One of the oldest and most centralissues in psychological science, atten-tion is the process of selecting for ac-tive processing ideas stored in mem-ory in our minds, or aspects of ourphysical environment, such as ob-jects. The study of attention has be-come a huge enterprise; last yearalone about 300 articles were pub-lished on attention (Fig. 1). Through-out history, many great minds havewrestled with the definition of atten-tion. In 370 B.C., Aristotle regardedattention as a narrowing of the senses.Centuries later, William James (1890)contended that “everyone knows whatattention is. It is the taking possessionof the mind in clear and vivid form of
one out of what seem several simulta-neous objects or trains of thought.”James’ account strongly joins atten-tion with subjective experience. More-over, James’s addressing of attentionboth to objects and to “trains ofthought” is important for understand-ing current approaches to sensory ori-enting and executive control (Driverand Frackowiak, 2001; Fernandez-Duque and Posner, 2001; Fan et al.,2003a,b). However, as outlined below,
attention in the sense of orienting tosensory objects can actually be invol-untary and can occur unconsciously.So attention is not, as the quote fromWilliam James implies, the same asbeing aware (see Table 1 for a list ofimportant terms and definitions inthis field of study).
Following a lull in the field duringthe early 1900s, Donald Broadbent re-sumed the quest after World War II todiscover attentional mechanisms. Ap-
plying formal information theory,Broadbent likened attention to a filter.He proposed that attention wasbounded by the amount of informa-tion located between parallel sensorysystems and a limited-capacity per-ceptual system (Broadbent, 1958).This view facilitated objective studiesof the limitations of the human abilityto deal with multiple signals at a timein a variety of practical tasks.
As psychology moved toward thestudy of cognitive mechanisms, newobjective methods allowed for inves-tigation into the processes of selec-tion. For example, studies showedthat words could activate their se-mantic associates without aware-ness of the word’s identity (i.e.,priming). The parallel organizationof sensory information extends to se-mantic processing. The act of select-ing a word meaning for active atten-tion appears to suppress theavailability of other meanings of theselected item and of competingitems. Consequently, attention hassince been viewed more as a mecha-nism for providing priority for mo-tor acts than as a filter or bottleneck.For example, the premotor theorysuggests that attention is simply apreparation for response, or a select-ing of the goal of an intended action(Rizzolatti et al., 1987). The focus on
Dr. Raz is on the faculty of the MRI Unitin the Department of Psychiatry, Divi-sion of Child and Adolescent Psychiatry,at Columbia University College of Phy-sicians and Surgeons and New YorkState Psychiatric Institute.*Correspondence to: Amir Raz, MRIUnit, Department of Psychiatry, Divisionof Child and Adolescent Psychiatry, Co-lumbia University College of Physiciansand Surgeons and New York State Psy-chiatric Institute, 1051 Riverside Drive,Box 74, New York, NY 10032. Fax: 212-543-6660; E-mail: [email protected]
DOI 10.1002/ar.b.20035Published online in Wiley InterScience(www.interscience.wiley.com).
Attention is not, as thequote from William
James implies, the sameas being aware.
THE ANATOMICAL RECORD (PART B: NEW ANAT.) 281B:21–36, 2004
response may explain why atten-tional systems seem to differ forspace that is within reach and spacethat lies beyond. However, intendedactions are probably not the onlysources of capacity limits and atten-tional selection. After all, even whenwe have no intention to act and pas-sively watch scenes go by (e.g., at themovies), we still select only a subsetof the information that reaches thesenses. Attention, therefore, was fur-ther operationalized as facilitatingsome kinds of memory and evenconsciousness.
Other views construe attention inthe context of resource limitation/selection. Attentional limits are typ-ically found in unpracticed tasks.However, with practice, most atten-tion-demanding tasks can be ren-dered attention-independent, or au-tomatized. One example of thisphenomenon involves visuallysearching for a set of letters amongother random letters. This task ini-tially demands attention and givessteep slopes of latencies against dis-play size. However, after weeks ofpractice, the slopes become flat(Spelke et al., 1976; Schneider andShiffrin, 1977). Another examplecomes from a demonstration involv-ing two verbal tasks, reading forcomprehension and writing to dicta-
tion. These similar and demandingtasks can at first only be done in alter-nation, but after months of practice canbe efficiently combined (Spelke et al.,1976). Given these examples, theoriesof attentional limits must also accountfor how these limits can be eliminated(e.g., following practice) when the bot-tleneck is seemingly bypassed and thecorrect action is effortlessly performed.
GROSS CHARACTERISTICS OFATTENTION
Visual attention often serves as a con-venient lens to examine the character-istics of attention. Researchers andclinicians have investigated the optics,anatomy, development, pathology,and underlying neural processes of vi-sion, making it the most widely stud-
Figure 1. Research into Attention. Relativenumbers of publications for each of five keywords, normalized to their relative frequen-cies in the 1960-1969 decade. The 2000data span only the years 2000 to 2003. Therelative frequency values were computedby adding up all the papers that used eachof these keywords in 1960–1969 in PubMedand finding the proportion using each key-word. These were the base proportions. Therelative proportions were then the propor-tions in each subsequent decade dividedby the original proportions from 1960–1969.From Cavanagh (2003, 2004).
TABLE 1. Terms and definitions related to attentional network research
Foveate To look at directly so the image being viewedfalls on the fovea of the retina
Attention The mental ability to select stimuli, responses,memories, and thoughts that arebehaviorally relevant among a host of othersthat are behaviorally irrelevant
Attentional networks Neural circuits subserving attentionalprocessing, which preserve a degree ofanatomical and functional independencebut interact in many practical situations
Executive network The mechanism for monitoring and resolvingconflict among thoughts, feelings, andresponses; an attentional system concernedwith such tasks as working memory,planning, switching, and inhibitory control
Orienting The process of selecting information fromsensory input
Alerting The process of achieving and maintaining astate of high sensitivity to incoming stimuli
Top-downmodulation
A downstream effect such as cognitivecontrol, as opposed to a bottom-up effect
Cognitive control Processes such as conflict resolution, errorcorrection, inhibitory control, planning, andresource allocation
Self-regulation A key mediator between geneticpredisposition, early experience, and adultfunctioning (e.g., in controlling the reactionto stress, the capacity to maintain focusedattention or the ability to interpret mentalstates both internally and in others)
Neuroimaging Technologically advanced, often noninvasive,tools for tapping neurophysiological aspectsof the behaving brain
Mismatch negativity(MMN)
An electrophysiological manifestation ofinvoluntary preattentive processing occurringwhen some regularity in the auditory streamis violated by a change (deviant stimulus);the MMN is advantageous with regard to theneural processing of unattended input
Anterior cingulatecortex (ACC)
Part of the brain’s limbic system classicallyrelated to affect but also active in manystudies of cognition; the ACC might be thebrain’s error detection and correctiondevice part of a circuit involved in a form ofattention that serves to regulate bothcognitive and emotional processing
Functional magneticresonanceimaging (fMRI)
A noninvasive neuroimaging technique thatregisters blood flow to functioning areas ofthe brain in high magnetic fields
Positron emissiontomography (PET)
A neuroimaging technique measuring the flowof blood containing radioactive atoms thatemit positrons
22 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
ied perceptual system. Studying visualattention allows us to explore how wemove the attentional “beam” aroundto various areas of the visual field andchange the detail with which we lookat any given area. For example, it ispossible to look at this page and payattention to its setup as a whole, orconcentrate on specific words and cer-tain letters therein. Paying attention
to single characters permits us tocatch typos, glean information aboutpunctuation marks, and even spotminute imperfections on the physicalpaper. However, at this level of detail,we may miss the bigger idea conveyedin a paragraph. As we shift our focus,we can change the target location ofour attention or the size of our atten-tional field. Many metaphors describe
visual attention: “spotlight”, “zoomlens,” “gating,” and “gradient,” alongwith the common usage of terms suchas “attentional gaze” or “attentionalfocus” (Shalev and Algom, 2000). Thenotion of a spotlight, which startedout as a crude metaphor and has sincebeen taken up as a serious viewpointto be tested, is a natural derivative ofthe spatial milieu in which attention is
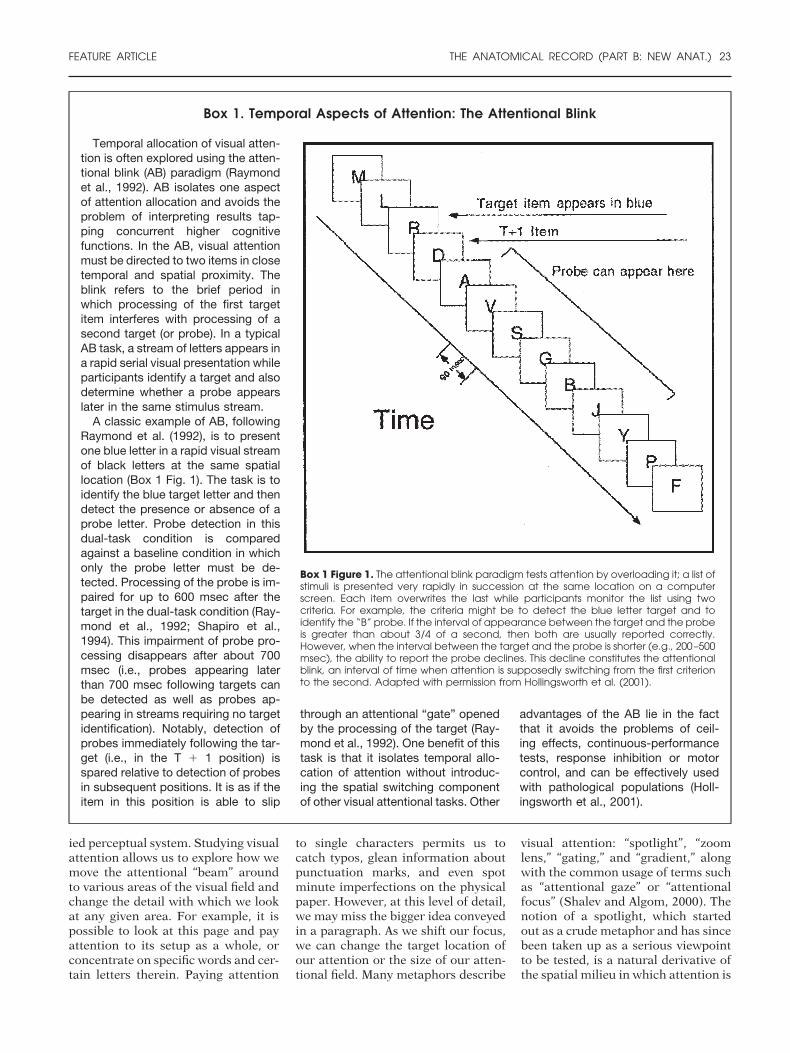
Box 1. Temporal Aspects of Attention: The Attentional Blink
Box 1 Figure 1. The attentional blink paradigm tests attention by overloading it; a list ofstimuli is presented very rapidly in succession at the same location on a computerscreen. Each item overwrites the last while participants monitor the list using twocriteria. For example, the criteria might be to detect the blue letter target and toidentify the “B” probe. If the interval of appearance between the target and the probeis greater than about 3/4 of a second, then both are usually reported correctly.However, when the interval between the target and the probe is shorter (e.g., 200–500msec), the ability to report the probe declines. This decline constitutes the attentionalblink, an interval of time when attention is supposedly switching from the first criterionto the second. Adapted with permission from Hollingsworth et al. (2001).
Temporal allocation of visual atten-tion is often explored using the atten-tional blink (AB) paradigm (Raymondet al., 1992). AB isolates one aspectof attention allocation and avoids theproblem of interpreting results tap-ping concurrent higher cognitivefunctions. In the AB, visual attentionmust be directed to two items in closetemporal and spatial proximity. Theblink refers to the brief period inwhich processing of the first targetitem interferes with processing of asecond target (or probe). In a typicalAB task, a stream of letters appears ina rapid serial visual presentation whileparticipants identify a target and alsodetermine whether a probe appearslater in the same stimulus stream.
A classic example of AB, followingRaymond et al. (1992), is to presentone blue letter in a rapid visual streamof black letters at the same spatiallocation (Box 1 Fig. 1). The task is toidentify the blue target letter and thendetect the presence or absence of aprobe letter. Probe detection in thisdual-task condition is comparedagainst a baseline condition in whichonly the probe letter must be de-tected. Processing of the probe is im-paired for up to 600 msec after thetarget in the dual-task condition (Ray-mond et al., 1992; Shapiro et al.,1994). This impairment of probe pro-cessing disappears after about 700msec (i.e., probes appearing laterthan 700 msec following targets canbe detected as well as probes ap-pearing in streams requiring no targetidentification). Notably, detection ofprobes immediately following the tar-get (i.e., in the T � 1 position) isspared relative to detection of probesin subsequent positions. It is as if theitem in this position is able to slip
through an attentional “gate” openedby the processing of the target (Ray-mond et al., 1992). One benefit of thistask is that it isolates temporal allo-cation of attention without introduc-ing the spatial switching componentof other visual attentional tasks. Other
advantages of the AB lie in the factthat it avoids the problems of ceil-ing effects, continuous-performancetests, response inhibition or motorcontrol, and can be effectively usedwith pathological populations (Holl-ingsworth et al., 2001).
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 23
assumed to operate and relates to thecommon experience concerning thekind of attention needed for readingversus proofreading. The bulk of theevidence suggests that attention canhave an influence quite early in thevisual system and scientists are stilllearning what visual operations atten-tion could potentially influence (Rob-ertson and Garavan, in press). How-ever, it is important to complementthe spatial aspects of attention with itstemporal limitations (see Box 1).
Given a large visual array of individ-ual features, one can choose either toexamine it globally or to investigate itsspecific features. Using compoundstimuli for studying global and localprocessing, it is possible to study howone can shift back and forth betweenthem by changing the attentional fo-cus (Navon, 2003). For example, cer-tain patients have difficulty examiningthe local features. These patients usu-ally have damage to the left tem-poroparietal lobe. Other patients maydo well with the local features, but failto appreciate the overall contour; theyusually have damage to the right tem-poroparietal lobe. Indeed, the parietallobe tends to emphasize the shiftingbetween local and global stimuli,while the temporal lobe seems to de-termine whether one can actually ex-amine a local or global feature of thestimulus.
We usually foveate, or look at, ex-actly the thing in which we are inter-ested, and that process generally re-lates our attention to where we fixate.However, it is easy to dissociate atten-tion from the fovea. We can cue peo-ple to attend to some location in spaceother than the center of gaze and thenshow that they are sensitive to infor-mation that occurs at the cued loca-tion and relatively slow or insensitiveto information at the fovea, as mea-sured by a low threshold or fast re-sponse time (Posner, 1980). It is be-lieved that these covert shifts inattention are used to select the part ofthe visual field to which one usuallywants to move the eyes because ineveryday life, one usually follows co-vert shifts of attention with an eyemovement. Attention to visual ele-ments can also apply to other modal-ities, such as the auditory system.
When multiple people talk simulta-neously, it is sometimes necessary to
select one out of these streams of con-versation to follow in detail. We usu-ally do that based on the location ofthe person: we may visually orient to-ward the person and/or hone in ontheir frequency of voice. For example,we can typically separate a male voicefrom a female voice more easily thanwe can separate two male voices. Al-ternatively, we may pay attention tothe content of the information by fol-lowing a story line. When we attend toone stream, the other informationcoming from conversations around usgoes into the background: it ispresent, but does not reach focal anal-ysis. Data suggest that much of thisunattended information is actuallyprocessed in subtle and complicatedways (Kihlstrom, 1996). Moreover,unattended information can suddenlybecome interesting (e.g., when our
name is mentioned), whereby we ori-ent to the new information. These at-tentional phenomena have been stud-ied experimentally in some detail andwill be described next.
ATTENTION AND PERCEPTION
Attention is not a panacea to percep-tion because there is a great deal at-tention cannot do (Raz et al., 2004b;Raz, 2004a,b). For example, while ori-enting to a location, attention can givepriority to that location (i.e., targetsthat appear there will be perceivedmore rapidly and with lower thresh-olds). However, attention cannot sub-stitute for the acuity provided by thefovea. While the fovea is critical foracuity, the costs in reaction time foran unexpected foveal stimulus are justas great as for an unexpected periph-eral event. Thus, visual attention in-
fluences priority or processing prefer-ence, but also modulates sensoryprocessing and top-down control(Pessoa et al., 2003; Raz, 2004) andmay even affect accommodation (Razet al., 2004a).
Compared to unattended stimuli,behavioral findings show that reac-tion time to attended stimuli is usuallyfaster. Neurophysiological data revealenhancement of electrical activityover extrastriate visual areas by about90 msec following visual presentation.Although performance may improveon increased attentional investment,great controversy has existed overwhat orienting attention to a sensorystimulus actually does. The psycho-physics literature provides reliable ac-counts of how visual thresholds corre-late with attentional investment.Studies show that improvement in vi-sual acuity is not synonymous withaltered thresholds for detection, bet-ter performance, or faster reactiontimes. Whereas acuity requires theresolution of detail, detection thresh-olds and reaction time can involve thesummation of luminance, whichmight obscure detail (Raz et al.,2004b).
Although investing attention is fre-quently associated with looking di-rectly at the scene of interest, covertattention is the ability to grant suchinformation priority in processingwithout eye movement. Researchershave shown that the performance im-provement at attended locations re-sults somewhat from an enhancedspatial resolution at the cued location(Yeshurun and Carrasco, 1998, 1999,2000; Carrasco et al., 2002). Studiesexploring the relationship between vi-sual attention and contrast sensitivityshow that covert attention not onlyimproves discrimination in a wide va-riety of visual tasks, but could alsoincrease the rate at which informationis processed. Some findings do indi-cate that contrast sensitivity is greaterin the lower versus higher visual me-ridian. However, most evidence setslimits to the effects of attention onspatial resolution and specifies thatcertain visual, not attentional, con-straints determine aspects of spatialresolution (Talgar and Carrasco,2002).
Researchers in the fieldagree that attention is
not a unitary term.Rather, we can
fractionate attention intosubsystems of more
circumscribed functionand anatomy.
24 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
ATYPICAL ATTENTION
Biological rhythms affect attentionalperformance. Diurnal reductions inattention normally occur during thehours of maximum sleepiness, 2:00a.m. to 7:00 a.m., which coincideswith the period of lowest body tem-perature. Attentional performance isusually enhanced in the evening,when body temperature peaks (Raz,1999; Manly et al., 2002). During thestates of sleep deprivation and sleep,voluntary attention is often markedlyattenuated or even absent. However,evidence suggests that certain atten-tional as well as preattentional mech-anisms remain intact albeit unstable(Raz, 1999; Raz and Posner, 2000; Do-ran et al., 2001; Raz et al., 2001).Dreaming is usually divorced from asense of controlled awareness. How-ever, purported accounts of luciddreaming, in which persons dreamwhile knowing that they are dream-ing, suggest that some control mech-anisms may be available during sleep.Other common anecdotes in supportof this concept include the incorpora-tion of ambient sound into the dreamcontent as well as the idea of sensitiv-ity to one’s own name.
To investigate information process-ing in sleep, researchers measure elec-trical recordings from the scalp usingelectroencephalography (EEG). Byaveraging the brain’s electrical re-sponse potentials to stimuli using atechnique called event-related poten-tials (ERPs), it is possible to examinethe processing capability of the sleep-ing brain. One such component to ex-amine is the mismatch negativity(MMN). The MMN is an electrophys-iological manifestation of involuntarypreattentive processing in response tooddball stimuli. In a typical MMNparadigm, a deviant auditory stimulusis infrequently interspersed within asequence of standard auditory stim-uli. The MMN is evident in the dif-ference waveform resulting from thesubtraction of the ERP elicited bythe standard stimulus from that elic-ited by the novel auditory stimuli(the deviants). The difference wave-form, occurring even without atten-tion, normally peaks between 100and 250 msec from the onset of thedeviant event. This value depends onthe dimension of deviance and its
magnitude. The MMN presumablyindicates a mechanism that com-pares the current auditory input tothe memory traces formed by previ-ous auditory inputs and signals theoccurrence of a mismatch (Naa-tanen, 2003).
In adults, MMN tends to declineduring drowsiness and its persis-tence into adult human sleep is stilldebated (Atienza et al., 2002). OtherEEG components do reflect thebrain’s reaction to novelty. Althoughactive midbrain inhibition blockscortical activity in the developedbrain, the sleeping infant brain isprobably not as capable of blockingand inhibiting information effi-ciently. Indeed, MMN is obtainablefrom newborns and young infantsand experimental data show thatduring those early developmentalstages, the brain can learn, even dur-ing sleep (Cheour et al., 2002).
Attention is sometimes associatedwith hypnosis, a special cognitivestate sometimes confused with sleep.Hypnosis has been used clinically forhundreds of years and is primarily aphenomenon involving attentive re-ceptive concentration. Following par-ticular suggestions, highly hypnotiz-able individuals may experienceattentional and perceptual changes,which may not typically occur duringcommon awareness. In a responsivesubject, suggestions for perceptual al-teration are accompanied by repro-ducible changes in brain action. Forexample, the activity of the anteriorcingulate cortex (ACC) to painfulstimuli or conflict resolution can bemodulated by hypnotic suggestion(Rainville et al., 1997; Raz and Sha-piro, 2002; Raz, 2004). Interestingly,most children are highly hypnotizableand more easily inducted into hypno-sis than are adults.
ATTENTION ANDNEUROIMAGING
Two recently developed tools havebeen particularly valuable in studyingattention as an organ system: brainimaging and genomics. Neuroimaginghas illuminated the anatomical areasinvolved in attention, while the Hu-man Genome Project is identifyinggenes involved in these networks. Thisarticle focuses on data concerning theneuroimaging of attention. The genet-ics of attention and the interaction be-tween the two methodologies are ad-dressed elsewhere (Fossella et al.,2002b; Fan et al., 2003c).
The past 2 decades have ushered ina new era of methodological advancesin tools for noninvasive imaging of theliving brain. The information gleanedfrom such advances has identifiedboth anatomical and functional as-pects of the neural processes underly-ing attention (Kastner and Ungerlei-der, 2000, 2001; Driver andFrackowiak, 2001; Pessoa et al., 2003).Functional neuroimaging methods al-low researchers to measure changesin brain activity associated with si-multaneous changes in behavior, or inresponse to a wide variety of stimuli.ERP, functional magnetic resonanceimaging (fMRI), magnetoencephalog-raphy (MEG), near-infrared spectros-copy (NIRS), positron emission to-mography (PET), and single photonemission computed tomography(SPECT) all measure aspects of phys-iological change in brain activity.
Early neuroimaging techniques ofthe 1970s allowed researchers torecord from cells in alert monkeysperforming an experimental cognitivetask. Data from these assays showedincreased firing rates for cells in anumber of brain regions when themonkeys paid attention compared towhen they did not (Wurtz and Gold-berg, 1972). These efforts directly re-lated to studies of other aspects of at-tention, such as research carried outin the 1960s to decipher the mecha-nism of brain information processing(e.g., Broadbent). Since the 1980s, thestudy of patients with focal brain le-sions and ERPs has established linksbetween the brain areas showing se-lective enhancement and specific as-pects of information processing. Re-cently, neuroimaging methods were
Two recently developedtools have been
particularly valuable instudying attention as an
organ system: brainimaging and genomics.
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 25
developed that expanded the types ofattention that could be studied physi-ologically. Some of these findings be-gan to influence computational mod-els of attention (LaBerge, 1994).
Psychological studies have eluci-dated the pertinent questions aboutattention. These studies created a sub-stantial number of experimental par-adigms, including dichotic listening,visual search, dual-task performance,flanker tasks, rapid serial visual pre-sentation, and negative priming.Through these models, researchershoped to capture the range of phe-nomena encompassed by attentionunder the controlled conditions re-quired by scientific scrutiny. Converg-ing evidence is vital to such analysis asscientists test hypotheses in a varietyof different ways so that artifacts ofone method are circumvented by an-other. Confidence in the conclusionsincreases with replications and thesystematic dismissal of potential andlingering caveats. Since inferences arenecessarily indirect (e.g., observationof behavior or measurement of brainactivations), by putting them to-gether, one further constrains the pro-posed theories.
While the psychology of attentionhas furnished a number of interestingresults regarding the limits of perfor-mance and of unconscious process-ing, there was no agreement onwhether attention involved separatemechanisms from those used to pro-cess data, nor did psychological stud-ies analyze the neural mechanisms ofattention. Furthermore, because ob-servable responses are needed as evi-dence, purely psychological studiesare limited in their ability to deter-mine how much information is pro-cessed concerning unattended mes-sages. Fortunately, brain imagingallows monitoring of incidental andinvoluntary processing of unattendedstimuli. Insights into the neural sys-tems underlying attention, therefore,markedly benefited from experimen-tal paradigms involving selection ofsensory information coupled with thetechnological innovation of imagingthe living brain.
Brain imaging, including data fromimplanted electrodes in both humanand nonhuman primates, has forgedan impressive link between psychol-ogy and neuroscience (Posner,
2004b). Neuroimaging data have con-firmed earlier notions that attention isnot a unitary concept and that the hu-man brain likely entertains several at-tentional systems of different thoughinterrelated functions (Posner and Pe-tersen, 1990). Indeed, for the last15–20 years, brain imaging has beenexamining questions of attention andmany other cognitive tasks. Major re-sults of this research can be summa-rized in three statements (Posner,2004c). One, networks of neural areas,often widely scattered over the brain,but not involving the whole brain,compute cognitive and emotionaltasks. Each node of the network com-putes a different aspect and togetherthe networks orchestrate the wholetask. Two, some networks are in-volved in the control of other net-works. These attentional networks areinvolved in the selection and controlof networks that process sensory in-
formation and information frommemory. Three, these networkschange with development, learning,brain injury, and pathology.
Given the increased description ofattentional networks in recent years,we can now use these networks asmodel systems for the exploration ofsymptoms arising from various formsof pathology (Berger and Posner,2000). Current research has demon-strated that distinct brain areas in-deed mediate different attentionalprocesses and thus it is now possibleto examine selective attention as anorgan system with its own functionalanatomy, circuitry, and cellular struc-ture (Posner and Fan, 2004). Althoughstill incomplete, this information hasilluminated important questions incognitive science and has provided in-sights into neurological and psychiat-ric disorders of both children andadults. Converging data now point out
that attention enables the exercise ofvoluntary control over thoughts, feel-ings, and actions as a means of self-regulation (Raz, 2004) in adulthoodand throughout development (Posnerand Rothbart, 1998, 2000; Bronson,2000; Rothbart et al., 2004). Varia-tions in the operational efficiency ofthese attentional systems explain dif-ferences in self-regulation and emo-tional control and promise to help de-scribe mechanisms of volition andsustained effort (Rueda et al., 2004b,in press). In a way, this network ap-proach owes a lot to the work carriedout about 50 years ago by Donald O.Hebb, who outlined in his cell-assem-bly theory the ensemble of areas in thebrain that might be involved in pro-cessing cognitive and emotional tasks(Posner and Rothbart, in press).
DIFFERENT TYPES OF ATTENTION
Attention does not imply a singularmechanism; rather, it is a complexsystem presiding over a number ofdistinct neuronal circuits. This under-standing comes from combining datagathered using various neuroimagingtechniques. Early PET data show thatdifferent attributes such as color, mo-tion, and shape are processed by atleast partially separate systems (Cor-betta et al., 1991a, 1991b). Addition-ally, a recent review of fMRI resultsreveals a distributed system of brainregions that control attention (Fan etal., 2003a; Pessoa et al., 2003). Ac-cordingly, researchers have formu-lated experimental paradigms thatconsiderably shape our understand-ing of attention. More recently, thenotion of attention has been extendedto the control of action, thought, andaffect both in adults and across devel-opment (Posner and Rothbart, 1998,2000; Rueda et al., 2002; Rothbart etal., 2003, 2004; Rueda et al., 2004a,2004b, in press).
Three largely disparate attentionalcontrol systems—select, orient, andalert—have been identified. Althoughthe three systems interact in manypractical contexts, the operations ofthese attentional networks have a cer-tain degree of functional and anatom-ical independence. Whereas the selectsystem involves choosing among con-flicting actions, orient refers to a pointof reference to sensory objects, and
Attention does not implya singular mechanism;rather, it is a complex
system presiding over anumber of distinctneuronal circuits.
26 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
alert involves changes in the internalstate in preparation for perceiving astimulus. Insult to each of these dis-tinct attentional circuits, irrespectiveof the source, produces dissimilarneuropsychological deficits. Correlat-ing the etiologies of such injuries withsubsequent changes in behavior andcognition allows us to extend the roleof attentional neuroimaging possiblyinto rehabilitation (Robertson, 1999;Fan et al., 2003b; Posner and Fan,2004; Rueda et al., in press). Re-searchers have illuminated the neuro-chemical substrates of these atten-tional networks and detailed analysesare now emerging for these orthogo-nal sensory systems (Raz and Shapiro,2002; Fan et al., 2003b; Posner,2004a).
Neuroimaging findings have shownactivation of frontal areas associatedwith paying attention in the context ofstrong mental effort. For example,when experimental tasks appear to in-volve significant exertion during tar-get detection or conflict resolution,the area of the ACC is consistently ac-tivated. Thus, the select network is ahigher-level metacognitive attentionalsystem related to the subjective im-pression of mental effort (Fernandez-Duque et al., 2000a, 2000b). This formof supervisory or executive attentioncomprises the mechanisms for moni-toring and resolving conflict amongthoughts, feelings, and responses.Therefore, this attentional system isconcerned with such tasks as workingmemory, planning, switching, and in-hibitory control. Since frontal pa-tients are less able to implement a setof instructed goals, the orbital frontalarea is probably important for this ex-ecutive faculty (Duncan et al., 1996).
Visual orienting is the process ofselecting information from sensoryinput. Orienting of attention towardvisual objects can be either triggeredby the stimuli (e.g., by brief presenta-tions) or shifted as a result of volun-tary control. Orienting has thus be-come a vehicle for separatingautomatic from voluntary mecha-nisms. Early PET results have under-lined the importance of the parietalarea together with thalamic and mid-brain areas for visual orienting. Theorienting network has been used tounderstand the effects of lesions thatproduce neglect of sensory informa-
tion either by brain damage or by re-stricting transmitter input. Likewise,frontal attentional networks have pro-vided similar understanding of pa-thologies at higher levels of cognition.For example, evidence exists relat-ing frontal networks to attention def-icit hyperactivity disorder (ADHD)(Berger and Posner, 2000). Recent re-search has linked the cholinergic sys-tem to orienting (Beane and Mar-rocco, 2004).
The alert, or vigilant, system func-tions to achieve and maintain a stateof high sensitivity to incoming stim-uli. The process of acquiring andmaintaining alertness in the face ofprolonged dull tasks has been an areaof vigorous research. Recent reportshave suggested some of the anatomyinvolved in how arousal, alertness,and vigilance affect performance
(Robertson and Garavan, in press).They also highlight the importance ofnorepinephrine modulation in thealerting that follows warning signals(Witte and Marrocco, 1997; Witte etal., 1997).
SUBSTRATES OF ATTENTIONALNETWORKS
Attention can enhance neural process-ing at multiple levels. As a selectiveaspect of information processing, at-tention allows some things to be priv-ileged while others are ignored. Vigor-ous studies into the modulation ofneural responses by attention have re-vealed that attention can be viewed asa system of anatomical areas carryingout the functions of alerting, orient-ing, and executive control (Posner andRaichle, 1996; Posner, 2004a). In linewith an attentional research agenda
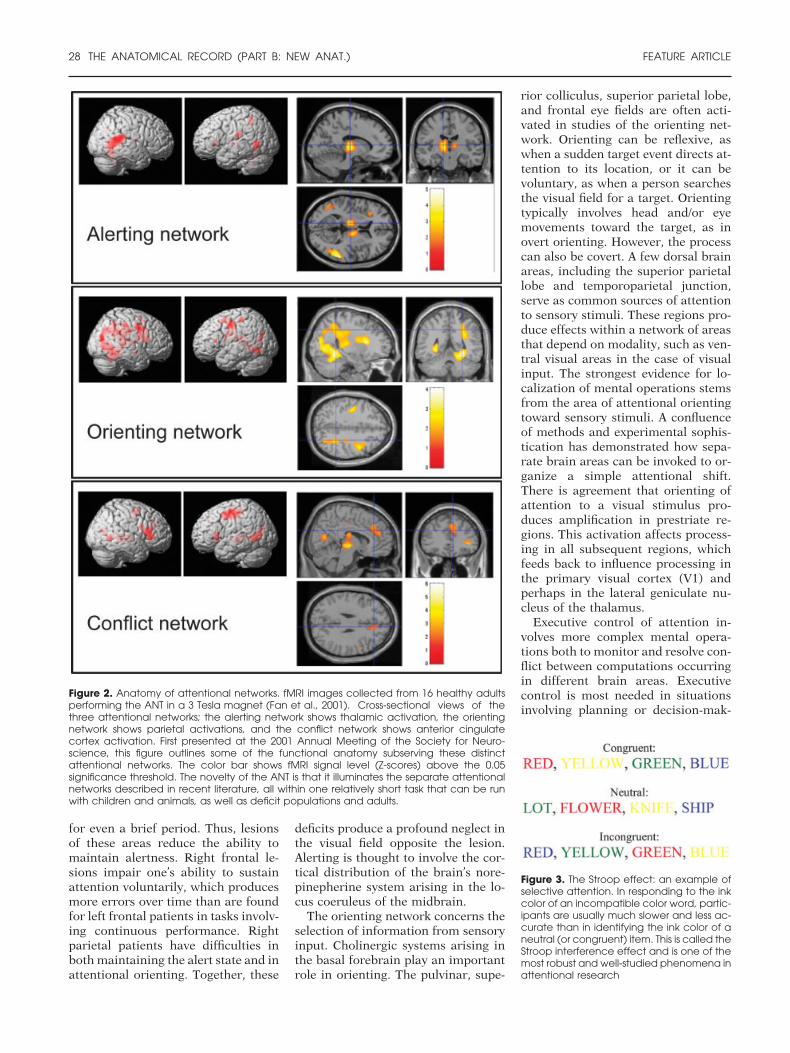
(Posner and Fan, 2004), we have re-cently devised a simple Attention Net-work Test (ANT) that can be per-formed by adults, children, patients,and even nonhuman animals (Fan etal., 2002). The ANT takes about 30min to administer and provides threenumbers that indicate the efficiencyof the networks that perform the alert,orient, and conflict-resolution func-tions. Previous work with this testprovided evidence on its reliability,heritability, and the independence ofthe results (Fan et al., 2001a, 2001b;Fossella et al., 2002a). Recent neuro-imaging findings have also used theANT to probe and assess these distinctattentional networks (Fan et al.,2001a). These behavioral and imagingdata suggest that the networks arelargely independent and further per-mit identification of the neuroanat-omy subserving these attentional net-works (Fig. 2).
Although previous studies have ex-amined areas of the brain involved invarious ANT components (Corbetta etal., 2000; Hopfinger et al., 2000), neu-roimaging the ANT has illuminatedareas involved in carrying out theseattentional networks as a whole. fMRIdata from the ANT indicate that threelargely orthogonal networks relate tocomponents of attention (Fan et al.,2001a). Pharmacological studies haverelated each of these networks withspecific chemical neuromodulators.First, cholinergic systems arising inthe basal forebrain play an importantrole in orienting (Beane and Mar-rocco, 2004). Second, the norepineph-erine system arising in the locus coer-uleus of the midbrain functions inalerting (Witte and Marrocco, 1997;Witte et al., 1997; Coull et al., 2001).Third, the ACC and lateral prefrontalcortex are target areas of the mesocor-tical dopamine system involved in ex-ecutive attention (Marrocco and Da-vidson, 1998; Deth et al., 2004;Robbins et al., 2004).
Alerting involves a change in the in-ternal state in preparation for perceiv-ing a stimulus. The alert state is criti-cal for optimal performance in tasksinvolving higher cognitive functions.Neuroimaging studies have shown ac-tivity in the frontal and parietal re-gions particularly of the right hemi-sphere when people are required toachieve and maintain the alert state
Alerting involves achange in the internalstate in preparation forperceiving a stimulus.
The alert state is criticalfor optimal performancein tasks involving higher
cognitive functions.
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 27
for even a brief period. Thus, lesionsof these areas reduce the ability tomaintain alertness. Right frontal le-sions impair one’s ability to sustainattention voluntarily, which producesmore errors over time than are foundfor left frontal patients in tasks involv-ing continuous performance. Rightparietal patients have difficulties inboth maintaining the alert state and inattentional orienting. Together, these
deficits produce a profound neglect inthe visual field opposite the lesion.Alerting is thought to involve the cor-tical distribution of the brain’s nore-pinepherine system arising in the lo-cus coeruleus of the midbrain.
The orienting network concerns theselection of information from sensoryinput. Cholinergic systems arising inthe basal forebrain play an importantrole in orienting. The pulvinar, supe-
rior colliculus, superior parietal lobe,and frontal eye fields are often acti-vated in studies of the orienting net-work. Orienting can be reflexive, aswhen a sudden target event directs at-tention to its location, or it can bevoluntary, as when a person searchesthe visual field for a target. Orientingtypically involves head and/or eyemovements toward the target, as inovert orienting. However, the processcan also be covert. A few dorsal brainareas, including the superior parietallobe and temporoparietal junction,serve as common sources of attentionto sensory stimuli. These regions pro-duce effects within a network of areasthat depend on modality, such as ven-tral visual areas in the case of visualinput. The strongest evidence for lo-calization of mental operations stemsfrom the area of attentional orientingtoward sensory stimuli. A confluenceof methods and experimental sophis-tication has demonstrated how sepa-rate brain areas can be invoked to or-ganize a simple attentional shift.There is agreement that orienting ofattention to a visual stimulus pro-duces amplification in prestriate re-gions. This activation affects process-ing in all subsequent regions, whichfeeds back to influence processing inthe primary visual cortex (V1) andperhaps in the lateral geniculate nu-cleus of the thalamus.
Executive control of attention in-volves more complex mental opera-tions both to monitor and resolve con-flict between computations occurringin different brain areas. Executivecontrol is most needed in situationsinvolving planning or decision-mak-
Figure 3. The Stroop effect: an example ofselective attention. In responding to the inkcolor of an incompatible color word, partic-ipants are usually much slower and less ac-curate than in identifying the ink color of aneutral (or congruent) item. This is called theStroop interference effect and is one of themost robust and well-studied phenomena inattentional research
Figure 2. Anatomy of attentional networks. fMRI images collected from 16 healthy adultsperforming the ANT in a 3 Tesla magnet (Fan et al., 2001). Cross-sectional views of thethree attentional networks; the alerting network shows thalamic activation, the orientingnetwork shows parietal activations, and the conflict network shows anterior cingulatecortex activation. First presented at the 2001 Annual Meeting of the Society for Neuro-science, this figure outlines some of the functional anatomy subserving these distinctattentional networks. The color bar shows fMRI signal level (Z-scores) above the 0.05significance threshold. The novelty of the ANT is that it illuminates the separate attentionalnetworks described in recent literature, all within one relatively short task that can be runwith children and animals, as well as deficit populations and adults.
28 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
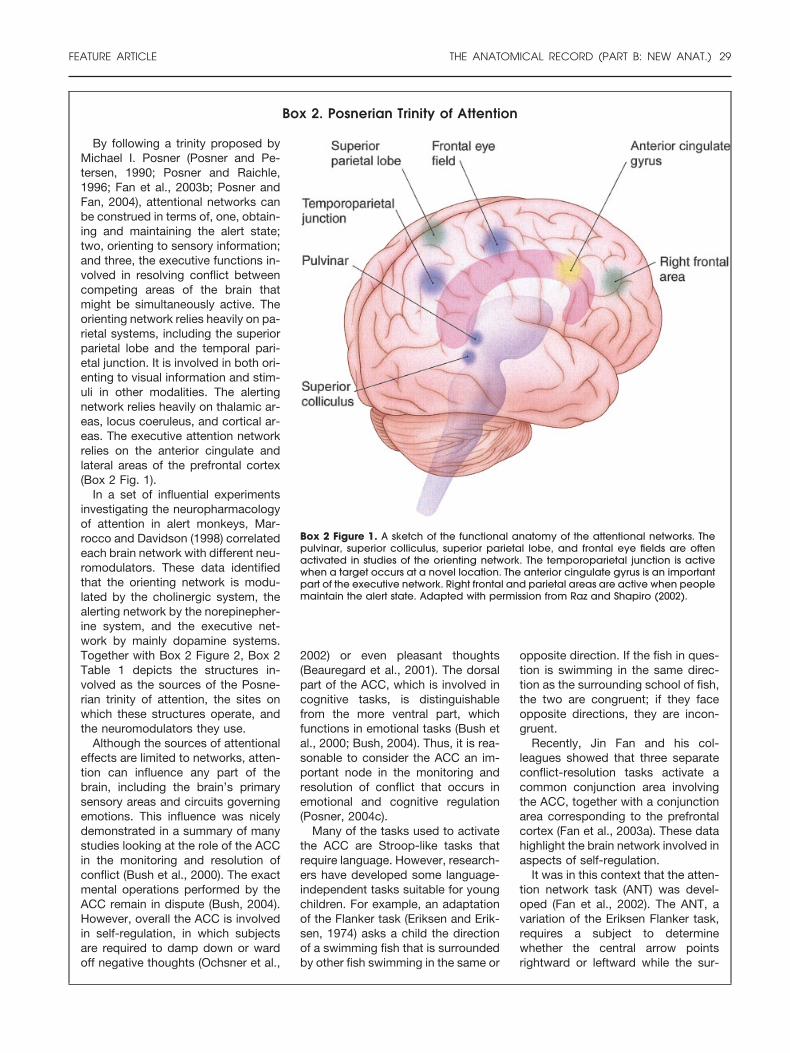
Box 2. Posnerian Trinity of Attention
Box 2 Figure 1. A sketch of the functional anatomy of the attentional networks. Thepulvinar, superior colliculus, superior parietal lobe, and frontal eye fields are oftenactivated in studies of the orienting network. The temporoparietal junction is activewhen a target occurs at a novel location. The anterior cingulate gyrus is an importantpart of the executive network. Right frontal and parietal areas are active when peoplemaintain the alert state. Adapted with permission from Raz and Shapiro (2002).
By following a trinity proposed byMichael I. Posner (Posner and Pe-tersen, 1990; Posner and Raichle,1996; Fan et al., 2003b; Posner andFan, 2004), attentional networks canbe construed in terms of, one, obtain-ing and maintaining the alert state;two, orienting to sensory information;and three, the executive functions in-volved in resolving conflict betweencompeting areas of the brain thatmight be simultaneously active. Theorienting network relies heavily on pa-rietal systems, including the superiorparietal lobe and the temporal pari-etal junction. It is involved in both ori-enting to visual information and stim-uli in other modalities. The alertingnetwork relies heavily on thalamic ar-eas, locus coeruleus, and cortical ar-eas. The executive attention networkrelies on the anterior cingulate andlateral areas of the prefrontal cortex(Box 2 Fig. 1).
In a set of influential experimentsinvestigating the neuropharmacologyof attention in alert monkeys, Mar-rocco and Davidson (1998) correlatedeach brain network with different neu-romodulators. These data identifiedthat the orienting network is modu-lated by the cholinergic system, thealerting network by the norepinepher-ine system, and the executive net-work by mainly dopamine systems.Together with Box 2 Figure 2, Box 2Table 1 depicts the structures in-volved as the sources of the Posne-rian trinity of attention, the sites onwhich these structures operate, andthe neuromodulators they use.
Although the sources of attentionaleffects are limited to networks, atten-tion can influence any part of thebrain, including the brain’s primarysensory areas and circuits governingemotions. This influence was nicelydemonstrated in a summary of manystudies looking at the role of the ACCin the monitoring and resolution ofconflict (Bush et al., 2000). The exactmental operations performed by theACC remain in dispute (Bush, 2004).However, overall the ACC is involvedin self-regulation, in which subjectsare required to damp down or wardoff negative thoughts (Ochsner et al.,
2002) or even pleasant thoughts(Beauregard et al., 2001). The dorsalpart of the ACC, which is involved incognitive tasks, is distinguishablefrom the more ventral part, whichfunctions in emotional tasks (Bush etal., 2000; Bush, 2004). Thus, it is rea-sonable to consider the ACC an im-portant node in the monitoring andresolution of conflict that occurs inemotional and cognitive regulation(Posner, 2004c).
Many of the tasks used to activatethe ACC are Stroop-like tasks thatrequire language. However, research-ers have developed some language-independent tasks suitable for youngchildren. For example, an adaptationof the Flanker task (Eriksen and Erik-sen, 1974) asks a child the directionof a swimming fish that is surroundedby other fish swimming in the same or
opposite direction. If the fish in ques-tion is swimming in the same direc-tion as the surrounding school of fish,the two are congruent; if they faceopposite directions, they are incon-gruent.
Recently, Jin Fan and his col-leagues showed that three separateconflict-resolution tasks activate acommon conjunction area involvingthe ACC, together with a conjunctionarea corresponding to the prefrontalcortex (Fan et al., 2003a). These datahighlight the brain network involved inaspects of self-regulation.
It was in this context that the atten-tion network task (ANT) was devel-oped (Fan et al., 2002). The ANT, avariation of the Eriksen Flanker task,requires a subject to determinewhether the central arrow pointsrightward or leftward while the sur-
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 29
Box 2. (Continued)
Box 2 Figure 2. Experimental procedure for the ANT. a: The four cue conditions. b: Thesix stimuli used in the experiment. c: The task as it unfolds in time. Adapted withpermission from Fan et al. (2002).
rounding arrows may be either con-gruent or incongruent (Box 2 Fig. 2).This design permits researchers tomeasure the subject’s ability to re-solve conflict, or cognitive regulation,by subtracting the congruent reactiontime from the incongruent reactiontime. However, before the task starts,the subject is given a cue, which maybe no cue at all, a double cue (alertingthe subject to get ready because thetarget is about to occur), or it may bea cue orienting the subject to the tar-get’s location. This design permitstwo additional subtractions in orderto extract both a measure of alertingand a measure of orienting. Analysisof reaction time data reveals little cor-relation between these three tasks.Although small interactions do occur,such as when the orienting cue oc-curs at the target location, there ap-pears to be relative independencebetween the congruency conditionand the alerting and orienting condi-tions. Using fMRI, Jin Fan has alsoshown activation of unique neuroana-tomical sites for each attentional net-work. The alerting network elicits astrong thalamic activation, while ori-enting activates superior and inferiorareas of the parietal lobe and the con-flict or executive network induces ac-tivation at the ACC and lateral pre-frontal cortex. Whereas many otherstudies have measured these atten-tional networks separately, the ANTallows measurement of all attentionalnetworks within one relatively shorttask that can be run with children andanimals, as well as adults and pa-tients.
Box 2. TABLE 1. Attentional networks: brain regions and neuromodulators
Alert (arousal, vigilant attention) Locus coeruleus: right frontal and parietal cortex Norepinepherine
30 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
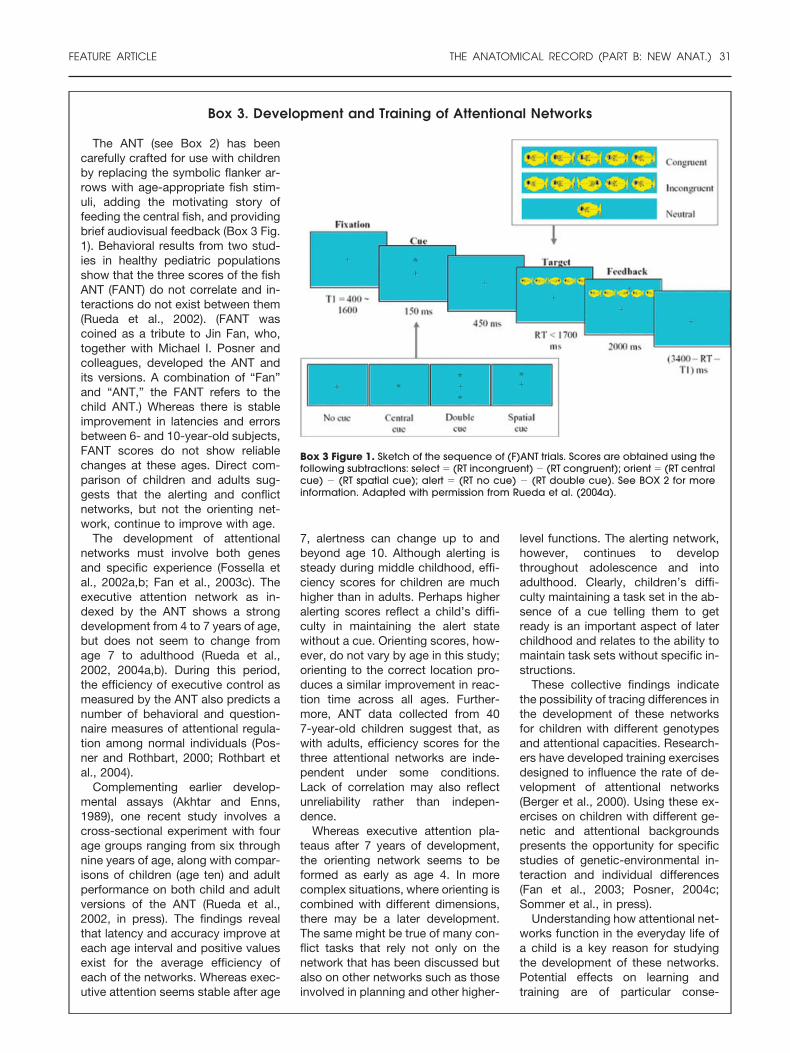
Box 3. Development and Training of Attentional Networks
Box 3 Figure 1. Sketch of the sequence of (F)ANT trials. Scores are obtained using thefollowing subtractions: select � (RT incongruent) � (RT congruent); orient � (RT centralcue) � (RT spatial cue); alert � (RT no cue) � (RT double cue). See BOX 2 for moreinformation. Adapted with permission from Rueda et al. (2004a).
The ANT (see Box 2) has beencarefully crafted for use with childrenby replacing the symbolic flanker ar-rows with age-appropriate fish stim-uli, adding the motivating story offeeding the central fish, and providingbrief audiovisual feedback (Box 3 Fig.1). Behavioral results from two stud-ies in healthy pediatric populationsshow that the three scores of the fishANT (FANT) do not correlate and in-teractions do not exist between them(Rueda et al., 2002). (FANT wascoined as a tribute to Jin Fan, who,together with Michael I. Posner andcolleagues, developed the ANT andits versions. A combination of “Fan”and “ANT,” the FANT refers to thechild ANT.) Whereas there is stableimprovement in latencies and errorsbetween 6- and 10-year-old subjects,FANT scores do not show reliablechanges at these ages. Direct com-parison of children and adults sug-gests that the alerting and conflictnetworks, but not the orienting net-work, continue to improve with age.
The development of attentionalnetworks must involve both genesand specific experience (Fossella etal., 2002a,b; Fan et al., 2003c). Theexecutive attention network as in-dexed by the ANT shows a strongdevelopment from 4 to 7 years of age,but does not seem to change fromage 7 to adulthood (Rueda et al.,2002, 2004a,b). During this period,the efficiency of executive control asmeasured by the ANT also predicts anumber of behavioral and question-naire measures of attentional regula-tion among normal individuals (Pos-ner and Rothbart, 2000; Rothbart etal., 2004).
Complementing earlier develop-mental assays (Akhtar and Enns,1989), one recent study involves across-sectional experiment with fourage groups ranging from six throughnine years of age, along with compar-isons of children (age ten) and adultperformance on both child and adultversions of the ANT (Rueda et al.,2002, in press). The findings revealthat latency and accuracy improve ateach age interval and positive valuesexist for the average efficiency ofeach of the networks. Whereas exec-utive attention seems stable after age
7, alertness can change up to andbeyond age 10. Although alerting issteady during middle childhood, effi-ciency scores for children are muchhigher than in adults. Perhaps higheralerting scores reflect a child’s diffi-culty in maintaining the alert statewithout a cue. Orienting scores, how-ever, do not vary by age in this study;orienting to the correct location pro-duces a similar improvement in reac-tion time across all ages. Further-more, ANT data collected from 407-year-old children suggest that, aswith adults, efficiency scores for thethree attentional networks are inde-pendent under some conditions.Lack of correlation may also reflectunreliability rather than indepen-dence.
Whereas executive attention pla-teaus after 7 years of development,the orienting network seems to beformed as early as age 4. In morecomplex situations, where orienting iscombined with different dimensions,there may be a later development.The same might be true of many con-flict tasks that rely not only on thenetwork that has been discussed butalso on other networks such as thoseinvolved in planning and other higher-
level functions. The alerting network,however, continues to developthroughout adolescence and intoadulthood. Clearly, children’s diffi-culty maintaining a task set in the ab-sence of a cue telling them to getready is an important aspect of laterchildhood and relates to the ability tomaintain task sets without specific in-structions.
These collective findings indicatethe possibility of tracing differences inthe development of these networksfor children with different genotypesand attentional capacities. Research-ers have developed training exercisesdesigned to influence the rate of de-velopment of attentional networks(Berger et al., 2000). Using these ex-ercises on children with different ge-netic and attentional backgroundspresents the opportunity for specificstudies of genetic-environmental in-teraction and individual differences(Fan et al., 2003; Posner, 2004c;Sommer et al., in press).
Understanding how attentional net-works function in the everyday life ofa child is a key reason for studyingthe development of these networks.Potential effects on learning andtraining are of particular conse-
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 31
ing, error detection, novel or not well-learned responses, difficult or danger-ous conditions, and in overcominghabitual actions. The ACC and lateralfrontal cortex are target areas of theventral tegmental dopamine system.Brain imaging data have repeatedlyshown that the ACC is an importantnode in this network. More specifi-cally, neuroimaging studies haveshown activation of the dorsal ACC intasks requiring people to respond ei-ther to a prepotent response or to arather strong conflicting dimension.For example, in the classic Strooptask, experienced readers name theink color of a displayed word (Stroop,1935). Subjects are usually slower andless accurate in responding to the inkcolor of an incompatible color word,such as the word “RED” displayed inblue ink, than in identifying the inkcolor of a control item (e.g., “LOT”inked in red; Fig. 3). Another suchtask involves subjects responding tothe direction of a central arrow whenflanking arrows could either point inthe same (congruent) or opposite (in-congruent) direction (see Box 2 Fig.2). The ANT uses this flanker task tomeasure conflict. Such conflict taskshave been shown by neuroimagingstudies to activate the lateral prefron-tal cortex, frontal areas such as the
dorsal ACC, and parts of the basalganglia. These experimental tasksprovide a means of fractionating thefunctional contributions of areaswithin the executive attention net-work. Chiefly, the ACC is more activeon incongruent than on congruent tri-als. This difference could reflect thegeneral finding that lateral areas areinvolved in representing specific in-formation over time, while medial ar-eas are more related to the detectionof conflict.
Patients with focal brain lesions ofthe ACC initially display deficits ofvoluntary behavior. The notion ofACC involvement in cognitive controland volition has been a topic of muchinterest lately. Based on behavioral,optical, and neuroimaging data, werecently reported that effective post-hypnotic suggestion, or verbal exhor-tation, to construe words as meaning-less symbols can modulate focal brainactivity in highly suggestible individ-uals (Raz et al., 2002, 2003; Raz,2004). This top-down influence is bothpotent and selective: it removesStroop conflict and reduces neuroim-aging signals in both the ACC and ex-trastriate cortex (Raz, 2004). Whereasa drop in ACC activity is likely relatedto the abrogation of conflict, the oc-cipital-parietal area may be related to
the visual word-form area, which ar-ranges visual letters into words. Oth-ers similarly report that in highlysuggestible individuals, hypnotic in-structions to view a color image ingray scale prevents normal activationof the color areas of the prestriate cor-tex (Kosslyn et al., 2000). These dataimply that, at least in highly suggest-ible individuals, attentional manipu-lations can influence aspects of self-regulation by affecting neural activityin specific brain areas. These findingssupport the use of special attentionalinterventions such as hypnosis tostudy related brain processes (Razand Shapiro, 2002).
ANT AND LANT
Aspects of laterality and developmentmay operate within attentional net-works. For example, human vigilancenetworks tend to be right-lateralized,while orienting appears to be specificto both modality and hemisphere.PET data collected from healthy indi-viduals show that the right parietallobe is involved in attentional shiftson either side, whereas the left pari-etal lobe was involved in attentionalshifts on the right side only. Patientswith right parietal damage exhibit im-paired leftward orienting for both vi-
Box 3. (Continued)
quence. Whereas training in non-human animals can persist for hun-dreds of thousands of trials, as-sessing the effects of training inhuman and especially in childrencan be difficult. Training for only afew days would be unlikely to havea large effect on behavior, but maycause minimal changes that wouldindicate greater potential benefitsof more extensive attentional train-ing. Given this premise, Posner andhis colleagues conducted a 5-dayattentional training pilot. On thefirst and last day, they examinedthe children’s EEG during the ANTtask, as well as their behavioraldata, IQ, and temperament mea-sures. Preliminary analyses indi-cate that even a brief 5-day atten-tional training shifts the attentionalnetworks of children toward a more
adult-likepattern. Additionally, thedata also show a striking increase inIQ, suggesting the generalization ofthese effects. Specifically, the exper-imental group improves significantlyin the matrix portion of the child ver-sion of the IQ test. Thus, attentionaltraining appears to improves the un-derlying neural networks involved inconflict resolution and may even gen-eralize to different tasks (Posner,2004c).
Similar findings have been reportedin 8-year-old children diagnosed withADHD. After completing a trainingprogram to strengthen working mem-ory, participants have significantlyimproved motor performances and IQresults on the nonverbal complex rea-soning task of the Raven’s progres-sive matrices (Klingberg et al., 2002,in press; Olesen et al., 2004). These
data suggest that working memorytraining potentially could be of clin-ical use for ameliorating the symp-toms in ADHD. Training effortssimilar to the above-mentioned at-tentional assays have been re-ported in adults and children withADHD (Posner, 2004c). As neuro-imaging begins to unravel the ef-fects of practice on brain sub-strates (Garavan et al., 2004;Landau et al., 2004), cumulativedata suggest that these attentionalnetworks can be modified. Intro-ducing attentional training in pre-schools or childcare centers maybe a valuable innovation. Atten-tional training might be of particularimportance because it forms spe-cific neural networks involved inmultiple domains of behavior (Pos-ner, 2004c).
32 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
sual fields. When the corpus callosumis intact and the two cerebral hemi-spheres are in normal communica-tion, a single focus of orienting ismaintained. However, when the cal-lossal commissures are disconnected,as in split-brain patients, each hemi-sphere has a partly separate focus fororienting (Zaidel, 1995). From a mo-dality perspective, visual, tactile, andauditory neglect seem to involve dif-ferent areas of the parietal lobe. None-theless, the phenomenon of cross-modal integration, in which visualinformation influences the locationfrom which we hear sounds, suggeststhat these areas do communicate.
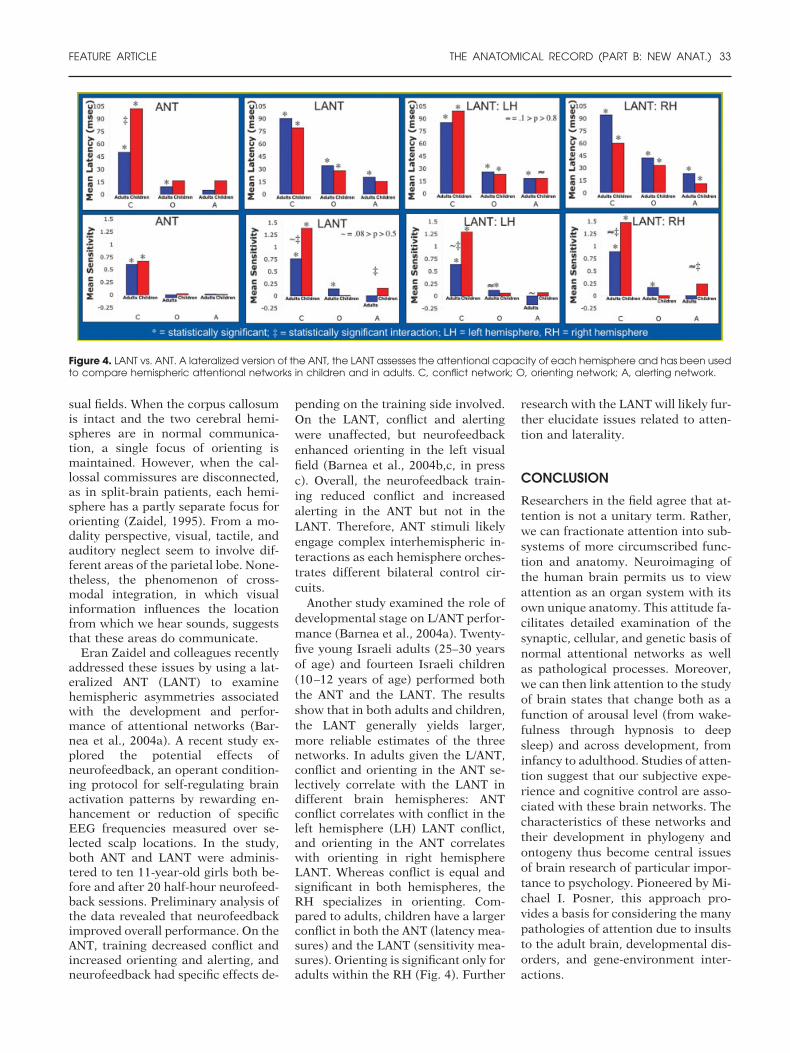
Eran Zaidel and colleagues recentlyaddressed these issues by using a lat-eralized ANT (LANT) to examinehemispheric asymmetries associatedwith the development and perfor-mance of attentional networks (Bar-nea et al., 2004a). A recent study ex-plored the potential effects ofneurofeedback, an operant condition-ing protocol for self-regulating brainactivation patterns by rewarding en-hancement or reduction of specificEEG frequencies measured over se-lected scalp locations. In the study,both ANT and LANT were adminis-tered to ten 11-year-old girls both be-fore and after 20 half-hour neurofeed-back sessions. Preliminary analysis ofthe data revealed that neurofeedbackimproved overall performance. On theANT, training decreased conflict andincreased orienting and alerting, andneurofeedback had specific effects de-
pending on the training side involved.On the LANT, conflict and alertingwere unaffected, but neurofeedbackenhanced orienting in the left visualfield (Barnea et al., 2004b,c, in pressc). Overall, the neurofeedback train-ing reduced conflict and increasedalerting in the ANT but not in theLANT. Therefore, ANT stimuli likelyengage complex interhemispheric in-teractions as each hemisphere orches-trates different bilateral control cir-cuits.
Another study examined the role ofdevelopmental stage on L/ANT perfor-mance (Barnea et al., 2004a). Twenty-five young Israeli adults (25–30 yearsof age) and fourteen Israeli children(10–12 years of age) performed boththe ANT and the LANT. The resultsshow that in both adults and children,the LANT generally yields larger,more reliable estimates of the threenetworks. In adults given the L/ANT,conflict and orienting in the ANT se-lectively correlate with the LANT indifferent brain hemispheres: ANTconflict correlates with conflict in theleft hemisphere (LH) LANT conflict,and orienting in the ANT correlateswith orienting in right hemisphereLANT. Whereas conflict is equal andsignificant in both hemispheres, theRH specializes in orienting. Com-pared to adults, children have a largerconflict in both the ANT (latency mea-sures) and the LANT (sensitivity mea-sures). Orienting is significant only foradults within the RH (Fig. 4). Further
research with the LANT will likely fur-ther elucidate issues related to atten-tion and laterality.
CONCLUSION
Researchers in the field agree that at-tention is not a unitary term. Rather,we can fractionate attention into sub-systems of more circumscribed func-tion and anatomy. Neuroimaging ofthe human brain permits us to viewattention as an organ system with itsown unique anatomy. This attitude fa-cilitates detailed examination of thesynaptic, cellular, and genetic basis ofnormal attentional networks as wellas pathological processes. Moreover,we can then link attention to the studyof brain states that change both as afunction of arousal level (from wake-fulness through hypnosis to deepsleep) and across development, frominfancy to adulthood. Studies of atten-tion suggest that our subjective expe-rience and cognitive control are asso-ciated with these brain networks. Thecharacteristics of these networks andtheir development in phylogeny andontogeny thus become central issuesof brain research of particular impor-tance to psychology. Pioneered by Mi-chael I. Posner, this approach pro-vides a basis for considering the manypathologies of attention due to insultsto the adult brain, developmental dis-orders, and gene-environment inter-actions.
Figure 4. LANT vs. ANT. A lateralized version of the ANT, the LANT assesses the attentional capacity of each hemisphere and has been usedto compare hemispheric attentional networks in children and in adults. C, conflict network; O, orienting network; A, alerting network.
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 33
ACKNOWLEDGMENTSI thank Michael I. Posner for hisgraceful mentorship, professional andpersonal inspiration, and continuedfriendship and support. I am also in-debted to Jin Fan and Mindy Tanzolafor their consistent collaboration con-cerning aspects of the content andpresentation, respectively.
LITERATURE CITED
Akhtar N, Enns JT. 1989. Relations be-tween covert orienting and filtering inthe development of visual attention. JExp Child Psychol 48:315–334.
Atienza M, Cantero JL, Dominguez-MarinE. 2002. Mismatch negativity (MMN): anobjective measure of sensory memoryand long-lasting memories during sleep.Int J Psychophysiol 46:215–225.
Barnea A, Rassis A, Neta M, Raz A, ZaidelE. 2004a. The lateralized attention net-work test (LANT) in children and adults.San Francisco: Cognitive NeuroscienceSociety.
Barnea A, Rassis A, Raz A, Othmer S,Zaidel E. 2004b. The effect of neurofeed-back on attention in children. San Fran-cisco: Cognitive Neuroscience Society.
Barnea A, Rassis A, Raz A, Othmer S,Zaidel E. 2004c. Effects of neurofeed-back on hemispheric attention networks.Tennet: an annual meeting of theoreticaland clinical neuropsychology. Montreal:Canada.
Barnea A, Rassis A, Raz A, Othmer S,Zaidel E. Effects of neurofeedback onhemispheric attentional networks. BrainCogn (in press c).
Beane M, Marrocco RT. 2004. Cholinergicand noradrenergic inputs to the poste-rior parietal cortex modulate the compo-nents of exogenous attention. In: PosnerMI, editor. Cognitive neuroscience of at-tention. New York: Guilford Press. p313–325.
Beauregard M, Levesque J, Bourgouin P.2001. Neural correlates of conscious self-regulation of emotion. J Neurosci 21:RC165.
Berger A, Jones L, Rothbart MK, PosnerMI. 2000. Computerized games to studythe development of attention in child-hood. Behav Res Methods Instrum Com-put 32:297–303.
Broadbent DE. 1958. Perception and com-munication. New York: Pergamon Press.
Bronson MB. 2000. Self-regulation in earlychildhood: nature and nurture. NewYork: Guilford Press.
Bush G, Luu P, Posner MI. 2000. Cognitiveand emotional influences in anterior cin-gulate cortex. Trends Cogn Sci 4:215–222.
Bush G. 2004. Multimodal studies of cin-gulate cortex. In: Posner MI, editor. Cog-
nitive neuroscience of attention. NewYork: Guilford Press. p 207–218.
Carrasco M, Williams PE, Yeshurun Y.2002. Covert attention increases spatialresolution with or without masks: sup-port for signal enhancement. J Vis 2:467–479.
Cavanagh P. 2003. Attention routines. In:International workshop on visual atten-tion. San Miniato, Italy.
Cavanagh P. 2004. Attention Routines andthe Architecture of Selection. In: PosnerMI, editor. Cognitive Neuroscience of At-tention. New York: Guilford Press. p 13–28.
Cheour M, Martynova O, Naatanen R,Erkkola R, Sillanpaa M, Kero P, Raz A,Kaipio ML, Hiltunen J, Aaltonen O,Savela J, Hamalainen H. 2002. Speechsounds learned by sleeping newborns.Nature 415:599–600.
Corbetta M, Miezin FM, Dobmeyer S,Shulman GL, Petersen SE. 1991a. Selec-tive and divided attention during visualdiscriminations of shape, color, andspeed: functional anatomy by positronemission tomography. J Neurosci 11:2383–2402.
Corbetta M, Miezin FM, Shulman GL, Pe-tersen SE. 1991b. Selective attentionmodulates extrastriate visual regions inhumans during visual feature discrimi-nation and recognition. Ciba FoundSymp 163:165–175.
Corbetta M, Kincade JM, Ollinger JM,McAvoy MP, Shulman GL. 2000. Volun-tary orienting is dissociated from targetdetection in human posterior parietalcortex. Nat Neurosci 3:292–297.
Coull JT, Nobre AC, Frith CD. 2001. Thenoradrenergic alpha2 agonist clonidinemodulates behavioural and neuroana-tomical correlates of human attentionalorienting and alerting. Cereb Cortex 11:73–84.
Deth RC, Kuznetsova A, Waly M. 2004.Attention-related signaling activities ofthe d4 dopamine receptor. In: PosnerMI, editor. Cognitive neuroscience of at-tention. New York: Guilford Press. p269–282.
Driver J, Frackowiak RSJ, editors. 2001.Imaging selective attention in the humanbrain. New York: Elsevier.
Duncan J, Emslie H, Williams P, JohnsonR, Freer C. 1996. Intelligence and thefrontal lobe: the organization of goal-directed behavior. Cogn Psychol 30:257–303.
Eriksen BA, Eriksen CW. 1974. Effects ofnoise letters upon the identification of atarget letter in a nonsearch task. PerceptPsychophys 16:143–149.
Fan J, McCandliss BD, Flombaum JI, Pos-ner MI. 2001a. Imaging attentional net-works. San Diego, CA: Annual Meetingof the Society for Neuroscience.
Fan J, Wu Y, Fossella JA, Posner MI.2001b. Assessing the heritability of at-tentional networks. BMC Neurosci 2:14.
Fan J, McCandliss BD, Sommer T, Raz A,Posner MI. 2002. Testing the efficiencyand independence of attentional net-works. J Cogn Neurosci 14:340–347.
Fan J, Flombaum JI, McCandliss BD,Thomas KM, Posner MI. 2003a. Cogni-tive and brain consequences of conflict.Neuroimage 18:42–57.
Fan J, Raz A, Posner MI. 2003b. Atten-tional mechanisms. In: Aminoff MJ, Da-roff RB, editors. Encyclopedia of neuro-logical sciences. New York: ElsevierScience. p 292–299.
Fan J, Fossella J, Sommer T, Wu Y, PosnerMI. 2003c. Mapping the genetic varia-tion of executive attention onto brain ac-tivity. Proc Natl Acad Sci USA 100:7406–7411.
James W. 1890. The principles of psychol-ogy. New York: Dover.
Kastner S, Ungerleider LG. 2000. Mecha-nisms of visual attention in the humancortex. Annu Rev Neurosci 23:315–341.
Kastner S, Ungerleider LG. 2001. The neu-ral basis of biased competition in humanvisualcortex.Neuropsychologia39:1263–1276.
Kihlstrom JF. 1996. Perception withoutawareness of what is perceived, learningwithout awareness of what is learned. In:Velmans S, editor. The science of con-sciousness: psychological, neuropsycho-logical and clinical reviews. London:Routledge. p 23–46.
Klingberg T, Forssberg H, Westerberg H.2002. Training of working memory inchildren with ADHD. J Clin Exp Neuro-psychol 24:781–791.
34 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE
Klingberg T, Fernell E, Olesen PJ, JohnsonM, Gustafsson P, Dahlstrom K, GillbergCG, Forssberg H, Westerberg H. Com-puterized training of working memory inchildren with ADHD: a aontrolled, ran-domized, double-blind trial. J Am AssocChild Adolesc Psych (in press).
Kosslyn SM, Thompson WL, Costantini-Ferrando MF, Alpert NM, Spiegel D.2000. Hypnotic visual illusion alterscolor processing in the brain. Am J Psy-chiatry 157:1279–1284.
LaBerge D. 1994. Quantitative models ofattention and response processes inshape identification tasks. J Math Psy-chol 38:198–243.
Landau SM, Schumacher EH, Garavan H,Druzgal TJ, D’Esposito M. 2004. A func-tional MRI study of the influence ofpractice on component processes ofworking memory. Neuroimage 22:211–221.
Manly T, Lewis GH, Robertson IH, WatsonPC, Datta AK. 2002. Coffee in the corn-flakes: time-of-day as a modulator of ex-ecutive response control. Neuropsycho-logia 40:1–6.
Marrocco RT, Davidson MC. 1998. Neuro-chemistry of attention. In: ParasuramanR, editor. The attentional brain. Cam-bridge, MA: MIT Press. p 35–50.
Naatanen R. 2003. Mismatch negativity:clinical research and possible applica-tions. Int J Psychophysiol 48:179–188.
Navon D. 2003. What does a compoundletter tell the psychologist’s mind? ActaPsychol (Amst) 114:273–309.
Ochsner KN, Bunge SA, Gross JJ, GabrieliJD. 2002. Rethinking feelings: an FMRIstudy of the cognitive regulation of emo-tion. J Cogn Neurosci 14:1215–1229.
Olesen PJ, Westerberg H, Klingberg T.2004. Increased prefrontal and parietalactivity after training of working mem-ory. Nat Neurosci 7:75–79.
Pessoa L, Kastner S, Ungerleider LG. 2003.Neuroimaging studies of attention: frommodulation of sensory processing to top-down control. J Neurosci 23:3990–3998.
Posner MI, Fan J. 2004. Attention as anorgan system. In: Pomerantz JR, CraiMC, editors. Topics in integrative neuro-science: from cells to cognition. Cam-bridge: Cambridge University Press.
Posner MI, Petersen SE. 1990. The atten-tion system of the human brain. AnnuRev Neurosci 13:25–42.
Posner MI, Rothbart MK. Hebb’s neuralnetworks support the integration of psy-chological science. Canadian Psycholo-gist (in press).
Posner MI. 1980. Orienting of attention. QJ Exp Psychol 32:3–25.
Posner MI. 2004a. Cognitive neuroscienceof attention. New York: Guilford Press.
Posner MI. 2004b. The achievement ofbrain imaging: past and future. In: Kan-wisher N, Duncan J, editors. Attentionand performance XX. New York: OxfordUniversity Press. p 505–528.
Posner MI. 2004c. Tenth annual George A.Miller distinguished lecture at the an-nual meeting of the Cognitive Neuro-science Society. San Francisco: Cogni-tive Neuroscience Society.
Raymond JE, Shapiro KL, Arnell KM.1992. Temporary suppression of visualprocessing in an RSVP task: an atten-tional blink? J Exp Psychol Hum PerceptPerform 18:849–860.
Raz A. 1999. The effects of total sleep de-privation on the spotlight of visual atten-tion and on pre-attentional processing.Jerusalem: Hebrew University.
Raz A, Posner MI. 2000. Attention undersleep deprivation: mismatch negativityaltered by attentional state. J Cogn Neu-rosci 85C:81.
Raz A, Deouell LY, Bentin S. 2001. Is pre-attentive processing compromised byprolonged wakefulness? effects of totalsleep deprivation on the mismatch neg-ativity. Psychophysiology 38:787–795.
Raz A, Shapiro T. 2002. Hypnosis and neu-roscience: a cross talk between clinicaland cognitive research. Arch Gen Psychi-atry 59:85–90.
Raz A, Shapiro T, Fan J, Posner MI. 2002.Hypnotic suggestion and the modulationof Stroop interference. Arch Gen Psychi-atry 59:1155–1161.
Raz A, Landzberg KS, Schweizer HR, Ze-phrani ZR, Shapiro T, Fan J, Posner MI.2003. Posthypnotic suggestion and themodulation of Stroop interference undercycloplegia. Conscious Cogn 12:332–346.
Raz A. 2004a. Atypical attention: hypnosisand conflict reduction. In: Posner MI,editor. Cognitive neuroscience of atten-tion. New York: Guilford Press. p 420–429.
Raz A. 2004b. Attention. In: SpielberegerC, editor. Encyclopedia of applied psy-chology. San Diego, CA: Elsevier ScienceAcademic Press. p 203–208.
Raz A. 2004c. Neural substrates of atten-tion. In: Adelman G, Smith BH, editors.Encyclopedia of neuroscience. NewYork: Elsevier Science Academic Press.
Raz A, Marinoff GP, Landzberg KS, Guy-ton D. 2004a. Substrates of negative ac-commodation. Binocul Vis StrabismusQ 19:71–74.
Raz A, Marinoff GP, Zephrani ZR,Schweizer HR, Posner MI. 2004b. Seeclearly: suggestion, hypnosis, attention,and visual acuity. Int J Clin Exp Hypn52:159–187.
Rizzolatti G, Gentilucci M, Fogassi L, Lup-pino G, Matelli M, Ponzoni-Maggi S.1987. Neurons related to goal-directed
motor acts in inferior area 6 of the ma-caque monkey. Exp Brain Res 67:220–224.
Robbins TW, Milstein JA, Dalley JW. 2004.Neuropharmacology of attention. In:Posner MI, editor. Cognitive neuro-science of attention. New York: GuilfordPress. p 269–282.
Rothbart MK, Ellis LK, Posner MI. 2004.Temperament and self regulation. In:Baumeister RF, Vohs KD, editors. Hand-book of self regulation. New York: Guil-ford Press. p 357–370.
Rueda MR, Fan J, Halparin J, Gruber D,Posner MI. 2002. Assaying the develop-ment of attentional networks in six to tenyear old children. San Francisco: AnnualMeeting of the Cognitive NeuroscienceSociety.
Rueda MR, Fan J, McCandliss BD, Halpa-rin JD, Gruber DB, Lercari LP, PosnerMI. 2004a. Development of attentionalnetworks in childhood. Neuropsycholo-gia 42:1029–1040.
Rueda MR, Posner MI, Rothbart MK.2004b. Attentional control and selfregulation. In: Baumeister RF, VohsKD, editors. Handbook of self regula-tion. New York: Guilford Press. p 283–300.
Rueda MR, Fan J, McCandliss BD, Halpa-rin JD, Gruber DB, Lercari LP, PosnerMI. Development of attentional net-works in childhood. Neuropsychologia(in press).
Schneider W, Shiffrin RM. 1977. Con-trolled and automatic human informa-tion processing: I, detection, search andattention. Psychol Rev 84:1–66.
Shalev L, Algom D. 2000. Stroop andGarner effects in and out of Posner’sbeam: reconciling two conceptions ofselective attention. J Exp Psychol 26:997–1017.
Shapiro KL, Raymond JE, Arnell KM.1994. Attention to visual pattern infor-mation produces the attentional blink inrapid serial visual presentation. J ExpPsychol Hum Percept Perform 20:357–371.
Sommer T, Fossella JA, Fan J, Posner MI.Inhibitory control: cognitive subfunc-tions, individual differences and varia-tion in dopaminergic genes. Proccedingsof the Hanse Institute Frankfurt, Ger-many. (in press).
Spelke E, Hirst W, Neisser U. 1976. Skillsof divided attention. Cognition 4:215–230.
FEATURE ARTICLE THE ANATOMICAL RECORD (PART B: NEW ANAT.) 35
Stroop JR. 1935. Studies of interference inserial verbal reactions. J Exp Psychol 18:643–661.
Talgar CP, Carrasco M. 2002. Vertical me-ridian asymmetry in spatial resolution:visual and attentional factors. PsychonBull Rev 9:714–722.
Witte EA, Marrocco RT. 1997. Alteration ofbrain noradrenergic activity in rhesusmonkeys affects the alerting componentof covert orienting. Psychopharmacol-ogy (Berl) 132:315–323.
Witte EA, Davidson MC, Marrocco RT. 1997.Effects of altering brain cholinergic activityon covert orienting of attention: comparisonof monkey and human performance. Psy-chopharmacology (Berl) 132:324–334.
Wurtz RH, Goldberg ME. 1972. The primatesuperior colliculus and the shift of visualattention. Invest Ophthalmol 11:441–450.
Yeshurun Y, Carrasco M. 1998. Attentionimproves or impairs visual performanceby enhancing spatial resolution. Nature396:72–75.
Yeshurun Y, Carrasco M. 1999. Spatial at-tention improves performance in spatialresolution tasks. Vision Res 39:293–306.
Yeshurun Y, Carrasco M. 2000. The locusof attentional effects in texture segmen-tation. Nat Neurosci 3:622–627.
Zaidel E. 1995. Interhemispheric transferin the split brain: long-term status fol-lowing complete cerebral commissurot-omy. In: Davidson RH, Hugdahl, K, edi-tors. Brain asymmetry. Cambridge, MA:MIT Press. p 491–532.
36 THE ANATOMICAL RECORD (PART B: NEW ANAT.) FEATURE ARTICLE