induction of H2O2, resulting in a significant rise in cat-alase (50–66 %) and glutathione peroxidase (175 %) activities, which were helpful in defending digestive tract cells against oxidative injury. These results help in understanding the anticancer mechanism of 4-MU based on its antioxidation in organisms.
Umbelliferones, which are widely found in plants, have gained significant attention due to their superior biological roles in many animal species and underlying tumor-inhib-itory effects (Morabito et al. 2010). In recent years, there
Abstract Umbelliferones have gained significant attention due to their tumor-inhibitory effects in vitro. This study was undertaken to examine the impact of umbelliferones in an invertebrate model organism, Bom-byx mori, to assess the underlying antimicrobial activi-ties via antioxidation in vivo. Oral administration of 4 mM 4-methylumbelliferone (4-MU), a model umbel-liferone drug, in B. Mori larvae caused a rapid increase in reactive oxygen species, such as hydrogen peroxide (H2O2) and antimicrobial activity in the digestive tract. In addition, a significant increase in total antioxidant capacity as well as superoxide anion radical-inhibiting activity and reduced glutathione were detected. The antioxidant defense system was activated following
Communicated by I. D. Hume.
Y. Fang and H. Wang are co-first authors, who contributed equally to this work.
Electronic supplementary material The online version of this article (doi:10.1007/s00360-014-0840-1) contains supplementary material, which is available to authorized users.
Y. Fang · H. Wang · W. Zhu · l. Wang · H. liu · X. Xu · W. Yin · Y. Sima (*) · S. Xu (*) Department of applied Biology, School of Biology and Basic Medical Sciences, Medical College, Soochow University, Suzhou 215123, Chinae-mail: [email protected]
Y. Fang · H. Wang · W. Zhu · l. Wang · H. liu · X. Xu · W. Yin · Y. Sima · S. Xu national engineering laboratory for Modern Silk, Institute of agricultural Biotechnology and ecology, Soochow University, Suzhou 215123, China
has been intense research on the tumor-inhibitory activities of umbelliferones using the model drug, 4-methylumbel-liferone (4-MU), in many in vivo and in vitro experiments with mammalian cells (Urakawa et al. 2012; Twarock et al. 2011; Oemardien et al. 2011; Bhattacharyya et al. 2009; nakazawa et al. 2006). To date, the mechanism of how 4-MU inhibits the growth of cancer cells as well as the occurrence and development of tumors is unknown. It is likely that the conjugated double bonds between the oxy-gen atoms in umbelliferones trigger the formation of free radicals in organisms (li et al. 2005), however, there are few comprehensive in vivo behavior and toxicity studies in animals.
For drug toxicity tests, the experimental animals used are typically mammals (such as mice). The orally admin-istered drug remains in the stomach for a relatively long period and is easily digested by gastric fluid, making observations on the influence of the drug on the diges-tive tract difficult. In this study, Bombyx mori (B. mori), an invertebrate, was used as the experimental animal. The advantage of this animal as a model is that the oral admin-istration of 4-MU results in the direct contact of 4-MU with the inner wall of the digestive tract. Moreover, 4-MU enters the blood circulation quickly in B. mori (as in mice) after oral exposure. 4-MU is detoxified in the fat body (analogous to the mouse liver) and discharged via the Mal-pighian tubules (analogous to the mouse kidney) (Hama-moto et al. 2009). Bombyx mori is suitable for the study of drug metabolic pathways in nutrient-absorbing tissues (Hamamoto et al. 2004).
Previous studies have shown that 4-MU has two modes of metabolic modification in mammals: glyco-sylation and sulfation. In B. mori, glycosylation is the dominant mode (Hamamoto et al. 2009), due to the strong activities of glycosylation enzymes in the diges-tive tract and blood (Xu et al. 2013). In our study, larvae from B. mori were orally exposed to 4-MU to investigate changes in the ability of the digestive tract to resist oxi-dative stress. Data on the effect of 4-MU on the exposed tissues in B. mori were obtained via the oxidation–reduction reaction.
Many anti-tumor drugs cause damage to normal cells (ryberg et al. 2008), but few studies have examined the toxicity of 4-MU as an anti-tumor drug. We sought to examine whether 4-MU injures normal cells and, specifi-cally, whether it has adverse impacts on the digestive tract when orally administered. We used B. mori as a model invertebrate organism to reveal the mechanism of 4-MU and its role in tissue protection through promoting the anti-oxidant ability of cells in the digestive tract. This work pro-vides a foundation for analyzing the effects of umbellifer-ones on the digestive tract.
Materials and methods
Preparation of animals
The Dazao strain of B. mori was reared in our laboratory. larvae were reared at 25 °C with a photoperiod of 12 h light and 12 h dark. Similar-sized larvae (body weight 1.0 ± 0.1 g) were orally exposed to 4-MU (Sigma, St. louis, MO, USa) on Day 3 of the 5th instar. 4-MU was dissolved in 10 % Dimethyl Sulfoxide (DMSO) to a final concentration of 4 mM. The instillation dose was 50 μl per individual. To minimize food residue, starvation was imposed for 3 h before exposure to 4-MU; the control group received the same starvation. Subsequently, digestive tracts were carefully isolated from the exposed and control larvae 8–120 min after injection and prepared for testing enzymatic activities and other indices. after removing the peritrophic membrane and bits of food residue, the pre-pared digestive tracts of both groups were approximately 0.1 g in both exposed and control groups. The homogen-ate was ground on ice. a diagram of the study timeline is shown in Figure S1. High-performance liquid chroma-tography analysis indicated the presence of 4-MU in the digestive tract 8 min after the 50 μl oral injection of 4 mM 4-MU.
Superoxide anion and reactive oxygen species (rOS) staining
The levels of superoxide anion and rOS were measured using a dihydroethidium kit (Beyotime, Shanghai, China) and rOS kit (Beyotime, Shanghai, China), respectively. The digestive tract was collected in diethylpyrocarbonate (DePC) (containing 0.7 % naCl) to avoid air exposure and was then vortexed for 20 s in normal saline which included 0.1 % DMSO. after that, the digestive tracts were quickly placed into a staining solution for 7 min (for dihydroethid-ium) or 15 min (for 2′,7′-dichlorofluorescin diacetate), and then washed for 5 min in the dark room with saline. The red fluorescence (for superoxide anions) or green fluores-cence (for rOS) was observed with a fluorescence micro-scope (Olympus Medical Systems Corp., Tokyo, Japan).
Hematoxylin and eosin (He) staining
During hours 2 after exposure to 4-MU, the digestive tracts of larvae were isolated on ice and fixed in 4 % paraformal-dehyde. after serial dehydration in ethanol at 70, 80, 90, 95 and 100 %, for 15 min each, the samples were treated twice with xylene, for 15 min, embedded in paraffin blocks, and sectioned at 5–10 μm thickness. For He staining, the sec-tions were dewaxed using xylene and rehydrated in an
701J Comp Physiol B (2014) 184:699–708
1 3
ethanol series. all sections were visualized using an Olym-pus BX51Tr U-UCD8a microscope (Olympus, Tokyo, Japan).
Biochemical assays
The activities of catalase (CaT) and superoxide dismutase (SOD) and the level of hydrogen peroxide (H2O2) were measured using their respective assay kits (Technology Co., ltd. nanjing Jiancheng, China). The activities of glu-tathione reductase (gr) and glutathione peroxidase (gPx) were measured using the gr assay Kit and the Total gPx-activity kit (Beyotime, Shanghai, China), respectively. The activity of γ-glutamate–cysteine ligase (gCl) and nicoti-namide adenine dinucleotide (naDH) oxidase, the level of gSH and naD(H) were measured using their respective assay kits (Comin, Suzhou, China) according to the manu-facturer’s instructions.
Determination of superoxide anion radical-inhibiting activity
The superoxide anion radical-inhibiting activity was meas-ured using an assay kit (Technology Co., ltd. nanjing Jiancheng, China) according to the method of Wang et al. (2012). an activity unit was defined as follows: when the sample per liter reacting at 37 °C for 40 min inhibits super-oxide anion radicals, this is equivalent to the variation value of 1 mg of vitamin C inhibiting the superoxide anion radical.
The antibacterial experiment in vivo
The silkworm larvae on Day 3 of the 5th instar were starved for 3 h, then orally exposed to 4-MU (4 mM, 50 μl), fol-lowing an oral exposure to Escherichia coli (cfu = 5×107, 50 μl) at 8-min intervals. The strain of E. coli used was maintained at our laboratory. a direct current of 16 V was used 8 min later to cause the larvae to disgorge its digestive juice, which was diluted with normal saline (0.7 %) 1,000 and 10,000 times before being cultured in agarose medium. For each culture dish (diameter, 10 cm), the culture amount was 200 µl. after culturing for 12 h at 37 °C, the colonies were counted.
analysis of gene expression profiles
Total rna was isolated from digestive tract of larvae using TrIzol reagent (life Technologies, Carlsbad, Ca, USa) and purified using an Oligotex® mrna Midi kit (Qia-gen, Valencia, Ca, USa), according to the manufacturer’s instructions. The real-time reverse transcription poly-merase chain reaction (qrT-PCr) was used to investigate
the expression level of genes in MaPK pathway with the primers listed in supplementary Table S1. an SYBr® Pre-mix ex Taq™ kit (Perfect real Time) was used for qrT-PCr and Rp49 gene of B. mori(nCBI accession number: nM_001126254) was set as the reference gene. The reac-tion program was: 95 °C degeneration for 1 min; followed by 40 cycles of 95 °C for 5 s, 58 °C for 10 s, and 72 °C for 10 s. The process was set automatically by the detection program and three replicates were used for each.
replication of experiments and statistical analyses
all experiments were performed at least three times and the significance of differences was determined using Student’s t test.
Results and analysis
4-MU induces the antimicrobial activities of the animal digestive tract increased
Umbelliferones reportedly show strong antibacterial effects on plant pathogenic bacteria in vitro (Zhang and Jiang 2010). In this research, silkworm larvae were orally exposed to 4-MU (4 mM, 50 μl), following an oral expo-sure to E. coli (cfu = 5×107, 50 μl) at 8-min intervals, and then take an antibacterial experiment in vivo. The results showed that the count of E. coli had remarkably reduced in the intestinal juice of 4-MU exposure group (Fig. 1) and indicated that 4-MU would induce the antimicrobial activi-ties of the animal digestive tract increased significantly.
4-MU induces an oxidative stress response but without damage to the digestive tract
To determine the mechanism underlying the increased anti-bacterial ability following exposure to 4-MU, the levels of superoxide anions stained by dihydroethidium (Fig. 2a) and rOS stained by 2′,7′-dichlorofluorescin diacetate (Fig. 2B) in the digestive tract were measured, as umbel-liferones are likely to trigger the formation of free radicals in organisms (li et al. 2005). To examine the contribution of the superoxide anion, a form of rOS (Bae et al. 2011), to oxidative stress, we measured the level of the anion. The results showed that the amount of both rOS and super-oxide anions increased after exposure to 4-MU, though the increase rate and duration differed (Fig. 2). at 8 min, the rOS and superoxide anions were significantly higher compared with the positive control group (10 % DMSO exposure). The increase in rOS continued until 60 min (Fig. 2B). after 32 min, the increase in superoxide anions stopped (Fig. 2a).
702 J Comp Physiol B (2014) 184:699–708
1 3
The results in Figs. 1, 2 show that antimicrobial activ-ity in the digestive tract increased as the level of rOS rose following 4-MU exposure. However, whether the rOS
caused damage to the tissue is unknown. Previous research has shown that rOS can oxidize intracellular nucleic acids and proteins, damage the cell membrane and accelerate the
Fig. 1 4-MU exposure increases the antibacterial ability of the digestive tract. larvae were starved for 3 h, then orally exposed to 4-MU (4 mM, 50 μl), following oral exposure to Escherichia coli (cfu = 5×107, 50 μl) at 8-min intervals. a direct current of 16 V was used 8 min later which caused the larvae to disgorge their digestive juice. This digestive juice was diluted with normal saline (0.7 %) 1,000 (A) and 10,000 times (B) before being cultured in agarose medium. (a) Colony count on the plate; (b) plate culture of intestinal juice after oral injection of 10 % DMSO; (c) plate culture of intestinal juice after oral injection of 4-MU. *P < 0.01 indicates a significant difference between the 4-MU treatment group and the positive control group (n = 3 for each group)
Fig. 2 exposure to 4-MU induced the level increase of rOS and superoxide anion in digestive tract. A Superoxide anion staining. B rOS staining. (a–c) represent 8, 32, and 60 min after oral injection of 10 % DMSO, respectively; (e–f) represent 8, 32, and 60 min after oral injection of 4-MU + 10 % DMSO. The scale is 100 μm
703J Comp Physiol B (2014) 184:699–708
1 3
aging and decomposition of cells (Hermenean et al. 2013), and cause damage to cellular structures and tissue function (Bae et al. 2011; Martindale and Holbrook 2002).
accompanied the rOS increased after 4-MU exposure during 8–120 min (Fig. 2), however, observations of the digestive tract at 2 h after 4-MU exposure by He staining indicated no obvious cellular damage in the digestive tract (Fig. 3a). Moreover, DaPI nuclear staining revealed no increase in autophagy or accelerated apoptosis (Fig. 3B). Similar results were observed in the digestive tract at 12 h after 4-MU exposure. Further investigation of the mrna level of some key genes in the mitogen-activated protein kinase (MaPK) pathway showed that p38(aB208585) was up-regulated after exposure to 4-MU. However, extracel-lular signal-related kinase (Erk)(DDBJ, eMBl, and gen-bank accession number, aB183869) and MaPK–erK
kinase(Mek)(aB183870) also increased, to some extent, instead of down-regulation which would be seen in apop-totic cells (Fig. 3C). These results indicated that tissue cells were not damaged by the rapid rise in rOS level in the digestive tract following 4-MU exposure.
exposure to 4-MU increases the antioxidant ability of the digestive tract in B. mori
given that there was no damage or apoptotic cells in the digestive tract following exposure to 4-MU, we deduced that 4-MU triggered rapid rOS production in B. mori and improved rOS decomposition in the digestive tract. The antioxidative ability of organisms is controlled by both enzymatic reactions and non-enzymatic reactions. The total antioxidant capacity (T-aOC) and the inhibition superoxide
Fig. 3 4-MU exposure does not cause structural damage to the diges-tive tract. The digestive tract of larvae were isolated at 2 h after oral injection 4-MU. A He results. B DaPI results. The red and yellow arrows represent the vacuole and the nuclei, respectively. The scale is 50 μm. C real-time PCr analysis of the mrna levels of Mek,
erk, p38 in digestive tract after exposure to 4-MU. *P < 0.05 and **P < 0.01 indicate significant differences between the 4-MU treat-ment group and the positive control group (n = 5 larvae, repeat observation 3 times) (color figure online)
704 J Comp Physiol B (2014) 184:699–708
1 3
anion radical capacity represent the sum of these antioxi-dative reactions. To measure the influence of 4-MU on the oxidation reaction in the digestive tract, first the T-aOC was measured. The results showed that T-aOC significantly increased 8–60 min after exposure to 4-MU (Fig. 4a). at 32 and 120 min after drug administration, inhibition superox-ide anion radical capacity significantly increased (Fig. 4b). The gSH/gSSg ratio at 8, 48, and 60 min significantly increased after administration with 4-MU compared with the positive control (Fig. 4c). These results demonstrate that the antioxidant ability of the digestive tract in B. mori was greatly enhanced after 4-MU exposure.
H2O2 is produced after 4-MU exposure in the digestive tract
The results (Fig. 4b) showed that the superoxide anion rad-ical-inhibiting activity was not significantly up-regulated like T-aOC (Fig. 4a).Therefore, the increase in superox-ide anions induced by 4-MU was only a minor contribu-tor to the increase of rOS. We infer that 4-MU exposure probably induced the increase of H2O2, another main form of rOS in organisms (Circu and aw 2010). The results showed that 8–120 min after 4-MU exposure, the H2O2 levels in the digestive tract were consistently significantly higher than that of the positive control (Fig. 5a). Thus, the increase in H2O2 proved to be the primary mechanism behind the increase of rOS.
Besides the increase of H2O2 in the digestive tract after 4-MU exposure, the activity of CaT, an enzyme that degrades H2O2, also significantly increased (Fig. 5b). In contrast, the activity of SOD, the key enzyme in the formation of H2O2,
was significantly decreased at 16 and 60 min (Fig. 5c). Simi-larly, a rapid change in SOD activity was also shown in mice exposed to alcohol at 30 min, which indicated that the stress oxidant response in animals is very fast (Chen et al. 1998). The decreased activity of SOD might be due to a feedback inhibition process related to H2O2, a phenomenon that has already been reported (liu et al. 2012). For this reason, we believe that the generation of large quantities of H2O2 after 4-MU exposure is not entirely due to catalysis by SOD.
To track other sources of H2O2 after 4-MU exposure, the metabolic level of the respiratory electron transfer chain was detected. We examined naDH oxidase (nOX-1), another key enzyme in the formation of H2O2. nOX-1 activities were similar to the control group at all time points, except at 60 min when nOX-1 activity was lower than the posi-tive control (Fig. 5d). The naD+ content, an indicator of catalytic production from nOX-1, was always higher than the positive control (Fig. 5d) as the H2O2 level continuously increased. However, the naDH (substrate) content signifi-cantly decreased (Fig. 5e). This further proved that a large amount of H2O2 induced by 4-MU was generated through catalysis by nOX-1. except at 48 min, the naDH/naD+ ratio (Fig. 5g) showed a significant decreasing trend. Thus, the respiratory electron transfer chain was the major source of H2O2 in the digestive tract after 4-MU exposure.
exposure to 4-MU elevates gSH levels in the digestive tract
The above results indicated that 4-MU exposure induces a large amount of H2O2 in the digestive tract of B. mori. The metabolic levels of enzymes related to H2O2 scavenging
Fig. 4 Change of the antioxidant levels in digestive tract after oral injection of 4-MU. a at 37 °C, one unit of total antioxidant capacity (T-aOC) was defined as a 0.01 increase in OD value per milligram of tissue protein per minute; b Superoxide anion radical-inhibiting activ-ity. c Fold changes of the gSH/gSSg ratio between the 4-MU treat-
ment group and the positive control group. *P < 0.05 and **P < 0.01 indicate significant differences between the 4-MU treatment group and the positive control group (n = 5 larvae, repeat observation 3 times)
705J Comp Physiol B (2014) 184:699–708
1 3
were also investigated (guttmann 2010). glutathione metabolism, a major pathway for H2O2 scavenging, con-sists of the γ-glutamine cycle (responsible for the synthesis and degradation of glutathione and the oxidation–reduction cycle [oxidation of reduced glutathione (gSH) and reduc-tion of oxidized glutathione (gSSg)] (guttmann 2010). gSH is involved in the scavenging of rOS, including H2O2. The generated product, gSSg, is finally reduced to gSH with naDPH (Jeon et al. 2012).
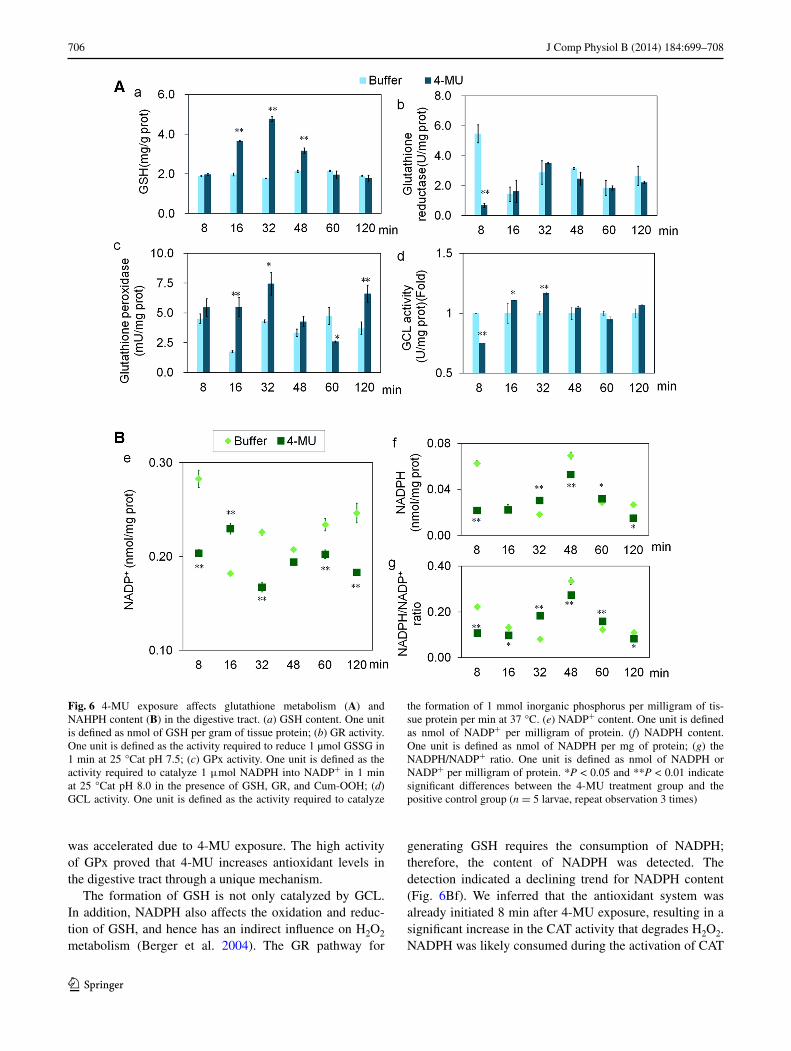
The gSH content was significantly higher at 16–48 min in the digestive tract (Fig. 6aa). However, gr, which
catalyzes the generation of gSH, did not demonstrate an increasing trend (Fig. 6ab). Meanwhile, gCl, an enzyme in another gSH synthetic pathway, showed a temporary decrease in activity 8 min after 4-MU exposure, but at 16–32 min, the activity significantly increased and then fell back to the level of the positive control (Fig. 6ad). The increased gSH was, therefore, unlikely to be due to the gr pathway and more likely to be due to the catalysis by gCl. In addition, the activity of gPx showed significant increases at 16, 32, and 120 min after exposure (Fig. 6ac); thus the oxidative metabolism of gSH in the digestive tract
Fig. 5 H2O2 is produced after 4-MU exposure. a H2O2 content. One unit is defined as mmol of H2O2 contained per gram of fresh tissue; b CaT activity. One unit is defined as 1 μmol of H2O2 degraded per gram of tissue protein during one second; c SOD activity. One unit is defined as the amount of SOD corresponding to a 50 % inhibition rate; d nOX-1 activity. One unit is defined as a 0.01 change in a600
per milligram of tissue protein per minute in the reaction system. e, f Content of naD+ and naDH, respectively. One unit was defined as nmol of naD+ or naDH per milligram of tissue; g the naDH/naD+ ratio. *P < 0.05 and **P < 0.01 indicate significant differ-ences between the 4-MU treatment group and the positive control group (n = 5 larvae, repeat observation 3 times)
706 J Comp Physiol B (2014) 184:699–708
1 3
was accelerated due to 4-MU exposure. The high activity of gPx proved that 4-MU increases antioxidant levels in the digestive tract through a unique mechanism.
The formation of gSH is not only catalyzed by gCl. In addition, naDPH also affects the oxidation and reduc-tion of gSH, and hence has an indirect influence on H2O2 metabolism (Berger et al. 2004). The gr pathway for
generating gSH requires the consumption of naDPH; therefore, the content of naDPH was detected. The detection indicated a declining trend for naDPH content (Fig. 6Bf). We inferred that the antioxidant system was already initiated 8 min after 4-MU exposure, resulting in a significant increase in the CaT activity that degrades H2O2. naDPH was likely consumed during the activation of CaT
Fig. 6 4-MU exposure affects glutathione metabolism (A) and naHPH content (B) in the digestive tract. (a) gSH content. One unit is defined as nmol of gSH per gram of tissue protein; (b) gr activity. One unit is defined as the activity required to reduce 1 µmol gSSg in 1 min at 25 °Cat pH 7.5; (c) gPx activity. One unit is defined as the activity required to catalyze 1 μmol naDPH into naDP+ in 1 min at 25 °Cat pH 8.0 in the presence of gSH, gr, and Cum-OOH; (d) gCl activity. One unit is defined as the activity required to catalyze
the formation of 1 mmol inorganic phosphorus per milligram of tis-sue protein per min at 37 °C. (e) naDP+ content. One unit is defined as nmol of naDP+ per milligram of protein. (f) naDPH content. One unit is defined as nmol of naDPH per mg of protein; (g) the naDPH/naDP+ ratio. One unit is defined as nmol of naDPH or naDP+ per milligram of protein. *P < 0.05 and **P < 0.01 indicate significant differences between the 4-MU treatment group and the positive control group (n = 5 larvae, repeat observation 3 times)
707J Comp Physiol B (2014) 184:699–708
1 3
and gPx. Thus, no massive accumulation of naDPH was detected at 8 min or within 2 h.
The above-mentioned gCl pathway (Fig. 6ad) is the major source of gSH after 4-MU exposure and gSH content increased at 16–32 min by enhancing gCl activity. The gr activity (related to the oxidative metabolism of naDPH) remained at the level of the control group within 120 min after 4-MU exposure. This implied that the gSH content in the tissue was also dependent on the oxidative metabolism of naDPH. The decline of the naDPH/naDP+ ratio in the beginning and later period (Fig. 6Bg) reflects the effect of 4-MU on the metabolism of naDPH. 4-MU is also poten-tially involved in the oxidation–reduction pathway of gSH by accelerating the oxidative metabolism of gSH.
Discussion
Our experimental results indicated that although there was an increase in the levels of superoxide anion radical-inhib-iting activity (Fig. 4b), T-aOC (Fig. 4a), and reduced gSH, the H2O2 content also significantly increased. Thus, 4-MU was not directly involved in improving the antioxidant level of digestive tract.
H2O2 was the major form of rOS induced after 4-MU exposure. It is also the most common type of rOS in aerobic organisms. as a second messenger, H2O2 is the key regula-tor for various reactions to oxidative stress. H2O2 is related to the metabolism of naD, naDP, and glutathione (Bae et al. 2011; Vurusaner et al. 2012; rhee 2006). The experi-mental results confirmed the close relationship between 4-MU-induced changes in H2O2 metabolism and the meta-bolic pathways for these three molecules (Figs. 5, 6).
The naDH/naD+ ratio is the key regulator in eMP-TCa and the respiratory electron transfer chain. The oxida-tion of naDH provides a main source of rOS (Houtkoo-per et al. 2010). We found that 4-MU exposure inhibits the respiratory electron transfer chain in the digestive tract of B. mori, and a significant increase in H2O2 resulted via the naDH–naD pathway (Fig. 5).
naDPH is involved in antioxidant reactions at various levels. Both the reactivation of the rOS-scavenger CaT and gSH generation (via gSSg) require naDPH. gSH, in turn, serves as a substrate for the reaction between gPx and thiore-doxin peroxidase for the scavenging of rOS. Drug, toxin, and hormone biotransformation are also enabled by the par-ticipation of naDPH + H+ in hydroxylation. In this experi-ment, naDPH decreased with increases in antioxidant levels in the digestive tract after 4-MU exposure. We attribute this to the initiation of the antioxidant defense system 8 min after 4-MU exposure. CaT activity, related to H2O2 degradation, significantly increased and naDPH was consumed for CaT activation. as the major source of gSH after 4-MU exposure, the gCl pathway induced an increase of gSH by enhanc-ing gCl activity at 16–32 min. The relatively constant gSH content before and after this period might be explained by the oxidative metabolism of naDPH. The gr activity, related to the catalysis of this reaction, remained at the level of the control group within 120 min; therefore, naDPH main-tained its effect as a reduction source. The detection of SOD, CaT, gCl, and gSH-Px activities (as part of the antioxidant defense system of the digestive tract) indicated a significant increase in free radicals, rOS scavenging, and H2O2 degra-dation after 4-MU exposure. The peroxidation chain reaction was effectively blocked, and the lipid peroxidation of tissue and cells was avoided (Sohal and Orr 2012).
Fig. 7 effect of 4-MU exposure on the antioxidant defense system in the digestive tract of Bombyx mori. 4-MU induced production of H2O2 and superoxide anion, and the main source of H2O2 was cata-lyzed by nOX-1. CaT and gPx removed H2O2. During the process of H2O2 removal, the main sources of gSH are generated by gCl
catalytic, naDPH also played a secondary role. The black bold lines indicate activated reactions and the black thick lines indicate slightly activated reactions. The red bold line indicates the restricted reactions (color figure online)
708 J Comp Physiol B (2014) 184:699–708
1 3
Figure 7 summarizes the effects of 4-MU exposure on the antioxidant defense system of the digestive tract in B. mori. 4-MU inhibits the respiratory electron transfer chain of the digestive tract and induces the generation of H2O2, which further activates the antioxidant defense system. Concurrently, 4-MU induces an increase in H2O2 degrada-tion and protects the digestive tract from rOS damage.
In conclusion, we found evidence that 4-MU increases antibacterial levels in the digestive tract of invertebrate animals in vivo and proposed a possible mechanism. This provides a further basis for studying the effects of umbel-liferones in the digestive tract.
Acknowledgments We wish to acknowledge associate Professor linchuan Zhao for his help and advice during our research. The pre-sent work was supported by the national High-Tech r&D Program of China (863 Program) (grant no. 2011aa100306), national natural Science Foundation of China (grant no. 31172264), Provincial Key Technology r&D Program of Jiangsu (Project no. Be2011327-1), and Priority academic Program Development of Jiangsu Higher edu-cation Institutions.
Conflict of interest The authors declare no conflict of interest.
References
Bae YS, Oh H, rhee Sg, Yoo YD (2011) regulation of reactive oxy-gen species generation in cell signaling. Mol Cell 32:491–509
Berger F, ramírez HMH, Ziegler M (2004) The new life of a cen-tenarian: signalling functions of naD (P). Trends Biochem Sci 29:111–118
Bhattacharyya SS, Paul S, Mandal SK, Banerjee a, Boujedaini n, Khuda Bar (2009) a synthetic coumarin (4-methyl-7 hydroxy-coumarin) has anti-cancer potentials against DMBa-induced skin cancer in mice. eur J Pharmacol 614:128–136
Chen JF, guo YH, Meng C, lu HD, lin Z, Cai YX (1998) Determi-nation of sod enzyme activity in the blood of rats suffering from alcoholism. J Fuzhou Univ 26:100–104 (In Chinese)
Circu Ml, aw TY (2010) reactive oxygen species, cellular redox systems, and apoptosis. Free radic Biol Med 48:749–762
guttmann rP (2010) redox regulation of cysteine-dependent enzymes. asian australas J anim Sci 88:1297–1306
Hamamoto H, Kurokawa K, Kaito C, Kamura K, razanajatovo IM, Kusuhara H, Santa T, Sekimizu K (2004) Quantitative evaluation of the therapeutic effects of antibiotics using silkworms infected with human pathogenic microorganisms. Int J antimicrob agents 48:774–779
Hamamoto H, Tonoike a, narushima K, Horie r, Sekimizu K (2009) Silkworm as a model animal to evaluate drug candidate toxicity and metabolism. Comp Biochem Physiol 149:334–339
Hermenean a, ardelean a, Stan M, Herman H, Mihali CV, Costache M, Dinischiotu a (2013) Protective effects of naringenin on carbon tetrachloride-induced acute nephrotoxicity in mouse kidney. Chem Biol Interact 13:00156–00157. doi:10.1016/j.cbi.2013.06.016
Houtkooper rH, Cantó C, Wanders rJ, auwerx J (2010) The secret life of naD+: an old metabolite controlling new metabolic sign-aling pathways. endocr rev 31:194–223
Jeon SM, Chandel nS, Hay n (2012) aMPK regulates naDPH homeostasis to promote tumour cell survival during energy stress. nature 485:661–665
li J, li n, Wu T, Wang C (2005) The coumarins substances chroma-tography detection progress. life Sci Instrum 5:34–38 (In Chinese)
liu Y, Shi QF, Ye YC, Tashiro S, Onodera S, Ikejima T (2012) activated O2
(•−) and H2O2 mediated cell survival in SU11274-treated non-small-cell lung cancer a549 cells via c-Met-PI3K-akt and c-Met-grb2/SOS-ras-p38 pathways. J Pharmacol Sci 119:150–159
Martindale Jl, Holbrook nJ (2002) Cellular response to oxidative stress: signaling for suicide and survival. J Cell Physiol 192:1–15
Morabito g, Trombetta D, Brajendra KS, ashok KP, Virinder SP, naccari C, Mancari F, Saija a, Cristani M, Firuzi O, Saso l (2010) antioxidant properties of 4-methylcoumarins in in vitro cell-free systems. Biochimie 92:1101–1107
nakazawa H, Yoshihara S, Kudo D, Morohashi H, Kakizaki I, Taka-gaki aK, Sasaki KM (2006) 4-methylumbelliferone, a hyalu-ronan synthase suppressor, enhances the anticancer activity of gemcitabine in human pancreatic cancer cells. Cancer Chemother Pharmacol 57:165–170
rhee Sg (2006) H2O2, a necessary evil for cell signaling. Science 312:1882–1883
ryberg M, nielsen D, Cortese g, nielsen g, Skovsgaard T, andersen PK (2008) new insight into epirubicin cardiac toxicity: compet-ing risks analysis of 1097 breast cancer patients. J natl Cancer Inst 15:1058–1067
Sohal rS, Orr WC (2012) The redox stress hypothesis of aging. Free radic Biol Med 52:539–555
Twarock S, Freudenberger T, Poscher e, Dai g, Jannasch K, Dullin C, alves F, Prenzel K, Knoefel WT, Stoecklein nH, Savani rC, Homey B, Fischer JW (2011) Inhibition of oesophageal squa-mous cell carcinoma progression by in vivo targeting of hyaluro-nan synthesis. Mol Cancer Ther 10:30
Urakawa H, nishida Y, Wasa J, arai e, Zhuo l, Kimata K, Kozawa e, Futamura n, Ishiguro n (2012) Inhibition of hyaluronan syn-thesis in breast cancer cells by 4-methylumbelliferone suppresses tumorigenicity in vitro and metastatic lesions of bone in vivo. Int J Cancer 130:454–466
Vurusaner B, Poli g, Basaga H (2012) Tumor suppressor genes and rOS: complex networks of interactions. Free radic Biol Med 52:7–18
Wang Yl, li K, Zhang J, He JH, li ZJ (2012) Optimization of pro-cessing conditions by using protease to hydrolyze bone powder to produce polypeptides and its antioxidative function. Int J Basic appl Sci 12:8–21
Xu X, Wang M, Wang Y, Sima Y, Zhang D, li J, Yin W, Xu S (2013) green cocoons in silkworm Bombyx mori resulting from the quercetin 5-O-glucosyltransferase of UgT86, is an evolved response to dietary toxins. Biol reprod 40:3631–3639
Zhang W, Jiang M (2010) Synthesis and antifungal activities of umbel-liferone analogs. Chin J Org Chem 30:254–259 (In Chinese)