53
Antigen recognition in innate and adaptive immunity Diversity in antigen recognition receptors
| Date post: | 18-Dec-2015 |
| Category: |
Documents |
| Upload: | chastity-harrell |
| View: | 219 times |
| Download: | 0 times |

Antigen recognition in innate and adaptive immunity
Diversity in antigen recognition receptors

Immune system
The ability to distinguish self from non-self

Innate immunity
• Very old. Every organism has some form of innate defence system.
• Does not adapt. Must rely of unique molecular features of pathogens.
• Must develop a large number of different receptor molecules, each with the capacity to distinguish some component unique to a pathogen.

Combination of innate and ad

Adaptive immunity
• Only found in jawed fish and above.• Happened 500 million years ago.• Uses VDJ recombination.• Small number of similar receptor molecules
(Ig, TcR and MHC) but the genes have been duplicated and segmented to allow recombination. Produces vast repertoire of “binding molecules”.
• TcR is unique because it has to bind MHC.

Adaptive immunity
• Humoral– B cells make soluble immunoglobulin.– 5 classes beginning with IgM that switch to other classes
depending on site, or function
• Cellular T cells
• Helper CD4 cells make cytokines to drive B-cells and T cells to develop
• Cytotoxic CD8 killer cells have mechanism to kill infected cells• NK T cells – unique TcR receptor for recognising non-peptide
antigens T cells have limited repertoire. Important for recognising
non-peptide antigens and non-MHC antigens.

Did the adaptive immune system evolve from the innate
system?
Two orders of agnathans - Hagfish and lamprey – the most disgusting animal (parasite) on the planet

Agnathans• Diverse haematological cells – heterogenous leukocytes• Produce opsonins and agglutinins• Allograft rejection• DO NOT have MHC, Ig, TcR or RAG-1/RAG-2 genes• Have their own adaptive immune receptors VLR-A (T
cell like) and VLR-B (B-cell like).• LRR receptors that can rearrange somatically.• Mechanism is unknown, but probably a transposase.• 35 VLR-A molecules and 38 VLR-B molecules
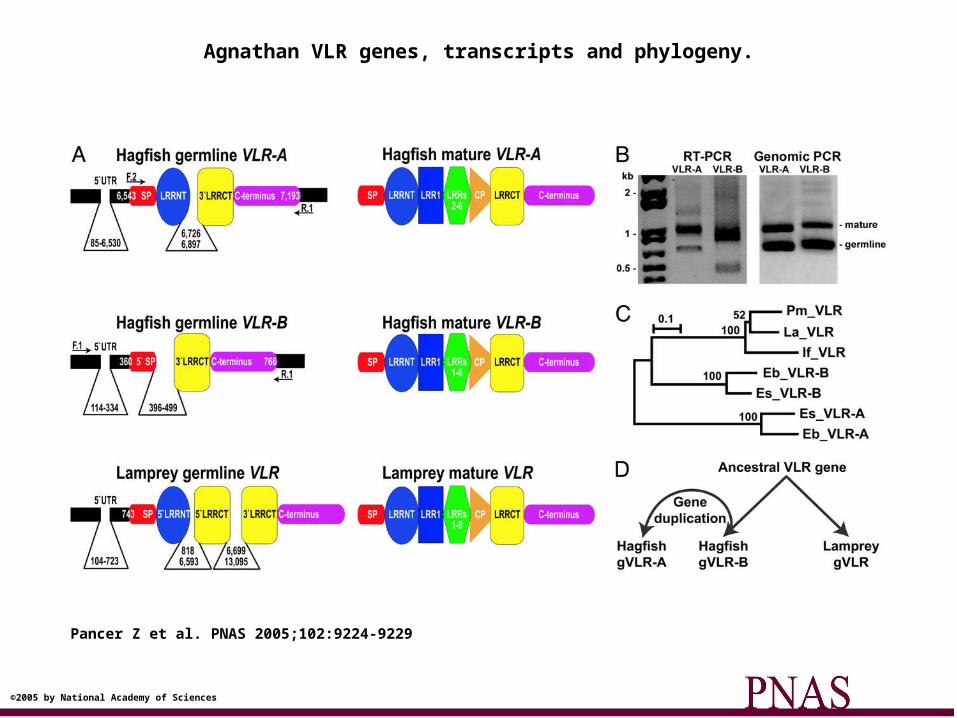
Agnathan VLR genes, transcripts and phylogeny.
Pancer Z et al. PNAS 2005;102:9224-9229
©2005 by National Academy of Sciences

Within 40 million years during the Cambrian period2 radically distinct adaptive immune systems (agnathans
and gnathostomes) evolved to generate a system for providing a set of diverse receptors.
Is this convergent evolution or ancestral evolution??i.e. do the VLRs represent the forerunners to the Ig and
TcR or did they evolve independently?

The molecules
• Innate immunity – Toll-like (TLR), CTL (C-type lectins) and NOD-like
(NLR) receptors– Invariant TcR (NK recognition) (Rapid)– Non-polymorphic MHC (i.e. CD1)– Non-classical MHC
• Adaptive immunity– T cell Receptor ( and )– Immunoglobulins– MHC – pathogen fragment presentation

Effective immune response
• Requires combination of both adaptive and innate response.
• Don’t get commitment without innate signals• Adjuvants promote formation of a protective
immune response to antigen. Merely molecules that trigger innate receptors e.g. killed mycobacterium.
• Whole new field has opened up using the various innate PAMPs as potential vaccine adjuvants.

Types of immune responses
• Complement – direct killing• Phagocytosis – complement opsonins• Defensins – secreted bactericidal compounds• Antibodies – block and direct receptor mediated
phagocytosis and complement killing.• T cells provide cellular immunity – kill virally
infected cells. Also essential in driving B cell and T cell maturation – cytokines

Central Cell• Dendritic cell or primary antigen presenting cell.
Found in many forms – at the very heart of the naïve immune response.

Classic cellular immunity
• Primary function is to respond to antigens presented by the APC.
• Antigens are fragments of intracellular or extracellular pathogens.
• Complex processing of antigens inside the APC and loading of MHC class I and class II in specific vesicles.
• Binding of peptides governed by motifs. Anchor residues in peptides that interact with floor and walls of the peptide groove.

m
HLA A2polymorphic

HLA DR1polymorphic
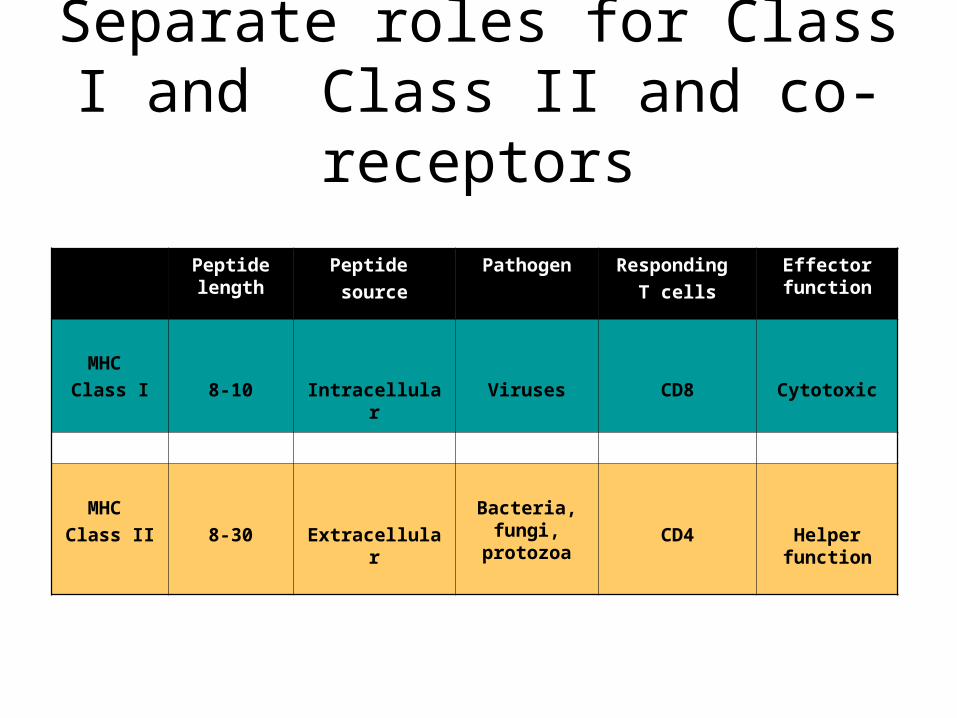
Separate roles for Class I and Class II and co-receptors
Peptide length
Peptide source
Pathogen Responding T cells
Effector function
MHC Class I 8-10 Intracellular Viruses CD8 Cytotoxic
MHC Class II 8-30 Extracellular
Bacteria, fungi,
protozoaCD4 Helper
function

CD4 CD8 binding sites
CD8SITE
CD4 site

Innate receptors
• Now a vast array of these discovered.• Absolutely critical in the immune response• LPS is an extremely powerful activator of B cells. First
discovered when mouse strain resistant to LPS. Found to have a defect in the CD14 molecule.
• Later evidence found presence of genes similar to drosophila Toll for pattern recognition in development.
• Found that these recognized various microbial componds.
• Extremely important to vaccine development and adjuvants.
• Appears they are also important in autoimmune disease.

Diversity of pattern recognition receptors

CD1 antigen presentation
• CD1 is a non-polymorphic MHC class I like molecule.
• Comes in 4 forms CD1a, b, c, d.
• Similar architecture to classical MHC class 1.
• Cleft is much narrower and deep with hydrophobic residues. Binds things other than peptides

CD1 ligands
-galactosyl ceramide (sponge)• Phophatidyl choline• Lipoarabinomannan (LAM)• Sulphatide• Many lipids (see review paper)• Mostly associated with mycobacteria• Some self antigens such as Ganglioside
GM1 – significance to autoimmunity.


CD1d structure with GALCerRecognized by iNK-T cells
Gal
Ceramide

CD1d structure with GALCerRecognized by iNK-T cells
SIDE TOP

iNK-T cell• Semi-invariant T cell Receptor• V24-J18 with V11 in humans• V14-J18 with V8.2, 4 and 2 in mice.• Restricted by CD1• Recognises -galactosyl ceramide.• Abundant in liver(~20%), BM (~3%)
spleen(~2%)• Massive release of IFN-, IL-4, TNF-, IL-13,
GMCSF, IL-2. Facilitates T cell and NK cell activation and expansion.

Innate immunity in plants flies and mammals is conserved – Toll/TLR

Specificity of TLR ligands
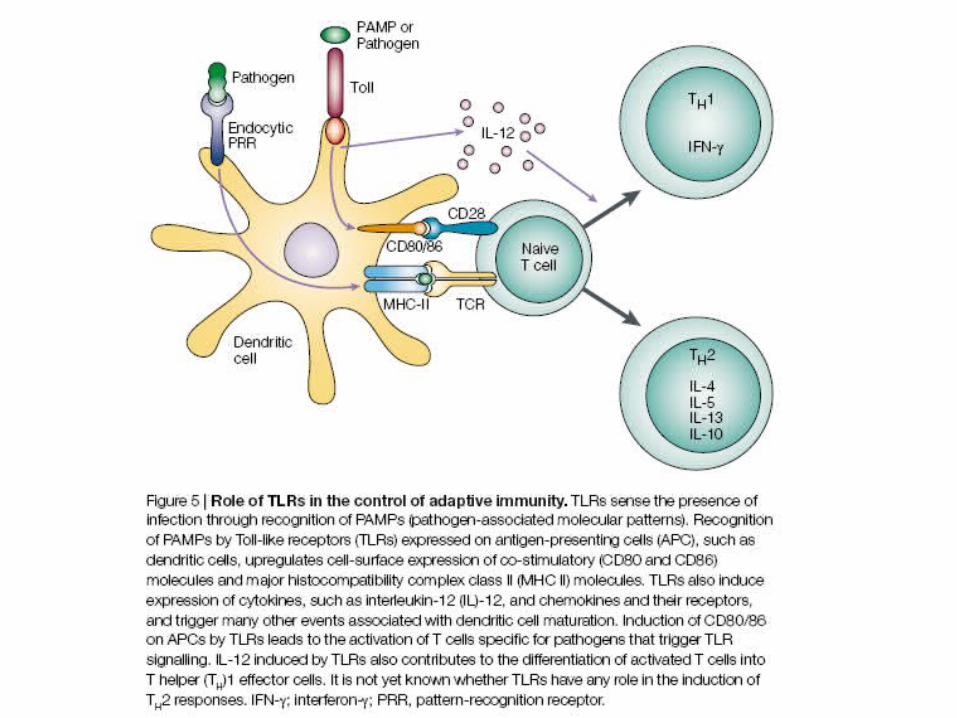

Basic structure of TLR
• Characterised by a LRR (leucine rich repeat external domain) and an intracellular TIR (Toll/IL-1 receptor) intracellular domain.
• LRR are a diverse set of proteins with consensus sequence in the domain
• L(X2)LXL (X2) NXL(X2)L(X7)L(X2)

Signalling of innate receptors
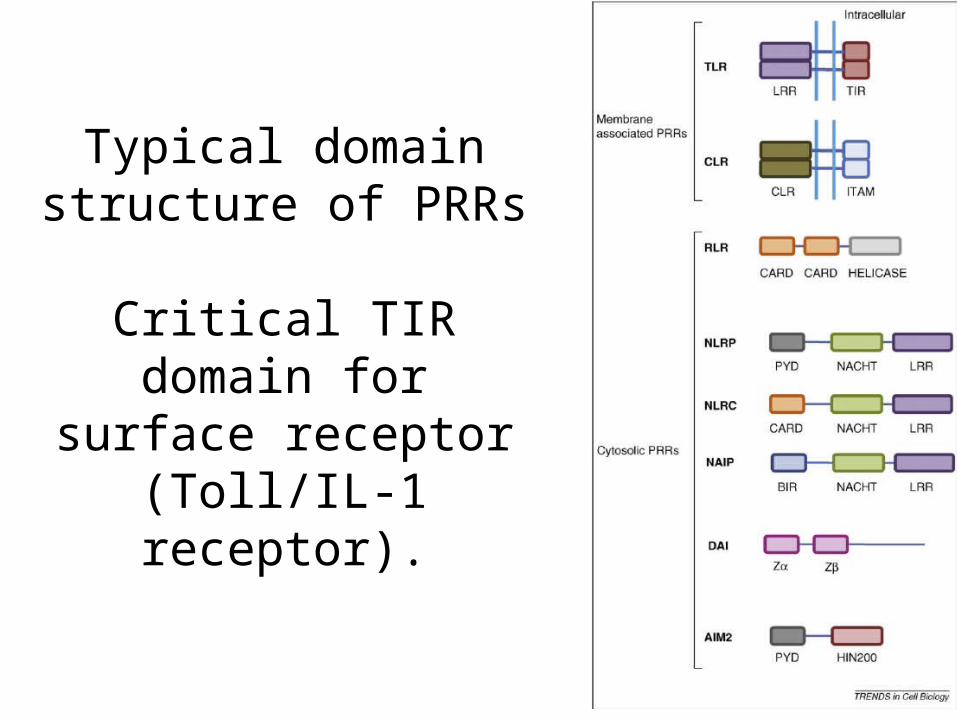
Typical domain structure of PRRs
Critical TIR domain for surface receptor
(Toll/IL-1 receptor).

Typical LRR structure
• Common horseshoe structure
• Concave surface formed by parallel -strands
• Convex surface formed by loops and 310 helices
• LRR are 20-30 amino acids long
• Binding region is typically the concave surface but not always

Ectodomain of TLR2 showing Leu rich repeat
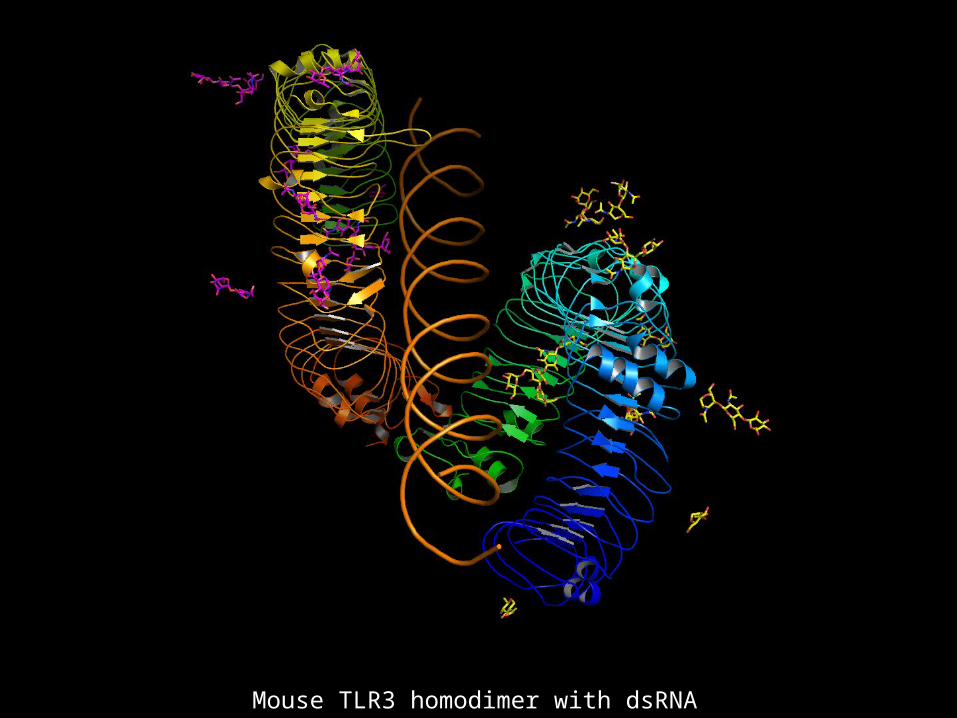
Mouse TLR3 homodimer with dsRNA

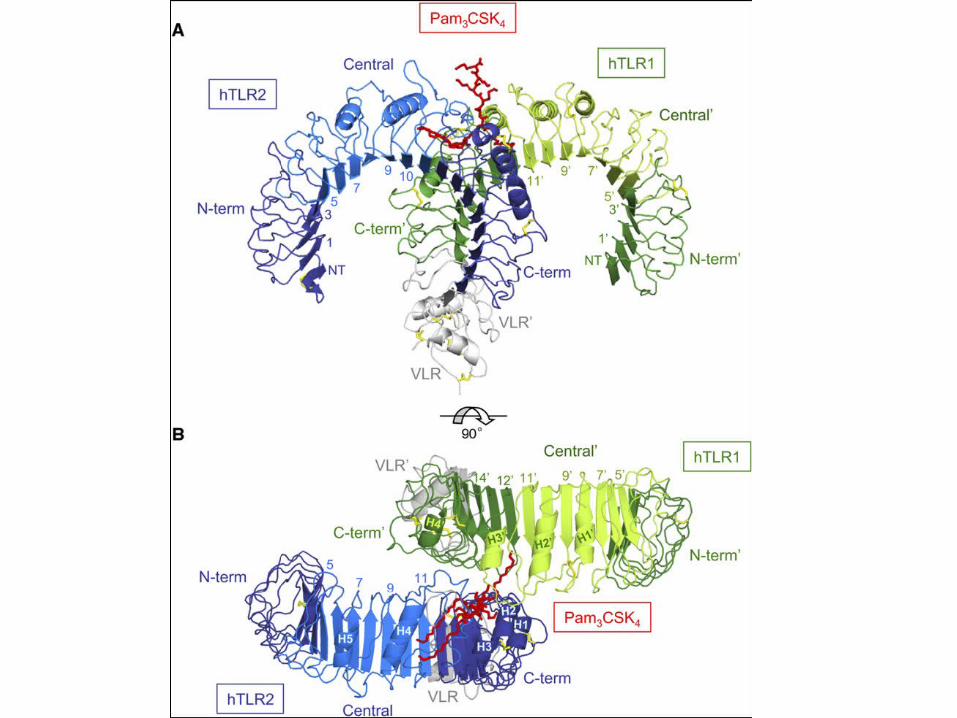
Pam3CSK4 is a potent PAMP
Activates through TLR1/TLR2
No activation
Triacylated lipopetide. 3 hydrophobic acyl chains and a charged head group
From: Jin et al, 2007 Crystal structure of the TLR1-TLR2 heterodimer induced by binding if a tracylated lipopeptide Cell 130 1071-1082

TLR2 with Pam3CSK4 bound
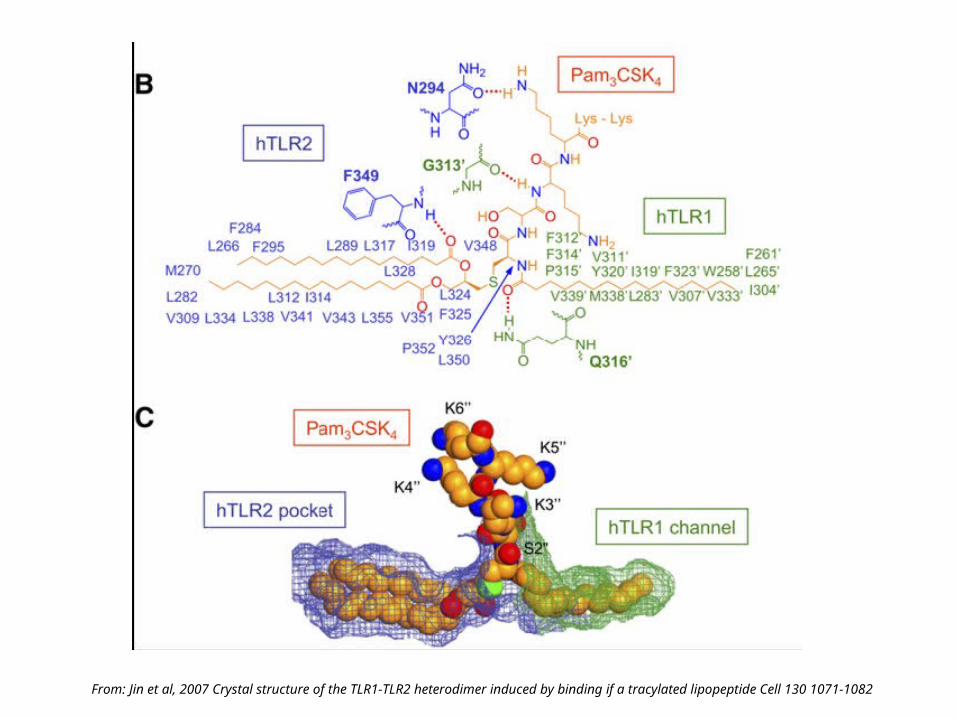
From: Jin et al, 2007 Crystal structure of the TLR1-TLR2 heterodimer induced by binding if a tracylated lipopeptide Cell 130 1071-1082
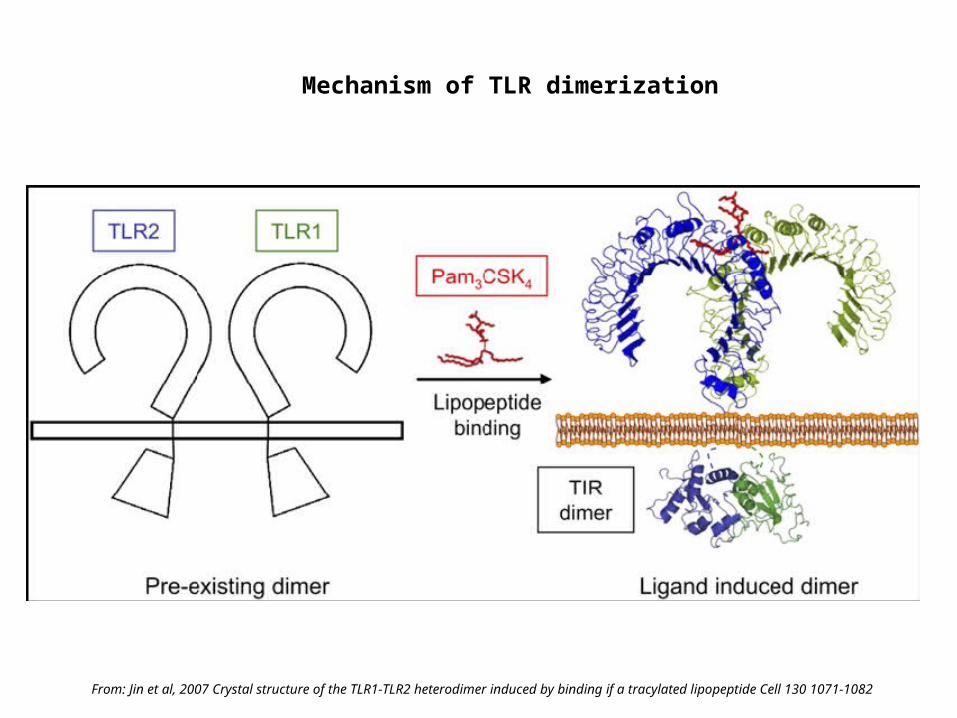
Mechanism of TLR dimerization
From: Jin et al, 2007 Crystal structure of the TLR1-TLR2 heterodimer induced by binding if a tracylated lipopeptide Cell 130 1071-1082

Lipopolysaccharide (LPS)Extremely potent Gram negative PAMP
This is the main immunogenic component
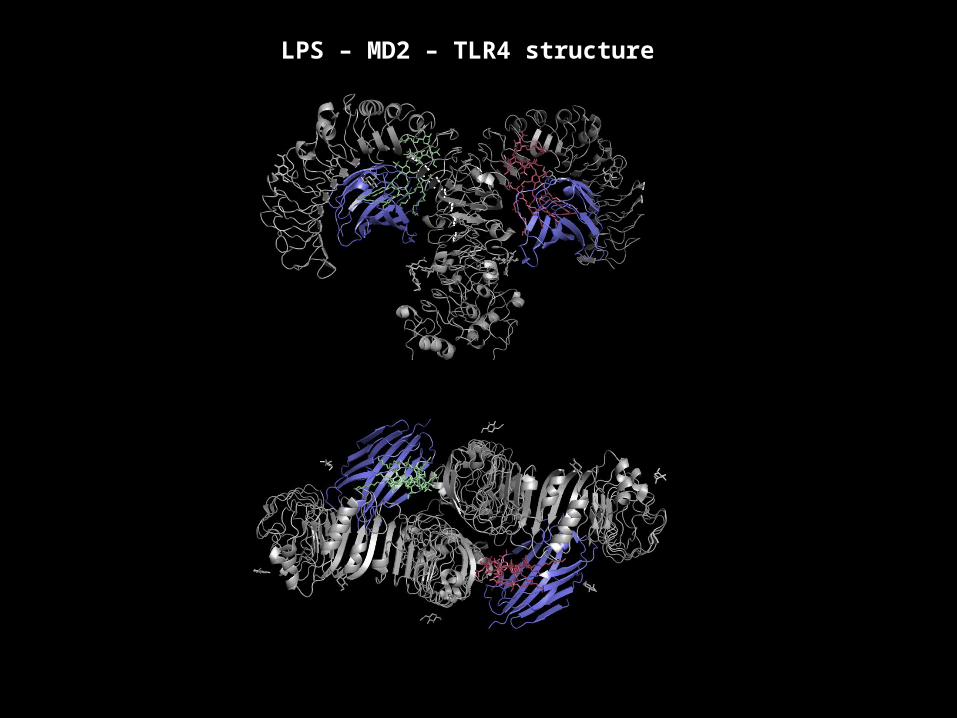
LPS – MD2 – TLR4 structure

Intracellular PRRs
A vast array of NLR specificities
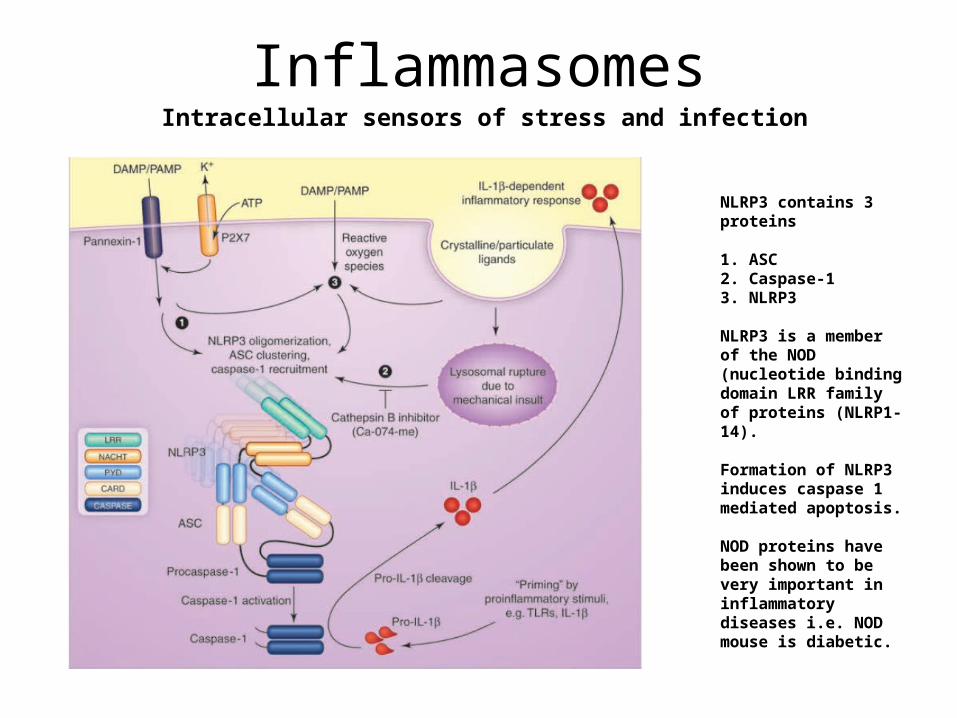
InflammasomesIntracellular sensors of stress and infection
NLRP3 contains 3 proteins
1. ASC2. Caspase-13. NLRP3
NLRP3 is a member of the NOD (nucleotide binding domain LRR family of proteins (NLRP1-14).
Formation of NLRP3 induces caspase 1 mediated apoptosis.
NOD proteins have been shown to be very important in inflammatory diseases i.e. NOD mouse is diabetic.

Known activators of NLRP3

Classical peptide recognition
• How does a T cell “see” such a small amount of peptide.• Each APC has ~105 MHC molecules on its surface.• Majority are filled with self peptides that do not elicit and
immune response.• Infection only likely to produce a small number of
relevant peptide-MHC complexes• These are likely to be spread across the surface, not
clustered.• How does the TcR “find” these.• How many peptide-MHC complexes.

Thymic +ve and –ve selection
• TcRs are selected on self MHC and self peptides in the thymus. 99% of all T cells never make it out of the thymus.
• Antagonist peptides positively select. Agonist peptides negatively select T cell.
• Same self peptides are required to keep T cells alive in the periphery.
• T cells normally tolerant to self peptides.

The immunological Synapse
• The interface between APC and T cell.• A lot going on over a small area in a short period of time.• Many molecules involved• Adhesion molecules (ICAM-1, LFA-1, B7.1/2 CD80,
CD86)• Antigen receptor• MHC• Co-receptors (CD4 & CD8)• Intracellular signalling molecules• Dynamic region that forms rapidly to concentrate events
into a tight junction between cells. • Clustering of intracellular kinases or phosphatases

Recent studies
• With more TcR/pepMHC systems studied, apparent that ½-life does not directly dictate agonist potential. In fact certain cases, there is no correlation. Antagonists have longer ½ lifes than their agonists in some cases.
• Now apparent that there is some steric change in TcR.• Latest is a two step twist-cap model of engagement• 1. Weak binding of TcR to MHC• 2. Change to fit peptide by TcR to strengthen the
binding.

Involvement of self-peptides in T cell Recognition
• Surprising recent finding is that agonist peptide is enhanced by the inclusion of endogenous self peptide MHC complexes.
• Model is now a pseudo-dimer where activation involves dimerization of agonist peptide MHC and self peptide MHC together.
• Can help to explain the role of self peptides in thymic learning and requirement in periphery to keep T cells alive.

Pseudo dimer model

Summary
• Multiple forms of immune recognition.• Invariant MHC (CD1) present non-peptides such as
lipids to g/d T cells and special NK T cells.• Innate receptors – vast complex patterns• Classical T cell activation involves intricate interaction of
T cell with pepMHC and “understanding” minor differences between agonist (pathogen) vs self-peptide on which is was selected.
• New model is one where TcR sees combination of non-self and self peptide. Critical is the co-receptor CD4 and CD8.

















