Angiogenesis is crucial for liver regeneration after partial hepatectomy Yugo Uda, MD, a Tadamichi Hirano, MD, PhD, a Gakuhei Son, MD, PhD, a Yuji Iimuro, MD, PhD, a Naoki Uyama, MD, PhD, a Junichi Yamanaka, MD, PhD, a Akira Mori, MD, PhD, b Shigeki Arii, MD, PhD, c and Jiro Fujimoto, MD, PhD, a Nishinomiya, Gifu, and Tokyo, Japan Background. Recent studies of hepatic regeneration have mainly focused on the growth of parenchymal cells. However, remodeling of liver vessels seems to be crucial during hepatic regeneration. In this study, we investigated the influence of antiangiogenesis on hepatic regeneration using sFlt-1, a soluble receptor for vascular endothelial growth factor that acts as a dominant negative receptor, and the hepatocyte growth factor antagonist NK4. Methods. A sFlt-1–expressing adenoviral vector, an NK4-expressing adenoviral vector, or both combined were infected into C57BL6 mice via the tail vein. A 70% partial hepatectomy was performed on all of the mice 48 hours after infection. The remnants of the liver were removed after the partial hepatectomy, and hepatic regeneration was assessed by measuring the remnant liver weight and hepatocyte mitosis, bromodeoxyuridine staining, immunohistochemical staining with anti–platelet endothelial cell adhesion molecule-1 antibodies, and real-time polymerase chain reaction studies for angiogenic factors. Results. The immunohistochemical staining for CD31 showed suppression of sinusoidal endothelial cells growth in sFlt-1–expressing adenoviral vector–and NK4-expressing adenoviral vector–infected mice. Increases in the remnant hepatic weight were significantly lower in the sFlt-1–expressing adenoviral vector–infected mice. The bromodeoxyuridine index and mitotic cell results revealed a significant decrease in hepatic regeneration in the sFlt-1–expressing adenoviral vector–and NK4-expressing adenoviral vector–infected mice. The suppressive effects on hepatic regeneration were significantly enhanced by combined sFlt-1–expressing adenoviral vector and NK4-expressing adenoviral vector infection. Real-time polymerase chain reaction results revealed the significant suppression of angiogenic growth factor receptors Tie-1 and Tie-2. Conclusion. The angiogenesis inhibitor significantly suppressed hepatic regeneration. These results suggest that hepatic regeneration after hepatectomy closely correlates with angiogenesis. (Surgery 2013;153:70-7.) From the Departments of Surgery at Hyogo College of Medicine, a Nishinomiya and Asahi University Mura- kami Memorial Hospital, b Gifu; and the Department of Hepato-Biliary-Pancreatic Surgery, c Tokyo Medical and Dental University, Tokyo, Japan THE LIVER HAS THE UNIQUE CAPACITY to regenerate itself after injury or surgical resection. Under nor- mal conditions, hepatocytes are quiescent cells with a long life span. Once a loss of hepatic tissues (such as after a hepatectomy) occurs, however, hepatocytes re-enter the cell cycle accompanied by various growth responses. Regarding the transition from quiescent to pro- liferative hepatocytes, recent publications have clearly shown cytokine signaling after partial hepa- tectomy (PH). Tumor necrosis factor–alpha (TNF- a) 1 binding to its receptor on Kupffer cells, along with C3a, C5a, and myeloid differentiation factor 88 (MyD88), also activates nuclear factor kappa beta (NF-kb). 2,3 Interleukin-6 is subsequently released and binds to its receptors gp80 and gp130, which leads to hepatocyte proliferation via the activation of the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) and mitogen-activated protein kinase (MAPK) path- ways. 4-6 After these cytokines have triggered the G0 to G1 transition, cell cycle progression is then regu- lated by several growth factors. Among the growth factors that regulate liver regeneration, hepatocyte growth factor (HGF), epidermal growth factor (EGF), and transforming growth factor–alpha (TGF-a) are known to stimulate hepatocytes. 7-9 In addition, transforming growth factor-beta Supported by a Grant-in-Aid for Scientific Research (No. 17390375) from the Japan Society for the Promotion of Science and the Ministry of Health. Accepted for publication June 8, 2012. Reprint requests: Jiro Fujimoto, MD, PhD, Department of Sur- gery, Hyogo College of Medicine, 1.1. Mukogawacho, Nishino- miya, 663, Japan. E-mail: [email protected]. 0039-6060/$ - see front matter Ó 2013 Mosby, Inc. All rights reserved. http://dx.doi.org/10.1016/j.surg.2012.06.021 70 SURGERY
Transcript
Support1739037and the
Accepte
Reprintgery, Hymiya, 66
0039-60
� 2013
http://d
70 SU
Angiogenesis is crucial for liverregeneration after partial hepatectomyYugo Uda, MD,a Tadamichi Hirano, MD, PhD,a Gakuhei Son, MD, PhD,a Yuji Iimuro, MD, PhD,a
Shigeki Arii, MD, PhD,c and Jiro Fujimoto, MD, PhD,a Nishinomiya, Gifu, and Tokyo, Japan
Background. Recent studies of hepatic regeneration have mainly focused on the growth of parenchymalcells. However, remodeling of liver vessels seems to be crucial during hepatic regeneration. In this study,we investigated the influence of antiangiogenesis on hepatic regeneration using sFlt-1, a soluble receptorfor vascular endothelial growth factor that acts as a dominant negative receptor, and the hepatocytegrowth factor antagonist NK4.Methods. A sFlt-1–expressing adenoviral vector, an NK4-expressing adenoviral vector, or both combinedwere infected into C57BL6 mice via the tail vein. A 70% partial hepatectomy was performed on all ofthe mice 48 hours after infection. The remnants of the liver were removed after the partial hepatectomy,and hepatic regeneration was assessed by measuring the remnant liver weight and hepatocyte mitosis,bromodeoxyuridine staining, immunohistochemical staining with anti–platelet endothelial cell adhesionmolecule-1 antibodies, and real-time polymerase chain reaction studies for angiogenic factors.Results. The immunohistochemical staining for CD31 showed suppression of sinusoidal endothelial cellsgrowth in sFlt-1–expressing adenoviral vector–and NK4-expressing adenoviral vector–infected mice.Increases in the remnant hepatic weight were significantly lower in the sFlt-1–expressing adenoviralvector–infected mice. The bromodeoxyuridine index and mitotic cell results revealed a significant decreasein hepatic regeneration in the sFlt-1–expressing adenoviral vector–and NK4-expressing adenoviralvector–infected mice. The suppressive effects on hepatic regeneration were significantly enhanced bycombined sFlt-1–expressing adenoviral vector and NK4-expressing adenoviral vector infection. Real-timepolymerase chain reaction results revealed the significant suppression of angiogenic growth factorreceptors Tie-1 and Tie-2.Conclusion. The angiogenesis inhibitor significantly suppressed hepatic regeneration. These resultssuggest that hepatic regeneration after hepatectomy closely correlates with angiogenesis. (Surgery2013;153:70-7.)
From the Departments of Surgery at Hyogo College of Medicine,a Nishinomiya and Asahi University Mura-kami Memorial Hospital,b Gifu; and the Department of Hepato-Biliary-Pancreatic Surgery,c Tokyo Medicaland Dental University, Tokyo, Japan
THE LIVER HAS THE UNIQUE CAPACITY to regenerateitself after injury or surgical resection. Under nor-mal conditions, hepatocytes are quiescent cellswith a long life span. Once a loss of hepatic tissues(such as after a hepatectomy) occurs, however,hepatocytes re-enter the cell cycle accompaniedby various growth responses.
Regarding the transition from quiescent to pro-liferative hepatocytes, recent publications have
ed by a Grant-in-Aid for Scientific Research (No.5) from the Japan Society for the Promotion of ScienceMinistry of Health.
d for publication June 8, 2012.
requests: Jiro Fujimoto, MD, PhD, Department of Sur-ogo College of Medicine, 1.1. Mukogawacho, Nishino-3, Japan. E-mail: [email protected].
60/$ - see front matter
Mosby, Inc. All rights reserved.
x.doi.org/10.1016/j.surg.2012.06.021
RGERY
clearly shown cytokine signaling after partial hepa-tectomy (PH). Tumor necrosis factor–alpha (TNF-a)1 binding to its receptor on Kupffer cells, alongwith C3a, C5a, and myeloid differentiation factor88 (MyD88), also activates nuclear factor kappabeta (NF-kb).2,3 Interleukin-6 is subsequentlyreleased and binds to its receptors gp80 andgp130, which leads to hepatocyte proliferation viathe activation of the Janus kinase (JAK)/signaltransducer and activator of transcription (STAT)andmitogen-activated protein kinase (MAPK) path-ways.4-6 After these cytokines have triggered the G0to G1 transition, cell cycle progression is then regu-lated by several growth factors. Among the growthfactors that regulate liver regeneration, hepatocytegrowth factor (HGF), epidermal growth factor(EGF), and transforming growth factor–alpha(TGF-a) are known to stimulate hepatocytes.7-9
1 (TGF-b1) is known to inhibit proliferation and ter-minate liver regeneration.10 In addition to thesegrowth factors, the role of nonparenchymal cellsduring liver regeneration has been a focus of inter-est. Vascular endothelial growth factor (VEGF)expression increases after PH, and sinusoidal endo-thelial cell proliferation has been shown to occurduring liver regeneration.11-13
Angiogenesis is the formation of new micro-vessels from pre-existing vessels. It is essential forwound healing, regeneration, and organ develop-ment. Angiogenesis has recently received muchattention because it plays a pivotal role in tumorgrowth and metastasis. Various antiangiogenic can-cer therapies have been extensively studied, andseveral authors have reported the remarkabletherapeutic effects of antiangiogenic gene therapyfor hepatocellular carcinoma.14,15 However, thereis a distinct difference between tumor-relatedangiogenesis and angiogenesis in physiologic re-sponses, and the relationship between angiogene-sis and the growth of sinusoidal endothelial cellsin the regenerating liver has been studied lessextensively.
In the present study, we investigated whetherangiogenesis is accompanied by liver regenerationand is indispensable for liver regeneration. To dothis, we used 2 antiangiogenic gene expressingadenoviral vectors: sFlt-1–expressing adenoviralvector (Ad.Fex), which expresses a soluble formof VEGF receptor 1 (sFlt-1) that is involved in thehigh-affinity binding of VEGF and indirect inhibi-tion of angiogenesis,14,16 and an NK4-expressingadenoviral vector (Ad.NK4), a proteolytic diges-tion product of HGF that acts as a specific antago-nist to HGF. HGF is a potent angiogenic factor thatstimulates endothelial cell motility and prolifera-tion. NK4 inhibits HGF-mediated angiogenesisand strongly inhibits angiogenesis driven byVEGF and basic fibroblast growth factor (bFGF).17
MATERIALS AND METHODS
Adenoviral vectors. Two adenoviral vectors wereused in this study. The Ad.Fex vector was based ona replication-deficient recombinant adenovirustype 5 (Ad5). It incorporates E1a and partial E1band E3 deletions that contains the cytomegalovirus(CMV) early enhancer/chicken b actin promoterand enhancer. The cDNA encodes the first 3immunoglobulin-like loops of sFlt-1, which areinvolved in high-affinity binding of VEGF, but lacksthe trans membrane and intracellular domains.16
The Ad.NK4 vector was based on a replication-deficient recombinant Ad5 with E1 and E3
deletions. It expresses the NK4 gene under thetranscriptional control of the CMV immediate-early enhancer and promoter.18 NK4 acts as a spe-cific antagonist to HGF, and it strongly inhibitsangiogenesis driven by VEGF and bFGF.17 A hu-man embryonic kidney cell line (293) was obtainedfrom the American Type Culture Collection (Man-assas, VA) and grown in Dulbecco’s modifiedEagle’s medium with 10% fetal calf serum. The re-combinant virus was grown in the 293 cells andthen purified by cesium chloride gradient ultra-centrifugation. The titers of the viral stocks weredetermined by plaque assays using the 293 cells.Ad.LacZ, which expresses the LacZ gene underthe control of CMV promoter, was used asa control vector.
Experimental animal models. Male C57/BL6mice, 18 g to 21 g and 6 to 7 weeks old, werepurchased from CLEA Japan, Inc (Tokyo Japan).First, the serum protein concentrations of sFlt-1 and NK4 were evaluated by injecting 109 pfu ofeach adenoviral vector (n = 8 each) via the tailvein. Blood was obtained from the tail vein at 1,2, 3, 7, and 14 days postinjection. The serum pro-tein concentrations of sfFt-1 and NK4 were deter-mined by enzyme-linked immunosorbent assayusing the Quantikine Mouse Soluble VEGF R1 Im-munoassay kit (R&D Systems, Minneapolis, MN) oran IMMUNIS human HGF enzyme immunoassaykit (Institute of Immunology, Tokyo, Japan).
Next, the mice were divided into 4 groups toreceive different treatments: Ad.NK4 (109 pfu);Ad.Fex (109 pfu); Ad.NK4 plus Ad.Fex (5 3 108
pfu each); and Ad.LacZ (109 pfu) as a control.Each adenoviral vector was injected via the tailvein. Two days after injecting the adenoviralvectors, a two-thirds partial hepatectomy was per-formed under ether anesthesia.19 The mice werekilled at 1, 1.5, 2, 3, 5, and 7 days after PH, andthe livers were removed for analysis. The micewere maintained in a temperature-controlledroom on a 12-hour light/dark cycle, with freeaccess to standard water and chow. All experimen-tal animal research protocols in the present studywere submitted and approved by the experimentalanimal review committee of the Hyogo College ofMedicine.
Immunohistochemistry. Bromodeoxyuridine expres-sions and mitotic cell counts: For bromodeoxyuridine(BrdU) staining, animals were intraperitoneallyinjected with BrdU (Sigma, St Louis, MO) (50mg/kg of a 0.2% solution in phosphate-bufferedsaline) 1 hour before their liver specimens wereharvested. The liver sections were incubated withmouse monoclonal anti-BrdU Ab (Novocastra,
SurgeryJanuary 2013
72 Uda et al
Newcastle upon Tyne, UK), as has been previouslydescribed.20 BrdU expression was examined with aNikon ECLIPSE E600 (Nikon, Tokyo, Japan). Thepercentage of BrdU-positive cells in 1000 hepato-cytes was termed the labeling index. To assess thenumber of hepatocyte mitoses, the liver sectionswere stained with hematoxylin–eosin. The mitotichepatocytes were counted in 50 high-power(3 200) fields on each slide, and the countswere expressed as mitotic cells per high-powerfield.
Sinusoidal endothelial cells: Endothelial cell stain-ing was performed with a rat anti-mouse CD31(platelet endothelial cell adhesion molecule-1;PECAM-1) monoclonal antibody (BD Biosciences,Franklin Lakes, NJ) at a 1:200 dilution. Theparaffin-embedded sections were cut at a thicknessof 4 mm. Endogenous peroxidase was blocked byincubating with 0.3% H2O2 for 30 minutes. Thesections were incubated with protein blockserum-free (DAKO, Glostrup, Denmark) for 30minutes, followed by the PECAM-1 antibody at48C overnight. Histofine Simplex Stain MouseMAX-PO (rat; Nichirei, Tokyo, Japan) was thenapplied for 60 minutes. The sections were incubatedwith 3-amino-9-ethylcarbazole chromogen, followedby counterstaining with Mayer’s hematoxylin.
Determination of the sinusoidal endothelial cell area:To assess angiogenesis associated with liver regen-eration, the percentage of the total liver area thatwas immunostained with PECAM was determined.Ten 200-fold magnification fields were randomlyselected in the portal and central areas. Theimages were scanned with a microscope viewingsystem (Digital Sight, Nikon, Japan) and evaluatedby image analyzer software (Scion Image; ScionCorp, Frederick, MD).
Measurement of angiogenic growth factor geneexpression by real-time polymerase chain reactionstudies. Total RNA was extracted from the liverspecimens using an ISOGEN kit (Nippon Gene,Tokyo, Japan), according to the manufacturer’sguidelines. The RNA concentrations were measuredspectrophotometrically at 260 nm. For each sample,1 mg of total RNAwas used. Reverse transcription wasperformed using the High Capacity cDNA ReverseTranscription Kit (Applied Biosystems, Foster City,CA) according to the manufacturer’s guidelines.The primers and probes for the real-time reversetranscription polymerase chain reaction (RT-PCR)were provided by Applied Biosystems as ready-to-usemixes. The product identification numbers of themixes were Mm00456499_m1 for angiopoietin-1,Mm01223051_m1 for angiopoietin-2, Mm01180914-ml for Tie-1, Mm01256904_m1 for Tie-2, and
Hs99999901_s1 for eukaryotic 18s rRNA. RT-PCRwas performed using the Applied Biosystems 7300Real Time PCR system (Applied Biosystems). Ahousekeeping gene, 18S rRNA, served as the endog-enous control.
Cell proliferation assay. A human hepatocytecell line, NMuli, was grown in a Dulbecco’s mod-ified Eagle’s medium with 10 % fetal calf serum.The NMuLi cells (2 3 105) were infected withAd.NK4, Ad.Fex, Ad.NK4 plus Ad.Fex, andAd.LacZ at a multiplicity of infection of 30. Thecells were harvested at 1, 3, 5, and 7 days after in-fection and counted using a crystal violet stainingprocedure.15
Statistical methods. The unpaired Student t testwas used for comparing the continuous variablesbetween groups, and P < .05 was considered statis-tically significant.
RESULTS
In vivo transduction of Ad.NK4 or Ad.Fex viathe tail vein. Serum Nk4 and sFlt-1 proteins weremeasured by enzyme-linked immunosorbent assay.In the mice receiving 109 pfu of Ad.NK4 via the tailvein, the serum NK4 protein reached a peak value>200 ng/mL at 3 days after transduction and grad-ually declined to 0 ng/mL by 14 days after trans-duction (Fig 1, A). In the mice receiving 109 pfuof Ad.Fex, the serum sFlt-1 protein peaked at>300 ng/mL at 24 hours after transduction andgradually declined to 0 ng/mL by 14 days aftertransduction (Fig 1, B).
Sinusoidal endothelial cell area. The change inthe percentage of sinusoidal endothelial cell areaout of the total liver tissue area after PH wasdetermined. In the control mice that were trans-fected Ad.LacZ, the sinusoidal endothelial cellarea gradually decreased until 72 hours after PHand then increased to the preoperative levels at120 hours after PH. In Ad.NK4- and Ad.Fex-transfected mice, however, a significant increasein the sinusoidal endothelial cell area was notobserved at 120 hours after PH (P < .05 for bothNK4 and sFlt-1; Fig 1, C). In the AdNK4 and Ad.-Fex–transfected mice, the percentage of sinusoidalendothelial cell area was significantly lower at72 hours (P < .05) and at 120 hours (P < .01)than in the Ad.LacZ-transfected mice.
Liver regeneration in the Ad.NK4 and Ad.sFlt-1–transfected mice. Remnant liver weight: Transfectionwith Ad.NK4 or Ad.Fex affected both the vasculardensity after PH and the liver regeneration kinet-ics. The liver/body weight ratios of the Ad.NK4-and Ad.sFlt-1–transfected mice were significantly
Fig 1. The protein expressions of NK4 (A) and sFlt-1 (B) in mouse plasma. (C) The percentage of endothelial cell areaafter partial hepatectomy. The values represent mean ± standard deviation (*P < .05; **P < .01). At each point, n = 6.(D) The liver/body ratio after partial hepatectomy. The values represent the mean ± standard deviation (*P < .05). Ateach point, n = 6.
SurgeryVolume 153, Number 1
Uda et al 73
lower than that of control mice (at 3 days and 5days for NK4 and at 3 days for sFlt-1). At 7 days, theratios in the 2 groups were similar (Fig 1, D, leftand middle). Transfection with combinedAd.NK4 and Ad.Fex decreased the recovery of livermass at 3 and 5 days after the PH (Fig 1, D, right).
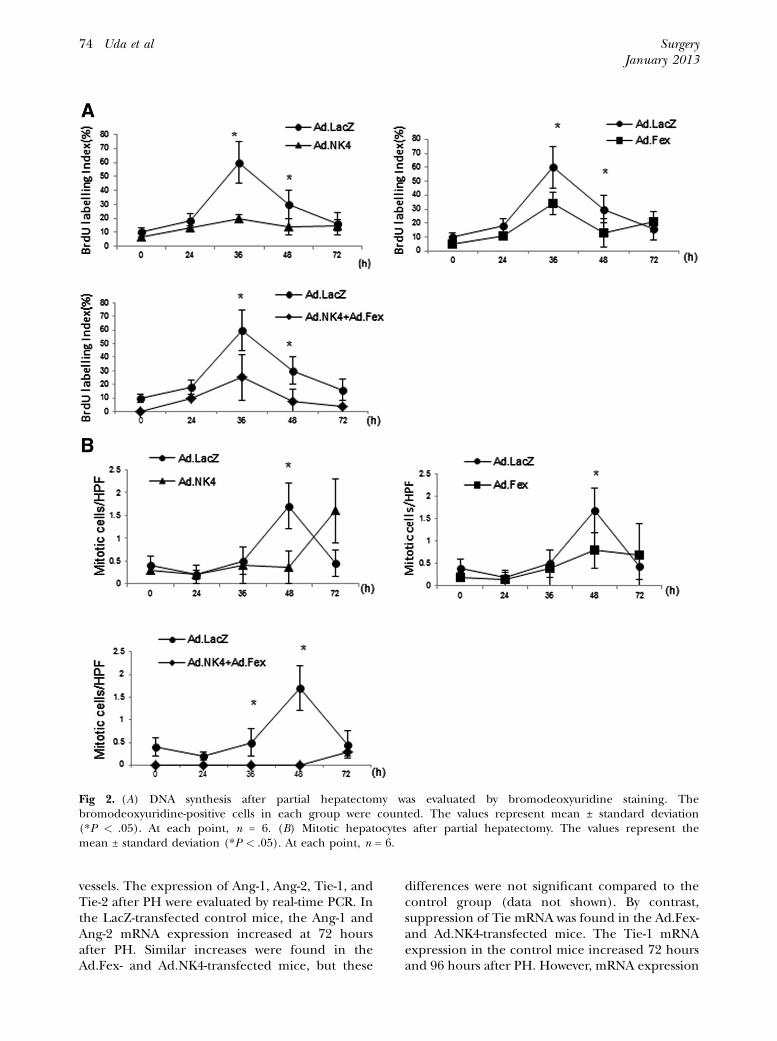
Hepatocyte DNA synthesis: We examined whetherthe antiangiogenic factors impaired hepatocytereplication by evaluating the DNA synthesis usingBrdU staining and a mitotic hepatocyte count afterPH. An increase in the proportion of hepatocytesthat were BrdU-positive after PH was seen in theLacZ-transfected mice. This proportion reached apeak at 36 hours and had subsequently decreasedby 72 hours. The Ad.NK4- and Ad.Fex-transfectedmice exhibited distinct BrdU uptake kinetics.BrdU uptake did not significantly increase at 36and 48 hours after PH compared with the controlmice (Fig 2, A). We also estimated the number ofmitotic hepatocytes at various times after PH. Asa previous report has shown, the kinetics of mitotichepatocytes are similar to those of BrdU staining,but with a 12- to 24-hour delay in the controlmice.4 In LacZ-transfected mice, the number of
mitotic hepatocytes reached a peak at 48 hoursand then decreased. However, the numbers of mi-totic hepatocytes were significantly lower at 48hours in the NK4- and sFlt-1–transfected mice,and at 36 and 48 hours in the mice transfectedwith both (Fig 2, B). These results indicate theimportance of angiogenesis in liver regenerationafter hepatectomy.
Effects of NK4 or sFlt-1 on hepatocyte prolifer-ation in vitro. To exclude any direct effects of NK4or sFlt-1 on hepatocyte proliferation, we evaluatedwhether NK4 or sFlt-1 affected the proliferation ofcultured NMuli hepatocytes. NMuLi cells(2 3 105) were infected with Ad.NK4, Ad.Fex,Ad.NK4 and Ad.Fex, and Ad.LacZ at a multiplicityof infection of 30. The cells were harvested at 1, 3,5, and 7 days after infection and counted using acrystal violet staining procedure. No significanteffect on cell proliferation was seen (Fig 3).
Real-time PCR quantification of angiopoietinand its receptor. The angiogenic growth factorangiopoietin (Ang) and its receptor (Tie) areknown to be crucial for the formation and remod-eling of a supportive structure around nascent
Fig 2. (A) DNA synthesis after partial hepatectomy was evaluated by bromodeoxyuridine staining. Thebromodeoxyuridine-positive cells in each group were counted. The values represent mean ± standard deviation(*P < .05). At each point, n = 6. (B) Mitotic hepatocytes after partial hepatectomy. The values represent themean ± standard deviation (*P < .05). At each point, n = 6.
SurgeryJanuary 2013
74 Uda et al
vessels. The expression of Ang-1, Ang-2, Tie-1, andTie-2 after PH were evaluated by real-time PCR. Inthe LacZ-transfected control mice, the Ang-1 andAng-2 mRNA expression increased at 72 hoursafter PH. Similar increases were found in theAd.Fex- and Ad.NK4-transfected mice, but these
differences were not significant compared to thecontrol group (data not shown). By contrast,suppression of Tie mRNA was found in the Ad.Fex-and Ad.NK4-transfected mice. The Tie-1 mRNAexpression in the control mice increased 72 hoursand 96 hours after PH. However, mRNA expression
Fig 3. A human hepatocyte line, NMuli, was transfectedwith an adenoviral vector and grown in medium. Over a7-day period, cell proliferation was not significantlysuppressed by adenoviral transfection. Each value repre-sents the mean ± standard deviation for triplicatesamples.
Fig 4. The time course of Tie-1 (A) and Tie-2 (B) mRNAexpression after partial hepatectomy is shown. Thevalues represent mean ± standard deviation (*P < .05).At each point, n = 6.
SurgeryVolume 153, Number 1
Uda et al 75
did not increase in the Ad.Fex- or Ad.NK4-transfected mice, and the amount of mRNA wassignificantly lower than for the controls (Fig 4, A).With regard to Tie-2 mRNA expression, the sup-pression phenomenon was profound and lastingto 120 hours (Fig 4, B).
DISCUSSION
All mammalian cells must be surrounded by avascular network within 200 mm to be supplied withoxygen and nutrition.21 When the cell numbers in-crease, the distance between cells and vessels phys-ically increases, leading to the formation of newvessels from the pre-existing vascular network.Folkman22 has previously shown the crucial roleof angiogenesis in tumor growth, and angiogenesishas received much attention in disease pathology.However, less is known about the role of angiogen-esis in physiologic responses, including liver regen-eration. The regulatory contribution of severalcytokines and growth factors to the transitionfrom quiescent to proliferative hepatocytes hasbeen extensively studied. Among them, the inter-leukin-6/gp130-dependent pathways that followthe activation of JAK/STAT and MAPK are keysignals for liver regeneration. However, Wuestefeldet al23 have reported on abundant DNA synthesisin hepatocytes after PH, implying that anotherfactors contribute to liver regeneration in the earlyphases after PH.
Our findings indicate a key role for angiogen-esis in liver regeneration after PH. Two differentantiangiogenic agents, sFlt-1 and NK4, significantlydecreased the number of endothelial cells found
after PH. DNA synthesis, as assessed by BrdUstaining, was suppressed at 36 and 48 hours afterPH, and hepatocyte proliferation also decreased at48 hours after PH, leading to a delay in liverregeneration. Several reports have shown the rela-tion of angiogenesis to liver regeneration. Whenantiangiogenic agents (such as anti-VEGF anti-bodies and angiostatin) are administered, theproliferative activity of hepatocytes is inhibitedand liver regeneration decreases.13,24 By contrast,angiogenic agents (such as bFGF) increase liver re-generation.25 These findings support our resultsand reinforce the importance of angiogenesis inliver regeneration. In our results, liver weight/body ratios and BrdU uptake were less in the treat-ment group but became equal with time. We thinkthis phenomenon may be caused by facilitation ofanother related factor with regard to liver regener-ation. In findings similar to our present study, wepreviously reported a crucial role of MyD88, whichis a common adaptor molecule of Toll-like recep-tors. The liver weight/body ratio of MyD88�/�
mice was significantly lower than that of wild-typemice at 48 and 72 hours after PH; by 96 hours,the ratios in the 2 groups were similar.2 Moreover,Ng et al recently reported that knockdown ofmiR-21 impaired progression of the hepatocyte’s
SurgeryJanuary 2013
76 Uda et al
cell cycle after PH, mainly through a decrease ofcyclin D1 expression. However, overexpression ofcyclin E compensates the lack of cyclin D1 and fa-cilitates cell cycle of hepatocytes.26 Therefore, theinhibition of angiogenesis truly suppressed liverregeneration; however, other factors may compen-sate and facilitate liver regeneration.
Al1 contribute to boost the effect. sFlt-1 binds toVEGF with high-affinity and indirectly inhibit VEGFsignaling.14,16 On the other hand, NK4 does notblock VEGF-induced angiogenic signals in the earlyphase because NK4 allowed VEGF-induced kinasedomain-containing receptor (KDR)/VEGF recep-tor tyrosine phosphorylation and VEGF-inducedactivation of extracellular signal-regulated kinases1 and 2. Therefore, a likely explanation is thatNK4may block angiogenic signals through putativebindingmolecules on endothelial cells, not throughKDR/VEGF receptor.17 The two-point blocking ofVEGF signals by combined transfection has thepossibility to boost the effect compared to singletransfection.
The Ang/Tie pathway is also crucial to theremodeling and formation of new vessels, and themRNA expression of the Ang-1 and Ang-2 angio-genic growth factors and of the Tie-1 and Tie-2receptors increases after PH.11 Our real-time PCRresults found increases in the mRNA expression ofAng-1 and Ang-2 and of the Tie-1 and Tie-2 recep-tors in the Ad.LacZ-transfected mice that weresimilar to those found in previous reports. NeithersFlt-1 nor NK4 influenced the expression of Ang-1 andAng-2, which regulate vessel stability by activat-ing or antagonizing signaling viaTie-2.However, theantiangiogenic agents significantly suppressedexpression of the Tie-1 and Tie-2 receptors. Theorphan receptor Tie-1 is thought to be involved inmaintaining vessel integrity.27 Because VEGFinduces and enhances the expression of Tie-1,28
Tie-1 suppression by sFlt-1 and NK4 is understand-able, considering its ability to inhibit VEGF. How-ever, the mechanisms of sFlt-1 and NK4 Tie-2suppression have not been elucidated, because littleis known about the signals regulating Tie-2 otherthan hypoxia, TNF-a, and IL-1b.29 Additional analy-sis will be required to clarify the precisemechanismsconnecting antiangiogenic factors and the Ang/Tiepathway.
As we and other authors have shown, there isa lag between sinusoidal cell proliferation andhepatocyte proliferation.11,12 During liver regenera-tion, a biphasic proliferative wave was seen in the he-patocytes 1 to 3 days after the PH, after which thesinusoidal cells began to proliferate. Unsolved ques-tions remain as to when and how the antiangiogenic
agents act to inhibit hepatocyte proliferation. Dindet al30 have recently reported that liver sinusoidalendothelial cells (LSECs) play a crucial role in trig-gering the proliferation of hepatocytes. They clearlyshowed that PH-induced VEGF receptor activationon the surfaces of LSECs, initiating Id1 upregula-tion in LSECs and secrete of HGF and Wnt2, thenhepatocytes were necessary conditions for proli-feration in the early stage after PH.30 Althoughadditional studies are required, it is possible thatantiangiogenic agents inhibit such an interactionbetween LSECs and hepatocytes in the early phasesafter PH, and then suppressed hepatocyte prolifera-tion results in the delay of liver regeneration, notonly inhibition of the supply of oxygen andnutrition.
In conclusion, the angiogenesis inhibitors sFlt-1 and NK4 significantly decreased the number ofendothelial cells and suppressed hepatic regener-ation. These results suggest that hepatic regener-ation after hepatectomy is closely correlated withangiogenesis.
We thank Ms Keiko Mitani, Ms Aiko Wakahara, andMrs Kazuko Aizawa for their technical assistance.
REFERENCES
1. Yamada Y, Kirillova I, Peschon JJ, Fausto N. Initiation ofliver growth by tumor necrosis factor: deficient liverregeneration in mice lacking type I tumor necrosis factorreceptor. Proc Natl Acad Sci U S A 1997;94:1441-6.
2. Seki E, Tsutsui H, Iimuro Y, Naka T, Son G, Akira S, et al.Contribution of Toll-like receptor/myeloid differentiationfactor 88 signaling to murine liver regeneration. Hepatol-ogy 2005;41:443-50.
3. Strey CW, Markiewski M, Mastellos D, Tudoran R, SpruceLA, Greenbaum LE, et al. The proinflammatory mediatorsC3a and C5a are essential for liver regeneration. J Exp Med2003;198:913-23.
4. Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G,Furth EE, Poli V, et al. Liver failure and defective hepato-cyte regeneration in interleukin-6-deficient mice. Science1996;274:1379-83.
5. Levy DE, Lee CK. What does Stat3 do? J Clin Invest 2002;109:1143-8.
6. Talarmin H, Rescan C, Cariou S, Glaise D, Zanninelli G, Bi-lodeau M, et al. The mitogen-activated protein kinase ki-nase/extracellular signal-regulated kinase cascadeactivation is a key signalling pathway involved in the regula-tion of G(1) phase progression in proliferating hepatocytes.Mol Cell Biol 1999;19:6003-11.
7. Fausto N. Growth factors in liver development, regenera-tion and carcinogenesis. Prog Growth Factor Res 1991;3:219-34.
8. Matsumoto K, Nakamura T. Hepatocyte growth factor:molecular structure, roles in liver regeneration, and otherbiological functions. Crit Rev Oncog 1992;3:27-54.
10. Braun L, Mead JE, Panzica M, Mikumo R, Bell GI, Fausto N,et al. Transforming growth factor beta mRNA increases dur-ing liver regeneration: a possible paracrine mechanism ofgrowth regulation. Proc Natl Acad Sci U S A 1988;85:1539-43.
11. Sato T, El-Assal ON, Ono T, Yamanoi A, Dhar DK, NagasueN, et al. Sinusoidal endothelial cell proliferation and ex-pression of angiopoietin/Tie family in regenerating ratliver. J Hepatol 2001;34:690-8.
12. Shimizu H, Miyazaki M, Wakabayashi Y, Mitsuhashi N, KatoA, Ito H, et al. Vascular endothelial growth factor secretedby replicating hepatocytes induces sinusoidal endothelialcell proliferation during regeneration after partial hepatec-tomy in rats. J Hepatol 2001;34:683-9.
13. Taniguchi E, Sakisaka S, Matsuo K, Tanikawa K, Sata M. Ex-pression and role of vascular endothelial growth factor inliver regeneration after partial hepatectomy in rats.J Histochem Cytochem 2001;49:121-30.
14. Graepler F, Verbeek B, Graeter T, Smirnow I, Kong HL,Schuppan D, et al. Combined endostatin/sFlt-1 antiangio-genic gene therapy is highly effective in a rat model ofHCC. Hepatology 2005;41:879-86.
15. Son G, Hirano T, Seki E, Iimuro Y, Nukiwa T, Matsumoto K,et al. Blockage of HGF/c-Met system by gene therapy(adenovirus-mediated NK4 gene) suppresses hepatocellularcarcinoma in mice. J Hepatol 2006;45:688-95.
16. Yang W, Arii S, Mori A, Furumoto K, Nakao T, Isobe N, et al.sFlt-1 gene-transfected fibroblasts: a wound-specific genetherapy inhibits local cancer recurrence. Cancer Res 2001;61:7840-5.
17. Kuba K, Matsumoto K, Date K, Shimura H, Tanaka M,Nakamura T, et al. HGF/NK4, a four-kringle antagonistof hepatocyte growth factor, is an angiogenesis inhibitorthat suppresses tumor growth and metastasis in mice.Cancer Res 2000;60:6737-43.
18. Maemondo M, Narumi K, Saijo Y, Usui K, Tahara M, TazawaR, et al. Targeting angiogenesis and HGF function using anadenoviral vector expressing the HGF antagonist NK4 forcancer therapy. Mol Ther 2002;5:177-85.
19. Higgins G, Anderson R. Experimental pathology ofthe liver. I. Restoration of the liver in the white rat
following partial surgical removal. Arch Pathol 1931;12:186-202.
20. Ogushi I, Iimuro Y, Seki E, Son G, Hirano T, Hada T, et al.Nuclear factor kappa B decoy oligodeoxynucleotidesprevent endotoxin-induced fatal liver failure in a murinemodel. Hepatology 2003;38:335-44.
21. Carmeliet P, Jain RK. Angiogenesis in cancer and otherdiseases. Nature 2000;407:249-57.
22. Folkman J. The role of angiogenesis in tumor growth.Semin Cancer Biol 1992;3:65-71.
23. Wuestefeld T, Klein C, Streetz KL, Betz U, Lauber J, Buer J,et al. Interleukin-6/glycoprotein 130-dependent pathwaysare protective during liver regeneration. J Biol Chem2003;278:11281-8.
24. Drixler TA, Vogten MJ, Ritchie ED, van Vroonhoven TJ,Gebbink MF, Voest EE, et al. Liver regeneration is anangiogenesis-associated phenomenon. Ann Surg 2002;236:703-11.
25. Greene AK, Wiener S, Puder M, Yoshida A, Shi B, Perez-Atayde AR, et al. Endothelial-directed hepatic regenerationafter partial hepatectomy. Ann Surg 2003;237:530-5.
26. Ng R, Song G, Roll GR, Frandsen NM, Willenbring H.A microRNA-21 surge facilitates rapid cyclin D1 translationand cell cycle progression in mouse liver regeneration.J Clin Invest 2012;122:1097-108.
27. Rodewald HR, Sato TN. Tie1, a receptor tyrosine kinase es-sential for vascular endothelial cell integrity, is not criticalfor the development of hematopoietic cells. Oncogene1996;12:397-404.
28. McCarthy MJ, Crowther M, Bell PR, Brindle NP. The endo-thelial receptor tyrosine kinase tie-1 is upregulated byhypoxia and vascular endothelial growth factor. FEBS Lett1998;423:334-8.
29. Willam C, Koehne P, Jurgensen JS, Grafe M, Wagner KD,Bachmann S, et al. Tie2 receptor expression is stimulatedby hypoxia and proinflammatory cytokines in humanendothelial cells. Circ Res 2000;87:370-7.
30. Ding BS, Nolan DJ, Butler JM, James D, Babazadeh AO,Rosenwaks Z, et al. Inductive angiocrine signals fromsinusoidal endothelium are required for liver regeneration.Nature 2010;468:310-5.