Volume 159 Number 4 11. Rosenfeld CR, Jackson GM. Estrogen-induced refracto- riness to the pressor effects of infused angiotensin I I. AM j 0BSTET GYNECOL 1984;148:429. 12. Naden RP, Rosenfeld CR. Systemic and uterine re- sponsiveness to angiotensin II and norepinephrine in estrogen-treated nonpregnant sheep. AM J 0BSTET GY- NECOL 1985;153:417. 13. Chesley LC, Tepper IH. Effects of progesterone andes- trogen on the sensitivity to angiotensin II. .J Clin Endo- crinol 1967;27:576. 14. Everett RB, Worley RJ, MacDonald PC, Gant NF. Modi- fication of vascular responsiveness to angiotensin II Hormone treatments and vascular reactivity in pregnant women by intravenously infused 5-alpha- dihydroprogesterone. AM .J 0BSTET GYNECOL 1978; 131: 352. 15. Everett RB, Worley RJ, MacDonald PC, Gant NF. Effect of prostaglandin synthetase inhibitors on pressor re- sponse to angiotensin II in human pregnancy . .J Clin En- docrinol Metab 1978;46: 1007. 16. Wilson ESB. A prospective controlled study of the effect on blood pressure of contraceptive preparations contain- ing different types and dosage of progestogen. Br J Obstet Gynaecol 1984;91: 1254. Arachidonic acid incorporation into lipids of term human amnion David M. Olson, PhD,* and Zofia Smieja, BSc(Hons) London, Ontario, Canada There were no differences in the rate or amount of (1- 14 C)-Iabeled arachidonic acid incorporated into triacylglycerides, diacylglycerides, or any phospholipid species of freshly dispersed term human amnion cells obtained before or after labor. Both phosphatidylcholine and phosphatidylethanolamine incorporated 14 C-arachidonic acid in proportion to their molar percent of total amnion phospholipids, but phosphatidylinositol incorporated three times as much 14 C-arachidonic acid, suggesting either a rapid turnover in this specific phospholipid pool or a greater specificity for the transfer of arachidonoyl-coenzyme A to lysophosphatidylinositol. No or little competition of 14 C-arachidonic acid incorporation into triacylglycerides or phospholipids occurred with palmitic acid, linoleic acid, or 'Y-Iinolenic acid. However, dihomo-'Y-Iinolenic acid, eicosapentaenoic acid, and unlabeled arachidonic acid were effective inhibitors. We conclude that the term amnion has high acyl transferase activity, that no change in the basal activity of this enzyme occurs with the onset of labor, and that a specific acyl transferase exists for 20-carbon polyunsaturated fatty acids. (AM J 0BSTET GYNECOL 1988;159:995-1001.) Key words: Amnion, human term labor, arachidonic acid incorporation The control of arachidonic acid mobilization and sub- sequent prostaglandin production is an important reg- ulatory process, both for the maintenance of gestation and the initiation and progression of labor at term. I. 2 It is now established that prostaglandin production by amnion increases with the onset of labor in women 2 From the Departments of Paediatrics and Physiology, The University of Western Ontario, The Lawson Research Institute, St. Joseph's Health Centre of London. Supported by March of Dimes Birth Defects Foundation Basil O'Con- nor Starter Research Grant No. 5-530 and by an operating grant from the Medical Research Council of Canada. Received for publication November 16, 1987; revised March 3, 1988; accepted April20, 1988. Reprint requests: Dr. David M. Olson, The Lawson Research Insti- tute, St. Joseph's Health Centre of London, 268 Grosvenor St., London, Ontario N6A 4V2, Canada. *Research Scholar of the Medical Research Council. and is a process controlled at two separate levels: ara- chidonate mobilization and metabolism. The amnion contains the enzymatic apparatus to release covalently bound arachidonic acid from phospholipids, 1 and a sig- nificant amount of arachidonate is lost from phospha- tidylethanolamine and phosphatidylinositol with the onset of labor in women.1. 3 The amount of arachidonic acid in mammalian cells that is free compared with the amount that is esterified is very low,' however, sug- gesting that enzymes that esterify arachidonic acid are highly active. Arachidonic acid is incorporated into di- and triacyl- glycerides during de novo synthesis. However, it is pri- marily incorporated into phospholipids through acyl- ation of lysophosphoglycerides. 4 The first step in this incorporation is the formation of arachidonoylcoen- zyme A by acyl-coenzyme A synthetase (EC 6.2.1.3). This step has a corequirement for adenosine tri- 995
Transcript
Volume 159 Number 4
11. Rosenfeld CR, Jackson GM. Estrogen-induced refractoriness to the pressor effects of infused angiotensin I I. AM j 0BSTET GYNECOL 1984;148:429.
12. Naden RP, Rosenfeld CR. Systemic and uterine responsiveness to angiotensin II and norepinephrine in estrogen-treated nonpregnant sheep. AM J 0BSTET GYNECOL 1985;153:417.
13. Chesley LC, Tepper IH. Effects of progesterone andestrogen on the sensitivity to angiotensin II. .J Clin Endocrinol 1967;27:576.
14. Everett RB, Worley RJ, MacDonald PC, Gant NF. Modification of vascular responsiveness to angiotensin II
Hormone treatments and vascular reactivity
in pregnant women by intravenously infused 5-alphadihydroprogesterone. AM .J 0BSTET GYNECOL 1978; 131: 352.
15. Everett RB, Worley RJ, MacDonald PC, Gant NF. Effect of prostaglandin synthetase inhibitors on pressor response to angiotensin II in human pregnancy . .J Clin Endocrinol Metab 1978;46: 1007.
16. Wilson ESB. A prospective controlled study of the effect on blood pressure of contraceptive preparations containing different types and dosage of progestogen. Br J Obstet Gynaecol 1984;91: 1254.
Arachidonic acid incorporation into lipids of term
human amnion
David M. Olson, PhD,* and Zofia Smieja, BSc(Hons)
London, Ontario, Canada
There were no differences in the rate or amount of (1-14C)-Iabeled arachidonic acid incorporated into
triacylglycerides, diacylglycerides, or any phospholipid species of freshly dispersed term human amnion
cells obtained before or after labor. Both phosphatidylcholine and phosphatidylethanolamine incorporated 14C-arachidonic acid in proportion to their molar percent of total amnion phospholipids, but
phosphatidylinositol incorporated three times as much 14C-arachidonic acid, suggesting either a rapid
turnover in this specific phospholipid pool or a greater specificity for the transfer of arachidonoyl-coenzyme
A to lysophosphatidylinositol. No or little competition of 14C-arachidonic acid incorporation into
triacylglycerides or phospholipids occurred with palmitic acid, linoleic acid, or 'Y-Iinolenic acid. However,
dihomo-'Y-Iinolenic acid, eicosapentaenoic acid, and unlabeled arachidonic acid were effective inhibitors.
We conclude that the term amnion has high acyl transferase activity, that no change in the basal activity of
this enzyme occurs with the onset of labor, and that a specific acyl transferase exists for 20-carbon
Key words: Amnion, human term labor, arachidonic acid incorporation
The control of arachidonic acid mobilization and subsequent prostaglandin production is an important regulatory process, both for the maintenance of gestation and the initiation and progression of labor at term. I. 2
It is now established that prostaglandin production by amnion increases with the onset of labor in women2
From the Departments of Paediatrics and Physiology, The University of Western Ontario, The Lawson Research Institute, St. Joseph's Health Centre of London.
Supported by March of Dimes Birth Defects Foundation Basil O'Connor Starter Research Grant No. 5-530 and by an operating grant from the Medical Research Council of Canada.
Received for publication November 16, 1987; revised March 3, 1988; accepted April20, 1988.
Reprint requests: Dr. David M. Olson, The Lawson Research Institute, St. Joseph's Health Centre of London, 268 Grosvenor St., London, Ontario N6A 4V2, Canada.
*Research Scholar of the Medical Research Council.
and is a process controlled at two separate levels: arachidonate mobilization and metabolism. The amnion contains the enzymatic apparatus to release covalently bound arachidonic acid from phospholipids, 1 and a significant amount of arachidonate is lost from phosphatidylethanolamine and phosphatidylinositol with the onset of labor in women.1. 3 The amount of arachidonic acid in mammalian cells that is free compared with the amount that is esterified is very low,' however, suggesting that enzymes that esterify arachidonic acid are
highly active. Arachidonic acid is incorporated into di- and triacyl
glycerides during de novo synthesis. However, it is primarily incorporated into phospholipids through acylation of lysophosphoglycerides.4 The first step in this incorporation is the formation of arachidonoylcoenzyme A by acyl-coenzyme A synthetase (EC 6.2.1.3). This step has a corequirement for adenosine tri-
995
996 Olson and Smieja October 1988 Am .J Obstet Gynecol
A 14C-AA INCORPORATION Time- Course
B 14C-AA INCORPORATION c % PHOSPHOLIPID 14C-AA Time -Course Time- Course
<(
.:! <(
j~ u :!
~ "0 i -------.tc J!1 44 0 "()
~L_Jp, 0
"' 24 "' <f-
r~T "0 18 i5. '5 -"' 12 a_
"' 0 -"' 6 a. '0 b---6--o\PS
* 0
0 30 120 180 MINUTES
Fig. 1. Time course of "C-arachidonic acid incorporation into amnion lipids. Open symbols represent cesarean section (CS) cells and closed symbols represent spontaneous labor (SL) cells. PC, Phosphatidylcholine (squares), PI, phosphatidylinositol (triangles); PE, phosphatidylethanolamine (circles); PS, phosphatidylserine (diamonds). A, Mean ::!: SEM (n = 3) ''C-arachidonic acid incorporation into lipid subclasses. As the label is removed from the incubation medium, it is incorporated into triglycerides. phospholipids. and other lipids. There is no significant difference in incorporation between cesarean section and spontaneous labor cells. B, Mean ::!: SEM (n = 3) 11C-arachidonic acid incorporation into amnion phospholipids. There is no significant difference in incorporation between cesarean section and spontaneous labor cells for any phospholipid. C, Mean ::!: SEM (n = 5) "C-arachidonic acid incorporation into amnion phospholipids expressed as a percentage of the total arachidonic acid incorporated into phospholipids at each time point. The data are derived from B and are combined at each time point for cesarean section and spontanoues labor cells. *Significantlv different (p < 0.05) compared wth 30-minute value.
phosphate. Second, acyl-coenzyme A (1-acylglycero-3-phosphoglyceride 0-acyl transferase; EC 2.3.1.23) adds the arachidonoyl moiety to the lysophospholipid. Fatty acids can be released from acyl-coenzyme A by acylcoenzyme A hydrolases and from glycerophospholipids by phospholipases; hence, to maintain low intracellular concentrations of free arachidonic acid, acvl transferase activity must be greater than the combined activities of phospholipase and acyl-coenzyme A hydrolase.'
Free arachidonic acid metabolism can be directed by the activities of acyl-coenzyme A synthetase and acyl transferase. In rat liver microsomal preparations, addition of adenosine triphosphate shifted free arachidonate away from metabolism by cyclooxygenase toward incorporation into phospholipids.' Coincubation of adenosine triphosphate with coenzyme A and the rat microsomal preparation further promoted arachidonic acid incorporation into phospholipids. Other agents such as indomethacin, acetylsalicylic acid, diclofenac and calcium ionophore A23187, on the other hand, inhibited arachidonate esterification in macrophages, suggesting an action on acyl transferase activity.6 Together, these observations show that arachidonic acid esterification into cell lipids can be pharmacologically manipulated, and that lysophospholipid acylation is one mechanism whereby free arachidonate levels may be regulated in cells.
The purpose of this investigation was to study the incorporation of arachidonic acid into amnion cell lipids to determine ( 1) the proportional amount incorporated into the major lipid and phospholipid species, (2) whether the amount or rate of incorporation varied with the onset of labor, and (3) the relative specificity of the pathway for arachidonate incorporation.
Material and methods
Material. The following items were purchased from the Sigma Ch~mical Company, St. Louis, Mo.: Arachidonic acid, 99% free acid, shipped on dry ice; collagenase, type V; trypsin inhibitor (from soya beans), type li-S; bovine serum albumin, essentiallv fatty acid free, prepared from fraction V; and N- 2- hydroxyethyl-piperazine-N' -2-ethanesulfonic acid. The phospholipids, neutral lipids, and most fatty acids were purchased from Serdarv Research Laboratories, London, Ontario, Canada. One fatty acid, cis-5,8,11,14,17-eicosapentaenoic acid, was purchased from Aldrich Chemical Co., Milwaukee, Wis. Thin-layer chromatography plates used were preadsorbent Si250-PA(l9C) from]. T. Baker Chemical Co., Phillipsburg, N.J. 1-"C-Arachidonic acid (specific activity 59.6 mCi/mmol) in toluene was obtained from Amersham, Mississauga, Ontario, Canada; and minimum essential medium (Earle's salts) was obtained from
Volume 159 Number 4
Table I. ''C-Arachidonic acid incorporation into amnion lipids
Triacvlglycerides Diacylglvcerides Cholestervl esters Phosphatidvlcholine Phosphatidvlinositol Total phosphatidylethanolamine Phosphatidylserine Free arachidonic acid and metabolites
Data are the mean ± SEM; n = 5 (three cesarean section and two spontaneous labor amnions). Cells were washed three times with bovine serum albumin (0.5%) after 3 hours of incubation in minimal essential ·medium.
"C-Arachidonic acid = 0.1 11Ci/O.S ml (3.3 j.l.mol/L) in incubation media.
Flow Labs, Mississauga, Ontario, Canada. All solvents were A.C.S. graqe and all other chemicals were reagent grade.
Methods Tissues. Term human amnions obtained at elective
cesarean section before the onset of labor or those g<lthered after spontaneous vaginal deliverv were used. All infants were delivered at term and all of them were healthy.
Dispersed amnion cell5. Amnion was obtained bv manually separating amnion from chorion. The tissue was washed with Dulbecco's phosphate-buffered saline solution and cut into small pieces. Cells were dispersed with collagenase (0.1 o/c wt/ vol) plus trypsin inhibitor (0.05% wt/vol) in Dulbecco's phosphate-buffered sali~e solution. The first digestion was 10 minutes at 37° C with 25 ml enzvme solution to remove cells damaged from the separation and mincing. Whole pieces of tissue were picked out of the mixture and added to fresh enzyme solution (25 ml) and digested for 60 to 90 minutes until the tissue pieces were completelv dispersed. The cell suspension was filtered through four lavers of gauze, and the filtrate was centrifuged for 10 minutes at 200 X gat ambient temperature. The dispersed cells in the pellet were washed in minimum essential medium buffered with 25 mmol/L N-2-hvdroxvethylpiperazine-N' -2-ethanesulfonic acid at pH 7 .4. With this technique, both individual cells and small "sheets" of epithelial amnion cells are formed. Without trvpsin, a homogeneous preparation of single cells is difficult to achieve, due most probablv to the tight junctions found between amnion epithelial cells.' In these preparations, diffusion distances should have been minimal, allowing maximum exposure of the cells to the fatty acids. Viability as assessed by trvpan blue staining was
consistently >85%. Cells were counted with a hemacvtometer.
"'C Q) -c 6 a. ~
0 g <{ <{
Amnion arachidonic acid incorporation 997
30
ARACHIDONIC ACID .lJM
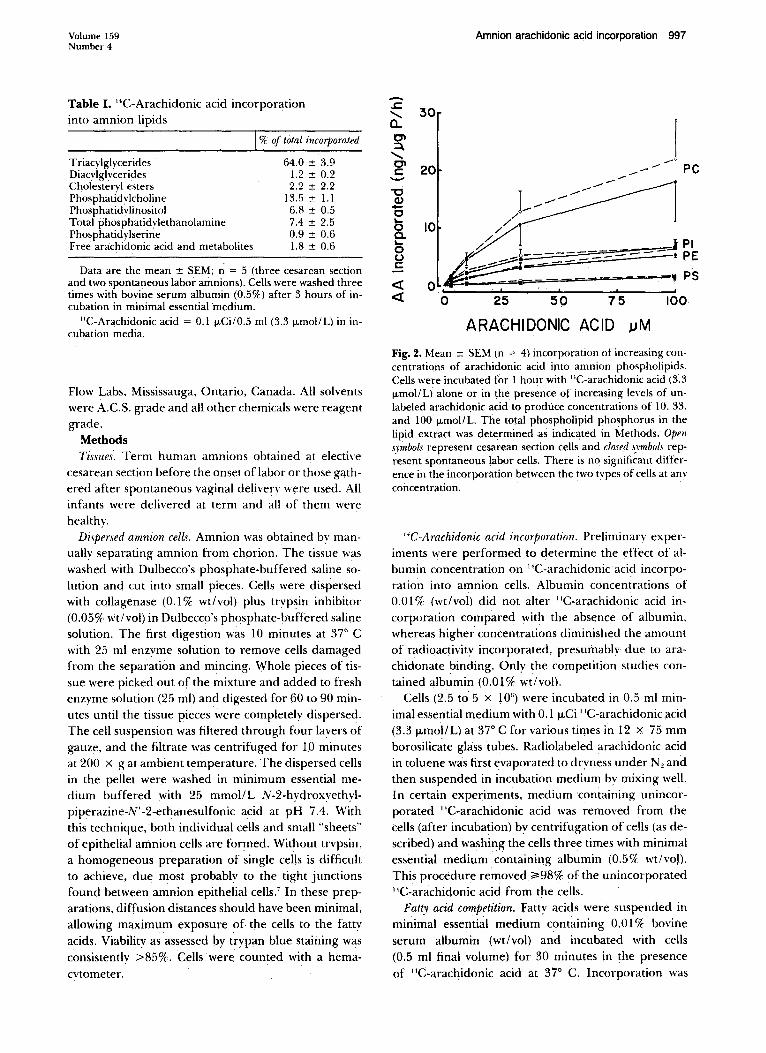
Fig. 2. Mean ± SEM (n = 4) incorporation of increasing concentrations of arachidonic acid into amnion phospholipids. Cells were incubated for l hour with "C-arachidonic acid (3.3 11mol/L) alone or in the presence of increasing levels of unlabeled arachidqnic acid to produce concentrations of I 0. 33. and 100 IJ.mol/L. The total phospholipid phosphorus in the lipid extract was determined as indicated in Methods. Open s_vmhols represent cesarean section cells and closed -~\·mbols represent spontaneous labor cells. There is no significant difference in the incorporation between the two tvpes of cells at anv concentration.
'·'C-Arachidonic acid incorporation. Preliminarv experiments were performed to determine the effect of albumin concentration on ''C-arachidonic acid incorporation into amnion cells. Albumin concentrations of 0.01% (wt/vol) did not alter "C-arachidonic acid incorporation compared with the absence of albumin, whereas higher concentrations diminished the amount of radioactivitv incorporated, presumablv due to arachidonate binding. Only the competition studies contained albumin (O.Ol% wt/vol).
Cells (2.5 to 5 x 10") were incubated in 0.5 ml minimal essential medium with 0.1 J.LCi ''C-arachidonic .acid (3.3 J.Lmol/L) at 37° C for various times in 12 X 75 mm borosilicate glass tubes. Radiolabeled arachidonic acid in toluene was first evaporated to drvness under N2 and then suspended in incubation medium bv mixing well. In certain experiments, medium containing unincor~ porated ''C-arachidonic acid was removed from the tells (after incubation) bv centrifugation of cells (as described) and washing the cells three times with minimal essential medium' containing albumin (0.5% wt/vo[).
This procedure removed. ;;;.98% of the unincorporated ''C-arachidonic acid from the cells.
Fall)' acid competition. Fatty acids were suspended in minimal essential medium containing 0.01% bovine
serum albumin (wt/vol) and incubated with cells (0.5 ml final volume) for 30 minutes in the presence of ''C-arachidonic acid at 37° C. Incorporation was
998 Olson and Smieja
Table II. Ratio of percent* of phospholipid-associated ''C-arachidonic acid determined at 3 hours of incubation to molar percentt of phospholipid in human amnion
Phosphatidylcholine Total phosphatidylethanolamine Phosphatidylinositol
Data are the mean ± SEM (n = .~).
Ratio
0.94 ± 0.15 0.72 ± 0.38 3.32 ± l.Ol
*Percent of phospholipid-associated "C-arachidonic acid is from Fif{. l , C.
+\1olar percent of phospholipid is from Okita et al.'
terminated by the addition of 3.0 ml of chloroforms: methanol (I : I) and extraction as described below.
Lipid rxlmction. The lipids were extracted by adding !tO ml of chloroform: methanol (I : I) to cells in 0 .. ? ml minimal essential medium. The phases were separated bv the addition of 0.5 ml I mol/L KC:l. The lower (organic) phase was removed, dried under N", and reconstituted in 200 I-ll chloroform: methanol (2: I)
for thin-layer chromatographic analysis or phospholipid phosphorus determination. This procedure extracted 9Wlr of ''C:-arachidonic acid found in washed cells.
Separation of lipids. To separate phospholipids. a portion (20 I-ll) of the reconstituted samples was spotted on Baker Si250-PA(l9C) preadsorbent plates. The plates were prewashed in methanol and activated at 160° C under vacuum for lO minutes before use. The plates were run in a one-dimensional system' with chloroform: methanol: petroleum ether: diethyl ether acetic acid: boric acid (40: 20:26:4: I 0: 1.8 gm) at 18° C
in paper-lined, preequilibrated tanks. The boric acid was dissolved first in acetic acid and methanol. Phospholipids were detected by I" vapor and cochromatography with authentic standards. The appropriate spots were scraped from the plates into scintillation counting vials, mixed with scintillation counting fluid, and counted by a scintillation spectrometer. This technique does not separate diacyl and plasmalogen phosphatidylethanolamine species. Therefore, total phosphatidylethanolamine is used to refer to both species.
The one-dimensional system of petroleum ether: diethyl ether: acetic acid (80: 20: 1) was used for determination of radioactivity associated with neutral lipids. The thin-layer chromatographic techniques for determination of the appropriate lipids were the same as those for phospholipid determinations.
Phospholipid phosphorus determination. The method of Goppelt and Resch" was used for the determination of total phospholipid phosphorus in extracted cell lipids, with the variation that the tubes were heated in an oven
October 1988 Am .J Obstet Gynecol
rather than an oil bath for 10 minutes. Potassium phosphate was used as the standard.
Statistical analysis. Data were analyzed by an appropriate analysis of variance. Significance was taken as a p value < 0.05. All values are expressed as the mean± SEM.
Results
The incorporation of "C-arachidonic acid into amnion lipids during a 3-hour incubation is given inTable I. The cells (from amnions associated with both cesarean section and spontaneous vaginal deliveries) were washed free of unincorporated ''C-arachidonic acid, extracted, and the lipids were separated by thin-layer chromatography in the two solvent systems described in Methods. Total radioactivity on the plates was determined and the percentage associated with each lipid species was calculated. Arachidonate was incorporated mostly into triacylglycerides, with about half as much incorporated into phospholipids. Very little was found in cholestervl esters. Less than 2% of arachidonate associated with the cells existed as a free acid or a metabolite.
The time course of ''C-arachidonic acid incorporation into the major lipid classes of the amnion is shown In Fig. l. In Fig. 1, A, the incorporation into triglycerides, phospholipids, and other lipids (diacylglycerols, cholestervl esters) is shown. Incorporation was the most rapid for the first 30 minutes of incubation, but continued to increase throughout the 3 hours of incubation. At 3 hours, a mean (±SEM) 65% ± 7.3% and 65o/c ± :~.7% of the "C-arachidonic acid in the incubation medium had been incorporated into all lipids in the cesarean section and spontaneous labor cells, respectively. By analysis of variance, no statistically significant differences in the percent of ''C-arachidonic acid incorporation into any lipid class at any time point were observed between cesarean section and spontaneous labor cells.
Fig. I, B, shows the percent of total incubated "Carachidonic acid incorporated into amnion phospholipids. These data indicate that most of the arachidonate incorporated rapidly into phosphatidylcholine during the first 30 minutes of incubation, whereas the amount associated with other phospholipids was not as great at 30 minutes, increasing more throughout the 3 hours of incubation. There was no statistically significant difference in ''C-arachidonic acid incorporation into any amnion phospholipid with the onset of labor.
Fig. I, C, represents the percentage of 11Carachidonic acid incorporated into total amnion phospholipids for each of four phospholipids at three different times of incubation, for combined cesarean section and spontaneous labor cells. The data indicate that the relative amount of ''C-arachidonic acid associated
Volume 159 Number 4
Amnion arachidonic acid incorporation 999
14c-AA Incorporation into Triglycerides
·-·ti:O A-A11:2 0-011:3 6-A20:3 0-020:4 -·20:5
QL---~----~--~----~--~ 0 25 50 75 100 125
Fatty Acid JI).A
Phosphatidylcholine Phosphotidylethanolomine c 120 .2 120 e 0 100 e- 100 0
~-:a 80 't>.l:;
80 ·- c
~· ~~ 60 60 c 0 ·-·11:0
~~ 40 40 ·-~11:2 6 0-011:3
"" A-A20:3 ~
" i=il:: 0 20 20 ~
I ~ 0 0
0 25 50 75 100 125 0 25 50 75 100 125
c Phosphatidylinositol Phosphatidylserine
i 120 120
0 100 100 e-0
~'i 80 't>.l:;
80 ·- c
~~ 60 60 --11.'() c 0 ~=alt§
~ ~.!!. 40 40 A-A20:3
"" 0-020:4 " ~ 20
-·20:1 20
I ~
0 25 50 75 100 125 0 25 50 75 100 125
Fatty Acid JI).A Fatty Acid ¢.A
Fig. 3. Mean effect of increasing concentrations of various fatty acids on the incorporation of 14C-arachidonic acid into amnion triglycerides. Only spontaneous labor amnions were used. Cells were incubated with 14C-arachidonic acid (0.1 J.LCi) only (control) or in the presence of unlabeled fatty acids for 30 minutes. The radioactivity incorporated into lipids in the presence of unlabeled fatty acid is expressed as percent of control incubations (100%). Each point is the mean of two to four determinations.
with phosphatidylcholine decreases with increasing incubation time (p < 0.05), whereas the reverse is true for total phosphatidylethanolamine (p < 0.05). No changes were observed in the percentage of phospholipid-associated 14C-arachidonic acid for either phosphatidylinositol or phosphatidylserine.
The ratios of the percent of phospholipid-associated 14C-arachidonic acid to the molar percent of phospholipid are listed in Table II. The values of percentage of phospholipid-associated 14C-arachidonic acid from Fig. I, C were divided by the molar percent of phospholipids from amnion obtained in early labor published by Okita eta!.' The molar percents of phos-
pholipid for phosphatidylcholine, phosphatidylethanolamine (both plasmalogen and diacyl forms), and phosphatidylinositol are 49.1 %, 30.9%, and 7.3%, respectively.3 The computed ratios in Table II indicate that for phosphatidylcholine and total phosphatidylethanolamine, arachidonic acid is incorporated in proportion to the amount of phospholipid species present. However, arachidonate incorporation into phosphatidylinositol is three times greater than the relative amount of phosphatidyinositol present in amnion.
The effect of dose on arachidonic acid incorporation into amnion phospholipids was explored in Fig. 2. The doses tested were 3.3 to 100 J.LmoiiL. The mass of ara-
1000 Olson and Smieja
chidonate incorporated per microgram of phospholipid phosphorus was determined. As observed earlier, the greatest incorporation of arachidonate was into phosphatidvlcholine, with similar amounts incorporated into total phosphatidvlethanolamine and phosphatidvlinositol and the least amount entering phosphatidvlserine. This relationship was consistent at each dose of arachidonate. Again, there was no significant difference in the amount incorporated in cesarean section and spontaneous labor cells for anv phospholipid.
To test the specificity of arachidonate incorporation into triacvlglycerides and phospholipids. several different fattv acids were added to cell incubations in the presence of "C-arachidonic acid (Fig. 3). The compounds tested were palmitic acid (16: 0), linoleic acid (18:2. n-6). '{-linolenic acid (18:3, n-6). dihomo-'{linolenic acid (20: 3, n-6), arachidonic acid (20: 4, n-fi). and eicosapentaenoic acid (20: 5, n-3). Three of these fattv acids, 18 : 2, 18 : 3. and 20 : 3. are anabolic precursors for arachidonic acid. whereas 20:5. a fattv acid found in high levels in marine animals. is a competitive antagonist of arachidonic acid for cvdooxvgenase in the amnion'" and is the precursor for prostaglandins of the three series. Increasing the concentration of unlabeled arachidonic acid decreased the incorporation of ''(>arachidonic acid into triacylglycerides and phospholipids. Each of the other 20-carbon fatty acids. 20:3 and 20:5, similarlv decreased the incorporation of ''C-arachidonic acid into triacvlglvcerides and phospholipids, although eicosapentaenoic acid was less effective in inhibiting the incorporation of ''C:-arachidonic acid into phosphatidvlinositol and phosphatidvlserine. The other three fattv acids tested were relatively ineffective in inhibiting ''C-arachidonic acid incorporation into lipids, with the exception of linoleic acicl. which decreased ''(>arachidonic acid incorporation into phosphatidylserine in a fashion similar to high concentrations of 20: 3 and 20:4.
Comment
In manv tissues. triacylglvcerols contain low amounts of arachidonic acid.' and relativelv little arachidonic acid is incorporated into triacvlglvcerols during in situ'' or in vitro studies.'" Amnion, along with adrenal cortex and adipose tissue. contains uncharacteristically high levels of arachidonovl moieties in triacvlglvcerols." ''· '' In this studv, triacylglvcerols took up more radiolabeled arachidonic acid than other lipids. A similar observation was made with avian shell gland tissues.'-, It is unlikely that triacylglvcerol acts as a source of releasable arachidonate'; hence this active incorporation may be a means bv which the cellular free arachidonate level is kept low to diminish prostaglandin svnthesis.
October 1988 Am .J Obstet Gynecol
Schwartz et al.'" observed that more "H-arachidonic acid was incorporated into phospholipids (especially phosphatidylcholine and total phosphatidylethanolamine) after the onset of labor when expressed as the percent of total arachidonic acid incorporation. However, when expressed as percent of arachidonate incorporated into only phospholipids, there was no difference in incorporation with the onset of labor. Our results do not show any changes with the onset of labor for arachidonate incorporation into anv lipid class, regardless of how the data are expressed. Any discrepancies between these two studies mav be due to methodology, because Schwartz et al. used whole tissue and we used dispersed cells. Our data suggest that no differences in acyl-coenzvme A synthetase or acyl transferase enzyme activities exist with the onset of labor. Phospholipase A" and phospholipase C plus diacvlglycerollipase and monoacylglycerollipase are responsible for the release of arachidonic acid. and, interestingly, no changes exist with labor in the activities of either phospholipase.' It is probable, therefore, that changes in the activities of enzymes that regulate cellular free arachidonate levels are acute in response to
specific stimuli. The percent of total phospholipid arachidonate in
corporated into phosphatidylcholine was greatest during the first 30 minutes and declined during the next 2.5 hours, while the reverse was true for total phosphatidylethanolamine when 1 'C-arachidonic acid was constantly present in the extracellular medium (Fig. l. C). This might simplv reflect the orientation of phosphatidylcholine toward the outside of the cell and, consequently, a variable rate at which arachidonate enters various lysophospholipid pools. However. we have found in pulse experiments with monolaver cultures of amnion epithelial cells in which ''C-arachidonic acid was added to cells for 2 hours and then removed, that this same pattern continues for 18 to 24 hours until equilibrium of the ''C-arachidonic acid lapel is reached in all phospholipids (data not shown). Therefore, this might also suggest a fattv acid transfer between the two phospholipids involving coenzyme A and a reversal of the acyl transferase reaction. 17
· "
Arachidonic acid was incorporated verv rapidly into phosphatidvlinositol. This rate did not diminish during the 3 hours of incubation and was approximately three times greater than the incorporation rate for other phospholipids on a molar percent basis. These data suggest that arachidonate entered a pool of rapidlv turning over phosphatidylinositol, that it entered a phosphatidylinositol pool metabolicallv isolated from other phospholipids, or that lysophosphatidylinositol has a higher affinity for arachidonovl-coenzyme A transferase than other lvsophospholipids."'
Volume 159 Number 4
The position on the glycerophospholipids to which 11C-arachidonic acid was bound was not determined by hydrolysis with phospholipases. However, Schwartz et al. 6 found that 14C-palmitate and 'H-arachidonate were both incorporated into amnion phospholipids. Because saturated fatty acids are mostly found in the sn-1 position and unsaturated fatty acids are found in the sn-2 position of glycerophospholipids,' these data suggest that palmitate was bound to the sn-1 position, whereas arachidonate was bound to the sn-2 position. The results from the current study would not contradict the earlier observations, because palmitate (16: 0) did not compete with 11 C-arachidonic acid for incorporation. The data shown in Fig. 3 would suggest that acyl transferase enzymes specific for the length of the fatty acid chains exist, because 18:3 (n-6) was not as competitive as 20:3 (n-6) in inhibiting 20:4 (n-6) incorporation. This observation may have a special application to people whose diets are high in marine fat (and therefore 20: 5) and who incorporate significant levels of 20:5 into lipids. One of these groups, the Faroese, have significantly higher birth weights among single infants of primiparous mothers than among the general Danish population; this has been attributed to their high level of n-3 polyunsaturated fatty acids, which may prolong gestation.20
In summary, these data indicate that archidonic acid is incorporated rapidly into amnion lipids, ensuring that the level of free arachidonate is kept very low in the cell. Specific enzyme pathways exist for this process. There is no difference in the pattern or rate of incorporation of arachidonic acid with the onset of labor in women, indicating that archidonate mobilization for prostaglandin synthesis at term is acutely controlled and not the result of a change in basal activity of enzymes regulating arachidonate storage.
We thank Mrs. Jean Weick and Mrs. Jan Challis for typing the manuscript and Drs. Fred Possmayer and Tamas Zakar for helpful comments.
REFERENCES I. Bleasdale JE, Johnston JM. Prostaglandins and human
parturition: regulation of arachidonic acid mobilization. Rev Perinat Med 1985;5:151-91.
2. Challis JRG, Olson DM. Parturition. In: Knobil E, Neill J, eds. The physiology of reproduction. New York: Raven Press, 1988:2177-216.
3. Okita JR, MacDonald PC, Johnston JM. Mobilization of arachidonic acid from specific glycerophospholipids of human fetal membranes during early labor. J Bioi Chern 1982;257: 10429-34.
Amnion arachidonic acid incorporation 1001
4. Irvine RF. How is the level of free arachidonic acid controlled in mammalian cells? BiochemJ 1982;204:3-16.
5. Morita I, Murota S-1. Prostaglandin-synthesizing system in rat liver homogenates: ATP shifts arachidonic acid away from cyclooxygenase into phospholipids. Biochim Biophys Acta 1980;619:428-31.
6. Kroner EE, Peskar BA, Fischer H. Ferber E. Control of arachidonic acid accumulation in bone marrowderived macrophages by acyltransferases. J Bioi Chern 1981 ;256:3690-7.
7. OkitaJR, Sagawa N, Casey ML, Snyder JM. A comparison of human amnion tissue and amnion cells in primary culture by morphological and biochemical criteria. In Vitro 1983; 19:117-26.
8. Gilfillan AM, Chu AJ, Smart DA, Rooney SA. Single plate separation of lung phospholipids including disaturated phosphatidylcholine. J Lipid Res 1983;24: 1651-6.
9. Goppelt M, Resch K. Densitometric quantitation of individual phospholipids from natural sources separated by one-dimensional thin-layer chromatography. Anal Biochem 1984;140:152-6.
I 0. Plosker SM, Patei-Christopher A, Powell W, Casper RF. Inhibition of human amnion production of PGE, and PGF2• by gamma linolenic acid and eicosapentaenuic acid. In: Proceedings of the forty-second annual meeting of the Society of Obstetricians and Gynaecologists of Canada, Charlottetown, Prince Edward Island, Canada, June 23-27, 1986.
II. Puustinen T, Dahl M-L, Uotila P. The distribution of 11C-arachidonic acid in hamster lungs is sensitive to quinacrine. Prostaglandins 1983 ;25: 829-3 7.
12. Hill EE, Lands WEM. Incorporation of long-chain and polyunsaturated acids into phosphatidate and phosphatidylcholine. Biochim Biophys Acta 1968;152:645-8.
13. Christ EJ, Nugteren DH. The biosynthesis and possible function of prostaglandins in adipose tissue. Biochim Biophys Acta 1970;218:296-307.
H. Vahouny GV, Chanderbhan R, Hodges VA, Treadwell CR. Cholesterol arachidonate as a prostaglandin precursor in adrenocortical cells. Prostaglandins 1978;16:207-20.
15. Toth M, Asboth G, Hertelendy F. Turnover of lipidbound arachidonate and biosynthesis of prostanoids in the endometrium and myometrium of the laying hen. Arch Biocehm Biophys 1983;226:27-36.
16. Schwartz AL, Forster CS, Smith PA, Liggins GC. Human amnion metabolism. II. Incorporation of fatty acids into tissue phospholipids in vitro. AM J 0BSTET GYNECOL
1977;127:475-81. 17. Irvine RF, Dawson RMC. Transfer of arachidonic acid
between phospholipids in rat liver microsomes. Biochem Biophys Res Commun 1979;91: 1399-405.
18. Trotter J, Ferber E. CoA-dependent cleavage of arachidonic acid from phosphatidylcholine and transfer to phosphatidylethanolamine in homogenates of murine thymocytes. FEBS Lett 1981;128:237-41.
19. Holub BJ, Kuksis A. Differential distribution of orthophosphate-32P and glycerol- 14C among molecular species of phosphatidylinositols of rat liver in vivo. J Lipid Res 1971; 12:699-705.
20. Olsen SF, Hansen H, Sorensen TIA, et al. Intake of marine fat, rich in (n-3)-polyunsaturated fatty acids, may increase birthweight by prolonging gestation. Lancet 1986;2:367-9.