemistry, genetics and
medical applications. San Diego: Academic Press.
7. Gabor, M., & Fred, K. (1957). The disulfide bonds of human
serum albumin and bovine c-globulin. Journal of American
Chemical Society, 79, 134–139.
8. Sugio, S., Kashima, A., Mockizuki, S., Noda, M., & Kobayashi,
K. (1999). Crystal structure of human serum albumin at 2.5 A
resolution. Protein Engineering, 12, 439–446.
9. Vorobey, P., Steindal, A. E., Vorobey, A., & Moan, J. (2006).
Influence of human serum albumin on photodegradation of folic
acid in solution. Journal of Photochemistry Photobiology, 82,
817–822.
10. Bhagvan, N. V., & Ha, C. E. (2013). Novel insight into the
pleiotropic effects of human serum albumin in health and disease.
Biochemica et Biophysica Acta General subjects, 12, 5486–5493.
11. Lode, H. (1999). Potential interactions of the extended spectrum
fluoroquinolones with the CNS. Drug Safety, 2, 123–135.
12. Benet, L. Z., & Hoener, B. A. (2002). Changes in plasma protein
binding have little clinical relevance. Clinical Pharmacology
Therapeutics, 71, 115–121.
13. Herve, F., Urien, S., Albengres, E., Duche, J. C., & Tillement, J.
P. (1994). Drug binding in plasma. A summary of recent trends in
the study of drug and hormone binding. Clinical Pharmacoki-
netics, 26, 44–58.
14. Levine, R. R. (2000). Pharmacology: Drug actions and reactions
(6th ed.). New York: The Parthenon Publishing Group.
15. Pratt, W. B., & Taylor, P. (1990). Principles of drug action: The
basis of pharmacology (3rd ed.). New York: Churchill Living-
stone Inc.
16. Khan, M. Y. (1986). Direct evidence for the involvement of
domain III in the N-F transition of bovine serum albumin. Bio-
chemical Journal, 236, 307–310.
17. Curry, S., Mandelkow, H., Brick, P., & Franks, N. (1998). Crystal
structure of human serum albumin complexed with fatty acid
reveals an asymmetric distribution of binding sites. Nature
Structural Biology, 5, 827–835.
18. He, X. M., & Carter, D. C. (1992). Atomic structure and chem-
istry of human serum albumin. Nature, 358, 209–215.
19. Wallevik, K. (1973). Reversible denaturation of human serum
albumin by pH, temperature, and guanidine hydrochloride fol-
lowed by optical rotation. Journal of Biological Chemistry, 248,
2650–2655.
20. Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J.
(1951). Protein measurement with the Folin phenol reagent.
Journal of Biological Chemistry, 193, 265–275.
21. Zunszain, P. A., Ghuman, J., Komatsu, T., Tsuchida, E., & Curry,
S. (2003). Crystal structural analysis of human serum albumin
complexed with hemin and fatty acid. BMC Structural Biology, 3,
1–9.
22. Kratochwil, N. A., Huber, W., Muller, F., Kansy, M., & Gerber,
P. R. (2002). Predicting plasma protein binding of drugs: a new
approach. Biochemical Pharmacology, 64, 1355–1374.
23. Kang, J., Liu, Y., Xie, M. X., Li, S., Jiang, M., & Wang, Y. D.
(2004). Interactions of human serum albumin with chlorogenic
acid and ferulic acid. Biochimica Biophysica Acta, 1674,
205–214.
24. Johansson, J. S. (1997). Binding of the volatile anesthetic chlo-
roform to albumin demonstrated using tryptophan fluorescence
quenching. Journal of Biological Chemistry, 272, 17961–17965.
25. Kragh-Hansen, U. (1988). Evidence for a large and flexible
region of human serum albumin possessing high affinity binding
sites for salicylate, warfarin, and other ligands. Molecular
Pharmacology, 34, 160–171.
26. Silva, D., Cortez, C. M., Cunha-Bastos, J., & Louro, S. R. W.
(2004). Methyl parathion interaction with human and bovine
serum albumin. Toxicology Letters, 147, 53–61.
27. Kelly, S. M., & Price, N. N. (2000). The use of circular dichroism
in the investigation of protein structure and function. Current
Protein and Peptide Science, 1, 349–384.
28. Ahmad, B., Parveen, S., & Khan, R. H. (2006). Effect of albumin
conformation on the binding of ciprofloxacin to human serum
albumin: a novel approach directly assigning binding site. Bio-
macromolecules, 7, 1350–1356.
Cell Biochem Biophys
123