Page 1
Assessment of Methods for On-Farm Euthanasia of Layer Chickens
by
Rathnayaka Mudiyanselage Amila Subhashinie Bandara
A Thesis
Presented to
The University of Guelph
In partial fulfilment of requirements
for the degree of
Doctor of Philosophy
in
Animal Biosciences
Guelph, Ontario, Canada
© Rathnayaka Mudiyanselage Amila Subhashinie Bandara, June 2019
Page 2
ABSTRACT
ASSESSMENT OF METHODS FOR ON-FARM EUTHANASIA OF LAYER CHICKENS
R. M. A. S. Bandara Advisor:
University of Guelph, 2019 Professor T. M. Widowski
Animal care guidelines for poultry require that the methods used for routine killing result in
rapid and irreversible loss of sensibility and cause minimal pain and distress. This thesis assessed
the efficacy of three types of physical on-farm euthanasia methods in different age groups of
layer chickens, and the degree of aversion and time to loss of sensibility for different CO2
concentrations (25%, 35%, 50%, and 70%) in laying hens. In Study 1, all three commercially
available non-penetrating captive bolt devices tested caused sufficient brain trauma to result in
rapid insensibility and brain death in four different age groups (10-11, 20-21, 30-35, 60-70
weeks) of layer chickens. This study also identified and corroborated practical behavioural
indicators of death in layer chickens that can be used in field conditions to achieve the animal
care guideline requirements of confirming the death before disposing of carcasses; onset of tonic
convulsions, last movement, and cloacal relaxation were good indicators of clinical death. A
second study assessed efficacy of a commercially available mechanical cervical dislocation
device (MCD) in comparison to manual cervical dislocation (CD) in different age groups (12,
27-29, and 65-70 weeks) of layer chickens. Killing methods were assessed in anesthetized
chickens to minimize welfare concerns. MCD resulted in a longer time to brain death than CD.
Radiographs revealed that the majority of birds killed by CD had ideal dislocation sites between
the skull and atlas (C1) or between cervical vertebrae C1-C2. The MCD resulted in a majority of
Page 3
dislocations at lower cervical vertebrae. There were few fractures in birds killed by either
method. A final study demonstrated that concentrations of 50% and 70% CO2 were significantly
more aversive to laying hens than 25% and 35%, based on an approach avoidance test. However,
hens demonstrated headshaking and open mouth breathing at all tested CO2 concentrations, and
some birds displayed conditioned place avoidance at the low concentrations. Loss of posture,
indicating insensibility, occurred in less than 25s in all CO2 concentrations with shorter latencies
at higher concentrations. The thesis provides important information for refinement of future
euthanasia guidelines for the layer chicken industry.
Page 4
iv
ACKNOWLEDGEMENTS
First and foremost, I would like to thank my advisor, Dr. Tina M Widowski for her tremendous
support and guidance during my graduate career. Her understanding of me as an international
student whose family lives thousands of miles away was unbelievable. I am grateful for the
opportunities she has provided, and this would not have been possible without her
encouragement and belief in my capabilities.
My special sincere thanks should go to Dr. Stephanie Torrey, a member of the advisory
committee, whose continued support and dedication has been invaluable. Her frequent
encouragement and guidance to develop my research and further my career were remarkable. A
big thank you to Dr. Suzanne Millman for her valuable guidance as the co-advisor throughout the
completion of my research work at Iowa State University, and for constructive feedback as co-
author. I would also like to thank my advisory committee – Dr. Patricia Turner and Dr. Karen
Schwan-Lardner for their expertise, feedback and continuous involvement.
Thank you to Linda Caston for the technical support and wonderful companionship given
throughout this project. Your understanding and advice were remarkable for me to develop my
personality and to adjust to a different culture. Thank you to Dr. Anna Bolinder and Dr. Alex
zur-Linden for your expertise and technical support. Thank you to Dr. Michelle Edwards for
your vast knowledge of statistics, and the given support on advising on statistical analysis
through this thesis.
Page 5
v
Thank you to Kahlee Latreille, Rebbeca Parsons, Alex Hurtado-Terminal, Dilan C. Waters, and
Daniel Rothschild for the given technical support, and wonderful companionship during the
research project. Thank you to everyone in the Widowski Lab for helping with data collection.
Thank you to the farm staff at the Arkell Poultry Research Station and ISU LAR technicians for
animal care and technical help with this project.
Thank you to Dr. M.A.J.P. Munasinghe, former Head of the Department of Livestock
Production, Sabaragamuwa University of Sri Lanka for assisting to obtain study leave to pursue
this PhD degree programme, and arranging staff to cover my duties in the department.
I have no words to thank my loving husband Dhanushka. Your willingness to share all our
responsibilities and make sacrifices were unbelievable during this difficult period. You are the
one who gave the biggest support in this journey, physically being across the oceans but keeping
me in your heart.
Finally, I would like to sincerely thank all the birds who sacrificed for this project, in order to
attempt to improve welfare for poultry as a whole.
Page 6
vi
TABLE OF CONTENTS
ABSTRACT ................................................................................................................................................. ii
ACKNOWLEDGEMENTS ...................................................................................................................... iv
TABLE OF CONTENTS .......................................................................................................................... vi
LIST OF TABLES ..................................................................................................................................... ix
LIST OF FIGURES .................................................................................................................................. xii
CHAPTER 1 ................................................................................................................................................ 1
Introduction ................................................................................................................................................. 1
1.1 Euthanasia needs in the layer industry ................................................................................................ 1
1.2 Welfare concerns associated with euthanasia ..................................................................................... 1
CHAPTER 2 ................................................................................................................................................ 4
Literature Review and Thesis Objectives ................................................................................................. 4
2.1. Avian anatomy that helps to explain the efficacy of different euthanasia techniques ....................... 4
2.1.1. Anatomy of the avian skull ......................................................................................................... 4
2.1.2. Neuroanatomy of avian brain and spinal cord ............................................................................ 5
2.1.3. Neuroanatomy of the reflex arc................................................................................................. 10
2.1.4. Traumatic brain injuries (TBI) .................................................................................................. 11
2.1.5. Anatomy of the avian neck ....................................................................................................... 12
2.1.6. Avian respiratory system .......................................................................................................... 14
2.2. Sensibility, death, and pain .............................................................................................................. 18
2.2.1. Sensibility ................................................................................................................................. 18
2.2.2. Death ......................................................................................................................................... 20
2.2.3. Pain and its mechanism ............................................................................................................. 21
2.3. Assessing insensibility and time of death ........................................................................................ 26
2.3.1. Brain stem and spinal reflexes .................................................................................................. 27
2.3.2. Physiologic measures and behaviours ....................................................................................... 29
2.4. Euthanasia methods and mode of action .......................................................................................... 33
2.4.1. Physical methods....................................................................................................................... 33
2.4.2. Inhaled gas ................................................................................................................................ 41
2.5. Motivation, preference and aversion tests ........................................................................................ 45
2.5.1. Introduction ............................................................................................................................... 45
2.5.2. Approach avoidance and conditioned place avoidance paradigms ........................................... 47
Page 7
vii
2.6. Thesis objectives .............................................................................................................................. 49
CHAPTER 3 .............................................................................................................................................. 52
Anatomical Pathology, and Behavioural and Physiological Responses Induced by Application of
Non-Penetrating Captive Bolt Devices in Layer Chickens .................................................................... 52
3.1. Abstract ............................................................................................................................................ 53
3.2. Introduction ...................................................................................................................................... 54
3.3. Materials and methods ..................................................................................................................... 56
3.3.1. Animals and facilities................................................................................................................ 57
3.3.2. Non-penetrating captive bolt devices ........................................................................................ 57
3.3.3. Ante mortem assessments ......................................................................................................... 59
3.3.4. Macroscopic assessment of tissue damage ............................................................................... 60
3.3.5. Statistical analyses .................................................................................................................... 61
3.4. Results .............................................................................................................................................. 62
3.4.1. Ante mortem assessments ......................................................................................................... 62
3.4.2. Pathology evaluations ............................................................................................................... 65
3.5. Discussion ........................................................................................................................................ 68
3.6. Conclusion ....................................................................................................................................... 74
CHAPTER 4 .............................................................................................................................................. 88
Efficacy of a Novel Mechanical Cervical Dislocation Device in Comparison to Manual Cervical
Dislocation in Layer Chickens ................................................................................................................. 88
4.1. Abstract ............................................................................................................................................ 89
4.2. Introduction ...................................................................................................................................... 90
4.3. Methods............................................................................................................................................ 92
4.3.1. Animals and facilities................................................................................................................ 93
4.3.2. Koechner Euthanizing Device (KED) ....................................................................................... 93
4.3.3. Anesthesia and killing procedures ............................................................................................ 94
4.3.4. Ante mortem assessment ........................................................................................................... 95
4.3.5. Post mortem assessment............................................................................................................ 96
4.3.6. Statistical analyses .................................................................................................................... 97
4.4. Results .............................................................................................................................................. 99
4.4.1. Assessment of Ante mortem measures ..................................................................................... 99
4.4.2. Assessment of postmortem measures...................................................................................... 101
4.5. Discussion ...................................................................................................................................... 105
4.6. Conclusion ..................................................................................................................................... 110
Page 8
viii
CHAPTER 5 ............................................................................................................................................ 125
Aversion to CO2 Gas in Laying Hens Using Approach-Avoidance and Conditioned Place Avoidance
Paradigms ................................................................................................................................................ 125
5.1. Abstract .......................................................................................................................................... 126
5.2. Introduction .................................................................................................................................... 127
5.3. Methods.......................................................................................................................................... 130
5.3.1 Animals, Housing, and Management ....................................................................................... 131
5.3.2. Experimental room and equipment ......................................................................................... 131
5.3.3. Experimental design ................................................................................................................ 133
5.3.4. Training procedures ................................................................................................................ 133
5.3.5. Testing protocol ...................................................................................................................... 134
5.3.6. Behavioural data collection ..................................................................................................... 135
5.3.7. Statistical analyses .................................................................................................................. 137
5.4. Results ............................................................................................................................................ 139
5.4.1. Approach and avoidance behaviour ........................................................................................ 139
5.4.2. Other observed behaviours ...................................................................................................... 141
5.4.3. Parameters associated with loss of sensibility ........................................................................ 143
5.5. Discussion ...................................................................................................................................... 144
5.6. Conclusions .................................................................................................................................... 153
CHAPTER 6 ............................................................................................................................................ 162
General Discussion .................................................................................................................................. 162
6.1. Evaluation of physical on-farm euthanasia methods ..................................................................... 162
6.1.1. Humaneness of the euthanasia methods .................................................................................. 162
6.1.2. Pathology caused by euthanasia methods and effectiveness ................................................... 165
6.1.3. Limitations of reflex and behaviour measures ........................................................................ 169
6.1.4. Device success and operator safety ......................................................................................... 172
6.2. Assessment of aversion to CO2 in laying hens ............................................................................... 175
6.2.1 Evaluation of the method ......................................................................................................... 179
6.2.2 Implications .............................................................................................................................. 181
6.3 In retrospect .................................................................................................................................... 183
6.4 Overall conclusion .......................................................................................................................... 184
BIBLIOGRAPHY ................................................................................................................................... 188
Page 9
ix
LIST OF TABLES
Table 3. 1 : List of number of birds killed with the different NPCB devices by age group, strain,
body weight, and sex. Number of failed birds are indicated in parentheses. ................................ 75
Table 3. 2 : Ante-mortem assessment measures, descriptions, and procedures used, listed in
order of observation after application of each killing method ...................................................... 76
Table 3. 3: Gross and microscopic pathology scoring criteria for macroscopic, and microscopic
hemorrhage ................................................................................................................................... 77
Table 3. 4: Mean time (± SE, s) to onset of specific measures after application of different
NPCB devices in different age groups of layer chickens ............................................................. 78
Table 3. 5: Pearson correlation coefficients to assess the relationship between the antemortem
measures for different NPCB devices for all ages of layer females and males (n = 94 for Zephyr
E, n=92 for Zephyr EXL and n=93 for TED) ............................................................................... 79
Table 3. 6: Regression and relative contribution (R2) for response of dependent variable (Y) for
independent variables (X) of different NPCB devices ................................................................. 80
Table 3. 7: Summary of gross scores for subcutaneous hemorrhage, skull fractures, and subdural
hemorrhage in birds killed by different NPCB devices. Number of birds with each score are
indicated. ....................................................................................................................................... 81
Table 3. 8: Summary of microscopic scoring of brains for trauma following application of each
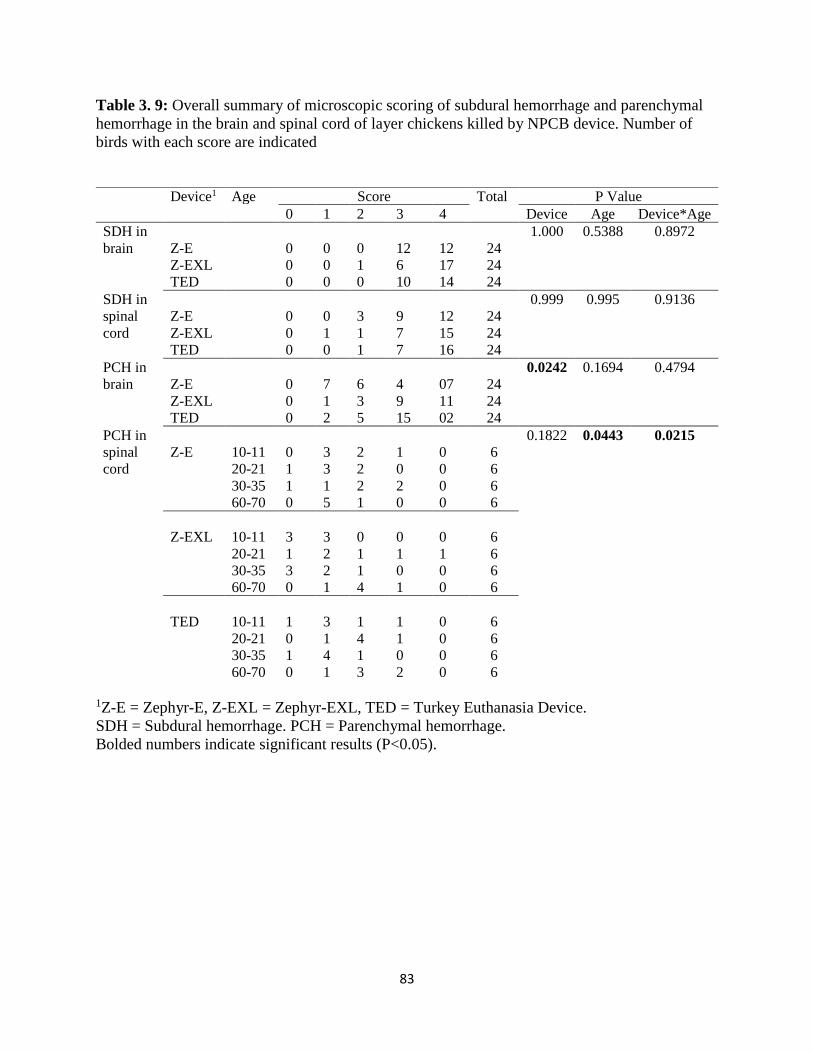
of the three NPCB devices in layer chickens. Number of birds with each score are indicated. ... 82 Table 3. 9: Overall summary of microscopic scoring of subdural hemorrhage and parenchymal
hemorrhage in the brain and spinal cord of layer chickens killed by NPCB device. Number of
birds with each score are indicated ............................................................................................... 83
Table 4. 1: Strain, sex, body weight and sample sizes for the different age classes of birds used
in the study .................................................................................................................................. 111
Table 4. 2: List of ante-mortem assessment measures, description, and procedure use, recorded
in order of observation after application of each killing method (based on Chapter 3) ............. 112
Table 4. 3: Definitions for the terminology used in radiograph evaluation ............................... 113
Page 10
x
Table 4. 4: Gross and microscopic pathology scoring criteria for macroscopic, and microscopic
hemorrhage (Woolcott et al., 2018ab; Chapter 3) ....................................................................... 114
Table 4. 5: Number of birds presenting with ante-mortem measures following application of the
killing methods............................................................................................................................ 115
Table 4. 6: Mean latencies to or durations of (± SE, s) ante-mortem measures in conscious and
anesthetized chickens killed by manual cervical dislocation in different age groups. P values are
given for effects of age, anesthesia and age by anesthesia interaction. ...................................... 116
Table 4.7: Mean latencies to or durations of (± SE s) ante-mortem measures in anesthetized
chickens killed by manual or mechanical cervical dislocation in different age groups. P values
are given for effects of age, method and age by method interaction. ......................................... 117
Table 4. 8: Presence and location of luxation/subluxation (from radiographs) and spinal cord
transections (from macroscopic evaluation) in conscious and anesthetized chickens killed by
manual cervical dislocation and KED1. Values indicate number of birds. ................................. 118
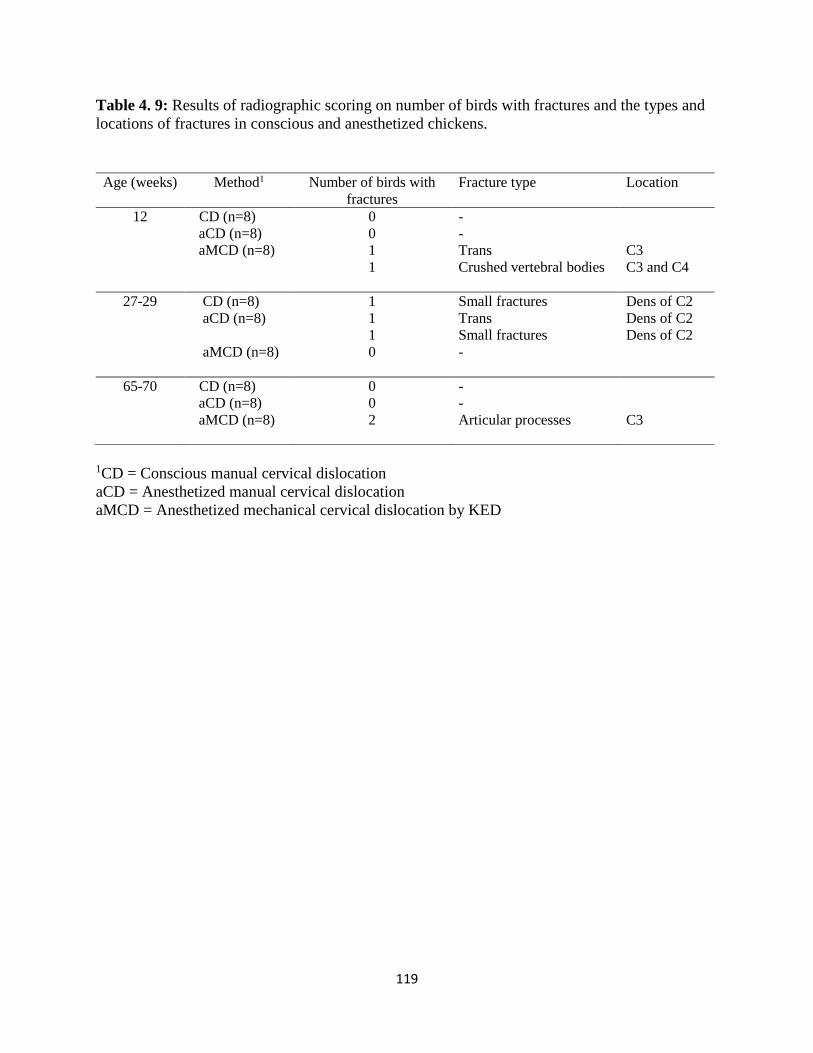
Table 4. 9: Results of radiographic scoring on number of birds with fractures and the types and
locations of fractures in conscious and anesthetized chickens. .................................................. 119
Table 4. 10: Macroscopic evaluation of subcutaneous hemorrhage (SCH) at the site of
dislocation. Number of birds with each score are indicated ....................................................... 120
Table 4. 11: Summary of microscopic scoring of brains for trauma following application of each
of the three killing methods in layer chickens. Number of birds with hemorrhage (any score >0)
in each section are indicated. ...................................................................................................... 121
Table 4. 12: Overall summary of microscopic scoring of subdural hemorrhage and parenchymal
hemorrhage in the spinal cord of layer chickens killed by three killing methods. Number of birds
with each score are indicated. ..................................................................................................... 122
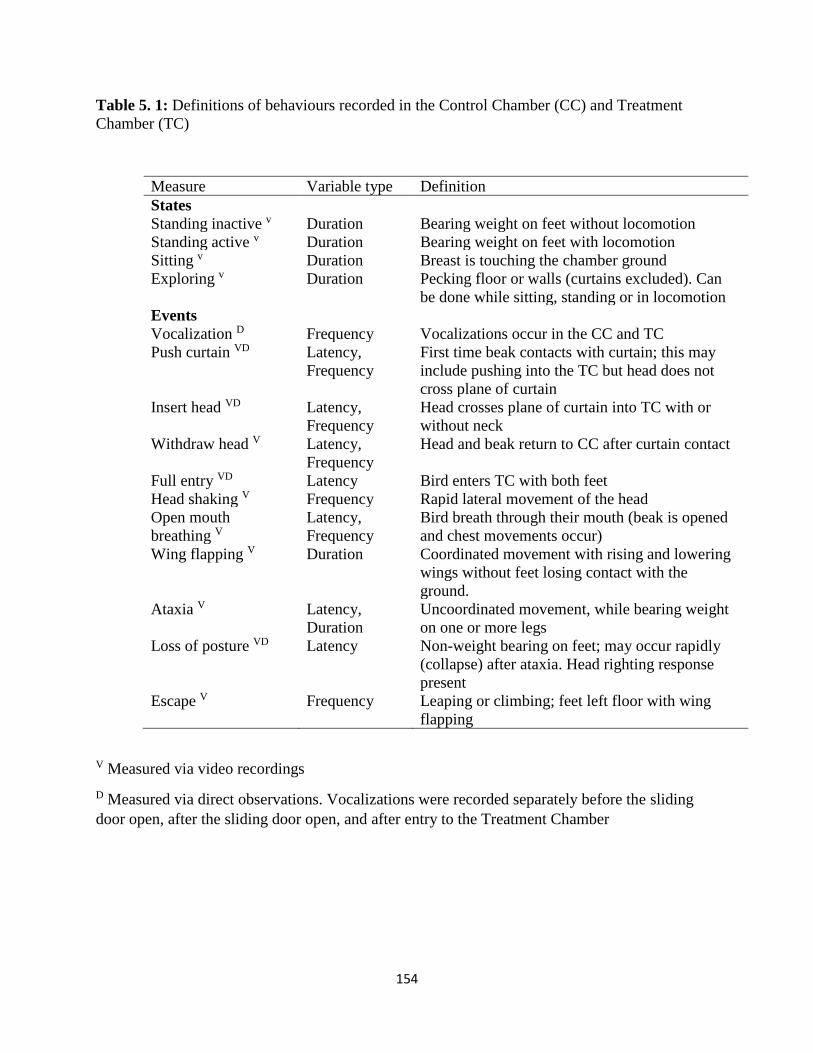
Table 5. 1: Definitions of behaviours recorded in the Control Chamber (CC) and Treatment
Chamber (TC) ............................................................................................................................. 154
Table 5. 2: Test order, assigned CO2 treatment and outcomes in each of the four rounds for the
12 hens enrolled in the study. ..................................................................................................... 155
Page 11
xi
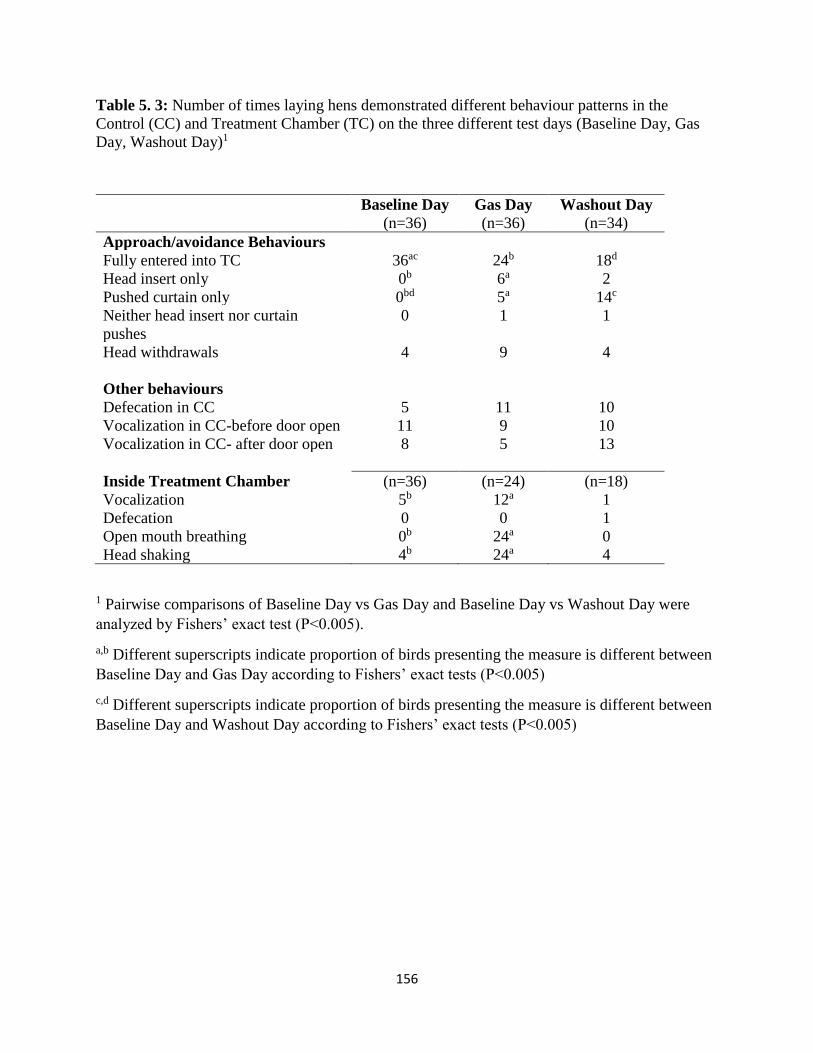
Table 5. 3: Number of laying hens that demonstrated different behaviour patterns in the Control
(CC) and Treatment Chamber (TC) on the three different test days (Baseline Day, Gas Day,
Washout Day)1 ............................................................................................................................ 156
Table 5. 4: Number of laying hens that demonstrated different behaviour patterns in the Control
(CC) and Treatment Chamber (TC) at the different CO2 concentrations on Gas Days. ............. 157
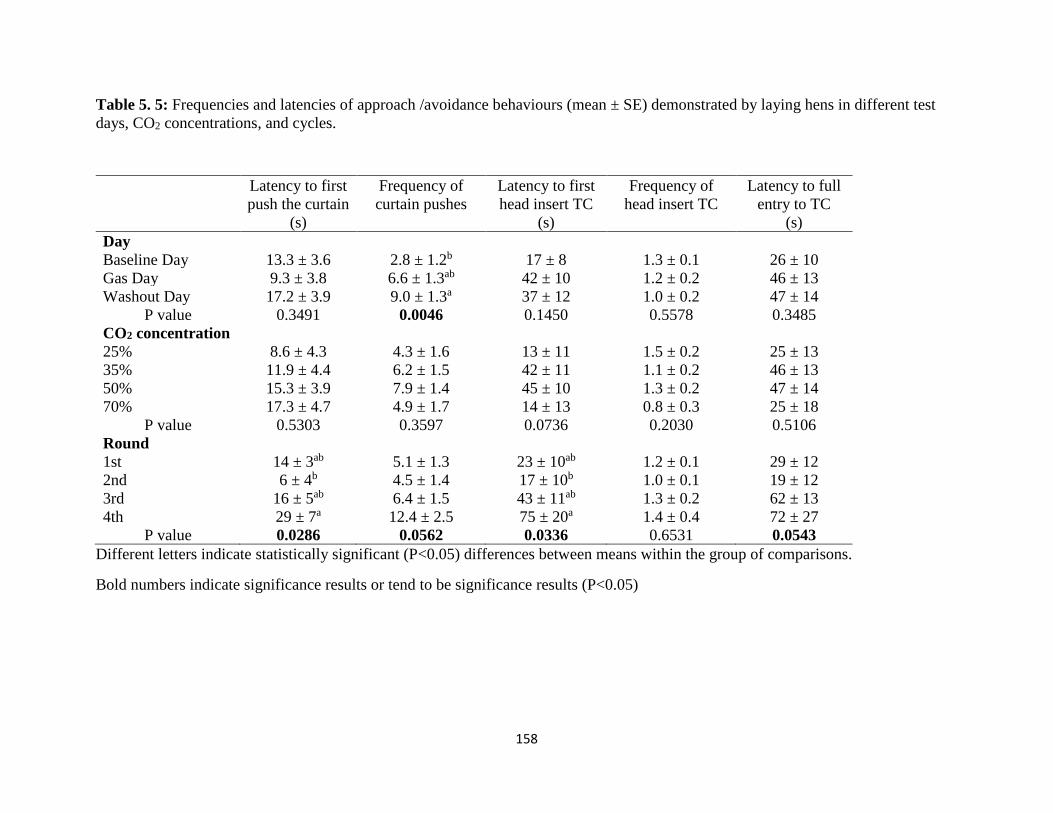
Table 5. 5: Frequencies and latencies of approach /avoidance behaviours (mean ± SE)
demonstrated by laying hens in different test days, CO2 concentrations, and cycles. ................ 158
Table 5. 6: Behaviours associated with loss of consciousness on Gas Day in laying hens: Mean
time ± SE s are presented for 25%, 35%, and 50% CO2 concentrations. Average time ± SD s are
presented for 70% CO2 concentration......................................................................................... 159
Table 5. 7: Time (s) at onset of behaviour during recovery in laying hens exposed to different
CO2 concentrations. Mean time ± SE s are presented for 25%, 35%, and 50% CO2 concentrations
..................................................................................................................................................... 160
Page 12
xii
LIST OF FIGURES
Figure 2. 1: (A) Schematic diagram of modern consensus view of avian brain according to the
conclusions of the Avian Brain Nomenclature Forum (Jarvis et al., 2005, with permission). (B)
Dorsal view of a chicken brain: C- cerebrum, D-cerebellum, H- hind brain, P- optic lobes, S-
spinal cord. ...................................................................................................................................... 7
Figure 2. 2 : Ventral view of the brain of a chicken with cranial nerves (Orosz, 2007) ............... 8
Figure 2. 3 : A radiograph of the cervical vertebrae of a layer chicken (21 weeks old hen. SK-
skull, O- occipital lobe, C1- first cervical vertebra, C2- second cervical vertebra, C3- third
cervical vertebra, C4- fourth cervical vertebra. ............................................................................ 14
Figure 3. 1: Non-penetrating captive bolt devices. (A) Zephyr-E standard: (B) Conical shape
bolt head, (C) Standard subject adapter. (D) Zephyr-E-layer: (E) Round shape bolt head, (F)
Chicken subject adapter. (G) Zephyr-EXL: (H) Conical shape bolt head, (I) chicken subject
adapter. (J) Turkey Euthanasia Device (TED): (K) Flat bolt head, (L) R-3 subject adapter. ....... 84
Figure 3. 2: Application of the Zephyr-EXL device on a 30 w.o. hen: The bird was restrained in
sternal recumbency with its neck resting ventrally on the ground, and the wings held gently
towards the body of the bird. Device was placed perpendicular to the top of the frontal bone just
behind the comb and on the mid line between the eyes and ears. ................................................ 85
Figure 3. 3 : Figure 3. Gross pathology scoring criteria for skull fractures. Arrows indicate the
fracture type [modified from Erasmus et al., (2010b) and Casey-Trott et al., (2013)]. (A) No
fracture, intact skull (score 0). (B) Depression fracture (score 1). (C) Penetrating fracture-no
imbedded fragments (score 2). (D) Penetrating fracture- with imbedded fragments (score 3). ... 86
Figure 3. 4 : Skin reflected to demonstrate gross subcutaneous hemorrhage. (A) Hemorrhage
with less than 25% of area covered (score 2) of a 65 w.o. bird killed by the TED. (B)
Hemorrhage completely covering area from the eyes to base of the skull (score 4) of a 10 w.o.
bird killed by the TED. (C) Gross subdural dorsal hemorrhage covering less than 25% of the
brain surface (score 1) of a 33 w.o. bird killed by the Zephyr-E. (D) Gross subdural dorsal
hemorrhage covering 51 -75% of the brain surface (score 3) of a 33 w.o. bird killed by the
Zephyr-E. ...................................................................................................................................... 87
Figure 4. 1: A- Koechner euthanasia device (KED model-C): S-single side blade; D-double
angle blade. B-application of KED model-C in a 65 week old anesthetized rooster. ................. 123
Page 13
xiii
Figure 4. 2 : Radiographs of chickens showing cervical dislocations. A- Luxation between the
skull and C1 vertebra in a 65 week old rooster killed by manual cervical dislocation: Sk-Skull,
C1- first cervical vertebra, C2- second cervical vertebra. B - Subluxation between C2 and C3
(letter S shows the site of subluxation) in a 65 week old rooster killed by KED: fractures (F) are
present on the articular processes of the C3 vertebra. ................................................................ 124
Figure 5. 1 : A- Lateral camera view of a laying hen inside control chamber (CC) waiting for the
sliding door (S) to open and to enter treatment chamber (TC). Wire mesh (W) can be seen on top
of the CC. B- Lateral camera view of hen pushing through the curtain (C) to enter treatment
chamber (TC) to access meal worms. Both images were collected on a Baseline Day, with
ambient air conditions in CC and TC. ........................................................................................ 161
Figure 6. 1 : The fabric chicken restraint device (fabric sleeve made of nylon): A- a toggle at the
head end to tighten it, B- Opening at the bottom end ................................................................. 187
Page 14
1
CHAPTER 1
Introduction
1.1 Euthanasia needs in the layer industry
On-farm euthanasia is considered to be a fundamental aspect of management in livestock
production. In the poultry industry, some of the most common reasons for killing birds on the
farm are (1) to prevent suffering from injury or sickness, (2) disease control, and (3) stock
management (e.g., of male layers). In Canada, all layer farms are required to have a euthanasia
plan and any injured or diseased birds found in layer flocks need to be identified and euthanized
in a timely manner (NFACC, 2017). Therefore, on-farm euthanasia is necessary for eliminating
suffering and distress and maintaining flock health and well-being.
1.2 Welfare concerns associated with euthanasia
The term euthanasia is used to describe ending the life of an individual to minimize or eliminate
pain and distress (AVMA, 2013). The term euthanasia is derived from the Greek word “eu”
meaning good and “thanos” meaning death. When an animal no longer has good welfare (when it
no longer has a life worth living; Mellor, 2016), the humane thing to do is to offer it a good
death. The desired outcome of euthanasia is relieving suffering (minimize pain, distress, and
negative affect to the animal). Therefore, the humaneness of the technique is considered to be an
important ethical issue. Many authors define the humaneness of a killing technique based on how
quickly the animal loses sensibility (Erasmus et al., 2010a; Martin et al., 2016; Woolcott et al.,
2018 a,b). Therefore, the preferred methods are ones that cause minimal pain and distress and
Page 15
2
result in rapid loss of sensibility followed by loss of brain function and, ultimately, respiratory
and cardiac arrest (AVMA, 2013).
Consideration must also be given to selecting a euthanasia method that is reliable, effective,
practicable, safe, easy to use and as esthetically acceptable as possible (AVMA, 2013). It is
mandatory for the personnel performing the euthanasia to demonstrate proficiency in the use of
the technique (NFACC, 2016). Experience in humane restraint of the animal is also important to
ensure that pain and distress are minimized. Moreover, proper physical handling is vital to ensure
the safety of the person performing the euthanasia and to protect other persons and animals.
On these grounds, the American Veterinary Medical Association classified euthanasia methods
as acceptable, acceptable with conditions, and unacceptable (AVMA, 2013). Acceptable
euthanasia methods for poultry include overdose of injectable anesthetic (barbiturates and
barbituric acid derivatives). Inhaled gases (CO2, CO, N2, Ar) are acceptable with the condition
that they cause rapid loss of sensibility and no or minimal aversiveness to the species. Manual
and mechanical cervical dislocation are acceptable, but with the condition of no crushing of
cervical bones and spinal cord unless the bird is first rendered insensible. Decapitation is
acceptable with the condition that it must be performed by a competent person with a sharp
instrument, ensuring rapid and unobstructed severing of the head from the neck. Penetrating and
non-penetrating captive bolts are acceptable for euthanasia of large poultry with appropriate
restraint and rapid loss of sensibility. Blunt force trauma applied manually to the head is not
listed as either acceptable or acceptable with conditions for poultry euthanasia in AVMA (2013)
guidelines, as this method could be displeasing to the personnel when performing it and repeated
Page 16
3
application can result in personnel fatigue, loss of efficacy, and humane concerns. However, in
the recent Code of Practice for the Care and Handling of Pullets and Laying Hens, NFACC
(2017) listed blunt force trauma as an on-farm euthanasia option.
Therefore, all euthanasia techniques must meet certain criteria to be considered humane.
However, not all existing methods have been scientifically evaluated to determine whether or not
they meet these criteria for laying hens, and new methods are being developed that also require
evaluation.
Page 17
4
CHAPTER 2
Literature Review and Thesis Objectives
Current on-farm euthanasia techniques for poultry involve separation of the body from brain and
central nervous system control (eg., mechanical/manual cervical dislocation or decapitation),
direct destruction of the brain (blunt force trauma, penetrative or non-penetrative captive bolts),
and use of inhaled gas leading to death by hypoxia. This chapter explains: 1. Basic anatomy and
related functions of the avian skull, brain, neck and cardiorespiratory system; 2. Consciousness,
sensibility, death and pain; 3. Assessing insensibility and time to death by using different
parameters; 4. Euthanasia methods and their modes of action; 5. Motivation, preference and
aversion tests, which explains methods for assessing aversiveness to gaseous methods in animals.
All five topics in the chapter help to gain a better insight into scientific assessment of efficacies
and welfare implications of poultry on-farm euthanasia techniques.
2.1. Avian anatomy that helps to explain the efficacy of different euthanasia
techniques
2.1.1. Anatomy of the avian skull
The avian skull is light weight and strong and is characterized by a beak which replaces the true
jaw of mammals (Hogg, 1982). The dorsal view of the skull is cone shaped where the beak
creates the point of the cone. However, the skull is flattened on the ventral side. Two large
orbital fossa help to accommodate relatively large eyes. The avian skull is divided into two
cranial and facial anatomical areas which are formed by collections of several bones. Facial
Page 18
5
bones are located at the front of the skull (nasal bone) and beak. The beak is formed by the
premaxillary, the maxillaries, and the nasal bones (these bones are completely fused in most
adult birds, and sometimes pneumatized). Cranial bones are those that form the back of the skull
and the cranial cavity, which accommodates the brain. The main cranial bones are the occipital
bone, the parietal bones, the temporal bones and the frontal bones. The “foramen magnum” is the
large opening located in the back of the skull (within the occipital bone) where the spinal cord
passes through to connect with the brain. The suture lines of the cranial bones and facial bones
are visible until 4-5 months of age, but fuse at older ages in domestic fowl (Hogg, 1982).
2.1.2. Neuroanatomy of avian brain and spinal cord
The avian brain is a delicate structure accommodated inside the cranial cavity. Anatomically it is
divided into three major regions: cerebrum (telencephalon), cerebellum (metencephalon), and
brain stem (compromised of medulla oblongata, pons, mesencephalon [mid brain] and
diencephalon) (Figure 2.1). The brain is encased in three protective connective tissue layers
known as meninges: dura mater, arachnoid mater and pia mater, all developed from the
mesoderm of the embryo. The subarachnoid space contains cerebrospinal fluid.
The largest area of the avian brain is the cerebrum. The avian cerebrum is divided into the left
and right hemispheres by a fissure, which is similar to the mammalian brain and is involved in
the processing of sensory information (visual and auditory) and control of motor activity (Jarvis
et al., 2005). Some differences exist in the cerebral cortex of a bird: the cerebral cortex has a
smooth surface and lacks gyri and sulci that are present in mammalian brains. In addition, the
cortical cells which process information are absent on the surface of avian brains but process the
information using subcortical nuclei located deep in the cortex (Orosz, 2007). The olfactory
Page 19
6
lobes of birds are relatively smaller than in mammals (Husband and Shimizu, 1999). The
cerebellum is a well-developed structure in birds and controls essential functions of locomotion
and balance (Pearson, 1972). Dorsally, the mid brain is covered by the cerebellum and cerebral
hemispheres. The mid brain is mainly responsible for coordinating sensory input, and possesses
two channels which connect left and right sides of the brain. It consists of the optic tectum (roof)
and tegmentum (floor) (Whittow, 2000). The avian optic tectum, which processes visual stimuli,
is comparatively enlarged and highly developed in comparison to mammals who have a tiny
superior colliculus instead (Whittow, 2000). The medulla oblongata (brain stem) connects the
spinal cord to the brain and is responsible for controlling respiration, blood circulation, motor
functions and movement related to food intake (Nickel, 1977). The reticular formation, formed
by a bewildering array of nuclei in the brain stem helps to integrate information between the
medulla, cerebellum and spinal cord through ascending and descending neural networks (Jones,
1995). It plays a crucial role in sensibility and arousal in both mammals and birds (Revzin,
1965). Therefore, loss of sensibility results from any damage to the reticular formation or to the
pathway that connects the reticular formation to the cerebral cortex. The pons is located above
the medulla oblongata which helps to integrate information in between cerebellum and cerebrum
by forming the nuclei bridge. The diencephalon consists of the epithalamus, thalamus, and
hypothalamus (Breazile and Heartwig, 1989). The epithalamus controls some functions in the
limbic system while the pineal gland, located in the epithalamus, regulates circadian rhythms.
The thalamus integrates sensory inputs which are received from different sensory nerves. The
hypothalamus regulates homeostasis by producing a variety of hormones responsible for various
functions in the body.
Page 20
7
A B
Figure 2. 1: (A) Schematic diagram of modern consensus view of avian brain according to the
conclusions of the Avian Brain Nomenclature Forum (Jarvis et al., 2005, with permission). (B)
Dorsal view of a chicken brain: C- cerebrum, D-cerebellum, H- hind brain, P- optic lobes, S-
spinal cord.
Birds have 12 cranial nerves (CN) which are similar to those of mammals (Orosz, 1996; Figure
2.2). The olfactory and optic nerves emerge from the cerebrum while the other ten nerves emerge
from the brain stem. The olfactory nerve (CN I) is a sensory nerve responsible for smell, while
the optic nerve (CN II) is responsible for vision. The oculomotor nerve (CN III) is a motor nerve
which controls the dorsal, ventral and medial rectus muscles, and the ventral oblique muscle of
the eye. It also innervates the muscle of the eyelid. The dorsal oblique muscle of the eye is
controlled by the trochlear nerve (CN IV). The trigeminal nerve (CN V) is a mixed nerve
consisting of two sensory branches (the ophthalmic branch and maxillary branches) and one
motor branch (mandibular). The sensory branches supply nerves to the upper eye lid, skin of the
forehead, nasal cavity, upper beak, lower eyelid, palate and infraorbital sinus. The mandibular
branch innervates the muscles for mastication and skin and the mucosa at the commissures of the
Page 21
8
beak. The abducent nerve (CN VI) controls the lateral rectus muscle and muscle of the third
eyelid. The facial nerve (CN VII) controls the hyoid and cutaneous neck muscle and is not
involved in the sense of taste in birds as it is in mammals. The vestibulocochlear nerve (CN VIII)
is responsible for posture in relation to movement of the head, and eye coordination. In addition,
CN VIII facilitates perception of hearing. The glossopharyngeal nerve (CN IX) receives sensory
input from taste fibres and controls larynx and trachea joining with the vagus nerve (CN X).
Superficial muscles of the neck are controlled by the spinal accessory nerve (CN XI) and vagus
nerve. The hypoglossal nerve (CN XII) and glossopharyngeal nerve also innervate the tracheal
muscles.
Figure 2. 2 : Ventral view of the brain of a chicken with cranial nerves (Orosz, 2007)
Page 22
9
The spinal cord is located within the vertebral column and is the primary neural pathway
connecting the body and the brain. The spinal cord connects to the caudal end of the brain stem.
The meninges covering the brain continue around the spinal cord and work as a protective
casing. Pia matter envelops the spinal cord as the innermost layer, and the middle layer is
arachnoid matter (Yew et al., 1996). There are spaces in between the meninges, and
cerebrospinal fluid and arteries which supply oxygenated blood to the spinal cord are located
inside the subarachnoid space. The outer layer of the spinal cord is formed by the myelinated
nerve tracts (white matter) while the inner layer consists of grey matter (cell bodies of inter
neurons and motor neurons, unmyelinated axons and neuroglia cells (Butler and Hodos, 1996).
A pair of right and left spinal nerves exit between each pair of vertebrae. Spinal nerves transmit
information about the internal and external environment of the body to the brain and vice versa.
The region of the cord from which one set of spinal nerves emerges is known as a spinal
segment. Each spinal segment is named for its corresponding vertebrae. These segments are
grouped into regions according to main body regions through which the vertebral column passes
(cervical- in the neck, thoracic- in the chest cavity, lumbar- in the abdominal region, sacral- in
the pelvic region). The avian spinal cord vertebral column are the same length, and birds do not
have a cauda equina as in mammals (Orosz, 1996). Spinal nerves emerge laterally rather than
caudally through the vertebral foramin. However, the dura is separated from the periosteal lining,
forming an epidural space in the cervical and thoracic regions. This epidural space is filled with a
fluid (gelatinous tissue) and acts as a shock absorber, and facilitates the flexibility of the neck in
birds (Orosz, 1996).
Page 23
10
2.1.3. Neuroanatomy of the reflex arc
Reflexes are automatic responses to internal or external stimuli. The reflex arc is the functional
unit of nerve reflexes. In mammals and birds, some sensory neurons synapse with the motor
neurons in the spinal cord without passing directly into the brain, and facilitate reflex actions
more quickly than signals that pass through the brain. However, the brain can receive sensory
impulse from the reflex.
Reflex arcs that consist of only two neurons (a sensory and a motor neuron) are considered
monosynaptic and are the simplest skeletal muscle reflexes; an incoming sensory axon from a
muscle receptor enters through the dorsal root and terminates on a motor neuron in the ventral
horn of the gray matter. Most reflex arcs consist of three neurons: a sensory neuron, an
intermediate neuron (a connecting or relay neuron) and a motor neuron. For example, a stimulus
activates the pain receptors (nociceptors) of the skin and initiates an impulse in a sensory neuron.
This impulse travels to the spinal cord via the sensory neuron and passes to the relay neuron in
the spinal cord. The relay neuron passes the impulse to the brain and the motor neuron that
transmits the impulse to the muscles, causing them to contract and/or pull away from the source
of pain (Butler and Hodos, 1996).
Reflexes whose arcs pass through the spinal cord are called spinal reflexes. The spinal reflexes
are important in providing an automatic muscle reaction in response to a stimulus eg: withdrawal
reflex (pedal reflex) in chickens. In the withdrawal reflex, stimuli that are painful or unexpected
result in the reflexive withdrawal of a limb or the entire body. This type of withdrawal is caused
Page 24
11
by positive excitatory reflexes where stimuli activate motor neuron to contract the muscle (Butler
and Hodos, 1996).
Brain stem reflexes are controlled by the reflex center in the hind brain. Brain stem reflexes are
associated with the cranial nerves. These reflexes help to indicate the source or location of brain
stem lesions and neurological disorders (Benett, 1994). Insensibility and complete loss of
function in the brain of an animal is assessed based on the absence of brain stem reflexes
(Erasmus et al., 2010; Sandercock et al., 2014; Martin et al., 2016; Terlouw et al., 2016b).
2.1.4. Traumatic brain injuries (TBI)
Brain injuries are categorized in to traumatic brain injuries (TBI) and non-traumatic brain
injuries. TBI is defined as an alteration in brain function, or other evidence of brain pathology,
caused by an external force (David et al. 2010). When the injury is caused by an internal force
(e.g., stroke, infectious disease, electrical shock, lack of oxygen) it is known as non-traumatic
brain injury.
There are two types of head injuries; these are classified as closed head injuries and penetrating
head injuries (Gerstenbrand and Stepan, 2001). The closed head injury is an injury to the brain
caused by an outside force without any penetration of the skull. When a foreign body penetrates
the skull and passes into the meninges, it is known as a penetrating brain injury. There are four
main pathoanatomical sequelae of TBI: contusions, subarachnoid hemorrhage, hematomas
(including epidural, subdural, and intra parenchymal lesions), and diffuse axonal injuries
(Gerstenbrand and Stepan, 2001). Other than those four, ischemic brain injury and cerebral
Page 25
12
edema might be included in a “pathoanatomic” classification scheme. Strong rotation of the head
or shaking of the head causes diffuse axonal injury (Shaw, 2002).
Cerebral concussion is one of the most common traumatic brain injuries which is caused by
violent physical shaking of the brain and is responsible for a sudden temporary loss of
sensibility. Two authors define the cerebral concussion as a temporary disturbance of neural
activities due to sudden acceleration or deceleration of the head (Rosenthal, 1993; Label, 1997).
Due to high speed impact to the skull, blood vessels in the brain are torn and cause internal
bleeding. Hemorrhages are classified according to the location in the brain by using the name of
the meninges as a guide. Hemorrhage above the dura matter is classified as epidural hemorrhage
(EDH). A subdural hematoma (SDH) is a collection of blood below the inner layer of the dura
but external to the brain and arachnoid membrane. EDH and SDH are the most common type of
traumatic intracranial lesions in humans (Haselsberger et al., 1988). Mortality in acute SDH is
reported between 57% to 90% in humans (Haselsberger et al., 1988; Wilberger et al., 1991).
Bleeding into the cerebrospinal fluid which is located below the arachnoid membrane is known
as subarachnoid hemorrhage.
2.1.5. Anatomy of the avian neck
The neck of the chicken consists of 14 cervical vertebrae (C1-C14) (McLeod et al., 1964; Figure
2.3). The bodies of the vertebrae consist of freely movable saddle joints making the vertebral
column flexible (McLeod et al., 1964). The outer surface of the vertebrae consists of articular
and transverse processes (McLeod et al., 1964; Whittow, 2000). The first vertebrae (Atlas) and
Page 26
13
the second (Axis) are morphologically different from the rest of the vertebrae. The atlas (C1) is a
small, ring-like structure with a deep cavity (McLeod et al., 1964). The atlas attaches to the skull
via the occipital condyle to form the occipito-atlantal joint and facilitates the head to turn on the
neck (McLeod et al., 1964). The Axis (C2) attaches to the Atlas by atlanto-axial joints and helps
for cervical rotation (McLeod et al., 1964). The rest of the cervical vertebrae are similar to each
other. Each vertebra has a body that is concave on its superior surface and convex on its inferior
surface (McLeod et al., 1964). The hollow center of the body is called as vertebral foramen
where the spinal cord is located (McLeod et al., 1964; Whittow, 2000). Intervertebral discs are
located between the vertebral bodies, and are involved in cervical spine motion, stability, and
weight-bearing. (McLeod et al., 1964; Whittow, 2000)
Dislocation or fracture in vertebrae can result from mechanical damage (flexion, rotation,
compression, or extension) (Taneichi et al., 2005; Veras et al., 2000). Mechanical damage to the
vertebrae commonly results in damage to the spinal cord. Carotid arteries supply the head and
neck with oxygenated blood. The chicken has paired carotid arteries that run one on top of the
other in ventral side of the neck (MacLelland, 1990). Each carotid artery divides into three
separate arteries: the occipital artery, the internal carotid artery and the external carotid artery at
the base of the skull, and provide blood to separate areas of the head.
Page 27
14
Figure 2. 3 : A radiograph of the cervical vertebrae of a layer chicken (21 weeks old hen. SK-
skull, O- occipital lobe, C1- first cervical vertebra, C2- second cervical vertebra, C3- third
cervical vertebra, C4- fourth cervical vertebra.
2.1.6. Avian respiratory system
The primary function of the respiratory system is gas exchange by delivering enough oxygen and
removing sufficient carbon dioxide for metabolic demand. There are marked differences between
the mammalian and avian respiratory systems. In chickens and turkeys, the nostril is closed
dorsally by a horny flap, the operculum. There is a vertical lamella of cartilage at the ventral
border of the nostril (McLelland, 1990). The avian trachea is typically longer and wider than the
trachea of mammals (Gleed et al., 2001), and lies on the right side of the neck. The tracheal
cartilage are complete rings and consist of broad and narrow parts (Gleed et al., 2001). The
Page 28
15
trachea divides into two primary bronchi at the syrinx which is responsible for vocalization in
birds (Gleed et al., 2001).
Avian lungs are paired and attached firmly to the dorsal ribs (Duncker, 1974). Each lung is
triangular or quadrilateral in shape, does not divide into lobes and does not change volume
during breathing (Duncker, 1974). Most birds have nine air sacs which are poorly vascularized
by the systematic circulation (Duncker, 1974). In the chicken, the nine air sacs include four pairs
of air sacs and one unpaired sac: two interclavicular air sacs, two abdominal air sacs, two
anterior thoracic air sacs, two posterior thoracic air sacs, and one in the cervical area (Maina,
2003). Air sacs do not directly contribute to gas exchange (Magnussen et al., 1979). However,
these air sacs contribute to effective respiration by helping to ventilate the lungs (Farmer, 2006).
The total volume of the respiratory system in birds is greater than in comparably sized mammals
due to the air sacs (Magnussen et al., 1979). In contrast to mammals, birds do not have a
diaphragm to control the air pressure inside the thoracic cavity, and the avian thoracic cavity is at
the atmospheric air pressure (Whittow, 2000).
There are three types of bronchi in birds: primary, secondary and tertiary (called parabronchi)
(Duncker, 1974). Tertiary bronchi are the functional unit of gas exchange (Duncker, 1974).
There are connections between the secondary bronchi and air sacs. Smooth muscle rings that
surrounded the parabronchi lumen generates the force to move air in and out of the air sacs and
through the parabronchial lung (Duncker, 1974).
Page 29
16
2.1.6.1. Pulmonary circulation
The avian cardiovascular system is considered a high-performance system in comparison to
mammals, due to a proportionally larger heart, larger stroke volume, greater cardiac output,
higher blood pressures, and a lower heart rate (Whittow, 2000). Oxygen and carbon dioxide
exchange is also more efficient in birds than in mammals (Altman et al., 1997). The functional
anatomy of the pulmonary circulation has been described for the domestic fowl (Duncker, 1974;
Abdalla and King, 1975). Interparabronchial arteries run in between the parabronchi and branch
out to the pulmonary blood capillaries near the outside edge of the parabronchial mantle forming
a meshwork of capillaries (Duncker, 1974). Pulmonary capillary blood (oxygenated blood) is
collected in intraparabronchial veins located near the outside edges of the parabronchus and
transport to the heart via pulmonary vein (Duncker, 1974).
2.1.6.2. Welfare perspective on breathlessness
Breathlessness is a negative subjective experience that can impact one’s welfare (Beausoleil and
Mellor, 2015). Breathlessness is well documented in humans and identified as an unpleasant
sensation (Evans et al., 2002; von-Leupoldt et al., 2008). Dyspnoea is the most common term
used to describe sensations and experiences of respiratory discomfort in humans. According to
the definition of the American Thoracic Society “Dyspnoea is a term used to characterize a
subjective experience of breathing discomfort that consists of qualitatively distinct sensations
that vary in intensity. The experience derives from interactions among multiple physiological,
social and environmental factors, and may induce secondary physiological and behavioural
responses” (Parshall et al., 2012). However, in the veterinary literature, the term dyspnoea is
defined as “difficult, laboured breathing” (Mellema, 2008).
Page 30
17
Dyspnea, or breathlessness, is described in terms of its 3 different qualities: respiratory effort, air
hunger, and chest tightness (Beausoleil and Mellor, 2015). The degree of unpleasantness varies
in different qualities of breathlessness (Banzett et al., 2008). Respiratory effort is an increased
effort (depth or rate) to achieve the necessary or desired level of ventilation (Beausoleil and
Mellor, 2015). In respiratory effort, a motor command is sent to the respiratory muscles to
increase its activity to achieve the desired level of ventilation (e.g., during exercise). Air hunger
is the increased urge or increased need to breathe (Beausoleil and Mellor, 2015) and reported as
more unpleasant than respiratory effort (Banzett et al., 2008). Poor coordination between
automatic motor command and the degree of lung inflation causes air hunger (Lansing et al.,
2009). In general, increased level of CO2 in blood (hypercapnia), reduced arterial oxygen tension
(hypoxaemia) or metabolic acidosis results in the sensation of air hunger (Beausoleil and Mellor,
2015). Chest tightness occurs due to bronchoconstriction which can be a result of respiratory
inflammation or allergic bronchitis (Beausoleil and Mellor, 2015).
When assessing the humaneness of different euthanasia methods, authors utilize different
terminology to describe the respiratory responses in animals (gasping in broilers exposed to
Argon or CO2 – Gerritzen et al., 2000; Lambooij et al., 1999; deep breathing (deeper than normal
inspiration through the mouth) in laying hens and broilers exposed to CO2 or Argon- Webster
and Flethcer, 2001, mouth breathing in layers and broilers- Webster and Fletcher, 2004;
respiratory disruption (apparent increased inhalation depth and duration) in broilers exposed to
controlled atmosphere stunning with nitrogen, CO2, or argon (McKeegan et al., 2005). Some
terminologies operationally describe the behaviour (e.g., open mouth breathing) whereas others
describe the association with pain and distress (e.g., respiratory disruption). However, it is most
Page 31
18
reasonable to utilize terminology that describes the behavioural response, since associated
feelings of breathlessness or pain are currently unknown in animals.
In hypoxia or ischaemia, the eupnoea (normal rhythmic breathing) fails and gasping (a second
pattern) occurrs (St-John, 2009). In gasping, frequency and tidal volume of respiration initially
increase and then decline and cease (St-John, 2009). The functional mechanism of gasping is
associated with autoresuscitation and return of eupnoea (St John & Paton, 2004). The
mechanisms of eupnoea and gasping are generated in the brainstem respiratory control system
(St-John, 2009).
In this thesis the term “gasping” is used to refer to paroxysmal opening of the beak in in Chapter
3 and Chapter 4. This behaviour was observed in the chickens killed by nonpenetrating captive
bolts and cervical dislocation. Chest movement did not occur in these birds during gasping
behaviour. The term “open mouth breathing” is used to describe the observed breathing
behaviour in chickens exposed to CO2 in the Chapter 5. In open mouth breathing, birds breath
through their mouth (beak is opened and chest movements occur). It was difficult to conclude the
observed pattern of breathing was normal or abnormal in the CO2 chamber. Thus, it is reasonable
to describe the observed breathing behavour as open mouth breathing.
2.2. Sensibility, death, and pain
2.2.1. Sensibility
Sensibility cannot be considered as one single phenomenon. In general, sensibility is described
as the state of wakefulness which integrates sensation, sight, vocalization, and feeling (Damasio
Page 32
19
and Meyer, 2009). Human consciousness is described as incorporating external (behavioural)
and internal (cognitive, mental) components (Damasio and Meyer, 2009). The external
component includes signs of wakefulness, background emotions, sustained attention towards
objects and events in the environment, and the internal component includes more about the
mental state representing objects and events in relation to onse’s self (Damasio and Meyer,
2009).The primary level of consciousness in humans is defined as "an awareness of one's
surroundings, of the self, and of one's thoughts and feelings" (Sommerhoff and MacDorman,
1994). Because wakefulness is always an inferred state in animals and can only be assessed
indirectly, the term sensibility is preferred over consciousness to describe animal wakefulness
and responsiveness to external cues. In this thesis, the term of insensibility is used over
consciousness throughout.
Sensibility is evaluated in different states from wakefulness to sleeping to being in a coma
(Zeman, 2005; Sandercock et al., 2014). In neurobiological definitions, sensibility refers to
mental responsiveness and is associated with the reticular activating system. The Ascending
Reticular Activating System (ARAS), which innervates nerve fibres to the reticular formation of
the brain stem, dorsal pons and cerebral cortex, is responsible for the wakefulness (Brown et al.,
2012). A good functioning reticular formation and ARAS are essential for the maintenance of
sensibility. The cell bodies of the neurons of the reticular activating system are arranged
diffusely throughout the telencephalon. The function of this system is associated with arousal,
and its destruction produces a permanent coma (Kinney et al., 1994; Turner and Knapp, 1995).
In general, loss of sensibility is a result of irreversible or reversible dysfunction of the ARAS
and/or the cerebral hemispheres. Insensibility is defined as: “a state of unawareness (loss of
Page 33
20
consciousness) in which there is temporary or permanent disruption to brain function” (Terlouw
et al., 2016a).
There is considerable scientific evidence based on avian brain physiology to suggest that birds
exhibit sentience and awake states much like mammals (Butler et al., 2005; Rattenborg et al.,
2009). Moreover, awake birds showed similar electrical activity patterns of the brain compared
to mammals (Edelman et al., 2005). Sandercock et al. (2014) studied different reflex responses
by using electroencephalograph (EEG) activity (as a measure of brain function) in four different
clinical states of sensibility in hens and turkeys. The clinical states of sensibility they studied
were awake, sedated (drowsy), anesthetized (insensible) and deep hypnotic (insensible). Their
results revealed different reflex responses and corresponding EEG activities for the different
awake states to death.
2.2.2. Death
The traditional concept of death is defined as the end of a life due to cessation of heart beat and
respiration. A new concept of brain death was first described clinically in 1959 by two French
physicians who identified the state as "coma depasse" (Mollaret and Goulon, 1959). In literal
terms, the state of "coma depasse" is beyond the state of coma. The report of the Ad Hoc
Committee of the Harvard Medical School in 1968 elaborated on the concept of brain death and
brought awareness of brain death to a much wider audience via the “Harvard criteria”. The
Harvard criteria to establish brain death included 1. lack of receptivity and responsivity (with the
most intensely painful stimuli evoking no vocal or other response); (2) no movements (during
observation of one hour); (3) no breathing (apnea was to be confirmed during three minutes off
Page 34
21
any mechanical respirator); (4) no reflexes (emphasis being on brain stem reflexes); and (5) flat
electroencephalogram (EEG). To confirm death by the Harvard criteria, all the above findings
should show no change 24 hours later. These criteria have been accepted for confirming the
death of an individual. However, three years later two neurosurgeons suggested that irreversible
damage to the brain stem was the "point of no return” and the state of brain death in a patient
with known irreparable pathology can be established solely on clinical grounds (Mohandas and
Chou, 1971). Furthermore, Mohandas and Chaw (1971) suggested that EEG is not mandatory to
detect brain death, and they stated that presence of spinal reflexes had no bearing on the question
of brain death. These criteria are known as “Minnesota Criteria” or the “first brain stem
criterion”. After the Minnesota Criteria were released, a statement on the diagnosis of brain death
was published at the Conference of Medical Royal Colleges and their Faculties in the United
Kingdom in 1976: brain death was defined as “complete, irreversible loss of brain-stem
function” (BMJ, 1976; Lancet, 1976).
2.2.3. Pain and its mechanism
Pain often signals injury or disease in the body and produces actions to treat its causes. The
International Association for the Study of Pain defined pain in humans as "an unpleasant sensory
and emotional experience associated with potential or actual tissue damage, or described in such
terms” (IASP, 1994). Melzack and Wall (1965) proposed the theory of pain. The theory
proposed that “injury activates specific pain receptors and fibres which, in turn, project pain
impulses through a spinal pain pathway to a pain center in the brain”. International Association
for the Study of Pain defined nociception as the “encoding of noxious stimuli” (IASP, 1994). “A
stimulus that is damaging or threatens damage to normal tissues” is known as a noxious stimulus
Page 35
22
(IASP,1994). Nociception involves detection and quantitation of noxious stimuli. The process of
nociception consists of modifying and conveying that information to the brain (transmission,
modulation, and projection), and recognizing of the stimulus.
Pain may be physiologic, pathologic or neurogenic; acute or chronic; visceral or somatic
(Goldberg, 2014). Physiologic pain sensation is elicited by noxious stimulation of high-threshold
receptors, called nociceptors, in the skin that sense changes in heat, pressure and chemical
stimuli (Goldberg, 2014). Nociceptors innervate skin, muscle, fascia, joints, tendons, blood
vessels, and visceral organs. Varieties of nociceptors exist based on their response to stimuli
(Goldberg, 2014). Mechanical nociceptors selectively respond to intense mechanical stimuli, and
polymodal nociceptors respond non-selectively to noxious mechanical, thermal, and chemical
stimuli (Goldberg, 2014). These activated nociceptors send their information to the spinal cord
via two different afferent fibres: rapid myelinated A-delta fibres and slower unmyelinated C-
fibres (Basbaum et al., 2009). In the spinal cord these fibres synapse with the second-order
neurons in the dorsal horn, which either send axons into ascending sensory pathways to transmit
pain information to higher centers of the brain or serve as interneurons in segmental reflex
pathways, which allows the body to rapidly withdraw from noxious stimuli (Basbaum et al.,
2009). Ascending pathways involved in the transmission of pain include the cerebral cortex,
limbic system, and reticular activating system (Basbaum et al., 2009).
Chemically-induced pain is known as chemesthesis. The trigeminal nerve (cranial nerve V) is the
main functional component of the chemesthetic system (Bryant and Silver 2000). The trigeminal
nerve contains chemoreceptive fibers which can detect chemical irritants (Silver and Maruniak,
Page 36
23
1981). The trigeminal nerve consists of three major branches which serve the different mucosal
regions of the head: ophthalmic nerve (a sensory nerve), maxillary nerve (a sensory nerve), and
mandibular nerve (a mixed nerve, both sensory and motor). In chickens, the ophthalmic branch
innervates the frontal region, the parts of the eye, the rostrodorsal part of the nasal cavity, and is
also responsible for the motor control of eye and associates with reflexive response to irritating
stimuli to the ocular region (Mason and Clark, 2000). The maxillary nerve supplies sensory
inputs mainly to the mucosa of the conjunctiva and the palate, and the floor of the medial wall of
the nasal cavity (Mason and Clark, 2000). The mandibular branch innervates the rest of the
mouth including oral mucosa and wattles (Getty, 1975). Recently, McKeegan found that nasal
and buccal trigeminal polymodal nociceptors in chickens respond to ammonia, acetic acid vapor,
and carbon dioxide (McKeegan et al., 2003, 2005; McKeegan, 2004). McKeegan (2004) reported
that avian nociceptive threshold for CO2 is in the region of 40 - 50% CO2 in air.
It is difficult to understand animals’ experience of pain due to the absence of language. In
general, pain is detected by observing animals’ general appearance and “nocifensive” (pain-like)
behaviours; most laboratory species react in a manner to avoid and reduce the impact of acute
noxious stimuli, suggesting that they experience pain (Carstens and Moberg, 2000). There is both
physiological and behavioural evidence that domestic fowl experience pain, but they also appear
to have the ability to suppress pain by changes in motivation (Gentle, 2011). Gentle (2001)
assessed the cognitive perception of pain by a series of studies on selective attention on pain
related behaviours in chickens. The author used a sodium urate model of gouty arthritis to cause
acute gouty attacks in a single joint of a chicken, which produced a painful inflammation lasting
for 3 hrs. Nesting, feeding, exploration, and social interactions were used as the motivational
Page 37
24
changes. The author reported that the degree of pain suppression (based on expression of pain
related behaviours) ranged from hypoalgesia to complete analgesia. Therefore, shift in attention
has an ability to suppress the existing tonic pain in chickens. This phenomenon is important in
planning of different killing methods to minimize associated tonic pain.
Animal euthanasia guidelines explained bone crushing prior to insensibility is painful and should
be avoided (AVMA, 2013). Bone fractures are reported to be acutely painful in humans (Bove et
al., 2009), and other mammals (Waran et al., 2010) and in birds (Nasr et al., 2012). Mechanical
nociceptors in the periosteum of the affected bone are activated due to fracture displacement
(Freeman et al., 2008).
2.2.3.1. Anesthesia as a pain management tool
Anesthesia is mainly used during operative procedures so that the animal does not experience
pain. Anesthesia is defined as the “state in which, as a result of drug-induced insensibility the
patient neither perceives nor recalls noxious stimulation” (Roberts., 1987). There are two main
types of delivery routes for anesthetic agents: intravenous agents (generally administered
together with sedatives or narcotics), and volatile agents. Anesthetics interact with the ion
channels that regulate synaptic transmission and membrane potentials in key regions of the brain
and spinal cord (Alkire et al., 2008). Anesthetics decrease or inhibit the excitation mechanism of
the neurons (Ries and Puil, 1999). With all these mechanisms, anesthetics produce insensibility
by preventing integration (blocking the interactions among specialized brain regions) or by
reducing information (shrinking the number of activity patterns available to cortical networks)
(Alkire et al., 2009).
Page 38
25
The spinal cord is an important site of anesthetic mechanism. Neurons within the dorsal horn of
the spinal cord are involved in the transmission of stimuli to other central nervous system sites.
Anesthetics depress dorsal horn neuronal responses to noxious stimuli. Moreover, anesthetics
depress the motor neuron excitability within the spinal cord causing anesthetic-induced
immobility in response to a noxious stimulus (Collins et al., 1995).
2.2.3.2. Anesthetic depth and reflex responses
Many of the homeostatic control systems in the body are suppressed by anesthetics drugs.
Different states of anesthesia involve different suppression levels of sensibility, pain perception,
muscle tone and reflexes, which are due to the effect of the anesthetic drugs on the central
nervous system. Death can occur due to anesthetic overdose and pain can be experienced when
too light of a dose is used. For example, anesthetic agents, which have narrow safety margins,
result in death in one in 679 cases during anesthesia of healthy dogs and cats (Clarke and Hall,
1990). Therefore, maintaining minimum anesthetic depth for the particular procedure is
important, through adjusting the anesthesia. In general, anesthetized patients are insensitive to
stimuli and passage of time and do not dream (Hameroff, 2001). In animals, anesthetic drugs are
generally administered as a single injection (intraperitoneal or intramuscular) providing
analgesia, hypnosis, and muscle relaxation.
Anesthetic depth in animals is primarily assessed based on observational techniques of muscle
relaxation, reflex activities, and physiologic responses (Sandercock et al., 2014). The pedal
withdrawal reflex is commonly used to assess the depth of anesthesia in animals. If the animal
withdraws the limb in response to a toe pinch, it is assumed that the animal still experiences pain
Page 39
26
and need more anesthesia (Nevarez, 2005). The ocular reflexes are commonly used to assess the
anesthetic depth, which includes palpebral response, ocular position and corneal reflex (Nevarez,
2005; Lierz and Korbel, 2012). Sandercock et al. (2014) studied the “readily observable reflexes
and behaviours that are reliably associated with different states of sensibility” in two poultry
species (turkeys and laying hens). The clinical states of consciousness that they studied were
fully awake, semi awake (sedated), insensible–optimal (general anesthesia), and insensible–sub-
optimal (deep hypnotic state). According to their results, pupillary and nictitating membrane
reflexes and muscle tone (jaw and neck tone) were always present in sedated semi awake birds.
Jaw tone, neck tone and response to nociceptive stimuli were absent, and only pupillary and
nictitating membrane reflexes were present both in general anesthetic and deep hypnotic state.
Moreover, there were no differences in respiration rate, blood pressure and rectal temperature
between anesthetic states or between species.
2.3. Assessing insensibility and time of death
When animals are euthanized, the intention is to induce immediate, irreversible insensibility to
ensure minimum pain and distress (AVMA, 2013). Therefore, it is necessary to have a set of
criteria to assess insensibility and brain death. Indicators of sensibility must be absent, and
indicators of insensibility must be present to confirm insensibility. However, these indicators
may be different for different stunning or euthanasia techniques (physical, electrical and gaseous/
modified atmospheric). The available indicators to assess the state of sensibility in relation to
brain functions are discussed below.
Page 40
27
2.3.1. Brain stem and spinal reflexes
Brain stem reflexes are regulated by 12 pairs of cranial nerves that are not under cortical control.
The presence of central reflexes indicates functioning of the brain stem and the spinal cord, and
thus, sensibility. Insensibility or complete loss of brain function is assessed based on the absence
of brain stem reflexes (Sandercock et al., 2014; Martin et al., 2016; Erasmus et al., 2010a;
Verhoeven et al., 2015; Terlouw et al., 2016 a,b). Authors often used pupillary light reflex,
corneal reflex, and palpebral reflex to assess insensibility (Erasmus et al., 2010a; Sandercock et
al., 2014; Woolcott et al., 2018). Absence of these ocular reflexes indicates loss of sensibility.
The pupillary light reflex is tested by flashing a light into the eye and checking the pupil for
constriction. The pupillary reflex is controlled by the cranial nerve II (optic) and cranial nerve III
(oculomortor). The integration center of the pupillary reflex is located in the midbrain, near the
reticular formation. The presence of fixed dilated pupils is considered an important criterion to
evaluate the prognosis of comatose patients (Thomas, 2000). A well-functioning retina is
necessary for the activation of the pupillary reflex. The pupillary reflex is not considered a
reliable reflex of assessing sensibility during exsanguination because the blood loss can reduce
the functioning of the retina (Blackman et al., 1986). Blood covering the cornea or a damaged
eye may also reduce the feasibility of the test. Therefore, some limitations occur in assessing
pupillary reflex as an indicator of sensibility.
The other two ocular reflexes used to assess sensibility are the corneal and palpebral reflexes.
Both the corneal and palpebral reflexes are controlled by afferent cranial nerve V (trigeminal)
and efferent cranial nerve VII (facial) (Adams and Sheridan, 2008). The corneal reflex is tested
by lightly touching the cornea. This sensory information passes through the trigeminal nerve to
Page 41
28
reach the trigeminal nucleus which is located next to the reticular formation (Cruccu and
Deuschl, 2000).The trigeminal nucleus is connected to the facial nucleus to stimulate the facial
motor nerve. Due to motor responses of the facial nerve, which innervates the orbicularis oculi
muscle, the eye lid closes. The connections between the trigeminal and facial nuclei pass through
the reticular formation (Aramideh and Visser, 2002). Any interruption of this neural circuit will
cause the modification or absence of the reflex. If the corneal reflex is absent, there is a
probability of dysfunction of the reticular formation, because the neural circuit of the corneal
reflex crosses the reticular formation (Zerari-Mailly et al., 2003). Therefore, the absence of
corneal reflex is considered as a reliable indicator of insensibility (Terlouw et al., 2016).The
palpebral reflex is also tested by tapping the edge of the eye lid, which results in blinking. The
neural circuit of the palpebral reflex is similar to that of the corneal reflex. The nictitating
membrane (third eyelid) is a transparent membrane present in many birds, reptiles, amphibians,
fish, and some mammals such as rabbits. This membrane also closes in response to touching the
cornea. The neuronal circuit is similar to that described for the corneal reflex (Desmond et al.,
1983).
These brain stem reflexes provide practical means for assessing insensibility in field conditions.
However, brain stem reflexes are difficult to assess in closed or damaged eyes. In a review,
Erasmus et al. (2010a) reported that loss of pupillary light reflex is an indicator of complete
insensibility. However, the relationships between ocular reflexes and brain function are not
always clear. For example, Anil (1991) studied the effects of head-to-back electrical stunning on
the incidence of post-stunning reflex activity and cortical evoked responses in sheep and reported
that the corneal reflex and respiratory gasps were present in 10 out of 12 sheep which did not
Page 42
29
show concomitant visual and somatosensory evoked responses following a stun. Moreover, one
study with poultry showed that EEG activity ceased prior to the loss of brain stem reflexes
(Gregory and Wotton, 1990a). Sandercock et al. (2014) studied the presence or absence of brain
stem reflexes in three awake stages induced by sevoflurane, and at death, which was induced by
overdose of barbiturate in hens and turkeys. Though many of the reflexes disappeared when
birds were insensible, the nictitating membrane reflex were present even after brain death had
been confirmed by EEG based on isoelectric wave forms. The authors suggested that this may be
an artifact of the birds being anesthetized prior to death. Therefore, positive eye reflexes alone
cannot be considered as an indicator of sensibility in some cases.
The pedal or withdrawal reflex involves activation of nociceptors, and is used to assess the state
of insensibility (Erasmus et al., 2010a; Verhoeven et al., 2015a) . The pedal reflex is indicated
by withdrawal of the foot in response to pressure applied to the toes. Nose and ear prick, which
cause head withdrawal, are also used to assess sensibility. However, withdrawal reflex is difficult
to assess, especially following physical killing methods where severe body convulsions occur
(Tidswell et al., 1987).
2.3.2. Physiologic measures and behaviours
The animals’ loss of standing posture or inability to remain in an initial standing or sitting
position are used as indicators of the potential loss of sensibility (Terlouw et al., 2016). Loss of
standing posture is often the first sign of successful stunning, and indicates inability of the
cerebral cortex to control posture (Llonch et al., 2012; Raj et al., 1992) and damage to the
reticular formation (Scheepens, 2004). Both mechanical and electrical stunning should lead to
Page 43
30
immediate collapse (AVMA, 2013). However, severing of the upper spinal cord also causes
paralysis of the animal without causing brain damage. Therefore, this indicator should be
interpreted with caution (Terlouw et al., 2016). Anoxia in the brain following bleeding of non-
stunned animals results in progressive loss of standing, due to dysfunction of cortical structures
(Terlouw et al., 2016). Further, loss of posture was suggested as a sign of early stages of
insensibility (Gibson et al., 2015a; Gregory et al., 2010).
Cessation of rhythmic breathing is another indicator of loss of sensibility or death (Erasmus et
al., 2010a; Verhoeven et al., 2015). The control centers of the respiratory muscles are located in
the medulla oblongata which is located in the lower area of the brain stem (Siegel and Sapru,
2006). Inspiration and expirations are controlled by the different groups of neurons, located in
the control center (Siegel and Sapru, 2006). These neurons are stimulated by the reticular
formation that receives information from the periphery and higher brain centers (Siegel and
Sapru, 2006). It is reported that captive bolt caused axonal injuries to the brain stem and
rhythmic breathing was immediately abolished after an effective shot (Finnie et al., 2000).
Grandin (2013) suggested that the presence of rhythmic breathing after stunning is an indication
to proceed with a second stunning. The presence of rhythmic breathing after stunning is
generally accepted to indicate that an animal may not be fully insensible. Regaining rhythmic
breathing is considered as one of the first signs of recovery of sensibility after CO2 and electrical
stunning in poultry (Anastasov and Wotton, 2012).
Normal rhythmic breathing is known as eupnea. Eupnea is controlled by cranial and spinal
nerves (St-John et al., 2004). If eupnea fails, the second pattern of breathing which is called as
Page 44
31
gasping occurs. Gasping is characterized by irregular unorganized breathing which can be
induced by ischemia or hypoxia (St-John, 2009). Gasping is accompanied by guttural sounds in
many cases that can be confused with vocalizations. Progressive changing in rhythmic breathing
to irregular breathing is reported following bleeding of non-stunned animals (Verhoeven et al.,
2015). Gasping has been observed in successful nonpenetrating captive bolt stunning of turkeys
(Erasmus et al., 2010a) and calves slaughtered by bilateral severance of the common carotid
arteries and jugular veins (Blackmore et al., 1983). Terlouw et al., (2016a) suggested that
damage to the reticular formation or the medulla could be the reason for cessation of breathing in
mechanical stunning.
Convulsions (neuromuscular spams) are uncontrolled or involuntary muscle contractions of the
body which occur in electrical, mechanical and gas-stunned animals (Prinz et al., 2010; Erasmus
et al., 2010a; Raj et al., 1992). In general, neuromuscular spasms consist of clonic and tonic
phases. In birds, the clonic phase is characterized by severe wing flapping behaviour (Prinz et al.,
2010), and the tonic phase consists of extension or outstretching of legs and wings and an arched
neck (Terlouw et al., 2016b). Body movements are controlled by an integration mechanism
which involves the cerebral cortex, cerebellum, brain stem, spinal cord and the skeletal muscle
(Vander et al., 2001).
Some studies suggested that neuromuscular spams are incompatible with sensibility due to the
absence of higher motor control. Convulsions were reported in insensible birds during electrical
stunning (Cook et al., 1995) and gas stunning (Raj et al., 1992). In contrast, neuromuscular
spams occurred before insensibility in controlled atmosphere stunned broilers (Coenen et al.,
Page 45
32
2009). Gerritzen et al.(2004) observed convulsions just after loss of posture in broilers stunned
by 100% CO2, and just before loss of posture when broiler chickens were stunned using two
other gas mixtures (50% N2 +50% CO2, and 30% O2 + 40% CO2 + 30% N2). Based on the
results, the authors suggested that it is not always possible to distinguish between neuromuscular
spasms that are incompatible with sensibility and other involuntary muscle contractions
(myoclonic jerks). Moreover, Erasmus et al. (2010a) reported violent wing-flapping began
immediately after cervical crushing and cervical dislocation in turkeys, although palpebral and
corneal reflexes persisted for some time. They also reported that brain stem reflexes were
abolished immediately after blunt trauma and nonpenetrating captive bolt application while wing
flapping persisted. However, reflexes returned in some birds even when the wing flapping was
observed. Therefore, onset of convulsions cannot be used as an indicator of detecting
insensibility.
Dawson et al. (2007) reported that cessation of convulsions (at the end of tonic phase) as
measured by the accelerometer, occurs at about the same time as brain death in broiler chickens
killed by manual cervical dislocation. Further, the authors reported that cardiac relaxation
typically occurs after the convulsive phase. Based on the study of gas stunning of chickens by
Raj et al. (1992), brain death occurs at or shortly after the end of the convulsive phase. Erasmus
et al. (2010a) reported that “no reflexes or breathing returned after tonic spasms had completely
ceased” in turkeys killed with nonpenetrating captive bolts and blunt force trauma. Therefore,
they suggested that cessation of final tonic convulsions can be used as a criteria of irreversible
brain death following physical killing methods.
Page 46
33
Jaw tone and neck tone are considered as indicators of sensibility. Jaw tone is the resistance that
occurs when a force applies downward towards the lower jaw/beak. Neck tone is the ability of
the neck to hold the head upright. Both jaw tone and neck tone are controlled by cranial nerves,
and loss of neck tone and jaw tone indicate dysfunction of the brain stem (Erasmus et al., 2010a;
Martin et al., 2016). However, jaw relaxation cannot be used as a single indicator to assess
insensibility and can support other indicators of insensibility (Grandin, 2002; Gregory, 2007).
Cloacal reflex is also controlled by cranial nerves, and can be used as an indicator of brain death
(Martin et al., 2016). The external sphincter muscle of the vent contracts causing sporadic
opening and closing of the cloaca sphincter.
2.4. Euthanasia methods and mode of action
2.4.1. Physical methods
2.4.1.1. Blunt force trauma to head
Blunt trauma is defined as “the result of the impact of a body against a blunt surface, the impact
of an object with a blunt surface against a body, or a combination of both” (Merck, 2008). The
amount of force applied to an object depends on its acceleration in the space and the mass,
according to Newton’s second law: Force = mass × acceleration. The energy that the object
acquires and retains as long as it is moving is defined as kinetic energy. This energy is the force
transferred to another body when blunt trauma occurs: Kinetic energy = ½ mass × velocity. Light
objects that are accelerated to high velocity or heavy objects that impact slowly can deliver the
same kinetic energy. Therefore, the ability of an object to induce mild or severe damage depends
on its weight and velocity. The brain is a delicate organ which is surrounded by three meninges
and encased in calvarium. The calvarium and the cerebrospinal fluid layer helps to protect the
Page 47
34
brain from blunt trauma. When a blow hits a skull, most of the tissues that are damaged develop
more severe hemorrhages due to shearing and tension applied to the meningeal vessels at the site
of direct contact, and there may be contusions of soft tissues and fracture of the skull (Cors et al.,
2015).
A blow to the head is accepted as a humane euthanasia method for neonatal animals with a thin
cranium such as piglets (AVMA, 2013). The force delivered to the head should be able to cause
immediate insensibility in animals. A blow to the head needs to be performed by a well-trained
person who is aware of its esthetic implications (AVMA, 2013). European Commission (2009)
stated that this method can be applied to birds up to 5 kg in conjunction with cervical dislocation,
should not be used as a routine method, and should not be used by one person on any more than
70 animals per day.
Erasmus et al. (2010a) assessed the effectiveness of blunt force trauma in turkeys and found that
the method induced immediate insensibility leading to death. The authors further reported that
there was a difference across the age groups in failure rate, where 22% of turkey broilers (3.9 kg)
and 3% of turkey toms (13.1 kg) needed reapplication. Whiting et al. (2011) suggested the
manual blunt force trauma caused severe and fatal skull fractures in weaned piglets which is
sufficient to render them immediately insensible. However, the authors reported that there was a
24% failure rate and was esthetically displeasing to personnel. These results suggest that blunt
force trauma has a potential to induce immediate insensibility if correct force and placement on
the skull has been used, but there is a high degree of risk of failure.
Page 48
35
2.4.1.2. Cervical dislocation
Cervical dislocation is a common method for killing individual birds in the context of
commercial or non-commercial practice. EC 1099/2009 (The Council of the European Union,
2009) defines cervical dislocation as stretching and twisting of neck causing cerebral ischemia
(deprivation of glucose and oxygen supply to the brain tissues). In cervical dislocation, the skull
is separated from the vertebral column while severing the spinal cord and main blood vessels
supplying to the brain (Erasmus et al., 2010; Bader et al., 2014). Based on welfare concerns,
crushing of the vertebrae must not occur as a result of cervical dislocation (AVMA, 2013). Ideal
cervical dislocation is in between the occipital lobe of the skull and the first cervical bone
(Atlas); this causes extensive damage to the brain stem which renders the birds instantaneously
insensible due to the concussive effect that dislocation has on the brain from stretching and
severing of the spinal cord (Sparrey et al., 2014; Bader et al., 2014). Manual cervical dislocation
is performed using only the hands to cause cervical dislocation. However, cervical dislocation
must be performed by a skilled and confident operator (AVMA.2013).
Manual cervical dislocation may be esthetically displeasing to the operator (AVMA, 2007).
Additionally, operator fatigue may lead to compromised bird welfare when a large number of
birds are being killed. However, Martin et al. (2018) found no evidence of reduced performance
over time when stock people applied manual cervical dislocation to 100 birds.
Manual cervical dislocation is more difficult in mature birds than in young birds due to their
increased neck muscle tone. To address the limitations around manual cervical dislocation,
different tools have been developed to perform cervical dislocation mechanically. Some devices
Page 49
36
(e.g. killing cone and heavy stick) involve stretching, whereas others attempt to dislocate the
cervical vertebrae by forcing a blunted edge between two vertebrae (e.g. pliers) without
stretching or twisting the neck (Sparrey et al., 2014). The ultimate result should be the same as in
manual cervical dislocation (severing of spinal cord and blood vessels). However, some
mechanical cervical dislocation devices are being used without being scientifically tested for
their welfare impact.
Both manual and mechanical cervical dislocation methods have been questioned for humaneness
based on evidence that loss of sensibility is not immediate in poultry species (Gregory and
Wotton, 1990a; Sparrey et al., 2014). Erasmus et al. (2010) found that nictitating membrane
reflex persisted in turkey hens killed with cervical crushing using a burdizzo and all broiler
turkeys killed with manual cervical dislocation.
Several studies compared the efficacy of manual cervical dislocation to mechanical cervical
dislocation in poultry. Gregory and Wotton (1990a) studied cervical dislocation by manual
stretching and cervical dislocation by crushing (using Semark neck pliers) on chickens. The
authors reported that birds killed by using neck pliers exhibited longer time to loss of visual
evoked responses compared to those killed by manual cervical dislocation, and concluded that
cervical dislocation by stretching was more effective than cervical dislocation by crushing.
Martin et al. (2017) assessed a modified version of pliers on cadavers and the authors determined
that the device had poor potential to be effective given that fewer than half of the birds’ vertebrae
were dislocated while more than half had damage to trachea and vertebrae. Martin et al. (2016)
evaluated the kill efficacy of two novel mechanical poultry killing devices, the Modified Rabbit
Page 50
37
Zinger and a novel mechanical cervical dislocation gloved device (NMCD) in comparison with
manual cervical dislocation. The authors reported that NMCD was more effective than manual
cervical dislocation at killing layers and broilers of various ages (100% killing rate); birds were
rapidly rendered insensible due to extensive trauma to the brainstem and/or spinal cord and died
from cerebral ischemia due to severing of carotid arteries. The commercially available Koechner
euthanasia device (KED) was evaluated in awake broilers (Jacobs et al., 2019) and in
anesthetized turkeys (Woolcott et al., 2018b) and both papers reported shorter latency to loss of
brain stem reflexes indicative of insensibility in birds successfully killed with manual cervical
dislocation in comparison to birds killed by the KED device.
2.4.1.3. Decapitation
In decapitation, the head of the animal is completely severed from the body by using a sharp
blade in one action with rapid blood loss to leading to cerebral schemia resulting insensibility
leading to death (Mason et al., 2009). This technique is commonly practiced in small laboratory
animals to obtain brain tissues and fluid that are not contaminated with euthanizing
chemicals/agents like gases, anesthetics, and electric currents. AVMA (2013) guidelines state
that for decapitation to be acceptable, the blade must be a sharp one to ensure clean and rapid cut
at once. Derr (1991) suggested decapitation is a powerful arousal stimulus based on the evidence
of EEG in rat brains. Cutting of the throat of cattle without stunning results in pain associated
with the cut due to stimulation of nociceptors located in the neck tissue (Gibson et al., 2009;
Zulkifli et al., 2012) and distress associated with delays in the time to onset of insensibility and
with aspiration of blood into the respiratory track (Gregory et al., 2010). In decapitation,
Page 51
38
nociceptors in the neck tissue are stimulated as it damages to the skin and other tissues.
Therefore, a rapid cut with a sharp blade is essential in decapitation.
2.4.1.4. Captive bolts
Penetrating captive bolts are designed with a retractable steel bolt which penetrates the cranium
and enters the brain while firing. Direct brain damage caused by the bolt and the cerebral
concussion resulting from the kinetic energy delivered to the head are sufficient to cause the
animal to become insensible (Blackmore, 1983; Gregory et al., 2007). Kinetic energy (KE) is
proportional to the mass (m) of the bolt and its velocity (v), where the formula is KE = ½ mv2.
The velocity of the moving object has a greater effect on its kinetic energy than the mass.
Therefore, if the velocity of the bolt is reduced, its ability to effectively stun will be impaired.
2.4.1.4.1. Penetrating captive bolts
Captive bolts are commonly powered by pressurized air, cartridge, spring or elastic. Penetrating
captive bolts (PCB) are recommended as a euthanasia method for horses, ruminants, swine and
large poultry (turkeys, broiler breeders, ratites, waterfowl) with conditions (AVMA, 2013). The
conditions are personnel skill and experience, operator safety, correct position of the device and
it is also recommended that the animal is immediately exsanguinated or pithed after applying the
PCB device. However, many authors reported incomplete concussion or return to sensibility in
livestock after using PCB (Gregory et al., 2007; Gibson et al., 2012; Atkinson et al., 2013).
Sharp et al. (2014) reported that the wound caused by spring powered PCB was the full length of
the brain, from the cerebellum into the brain stem in joey cadavers. Authors suggested that
though the spring powered PCB can cause damage to a large area of the brain in joey cadavers, it
Page 52
39
is not reliable for induction of insensibility and death. Gibson et al. (2015b) studied the
mechanical factors related to the performance of PCB in cattle and concluded that appropriate
PCB/cartridge combination, the kinetic energy delivered to the head of the animal, bolt
penetration depth, and species/animal type must be considered. Other than these mechanical
factors, skull thickness is also an important factor which affects the bolt penetration depth
(Gibson et al., 2015b). Therefore, correct placement of the bolt on the head is essential in order
to deliver sufficient force to cause brain damage which helps to initiate effective stun in the
animal. The best place for the position of the device is where the skull is at its thinnest and the
brain is closest to the surface of the head. Raj and O’Callaghan (2001) reported that the
penetrative captive bolt should be fired perpendicularly to the skull to achieve effective stunning
in broilers. The authors explained that deviations of more than 20° from the right angle caused
failures in stunning.
2.4.1.4.2. Non-penetrating captive bolts
The non-penetrating captive bolts (NPCB) are designed with a flat, round or mushroom shaped
head (blunt bolt) which does not penetrate the brain. Therefore, although NPCB may be used to
stun large mammals such as cattle, slaughter weight pigs and adult sheep, it is not recommended
as a sole method of euthanasia (AVMA, 2013). Nonpenetrating captive bolts have the advantage
of consistently administering the same force to the skull independent of the strength of the
operator in comparison to manual blunt force trauma. The correct positioning of the device on
the head of the animal is important for an effective stunning. Finnie et al. (2003) tested a
cartridge powered NPCB (model MKL Karl Schermer and Co, Karlsruhe, Germany) on 4-5
weeks old lambs and 7-8 weeks old pigs. The authors reported minimal axonal injury, mild basal
subarachnoid hemorrhage and no contusional injury in pigs, and more severe axonal injuries,
Page 53
40
complicated contusions and subarachnoid and intraparenchymal hemorrhage in lambs.
Moreover, they explained that shape of the skull and thickness were the critical factors to cause
less damage in pigs than in lambs. Widowski et al. (2008) compared a prototype Zephyr
(modified pneumatic nail gun with a round bolt head, 120 psi) with manual blunt trauma on low
viability piglets (less than 24 hrs old) and concluded that manual blunt trauma was more
effective and humane than the non-penetrating captive bolt because piglets showed signs of
returning to sensibility with the Zephyr. However, Casey-Trott et al. (2013) studied the same
NPCB modified with conical nylon bolt head (115-120 psi) on neonatal piglets (less than 72 hr
old, < 3 kg) and concluded that the modified design of Zephyr-E reliably caused immediate,
sustained insensibility followed by death (in a single step) in 94% of neonatal piglets. Zephyr-E
was also effective on 3-9 weeks old piglets killing 99% in single discharge (Casey-Trott et al.,
2014).
Erasmus et al. (2010a) compared the same model of Zephyr-E as used in Casey-Trott et al.
(2013, 2014) to mechanical cervical dislocation, manual cervical dislocation and blunt force
trauma on turkeys. Their results revealed that Zephyr-E and blunt force trauma were similarly
effective in terms of time to loss of sensibility and the end time of convulsions in large turkeys.
Moreover, they concluded that the prototype Zephyr-E was more consistent than blunt trauma at
causing insensibility leading to death in small turkeys, and the Zephyr-E appeared to be a
humane on-farm killing method for cull turkeys.
The Zephyr is now commercially available from Bock Industries in several variations (Zephyr-E
and Zephyr EXL). The commercially available models of Zephyr-EXL and Turkey Euthanasia
Page 54
41
Device (TED) were evaluated in turkeys and shown to be highly effective based on rapid, almost
immediate, loss of brain stem reflexes and loss of EEG activity (Gibson et al., 2018; Woolcott et
al., 2018a).
2.4.2. Inhaled gas
Carbon dioxide is popular for euthanasia of laboratory rodents because of its relative safety, low
cost, ease of use and ability to be used as a mass euthanasia agent (Ambrose et al., 2000). The
AVMA (2013) guidelines state that CO2 should be administered by gradual-fill using a flow rate
between 10-30% chamber volume per minute (vol/min), until the animal is insensible, then the
flow rate can be increased to reduce the time to death. At room temperature and pressure, CO2 is
odorless, colorless, nonflammable and heavier than air gas. CO2 is dissolved in water according
to the following chemical reaction: CO2 + H2O ↔ H2CO3 ↔ H+ + HCO3-. This reaction can
prevent the acid-base balance in the body: pH = pK + log [HCO3- / CO2] (Henderson-Hasselbach
equation). Cellular respiration produces CO2 as a waste in animal’s metabolic tissues. CO2
diffuses into the blood capillaries from the metabolic cells and transported via blood either bound
to hemoglobin or dissolved as CO2, carbonic acid or bicarbonate iron (Baggott, 1982). A minor
amount of CO2 binds to plasma proteins to form carbamino compounds (Guais et al., 2011).
2.4.2.1. Hypercapnia and its influence
The partial pressure of CO2 (PCO2) inside the pulmonary capillary blood under normal condition
(7% or 46 mmHg) is always higher than that in the alveolar air (6% or 40 mmHg), which creates
a concentration gradient between blood and air in the alveoli. Therefore, CO2 freely diffuses via
the alveolar membrane and is released from the lungs (Guais et al., 2011). The toxic effect of
Page 55
42
CO2 can occur rapidly due to its free diffusion and is mainly associated with blood pH, heart and
central nervous system (Guais et al., 2011). Excess CO2 in the blood (hypercapnia) induces the
CO2 tension in arterial blood (PaCO2) causing acute or chronic respiratory acidosis (Guais et al.,
2011). Detection of blood CO2, O2 and pH are critical for maintaining spontaneous breathing
(Feldman et al., 2003). Although the O2 sensitive chemoreceptors are located peripheral to the
brain (peripheral chemo receptors), CO2/pH sensitive receptor sites are mainly in the brain
(central chemoreceptors). Central chemoreceptor sites contain chemosensitive neurons, and are
highly sensitive to CO2. In humans, a 1 mmHg increase in PCO2 increases ventilation rates by 20
to 30% (Feldman et al., 2003).
CO2 permeates the blood brain barrier and decreases the pH in cerebral spinal fluid (CSF) due to
increases in cellular bicarbonate and hydrogen ions (Martoft et al., 2003). CSF contains fewer
buffering components than in blood. Therefore, pH is decreases more rapidly in CSF than in
arterial blood (Guais et al., 2011). The decreased pH in CSF causes decreased nerve cell function
and cerebral electrical activity inducing anesthesia and analgesia (Lee et al., 1996). Some authors
reported mild effects to the nervous system in humans due to slightly higher concentrations of
CO2: visual impairment due to 1% of CO2 and headache due to 2% of CO2 (Jiang et al., 2005);
decrease in stereo acuity (Sun et al., 1996) and decrease in the ability to detect motion in more
than 2.5% of CO2 (Yang et al., 1997). Hypercapnia also affects cardiovascular functions. A level
of 5% CO2 in air caused increased heart rate, arterial pressure and peripheral vasodilation in
humans (Guais et al., 2011). Ventricular failure in dogs has been reported due to hypercapnia
(Stinson and Mattsson, 1970).
Page 56
43
2.4.2.2. Pain and distress associated with CO2
Distress is defined as “a negative state that develops when an organism is unable to adapt to a
stressor” (Moberg, 2000). There are many ways that CO2 can cause distress in an animal. CO2
reacts with the water molecule in the nasal mucous membranes and forms carbonic acid, which
stimulates trigeminal nociceptors and causes pain ( Leach et al., 2002; McKeegan, 2004). CO2
can also activate the nociceptors in the conjunctiva and cornea causing the sensation of burning
and stinging in humans (Feng and Simpson, 2003).
Different species detect different gasses in different concentrations in different ways. In general,
the olfactory threshold may not be aversive or cause pain. Humans can detect 10% CO2 by smell
and reported that this level was not pungent to inhale (Patterson et al., 1962). Moreover,
inhalation of 7% CO2 did not induce respiratory symptoms or irritation in humans (Patterson et
al., 1962). When humans are exposed to high CO2 concentrations breathlessness is induced and
suggested that it could be pungent (as cited in Gregory and Wotton, 1990b). Hypercapnia and
hypoxaemia results in air hunger in humans (as reviewed in Beausoleil and Mellor, 2015) and
signs of asphyxia and behavioral excitation have been observed because of both hypercapnia and
hypoxia (Forslid et al., 1986). CO2 has been identified to be painful to nasal mucosa when the
concentration is more than 65% (Hari et al., 1997). Danneman et al., (1997) also reported that
increasing CO2 was highly unpleasant at 50% and painful at 100% based in a human study. Pain
threshold level for CO2 in humans has been identified as concentrations of between 40% and
55% (Anton et al., 1992).
Page 57
44
Evidence exists that that chickens can also detect CO2 at lower concentrations that are not
necessarily aversive to them. For example, Raj and Gregory (1991) assessed hens’ behavioural
responses to feeding in chambers at 5%, 7.5%, and 10% CO2 concentrations. Authors observed
that when given a free choice, chickens avoided feeding at 7.5% and 10% CO2. However, when a
dominant hen occupied the chamber at the lower concentration, subordinate birds fed at the 7.5%
level. Dominant birds detected the levels and avoided feeding at these levels. Subordinate birds
selected to avoid the dominants birds and feed in CO2. Authors suggested that chickens could
detect 7.5% CO2 but that is was not aversive, or at least less aversive than the social pressure.
McKeegan et al., (2005) studied behavioural responses that may indicate detection of and
aversion to CO2 in chickens. Hens were exposed to short pulses of gas and their responses were
observed to indicate detection or aversion. Authors concluded hens detected CO2 at 10% as
indicated by mandibulation, orienting toward and interruption of ongoing behaviours, and
displayed head shaking at 20%. But authors further explained that behaviour threshholds were
not definitive measures of detection ability. Nasal and oral mucosa of the chicken contains
trigeminal polymodal nociceptors that can detect the presence and concentration of potentially
noxious chemical stimuli (ammonia, CO2 and acetic acid vapors) (McKeegan et al., 2004).
However, there is no olfactory response to CO2. As mentioned previously, the avian nociceptive
threshold is in the region of 40 - 50% CO2 (McKeegan et al., 2004), which is in the same range
as the electrophysiological thresholds reported for rats and humans (Anton et al., 1991, 1992).
Several behaviour studies have been conducted to assess the aversion to CO2 in poultry species.
Exposure to CO2 could cause distress to poultry because it may result in a sense of breathlessness
Page 58
45
as well as causing pain. Raj (1996) reported behaviours of head shaking, gasping and
vocalization at 72% CO2 in turkeys. Gerritzen et al. (2000) suggested that gasping and head
shaking in broiler chickens exposed to 60% CO2, and gas mixtures containing 30% and 40% CO2
were possible indicators of breathlessness and the pungency of gas mixtures. Webster and
Fletcher (2001) also observed deep breathing (breathing through the mouth) followed by head
shaking in layer chickens and broilers exposed to different CO2 concentrations (30%, 45%,
60%). These authors suggested that head shaking was an alerting response during early induction
of anesthesia due to CO2. McKeegan et al., (2005) reported respiratory disruption (apparent
increased inhalation depth and duration) in broiler chickens exposed CO2 concentrations of 10,
25, 40, 55, 70%) for 10 s. Heistand and Randall (1941) found that 6% CO2 caused slight
inhibition of breathing, whereas 10% CO2 caused increased depth of respiration in chickens.
Therefore, low levels of CO2 such as 10 % may result in increased respiratory effort but levels
over 40% levels are probably painful, possibly with increased respiratory effort combined.
2.5. Motivation, preference and aversion tests
2.5.1. Introduction
Growing interest in the ethical issues of animal care has led to a focus on the subjective feelings
of animals. Thus, during the past 30 years, welfare scientists have investigated numerous
scientific methods to understand the subjective states of animals (Dawkins, 1980). Most
scientists started to describe the importance of affective state in animal welfare (Fraser et al.,
1997; Broom, 1998) while some scientists stated that affective state is the sole criterion of
welfare (Duncan, 1996). Similarities across vertebrate species in their neuroanatomy and
physiological response to stimuli suggest a capacity for feelings in non-human animals
Page 59
46
(Dawkins, 1980). However, the direct observation of an animal’s feelings is not possible.
Cabanac (1979) stated that the correlation between reported feelings and behavioural responses
to stimuli in humans suggests that behavioural and physiological responses reflect the strength of
associated feelings.
Animals’ motivation is defined in several ways by animal welfare scientists. Simply,
‘motivation is the strength or willingness with which an animal engages in behaviour’ (Toates,
1986). In general, people associate motivation with feelings. For example, motivaton to drink is
often described as the feeling of being thirsty. Fraser and Duncan (1998) introduced the term of
motivational affective state (MAS) to refer to subjective affective states. They considered
pleasant or unpleasant experiences as the positive or negative affects leading to motivating
specific types of behaviours. They included as negative MASs states such as hunger and fear
which presume to play a role in the motivation of eating and escape, respectively, and as positive
MASs the pleasure that may accompany such behaviours as eating, playing and mating. Kirkden
and Pajor (2006) defined positive motivation as the desire to consume a commodity or perform a
behaviour, and negative motivation as the desire to avoid a painful or frightening stimulus.
Motivation is varied in strength. Studying the strength of motivation is important to determine
what animals truly want. If animals are unable to do what they are strongly motivated to do, it
can cause poor welfare (Dawkins, 1990). The term preference has been described as a difference
between the strength of motivation to obtain or avoid one resource or stimulus and the strength
of motivation to obtain or avoid another (Kirkden and Pajor, 2006).
Page 60
47
2.5.2. Approach avoidance and conditioned place avoidance paradigms
Approach avoidance paradigms are used to assess attraction or aversion to a stimulus. Animals
have a high motivation to reach food, and are capable of making the association of a specific
stimulus with food. When animals are rewarded with food inside an unconditioned environment,
they will start to associate that environment with food and the unconditioned environment will
become a conditioned stimulus. Once animals consistently choose to be in the conditioned
environment, it is possible to apply a different stimulus in the same environment and determine if
exposure to the new stimulus is neutral (the animal still enters the environment) or aversive (the
animal refuses to enter the environment).
Scientists have used approach-avoidance paradigms to evaluate the aversiveness to CO2 by
incorporating CO2 in an environment which has been conditioned with food. Webster and
Fletcher (2004) studied aversion of hens to different gas atmospheres by using an approach
avoidance paradigm. Their test apparatus consisted of two plywood chambers. One chamber was
elevated 30 cm above the other and connected by a corridor which was constructed of plywood.
Gas was injected into the lower chamber where hens associated with feed. Hens were feed
deprived to motivate them to approach food placed in the atmospheres. The number of hens that
approached and entered the lower chamber in different atmospheric conditions was observed to
detect the aversiveness of different concentrations of CO2. The authors concluded that there was
no welfare disadvantage to the use of up to 60% CO2.
Sandilands et al. (2011) examined the aversion of broiler chicks to three lethal gas mixtures at
various concentrations. In this study chicks were allowed to place their heads inside three
Page 61
48
feeding and drinking stations (FDS) in order to access food and water. Each FDS was then filled
with a different gas mixture. Aversion was assessed based on the time birds spent with their head
in each FDS (with more time indicating less aversion). The authors concluded that 60% CO2 was
highly aversive in comparison to 50% and 55% CO2 concentrations. Other studies also support
the finding that chickens given a free choice avoid a feeding chamber containing CO2. Raj and
Gregory (1991) studied 3 different CO2 concentrations (5%, 7.5%, 10%), and reported that hens
avoid the feeding chamber when the CO2 concentration was increased up to 5%. McKeegan et al.
(2006) assessed the time to cessation of feeding and withdrawal from the feeding chamber to
detect the degree of aversion in broiler chickens exposed to different CO2 concentrations (10%,
25%, 40%, 55%, 70%) for 10 s, and reported that chickens exhibited aversion to 40% and above
CO2 concentrations.
Conditioned place preference (CPP) or conditioned place aversion (CPA) paradigms are
potentially useful tools in animal welfare assessment, and mainly based on the principles of
classical (Pavlovian) conditioning (Pavlov, 1927). These paradigms measure the reinforcing
properties of a stimulus in the absence of the stimulus itself. Conditioning is done by pairing a
distinct set of environmental cues (usually in a location or a place) with a particular treatment,
and a distinctly different set of environmental cues (usually in an adjacent location or place) is
paired with a control treatment. Then, treatment and control pairings with location are repeated
several times so that the animal learns to associate the respective reinforcements or feelings
elicited by the treatments with the locations. In a post-conditioning test trial, the animal will
approach the environmental cues (location) that had positive or rewarding effects or avoid the
Page 62
49
location when the treatment had negative or aversive effects, resulting in either a conditioned
place preference or a conditioned place aversion for the treatment-paired environment.
Conditioned place aversion has been used to study the subjective experiences of animals exposed
to different gas concentrations. Ramsaya et al. (2003) studied the rewarding/aversive effects of
nitrous oxide (N2O) using the conditioning place paradigm in male Long–Evans rats. After
exposure to gas for 8 consecutive days, a conditioned place aversion was found in rats exposed
to 30% and 60% N2O. Place aversions were demonstrated during a 20-min test session on day 9
when placebo gas was delivered. Withrock (2015) and KC et al. (2016) studied aversion to three
different CO2 concentrations in goats and pigs, respectively, by using approach avoidance and
conditioned place avoidance paradigms. However, they concluded that aversion was not
sufficient to provoke a conditioned place avoidance in their experiments.
2.6. Thesis objectives
This thesis explores the efficacies of three different non-penetrating captive bolt devices
(Zephyr-E, Zephyr-EXL, and Turkey Euthanasia Device [TED]) (Bock-Industries.com) and a
novel mechanical cervical dislocation device (Koechner Euthanasia Device [KED]) (Koechner
MFG. CO., INC) as on-farm euthanasia techniques in layer chickens. Replication of previous
studies on assessment of aversiveness to CO2 in chicken using different testing protocols and
assessing different CO2 concentrations as possible euthanasia agents are important to refine
euthanasia protocols. Thus, aversion and time to loss of sensibility for four different CO2
concentrations (25%, 35%, 50%, and 70%) in layer chickens were studied.
Page 63
50
Objective 1: To assess the efficacies of the Zephyr-E, Zephyr-EXL and TED as on-farm
euthanasia methods for layer chickens.
-To assess the efficacies of Zephyr-E, Zephyr-EXL and TED for different production
stages (rearing, growing, laying and end of production) of layer chickens based on
measures of time to insensibility and death using eye reflexes and behavioural measures.
-To assess the efficacies of Zephyr-E, Zephyr-EXL and TED for different production
stages (rearing, growing, laying and end of production) of layer chicken based on post-
mortem assessment of traumatic brain injury.
-To identify behavioural and physiological indicators to confirm the death in layer
chickens before carcass disposal.
Objective 2: To assess the efficacies of mechanical cervical dislocation using Koechner
Euthanasia Device (KED model C) for three different age groups (12, 27-29, and 65-70 weeks
old) of layer chickens in comparison to manual cervical dislocation based on brain stem reflexes,
behavioural responses, physiological parameters, and post mortem pathology.
Due to ethical concerns, to avoid possibility of any pain associated with a novel killing method,
the KED was tested in anesthetized birds. Thus, the following objective also arose:
To understand the effect of anesthesia on brain stem reflexes, behavioural responses, and
physiological parameters by comparing awake versus anesthetized birds killed by manual
cervical dislocation in order to determine which measures would be valid for use in anesthetized
birds.
Page 64
51
Objective 3: Assessing CO2 as a humane killing method for poultry
- To assess the aversion for different CO2 concentrations (25%, 35%, 50%, and 70%) in
layer chickens (25- 27 weeks old) based on approach avoidance test and conditioned
place avoidance test.
- To assess the time to insensibility for different CO2 concentrations (25%, 35%, 50%, and
70%) in layer chickens (25- 27 weeks old)
Page 65
52
CHAPTER 3
Anatomical Pathology, and Behavioural and Physiological
Responses Induced by Application of Non-Penetrating Captive Bolt
Devices in Layer Chickens
This manuscript was published in Frontiers of Veterinary Science:
Bandara RMAS, Torrey S, Turner PV, Schwean-Lardner K and Widowski TM (2019).
Anatomical Pathology, Behavioral, and Physiological Responses Induced by Application of
Non-penetrating Captive Bolt Devices in Layer Chickens. Front. Vet. Sci. 6:89.
doi: 10.3389/fvets.2019.
Page 66
53
3.1. Abstract
This chapter evaluated three models of non-penetrating captive bolt devices, Zephyr-E, Zephyr-
EXL and Turkey euthanasia device (TED) for time to loss of sensibility and degree of brain
damage during euthanasia in four age groups of male and female layer chickens (10-11, 20-21,
30-35, 60-70 weeks respectively). Latencies to onset of insensibility and cardiac arrest were
assessed to detect whether killing birds via these devices was humane and effective. Both gross
and microscopic pathology evaluations were conducted to score skull and brain trauma post
mortem. All three NPCB devices induced loss of breathing, pupillary reflex and nictitating
membrane reflex within 5s after application in most chickens. Latencies to loss of jaw tone and
neck muscle tone were longer in 60-70 weeks old roosters (P<0.05). Younger birds (10-21 week-
old) demonstrated the longest time (P<0.0001) to onset of tonic convulsions, time at last
movement, cloacal relaxation and cessation of heart beat. A positive correlation (P<0.0001) was
found for all three devices between time of cardiac arrest and time to onset of tonic convulsions,
last movement, and cloacal relaxation. More than 80% of birds had skin lacerations with external
bleeding following application of all 3 devices. Device type did not affect the incidence of skull
fractures but higher skull fracture scores were noted in 10-11 week-old birds compared to other
ages. Regardless of device type and age, microscopic SDH was most apparent in the brain and
proximal spinal cord of all birds. In summary, all three devices caused significant trauma to the
midbrain and spinal cord. Results demonstrated that all three devices induce rapid insensibility
after application and can be used as a single-step method that results in a humane death in all age
groups of layer chickens.
Page 67
54
3.2. Introduction
In the poultry industry, there are several reasons for killing birds during production: to prevent
suffering from sickness or injury, for disease control, and for stock management. Therefore, on-
farm killing is a routine procedure on commercial poultry farms. Animal care guidelines for
livestock and poultry require that the methods used for routine killing cause minimal pain and
distress (NFACC, 2016). Moreover, the killing method should result in rapid and irreversible
loss of sensibility (or consciousness) to be considered humane (AVMA, 2013).
The most common method for killing poultry on farms is manual cervical dislocation which
involves stretching and separating the cervical vertebrae by hand. However, manual cervical
dislocation is considered esthetically displeasing to personnel performing it (AVMA, 2013), and
there is evidence that both manual and mechanical cervical dislocation methods may not cause
immediate unconsciousness (Erasmus et al., 2010a; Woolcott et al., 2018b). Newly designed
euthanasia devices are commercially available and, non-penetrating captive bolt devices (NPCB)
have been designed with a blunt bolt head that does not penetrate the brain. A NPCB device is
commonly used to stun large mammals such as cattle, slaughter weight pigs and adult sheep, but
is not recommended as a sole method of euthanasia for large animals, as it may not cause death
as a one-step method of euthanasia and another method is not applied animals may return to
sensibility (Finnie et al., 2003; AVMA, 2013).
The efficacy of some NPCB devices has been determined for turkeys. Erasmus et al. (2010b)
compared the prototype Zephyr-E to mechanical cervical dislocation, manual cervical dislocation
and blunt force trauma in turkeys. That study demonstrated that the prototype Zephyr-E device
Page 68
55
and blunt force trauma were more effective in terms of time to loss of sensibility in turkeys
compared to manual and mechanical cervical dislocation. These authors further suggested that a
NPCB device was more consistent than blunt force trauma at causing insensibility and death in
small turkeys. Woolcott et al. (2018a) studied two commercial models of NPCB devices:
Zephyr-EXL and Turkey Euthanasia Device (TED) on turkeys at three stages of production and
concluded that both devices were highly effective and reliable at inducing immediate
insensibility. Gibson et al. (2018) evaluated electroencephalographic (EEG) and behavioural
responses of turkeys stunned with three different NPCB devices (Cash Poultry Killer, TED, and
Zephyr EXL), and concluded that all devices were effective in causing insensibility in turkeys,
provided they were positioned correctly with the correct power load. Insensibility is often
assessed using brain stem reflexes including pupillary light and nictitating membrane reflexes in
poultry (Erasmus et al., 2010a; Sandercock et al., 2014; Martin et al., 2016). NPCB devices are
used to cause damage to the regions of the brain which control consciousness and vital functions.
Due to the acceleration force of the physical technique, cerebral contusion results with neuronal
damage and internal bleeding. Brain pathological lesions associated with hemorrhage were
associated with immediate loss of consciousness and indicative of immediate and irreversible
loss of central regulation of breathing and heart function in poultry (Erasmus et al., 2010b; Bader
et al., 2014; Woolcott et al., 2018a).
There are fewer published scientific evaluations of NPCB devices in layer chickens. Martin et al.
(2018) evaluated a cartridge-powered NPCB device (Accles and Shelvoke Cash Poultry Killer:
Model .22 CPK 200) in layer chickens, and reported 99.1% kill success with the shortest
duration to loss of brain stem reflexes compared to manual and mechanical cervical dislocation.
Page 69
56
However, the Cash Poultry Killer is cartridge- based, heavier, and more difficult to use compared
to other NPCB devices, a significant disadvantage. An evaluation of other lighter weight
commercially available pneumatically-powered NPCB devices is needed for layer chickens as an
alternative for on-farm euthanasia.
The objective of the present study was to compare the efficacy of the Zephyr-E, Zephyr-EXL,
and TED, all pneumatically powered NPCB devices, for on-farm euthanasia of four different
ages of layer chickens. Latencies to onset of insensibility and cardiac arrest were assessed as to
determine whether the techniques were humane and effective. Gross and microscopic evaluation
and scoring of the skull and brain were used to assess induced trauma.
Animal care guidelines require that death be confirmed before leaving birds and disposing of
carcasses (AVMA, 2013; NFACC, 2016). Common criteria include lack of breathing, pulse, and
cessation of the heart. Auscultation may be used to monitor heart beat but this can be difficult for
stock persons in the field as it requires a stethoscope, skill and practice. Therefore, identification
of behavioural and physiological reflexes that are correlated with cardiac arrest is needed to
readily confirm death before carcass disposal. Therefore, this study also investigated practical
behavioural and physiological indicators to confirm the death in layer chickens.
3.3. Materials and methods
The procedures and protocol for this research were reviewed and approved by the University of
Guelph Animal Care Committee (AUP 3321), which holds a Good Animal Practice certificate
issued by the Canadian Council on Animal Care.
Page 70
57
3.3.1. Animals and facilities
All chickens enrolled in this study were obtained from different research projects that were
conducted at the Arkell Poultry Research Station of the University of Guelph. All the birds had
been targeted for euthanasia by researchers or staff, because they had reached end of study or
because of routine flock depopulation. The Zephyr-E, Zephyr-EXL and TED devices were
assessed for killing efficacies and humaneness on four age groups of layer chickens: 10-11
weeks (1.1 ± 0.2 kg), 19-20 weeks (1.7 ± 0. 2 kg), 30-35 weeks (1.8 ± 0.2 kg), and 60-70 weeks
(2.2 ± 0.2 kg). Approximately 25 birds in each age group were evaluated with each of the three
devices, assessing 279 chickens in 4 different strains of White Leghorn, Brown Leghorn,
Columbian Rock, and Plymouth Rock (Table 3.1). Different age groups of birds were available
on different days. Birds were randomly assigned to a device, and a random order of application
for each device was followed on a trial day. Sex was not balanced among the treatments due to
fewer available male layer chickens. All the birds were euthanized at the Arkell Poultry Research
Station, University of Guelph.
3.3.2. Non-penetrating captive bolt devices
All three NPCB devices were commercially manufactured by Bock Industries, Inc., Philipsburg,
PA, USA. The Zephyr-E used in this study is the commercial model weighing 0.75kg. Two
models of the Zephyr-E device were used in this study: Zephyr-E-standard (Figure 3.1-A) and
Zephyr-E-layer (Figure 3.1-D). Zephyr-E consists of a modified pneumatic nail gun fitted with a
nylon head (diameter: Zephyr-E standard = 25 mm; Zephyr-E layer = 19 mm) attached to a
cylindrical metal bolt (diameter: 9.5 mm). The shape of the bolt head is conical in Zephyr-E-
standard (Figure 3.1-B) and round in Zephyr-E-layer (Figure 3.1-E). When fully extended, the
Page 71
58
bolt protrudes (Zephyr-E-standard = 19 mm; Zephyr-E-layer = 11.3 mm) past the barrel. In both
Zephyr-E devices, bolt velocity is 20 m/s, delivering 11 Joules when used at 120 psi (Bock-
Industries.com). The Zephyr-E-standard was used with standard subject adapter (Figure 3.1-C),
and the Zephyr-E-layer was used with chicken subject adapter (Figure 3.1-F).
The Zephyr-EXL is another commercially available pneumatic-powered NPCB device (Figure
3.1-G). The Zephyr-EXL uses a modified pneumatic nail gun that was fitted with a nylon head
(diameter: 25 mm) attached to a conical tipped cylindrical metal bolt (diameter: 9.5 mm) (Figure
3.1-H). The Zephyr-EXL bolt velocity is 27 m/s and delivers (Bock-Industries.com). The
Zephyr-EXL weighs 0.91 kg. The Zephyr-EXL was used with the chicken subject adapter
(Figure 3.1-I).
The Turkey Euthanasia Device (TED) (Figure 3.1-J) is a propane-powered NPCB device. The
TED consists of a gas-powered modified nail gun fitted with a flat metal head (diameter: 19.1
mm, length: 4mm) (Figure 3.1-K) attached to a cylindrical metal bolt (9.5mm) weighing 1.8 kg
(including battery and fuel cell). The TED delivers 28 Joules with a bolt velocity of 30 m/s
(Bock-Industries.com). The TED was used with adapter R3 (Figure 3.1-L). When the bolt is
fully extended, it protrudes 7.1 mm with the R-3 subject adapter.
3.3.2.1. Application of the NPCB devices
Per manufacturer instructions (Bock Industries Inc., 2016), all devices were applied
perpendicular to the top of the frontal bone just behind the comb and on a mid-line between the
eyes and ears (Figure 3.2). The Zephyr-E was set to 120 psi by connecting to a compressed air
Page 72
59
power supply (Hitachi EC 510, Hitachi Koki U.S.A., Ltd: running horse power = 0.8 kW, tank
capacity = 22.7 L, maximum pressure = 145 psi). The Zephyr-EXL was set to the 98-100 psi. For
consistency, the NPCB device operator was constant throughout all trials. Each bird was
positioned in sternal recumbency with its keel on a flat hard surface and restrained by another
researcher holding the wings gently towards the body of the bird during the application of the
device. Restraint cones were not used in order to facilitate observation of convulsions. A single
discharge was administered for each device.
3.3.3. Ante mortem assessments
The measures and procedures used for ante-mortem assessment are presented in Table 3.2. Time
to loss of brain stem reflexes (pupillary reflex and nictitating membrane reflex), jaw tone, and
neck muscle tone were assessed to detect insensibility. The parameters were assessed
immediately after device application and then at 10s intervals until cessation. Pupillary reflex
was checked for another 30 s (three times) after noting its absence to confirm brain death. If eye
reflexes and/or breathing were present more than 60 s following device application, a second
discharge was applied immediately, and the euthanasia trial was deemed a failure. Time at onset
of tonic convulsions (rigid extension of legs and neck), time at last movement, first feather
erection and cloacal relaxation following sporadic opening and closing were also recorded.
Presence and duration of heartbeat was monitored through stethoscope, and cardiac arrest was
determined to have occurred when no discernible heartbeat could be heard. All procedures to and
responses of each bird were video recorded after device application. Times to first feather
erection and cessation of all convulsions were based on blinded video recordings. Other
Page 73
60
measures were collected unblinded by live observations, and the time of each event was
reconfirmed using the video recordings.
3.3.4. Macroscopic assessment of tissue damage
Gross pathologic assessment was performed immediately after death in all chickens successfully
killed on the first attempt. Degree of external injury caused by each device was assessed (0-2
scale system) based on lacerations of the skin and presence of external hemorrhage at the site of
device application: 0=no laceration of the skin, 1=laceration of the skin with no external
bleeding, 2=laceration of the skin with external hemorrhage. Presence or absence of bleeding
from mouth and nose were also recorded as binary (yes/no) responses. Macroscopic scoring of
the degree of skull damage was based on a 0-3 scale system (Figure 3.3). Subcutaneous
hemorrhage (SCH) and subdural hemorrhage (SDH) were assessed based on a 0-4 scale (Table
3.3) (Walsh et al., 2017; Woolcott et al., 2018a). The degree of SCH was assessed by removing
the scalp and examining the amount of hemorrhage under the scalp. Then the dorsal surface of
the skull was cautiously cleaned, removing blood and tissues to examine the severity of skull
damage. Following this, the skull was lifted, and dura were removed to assess the SDH on the
brain.
3.3.4.1. Microscopic assessment of brain trauma
Following macroscopic assessment, brains and the cervical spinal cord (from 1st to 3rd cervical
bone) were collected from six randomly selected birds in each age group per device for
microscopic evaluation. Tissues were placed in 10% buffered formalin for at least 14 days before
trimming. For consistency, all trimming was performed by one individual. Three sections of the
Page 74
61
brain (A: cerebrum, B: mid brain and thalamus, C: cerebellum) and the spinal cord (C1: portion
under the first cervical bone, C2: portion under the second cervical bone, C3: portion under the
third cervical bone) were sampled. Tissue sections were embedded in paraffin, cut 4µm and
stained with hematoxylin and eosin (Animal Health Laboratory, University of Guelph) prior to
assessment. Sections were evaluated by a veterinary pathologist blinded to bird age, breed or
treatment to determine the degree of SDH and parenchymal hemorrhage (PCH) using a score
from 0 to 4: no hemorrhage (0), minimal (<5%) hemorrhage (1), mild (5-10%) hemorrhage (2),
moderate (10-30%) hemorrhage (3) and marked (>30%) hemorrhage (4) (Erasmus et al., 2010b;
Casey-Trott., 2013; Walsh et al., 2017; Woolcott et al., 2018a).
3.3.5. Statistical analyses
Statistical analyses were conducted using SAS 9.4 (SAS Institute Inc., Cary, NC, USA).
Generalized linear mixed models (GLMM) were used to analyze the fixed effects of device, age,
and their interactions on ante mortem evaluations. Least significant means separation was
conducted by using the Tukey-Kramer test.
Pearson product-moment correlation coefficients were used to determine the relationships
between the different antemortem measures for each device. Regression analysis was conducted
to establish the functional relationship between the highly correlated variables by generating the
estimates for the intercept and the regression coefficient by using REG procedure of SAS. Heart
beat end time was considered as the dependent variable (Y) and the independent variables (X)
were time to onset of tonic convulsions, time at cessation of convulsions, and time at cloacal
relaxation. The estimated intercept and the regression coefficient for each variable were used to
Page 75
62
generate the relevant fixed-effect equation to predict the value of heart beat end time for the
given level of the independent variable (cloaca contractions, cessation of convulsions and time at
onset of tonic convulsions).
Generalized linear mixed models (GLMM) with multinomial distribution and cumulative logit
link functions were used to analyze the effect of the device, age, and their interaction on
macroscopic and microscopic trauma assessments (multinomial ordinary data). Odds ratios were
computed to compare differences in the levels of fixed effects. Data from the three sections of
brain from each bird were pooled and the highest score of the three sections of each brain was
used for SDH and PCH analyses (Walsh et al., 2017, Woolcott et al., 2018a). The same
procedure was applied to the SDH and PCH analyses for the spinal cord.
3.4. Results
3.4.1. Ante mortem assessments
All three NPCB devices induced loss of breathing, pupillary reflex and nictitating membrane
reflex within 5s after application in most chickens. Overall, 100% successful killing was
observed for the TED with immediate and irreversible insensibility in all age groups. One failure
(a 29 w.o. hen) was noted of 93 birds for the Zephyr-EXL for unknown reasons. The Zephyr-E
resulted in 7 failures of 101 birds. Two failures occurred in 35 w.o. hens (one with the Zephyr-E-
standard and the other with the Zephyr-E-layer) and 5 birds were 65 w.o. (three roosters with the
Zephyr-E-standard and two hens with the Zephyr-E-layer) (Table 3.1). These birds continued to
demonstrate a pupillary light reflex, gasping, and rhythmic breathing. After 60s, the TED was
Page 76
63
applied as the second killing method. Ante mortem and pathology assessments were not
conducted for failed trials.
Clonic convulsions, with severe wing flapping and leg paddling, were observed immediately
following device application, regardless of device type or age group. Feather erection was
observed in all successfully killed birds. First feather erection was commonly observed in the
neck region, followed by intermittent feather erections in different areas of the body. Time to
onset of first feather erection was remarkably consistent between all devices and age groups
(Zephyr-E- 41±2s, Zephyr-EXL- 41±2s, TED - 43±2s, P=0.773). Gasping was observed in some
successfully killed birds for all three devices (Zephyr-E: 9 out of 94, Zephyr-EXL: 6 out of 91,
TED: 3 out of 93). The average duration ± SD duration of gasping for Zephyr-E= 51±28s,
Zephyr-EXL=29±17s and TED= 16±2s. Two birds killed with the Zephyr-EXL started gasping
at 50s and 90s after the application of device while others started at 10s after application of the
device. None of these birds demonstrated concurrent pupillary or nictitating eye reflexes.
A summary of ante mortem responses is shown in Table 3.4. Both time to loss of neck tone and
time to loss of jaw tone showed a difference for device, age, and device by age interaction
(P<0.05). The longest times to loss of jaw tone (31±3s) and neck muscle tone (28±3s) were
observed in 60-70 w.o. roosters. The TED had the longest times (P<0.05) to loss of jaw tone
(28±3s) and neck muscle tone (36±6s). Overall, time to loss of jaw tone and neck muscle tone
were longer in 60-70 w.o. roosters for TED (39±6s and 53±9s respectively, P<0.05). Younger
birds (10-11 week and 20-21 w.o.) showed longest times (P<0.0001) for onset of tonic
convulsions and cloacal relaxation (Table 3.4). A device effect was not observed for times to
Page 77
64
onset of tonic convulsions or last movement. Onset of tonic convulsions was easily observed
(stretched neck and legs with sudden feather erection) in 10-11 w.o. birds compared to the older
groups. A longer time to cloacal relaxation (P<0.05) was observed for the TED (186±24s)
compared to the Zephyr-E (167±24s), and no difference was found between the TED and the
Zephyr-EXL. Some birds defecated during cloacal relaxation regardless of the device. As the last
measure, longest time for cessation of heart beat (P<0.0001) was observed in younger birds (10-
11 w.o.: 235±26s). The shortest times for cloacal relaxation (P=0.033) and cessation of heart beat
(P=0.035) were recorded for the Zephyr-E.
The Pearson product-moment correlation coefficient was computed to assess the relationship
between the ante mortem measures for each device. Table 3.5 summarizes the results. Overall,
there were strong, positive significant correlations between time to cardiac arrest and time to
cloacal relaxation (Zephyr-E: r = 0.935, Zephyr-EXL: r = 0.906, TED: r = 0.906), time to last
movement (Zephyr-E: r = 0.951, Zephyr-EXL: r =0.934, TED: r = 0.948), and time to onset of
tonic convulsions (Zephyr-E: r = 0.917, Zephyr-EXL: r =0.856, TED: r =0.808). There was a
positive relationship between times to cardiac arrest and loss of neck muscle tone (r = 0.34, P=
0.0008) and between cardiac arrest and loss of jaw tone (r = 0.30, P=0.003) for TED. However,
correlations between cardiac arrest versus loss of jaw tone or loss of neck muscle tone were not
significant for the Zephyr-E and Zephyr-EXL devices. Loss of jaw tone and loss of neck muscle
tone showed a positive (P<0.0001) correlation for all three devices. Strong positive correlations
(P=0.0001) were also found between onset of tonic convulsions and cloacal contraction, onset of
tonic convulsions and time to last movement, and cloacal relaxation and time to last movement
Page 78
65
for all three devices (Table 3.5). Feather erection was not associated with any other measure for
any of the devices evaluated.
Regression analysis was conducted for the highly correlated variables considering time to cardiac
arrest as a dependent variable. Regression equations and relevant coefficient of determinations
(R2) are presented in Table 3.6.
3.4.2. Pathology evaluations
3.4.2.1. Macroscopic evaluation
The degree of external damage (external hemorrhage and skin lacerations) was not different
among the devices (P=0.595) or; age groups (P=0.062), nor was there an interaction (P = 0.689).
Skin lesions with external bleeding were noted in >80% of birds for all 3 devices (Zephyr-E:
80.6%, Zephyr-EXL: 85.8%, TED: 82.8%) and more than 70% in all age groups (10-11 w.o.:
85.3%, 20-21 w.o.: 72.2%, 30-35 w.o.; 81.3%, 60-70 w.o.: 90.4%). No external damage was
observed in a few birds killed by the different devices (Zephyr-E: 11.8%, Zephyr-EXL: 4.3%,
and TED: 12.9%) and in the different age groups (10-11 w.o.: 5.3%, 20-21 w.o.:20.3%, 30-35
w.o.;12 %, 60-65 w.o.:11.1%). Some birds showed both nasal and mouth bleeding (Zephyr-
E=7/94, Zephyr-EXL=3/92, TED=0/93) and protruding eyes with the cornea covered with blood
(Zephyr-E=10/94, Zephyr-EXL=11/92, TED=18/93).
Macroscopic lesion scores are presented in Table 3.7. All birds had subcutaneous hemorrhage on
the skull regardless of the device and age. The highest score of 4 was observed in 99% of birds in
the 10-11 w.o. group. Figure 3.4 shows two skulls with 25% and 100% macroscopic
Page 79
66
subcutaneous hemorrhage. Macroscopic subcutaneous hemorrhage found on the skull was
different among the devices (P<0.0001), age (P<0.0001) and device by age interaction
(P=0.029). Lower SCH scores were noted to occur 3.6 times more in birds killed with the
Zephyr-E than the TED and 0.1 times more likely for the Zephyr-EXL than the TED. No
difference was found between the Zephyr-E and Zephyr-EXL.
Skull fracture scores differed among the age groups (P=0.0001) and there was an age by device
interaction (P=0.0017). Skull fractures were common in the 10-11 w.o. group and more than
98% of birds of this age had penetrating fractures from all devices. Birds in the 20-21 and 60-70
w.o. groups had lower fracture scores than 11 w.o. birds. No difference was found between the
10-11 w.o. vs 30-35 w.o., and 20-21w.o. birds vs 60-70 w.o. birds. In addition, there was a 0.4
times greater chance of lower scores in 60-70 w.o. birds than in 30-35 w.o. birds. Some birds in
the 60-70 w.o. group did not have any skull fractures (Zephyr-E=4%, Zephyr- EXL=29%, TED=
20%). An age difference was found only for TED and not for the Zephyr devices.
Subdural macroscopic hemorrhage on the brain was substantial and 100% of birds had a score of
1 or more for all three devices (Table 3.7). Over 85% of birds had a score 2 or above for all
devices. The highest score of 4 was observed in 12% of birds killed by the Zephyr-E, 21% of
birds killed by the Zephyr-EXL and 31% of birds killed by the TED. Figure 3.4 shows one brain
covered by less than 25%, and another covered by 50-75% of subdural hemorrhage. Subdural
macroscopic dorsal hemorrhage was affected by device (P<0.0001), age (P<0.0001), and device
x age interaction (P=0.020, Table 3.7). The Zephyr-E generally resulted in lower scores
compared to the TED or Zephyr-EXL. There was no difference in degree of subdural
Page 80
67
hemorrhage between the TED and Zephyr-EXL. Birds at 10-11 w.o. generally had higher injury
scores than all the other age groups. A higher chance of having higher injury scores was
observed in birds at 20-21 w.o. than in birds at 30-35 w.o. Birds at 30-35 w.o. had 1.8 times
higher chance of having lower trauma scores than birds at 60-70 w.o.
3.4.2.2. Microscopic evaluation
Table 3.8 provides a summary for microscopic scores for subdural (SDH) and parenchymal (PH)
hemorrhage in three brain sections (cerebrum, mid brain, and cerebellum) killed with different
NPCB devices. SDH was observed in the cerebrum of all 24 birds killed by the TED, but only
some birds killed by Zephyr-E and Zephyr-EXL. Similarly, PCH was absent in the cerebrum of
some birds killed by Zephyr-E (n=4), Zephyr-EXL (n=3) and TED (n=1). SDH was substantial
in the midbrain and observed in all assessed birds killed by all three devices. PCH was also
found in the mid brains of all birds killed by Zephyr-EXL and TED except one bird killed by
Zephyr-E. All birds had SDH in the hind brain except one bird killed by Zephyr-EXL and TED.
Nearly 50% of birds did not have PCH in the hind brain when killed by the Zephyr-E (n=11),
Zephyr-EXL (n=7) and TED (n=12). PCH was absent in the hind brain of all 6 birds in 10-11
w.o. group. Four of six birds killed by the TED and one of six killed by the Zephyr-EXL in 10 -
11 w.o. group did not have hind brain PCH. Overall all three devices caused the highest degree
of trauma to the mid brain.
There were no device, age or their interaction effects on SDH in the brain or spinal cord (Table
3.9). However, SDH was observed in both the brain and spinal cord for all three devices in all
age groups. Similarly, PCH was observed in the brain in all ages for all three devices, but there
Page 81
68
was a device effect (P=0.024). Birds killed with the Zephyr-EXL had a greater chance of having
higher scores for PCH in the brain than those killed with the Zephyr-E or TED. Overall, 46% of
birds received a score of 4 for PCH when killed with Zephyr-EXL, compared to 8% for the TED
and 29% for the Zephyr-E. There was no difference between the TED and Zephyr-E. In contrast
to brain PCH, a device effect was not observed for PCH in the spinal cord (P=0.182). However,
PCH in the spinal cord was different (P=0.044) among the age groups and showed an age by
device interaction (P=0.021). Birds in the 60 w.o. group had a greater chance of having higher
scores for PCH in the spinal cord than birds in the 10-11or 30-35- w.o. groups.
3.5. Discussion
To the author’s knowledge, this is the first study to evaluate the commercially available
pneumatic NPCB devices (Zephyr-E, Zephyr-EXL, TED) for on-farm killing of layer chickens.
Our results demonstrated that all three devices were similarly effective at inducing insensibility
and causing death in all age groups.
All devices caused loss of pupillary light reflex, nictitating membrane reflex, and breathing
within 5s after application. Direct or indirect (contrecoup) trauma to the brain can result in
impairment of brain stem reflexes (White and Krause, 1993; Drew and Drew, 2004). The area
that controls the pupillary light reflex (cranial nerve II and III) is located in the mid brain. The
nictitating membrane reflex is controlled by both cranial nerve III (located in the mid brain) and
cranial nerve V (located in the pons). It is possible that direct damage to the corresponding
cranial nerves caused loss of these reflexes. However, the substantial microscopic parenchymal
hemorrhage (PCH) demonstrated that all three devices caused severe trauma to the mid brain.
Page 82
69
The medulla oblongata in the hind brain contains important regions that regulate the
cardiovascular and respiratory systems. The control centers of jaw tone and neck muscle tone are
also located in the hind brain. The majority of birds had microscopic SDH (99%) and PCH (more
than 50%) in the hind brain. The damage caused by the devices to the hind brain was enough to
cause impairment of breathing. Jaw tone and neck muscle tone disappeared in less than 20s and
50s respectively, in all birds indicating that all three devices caused hind brain damage. Overall,
all three devices disrupted brain function and caused rapid brain death.
Sandercock et al. (2004) reported that jaw tone and neck muscle tone were the most reliable
reflexes distinguishing between sensible and insensible states in poultry based on EEG studies.
Loss of jaw tone has been used as an indicator of loss of sensibility in poultry under field
conditions (Erasmus et al., 2010a; Martin et al., 2016). All birds in our study showed jaw tone
and neck muscle tone for a few seconds following application of all three devices, and muscle
tone did not disappear as quickly as did the eye reflexes. Martin et al. (2016) also reported longer
time to loss of jaw tone (21.7s) than pupillary reflex (11.6s) in chickens killed with a penetrating
captive bolt (Modified rabbit zinger). Results of the present study revealed longer time to loss of
jaw tone and neck muscle tone for TED compared to the Zephyr devices, which corresponded to
differences in PCH scores in the hind brain between devices. Moreover, the shortest time for
cloacal relaxation and cardiac arrest, two responses controlled by the hind brain, were recorded
for Zephyr-E. We suggest that the force caused to the hind brain by the TED is different
compared to the Zephyr devices. Differences between the TED and Zephyr devices could be due
to the shape of the bolt heads which deliver force differentially across and through the skull; the
TED has a flat bolt head and the Zephyr devices have either round or conical bolt head.
Page 83
70
Erasmus et al. (2010a) did not report any gasping in turkeys effectively stunned with the
Zephyr-E or blunt force trauma. In contrast, a few successfully killed birds (~10%) showed
gasping following application all three devices nearly for 60s. However, their eye reflexes were
completely absent. In the current study, paroxysmal opening of the beak without any chest
movement associated with breathing was recorded as gasping. Gasping is not indicative of
sensibility and can be present in the absence of auditory evoked potentials (Cors et al., 2015).
Therefore, gasping can be observed both in awake and insensible birds, and while unpleasant to
watch, is not necessarily indicative of device failure.
Seven birds failed to lose brain stem reflexes and breathing within 60s for Zephyr-E. Four of
them were killed with the Zephyr-E standard with a common subject adapter and 3 with the
Zephyr-E layer with the chicken subject adapter. As the failures with the common subject
adapter came first, we considered that the adapter was the cause of the failures. Then the device
was switched to Zephyr-E-layer with a chicken subject adapter. The chicken subject adapter
allows for a better alignment of the device around the comb of the bird. For the Zephyr-E and
Zephyr EXL the bolt velocity is adjustable as a function of air pressure. The pressure in the air
compressor was slightly lower than 120 psi when killing these seven birds, and we suspect this
could be a cause of the failed euthanasia. Based on this finding, we recommend that the Zephyr-
E be used exactly at 120 psi in layer chickens of all ages and weights.
Determining reliable indicators of irreversible brain injury is important for detecting clinical
death under field conditions since animal care guidelines require confirmation of death before
birds are disposed (NFACC, 2016). We studied times to onset of tonic convulsions, last
Page 84
71
movement, and cloacal relaxation since these are readily observed indicators that can be used to
confirm the irreversible brain injury. Tonic convulsions were observed in all successfully killed
birds regardless of the device and thus tonic convulsions can be used to indicate a successful
euthanasia. Sudden feather erection was observed first around 60s after device application and
also during the tonic phase in all birds killed successfully. Gerritzen et al. (2007) confirmed the
death of poultry killed with CO2 based on sudden feather erection along with occurrence of tonic
convulsions followed by complete muscle relaxation. Heard (2000) stated that sudden feather
erection during anesthesia of birds is indicative of cardiac arrest. However, in the current study,
time at first feather erection was not correlated with cessation of heart beat after killing with two
of the three devices, and cannot be considered a reliable indicator of time at cardiac arrest.
Similar results were determined by Hernandez et al. (2018) who also found that feather erection
was not associated with cardiac arrest or isoelectric point using EEG. Cessation of movement has
been used to estimate irreversible brain death (Erasmus et al., 2010a; Dawson et al., 2007;
Gerritzen et al., 2007). Time to last movement in the present study was highly correlated with
time of cardiac arrest, thereby serving as a reliable on-farm indicator of cardiac arrest. Cloacal
relaxation was the last reflex observed before cardiac arrest in all birds, which agrees with
Martin et al. (2016), and demonstrates its utility as a conservative indicator of death. Cardiac
arrest typically occurs after all motion has ceased (Dawson et al., 2007; Turner et al., 2012). This
coincides with the results in present study as all the birds ceased heart beat after cessation of all
movements and reflexes. The presence of a heart beat does not indicate sensibility, and is, itself,
a conservative measure. Turner et al. (2012) also reported the presence of a heartbeat in poultry
several minutes after brain death, as confirmed by use of an EEG. Auscultation of the heart with
a stethoscope can be difficult under the field conditions so it can be difficult to be certain of the
Page 85
72
exact moment of cardiac arrest. Additionally, the heart may continue to beat irregularly for some
time (Hernandez, 2018). Onset of tonic convulsions, last movement, and cloacal contractions can
be visually observed in the field. The relationship analysis in this study suggest that cardiac
arrest was highly positively correlated with onset of tonic convulsions, time to last movement,
and cloacal relaxation for all three NPCB devices and thus, onset of tonic convulsions, last
movement, and cloacal relaxation can be used by stock persons to make accurate decisions of
successful euthanasia under field conditions.
Younger birds (10-11 w.o. and 20-21 w.o.) had a longer latency to onset of tonic convulsions,
last movement, cloacal relaxation, and cardiac arrest. In addition, one, four and six of the six 10-
11 weeks old birds killed by the Zephyr-EXL, TED and Zephyr-E, respectively, had no PCH in
the hind brain, indicating less trauma to the hind brain. This might explain the longer latencies to
onset of tonic convulsions, last movement, cloacal relaxation, and cessation of heart beat in the
10-11 w.o. group. The oldest birds in the 60-70 w.o. group, with more mature anatomy (fused
and larger size skull, large comb) experienced a longer time to loss of jaw tone and neck muscle
tone. Therefore, the placement of the device on the head (the place of the skull where the bolt
hit), device configurations and anatomic structure of the head may have affected the degree of
damage cause to different regions of the brain.
Overall more than 80% birds showed external damage for all three devices. Some birds had
mouth and nose bleeding, and damaged eyes. External bleeding is important for biosecurity
measures and aesthetic concerns. The fine balance between effectiveness and aesthetics is the
key to selection of an appropriate method of euthanasia. Higher external damage caused by the
Page 86
73
NPCB devices may indicate a need for lower air pressure levels for the Zephyr-EXL based on
the age group. However, the Zephyr-E should only be used with the manufacturer-recommended
120 psi to minimize the chance of failures.
Results of the macroscopic pathology assessment indicated that lower subcutaneous and subdural
hemorrhage scores were more likely for the Zephyr-E. This may be due to lower force generated
by the Zephyr-E than for the Zephyr-EXL and TED. All three devices caused penetrating
fractures with no embedded fragments in most birds. Skull fractures are highly associated with
severe traumatic brain injuries leading to death in humans (Tseng et al., 2011). Studies in other
animal species have reported that skull fractures are often present in animals that were
effectively stunned with non-penetrating captive bolt devices (Finnie et al., 2000; Casey-Trott et
al., 2013). However, Erasmus et al. (2010b) reported that presence or absence of skull fractures
did not affect onset of insensibility in turkeys killed by NPCB or blunt trauma. Our results also
showed that presence or absence of skull fractures did not influence the effectiveness of inducing
insensibility and irreversible brain death. Overall, 15 birds in our study had an intact skull, and
all of them showed rapid loss of eye reflexes and irreversible brain death.
There was no difference found for the microscopic assessment of subdural hemorrhage in brain
sections among the devices. A device effect was found for PCH in that the Zephyr-EXL (at 98-
100 psi) caused higher scores of PCH in the brain than that seen with the TED or Zephyr-E. PCH
indicates traumatic brain injuries (TBI). Researchers have suggested that immediate insensibility
and irreversible loss of vital functions are associated with subdural and parenchymal hemorrhage
in poultry (Erasmus et al., 2010b; Bader et al., 2014). Results from all three devices evaluated in
Page 87
74
this study confirm this. It is important to note that all three devices also caused SDH and PCH in
the cervical area of the spinal cord. Therefore, the force generated by all three devices is
sufficient to cause intensive traumatic damage to the brain and to the cervical portion of the
spinal cord.
An unexpected problem was encountered with the Zephyr devices for Plymouth Barred Rock
chickens. In two chickens of this strain, the feathers became stuck between the bolt and the
adapter for both Zephyr-E and Zephyr-EXL. Despite this, the birds were both killed effectively
with these devices. We suggest that plumage type should be considered when using Zephyr
devices with chicken subject adapter.
3.6. Conclusion
This study demonstrated that brain trauma cause by all three NPCB devices was sufficient to
rapidly render the birds insensible, leading to irreversible brain death in all age groups of layer
chickens. The Zephyr-E, Zephyr-EXL, and TED devices can be used as a humane single-step
euthanasia method for layer chickens. Additionally, we suggest onset of tonic convulsions, last
movement, and final cloacal relaxation are good indicators of clinical death in layer chickens in
field conditions.
Page 88
75
Table 3. 1 : List of number of birds killed with the different NPCB devices by age group, strain,
body weight, and sex. Number of failed birds are indicated in parentheses.
Age
(weeks)
Device Body wt
(kg)
Strain Male Female Total
10-11 Z-E-std 1.1 ± 0.2 White Leghorn 13 12 25
Z-EXL 1.1 ± 0.2 White Leghorn 13 12 25
TED 1.1 ± 0.2 White Leghorn 13 25 25
20-21 Z-E-std 1.5 ± 0.2 White Leghorn 1 1 2
Z-E-layer 1.7 ± 0.2 White Leghorn 2 12
1.9 ± 0.3 Plymouth Rock 1 2 17
Z-EXL 1.6 ± 0.1 White Leghorn 3 8
1.9 ± 0.3 Plymouth Rock 3 4 18
TED 1.7 ± 0.2 White Leghorn 3 12
2.2 ± 0.9 Brown Leghorn 2 0
1.8 Plymouth Rock 0 1 18
30-35 Z-E-std 1.8 ± 0.2 White Leghorn 0 12+(1) 13
Z-E-layer 1.9 ± 0.2 Brown Leghorn 0 10+(1)
2.0 ± 0.1 Columbian Rock 0 3 14
Zephyr-EXL 1.7 ± 0.2 White Leghorn 0 12
1.9 ± 0.1 Brown Leghorn 0 4
1.8 ± 0.1 Columbian Rock 0 9 25
TED 1.6 ± 0.2 White Leghorn 0 12
1.9 ± 0.1 Brown Leghorn 0 6
1.8 ± 0.2 Columbian Rock 0 7 25
60-75 Z-E-std 2.2 ± 0.1 White Leghorn 2+(3) 0
2.4 ± 0.4 Brown Leghorn 0 4
1.8 ± 0.1 Columbian Rock 0 3
2.2 ± 0.1 Plymouth Rock 0 2 14
Z-E-layer 2.4 ± 0.3 Brown Leghorn 0 8+(2)
2.1 Columbian Rock 0 1
2.3 ± 0.1 Plymouth Rock 0 5 16
Zephyr-EXL 2.2 ± 0.1 White Leghorn 14+(1) 0
2.4 ± 0.7 Brown Leghorn 0 2
2.2 ± 0.2 Columbian Rock 0 7
1.9 Plymouth Rock 0 1 25
TED 2.0 ± 0.1 White Leghorn 14 0
2.2 ± 0.2 Brown Leghorn 0 6
2.1 ± 0.7 Columbian Rock 0 3
2.4 ± 0.1 Plymouth Rock 0 2 25
Z-E-std=Zephyr-E with standard subject adapter and conical shape bolt head.
Z-E-layer= Zephyr-E with chicken subject adapter and round shape bolt head.
TED= Turkey Euthanasia Device.
Page 89
76
Table 3. 2 : Ante-mortem assessment measures, descriptions, and procedures used, listed in
order of observation after application of each killing method
Measures Description Procedure
Pupillary light reflex Constriction of the pupil in response to
light
Light from a medical penlight
was directed into the eye and
pupil constriction was
examined
Nictitating
membrane reflex
Transient closure of the nictitating
membrane in response to mechanical
stimulation
The medial canthus of the eye
or the cornea was lightly
touched with a fingertip
Jaw tone Resistance to downward pressure
applied to the jaw
Gentle pressure was applied
to the lower jaw with a finger
Neck muscle tension Change in neck muscle tone or
movement of the head when the neck
is lifted
The neck was lifted with the
fingers of one hand
Gasping Paroxysmal opening of the beak Visual observation for paroxysmal opening of the
beak
Feather erection
Sudden erection of feathers, not in
response to external stimuli
Visual observation of sudden
feather erection on some part
of the body
Tonic convulsions Muscle rigidity with the legs and
wings outstretched
Visual observation of the
time of onset of legs and neck
outstretched
Cloacal relaxation
Cloaca opening following
contractions of cloaca
Visual observation for cloaca
opening following
contractions
Cardiac arrest Cessation of heart beat
Auscultation by using a
stethoscope
Breathing Rhythmic inhalation and exhalation Visual observation for
rhythmic movement of the
chest area
Page 90
77
Table 3. 3: Gross and microscopic pathology scoring criteria for macroscopic, and microscopic
hemorrhage
Score Macroscopic Microscopic
Subcutaneous or subdural hemorrhage
Subdural or parenchymal hemorrhage
0 None None
1 <25% of surface area Minimal (<5% of section)
2 26–50% of surface area Mild (5–10% of section)
3 51–75% of surface area Moderate (11–30% of section)
4 76–100% of surface area Marked (>30% of section)
Page 91
78
Table 3. 4: Mean time (± SE, s) to onset of specific measures after application of different
NPCB devices in different age groups of layer chickens
Measure Age
(weeks)
Device1 All
devices
P value
Z-E Z-EXL TED Device Age Device*Age
Loss of jaw <0.0001 <0.0001 0.0063
tone 10-11 13±2bc 22±2bc 25±6b 19±3b 20-21 21±3bc 21±2bc 29±6b 23±3b 30-35 20±3bc 19±2bc 24±6bc 21±3b 60-70 28±3b 27±2b 39±6a 31±3a All ages 21±3b 22±3b 28±3a Loss of neck <.0.0001 <0.0001 0.0297
muscle tone 10-11 18±4e 23±5de 32±9dc 24±6c
20-21 25±4dce 23±5de 33±10dc 26±6b
30-35 27±4dc 29±5dc 33±10dc 30±6b
60-70 35±4bc 44±5ab 53±9a 46±6a
All ages 27±6b 30±6b 36±6a
Time at first 0.7733 0.2154 0.0546
feather 10-11 38±2
erection 20-21 45±2
30-35 39±2
60-70 44±2
All ages 41±2 41±2 43±2
Onset of 0.2909 <0.0001 0.8839
tonic 10-11 153±22a
20-21 141±23a
30-35 111±23b
60-70 103±22b
All ages 122±22 126±22 132±22
Last 0.1042 <0.0001 0.9026
movement 10-11 206±25a
20-21 189±25a
30-35 146±25b
60-70 147±25b
All ages 164±25 171±25 181±25
Cloacal 0.0335 <0.0001 0.8349
relaxation 10-11 203±24a
20-21 199±25a
30-35 153±25b
60-70 150±24b
All ages 167±24b 175±24ab 186±24a
Cessation of 0.0354 <0.0001 0.8866
heart beat 10-11 235±26a
20-21 209±27b
30-35 173±27c
60-70 178±26c
All ages 189±26b 198±26ab 209±26a
Different letters indicate statistically significant (P<0.05) differences between the comparisons
Bold numbers indicate significance results (P<0.05)
1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device
Page 92
79
Table 3. 5: Pearson correlation coefficients to assess the relationship between the antemortem
measures for different NPCB devices for all ages of layer females and males (n = 94 for Zephyr
E, n=92 for Zephyr EXL and n=93 for TED)
Variable Device1 Loss of
neck
muscle
tone
Loss of jaw
tone
First
feather
erection
Onset of
Tonic
convulsions
Cloacal
relaxations
Last
movement
Cessation of
heart
Z-E -0.1082
p=0.2991
-0.1056
p=0.3111
0.1088
p=0.296
0.9174
p<0.0001
0.9352
p<0.0001
0.9519
p<0.0001
beat Z-EXL 0.0917
p=0.387
0.0723
p=0.495
0.0703
p=0.5078
0.8566
p<0.0001
0.9067
p<0.0001
0.9342
p<0.0001
TED 0.3422
p=0.0008
0.3003
p=0.0034
0.2065
p=0.047
0.8081
p<0.0001
0.9068
p<0.0001
0.9482 p<0.0001
Loss of neck
muscle tone
Z-E 0.7948
p<0.001
-0.0744
p=0.475
-0.2007
p=0.052
-0.1003
p=0.3383
-0.1622
p=0.1183
Z-EXL - 0.7089
p<0.0001
0.2021
p=0.054
-0.026
p=0.8047
0.058
p=0.5912
0.0462
p=0.663
TED 0.7660
p<0.0001
0.2971
p=0.0038
0.2371
p=0.0221
0.2134
p=0.0399
0.2832
p=0.0059
Loss of jaw
tone
Z-E -0.1461
p=0.1599
-0.1467
p=0.1583
-0.0639
p=0.5427
-0.1299
p=0.2119
Z-EXL - -0.1128
p=0.286
-0.036
p=0.7289
0.0259
p=0.8102
-0.0105
p=0.9212
TED 0.1880
p=0.0711
0.2371
p=0.0221
0.2138
p=0.0396
0.2763
p=0.0073
First
feather
Z-E 0.1148
p=0.2705
0.1903
p=0.067
0.1235
p=0.2374
erection Z-EXL - 0.0251
p=0.8128
0.0487
p=0.6522
0.0894
p=0.3989
TED 0.0588
p=0.5755
0.1013
p=0.333
0.1914
p=0.066
Onset of
Tonic
Z-E 0.9341
p<0.0001
0.9519
p=0.0001
Z-EXL - 0.9049
p<0.0001
0.9056
P<0.0001
TED 0.7915
p<0.0001
0.8195
p<0.0001
Cloacal
relaxation
Z-E 0.960
p<0.0001
Z-EXL - 0.94877
p=<0.0001
TED 0.9428
p<0.0001 1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device.
Bolded numbers indicate significant results (P < 0.05)
Page 93
80
Table 3. 6: Regression and relative contribution (R2) for response of dependent variable (Y) for
independent variables (X) of different NPCB devices
Independent
variable (X)
Dependent
variable
(Y)
Device1 Regression
equation
Coefficient of
determination (R2)
Time at onset Heart beat Z-E Y= 35.77 + 1.1X 0.8416
of tonic
convulsions
end time Z-EXL Y= 57.15 + 1.1X 0.7339
TED Y= 84.52 + 0.9X 0.6532
Time at cloacal Heart beat Z-E Y= 19.1 + 1X 0.8746
relaxation end time Z-EXL Y= 33.6 + 1X 0.8222
TED Y= 26.2 + 1X 0.8224
Time at last Heart beat Z-E Y= 24.6 + 1X 0.9062
movement end time Z-EXL Y= 40.5 + 0.9 X 0.8728
TED Y= 34.6 + 1X 0.8991
All regression coefficients were significant (P < 0.05). 1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device.
Page 94
81
Table 3. 7: Summary of gross scores for subcutaneous hemorrhage, skull fractures, and subdural
hemorrhage in birds killed by different NPCB devices. Number of birds with each score are
indicated.
Bold numbers indicate significance (P<0.05). 1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device.
SCH= subcutaneous hemorrhage, SDH= Subdural hemorrhage
Device1 Age Number of birds with gross score P-value
0 1 2 3 4 Total Device Age Device*
Age
SCH <0.0001 <0.0001 0.0295
Z-E 10-11 0 0 0 2 23 25
20-21 0 3 5 4 7 19
30-35 0 5 12 6 2 25
60-70 0 3 2 10 10 25
Z-EXL 10-11 0 0 0 4 21 25
20-21 0 2 2 7 7 18
30-35 0 2 15 5 3 25
60-70 0 7 7 4 6 24
TED 10-11 0 0 1 2 22 25
20-21 0 0 0 1 17 18
30-35 0 2 3 9 11 25
60-70 0 1 4 6 14 25
Skull 0.1485 0.0001 0.0017
fractures Z-E 10-11 0 0 20 5 N/A 25
20-21 1 4 11 3 N/A 19
30-35 0 3 19 3 N/A 25
60-70 1 2 13 9 N/A 25
Z-EXL 10-11 0 2 14 9 N/A 25
20-21 0 2 11 5 N/A 18
30-35 0 0 15 10 N/A 25
60-70 7 4 5 8 N/A 24
TED 10-11 0 0 13 12 N/A 25
20-21 0 7 11 0 N/A 18
30-35 1 4 13 7 N/A 25
60-70 5 5 13 2 N/A 25
Brain <0.0001 <0.0001 0.0203
SDH Z-E 10-11 0 2 13 8 2 25
20-21 0 5 7 4 3 19
30-35 0 7 10 7 1 25
60-70 0 1 12 7 5 25
Z-EXL 10-11 0 0 6 10 9 25
20-21 0 2 4 5 7 18
30-35 0 4 9 10 2 25
60-70 0 4 11 7 2 24
TED 10-11 0 0 4 6 15 25
20-21 0 1 4 10 3 18
30-35 0 3 11 9 2 25
60-70 0 4 5 7 9 25
Page 95
82
Table 3. 8: Summary of microscopic scoring of brains for trauma following application of each
of the three NPCB devices in layer chickens. Number of birds with each score are indicated.
Device1 Brain
section SDH Score PCH Score
0 1 2 3 4 0 1 2 3 4
Z-E Cerebrum 5 1 6 6 6 4 6 7 6 1
Mid brain 0 0 8 5 11 1 9 7 2 5
Hind brain 0 1 4 17 2 11 6 2 0 5
Z-EXL Cerebrum 3 0 2 10 9 3 3 4 8 6
Mid brain 0 2 2 7 13 0 4 4 7 9
Hind brain 1 4 5 7 7 7 7 7 2 1
TED Cerebrum 0 0 5 10 9 1 7 6 10 0
Mid brain 0 2 3 11 8 0 2 5 15 2
Hind brain 1 1 6 9 8 12 8 4 0 0
1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device.
SDH = Subdural hemorrhage.
PCH = Parenchymal hemorrhage
Page 96
83
Table 3. 9: Overall summary of microscopic scoring of subdural hemorrhage and parenchymal
hemorrhage in the brain and spinal cord of layer chickens killed by NPCB device. Number of
birds with each score are indicated
1Z-E = Zephyr-E, Z-EXL = Zephyr-EXL, TED = Turkey Euthanasia Device.
SDH = Subdural hemorrhage. PCH = Parenchymal hemorrhage.
Bolded numbers indicate significant results (P<0.05).
Device1 Age Score Total P Value
0 1 2 3 4 Device Age Device*Age
SDH in
brain
1.000 0.5388 0.8972
Z-E 0 0 0 12 12 24
Z-EXL 0 0 1 6 17 24
TED 0 0 0 10 14 24
SDH in
spinal
cord
0.999 0.995 0.9136
Z-E 0 0 3 9 12 24
Z-EXL 0 1 1 7 15 24
TED 0 0 1 7 16 24
PCH in 0.0242 0.1694 0.4794
brain Z-E 0 7 6 4 07 24
Z-EXL 0 1 3 9 11 24
TED 0 2 5 15 02 24
PCH in
spinal
cord
0.1822 0.0443 0.0215
Z-E 10-11 0 3 2 1 0 6
20-21 1 3 2 0 0 6
30-35 1 1 2 2 0 6
60-70 0 5 1 0 0 6
Z-EXL 10-11 3 3 0 0 0 6
20-21 1 2 1 1 1 6
30-35 3 2 1 0 0 6
60-70 0 1 4 1 0 6
TED 10-11 1 3 1 1 0 6
20-21 0 1 4 1 0 6
30-35 1 4 1 0 0 6
60-70 0 1 3 2 0 6
Page 97
84
Figure 3. 1: Non-penetrating captive bolt devices. (A) Zephyr-E standard: (B) Conical shape bolt head, (C) Standard subject adapter.
(D) Zephyr-E-layer: (E) Round shape bolt head, (F) Chicken subject adapter. (G) Zephyr-EXL: (H) Conical shape bolt head, (I)
Chicken subject adapter. (J) Turkey Euthanasia Device (TED): (K) Flat bolt head, (L) R-3 subject adapter.
Page 98
85
Figure 3. 2: Application of the Zephyr-EXL device on a 30 w.o. hen. The bird was restrained in
sternal recumbency with its neck resting ventrally on the ground, and the wings held gently
towards the body of the bird. Device was placed perpendicular to the top of the frontal bone just
behind the comb and on the mid line between the eyes and ears.
Page 99
86
Figure 3. 3 : Figure 3. Gross pathology scoring criteria for skull fractures. Arrows indicate the
fracture location [modified from Erasmus et al., (2010b) and Casey-Trott et al., (2013)]. (A) No
fracture, intact skull (score 0). (B) Depression fracture (score 1). (C) Penetrating fracture-no
imbedded fragments (score 2). (D) Penetrating fracture- with imbedded fragments (score 3).
Page 100
87
Figure 3. 4 : Skin reflected to demonstrate gross subcutaneous hemorrhage. (A) Hemorrhage
with less than 25% of area covered (score 2) of a 65 w.o. bird killed by the TED. (B)
Hemorrhage completely covering area from the eyes to base of the skull (score 4) of a 10 w.o.
bird killed by the TED. (C) Gross subdural dorsal hemorrhage covering less than 25% of the
brain surface (score 1) of a 33 w.o. bird killed by the Zephyr-E. (D) Gross subdural dorsal
hemorrhage covering 51 -75% of the brain surface (score 3) of a 33 w.o. bird killed by the
Zephyr-E.
Page 101
88
CHAPTER 4
Efficacy of a Novel Mechanical Cervical Dislocation Device in
Comparison to Manual Cervical Dislocation in Layer Chickens
This manuscript was accepted to journal of Animals with the following authors:
R.M.A.S. Bandara, S. Torrey, P.V. Turner, A. zur Linden, A. Bolinder, K. Schwean-Lardner and
T. M. Widowski
Page 102
89
4.1. Abstract
The main objective of this study was to assess the efficacy of mechanical cervical dislocation
using the Koechner Euthanasia Device Model C (KED) in comparison to manual cervical
dislocation in layer chickens. Laying hens and/or roosters in three different age groups (12, 27-
29, and 65-70 weeks old) were randomly assigned to one of three experimental groups: manual
cervical dislocation in conscious birds (CD), manual cervical dislocation in anesthetized birds
(aCD), or mechanical cervical dislocation by KED in anesthetized birds (aMCD). Anesthetized
birds received an intramuscular dose of 0.3 mg/kg medetomidine and 30 mg/kg of ketamine to
achieve a clinical anesthesia. A comparison of CD vs. aCD responses confirmed that the
anesthetic plane abolished or reduced clonic convulsions, nictitating membrane reflex, tonic
convulsions, and cloacal relaxation. Time to loss of the pupillary light reflex (~123 s), and time
to cardiac arrest (~172 s) were longer (P < 0.001) in the birds in aMCD group than aCD (~71 and
~137s, respectively). Radiographs revealed that the majority of the birds killed by manual
cervical dislocation (CD + aCD) had dislocations between the skull and atlas (C1) or between
cervical vertebrae C1-C2. The KED resulted in a majority of dislocations at C2-C3. Birds killed
by manual cervical dislocation presented more subdural and parenchymal hemorrhage in the
brain stem compared to birds killed by KED. Radiographs indicated the presence of fractures in a
few birds killed by either method (CD + aCD vs aMCD). Compared to manual CD, KED
resulted in less brain trauma and a longer latency to brain death indicating lower efficacy of KED
as an on-farm killing method.
Page 103
90
4.2. Introduction
Over the last decade, techniques used to kill poultry on farms have come under public and
scientific scrutiny due to concern for animal welfare. For the welfare of the animal, the goal of
any euthanasia technique is to achieve insensibility as quickly as possible, for the process to
cause minimal pain, and for death to follow quickly. Manual cervical dislocation is the most
common method for killing poultry on farms. Manual cervical dislocation involves stretching
and separating the cervical vertebrae by hand, rupturing the blood vessels and causing death by
cerebral ischaemia and extensive damage to the spinal cord and brainstem (Gregory and Wotton
1990a; Mason et al., 2009; Bader et al., 2014). Optimal application of cervical dislocation
separates the cervical vertebrae between the skull and the first cervical vertebra (C1) and
completely transects the spinal cord (HSA, 2004). However, cervical dislocation may be
esthetically displeasing to personnel performing it. Additionally, use of manual cervical
dislocation to kill poultry on-farm has been restricted to birds weighing less than 3 kg and to 70
birds per person per day through European Union (EU) legislation (EC, 2009).
To address the limitations around manual cervical dislocation, different tools have been
developed to perform cervical dislocation mechanically. Some devices (e.g. killing cone and
heavy stick) involve stretching, whereas others attempt to dislocate the cervical vertebrae by
forcing a blunted edge between two vertebrae (e.g. pliers, and Burdizzo) without stretching or
twisting the neck (Sparrey et al., 2014). Both manual and mechanical cervical dislocation
methods have been questioned for humaneness based on evidence that loss of sensibility is not
immediate in poultry species (Gregory and Wotton, 1990a; Sparrey et al., 2014). In their
guidelines, the AVMA (2013) specify that cervical dislocation “must result in luxation of the
Page 104
91
cervical vertebrae without primary crushing of the vertebrae and spinal cord”. However, there is
evidence that vertebral separation with the aid of a tool can cause fractures to the cervical
vertebrae in chickens (Bader et al., 2014) and turkeys (Bader et al., 2014; Woolcott et al., 2018b;
Widowski et al., 2018). At present, different devices are manufactured for cervical dislocation
and marketed to provide poultry producers with options for on-farm euthanasia. However, there
is a lack of evidence on the different devices’ efficacy in causing a humane death. The Koechner
Euthanizing Device (KED; Koechner Mfg. Co., Inc. Tipton, MO) is commercially available for
poultry as a mechanical cervical dislocation device. Woolcott et al. (2018b) assessed the efficacy
of the KED (Model S designed for chicks and poults) in anesthetized poults and young turkeys.
Their results revealed longer times to cessation of brain stem reflexes and more fractures in
cervical vertebrae in the birds killed by KED-S in comparison to manual cervical dislocation.
Jacobs et al. (2019) also found longer times to loss of brain stem reflexes when broiler chickens
were killed by KED-model-C compared to manual CD. Therefore, there is a need to evaluate the
KED in layer chickens before recommending its use on farms.
Ethical concerns arise in the evaluation of novel killing devices in conscious animals. Therefore,
it is necessary to initially evaluate novel killing devices either using cadavers (Martin et al.,
2017) or anesthetized animals (Woolcott et al., 2018b; Martin et al., 2019). The use of anesthetic
agents reduces the distress and pain associated with killing technique by abolishing the
awareness of the animal. Anesthetics produce insensibility by preventing integration (blocking
the interactions among specialized brain regions) or by reducing information (reducing the
number of activity patterns available to cortical networks) (Alkire et al., 2008). Efficacy and
humaneness of physical killing methods in poultry are often assessed based on time to loss of
Page 105
92
brain stem reflexes and degree of physiological trauma that is likely to result in rapid death (e.g.
Erasmus et al., 2010ab; Woolcott et al., 2018ab; Chapter 3). Previous studies have reported some
brain stem reflexes present under anesthesia in poultry (Woolcott et al., 2018b; Martin et al.,
2019), whereas others are not (Sandercock et al., 2014), thus the need to compare conscious and
anesthetized manual CD to understand the effect of anesthesia on the measures used to determine
efficacy.
The main objective of the current study was to assess the efficacy of mechanical cervical
dislocation using the Koechner Euthanasia Device (KED model-C) in comparison to manual
cervical dislocation based on behavioural responses, brain stem reflexes and postmortem analysis
of the physiological damage produced in three different age groups of layer chickens. For ethical
reasons, the KED was tested on anesthetized birds and compared with anesthetized birds killed
by manual cervical dislocation. In order to determine which measures would be valid for use in
anesthetized birds, our secondary objective was to determine the effect of the anesthetic agents
used in the study on behavioural and reflex responses by comparing anesthetized and conscious
birds killed by manual cervical dislocation.
4.3. Methods
The procedures and protocol for this research were reviewed and approved by the University of
Guelph Animal Care Committee (AUP 3321) which holds a Good Animal Practice certificate
101 issued by the Canadian Council on Animal Care.
Page 106
93
4.3.1. Animals and facilities
The birds were obtained from different research projects that were conducted at the Arkell
poultry research station of the University of Guelph. All the birds were targeted for euthanasia as
they had reached scheduled end of study, or because of routine flock depopulation. Different
strains and age groups of birds were available on different days. A health assessment was
performed on the birds prior to the experiment because we used anesthesia model, and only
healthy birds were used to reduce variable outcomes. Laying hens and/or roosters in three
different age groups (12, 27-29, and 65-70 weeks old) were randomly assigned to either manual
cervical dislocation (CD), anesthetized manual cervical dislocation (aCD) or anesthetized
mechanical cervical dislocation by KED (aMCD). Order of application of assigned treatments on
a trial day was determined by using the random number generator in Excel. Sex was not
balanced among the treatments due to the disproportionate availability of female and male layer
chickens in the different age groups. A total of 72 chickens were enrolled in this study: eight
birds in each age group were evaluated with each of the three killing methods. Strain, sex, body
weight and sample sizes for the different age classes of birds used in the study are given in Table
4.1.
4.3.2. Koechner Euthanizing Device (KED)
The Koechner Euthanizing Device (KED) is manufactured as a mechanical cervical dislocation
device for poultry by Koechner MFG. CO., INC (2016) (U.S. Patent No. 8,152,605). Four
different commercial models of the KED are manufactured for different weight classes (KED).
The model-S is designed for birds weighing up to 1.8 kg and the model-T is for birds weighing
up to 20.5 kg. The model-Txl is designed for birds weighing over 29.5 kg. The KED model
Page 107
94
evaluated in the current study was the model-C consisting of a 69 cm handle, marketed for birds
weighing up to 13.5 kg (Figure 4.1A).
4.3.3. Anesthesia and killing procedures
Anesthesia was induced in the birds by using a combination of medetomidine (1 mg/ml)
(CepetorTM, DIN:02337177, Modern Veterinary Therapeutics, LLC, Miami, FL, USA) and
ketamine (100 mg/ml) (Ketaset®, DIN: 02173239, Pfizer Animal Health, Kirkland, QC, CAN).
Appropriate drug doses to induce surgical anesthesia in chickens were established in a pilot
study. The chosen doses (medetomidine=0.3 mg/kg body weight; ketamine= 30 mg/kg body
weight) were administered as separate injections into the breast muscle by a veterinarian
monitoring the anesthesia.
After injecting the drugs, the birds were kept in a crate in a quiet and dark room for ten minutes
for the anesthesia to take effect. After another 5 minutes without interference, breathing,
heartbeat and pedal reflex were assessed. Fifteen minutes after the drug application, birds were
assessed for breathing, pupillary reflex, pedal reflex, neck muscle tone, jaw tone, and heartbeat.
The same responses were checked directly prior to application of the killing method. Birds were
determined to be ready to apply the killing method when they were non-responsive to handling,
and when assessment of breathing pattern, heart auscultation, jaw tone, and pedal reflex by the
monitoring veterinarian indicated surgical anesthesia.
Manual cervical dislocation was performed by a trained and experienced technician. The bird’s
head was held in the operator’s palm, with the neck between the index finger and thumb. Manual
Page 108
95
cervical dislocation was performed in one swift movement with the operator pulling down on the
bird’s head, stretching the neck, while rotating the bird’s head upwards into the back of the neck.
A trained and experienced technician applied the KED device. The KED was applied according
to the manufacturer’s instructions (Koechner MFG. Co., INC. 2016). Prior to the experiment, the
KED device was applied to cadavers in different age groups and radiographs were used to
confirm correct placement and pressure. All of the birds were manually restrained on a table in a
sternal recumbent position when applying the KED. The double angle blade was placed under
the neck of the bird while the single side blade was placed dorsally above the top of the neck at
the base of the skull. The handles were brought together quickly and firmly to cause dislocation
(Figure 4.1B). Then the device was removed from the neck of the bird.
4.3.4. Ante mortem assessment
The measures used in ante-mortem assessment are described in Table 4.2. Each measure was
checked immediately after application of the killing method and then at 10s intervals until
cessation. Presence of heartbeat was monitored through a stethoscope, and cardiac arrest was
determined to have occurred when no discernible heartbeat could be heard. All of the procedures
and responses of each bird were video recorded. Time to first feather erection was based on
blinded video recordings. The other measures were collected unblinded by live observations, and
the time at each event was confirmed with the video records.
Page 109
96
4.3.5. Post mortem assessment
4.3.5.1. Assessment of radiographs
Radiographs were performed on all birds immediately following euthanasia in an adjacent room.
Extreme care was taken with minimum handling to prevent further damage to the cervical
dislocation site. Four views were performed of each bird (dorsal ventral, ventral dorsal, right
lateral, left lateral) using a portable x-ray unit (Poskom VET-20BT X-ray unit with 80 kVp, 2.5
mAs power). The birds were placed directly on the wireless direct digital imaging receptor panel
(ULTRAMAXX, Promark imaging; 19.65 x 23.6 cm image matrix, a-SI TFT-PIN (thin filament
transistors), 77-micron pixel pitch, 6.0 to 7.0 lp/mm resolution) for radiograph acquisition.
Radiographs were assessed for site of luxation/subluxation, and for presence, type, and site(s) of
fractures (Table 4.3) by a board-certified veterinary radiologist blinded to the treatment. Digital
Imaging and Communications in Medicine (DICOM) formatted radiographs were interpreted
after importing the images into a DICOM viewer (Osirx MD, version 8.0.2, Pixmeo SARL,
Bernex, Switzerland).
4.3.5.2. Macroscopic assessment of tissue damage
Degree of external damage and bleeding caused by each killing method was assessed (0-2 scale)
based on laceration of the skin and presence of external hemorrhage on the neck: 0=no laceration
of the skin, 1=laceration of the skin with no external bleeding, 2=laceration of the skin with
external bleeding.
Dissection was performed on all birds to assess the degree of subcutaneous hemorrhage (SCH) at
the site of cervical dislocation, damage to the trachea, transection of the spinal cord, and degree
Page 110
97
of subdural hemorrhage (SDH) on the brain. Scoring criteria for macroscopic SCH and SDH are
presented in Table 4.4 (Woolcott et al., 2018ab; Chapter 3). SCH was scored by excising the skin
around the neck of the bird to measure the degree of hemorrhage at the site of cervical
dislocation. The skull was lifted, and dura were removed to assess the SDH on the brain.
4.3.5.3. Microscopic assessment of brain trauma
Once the macroscopic assessment was completed, brains and a sample of spinal cord from the
site of cervical dislocation were collected from six randomly selected birds in each age group per
killing method for microscopic evaluation. The brains and spinal cord samples were placed in
10% buffered formalin for at least 14 days before trimming. For consistency, all trimming was
performed by one individual. Three sections of the brain (cerebrum, mid brain and thalamus,
cerebellum/hind brain) and a section of the spinal cord at the site of cervical dislocation was
sampled (Woolcott et al., 2018b). The tissue sections were embedded in paraffin, cut 4µm and
stained with hematoxylin and eosin using standard techniques to make microscopic slides
(Animal health laboratory, University of Guelph). The sections on the slides were assessed
microscopically by a veterinary pathologist blinded to the treatments to determine the degree of
SDH and parenchymal hemorrhage (PCH). The degree of SDH and PCH were scored for each
microscopic slide using the same procedure as Chapter 3 (Table 4.4).
4.3.6. Statistical analyses
Statistical analyses were conducted using SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Two
separate statistical analyses were used. The first analysis was used to compare the behavioural
responses and reflex variables of anesthetized birds killed by manual cervical dislocation (aCD)
Page 111
98
to conscious birds killed by manual cervical dislocation (CD) in order to determine the effects of
anesthesia on each outcome variable. Then the response variables of the aCD were statistically
compared to those of the anesthetized birds killed by KED (aMCD).
Fisher’s exact tests were performed on the numbers of birds pooled across age groups that
showed pupillary light reflex, nictitating membrane reflex, gasping, feather erection, clonic
convulsions, tonic convulsions, and cloacal relaxation to find the effect of anesthesia (CD vs
aCD) and killing method (aCD vs aMCD). The numbers of birds presenting with luxation,
fractures, macroscopic SDH, and transection of spinal cord were similarly analyzed by Fisher’s
exact tests.
Generalized linear mixed models (GLMM) were used to analyze the fixed effects of killing
method, age, and killing method by age interaction of selected antemortem measurements (time
to loss of pupillary light reflex, duration of gasping, time at first feather erection, time to
cessation of heart beat). Nictitating membrane reflex, clonic convulsion, tonic convulsion, and
cloacal relaxation were excluded from the analysis due to low numbers of birds exhibiting these
measures in the anesthetized groups. Least significant means separation was conducted by using
the Tukey-Kramer test.
Generalized linear mixed models (GLMM) with multinomial distribution and cumulative logit
link functions were used to analyze the effect of the killing method, age, and their interaction on
postmortem macroscopic SCH at the site of dislocation, microscopic SDH and PCH of the brain
and spinal cord, and dislocation site in the neck (multinomial ordinal data). Odds ratios were
Page 112
99
computed to compare differences in the levels of fixed effects. Data from the three sections of
brain from each bird were pooled and the highest score of the three sections of each brain was
used for SDH and PCH analyses (Woolcott et al., 2018 ab; Chapter 3). The same procedure was
applied to the SDH and PCH analyses for the spinal cord. Dislocation site was analyzed by
conversion to a numerical category (skull–C1 = 1; C1-C2= 2; C2–C3 = 3; etc.); luxation and
subluxation were pooled for the analyses.
4.4. Results
4.4.1. Assessment of Ante mortem measures
4.4.1.1. Effects of anesthesia on birds killed by manual cervical dislocation
The proportion of birds presenting with ante-mortem measures following application of the
killing methods are given in Table 4.5. All conscious and anesthetized birds demonstrated
pupillary light reflex before application of the killing method. Pupillary light reflex was observed
in all conscious and anesthetized birds after application of the killing method indicating that
there was no anesthesia effect on pupillary light reflex. All conscious birds had nictitating
membrane reflex before application of manual cervical dislocation but nictitating membrane
reflex was present in only one anesthetized bird before application of manual cervical dislocation
(P=0.001).
The proportion of birds observed gasping was not different between conscious birds killed by
manual CD (75%) and anesthetized birds killed by manual CD (45%, P=1). The anesthetic did
not affect occurrence of feather erection in conscious birds killed by manual CD (conscious
manual CD = 87%, and anesthetized manual CD = 83%, P=1). Some anesthetized birds killed by
Page 113
100
manual CD did not exhibit clonic convulsions, tonic convulsions, or cloacal relaxation indicating
that there was anesthetic effect on these measures (Table 4.5). The number of birds exhibiting
clonic convulsions was significantly lower (P=0.001) in the anesthetized manual CD group than
the conscious manual CD group. Clonic convulsions with severe wing flapping and paddling
were observed in all conscious birds killed by manual CD. However, in 50% of anesthetized
birds killed by manual CD, clonic convulsions were absent except at the time of application of
the killing method (considered as no clonic convulsions in the analysis, only 2-3 wing flaps were
observed at the time of killing). An effect of anesthesia was found in the number of birds that
showed tonic convulsions (P<0.001) and cloacal relaxation (P<0.001).
Comparison of times to cessation of pupillary light reflex and heartbeat, time at first feather
erection, and duration of gasping in conscious and anesthetized birds killed by manual CD are
presented in Table 4.6. Durations were calculated for only the birds that showed the responses.
Overall, time to cessation of pupillary light reflex was longer (P=0.016) in conscious birds than
in anesthetized birds. There was a significant age by treatment interaction (P=0.036) with a
longer duration for the cessation of pupillary light reflex in conscious birds killed by manual CD
compared to the anesthetized birds killed by manual CD birds in the 65-70 week group. Duration
of gasping did not differ between the conscious and anesthetized groups (P=0.349). Anesthesia
did not affect time to first feather erection (P=0.226). There was no difference in time to
cessation of heart beat in conscious versus anesthetized birds killed by manual CD (P=0.137).
Time to cessation of heart beat took longer in the younger birds compared to older ones
(P<0.001), but there was no anesthesia by age interaction (P=0.596).
Page 114
101
4.4.1.2. Effect of manual CD versus mechanical CD in anesthetized birds
The number of anesthetized birds killed by manual CD or mechanical CD and their responses are
shown in Table 4.5. All anesthetized birds, regardless of treatment, presented pupillary light
reflex. The number of birds that demonstrated gasping was significantly higher (P<0.001) in
anesthetized birds killed by mechanical CD than anesthetized birds killed by manual CD. Feather
erection occurred in 83% of anesthetized birds killed by manual CD and in 75% of birds killed
by mechanical CD (P=0.723). There was no difference in the number of birds that showed clonic
convulsions in anesthetized manual CD and anesthetized mechanical CD groups (P=1). Number
of birds exhibiting tonic convulsion (P=1) and cloaca relaxations (P=0.740) was not different in
anesthetized birds killed by manual CD and mechanical CD.
Table 4.7 shows mean latencies to loss of antemortem measures or their durations in anesthetized
birds killed by manual CD versus mechanical CD in different age groups. Time to cessation of
pupillary reflex (P<0.001), duration of gasping (P=0.011), time to first feather erection (P=0.042)
and time to cessation of heart beat (P=0.012) were all longer in the anesthetized birds killed by
mechanical CD compared to anesthetized birds killed by manual CD in all age groups.
4.4.2. Assessment of postmortem measures
4.4.2.1. Assessment of cervical vertebrae radiographs
Results of radiographic scoring for presence and site of cervical dislocation are tabulated in
Table 4.8. The cervical vertebrae were assessed for any sites of luxation and/or subluxation. All
birds killed by manual CD had luxation (complete dislocation) irrespective of whether they were
conscious or anesthetized, whereas only 54% of birds killed by mechanical CD had luxation
Page 115
102
(P<0.001). The site of dislocation was significantly different for treatment (P<0.001) but not for
age (P=0.655) or age by treatment interaction (P=0.915) in birds pooled across luxation and
subluxation. There was a greater chance of causing dislocation further down the vertebrae than
the recommended skull to C1 in chickens killed with mechanical CD than manual CD. Overall
58% of the birds (conscious and anesthetized) killed by manual cervical dislocation had luxation
in between the skull and C1. In contrast, the majority of birds (75%) killed with mechanical CD
had luxation or subluxation between the C2 and C3 vertebrae.
The number of birds scored with fractures and the different types of fractures observed in the
cervical vertebrae are presented in Table 4.9. Fractures were observed in less than 10% of the
birds regardless of treatment or whether they presented with luxation or subluxation. There was
no difference between manual cervical dislocation and MCD in number of birds with fractures (P
= 0.195). Overall, 4 birds out of 24 (17%) killed with MCD and 3 out of 48 (6%) killed by
manual cervical dislocation had fractures of the cervical vertebrae. Figure 5.2 shows cervical
luxation (5.2-A), and subluxation with a fracture (5.2-B) in a layer chicken.
4.4.2.2. Macroscopic tissue damage assessment
Spinal cords were transected in all conscious and anesthetized birds killed by manual cervical
dislocation in all age groups (Table 4.8). The number of birds that had transected spinal cords
was significantly different between manual CD group (both conscious and anesthetized) and the
mechanical CD (P<0.001). Spinal cords were intact in 25% of birds killed by mechanical CD;
3/8 in both the 12 and 27-29 week groups, respectively. All spinal cords were transected in birds
killed by mechanical CD in the 60-75 week group.
Page 116
103
Trachea damage was not observed in any of the birds irrespective of the killing method. External
skin damage at the site of cervical dislocation was greater in the birds killed with mechanical CD
(12 week: 25%; 27-29 week:75%; 60-65 week:50%) than the birds killed by manual CD
(P<0.001). External skin damage at the site of cervical dislocation was observed in 8% of birds
killed by manual CD (conscious and anesthetized combined). Mechanical CD resulted in
external bleeding in 33% of the birds pooled across age groups whereas 4% of the pooled manual
CD birds had external bleeding (P=0.002).
Subcutaneous hemorrhage (SCH) at the site of dislocation was different among the treatments
(P<0.001) but there was no age effect (P=0.76) or age by treatment interaction (P=0.23) (Table
4.10). Except for one, all conscious and anesthetized birds killed by manual cervical dislocation
had a score of 2 or more for SCH. The highest score of 4 was found in over half of the birds
killed by manual cervical dislocation (conscious= 54%, anesthetized= 56%). Score 4 was absent
in all birds killed by the mechanical CD device. Macroscopic subdural hemorrhage (SDH) (score
1) in the brain was observed in only one bird out of 24 (12 w.o.) killed with MCD. Overall, 21%
of the birds killed by manual CD (10/48) had macroscopic SDH on the brain (P=0.086).
4.4.2.3. Microscopic evaluation
Table 4.11 gives descriptive statistics for SDH and PCH (all scores > 0) in the brain of birds
killed by the different methods in the three age groups. None of the birds in any group had SDH
of the cerebrum. Birds killed by manual CD (conscious and anesthetized combined) had SDH
both of the mid brain (30%) and the hind brain (63%) whereas only one bird out of 24 killed with
mechanical CD had SDH of the hind brain. Parenchymal hemorrhage (PCH) was observed in
Page 117
104
few birds overall. Six of 36 killed by manual CD had PCH in the hindbrain. No PCH was
observed in any of the birds killed by MCD. Although more birds killed by manual CD showed
microscopic damage compared to those killed by MCD, killing method, age, and method ˟ age
interaction did not affect brain SDH or PCH scores.
The number of birds with different scores for subdural hemorrhage (SDH) and parenchymal
hemorrhage (PCH) in spinal cord are presented in Table 4.12. Over 80% of all birds sampled had
SDH of the spinal cord (method P=0.168), but there was an effect of age (P=0.006), and
tendency for an age by killing method interaction (P=0.057). Birds at 12 weeks had 22 times of a
greater chance of having lower scores than birds at 65-70 weeks and 6 times greater chance of
having lower scores than birds at 27-29 weeks. Birds at 27-29 weeks had 4 times greater chance
of having lower scores for SDH of the spinal cord than birds at 65-70 weeks. There was a trend
of having lower scores of SDH of the spinal cord in the birds killed by mechanical CD in
younger age (12-29 weeks) groups. All the birds killed by mechanical CD had a score of 3 or 4 at
65-70 weeks. PCH in the spinal cord was significantly different among the killing methods
(P=0.041). Age and age by killing method interaction tended to be significant for the PCH
(P=0.088 and P=0.071 respectively). Conscious and anesthetized birds killed by manual CD had
a greater chance of having lower scores for PCH than birds killed by mechanical CD. Of the 18
anesthetized birds killed by mechanical CD, 13 birds (72%) had mild (score 2) to marked (score
4) PCH. Mild to marked PCH in the spinal cord was found in 50% of birds in conscious manual
CD group and 27% in anesthetized manual CD group.
Page 118
105
4.5. Discussion
Due to ethical concerns about testing the mechanical device on conscious birds, general
anesthesia was induced by using a combination of medetomidine and ketamine in order to
abolish any pain associated with the killing technique. Brain stem reflexes, behavioural
responses, and physiological parameters could be affected by the anesthetics used, which is why
we chose to study both the effect of the anesthesia and the effect of the method.
In the current study medetomidine was used in combination with ketamine. Medetomidine is
rapidly absorbed causing reliable sedation, analgesia, muscle relaxation, and anxiolysis (Sinclair,
2003). However, medetomidine decreases cerebral blood flow (Zornow et al., 1990; Keegan et
al., 1995) and heart rate (Cullen, 1996). Thus, medetomidine is combined with other anesthetics
to reduce the side effects (Sinclair, 2003). Ketamine is often used in combination with
medetomidine for avian anesthesia (Paul-Murphy et al., 2001). The major effects of ketamine
administration involve the central nervous system. Ketamine can affect eye reflexes and degree
of muscle tone and poses a problem when assessing the level of sedation or anesthesia (Hass et
al., 1992). Therefore, our anesthesia protocol had the potential to affect both antemortem
measures (eye reflexes and heart rate) and postmortem measures (brain hemorrhage), and needed
to be evaluated.
The anesthetic protocol used did not abolish the pupillary light reflex, which allowed us to use
this as an indicator of brain stem death in all birds. However, latency to loss of pupillary light
reflex was longer when birds were anesthetized. This is different than Woolcott et al., (2018b)
who found that pupillary light reflex was no longer in turkeys using the same anesthesia protocol
Page 119
106
as in this study. The anesthetics used in the current study also abolished or reduced four of the
other observed behavioural responses and reflexes (nictitating membrane reflex, clonic
convulsions, tonic convulsions, and cloacal relaxation) precluding their use for comparing killing
methods. Woolcott et al. (2018b) reported that presence of the nictitating membrane reflex,
presence of clonic convulsions, and duration of gasping were affected by the same anesthetic
protocol in turkeys. Sandercock et al. (2014) observed nictitating membrane reflex until after
respiratory arrest and brain death in laying hens anesthetized with sevoflurane, and suggested
that the longer time to cessation of nictitating membrane reflex may have been due to combined
effects of the anesthetic and sedative they used. In the current study, nictitating membrane reflex
was not present in many of the chickens as a result of the anesthetic protocol. We previously
found that onset of tonic convulsion and cloacal relaxation correlated with cessation of heart beat
in laying hens and can be used as a practical measure of clinical death (Chapter 3). However, our
current results revealed that both tonic convulsions and cloacal relaxation were reduced in many
of the anesthetized birds. Thus, latency to pupillary reflex, nictitating membrane reflex, clonic
convulsions, tonic convulsions, and cloacal relaxation were affected by our anesthetic protocol.
The anesthetics did not affect time to cessation of heart beat in chickens. Since latency to loss of
pupillary reflex was shortened by the anesthetic, our estimate for time of brain death using the
anesthetized bird as a model is conservative.
The main objective of the current study was to assess the efficacy of mechanical cervical
dislocation using the KED (mechanical CD) in comparison to manual cervical dislocation
(manual CD). The results of the current study reveal that time to loss of pupillary light reflex, an
indicator of brain stem death, was longer in the anesthetized birds killed with mechanical CD
Page 120
107
than by manual CD. Previous studies also reported shorter latency to loss of eye reflexes in
poultry successfully killed with manual cervical dislocation in comparison to birds killed by
mechanical CD using the KED (conscious broilers: Jacobs et al., 2019; anesthetized turkeys:
Woolcott et al., 2018b). Gregory and Wotton (1990a) reported that birds killed by mechanical
CD (using Semark neck pliers) exhibited longer time to loss of visual evoked responses
compared to manual CD, and concluded that cervical dislocation by stretching was more
effective than cervical dislocation by crushing. Hernandez (2018) also reported a longer time to
onset of isoelectric EEG indicating brain death in layers (60 weeks of age; anesthesia was
induced by isoflurane at 5% concentration with oxygen via a face mask) killed by mechanical
CD by KED in comparison to manual CD. Results from the current study reveal that time to
cessation of heart beat and time at feather erection were also longer in the anesthetized birds
killed by mechanical CD compared to manual CD. Heard (2000) suggested sudden feather
erection in anesthetized birds as an indicator of cardiac arrest or reduced blood flow to the heart.
In the current study, >75% of birds exhibited feather erection irrespective of the killing method.
Recent studies reported that there is no association between time at feather erection and cardiac
arrest in chickens (Hernandez 2018; Chapter 3). Thus, the delayed feather erection in the birds
killed by mechanical CD is difficult to explain.
Ideally, cervical dislocation separates the cervical vertebrae in between the skull, and the atlas
(C1), completely transecting the spinal cord and disrupting blood vessels. In the current study,
the vast majority of the birds killed by manual CD had dislocations in between the skull and atlas
(C1) or between C1-C2. However, the KED resulted in a majority of dislocations between C2
and C3. Thus, manual CD caused more cranial dislocations than the KED. Previous studies also
Page 121
108
reported similar results indicating that manual cervical dislocation resulted in the majority of
dislocations between the skull and atlas (C1) in chickens (Bader et al., 2014; Martin et al., 2016,
2017, 2018a) and turkeys (Woolcott et al., 2018b). Martin et al. (2018b) reported that more
cranial dislocation was associated with improved kill success and more rapid loss of reflexes
compared to more caudal dislocations for cervical dislocation.
Anatomical damage to the top of the spinal cord and the base of the brain stem is possible with
more cranial dislocation, which is associated with spinal cord concussion, neurogenic shock, and
loss of consciousness in humans (Dumont et al., 2001; Harrop et al., 2001). Manual CD resulted
in all birds having cervical luxation, more cranial luxation locations, more spinal cord
transections, greater SDH and PCH in the brain stem, and shorter time to loss of brain stem death
and cardiac arrest. Except for one bird, none of the birds killed with mechanical CD using the
KED had SDH or PCH in the brain, indicating that KED did not cause severe brain trauma.
Previous studies suggested that immediate insensibility and irreversible loss of vital functions are
associated with SDH and PCH in the brain (Erasmus et al., 2010b; Bader et al., 2014). We
suggest the absence of SDH and PCH may be the reason for the observed longer time to loss of
pupillary light reflex and clinical death in the birds killed by mechanical CD.
The majority of birds killed by either manual CD or mechanical CD showed transected spinal
cords and SDH and PCH in the spinal cord, indicating both methods were sufficient to cause
traumatic damage to the spinal cord. The spinal cord was transected in all 65-70 week old birds
killed by mechanical CD irrespective of whether there was luxation or subluxation. This
indicates that KED model-C caused severe damage to the cervical area of the spinal cord of 65-
Page 122
109
70 week old roosters. Some muscles responsible for inspiration are innervated by cervical nerves
and the accessory cranial nerve (CN X1) (DeTroyer and Estenne, 1984). Some authors reported
the evidence that damage to the cervical area of the spinal cord impaired the functioning of these
respiratory muscles resulting in hypoxia due to failures in respiration process, and finally cause
death in humans (Winslow and Rozovsky, 1983). A higher number of birds killed by mechanical
CD exhibited longer duration of gasping in comparison to manual CD. Therefore, we suggest
that hypoxia resulting from spinal cord trauma and leading to cerebral ischemia could be a main
reason for the observed gasping reflex and death in mechanically cervical dislocated birds; they
also had minimal levels of brain trauma.
There were few fractures in cervical vertebrae of the chickens irrespective of the killing method.
This is in contrast to previous studies that reported prevalence of fractures in poultry killed by
manual CD and mechanical CD. Woolcott et al. (2018b) found considerably more fractures in 3
week old turkeys killed by mechanical CD (KED model-S), with 78% having transverse or
comminuted fractures or fragmentation. In their study, turkeys killed via manual CD also had
fractures, although fewer (44%) than with KED. Bader et al. (2014) also reported different types
of fractures (transverse, comminuted, longitudinal) in chickens by manual cervical dislocation
and turkeys killed by mechanical cervical dislocation using a nonpenetrating forcep with three
blunt shear arms.
Less external blood loss and fewer skin tears are preferred in the commercial environments due
to biosecurity and esthetic concerns. Similar to the findings of Woolcott et al. (2018b) in turkeys
and Jacobs et al. (2019) in broiler chickens, the KED resulted in more external skin damage in
Page 123
110
layer chickens than manual CD. Other important factors when assessing killing methods for
practical on-farm use, are safety for stock people and ease of training. Cervical dislocation is
easy to perform manually, practical in any emergency killing, and can be applied immediately
during barn walkthroughs. However, manual CD is associated with variability in operators’
performance (Martin et al., 2018b). We assume a person can be easily trained on operating the
KED device, and there is little risk associated with mis-use of the device for the operator or
technical difficulties.
4.6. Conclusion
Compared to manual cervical dislocation, mechanical CD using the KED caused more external
skin trauma but less trauma to the brain with more incomplete dislocations located in the lower
regions of the cervical vertebrae. Birds killed with the KED also showed longer times to loss of
pupillary light reflex and cessation of heart beat. Overall, mechanical cervical dislocation by
KED resulted in lower efficacy in comparison to manual cervical dislocation as on-farm killing
method for layer chickens. The anesthetic protocol abolished or reduced clonic convulsions,
nictitating membrane reflex, tonic convulsions, and cloacal relaxation in the current study
suggesting that these behavioural responses or reflexes are not useful as an approximate measure
of brain death in cases where ketamine and medetomidine have been administered in chickens.
Page 124
111
Table 4. 1: Strain, sex, body weight and sample sizes for the different age classes of birds used
in the study
Age
(week)
Killing
method
Number
of birds
Strain Sex Body weight
(Kg)
(Avg wt ± SD)
12 CD 8 White Leghorn All females 1.00 ± 0.10
aCD 8 White Leghorn All females 0.94 ± 0.09
aMCD 8 White Leghorn 2 Males + 6 Females 1.02 ± 0.62
27-29 CD 8 White Leghorn 1 Male + 1 Females 1.77 ± 0.03
Columbian Rock 1 Male + 5 Females 1.75 ± 0.36
aCD 8 White Leghorn 3 Males + 3 Females 1.79 ± 0.29
Columbian Rock 2 females 2.03 ± 0.29
aMCD 8 White Leghorn 3 Males + 4 Females 1.86 ± 0.35
Columbian Rock 1 female 1.98
65-70 CD 8 White Leghorn All males 2.16 ± 0.18
aCD 8 White Leghorn All males 2.10 ± 0.19
aMCD 8 White Leghorn All males 2.29 ± 0.29
CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Page 125
112
Table 4. 2: List of ante-mortem assessment measures, description, and procedure use, recorded
in order of observation after application of each killing method (based on Chapter 3)
Measures Description Procedure
Pupillary light reflex Constriction of the pupil in
response to light
Light from a medical penlight
was directed into the eye and
pupil constriction was examined
Nictitating membrane reflex Transient closure of the
nictitating membrane in
response to mechanical
stimulation
The medial canthus of the eye or
the cornea was lightly touched
with a fingertip
Gasping Paroxysmal opening of the beak Visual observation of
paroxysmal opening of the beak Feather erection
Sudden erection of feathers, not
in response to external stimuli Visual observation of first
occurrence of feather erection
Clonic convulsions Rapid, uncoordinated movement
of the body and wings Visual observation of rapid wing
flapping and foot paddling Tonic convulsions Muscle rigidity with the legs
and wings outstretched Visual observation of the time
of onset of legs and neck
outstretched Cloacal relaxation
Cloacal opening following
contractions of cloaca Visual observation for cloacal
opening following contractions
Cardiac arrest Cessation of heart beat
Auscultation by using a
stethoscope
Page 126
113
Table 4. 3: Definitions for the terminology used in radiograph evaluation
Terminology Definition
Luxation Complete dislocation of two vertebrae at the articular
process joints.
Subluxation Partial (incomplete) dislocation of two vertebrae at
the articular process joints.
Fracture type
Sagittal Parallel to the long axis of the vertebra on midline.
Trans (transverse) Perpendicular to the long axis of the vertebra.
Articular processes Fractures of the articular processes of a vertebra.
Dens Fracture of the dens (tooth-like process that projects
from the cranial aspect of the centrum of the axis (C2)
to articulate with the atlas (C1).
Crushed vertebral bodies Multiple fractures of a vertebra that cannot be
classified as a fragment, articular process or dens
fracture.
Page 127
114
Table 4. 4: Gross and microscopic pathology scoring criteria for macroscopic, and microscopic
hemorrhage (Woolcott et al., 2018ab; Chapter 3)
Score Macroscopic Microscopic
Subcutaneous or subdural hemorrhage
Subdural or parenchymal hemorrhage
0 None None
1 <25% of surface area Minimal (<5% of section)
2 26–50% of surface area Mild (5–10% of section)
3 51–75% of surface area Moderate (11–30% of section)
4 76–100% of surface area Marked (>30% of section)
Page 128
115
Table 4. 5: Number of birds presenting with ante-mortem measures following application of the
killing methods.
Method
CD aCD aMCD
N=24 N=24 N=24
Pupillary light reflex 24 24 24
Nictitating membrane reflex 16a 1b, d 9c
Gasping 18 11d 24c
Feather erection 21 20 18
Clonic convulsions 24a 12b 11
Tonic convulsions 15a 1b 1
Cloacal relaxation 24a 7b 5
a,b Different superscripts indicate numbers of birds presenting the measure is different between
CD and aCD according to Fishers’ exact tests (P<0.005)
c,d Different superscripts indicate numbers of birds presenting the measure is different between
aCD and aMCD according to Fishers’ exact tests (P<0.005)
CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Page 129
116
Table 4. 6: Mean latencies to or durations of (± SE, s) ante-mortem measures in conscious and
anesthetized chickens killed by manual cervical dislocation in different age groups. P values are
given for effects of age, anesthesia and age by anesthesia interaction.
P value
Age
(week)
CD aCD
Age
Anesthesia
Age × Anesthesia
Time to 12 81 ± 7 70 ± 7 0.8507 0.0166 0.0369
loss of 27-29 77 ± 5 80 ± 5
pupillary 65-70 96 ± 7a 62 ± 7b
reflex Overall 85 ± 3 71 ± 3
Gasping 12 22 ± 7 50 0.1497 0.3490 0.7706
duration 27-29 57 ± 8 68 ± 8
65-70 73 ± 13 74 ± 17
Overall 51 ± 7 64 ± 11
Time to 12 82 ± 15 104 ± 16 0.8942 0.2264 0.3785
first feather 27-29 78 ± 19 114 ± 17
erection 65-70 93 ± 15 84 ± 11
Overall 85 ± 10 101 ± 10
Time to 12 191 ± 20 173 ± 20 0.0005 0.1377 0.5960
cessation 27-29 122 ± 9 119 ± 9
of heart 65-70 152 ± 12 119 ± 12
beat Overall 155 ± 3 137 ± 8
Durations calculated for only the birds that showed the responses
Superscript letters show the significant difference (P<0.05)
CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
Bolded numbers indicate significant results (P<0.05)
Page 130
117
Table 4.7: Mean latencies to or durations of (± SE s) ante-mortem measures in anesthetized
chickens killed by manual or mechanical cervical dislocation in different age groups. P values
are given for effects of age, method and age by method interaction.
P value
Age aCD aMCD
Age
Method
Age ×
Method
Time to 12 70 ± 14 142 ± 14 0.2014 <0.0001 0.3091
loss of 27-29 80 ± 8 118 ± 8
pupillary 65-70 62 ± 10 107 ± 10
reflex Overall 71 ± 7b 123 ± 7a
Gasping 12 50 117 ± 18 0.5367 0.0109 0.7338
duration 27-29 63 ± 13 102 ± 10
65-70 70 ± 20 132 ± 16
Overall 64 ±16b 117 ± 9a
Time to first 12 104 ±25 146 ± 30 0.7083 0.0423 0.5599
feather 27-29 114 ± 10 125 ± 11
erection 65-70 87 ± 15 135 ± 14
Overall 101 ± 12b 135 ± 11a
Time to 12 173 ± 23 172 ± 23 0.0998 0.0125 0.0945
cessation of 27-29 119 ± 10 153 ± 10
heart beat 65-70 119 ± 13 193 ± 13
Overall 137 ± 9b 172 ± 9a
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Durations calculated for only the birds that showed the responses
Superscript letters show the significant difference (P<0.05)
Bolded numbers indicate significant results (P<0.05)
Page 131
118
Table 4. 8: Presence and location of luxation/subluxation (from radiographs) and spinal cord transections (from macroscopic
evaluation) in conscious and anesthetized chickens killed by manual cervical dislocation and KED1. Values indicate number of birds.
Luxation Sub-Luxation Spinal
cord
transection
Age Method Total
number of
birds
assessed
# birds
with
luxation
Sk-C1 C1-C2 C2-C3 C3-C4 # birds
with sub-
luxation
Sk-C1 C1-C2 C2-C3 C3-C4 Number
of birds
12 CD 8 8 8 0 0 0 0 0 0 0 0 8 aCD 8 8 7 0 0 1 0 0 0 0 0 8 aMCD 8 5 2 1 2 0 3* 1 1 3 0 5 27-29 CD 8 8 1 7 0 0 0 0 0 0 0 8 aCD 8 8 3 5 0 0 0 0 0 0 0 8 aMCD 8 5 0 0 5 0 3 0 0 1 2 5 65-70 CD 8 8 4 3 1 0 0 0 0 0 0 8 aCD 8 8 5 3 0 0 0 0 0 0 0 8 aMCD 8 3 0 0 3 0 5 0 0 4 1 8
1Number of birds presenting luxation and subluxation were pooled for statistical analysis. Results of statistical analyses are given in
the text.
CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
*Two birds had sub-luxation in multiple places
Page 132
119
Table 4. 9: Results of radiographic scoring on number of birds with fractures and the types and
locations of fractures in conscious and anesthetized chickens.
1CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Age (weeks) Method1 Number of birds with
fractures
Fracture type Location
12 CD (n=8) 0 -
aCD (n=8) 0 -
aMCD (n=8) 1 Trans C3
1 Crushed vertebral bodies C3 and C4
27-29 CD (n=8) 1 Small fractures Dens of C2
aCD (n=8) 1 Trans Dens of C2
1 Small fractures Dens of C2
aMCD (n=8) 0 -
65-70 CD (n=8) 0 -
aCD (n=8) 0 -
aMCD (n=8) 2 Articular processes C3
Page 133
120
Table 4. 10: Macroscopic evaluation of subcutaneous hemorrhage (SCH) at the site of
dislocation. Number of birds with each score are indicated
1CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Method1 Age Score P value
0 1 2 3 4 Method Age Method*Age
SCH CD 12 0 0 1 2 5 0.0002 0.7603 0.2330
27-29 0 0 1 3 4
65-70 0 1 3 0 4
aCD 12 0 0 2 2 4
27-29 0 0 0 0 8
65-70 0 0 2 5 1
aMCD 12 0 5 2 1 0
27-29 0 2 0 6 0
65-70 0 2 5 1 0
Page 134
121
Table 4. 11: Summary of microscopic scoring of brains for trauma following application of each
of the three killing methods in layer chickens. Number of birds with hemorrhage (any score >0)
in each section are indicated.
Age Method1 Subdural hemorrhage
(SDH)
Parenchymal hemorrhage
(PCH)
(Week) Cerebrum Mid
brain
Hind
brain
Cerebrum Mid
brain
Hind
brain
12 CD 0 0 5 0 0 1
aCD 0 0 4 0 0 0
aMCD 0 0 0 0 0 0
27-29 CD 0 5 6 0 0 2
aCD 0 1 3 0 0 2
aMCD 0 0 1 0 0 0
65-70 CD 0 1 4 0 0 0
aCD 0 4 0 0 0 1
aMCD 0 0 0 0 0 0
Six randomly selected brains were assessed for each device in each age group. 1CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation by KED
Page 135
122
Table 4. 12: Overall summary of microscopic scoring of subdural hemorrhage and parenchymal
hemorrhage in the spinal cord of layer chickens killed by three killing methods. Number of birds
with each score are indicated.
Randomly selected 6 brains with spinal cords were assessed for each killing method in each age
group. 1CD = Conscious manual cervical dislocation
aCD = Anesthetized manual cervical dislocation
aMCD = Anesthetized mechanical cervical dislocation
Method1 Age Score P value
0 1 2 3 4 Method Age Method*Age
SDH of CD 12 4 2 0 0 0 0.1680 0.0006 0.0572
Spinal 27-29 1 1 2 2 0
cord 65-70 0 0 2 2 2
aCD 12 2 0 2 1 1
27-29 0 0 2 3 1
65-70 0 2 1 2 1
aMCD 12 1 3 1 1 0
27-29 2 0 4 0 0
65-70 0 0 0 4 2
PCH in CD 12 3 1 1 0 1 0.0407 0.0885 0.0712
spinal 27-29 2 1 2 2 0
cord 65-70 2 0 2 1 0
aCD 12 2 0 1 3 0
27-29 3 3 0 0 0
65-70 2 3 0 1 0
aMCD 12 0 2 0 3 0
27-29 3 0 4 0 0
65-70 0 0 1 3 2
Page 136
123
Figure 4. 1: A- Koechner Euthanasia Device (KED model-C): S-single side blade; D-double
angle blade. B-application of KED model-C in a 65 week old anesthetized rooster.
Page 137
124
Figure 4. 2 : Radiographs of chickens showing cervical dislocations. A- Luxation between the
skull and C1 vertebra in a 65 week old rooster killed by manual cervical dislocation: Sk-Skull,
C1- first cervical vertebra, C2- second cervical vertebra. B - Subluxation between C2 and C3
(letter S shows the site of subluxation) in a 65 week old rooster killed by KED: fractures (F) are
present on the articular processes of the C3 vertebra.
Page 138
125
CHAPTER 5
Aversion to CO2 Gas in Laying Hens Using Approach-Avoidance
and Conditioned Place Avoidance Paradigms
This manuscript is in preparation for submission to Poultry Science with the following
authors:
R. M.A.S. Bandara, T. M. Widowski, S. Torrey, R. L. Parsons, and S. T. Millman
Page 139
126
5.1. Abstract
Carbon dioxide (CO2) has been evaluated as an inhalant euthanasia agent in poultry but animal
welfare concerns have been raised on associated pain and aversion. This study assessed the
aversion to four different CO2 concentrations (25%, 35%, 50%, 70%) in laying hens based on
approach avoidance test and conditioned place avoidance test. An apparatus consisting of two
identical chambers separated by a sliding door and a curtain was used. The control chamber (CC)
maintained ambient air conditions; the treatment chamber (TC) maintained predetermined CO2
concentrations. Twelve laying hens (1.2 ± 0.1 kg; 24 week-old) were individually trained to
access rewards in the TC. Hens were randomly assigned to the first CO2 treatment and then
assigned to the remaining rounds to balance treatments. Each round of testing consisted of one
Gas Day, preceded by one Baseline Day (1% CO2) and followed by one Washout Day (1% CO2).
On Gas Days, the TC was maintained at one of four CO2 levels. Hens were considered to show
aversion if they pushed the curtain and/or inserted their heads into the TC without fully entering.
Hens were considered to display CPA if they did not enter the TC on Washout Day after entering
the TC on the previous Gas Day. Eight of 9 hens entered the TC at 25% and 35% CO2, 6 of 11
entered at 50%, and 2 of 7 entered at 70% CO2. Head shaking, open mouth breathing, ataxia and
loss of posture were observed in all hens at all concentrations. Latency to onset of ataxia (less
than 15s; P=0.09) tended to decrease and loss of posture (less than 25s; P=0.02) decreased with
increased CO2 concentrations. Some hens displayed conditioned place avoidance following loss
of posture at 25% and 35%, and more hens at 50% CO2.We suggest 50% and above CO2
concentrations are more aversive to hens than 25% and 35%. Therefore, euthanasia with CO2
should target concentrations less than 50% to achieve welfare advantages in layer hens.
Page 140
127
5.2. Introduction
Over the last three decades, carbon dioxide (CO2) has been evaluated by various researchers as a
method of gas stunning and as an inhalant euthanasia agent for poultry using either static
(prefilled) or gradual fill techniques. Animal welfare concerns have been raised regarding pain,
distress and associated aversion experienced by animals exposed to high concentrations of CO2
from the time of gas introduction to the onset of insensibility (Raj, 1996; McKeegan, 2005;
Sandilands et al., 2011). To be considered acceptable as an inhalant euthanasia agent, established
guidelines require that the concentration not be aversive to the species, and that the agent induce
rapid loss of sensibility without recovery (AVMA, 2013; NFACC, 2017).
Carbon dioxide is an anesthetic gas (Kohler et al., 1999) that produces insensibility by
suppressing afferent transmission of sensory stimuli to the brain (Raj et al., 1992). Carbon
dioxide causes death by inducing hypercapnia (excessive CO2 in the bloodstream) and hypoxia.
Carbonic acid is formed when CO2 contacts moisture on respiratory and ocular membranes,
which can cause pain and distress in animals due to stimulation of nociceptors in respiratory and
ocular membranes (Anton et al., 1992; Chen et al., 1995; Feng and Simpson, 2003; McKeegan et
al., 2004).
Different species can detect different gas levels through gustation, olfaction and nociception
(McKeegan et al., 2005). Humans can detect CO2 at the 10% level even though it is not pungent
to inhale (Patterson et al., 1962). Pain threshold level for CO2 in humans has been identified at
concentrations between 40% and 55% (Anton et al., 1992). Laying hens appear to be able to
detect CO2 concentrations as low as 7.5% (Raj and Gregory, 1991). Different behaviourl
Page 141
128
responses to CO2 have been suggested as indicators of distress, aversion and pain perception.
Gerritzen et al. (2000) suggested that gasping and head shaking in broiler chickens exposed to
60% CO2 and gas mixtures containing 30% and 40% CO2 as possible indicators of breathlessness
and the pungency of gas mixtures. The avian nociception threshold for CO2 is in the 40-50%
which is in the same range as reported in humans.
Cognitive paradigms of approach avoidance (AA) have been used to measure the aversion to a
negative experience associated with different gas types (CO2 in ewes: Grandin et al., 1986; NH3
in laying hens: Kristensen et al., 2000; CO2 and Ar in laying hens: Webster and Fletcher, 2004).
These tests directly ask the animal whether they are willing to enter an environment filled with
an inhalant agent. In an AA test, CO2 (potentially causing a negative experience) is paired with a
positive stimulus (reward) to elicit motivational conflict, and the motivation to approach the
reward (e.g. feed and enrichment) is compared against the motivation to avoid gas exposure
(layer chickens: Raj and Gregory, 1991, Webster and Fletcher, 2004; broilers: Sandilands et al.,
2011).
Previous studies that assessed aversion in chickens to concentrations of 30% CO2 and higher by
using AA tests have produced conflicting results. Webster and Fletcher (2004) tested aversion to
30%, 45% and 60% CO2 and concluded that there is no welfare disadvantage to the use of up to
60% CO2. They assessed avoidance of CO2 in a test apparatus consisting of a raised entry
chamber connected by a descending chute to a lower chamber into which gas was injected. Hens
had to go down a chute to the test chamber and were able to detect gas before reaching the
chamber. Hens made a number of stops and retreats during the approach to the lower chamber,
Page 142
129
so the level of gas they avoided was uncertain. Gerritzen et al. (2000) assessed aversion to 60%
CO2 in broiler chickens by using an AA test. In this study, chickens were isolated from the flock
and then allowed to reinstate social contact by walking through a CO2-filled tunnel towards the
companions placed behind a glass panel. No difference was found in numbers of birds that
entered the tunnel for different CO2 concentrations, and the authors concluded that the birds
could not detect CO2 concentrations. However, social isolation is highly distressing for chickens,
and the motivation for social reinstatement may have confounded the results. Additionally, very
few control birds entered the tunnel limiting the value of this test.
Previous studies also evaluated the aversion to CO2 in chickens based on interruption of on-
going feeding behaviour when increasing CO2 concentration. Only sub-stunning levels of CO2
(5%, 7.5%, 10%) were assessed by Raj and Gregory (1991) who reported that hens spent less
time in a feeding chamber when the concentration was above 5%. McKeegan et al. (2006)
studied the immediate aversion to CO2 (10%, 25%, 40%, 55%, 70%) in broiler chickens. The
chickens were exposed to gas for 10 s and the authors concluded that the birds demonstrated a
moderate immediate aversion to 40% or above CO2 based on the time at cessation of feeding and
withdrawal from the food dish. Sandilands et al. (2011) assessed feeding withdrawal time of
broiler chickens in higher CO2 concentrations of 50%, 55%, and 60%, and concluded that all
CO2 concentrations were aversive.
Conditioned place avoidance (CPA) tests measure animal responses when they are re-exposed to
an environment previously associated with a negative experience. CPA tests have been used to
evaluate the responses in goats (Withrock, 2015), swine (KC et al., 2016), and turkeys (Bandara
Page 143
130
et al., unpublished) exposed to different CO2 concentrations. While they did not measure CPA
directly, Webster and Fletcher (2004) observed that laying hens exposed to different gas
concentrations had significantly longer latencies to leave the control chamber on the day
following testing (retraining day) indicating that their previous experience affected subsequent
behavioural responses.
Replication of previous studies on assessment of aversiveness to CO2 in chickens using different
testing protocols, and assessing different CO2 concentrations as possible euthanasia agents are
important to refine euthanasia protocols. Therefore, our first objective was to assess the degree of
aversion in laying hens exposed to four different CO2 concentrations (25%, 35%, 50%, 75%)
based on AA test and distress behaviours. We hypothesized that birds would avoid entering the
treatment chamber when CO2 concentrations were aversive. The second objective was to assess
the degree of aversion associated with previous exposure and loss of sensibility based on CPA
and distress behaviours. We hypothesized laying hens would avoid the environment previously
associated with aversive CO2 concentrations and where loss of sensibility was experienced. The
third objective was to estimate the time to insensibility by assessing time to loss of posture in
laying hens exposed to different CO2 concentrations.
5.3. Methods
The protocol for this experiment was approved by the Institutional Animal Care and Use
Committee (IACUC) at Iowa State University.
Page 144
131
5.3.1 Animals, Housing, and Management
Twenty 36-week-old laying hens (White Leghorn) were obtained from a research flock at Iowa
State University. Upon arrival at Laboratory Animal Research (LAR) facility at Iowa State
University, the birds were housed in a single pen in a temperature-controlled room (2.5 m x 3.65
m) in the LAR facility. Room temperature was 24°C and relative humidity was maintained
between 60 – 65% with a photoperiod of 12 hrs (6.00 a.m to 6.00 p.m). The concrete floor of the
room was covered with wood shavings. Five nest boxes were provided inside the pen.
Environmental enrichment included two metal chains, two hanging compact discs, and a perch.
Birds were provided with ad libitum access to a commercial layer diet (Heartland Co-op, Prairie
City, IA) and water. Birds were individually identified by using colored leg bands. During a 4-
week acclimation period, pullets were habituated to the researcher’s presence and handling, and
birds were familiarized with features of the test apparatus (e.g. the same plastic curtain made of
transparent PVC strips, which was used in the test apparatus) and food rewards (live meal
worms) within the home pen.
5.3.2. Experimental room and equipment
The experiment was conducted in a room located in the same building where the hens were
housed. Tests were performed using a custom designed apparatus, as described by Withrock
(2015) and KC (2018). In brief, the apparatus consisted of two identical chambers (61cm x 61cm
x 76 cm) separated by a sliding door and a plastic curtain made of transparent PVC strips (Figure
5.1). Birds were able to see through the curtain and could easily push through it to enter the
treatment chamber. Each chamber was outfitted with a hinged door through which birds could be
placed or removed from the chamber. The doors and top panels were made of Plexiglass to
Page 145
132
facilitate behavioural observations. The side panels of the box were made of opaque, hard plastic
to avoid the researchers being seen by the birds. Chambers floors were covered with black rubber
mats.
The test apparatus was engineered for independent atmospheric control in each chamber, using
recirculating positive pressure systems to maintain separate designated CO2 concentrations. This
included two small square axial fans (Model 4WD47, Dayton Electric Mfg. Co. IL) that drew air
into the right (double) sidewall of the chamber where CO2 gas was injected and mixed into the
air stream. The CO2 enriched air was then delivered to the chamber through perforations evenly
distributed in the left side wall of the treatment chamber (TC). A continuous flow of ambient air
was similarly introduced into the control chamber (CC). To maintain the separate CC and TC
environments, a negative pressure exhaust sink was located between the two chambers where
any excess CO2 was evacuated.
To further facilitate maintaining CO2 concentration at or near ambient levels (<1%) within the
CC during all treatments, the top panel was opened to release air and was covered with a plastic
mesh (5.08 x 5.08 cm) to prevent birds escaping. The opened top panel also facilitated recording
of vocalizations inside CC.
To ensure that hens would be highly motivated to enter TC, enrichments were provided to
stimulate foraging and exploration. The rubber mat inside the treatment chamber (TC) was
covered by a waterproof disposable underpad (Pet All Star Training Pad, Pet All Stars, ASIN,
B014RDSMFU) to facilitate cleaning between birds. Wood shavings were scattered over the
Page 146
133
underpad. Enrichments included a compact disc, a plastic lid (diameter = 12.7 cm), some plastic
strings (green and white color, 30 cm in length, diameter = 2.5 mm), and a gray plastic carpet (12
x 50 cm). Green vegetables (broccoli and spinach, approximately 10g together) were scattered
over the plastic carpet to facilitate foraging. Commercial feed and live meal worms in plastic
cups were provided inside the TC to attract the birds. We assumed that hens would prefer the
enriched environment in the TC in the absence of an aversive stimuli.
5.3.3. Experimental design
Individual birds were initially assigned to receive each of four concentrations of CO2 (25%, 35%,
50%, 70%) in four rounds, and each bird acted as its own control. Birds received one gas
treatment per day, and were randomly assigned to gas treatment in the first round, then
systematically assigned to remaining gas treatments in subsequent rounds (Table 5.2). A
treatment round consisted of three consecutive days. On Baseline Day, both CC and TC were
maintained at ambient air conditions (<1% CO2). On Gas Day, TC was maintained at the
predetermined CO2 concentration, while CC was maintained at near ambient levels (<1%). On
Washout Day, TC and CC were again, both maintained at ambient air conditions (<1% CO2).
5.3.4. Training procedures
Home pen training was conducted during the 4-week acclimation period to habituate the birds to
researchers, handling, and to the act of acquiring meal worms by pushing through a curtain. A
covered area of 45 x 45 cm was established inside the home pen and the entrance to this area was
covered with the same curtain material used in the test apparatus. Meal worms were provided
Page 147
134
inside this covered area and hens were trained to enter the covered area by pushing through the
curtain.
The hens were individually trained to the experimental procedures within the apparatus. In all
tests and training sessions, feed was withheld from birds for one hour prior to training to increase
their motivation to access the food rewards in TC. As the first step, the bird was introduced into
the CC. The sliding door separating the two chambers was kept opened, after nearly 60 s the bird
was gently pushed through the slit in the middle of the curtain, and then allowed five minutes to
access the feed rewards and enrichments in the TC. On the second day, the bird was placed in
CC with the sliding door closed. After 5 minutes, the sliding door was opened, allowing access
to TC. If the bird did not enter TC within five minutes, a researcher gently guided it through the
curtain. Upon entry to the TC, the bird was again, provided five minutes during which it could
access the enrichments in the TC and freely move about the chambers. After the 5 minutes, the
bird was returned to the home pen.
All hens were trained individually in the apparatus for 5-6 consecutive days to achieve the
enrollment criteria for the testing phase. Training was concluded when the bird learned to
independently enter the TC on two consecutive days.
5.3.5. Testing protocol
Testing order of the birds was assigned based on their level of motivation to access the feed and
enrichment in treatment chamber during the training sessions. Hens who accessed the
enrichments fastest were placed first in the testing order in order to ensure longer feed
Page 148
135
deprivation for the less motivated hens. A health assessment was performed on each bird to
identify any wounds on the body prior to the test, and birds who were in good health were
eligible for the testing phase.
The testing procedures on Baseline Day and Washout Day were identical to those described for
final training with the exception that birds that chose to not enter TC after five minutes were not
guided through the sliding door; the test was concluded, and the bird was returned to the home
pen. On Gas Day, the TC was prefilled and stabilized at the designated CO2 treatment
concentration prior to placing each bird in the CC. If the bird did not enter the TC within the
given five minutes, the test was concluded, and the bird was returned to the home pen. When loss
of posture (LOP) occurred inside the TC, the bird was immediately removed and placed into a
recovery pen located within the testing room. During recovery, affected birds were observed
until they regained full sensibility and standing posture. Feed and live meal worms were
provided inside the recovery pen, and the bird was returned to the home pen after normal
foraging behaviour occurred. The apparatus was cleaned and disinfected with Accel
(ACCDISC1G-US, Virox Technologies Inc, Ontario, Canada) between tests, and the underpad
and feed enrichments were replaced in the TC.
5.3.6. Behavioural data collection
Behaviour data were collected via live observations and video recordings (Table 5.1). Frequency
of vocalizations in CC and TC, latency to fully enter TC, latency to loss of posture, and latency
to loss of neck tone were recorded during live observation. Live observations were collected by
two researchers, with Observer 1 sitting approximately 0.5 m from the hinged door of the TC
Page 149
136
from which the bird was visible. A black fabric curtain (2.1 m x 0.9 m) and lighting placement
were used to obscure Observer 1 from the bird’s view. Observer 1 recorded latencies using a
digital timer (National Presto IND. Inc., Eau Claire, WI). On Gas days, Observer 1 determined
time at loss of neck tone, and was responsible for moving the bird as quickly as possible from
TC the recovery pen. Observer 2 was positioned beside the chambers, out of the test bird’s view,
and operated the sliding door. Observer 2 listened for and counted all vocalizations using a
manual tally counter (Great Star Tally counter, Model 30665, Great Star Industrial USA LLC,
Huntersville, NC). All live observations were confirmed with video recordings, except for
vocalizations which were not audible by the digital recordings.
Continuous video was recorded using four color digital video cameras (Panasonic, Model WV-
CP-484, Matsushita Co. Ltd., Kadoma, Japan), which were positioned to provide views from the
top and side of the CC and TC. Cameras were fed into a multiplexer using a Noldus Portable Lab
(Noldus Information Technology, Wageningen, The Netherlands) that enabled capture of a dual
recording at 30 frames/s onto a computer using Handy AVI software (version 4.3 D, Anderson’s
AZcendant Software, Tempe, AZ). Data were collected from videos by a trained observer,
blinded to the animal ID, date and CO2 concentrations, using The Observer software (The
Observer version 10.1.548; Noldus Information Technology, Wageningen, The Netherlands). A
neutral individual performed the blinding procedures for the video recordings from all tests. The
blinding procedures involved cutting the video recordings to remove identification presented at
the beginning of each video, assigning a random number to each video segment and sorting for
the purpose of providing a random sequence in which videos were to be scored. Seven videos
were selected at random and duplicated within this sequence for the purpose of determining
Page 150
137
intra-observer reliability. Prior to data collection, the observer was trained to use the Observer
XT program by repeatedly scoring 2 videos and an ethogram from an unrelated CO2 study until
reaching an intra-observer reliability score of k ≥ 0.90 as calculated by the Observer program.
After reaching the desired level of competence, data collection began using the blinded videos
and ethogram shown in Table 5.1.
Latencies to first curtain pushing, first head insert, and full entry were calculated relative to the
time the sliding door was opened. Latencies for onset of ataxia and loss of posture were
calculated relative to the time of head insert at full body entry to TC. Duration was calculated as
total time recorded over all bouts during testing. Hens spent different amounts of time in CC
before entering the TC, and amounts of time spent in TC until loss of posture was also differed.
Therefore, frequency of vocalizations was calculated as a rate (number of vocalizations/minute)
for the analysis.
5.3.7. Statistical analyses
Fisher’s exact tests (SAS 9.4) were used to determine whether the number of hens that entered
TC and the number of hens that presented avoidance behaviours was independent of test day
(Baseline Day, Gas Day, Washout Day) using 2 × 3 tests. If the results showed significant
difference among the days, then pairwise comparisons between Baseline Day vs Gas Day (to
find any proximate effect), and Baseline Day vs Washout Day (to find any conditioned place
avoidance/ any learning behaviour) were performed for number of birds that entered TC
(yes/no), and presented behaviour (yes/no) using Fisher’s exact tests. The same analysis was
performed to determine whether entry to TC was independent of CO2 concentration (25%, 35%,
Page 151
138
50%,70%) using 2 × 4 tests. If the results showed significant difference among the CO2
concentrations, then pairwise comparisons between every combination of concentration (25% vs
35%, 25% vs 50%, 25% vs 70%, 35% vs 50%, 35% vs 70%, 50% vs 70%) were performed for
number of birds that entered TC (yes/no) using Fisher’s exact tests (Zar, 1998).
Frequencies and durations of behaviour during observations in CC and TC were analyzed by
using PROC GLIMMIX (SAS 9.4, SAS Inst. Inc., Cary, NC) with identity link function and
Gaussian distribution. Degrees of freedom and fixed effects standard errors were adjusted by
using Kenward-Roger method. Bird was included as a random effect.
The model for analyzing all behaviours observed in Control Chamber and Treatment Chamber
included the fixed effects of day (BD, GD, WD), CO2 concentration (25%, 35%, 50%, 70%,),
and round (1st, 2nd, 3rd, 4th). Interactions were not tested due to small sample size and too few
degrees of freedom available among the model factors.
The model for analyzing recovery behaviour inside the recovery pen included the fixed effects of
CO2 concentration (25%, 35%, 50%, 70% CO2).
Least square means estimates and standard errors (SE) are reported in the results. Least
significant means separation was conducted by using the Tukey-Kramer test.
Page 152
139
5.4. Results
Eighteen hens met the performance criteria for enrollment in the training phase within the home
pen. Among them, eight hens successfully completed apparatus training within 5 days. Four
other hens needed 6 days to complete the apparatus training. Therefore, 12 hens were enrolled in
the testing phase (1.2 ± 0.1 kg; 24 week-old).
Some birds died or failed to enter TC following gas exposure, resulting in uneven treatment
allocations (Table 5.2). One (F) hen never re-entered TC after her first gas experience. Another
(G) never entered TC after her 2nd gas experience, while 2 other hens (B and K) stopped
entering TC after their 3rd gas experience. Two birds (L and H) died during exposure to 50% and
70% CO2, respectively, during the 3rd round. Therefore, only 3 hens completed all 4 treatment
rounds.
5.4.1. Approach and avoidance behaviour
The numbers of hens that entered TC differed across days (p= 0.001; Table 5.3). According to
pairwise comparisons, fewer hens entered TC on Gas Days than Baseline Days (P<0.001) and on
Washout Days than Baseline Days (P<0.001). The numbers of hens that entered TC at the
different CO2 concentrations are presented in Table 5.4. The number of hens that entered TC on
Gas Day differed for different CO2 concentrations (P=0.028) with fewer birds entering TC as
CO2 concentration increased. Pairwise comparisons showed that the number of hens that entered
TC on Gas Day was significantly different between 25% and 70% CO2 (P=0.035), and between
35% and 70% CO2 (P=0.035). None of the birds that entered TC on Gas Day returned to CC
although they were able to freely move between the two chambers.
Page 153
140
The hens who did not fully enter TC on Gas Day always pushed at the curtain and/or inserted
their head into the TC (Table 5.3). In this way, they were exposed to CO2 concentrations but
chose not to enter. The proportion of hens who demonstrated head insert into TC without
entering into TC was significantly higher for Gas Days than Baseline Days (P=0.025). The
proportion of hens who showed curtain pushes without head insert and entering into TC tended
to be higher on Gas Days than baseline Days (P<0.054). Some hens also performed curtain
pushes and/or head inserts into TC on Washout Day without entering into TC (Table 5.3). More
hens pushed the curtain without head insert and entering into TC on Washout Day than Baseline
Days (P<0.001).
The numbers of hens that displayed approach/avoidance behaviours in the Control Chamber at
the different CO2 concentrations are presented in Table 5.4. The proportion of hens that entered
the TC on Gas Day was significantly lower for 70% CO2 compared to 25% and 35% CO2
(P=0.035). The proportion of hens that displayed other approach/ avoidance behaviours (e.g.
curtain pushes, head inserts, head withdrawals), and other behaviours (e.g. defecation,
vocalization, head shaking, open mouth breathing) did not differ between the CO2 concentrations
(Table 5.4).
Some hens who entered to TC on Gas Day did not enter in to TC on Washout Day. Number of
birds entered to TC on Washout Day was same for 25%, 35%, and 50 % CO2. Proportion of hens
(number of hens entered to TC on Washout Day out of number of hens entered to TC on Gas
Day) who showed conditioned place avoidance did not differ (P>0.05) among the CO2
Page 154
141
concentrations (3/8 for 25%; 3/8 for 35%; 3/6 for 50% CO2). Two birds entered to 70% CO2 and
one hen died. The remaining hen entered the TC on Washout Day.
Assessment of the motivation to enter the TC was based on latency to first push the curtain, the
frequency of curtain pushes, latency to first instance of the inserting head into the TC, frequency
of inserting head into the TC, and latency to full entry into the TC. Frequencies and latencies of
approach/avoidance behaviours are presented in Table 5.5. The frequency of curtain pushes was
higher on Washout Days (9.0 ±1.3) than Baseline Days (2.8 ± 1.2; P=0.0046). Hens
demonstrated more curtain pushes in the 4th round in comparison to 1st and 2nd rounds and this
tended to be significant (P=0.0562). Latency to first push the curtain and latency to first head
insert were significantly longer in the 4th round compared to the 2nd round (P=0.0286 and
P=0.0336 respectively). Latency to full entry to TC tended to be different among the rounds
(P=0.0543) with longer latencies in 3rd and 4th rounds.
5.4.2. Other observed behaviours
Open mouth breathing was observed only on Gas Day (Table 5.3) and by all the birds that
entered the TC (Table 5.4). The average frequency ± SD of open mouth breathing (the number of
gasps for the entire time) was 6 ± 2 in 25% CO2 (n=8), 5 ± 2 in 35% CO2 (n=8), 3 ± 1 in 50%
CO2 (n=6), and 2 ± 0.7 in 70% CO2 (n=2). Hens demonstrated head shaking on all 3 test days.
More hens demonstrated head shaking inside the TC on Gas Days than Baseline Days (P=0.001).
All hens that entered the gas showed head shaking, and this was not different between
concentrations (Table 5.4). The average frequency ± SD of head shaking until loss of posture
Page 155
142
inside the TC was 9.5 ± 3 in 25% CO2 (n=8), 9 ± 3 in 35% CO2 (n=8), 7.7 ± 3.9 in 50% CO2
(n=6), and 6 ± 2.8 in 70% CO2 (n=2).
The number of hens that vocalized on different days, and different CO2 concentrations are
presented in Table 5.3 and Table 5.4, respectively. The proportion of hens that vocalized inside
the CC did not differ for all 3 tests days and CO2 concentrations. The proportion of hens that
vocalized inside the TC was higher for the Gas Day in comparison to Base line Day (P=0.004)
but no difference was found between Washout Day and Baseline Day (P=0.42). Rates of
vocalizations (mean vocalizations per minute) before the sliding door was opened did not differ
among the 3 different test days (P=0.48), CO2 concentrations (P=0.12), and rounds (P=0.30). The
rate of vocalizations after the sliding door was opened among the 3 different test days tended to
be different (P=0.08), and a higher rate was observed on Washout Days (5.6 ± 1.3 per minute)
than Baseline Days (2.1 ± 1.2 per minute). However, no differences were found in the rate of
vocalizations after the sliding door was opened for CO2 concentration (P=0.76) or round
(P=0.46). Hens demonstrated higher rates of vocalizations inside TC on Gas Days (4.1 ± 0.7 per
minute) than Baseline Days (0.2 ± 0.5 per minute) and Washout Days (0.5 ± 0.8 per minute;
P=0.0001). Rate of vocalizations inside the TC did not differ between CO2 concentrations
(P=0.79) or rounds (P=0.19).
Duration of standing active in the CC was significantly longer (P=0.0011) on Gas Days (76±16
s) and Washout Days (98±15 s) than Baseline Days (20±14 s). Duration of standing active in the
CC tended to be significant for CO2 concentrations (P=0.0990). Hens performed standing active
in the CC for longer durations at 50% CO2 (95±16 s) than 25% CO2 (37±18 s) on Gas Days.
Page 156
143
Duration of standing active in CC did not differ among the rounds (P=0.41). Duration of
standing inactive in the CC was not different among the CO2 concentrations (P=0.2476), or
rounds (P=0.13). Duration of standing inactive in the CC was higher in 4th round (122 ± 27 s) in
comparison to the 1st round (36 ± 16 s; P=0.04). Except for two hens, all the other hens did not
sit inside the CC on any of the 3 test days. However, some hens demonstrated sitting inside CC
during the retraining period. Escape behaviour was absent in all hens.
5.4.3. Parameters associated with loss of sensibility
All the birds that entered TC on Gas Day showed ataxia and loss of posture irrespective of CO2
concentration. Mean time ± (SE) s for latencies (ataxia, loss of posture) and durations (ataxia)
are presented for 25%, 35%, and 50% in Table 5.6. Latency to onset of ataxia was tended to
decrease as concentration increased (P=0.09). Longer latency to onset of loss of posture was
observed at 25% CO2 than 50% CO2 (P<0.05). Table 5.7 shows behaviour observed during
recovery from exposure to CO2. Hens that lost sensibility at 50 % CO2 demonstrated the longest
average time to raise their head (175 ±11 s, P<0.0001), stand up (510 ± 77, P=0.0118), walk
(1077 ± 145; P=0.0006), and begin feeding (1346 ± 58, P<0.0001) compared to the birds
exposed to 25% and 35% CO2.
Only one hen who entered 70% CO2 was recovered and the relevant data inside the recovery pen
are shown in the Table 5.7. One hen that entered at 50% CO2 and another that entered at 70%
CO2 in their 3rd round died inside the recovery pen without regaining sensibility.
Page 157
144
5.5. Discussion
In approach avoidance paradigms, the motivation to approach a desirable stimulus is gauged
against the motivation to avoid an unpleasant stimulus. Results are expected to vary according to
strengths of competing motivations.
In the current study, all hens who were eligible for the apparatus training were able to learn the
assigned task within 5-6 days. Therefore, the assigned task to the hens (entering the TC pushing
through a curtain) is within hens’ cognitive capacity, and hens were sufficiently rewarded inside
the TC. Hens were able to see through the curtain which was between the CC and TC, and could
easily push through it to enter the chamber. In the current study, all the hens were interested in
meal worms. Upon entering the chamber, they first approached the meal worms and spent time
eating them. They were also interested in the commercial feed and spent the second longest time
on feeding. This suggests that the one hour feed was withheld induced feeding motivation of the
hens. None of the hens went back to CC on Baseline Days during the given 5 minutes, indicating
their willingness to stay inside the TC, and a preference for it compared the CC. All hens entered
TC in less than 60s on Baseline Days in the first round. We suggest that all hens were motivated
to enter the TC to acquire the provided feed and enrichments.
In the current study, refusal to enter the TC on Gas Day reveals that the motivation of a bird to
avoid the aversive environment inside TC was higher than the motivation to acquire the rewards
inside the TC. Except for one bird, all birds voluntarily entered to 25% and 35% CO2. The same
bird avoided 35% and 25% CO2 in its 1st and 2nd rounds respectively, and this might be due to
the difference in individual sensitivity of the hens. However, this bird entered into 50% CO2 in
Page 158
145
its 3rd round and died on Gas Day without regaining consciousness. The reason that this bird
avoided lower CO2 concentrations but entered at higher CO2 concentration is difficult to explain.
Overall, we observed that the number of hens entering into the TC decreased with increasing
CO2 concentrations. Webster and Fletcher (2004) observed similar results in layer hens in an
approach avoidance test, which used 30%, 45%, and 60% CO2 in a feeding chamber. The authors
reported that the percentage of birds that entered the feeding chamber decreased at higher
concentrations of CO2, and further suggested that hens could detect the presence of CO2 in the
corridor leading to the feeding chamber and tended to hesitate to push through the entrance of the
gas chamber as a result. In the current study, nearly half of the birds tested at 50% CO2 avoided
entering the TC on Gas Day indicating that half of the hens were able to detect 50% CO2 and
avoided it. Sandilands et al. (2011) assessed 50%, 55%, and 60% CO2 in broilers and reported
that all gas levels were aversive to some extent in broiler chickens based on time spent with head
in a feeding station filled with CO2 gas. Further the authors explained that 60% CO2 was
evidently the most aversive option among the gas mixtures tested, but even this concentration of
CO2 was not totally refused by broilers. In the current study, only two of 7 hens entered into 70%
CO2. Therefore, we suggest that 70% CO2 is relatively more aversive to layer chickens based on
avoidance. In the current study, 25% was the lowest CO2 concentration tested, and this is slightly
higher than the proposed aversion threshold level of 25% (McKeegan et al. 2003) for the
chickens. Except one, all hens chose to enter into 25% and 35% CO2 levels. Therefore, we
suggest that hens entered 25% and 35% CO2 with minimal or no aversion and that their
motivation to access the rewards inside the TC was higher than the motivation to avoid 25% and
35% CO2.
Page 159
146
The number of curtain pushes and head withdrawals before entering or avoiding TC on Gas Day
could be considered to reflect conflict or competing motivation between entering TC to acquire
the resources and avoidance of gas. All hens that did not enter into 50% CO2 showed a number
of curtain pushes and head withdrawals. Except for two hens, all other hens who did not enter
into 70% CO2 demonstrated curtain pushes without inserting their head into TC. The absence of
head insert into TC could be due to higher aversiveness of 70% CO2. All birds did push the
curtain several times, and many birds made head withdrawals prior to avoiding 50% and 70%
CO2 concentrations on the Gas Day, indicative of several attempts to enter the TC. Head
withdrawals on Baseline Day were only observed in 4 hens in their later rounds of testing.
Except for two hens at 25%, and two hens at 35% CO2, all other hens who voluntarily entered
into either 25% or 35% CO2 did not demonstrate head withdrawals prior to entering TC,
supporting our suggestion that these two concentrations were only minimally aversive to the
hens. However, it is difficult to say whether layer hens can detect 25% and 35% CO2 or not
based on the current results of the approach-avoidance test. Raj and Gregory (1991) reported
concentrations above 7.5% CO2 were detectable to laying hens and given a free choice, the hens
learned to avoid such atmospheres. In contrast, all except for one hen entered into 25% and 35%
CO2 in the current study.
In the current study, we hypothesized that a bird that entered into the gas and experienced loss of
consciousness would demonstrate conditioned place avoidance (CPA). Although the majority of
hens entered at 25% and 35% CO2, some of those hens did not enter TC on Washout Day.
Among them, one hen that entered 25% needed 4 days of retraining, and another hen that entered
35% CO2 needed 8 days of retraining to resume entering the TC voluntarily, providing evidence
Page 160
147
that they demonstrated CPA. None of the hens who entered and lost consciousness at 50% CO2
entered TC on Washout day, and all needed 4-5 days of retraining before re-entering TC
voluntarily. Moreover, all hens who demonstrated CPA did not insert their head into TC on
Washout Day and some did not even push the curtain. Many of the birds who demonstrated CPA
vocalized 50-90 times (within the given 5 minutes) after the sliding door was opened on
Washout Days showing evidence of either fearfulness or conflict to reenter the TC. Therefore, I
observed a trend of higher rate of vocalizations after the sliding door was opened on Washout
Days in comparison to Baseline Days.
The majority of hens who lost consciousness at 25% and 35% CO2 did not demonstrate CPA,
and this could be due to less aversion experienced at these two CO2 concentrations prior to loss
of consciousness. In this study, three birds who experienced 50% CO2 (by pushing the curtain
and head inserting) without full entry to the TC on Gas Day never reentered the TC, either on the
Washout Day or the next 4 days of retraining. Three other birds tested at 70% CO2 pushed the
curtain several times (two birds inserted their heads once into TC) but did not enter the TC on
Gas Day, and they did not enter the TC on Washout Day. Therefore, we suggest that exposure to
50% or higher CO2 concentration by pushing the curtain and/or inserting their heads into the TC
without experiencing loss of consciousness was enough to create CPA in the laying hens.
McKeegan et al. (2005) reported that chickens detected CO2 at 10% while Raj and Gregory
(1991) reported that hens can detect CO2 concentrations at 7.5%. Gerritzen et al. (2007)
compared the behaviour of ducks, broilers, laying hens and turkeys to assess differences in
susceptibility among species following exposure to CO2. increasing from 0% to 45% CO2 at a
Page 161
148
rate of 14 l/min. The authors reported that the first signs that birds noticed a change in their
environment was recorded from as little as 2% CO2 in the air within the test box for broilers,
ducks and turkeys which was at a significantly (P<0.05) lower concentrations than for laying
hens (6.6% CO2). The used least concentration of 25% CO2 in the current study is higher than
the detection levels in chickens. Thus, it is suggested that all tested concentrations were detected
by the hens.
Breathlessness is induced in humans when they exposed to high CO2 concentrations (Gregory
and Wotton, 1990b). Breathlessness is a negative experience and can activates the same regions
of the brain as other negative sensory experiences such as pain, hunger, and thirst in mammals
(Beausoleil and Mellor, 2015). Breathlessness is described in terms of its 3 different qualities:
respiratory effort, air hunger, and chest tightness (Beausoleil and Mellor, 2015). Respiratory
effort and air hunger are induced in CO2 euthanasia. Lack of available O2 resulted in tissue
hypoxia in the respiratory muscle, thus decreasing the muscle activity. Therefore, respiratory
effort (an increased conscious effort for respiration) is induced to achieve the necessary or
desired level of ventilation for the respiratory muscular functioning and respiratory ability
(Beausoleil and Mellor, 2015). Air hunger is the increased urge to breathe. Air hunger is more
unpleasant than respiratory effort (Banzett et al., 2008) and induced by hypercapnia and
hypoxaemia when there is no proper coordination between increased automatic command to
breath and degree of lung inflation (Beausoleil and Mellor, 2015). In humans, signs of asphyxia
and behavioral excitation have been observed because of both hypercapnia and hypoxia (Forslid
et al., 1986). McKeegan et al. (2005) conducted a behaviour study, which hens were exposed to
Page 162
149
short pulses of gas and their responses were observed to indicate aversion, and concluded that
aversion to CO2 was seen at 24%.
Exposure to CO2 could be distress to poultry because it may result a sense of breathlessness.
Different behaviour studies have been conducted to assess the aversion to CO2 in poultry species.
Gerritzen et al. (2000) suggested gasping and head shaking in broiler chickens exposed to 30%
and 60% CO2 as indicators of breathlessness and the pungency of gas mixtures. Webster and
Fletcher (2001) also observed head shaking and deep breathing (breathing through the mouth) in
layer chickens and broilers exposed to different CO2 concentrations (30%, 45%, 60%).
McKeegan et al., (2005) reported respiratory disruption (apparent increased inhalation depth and
duration) in broiler chickens exposed CO2 concentrations (10, 25, 40, 55, 70%) for 10 s. It is
reasonable to say all tested concentrations in the current study were aversion to the hens up to
some extent. Hens demonstrated open mouth breathing in all CO2 concentrations, suggesting that
they were experienced air hunger and may be with respiratory efforts in lower concentrations.
McKeegan et al. (2004) reported that the nasal and oral mucosa of the chicken contain trigeminal
polymodal nociceptors that can detect and transmit information relating to the presence and
concentration of potentially noxious chemical stimuli (ammonia, CO2 and acetic acid vapours).
The authors further suggested that the avian nociceptive threshold is in the region of 40 - 50%
CO2, which is in the same range as the electrophysiological thresholds reported for rats and
humans (Anton et al., 1991, 1992). Therefore, low levels of CO2 such as 25 % seems to be
probably air hunger but high levels are probably pain, possibly with air hunger combined.
Page 163
150
All hens in the current study entered the TC on Gas Day and showed head shaking and open
mouth breathing irrespective of CO2 concentration. Raj (1996) studied the degree of aversiveness
to 72% CO2 in air in turkeys by using 1-5 scale system based on behaviours of head shaking,
gasping and vocalization. He stated that all three behaviours were clearly observed in all turkeys
in the CO2 environment. However, interpretation of head shaking on animal welfare is
controversial. Head shaking has been described as indicative of an aversive reaction to CO2 and
respiratory distress in poultry (Raj and Gregory, 1994). However, other researchers suggested
head shaking as an alerting response to unexpected or novel events in poultry (Hughes, 1983) or
an attempt to regain an alert state when birds are feeling dizzy or start to lose consciousness
during the exposure to CO2 (Webster and Fletcher, 2001; Gerritzen et al., 2007). We observed
head shaking in some hens on Baseline Days and Washout Days when the CO2 concentration
was the same as ambient air. Therefore, the argument that head shaking is an aversive reaction to
CO2 or that it indicates respiratory distress is not supported. McKeegan et al. (2005) reported
that gustatory mandibulation was associated with head shaking, and suggested that the acidity of
CO2 may provide a gustatory stimulus which causes head shaking. Significantly more hens
showed head shaking on Gas Days than Baseline Days for all CO2 concentrations. Therefore,
hens are more likely to perform head shaking when exposed to CO2.
By rotating the same hens through different CO2 concentrations, there is a possibility that
experience of one CO2 concentration could carry over to influence the results of a test with
another CO2 concentration. Webster and Fletcher (2004), who also used repeated tests at
different concentrations of gas in laying hens, explained the possibility of carry-over effect due
to hens being exposed to more than one stunning gas atmosphere, and the suggested reasons
Page 164
151
were inability of complete physiological recovery between tests, or learned responses associated
with recovery from loss of consciousness or from exposure to the stunning atmosphere itself. In
the current study, there was a significant effect of round in some observed behaviours. Hens
tested in 50% and 70% CO2 in their previous rounds demonstrated longer latency to insert head
into TC and fully enter into TC for even lower concentrations of 35% and 25% CO2 in 3rd and
4th rounds. Overall, latency to first push the curtain was longer in 3rd and 4th rounds and this
could be also due to previous aversive experience in the TC. A higher number of curtain pushes
were also observed in the 4th round. This may be due to hens testing the air in TC due to their
previous experience with higher CO2 concentrations. Head withdrawals (2-3 times) on Baseline
Day were only observed in 4 hens in their 3rd round and one hen in its 2nd round. The hens who
demonstrated head withdrawals for 25% and 35% CO2 were tested for 50% and 70% CO2 in
their previous rounds, and this previous experience may be the reason for head withdrawals prior
to entering the lower concentrations of 25% and 35% CO2. More hens demonstrated CPA in the
3rd round for different CO2 concentrations compared to the first and second rounds, and needed
retraining, indicating that more hens learned to avoid TC after the 3rd gas experience.
For optimal welfare of the animal, the goal is to achieve insensibility and loss of consciousness
as quickly as possible, for the process to be pain-free, and for death to follow quickly. CO2
causes hypercapnia, suppressing afferent transmission of sensory stimuli to the cortex of the
brain, producing anesthesia (Forslid et al., 1986; Raj et al., 1992; Raj and Gregory, 1993). Loss
of posture (LOP) has been proven to be the reliable indicator of onset of unconsciousness in
poultry based on electroencephalography (EEG) studies in broiler chickens (Coenen et al., 2000;
Gerritzen et al., 2004; Benson et al., 2012). In the current study, onset of ataxia occurred
Page 165
152
regardless of CO2 concentration. However, there was a tendency of decreasing latency to onset
of ataxia with increased CO2 concentrations. Latency to onset of LOP was significantly different
among the CO2 concentrations. Longer latency to onset of LOP was observed in 25% CO2 than
50% CO2. Similar to the current study, previous research reported shorter times to LOP in higher
CO2 concentrations. Webster and Fletcher (2001) reported that higher CO2 concentrations
reduced the time to loss of consciousness based on time to LOP in layer hens and broilers.
Gerritzen et al. (2007) reported slight and occasionally significant differences in time to LOP
between different poultry species following exposure to CO2 treatments that increased from 0%
to 45% at a rate of 14 l/min. Broilers lost posture at 19.0% CO2, which was significantly
different than in ducks (23.8% CO2), while loss of posture occurred in layers and turkeys at
19.9% and 19.3% CO2, respectively. Results of our first study with turkeys (Bandara et al.,
unpublished data) are in accordance with these findings, where laying hens showed faster LOP in
comparison to turkeys. In the current study we could not observe loss of neck tone, an indicator
of loss of posture (Sandercock et al., 2014) in hens because the time between LOP and loss of
neck tone was short (~2 s) but in the turkeys, recording of loss of neck tone was possible due to
longer time (> 25 s) between LOP and loss of neck tone. Therefore, these findings revealed the
need to set different criteria for different poultry species using CO2 as a humane killing agent.
Recovery pen data revealed that birds that lost consciousness at 50% CO2 demonstrated the
longest time to raise their head, stand up, walk, and start feeding compared to the birds exposed
to 25% and 35% CO2. Overall, all hens regained consciousness within 6 minutes. We suggest at
least ~12 minutes to observe hens to confirm that they are dead following CO2 euthanasia if
using 25% or above CO2 concentration.
Page 166
153
5.6. Conclusions
The majority of hens avoided entering into 50% and 70% CO2. Except for one, all hens entered
into 25% and 35% CO2. Therefore, hens found 50% and 70% CO2 concentrations more aversive
than 25% and 35% CO2 concentrations. Hens demonstrated conditioned place avoidance for all
CO2 concentrations and needed several days of retraining after gas experience to start voluntarily
entering into TC irrespective of CO2 concentration. Therefore, all concentrations tested in the
current study were aversive to a certain extent. Loss of posture indicating loss of consciousness
occurred in less than 20s in all CO2 concentrations. Latency to LOP was shorter in 50% than
20% CO2.
Page 167
154
Table 5. 1: Definitions of behaviours recorded in the Control Chamber (CC) and Treatment
Chamber (TC)
Measure Variable type Definition
States
Standing inactive v Duration Bearing weight on feet without locomotion
Standing active v Duration Bearing weight on feet with locomotion
Sitting v Duration Breast is touching the chamber ground
Exploring v Duration Pecking floor or walls (curtains excluded). Can
be done while sitting, standing or in locomotion
Events
Vocalization D Frequency Vocalizations occur in the CC and TC
Push curtain VD Latency,
Frequency
First time beak contacts with curtain; this may
include pushing into the TC but head does not
cross plane of curtain
Insert head VD Latency,
Frequency
Head crosses plane of curtain into TC with or
without neck
Withdraw head V Latency,
Frequency
Head and beak return to CC after curtain contact
Full entry VD Latency Bird enters TC with both feet
Head shaking V Frequency Rapid lateral movement of the head
Open mouth
breathing V
Latency,
Frequency
Bird breath through their mouth (beak is opened
and chest movements occur)
Wing flapping V Duration Coordinated movement with rising and lowering
wings without feet losing contact with the
ground.
Ataxia V Latency,
Duration
Uncoordinated movement, while bearing weight
on one or more legs
Loss of posture VD Latency Non-weight bearing on feet; may occur rapidly
(collapse) after ataxia. Head righting response
present
Escape V Frequency Leaping or climbing; feet left floor with wing
flapping
V Measured via video recordings
D Measured via direct observations. Vocalizations were recorded separately before the sliding
door open, after the sliding door open, and after entry to the Treatment Chamber
Page 168
155
Table 5. 2: Test order, assigned CO2 treatment and outcomes in each of the four rounds for the
12 hens enrolled in the study.
Order of
testing Bird ID CO2 concentration (%)
1st Round 2nd Round 3rd Round 4th Round
1 A 70r 50r 35e 25e
2 B 25e 70e 50r* -
3 C 35e 25e 50ec* -
4 D 25e 35ec 70r 50r
5 E 50r* 35e 25ec* -
6 F 35ec* - - -
7 G 70r 50ec* - -
8 H 25e 50e 70ed -
9 I 70r 35ec 50ec* -
10 J 50e 70r 35e 25ec
11 K 50r* 35e 25ec* -
12 L 35r 25r 50ed -
e Birds entered TC on Gas Day. r Birds did not enter TC on Gas Day. c Birds did not enter TC on Washout Day d Birds did not recover from CO2 exposure on Gas Day (died)
*Birds required retraining (did not enter on subsequent Baseline Day)
Page 169
156
Table 5. 3: Number of times laying hens demonstrated different behaviour patterns in the
Control (CC) and Treatment Chamber (TC) on the three different test days (Baseline Day, Gas
Day, Washout Day)1
Baseline Day
(n=36)
Gas Day
(n=36)
Washout Day
(n=34)
Approach/avoidance Behaviours
Fully entered into TC 36ac 24b 18d
Head insert only 0b 6a 2
Pushed curtain only 0bd 5a 14c
Neither head insert nor curtain
pushes
0 1 1
Head withdrawals 4 9 4
Other behaviours
Defecation in CC 5 11 10
Vocalization in CC-before door open 11 9 10
Vocalization in CC- after door open 8 5 13
Inside Treatment Chamber (n=36) (n=24) (n=18)
Vocalization 5b 12a 1
Defecation 0 0 1
Open mouth breathing 0b 24a 0
Head shaking 4b 24a 4
1 Pairwise comparisons of Baseline Day vs Gas Day and Baseline Day vs Washout Day were
analyzed by Fishers’ exact test (P<0.005).
a,b Different superscripts indicate proportion of birds presenting the measure is different between
Baseline Day and Gas Day according to Fishers’ exact tests (P<0.005)
c,d Different superscripts indicate proportion of birds presenting the measure is different between
Baseline Day and Washout Day according to Fishers’ exact tests (P<0.005)
Page 170
157
Table 5. 4: Number of laying hens that demonstrated different behaviour patterns in the Control
(CC) and Treatment Chamber (TC) at the different CO2 concentrations on Gas Days.
Numbers within a row with different superscripts indicate proportion of birds presenting the
measure is different among the concentrations according to Fishers’ exact tests (P<0.005)
CO2 concentration
25%
35%
50%
70%
(n=9) (n=9) (n=11) (n=7)
Approach/avoidance Behaviours
Fully entered into TC 8a 8a 6ab 2b
Head insert only 0 0 4 2
Pushed curtain only
Neither head insert nor curtain pushes
0
1
1
0
1
0
3
0
Head withdrawals 2 1 4 2
Other behaviours
Defecation in CC 2 2 4 3
Vocalization in CC- before door open 3 2 4 0
Vocalization in CC- after door open 1 0 3 1
Inside Treatment Chamber (n=8) (n=8) (n=6) (n=2)
Open mouth breathing 8 8 6 2
Head shaking 8 8 6 2
Vocalization 4 4 4 0
Page 171
158
Table 5. 5: Frequencies and latencies of approach /avoidance behaviours (mean ± SE) demonstrated by laying hens in different test
days, CO2 concentrations, and cycles.
Latency to first
push the curtain
(s)
Frequency of
curtain pushes
Latency to first
head insert TC
(s)
Frequency of
head insert TC
Latency to full
entry to TC
(s)
Day
Baseline Day 13.3 ± 3.6 2.8 ± 1.2b 17 ± 8 1.3 ± 0.1 26 ± 10
Gas Day 9.3 ± 3.8 6.6 ± 1.3ab 42 ± 10 1.2 ± 0.2 46 ± 13
Washout Day 17.2 ± 3.9 9.0 ± 1.3a 37 ± 12 1.0 ± 0.2 47 ± 14
P value 0.3491 0.0046 0.1450 0.5578 0.3485
CO2 concentration
25% 8.6 ± 4.3 4.3 ± 1.6 13 ± 11 1.5 ± 0.2 25 ± 13
35% 11.9 ± 4.4 6.2 ± 1.5 42 ± 11 1.1 ± 0.2 46 ± 13
50% 15.3 ± 3.9 7.9 ± 1.4 45 ± 10 1.3 ± 0.2 47 ± 14
70% 17.3 ± 4.7 4.9 ± 1.7 14 ± 13 0.8 ± 0.3 25 ± 18
P value 0.5303 0.3597 0.0736 0.2030 0.5106
Round
1st 14 ± 3ab 5.1 ± 1.3 23 ± 10ab 1.2 ± 0.1 29 ± 12
2nd 6 ± 4b 4.5 ± 1.4 17 ± 10b 1.0 ± 0.1 19 ± 12
3rd 16 ± 5ab 6.4 ± 1.5 43 ± 11ab 1.3 ± 0.2 62 ± 13
4th 29 ± 7a 12.4 ± 2.5 75 ± 20a 1.4 ± 0.4 72 ± 27
P value 0.0286 0.0562 0.0336 0.6531 0.0543
Different letters indicate statistically significant (P<0.05) differences between means within the group of comparisons.
Bold numbers indicate significance results or tend to be significance results (P<0.05)
Page 172
159
Table 5. 6: Behaviours associated with loss of consciousness on Gas Day in laying hens: Mean
time ± SE s are presented for 25%, 35%, and 50% CO2 concentrations. Average time ± SD s are
presented for 70% CO2 concentration.
CO2 concentration
25% 35% 50% P value 70%
(n=8) (n=8) (n=6) (n=2)
Latency to ataxia 13 ± 0.8 11 ± 0.8 10 ± 0.9 0.096 11 ± 1*
Latency to loss of posture 19 ± 1a 17 ± 1ab 13 ± 1b 0.022 17 ± 3*
Different letters indicate statistically significant (P<0.05) differences between concentrations
within a row
*Did not statistically analyze due to only two data points
Page 173
160
Table 5. 7: Time (s) at onset of behaviour during recovery in laying hens exposed to different
CO2 concentrations. Mean time ± SE s are presented for 25%, 35%, and 50% CO2 concentrations
CO2 concentration
Behaviour 25%
(n=8)
35%
(n=8)
50%
(n=5)
P value 70%
(n=1)
Raised head 35±9b 37±9b 175±11a <0.0001 327*
Stand up 273±60b 192±60b 510±77a 0.0118 1703*
Start walking 267±115b 179±115b 1077±145a 0.0006 1803*
Feeding 295±45b 312±58b 1346±58a <0.0001 1860*
*Did not statistically analyze due to only one data point available
Different letters indicate statistically significant (P<0.05) differences between concentrations
within a row
Page 174
161
Figure 5. 1 : A- Lateral camera view of a laying hen inside control chamber (CC) waiting for the
sliding door (S) to open and to enter treatment chamber (TC). Wire mesh (W) can be seen on top
of the CC. B- Lateral camera view of hen pushing through the curtain (C) to enter treatment
chamber (TC) to access meal worms. Both images were collected on a Baseline Day, with
ambient air conditions in CC and TC.
W
S
C
A B
Page 175
162
CHAPTER 6
General Discussion
On-farm euthanasia is a common task within the poultry industry. The traditional methods by
which poultry are killed on-farm need to be evaluated for humaneness due to the increasing
concern for animal welfare. Additionally, new euthanasia methods continue to be developed,
often without scientific assessment for their effectiveness and humaneness. Therefore, there is an
urgent need to develop appropriate euthanasia methods that cause rapid, irreversible insensibility
and minimum pain or distress (AVMA, 2013; NFACC, 2016).
In this thesis, I assessed the efficacy of three physical euthanasia methods (non-penetrating
captive bolts; manual cervical dislocation; mechanical cervical dislocation), and the aversion to
different CO2 concentrations in layer chickens. The methods were compared for effectiveness
and humaneness allowing for recommendations to be made.
6.1. Evaluation of physical on-farm euthanasia methods
6.1.1. Humaneness of the euthanasia methods
In Chapters 3 and 4, I assessed three different commercially available non-penetrating captive
bolt (NPCB) devices and a mechanical cervical dislocation device (KED) in comparison to
manual cervical dislocation, respectively. Overall, all three non-penetrating captive bolts caused
rapid loss of sensibility in all age groups. All eye reflexes and rhythmic breathing were abolished
in ~5 s of application for all NPCB devices. Loss of jaw tone and neck muscle tone were
observed at ~ 30 s, while time to last movement and cardiac arrest occurred at less than 250 s.
Page 176
163
Previous research also reported rapid insensibility in poultry killed by similar NPCB devices
(Erasmus et al., 2010b; Woolcott et al., 2018a; Gibson et al., 2018).
Chapter 4 revealed that awake chickens killed by manual cervical dislocation demonstrated a
loss of pupillary light reflex at ~85 s, indicating that they may be sensible for a longer time in
comparison to birds killed by NPCB devices. Anesthetized birds killed by KED demonstrated
loss of pupillary light reflex at ~140 s, and this value is longer than for conscious birds killed via
NPCB devices and manual cervical dislocation. I found an anesthetic effect on eye reflexes and
other behavioural responses. Time to loss of pupillary light reflex was significantly reduced in
anesthetized birds compared to awake birds killed by manual cervical dislocation. The results
revealed a shorter latency to loss of pupillary light reflex indicative of brain death in the
anesthetized birds killed by manual cervical dislocation than in the anesthetized birds killed by
KED. Previous studies also revealed shorter latencies to insensibility and brain death in poultry
killed with manual cervical dislocation compared to other different mechanical cervical
dislocation devices (chickens killed by semark neck pliers: Gregory and Wotton, 1990a; turkeys
killed by burdizzo: Erasmus et al., 2010b; turkeys killed by KED: Woolcott et al., 2018b and
Jacobs et al., 2019; layer chickens killed by KED: Hernandez, 2018).Overall, the ability to cause
rapid loss of sensibility, shown through loss of reflexes and behaviours, was faster in birds killed
with NPCB devices than the cervical dislocation methods. Erasmus et al., (2010b) also reported
shorter latencies to insensibility in turkeys killed by a NPCB than by manual cervical dislocation.
However, a few birds failed to lose sensibility within 60 s for the Zephyr-E device in the current
study, and the cause was suspected to be due to a mechanical failure of the air compressor.
Results of the current study revealed that device success was 100% for TED and 99% for the
Page 177
164
Zephyr-EXL. Therefore, NPCB devices were shown to be relatively consistent in killing layer
chickens in an on-farm context. I suggest NPCB devices are more humane for killing of layer
chickens than the cervical dislocation methods.
AVMA (2013) guidelines indicate that cervical dislocation must result in luxation of the cervical
vertebrae without primary crushing of the vertebrae and spinal cord. Previous research reported
prevalence of vertebral fractures during mechanical cervical dislocation. Bader et al. (2014)
reported cervical fractures (in C1 and C2) in 43% of turkeys killed by using a non-penetrating
custom-made forceps with three blunt shear arms, and in 33% of turkeys killed by manual
cervical dislocation. Further, the authors reported cervical fractures (in C1 and C2) in 48% of
chickens killed by manual cervical dislocation. However, all the birds were stunned by a sharp
blow to the head from a wooden pole prior to cervical dislocation. Thus, the observed cervical
fractures may not be solely due to cervical dislocation and some may have resulted from the
blunt force trauma to the head. Woolcott et al. (2018b) found cervical fractures in 78% of 3
weeks old turkeys killed by mechanical CD (KED model-S), compared to 44% of turkeys killed
by manual CD. In Chapter 4, I found comparatively less vertebral fractures both in the layer
chickens killed by manual CD (6%) and KED (16%), which indicates differences across specific
cervical dislocation tools and across poultry species. When AVMA guidelines were established
in 2013, no evidence was available on cervical fractures in poultry killed by manual cervical
dislocation. In my study, cervical fractures were found in layer chickens killed by manual
cervical dislocation for the first time. These cervical fractures were found in 12-29 weeks old
birds and not in 60-75 weeks old birds. Thus, further studies need to assess the fractures in
different age groups of layer chickens killed by manual cervical dislocation.
Page 178
165
For humane reasons, we decided to assess the KED device in awake chickens only after
assessing it on anesthetized birds. However, based on the results of the trials using anesthetized
chickens (Chapter 4), we decided not to apply the KED device in awake chickens due to the
prolonged time for brain death, prevalence of sub-luxations versus full luxations and resultant
cervical fractures.
6.1.2. Pathology caused by euthanasia methods and effectiveness
The degree of tissue trauma caused by different euthanasia methods can be used to corroborate
the antemortem measures of loss of sensibility and brain death. The preferred killing methods are
ones that cause direct trauma to the brain disturbing normal electrical activity (Alexander, 1995;
Claassen et al., 2002) and result in rapid loss of sensibility and brain death. NPCB are
manufactured to cause direct damage to the brain and to result in loss of sensibility. However,
cervical dislocation methods aim to transect the spinal cord and destroy the blood vessels which
supply blood into the brain, causing death by cerebral ischemia (White and Krause, 1993;
Claassen et al., 2002). It has been suggested that the stretching and twisting motion in manual
cervical dislocation results in a concussive effect and brain stem damage (Shi and Whitebone,
2006; Cartner et al., 2007).
Skin laceration and external bleeding, skull fractures, and macroscopic and microscopic
hemorrhages and trauma of the brain and spinal cord were assessed for chickens killed with
NPCB. For birds killed by manual and mechanical cervical dislocation by KED, skin laceration
Page 179
166
and external bleeding, macroscopic and microscopic hemorrhage were also assessed, in addition
to dislocation site and fracture of the cervical vertebrae, and transection of the spinal cord.
Brain hemorrhage and skull fractures are frequently used to score the damage caused by physical
methods of euthanasia (Erasmus et al., 2010c; Casey-Trott et al., 2013; Bader et al., 2014;
Woolcott et al., 2018a,b). Researchers have suggested that immediate insensibility and
irreversible loss of vital functions are associated with evidence of subdural and parenchymal
hemorrhage in poultry (Erasmus et al., 2010c; Bader et al., 2014). Chapter 3 revealed that all
three NPCB devices caused substantial subdural and parenchymal hemorrhage in the brain
tissues. All birds sampled for microscopic scoring showed both parenchymal and subdural
hemorrhage in the mid brain for all NPCB devices except one bird for Zephyr-E. Subdural
hemorrhage (SDH) was substantial in the hind brain for all three NPCB devices with the vast
majority given the highest scores (moderate to marked levels). Nearly 50% of birds had
parenchymal hemorrhage (PCH) in the hind brain for all three NPCB devices. In contrast,
Chapter 4 revealed that brain damage was minimal for mechanical cervical dislocation method.
None of the birds killed by KED had PCH in the brain whereas only 1 of 18 birds sampled for
microscopic evaluation had minimal level of SDH in the hind brain. Brain damage was also
comparatively less in birds killed by manual cervical dislocation in comparison to NPCB. In the
birds killed by manual CD, minimal levels (<1%) of PCH were observed only in the hind brain
in 14% birds sampled whereas 76% of birds had SDH in the hind brain. A few birds (30%)
killed by manual CD had SDH also in the mid brain. Thus, in contrast to mechanical CD by
KED, stretching and twisting the neck during manual CD caused brain stem damage as
suggested in previous studies (Shi and Whitebone, 2006; Cartner et al., 2007). The neuronal
Page 180
167
network of the reticular formation which is responsible for consciousness (Seth et al., 2005;
Erasmus et al, 2010a) is located in the brain stem, and its ascending pathway innervates the fore
brain (Starzl et al., 1951) to process information of consciousness. Thus, trauma to the brain stem
region was suggested to disconnect the neuronal pathway that is responsible for sensibility
(Bader et al., 2014). In the current study, brain stem trauma was minimal in the birds killed with
KED, and less than in the birds killed with manual CD. Brain stem damage was higher in the
birds killed by NPCB in comparison to cervical dislocation methods, suggesting that this could
be the reason for rapid loss of sensibility in the birds killed with NPCB. The longest time to brain
death was observed in the birds killed with KED, corresponding to the minimal brain stem
trauma.
Gregory and Wotton (1990a) reported that cervical dislocation by pliers did not result in
immediate loss of sensibility in broiler chickens based on visual evoked responses, and the spinal
cord remained intact in 21% of birds. Carotid arteries also remained intact but left and right
arteries were damaged. The authors concluded that the birds killed by the pliers died from
asphyxia. In Chapter 4, subcutaneous hemorrhage in the neck was observed in the birds killed by
KED but brain trauma was absent in all except one bird. Thus, these findings support the
suggestions that death results from cerebral ischemia in the birds killed by mechanical cervical
dislocation and not because of the brain trauma. Overall, the minimal brain trauma could be the
reason I observed longer latencies to insensibility and clinical death in chickens killed with
mechanical cervical dislocation than the chickens killed with manual CD. The highest scores (3
and 4) for subcutaneous hemorrhage in the neck was found in more than 70% of the birds killed
by manual cervical dislocation. Therefore, I suggest that loss of sensibility and death results from
Page 181
168
both cerebral ischemia and brain trauma in the birds killed by manual CD. The killing methods
which do not stretch and twist the neck, resulting in cerebral ischemia alone (e.g. burdizzo or
pliers), have been prohibited since 2013 under the Council Regulation 1099/2009 (European
Commission, 2009).
Anatomical damage to the top of the spinal cord and the base of brain stem is associated with
spinal cord concussion, neurogenic shock, and loss of sensibility (Dumont et al., 2001; Harrop et
al., 2001). In all of the birds killed by manual cervical dislocation, the spinal cord was transected
at the top (between the skull and C1 or C1-C2). For the mechanical cervical dislocation, 75% of
birds had a transected spinal cord but further down the vertebrae than the recommended skull to
C1. I suggest that lower level transected spinal cord or dislocation could be the reason for
minimal brain stem trauma in the birds killed by KED. Martin et al. (2018) reported that high
neck dislocation was associated with improved kill success and more rapid loss of reflexes
compared to lower level dislocations for cervical dislocation. Overall 58% of the birds (awake
and anesthetized) killed by manual cervical dislocation had luxation in between the skull and C1.
In contrast, birds killed with mechanical CD presented dislocation with the majority (71%) of
sites occurring caudal to C2.
Subdural and parenchymal hemorrhage in the spinal cord were observed, indicating traumatic
damage to the spinal cord in birds after both manual and mechanical cervical dislocation. Some
muscles that are responsible for inspiration are innervated by cervical nerves and the accessory
cranial nerve (CN X1) (DeTroyer and Estenne, 1984). Some authors reported that damage to the
cervical area of the spinal cord impaired the functioning of these respiratory muscles resulting in
Page 182
169
hypoxia due to respiratory failure, and death in humans (Winslow and Rozovsky, 1983). The
observed traumatic damage to the spinal cord resulted in hypoxia causing death in the birds
killed by cervical dislocation. I suggest that hypoxia could be a reason for the observed open
mouth breathing in birds killed via cervical dislocation. This is also supported by the longer
duration of open mouth breathing in the birds killed by KED. Therefore, trauma to the spinal
cord could be the main reason for death in the birds killed by KED as they had minimal brain
trauma.
Erasmus et al. (2010c) suggested that the presence or absence of skull fractures did not affect the
onset of insensibility in turkeys killed by NPCB or blunt trauma. Chapter 3 revealed that some
birds in the 60-70 week old group did not have any skull fractures (Zephyr-E=4%, Zephyr-
EXL=29%, TED= 20%). However, all these birds were successfully killed and had a rapid loss
of sensibility, similar to those with skull fractures. Therefore, my results agree with Erasmus et
al. (2010c) that there is no association with degree of skull damage and time to insensibility or
brain death.
6.1.3. Limitations of reflex and behaviour measures
In this thesis, euthanasia methods were assessed based on how quickly insensibility and
subsequent brain death was induced (AVMA, 2013; NFACC, 2016). However, determining the
exact onset of brain death is highly debated in both human and veterinary medicine. Verhoeven
et al., 2015 stated that insensibility and brain death are best measured through
electroencephalogram (EEG), and it is the most objective measure. But EEG is not practical for
on-farm assessment of brain death. Animal care guidelines require that clinical death be
Page 183
170
confirmed before leaving birds and disposing of carcasses (AVMA, 2013; NFACC, 2016). Some
authors reported birds regaining sensibility or brain stem reflexes after application of a NPCB or
cervical dislocation (turkeys killed with NPCB- Erasmus et al., 2010b; broiler chickens killed by
KED and manual cervical dislocation- Jacobs et al., 2019). Thus, loss of brain stem reflexes is
not enough to confirm clinical death. If a bird regains sensibility, it may suffer until its clinical
death. Stockpeople need to monitor birds until clinical death to minimize the birds’ suffering; if
there is a return to sensibility, they need to apply a second killing method immediately.
Therefore, there is the need for behavioural and physiological reflexes, measures other than
EEG, to be identified as reliable indicators of clinical death, to be applied on birds prior to
carcass disposal.
Brainstem reflexes and behavioural measures have been validated as conservative measures of
sensibility and brain death (Gregory and Wotton, 1990a; Sandercock et al., 2014). I assessed the
relationships between time to loss of brain stem reflexes, some behavioural observations and
time of cessation of heart beat following application of the NPCB. My study suggests that
cardiac arrest was positively correlated with onset of tonic convulsions, time to last movement,
and cloacal relaxation for all three NPCB devices and thus, onset of tonic convulsions, last
movement, and cloacal relaxation can be used by stockpeople to make accurate decisions of
successful euthanasia under field conditions. Cessation of movement has been suggested to
estimate irreversible brain death (Dawson et al., 2007; Erasmus et al., 2010a). Raj et al. (1990)
studied the effect of CO2 stunning of hens on electroencephalogram (EEG) suppression and
found that cessation of convulsions happened (at 93 s) just before brain death (at 101s).
Moreover, Raj et al. (1992) reported brain death in hens exposed to CO2 and Ar mixture at 58 s
Page 184
171
based on EEG, and cessation of visible movements at 60s. My results agree with these findings,
that the last visible movement occurred immediately after the brain death. In more a recent study,
Jacobs et al. (2019) assessed the time at cessation of musculoskeletal movements (vent, legs, tail)
to interpret onset of brain death in broiler chickens. They reported that cessation of
musculoskeletal movements were the last visible measures to confirm brain death.
During this project, reflexes were assessed every 10 s, which is more frequent than previous
studies that used 15 s intervals to measure the reflexes (Martin et al., 2016, Woolcott et al, 2018
a,b). A 15s interval may have reduced the accuracy in determining when reflexes were lost, and
may have led to their overestimation. The 10 s interval used in this study was chosen to give
better accuracy, and still allow for multiple measures to be made sequentially within each
interval, providing a large amount of information.
In Chapter 4, we used anesthetics (ketamine and medetomidine) to reduce distress or pain
associated with the killing technique by abolishing the awareness of the animal. However, there
was a significant effect of the anesthetics on several brain stem reflexes and behaviour measures.
The anesthetic protocol used allowed measurement of pupillary reflex as a conservative indicator
of insensibility (due to brain death) in all birds. However, the anesthetics abolished or reduced
nictitating membrane reflex, clonic convulsions, tonic convulsions, and cloacal relaxation,
precluding their use for comparing killing methods. Woolcott et al. (2018b) also reported that the
nictitating membrane reflex, clonic convulsions, and gasping were affected by the same
anesthetic protocol in turkeys, although jaw tone was unaffected. In contrast, Sandercock et al.
(2014) reported observing nictitating membrane reflex even after respiratory arrest and brain
Page 185
172
death in laying hens anesthetized with sevoflurane. The authors suggested that the long time to
cessation of nictitating membrane reflex may have been due to combined effects of the used
anesthetic and sedative. Combined, these studies suggest that, nictitating membrane reflex,
clonic convulsions, tonic convulsions, and cloacal relaxation are more sensitive to the injectable
anesthetics and may not be useful as an approximate measure of brain death in cases where
ketamine and medetomidine have been administered in chickens.
In the current study the anesthetic protocol resulted in faster brain death in chickens. Thus, the
observed time for brain death in chickens killed by KED (while under anesthesia) is
conservative. Since anesthesia resulted in a faster time to brain death in birds killed via manual
CD, the time to death for birds killed via KED would likely be longer in awake chickens.
Sandercock et al. (2014) suggested that semi-awake (sedated) chickens demonstrated jaw tone
and neck muscle tone but these two measures were absent when birds were under general
anesthesia. Most of the birds in the current study were under general anesthesia and did not have
jaw tone. Therefore, we did not assess jaw tone as a measure of insensibility in anesthetized
birds. However, we found that individual variation occurred in depth of anesthesia in the
chickens for the same anesthetic protocol.
6.1.4. Device success and operator safety
All three NPCB devices used in this study caused rapid, irreversible insensibility and subsequent
death in layer chickens. Kill success was 100% for the TED device whereas only one failure was
observed for Zephyr-EXL device (one bird out of 93 birds). However, a few device failures
occurred for Zephyr -E-standard (4 birds out of 54 did not lose sensibility within 60 s). At the
Page 186
173
beginning of the experiment, birds were killed by using Zephyr-E-standard with a common
subject adapter and the conical shape bolt head. This common subject adapter is mainly designed
for swine and rabbits. Three birds killed by Zephyr-E-standard failed to lose brain stem reflexes
within 60s. Therefore, we switched to using the Zephyr-E-layer with a chicken subject adapter
and the round shape bolt head. The Zephyr-E-layer differs from the standard Zephyr-E in that
round shape bolt head and the chicken subject adapter. The chicken subject adapter allows for a
better alignment of the device around the comb of the bird. Selection of the correct subject
adapter for the species is an important factor for the killing success of Zephyr devices. However,
another three birds out of 47 killed by the Zephyr-E-layer also failed to lose brain stem reflexes
within 60s. The bolt velocity of Zephyr devices is adjustable as a function of air pressure.
Therefore, any pressure drop will result a lower bolt velocity and generate lower kinetic energy
from the device. The pressure in the air compressor was decreased below the recommended level
of 120 psi when killing these 7 birds by using Zephyr-E (4 birds by Zephyr-E standard + 3 birds
with Zephyr-E- layer). Therefore, I suspect this could be a cause for the observed failures. The
bolt velocity of the Zephyr-E is 20 m/s and generates 11 J of kinetic energy. The bolt velocity of
Zephyr-EXL is 27 m/s and generates 26 J kinetic energy. Therefore, the energy generated by
Zephyr-E is comparatively low. For the Zephyr-E, any air pressure level below 120 psi is not
enough to generate sufficient force to successfully kill a chicken. Therefore, Zephyr-E should be
used with the manufacturer-recommended exact air pressure level of 120 psi.
Another problem was encountered with Zephyr devices for one strain of chickens, the Plymouth
Barred Rock. After device application, the feathers of the Plymouth Barred Rock became lodged
between the bolt and the adapter for both Zephyr-E and Zephyr-EXL. Despite this, the birds
Page 187
174
were killed effectively with these devices. The Plymouth Barred Rock, a heritage strain of
chicken, differs from the other tested strains in feathering with easily removable feathers.
Seventeen of the birds used in this study were Plymouth Barred Rock, and two of them resulted
in this problem, when killed using the chicken subject adapter. Therefore, extra care needs to be
taken when using Zephyr devices with chicken adapters on chickens in different plumage types.
When evaluating killing methods for practical on-farm use, the methods need to be safe for
stockpeople. Sufficient training and skill are required for the method to be effective. Cervical
dislocation is widely used as an on-farm euthanasia technique by poultry producers because it is
easy to do, and practical and can be applied immediately during barn walk-throughs. People can
be easily trained on how to do cervical dislocation and there is minimal risk to the safety of the
operator. Operating the KED device is also easy, with minimal risk associated for the operator.
Comparatively, the potential for injury may be higher with operating NPCB devices. However,
all three devices are designed with a safety pin to minimize risk, and the safety pin needs to be
removed for the devices to function. The Zephyr devices additionally have a secondary safety
switch as an extra precaution, and the TED requires depression of the activator for the device to
function. Every individual using these devices should follow the steps of manufacturer’s operator
safety guide and should receive proper training in order to ensure the device is used in an
appropriate manner.
Additionally, proper restraint of the bird during device application is important, both for operator
safety and for the welfare of the bird. We did not use restraining devices in the current study
Page 188
175
because we needed to observe the reflexes and behavioural measures. Two people were involved
during the killing process: one person applied the device while the other one restrained the bird.
However, one person can kill a bird with NPCB devices or KED using a restraint device without
having assistance from a second person. A cone-shaped fabric (made of nylon) sleeve with an
opening at the top and bottom ends was designed for the purpose of our study, but also to
facilitate producers in restraint during euthanasia (Figure 6.1). This sleeve restrains the wings
towards the body of the bird during convulsions, and one person can easily apply the restraining
device on a bird.
Overall, more that 80% of birds showed external damage after application of the three NPCB
devices. Some birds had bleeding from the mouth and nose, and damaged eyes. Minimizing
external bleeding is important for biosecurity measures and aesthetic concerns. Serpa et al.
(2017) evaluated the efficacy of the Zephyr-EXL, powered at different air pressures levels (80,
90, 100, 110 and 120 psi) in killing broiler breeder roosters. They found that pressures between
80-120 psi are were equally effective at humanely killing broiler breeder roosters, and pressures
from 80-110 psi reduced the external damage compared to 120 psi.
6.2. Assessment of aversion to CO2 in laying hens
In Chapter 5, I presented an assessment of the degree of aversion in laying hens exposed to 4
different CO2 concentrations (25%, 35%, 50%, 70%) based on approach avoidance and
conditioned place avoidance paradigms with addition of a distress behaviour assessment. Results
revealed that approach avoidance and conditioned place avoidance paradigms were successful in
Page 189
176
assessing aversion to CO2 in layer chickens. Some hens avoided entering into some CO2
concentrations. Some hens displayed conditioned place avoidance to some CO2 concentrations.
I hypothesized that birds would avoid entering the treatment chamber when CO2 concentrations
were aversive. To support this hypothesis, I observed some proximate effects by comparing the
behaviours on Baseline Day versus Gas Day. Fewer hens entered the treatment chamber (TC) on
Gas Days than on Baseline Days. Avoidance of entering the TC on Gas Day revealed that the
motivation of a bird to avoid the aversive environment inside the TC was higher than the
motivation for acquiring the rewards inside the TC. More birds showed a higher number of
curtain pushes and head insertions into TC before entering or during avoidance on Gas Days than
Baseline Days. The number of curtain pushes and head withdrawals before entering / avoiding
TC on Gas day reflect conflict or competing motivation between entering the TC to acquire the
rewards and avoidance of gas. Overall, more hens demonstrated higher rates of vocalizations
inside the control chamber (CC) on Gas Day than Baseline Day, showing evidence of either
fearfulness or conflict to enter the TC.
All hens in the current study that entered the TC on Gas Day showed head shaking and open
mouth breathing irrespective of CO2 concentration. Head shaking and open mouth breathing,
previously observed in poultry for varied CO2 concentrations, have been suggested to be
indicators of aversion (Webster and Fletcher, 2001) and respiratory distress (turkeys: Raj, 1996;
broilers: Gerritzen et al., 2000; laying hens: Webster and Fletcher, 2001). Interpretation of head
shaking in this regard is difficult. Some researchers suggested head shaking served as an alerting
response to unexpected or novel events in poultry (Hughes, 1983) or an attempt to regain an alert
Page 190
177
state when birds are feeling dizzy or start to lose sensibility during the exposure to CO2
(Gerritzen et al., 2007). In the current study, some hens demonstrated head shaking on Baseline
Days and Washout Days when the CO2 concentration was the same as ambient air. Therefore,
the arguments that head shaking is an aversive reaction to CO2 or that it indicates respiratory
distress is not fully supported. McKeegan et al. (2005) suggested that the acidity of CO2 may
provide a gustatory stimulus which causes head shaking. Significantly more hens showed head
shaking on Gas Days than Baseline Days for all CO2 concentrations. Therefore, hens are more
likely to perform head shaking when exposed to CO2, but it is not necessarily indicative of
respiratory distress.
In the current study, all except one bird voluntarily entered to 25% and 35% CO2. Overall, the
number of hens entering into the TC decreased with increasing CO2 concentrations. Webster and
Fletcher (2004) suggested that hens could detect the presence of CO2 in the chute leading to a
chamber filled with 30%, 45%, and 60%. In the current study, the purpose-built chamber allowed
for greater control over the concentration of CO2 both within and outside the test chamber, so
that birds were exposed to the appropriate concentrations. The TC was monitored in real-time
with a wide span CO2 sensor and a real-time O2 sensor connected to an automatic data recording
system. The CC was monitored with a wide span CO2 sensor and an ambient range CO2 sensor.
Further, a negative pressure exhaust sink was located between the two chambers to evacuate
excess CO2 and maintain separate TC and CC environments. Some hens who did not enter into
the TC demonstrated a number of curtain pushes and head insertions indicating several attempts
to enter TC for 50% and 70% CO2 concentrations. Hens were exposed to the CO2 concentration
inside the TC when they performed head inserts and curtain pushes, and were able to detect the
Page 191
178
particular concentration inside the TC. Nearly half of the birds tested for 50% CO2 avoided
entering the TC on Gas Day suggesting that half of the hens found 50% CO2 as an aversive
stimulus. Only two hens entered into 70% CO2, and all the other hens tested avoided 70% CO2.
Therefore, the vast majority of hens were able to detect 70% CO2 and avoided entering into it.
McKeegan et al. (2003) observed an aversion threshold (based on an electrophysiological
assessment) to CO2 of 24% in air for chickens and further reported that chickens can detect 11%
CO2 based on a behaviour study. The thresholds of hen nasal and buccal trigeminal nociceptors
to CO2 have been investigated (McKeegan, 2004) and the authors suggest thresholds of 40–50%.
Therefore, in principle, hens should be able to detect the 25% and 35% CO2 concentrations, and
should experience pain and/or discomfort when inhaling 40% or above CO2 concentrations. The
hens who that did not enter into 50% CO2 and 70% CO2 showed a number of curtain pushes and
head withdrawals indicating that hens found these CO2 concentration aversive, and quite
possibly painful. Except for two, all other hens who voluntarily entered into either 25% or 35%
CO2 did not demonstrate head withdrawals prior to entering TC suggesting that hens entered
25% and 35% CO2 with minimal or no aversion, or that their motivation to access the rewards
inside the TC was higher than the motivation to avoid 25% and 35% CO2.
Loss of posture (LOP) has been proven to be the reliable indicator of onset of insensibility in
poultry based on electroencephalography (EEG) studies in broiler chickens (Coenen et al., 2000;
Gerritzen et al., 2004; Benson et al., 2012). In the current study, onset of ataxia occurred
regardless of CO2 concentration. Latency to LOP was significant among the CO2 concentrations.
Overall latency to LOP decreased with increasing CO2 concentration. Similar results were
Page 192
179
observed by Webster and Fletcher (2001) who reported that higher CO2 concentrations reduced
the time to loss of sensibility based on time to LOP in layer hens.
6.2.1 Evaluation of the method
Hens who were in the control chamber were given a free choice to enter or avoid the Treatment
Chamber (TC) filled with a CO2 concentration. Prior to the testing phase hens were trained to
enter TC pushing through a curtain. Overall, the vast majority of laying hens were able to learn
the task by the fifth day of training. This indicates that the assigned task was within the cognitive
capacity of laying hens.
In this experiment 12 birds were used for the study. Our plan was to use the same birds to check
the aversiveness for four gas concentrations. We randomly assigned the birds for different CO2
concentrations. But some hens never entered the TC after their first, second or third gas
treatments. Because of this, the number of birds tested for each concentration was different. Most
of the hens avoided entering TC after their third gas experience. It seems that hens were able to
learn the adverse effect inside the TC, with increasing exposure to any concentration. By
rotating the same hens through different CO2 concentrations, there is a possibility that experience
of a previous CO2 concentration could carry over to influence the results of a test with another
CO2 concentration. Birds that have experienced exposure to CO2 may not be representative of
naïve birds. When the same bird was exposed to the next gas level, their sensitivity or tolerance
to a gas concentration may have changed. Two hens who were exposed to 50% and 70% CO2 in
their third round died without regaining of consciousness. Previously they had been exposed to
Page 193
180
two other CO2 concentrations in their first and second rounds. The deaths could have been due to
lack of physiological recovery or anatomical damage due to previous gas exposures.
In this experiment I assumed that the strength of motivation to get food (commercial food and
meal worms) and motivation to avoid CO2 were additive. But this assumption does not always
hold true. In the current trial, I assigned the order of birds based on their motivation to enter the
treatment chamber during the training period. The most motivated birds were tested first. Food
was withheld 1 hour prior to testing the first bird. All the birds were tested within 5 hours of feed
deprivation. Kirkden et al. (2008) found that tolerance to the aversion of CO2 in rats was
increased with the deprivation time and then started to decrease after a certain point. They found
the least level of aversiveness at the intermediate feed deprivation. I provided both commercial
feed and meal worms in the treatment chamber, with meal worms only provided in the treatment
chamber except during the training. I observed that hens had a higher motivation to enter the TC
to eat meal worms. I also assumed that providing other enrichments for foraging helped to
increase their motivation to enter TC.
During the testing phase, we allowed 5 minutes for acclimatization in the control chamber prior
to opening the sliding door which was in between the two compartments. We allowed another
five minutes for the bird to enter the treatment chamber after the door was opened. The vast
majority of the birds entered TC within 3 minutes from opening the sliding door in their training
sessions. Therefore, I conclude that the allocated 5 minutes time was sufficient for the hens to
make the decision on whether to enter the TC or not.
Page 194
181
Time at egg laying was not considered in the current trial. However, most hens laid before 1.00
p.m., and the test began at 1.30 p.m. every day. We assumed all hens completed egg laying by
that time, but we did not manage hens individually for their egg laying. This confounding effect
could have interfered with the results of distress behaviour or avoidance. Every day we
performed a health assessment for all the birds before the test. Therefore, it can be confirmed that
health issues did not interfere with the results of the current study.
6.2.2 Implications
This thesis assessed available on-farm euthanasia methods for laying hens. However, in Chapter
5, aversion to different CO2 concentrations was assessed with their possibility to induce rapid
insensibility in layer chicken. Thus, it is important to discuss how does the results of Chapter 5
apply to on-farm euthanasia context.
Carbon dioxide is used in two different techniques to euthanise chickens. One method is gradual
filling the euthanasia box with CO2 and the other one is immersed birds in pre-filled CO2
chambers (Close et al., 1996). Gradual filling of 100% CO2 is accepted to achieve minimum pain
and distress (AVMA, 2013). Geritzen et al. (2004) suggested that anesthesia is induced in a bird
before CO2 level reach painful level. Immersion in CO2 is acceptable at low CO2 concentrations
to avoid associate distress (AVMA, 2013). Gentle death in adult bird with minimal pain that take
longer is preferred than quick death with distress (AVMA, 2013).
I studied 25% and 35 % CO2 concentrations, and found that these levels were relatively less
aversive to the hens. Onset of insensibility occurred less than 20 s at 25% and 35% CO2
Page 195
182
concentrations. Thus, suggested that there are welfare advantages to the hens if use 20% and
30% CO2 concentrations as prefilled technique in euthanasia. However, practical issues arise in
on-farm context to use lower level of CO2. Many farms used gradual filling technique and it is
not sure whether a lower CO2 concentration is possible to hold for a particular time period inside
the CO2 chamber. The commercially available euthanasia boxes are mainly designed to gradual
increase up to 60% CO2 or above levels.
Gerriteze et al. (2013) reported that multistage gas stunning has distinct advantages for bird
welfare. Authors started with a lower concentration (30% or 40% CO2) and increased to 65%
CO2. Birds were exposed to 45 or 60 s at the particular CO2 concentration. Authors concluded
that CO2 concentration did not exceed 40% before all the birds expressed loss of posture
indication of unconsciousness. Moreover, it was reliable in both the experimental and the
semicommercial setup. I did not find difference in latency to onset of LOP at 35% and 50% CO2.
Therefore, it is reasonable to start with 30% CO2 concentration and hold the level until bird loss
its sensibility, and then increase to a higher concentration to achieve a rapid death in layer
chicken euthanasia to achieve more welfare benefits.
However, these lower CO2 concentrations cannot recommend for chick euthanasia. The atrial
pCO2 level is comparatively higher (60 mm Hg) in the embryo (Freeman and Vince, 1974).
Thus, day-old chicks are more resistant to hypercapnia and anoxia (Jaksch, 1981), Therefore,
higher CO2 concentrations need to be targeted in day old chick euthanasia and the levels
recommended for adult chickens are not sufficient to achieve rapid insensibility or death (Jaksch,
1981). Bethany et al., (2019) concluded that shortest time of distress and fastest time of death
Page 196
183
occurred in old chicks immersed in 100% CO2 concentration in comparison to gradual filling
technique. Therefore, results of the current study can only apply in on-farm context by targeting
the welfare advantages of adult birds and not the chicks.
6.3 In retrospect
The results of the current study demonstrate that non-penetrating captive bolt (NPCB) devices
caused rapid, irreversible insensibility and subsequent death in layer chickens. However, external
skin damage and bleeding was higher for NPCB devices in comparison to cervical dislocation
methods. Some birds killed by NPCB devices had mouth and nose bleeding, and damaged eyes.
External bleeding is important when considering biosecurity and aesthetic concerns. I would
recommend doing a pilot study to identify the degree of external damage caused by different
NPCB devices using different pressures. In that way, it will be possible to come up with a more
suitable scoring system to evaluate the external damage. In the current study we did not score the
degree of external bleeding. Nose bleeding and mouth bleeding should also be included in the
external damage and bleeding scoring system. Future studies to evaluate suitable air pressure
levels for Zephyr-EXL, which cause minimal external damage and rapid insensibility leading to
death should be based on a comprehensive external damage scoring system.
I would have liked to use pentobarbital sodium, which is considered the gold standard in
euthanasia to induce rapid insensibility and brain death (Hernandez, 2018) as the secondary
killing method instead of having to apply a second shot with the TED in the failed birds. This
would preserve brain architecture better for macroscopic and microscopic assessment. In that
Page 197
184
way, I would have been able to get a better idea about the degree (or lack) of pathology of the
brains of the birds that failed to be killed with the tested method.
Chapter 5 revealed that layer hens showed learned behaviours going through the treatment
rounds during testing for aversion to CO2 concentrations. Some birds might not have fully
recovered physically when they were tested in the next immediate round. Therefore, I would
have preferred to have 12 naïve birds for each concentration rather than rotating the same 12
hens through different concentrations. However, that would increase the number of animals
necessary for the trial, and this may not fit with the 3Rs concept. This would also add
considerable time and expense for training the birds. An alternative would be to permit
additional days of rest to the hens between testing rounds. The other issue of repeating the hens
through the rounds was that a hen may learn that she will be returned to her home pen if she does
not enter TC. She also learnt that the TC is an unpredictable environment. This could be the
reason that a greater number of hens avoided entering the TC in the 3rd round. However, I did not
provide food in the home pen until the last hen completed the test in order to decrease the
motivation of a hen to return to the home pen Meal worms were never provided inside the home
pen after the habituation period, and a hen’s only chance to get meal worms was by entering the
TC.
6.4 Overall conclusion
The death of an animal at human hands, is an important element of an animal’s welfare.
Therefore, to achieve a good death in an animal, all possible measures need to be taken. People
argue that euthanasia means ending of one’s life to eliminate pain and distress. However, many
Page 198
185
of the available killing methods cause pain and distress. This thesis addresses how painful the
available killing methods and their degree of suitability as a humane method in layer chickens,
thus giving a chance to think and argue on whether we can achieve a good death without a
humane killing method. It is our responsibility to ensure that the existing killing methods truly
result in a good death to animals without inducing unnecessary pain or distress.
Available euthanasia guidelines and codes of practice listed several conditions which should be
achieved during killing an animal in order to offer a humane death. These euthanasia protocols
are based on previous research studies conducted on few species. A killing method assessed for
one species or one age group cannot be generalized to the other species or other age groups.
Therefore, available killing methods need to be assessed on target species and target age groups.
Euthanasia guidelines should focus on species and age categories to achieve accurate
recommendations.
The series of experiments conducted in this thesis have identified the potential of new physical
killing devices (NPCB and KED) in layer chicken industry. Assessments were done in different
age groups facilitating more accurate recommendations. Three different types of commercially
available NPCB devices were assessed in layer chickens assuming that device differences could
cause a different degree of welfare advantages in birds. This facilitates the most accurate
recommendation of devices to use on layer chickens rather than recommending all NPCB
devices in general. KED-model-C was assessed in different age groups as there were no
recommended minimum body weight for application of the device. Manufacturers should clearly
indicate age or weight limits (both minimum and maximum) for different poultry species.
Page 199
186
Different poultry species have anatomical differences, thus using only the weight category may
not be enough to select the correct model of the device.
Existing euthanasia protocols or guidelines have not recommended specific CO2 concentrations
for layer hens to achieve welfare advantages. In Chapter 5, I identified relatively less aversive
CO2 concentration (25% and 35%) which resulted in insensibility less than 20s in layer chickens.
Results of approach avoidance tests are always relative and depend on the cost and rewards to
the animal. The design of the current study provided an advantage in the terms of welfare as
birds were not highly motivated to enter the TC. I provided luxury treat (meal worms) and some
environmental enrichments which resulted in a positive motivation. Since the bird does not need
them they can easily avoid entering TC. On the other hand, I withheld the feed, and this is a
negative motivation. However, the feed withdrawal period was short and may not resulted in
strong negative motivation in birds. Overall, the cost was not great for the bird and suggest of
that 25% and 35% levels were not that aversive.
This thesis provides important information for euthanasia guidelines to be considered in the
future with special attention to laying hen industry. Comprehensive trauma analysis of each
physical killing method gives information for manufacturers to consider in further improvement
of the existing devices to be more efficient as euthanasia methods. Every method had limitations,
thus selecting of the most appropriate method for the existing situation is important in an on-
farm context.
Page 200
187
Figure 6. 1 : The fabric chicken restraint device (fabric sleeve made of nylon): A- a toggle at the
head end to tighten it, B- Opening at the bottom end
Images curtesy of Institute for Applied Poultry Technologies - euthanasia techniques
http://www.iaptwest.org/welfare-and-pain-management/
Page 201
188
BIBLIOGRAPHY
Abdalla M. A. and King A. S. (1975). The functional anatomy of the pulmonary circulation of
the domestic fowl. Respiration Physiology 23(3): 267–290.doi: 10.1016/0034-
5687(75)90078-X.
Adams D. B. and Sheridan A. D. (2008). Specifying the Risks to Animal Welfare Associated
with Livestock Slaughter without Induced Insensibility. Australian Government
Department of Agriculture, Fisheries and Forestry, (November), pp. 1–100. Available at:
http://www.daff.gov.au/animal-plant----eealth/
welfare/aaws/a_scientific_comment_on_the_welfare_of_sheep_slaughtered_without_stu
nning/specifying_the_risks_to_animal_welfare_associated_with_livestock_slaughter_wit
hout_induced_insensibility.
Alexander M.P. (1995). Mild traumatic brain injury: pathophysiological, natural history, and
clinical management. Neurology 45, 1253-1260.
Alkire M. T., Hudetz A. G. and Tononi G. (2008). Consciousness and Anesthesia. Science
322(5903): 876–880. doi:10.1126/science.1149213.
Altman R.B. (1997). Avian Medicine and Surgery. Clubb SL, Dorrestein GM, et al (eds)
Philadelphia, PA, Saunders
Ambrose N, Wadham J, Morton D (2000) In: Refinement of Euthanasia. Progress in the
Reduction, Refinement and Replacement of Animal Experimentation (Balls M, van
Zeller AM, Halder ME, eds). Amsterdam: Elsevier Science. pp.1159-1169.
American Veterinary Medical Association AVMA Guidelines for the Euthanasia of Animals:
2013 Edition (2013). Available online:
https://www.avma.org/KB/Policies/Documents/euthanasia.pdf (accessed on 28 March
2016).
Anastasov M. I. and Wotton S. B. (2012). Survey of the incidence of post-stun behavioural
reflexes in electrically stunned broilers in commercial conditions and the relationship of
their incidence with the applied water-bath electrical parameters. Animal Welfare, 21(2):
247–256. doi: 10.7120/09627286.21.2.247.
Anil M. H. (1991). Studies on the return of physical reflexes in pigs following electrical
stunning. Meat Science, 30(1): 13–21. doi: 10.1016/0309-1740(91)90030-T.
Animal Production Science, 54: 187–193.http://dx.doi.org/10.1071/AN12128
Anton F., Euchner I., and Handwerker H. O. (1992). Psychophysical examination of pain
induced by defined CO2 pulses applied to the nasal mucosa. Pain, 49: 53-60.
Page 202
189
Anton F., Peppel P., Euchner I., and Handwerker H.O. (1991). Controlled noxious chemical
stimulation: responses of rat trigeminal brainstem neurons to CO2 pulses applied to the
nasal mucosa. Neuroscience Letters, 123: 208-211.
Anton F., Euchne, I., Handwerker H.O., (1992). Psychophysical examination of the pain induced
by defined CO2 pulses applied to the nasal mucosa. Pain 49 (1):53–60.
Aramideh M. and Ongerboer De Visser B. W. (2002). Brainstem reflexes: Electrodiagnostic
techniques, physiology, normative data, and clinical applications. Muscle and Nerve,
26(1): 14–30. doi: 10.1002/mus.10120.
Atkinson S., Velarde A., and Algers B. (2013). Assessment of stun quality at commercial
slaughter in cattle shot with captive bolt. Animal Welfare, 22: 473–481
Bader S., Meyer-Kuhling B., Gunther R., Breithaupt A., Rautenschlein S. and Gruber A. D.
(2014). Anatomical and histologic pathology induced by cervical dislocation following
blunt head trauma for on-farm euthanasia of poultry. The Journal of Applied Poultry
Research, 23(3): 546–556. doi: 10.3382/japr.2014-00977.
Baggott J. (1982). Gas Transport and pH Regulation, in Textbook of Biochemistry with Clinical
Correlations (Devlin, T. M., Ed.) pp 10981123, John Wiley and Sons, New York
Baker B.I, Torrey S., Widowski T.M.,Turner P.V., Knezacek T.D., Nicholds J., Crowe T.G
Schwean-Lardner K. (2019). Evaluation of carbon dioxide induction methods for the
euthanasia of day-old cull broiler chicks. Poultry Science 98:2043–2053.
http://dx.doi.org/10.3382/ps/pey581
Banzett R. B, Pedersen S. H, Schwartzstein R. M., Lansing R.W. 92008). The affective
dimension of laboratory dyspnea: Air hunger is more unpleasant than work/effort.
American Journal of Respiratory and Critical Care Medicine 177, 1384–90
Basbaum A. I., Bautista D M, Scherrer G, and Julius D. (2009). Cellular and Molecular
Mechanisms of Pain. Cell, 139:267-284. DOI 10.1016/j.cell.2009.09.028
Beausoleil N.J., and Mellor D.J. (2015). Introducing breathlessness as a significant animal
welfare issue. New Zealand Veterinary Journal 63:44-51.
Benson, E.R.1., Alphin, R.L., Rankin, M.K., Caputo M.P., Kinney C.A. and Johnson A.L.
(2012). Evaluation of EEG based determination of unconsciousness vs. loss of posture in
broilers. Research in Veterinary Science. 93(2):960-964. doi: 10.1016/j.rvsc.2011.12.008
Blackman N., Cheetham K. and Blackmore D. (1986). Differences in blood supply to the
cerebral cortex between sheep and calves during slaughter. Research in Veterinary
Science, 40:252–254.
Blackmore D. K., Newhook J. C. and Grandin T. (1983). Time of onset of insensibility in four-
to six-week-old calves during slaughter. Meat Science. 9(2):145–149. doi: 10.1016/0309-
1740(83)90024-4.
Page 203
190
BMJ (1976). Diagnosis of brain death: statement issued by the honorary secretary of the
Conference of Medical Royal Colleges and their Faculties in the United Kingdom on 11
October 1976. BMJ 1976; 2: 1187-1188.29.
Bock Industries Inc. Captive Bolt Devices for Humane Livestock Euthanasia (2016). Available
online: http://www.bock-industries.com/
Bove S. E., Flatters S. J. L., Inglis J. J. and Mantyh P.W. (2009). New advances in
musculoskeletal pain. Brain Research Reviews. Elsevier B.V., 60(1): pp. 187–201. doi:
10.1016/j.brainresrev.2008.12.012.
Breazile J. E. and Hartwig H.G. (1989). Central nervous system. In: King AS, McLelland J (eds)
Form and function in birds. Academic, London, pp 485–566
Brown R. E., Basheer R., McKenna J. T., Strecker R. E. and McCarley R. W. (2012). ‘Control of
Sleep and Wakefulness’, Physiological Reviews, 92(3), pp. 1087–1187. doi:
10.1152/physrev.00032.2011.
Bryant B., Silver W.L. (2000). Chemesthesis: the common chemical sense. In: Finger TE, Silver
WL, Restrepo D, editors. The neurobiology of taste and smell. New York: Wiley-Liss. p
73–100.
Butler A. B. and Hodos W. (2005a). Comparative Vertebrate Neuroanatomy- Evolution and
Adaptation. John Wiley & Sons INC. publications
Butler, A. B., Manger, P. R., Lindahl, B. I. B. and Arhem, P. (2005b) ‘Evolution of the neural
basis of consciousness: A bird-mammal comparison’, BioEssays, 27(9), pp. 923–936.
doi: 10.1002/bies.20280.
Cabanac, M., (1979). Sensory pleasure. Quarterly Review of Biology 54,1–129.
Carstens E. and Moberg G. P. (2000). ‘Recognizing pain and distress in laboratory animals’,
ILAR Journal, 41(2), pp. 62–71. doi: 10.1093/ilar.41.2.62.
Casey-Trott T. M., Millman S. T., Turner P. V., Nykamp S. G. and Widowski T. M. (2013).
Effectiveness of a nonpenetrating captive bolt for euthanasia of piglets less than 3 d of
age. J Anim Sc., 91:5477-5484. doi: 10.2527/jas.2013-6320
Casey-Trott T. M., Millman S. T., Turner P. V., Nykamp S. G., Lawlis P. C. and Widowski T.
M. (2014). Effectiveness of a nonpenetrating captive bolt for euthanasia of 3 kg to 9 kg
pigs. J Anim Sc., 92:5166-5174. doi: 10.2527/jas.2014-7980
Chapman C. R. (2005). Pain perception and assessment. Minerva Anestesiologica, 71(7-8):413-
417
Chen, X., J. Gallar, M.A. Pozo, M. Baeza, and C. Belmonte (1995). CO2 stimulation
of the cornea: a comparison between human sensation and nerve activity in polymodal
nociceptive afferents of the cat. European Journal of Neuroscience 7: 1154-1163.
Page 204
191
Claassen, J., Carhuapoma, J.R., Kreiter, K.T., Du, E.Y., Connolly, E.S., Mayer, S.A. (2002).
Global cerebral edema after subarchnoid hemorrhage: frequency, predictors, and impact
on outcome. Stroke 33, 1225-1232
Clark L, Hagelin Julie, and Werner S J. (2014). "The Chemical Senses in Birds". USDA National
Wildlife Research Center -Staff Publications. 1618.
https://digitalcommons.unl.edu/icwdm_usdanwrc/1618
Clarke K. W. and Hall, L. W. (1990). A survey of anaesthesia in small animal practice:
AVA/BSAVA report’, Veterinary Anaesthesia and Analgesia, 17(1): 4–10. doi:
10.1111/j.1467-2995.1990.tb00380.x.
Close, B., Banister K., Baumans V., Bernoth E. M., Bromage N., Bunyan J., Erhardt W.,
Flecknell P., Gregory N., Hackbarth H., Morton D., Warwick C. (1997).
Recommendations for euthanasia of experimental animals: part 2. Lab. Anim. 31:1–32
Coenen, A., A. Smith, L. Zhonghua, and G. Vanluijtelaar (2000). Gas mixtures for anesthesia
and euthanasia in broiler chickens. World’s Poultry Science Journal 56: 225-234.
Coenen A. M. L., Lankhaar, J., Lowe, J. C. and McKeegan, D. E. F. (2009). Remote monitoring
of electroencephalogram, electrocardiogram, and behavior during controlled atmosphere
stunning in broilers: implications for welfare.’, Poultry science, 88(1), pp. 10–19. doi:
10.3382/ps.2008-00120.
Collins J. G., Kendig J. J. and Mason P. (1995). Anesthetic actions within the spinal cord:
contributions to the state of general anesthesia. Trends in Neurosciences, 18(12): 549–
553. doi: 10.1016/0166-2236(95)98377-B.
Cook C. J., Devine C. E., Gilbert K.V., Smith D. D. and Maasland S. A. (1995). The effect of
electrical head-only stun duration on electroencephalographic-measured seizure and brain
amino acid neurotransmitter release. Meat Sci. 40:137–147.
Cors J.C., Gruber A.D., Gunther R., Meyer-Kuhling B., Esser K.H., Rautenschlein S. (2015).
Electroencephalographic evaluation of the effectiveness of blunt trauma to induce loss of
consciousness for on-farm killing of chickens and turkeys. Poult. Sci. 94:147–155. doi:
10.3382/ps/peu038
Cruccu G. and Deuschl G. (2000).The clinical use of brainstem reflexes and hand-muscle
reflexes. Jounal of animal Sc. 91 (11):5477-87.doi: 10.2527/jas.2013-6320
Cullen L.K. (1996). Medetomidine sedation in dogs and cats: a review of its pharmacology,
antagonism and dose. British Veterinary Journal 152:519–535
Damasio A. and Meyer K. (2009). Consciousness: An Overview of the Phenomenon and of Its
Possible Neural Basis. S. Laureys & G. Tononi (Eds.) The Neurology of Consciousness:
3-14. Online access : https://www.sciencedirect.com/sdfe/pdf/download/eid/3-s2.0-
B9780123741684000010/first-page-pdf
Page 205
192
Danneman, P. J., Stein S, Walshaw S.O. (1997). Humane and practical implications of using
carbon dioxide mixed with oxygen for anesthesia or euthanasia of rats. Lab. Anim. Sci.
47:376–385
Dawkins M. S. (1980). Animal Suffering. Chapman and Hall: London, UK
Dawkins M. S. (1990). From an animal's point of view: Motivation, fitness, and animal welfare.
Behavioral and Brain Sciences 13(1):1-9
Dawson M. D., Lombardi M. E., Benson E. R., Alphin R. L. and Malone G. W. (2007). Using
accelerometers to determine the cessation of activity of broilers. Journal of Applied
Poultry Research, 16(4), pp. 583–591. doi: 10.3382/japr.2007-00023.
Derr R. F. (1991). Pain perception in decapitated rat brain. Life Sciences, 49(19): 1399–1402.
doi: 10.1016/0024-3205(91)90391-N.
Desmond J. E., Rosenfield M. E. and Moore J. W. (1983). An HRP study of the brainstem
afferents to the accessory abducens region and dorsolateral pons in rabbit: Implications
for the conditioned nictitating membrane response. Brain Research Bulletin, 10(6): 747–
763. doi: 10.1016/0361-9230(83)90208-3.
DeTroyer A., Estenne M., (1984). Coordination between rib cage muscles and diaphragm
during quiet tidal breathing in humans. J Appl Physiol, 57:899–906.
Drew L.B. and Drew W.E. (2004). The contrecoup-coup phenomenon. Neurocritical Care 1:
385-390. doi.org/10.1385/NCC:1:3:385
Dumont R.J., Okonkwo D.O., Verma S., Hurlbert R.J., Boulos P.T., Ellegala D.B., Dumont A.S.
(2001). Acute Spinal Cord Injury, Part I: Pathophysiologic Mechanisms. Clinical
Neuropharmacology 24: 254–264
Duncan I. J. H. (1996). Animal welfare defined in terms of feelings. Acta Agriculturae
Scandinavica Section A, Animal Science, Supplement 27: 29-35.
Duncker H. R. (1974). Structure of the avian respiratory tract. Respiration Physiology, 22(1–2):
1–19. doi: 10.1016/0034-5687(74)90044-9.
Eccleston C. (1995). Chronic pain and distraction: an experimental investigation in to the role of
sustained and shifting attention in the processing of chronic persistent pain. Behaviour
research therapy 33:391-405.
Edelman D. B., Baars B. J. and Seth A. K. (2005). Identifying hallmarks of consciousness in
non-mammalian species. Consciousness and Cognition, 14(1): 169–187. doi:
10.1016/j.concog.2004.09.001.
Erasmus M. A., Turner P. V. and Widowski T. M. (2010a). Measures of insensibility used to
determine effective stunning and killing of poultry. Journal of Applied Poultry Research,
19(3): 288–298. doi: 10.3382/japr.2009-00103.
Page 206
193
Erasmus M. A., Lawlis P., Duncan I. J. H. and Widowski T. M. (2010b). Using time to
insensibility and estimated time of death to evaluate a nonpenetrating captive bolt,
cervical dislocation, and blunt trauma for on-farm killing of turkeys. Poultry Science,
89(7): 1345–1354. doi: 10.3382/ps.2009-00445.
Erasmus M.A., Turner P.V., Nykamp S.G., Widowski T.M. (2010c). Brain and skull lesions
resulting from use of percussive bolt, cervical dislocation by stretching, cervical
dislocation by crushing and blunt trauma. Veterinary Records 167: 850–858.
European Council. European Council Regulation (EC) 1099/2009 of 24 September 2009 on the
Protection of Animals at the Time of Killing. Available online:
https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/325000/re
gulation_1099_2009_en.pdf
Evans K.C., Banzett R.B., Adams L., McKay L., Frackowiak R.S., Corfield D.R., (2002). BOLD
fMRI identified limbic, paralimbic, and cerebellar activation during air hunger. Journal of
Neurophysiology 88, 1500–11, 2002
Farmer C.G. (2006). On the origin of avian air sacs. Respiratory Physiology & Neurobiology
154: 89–106.
Feldman J. L., Mitchell G. S. and Nattie E. E. (2003). Breathing: Rhythmicity, Plasticity,
Chemosensitivity. Annual Review of Neuroscience, 26(1): 239–266. doi:
10.1146/annurev.neuro.26.041002.131103.
Feng Y., Simpson T.L. (2003). Nociceptive sensation and sensitivity evoked from human cornea
and conjunctiva stimulated by CO2. Invest. OPhth. Vis. Sci. 44., 529-532.
Feng, Y., and T. L. Simpson (2003). Nociceptive sensation and sensitivity evoked from
human cornea and conjunctiva stimulated by CO2. Investigative Ophthalmology &
Visual Science 44: 529-532.
Finnie J. W., Blumbergs P. C., Manavis J., Summersides G. and Davies R. (2000). Evaluation of
brain damage resulting from penetrating and non‐penetrating captive bolt stunning using
lambs. Australian Veterinary Journal 78: 775–778.
Finnie J.W., Manavis J., Summersides G.E., Blumbergs P.C. (2003). Brain damage in pigs
produced by impact with a non-penetrating captive bolt pistol. Aust. Vet. J. 81:153–155.
doi: 10.1111/j.1751-0813.2003.tb11078.x
Forslid A., Ingvar M., Rosen I., Ingvar D.H. (1986). Carbon dioxide narcosis: influence of short-
term high concentration carbon dioxide inhalation on EEG and cortical evoked responses
in the rat. Acta Physiolgica 127: 281-287.
Fraser D. and Duncan I. J. (1998). Pleasures, Pains and Animal Welfare: Toward a Natural
History of Affect. Animal Welfare, 7: 383–396.
Page 207
194
Fraser D., Weary D. M., Pajor E. A., and Milligan B. N. (1997). A scientific conception of
animal welfare that reflects ethical concerns. Animal Welfare, 6:187-205.
Freeman, B. M. and Vince M. A. (1974). Development of the Avian Embryo. Chapman and
Hall, London.
Freeman K. T., Koewler N. J., Jimenez-Andrade J. M., Buus R. J., Herrera M. B., Martin C. D.,
Ghilardi J. R., Kuskowski M. A., Mantyh P. W. (2008). A fracture pain model in the rat:
adaptation of a closed femur fracture model to study skeletal pain. Anesthesiology,
108(3): 473–483. doi: 10.1097/ALN.0b013e3181649351.
Gentle M.J. (2001). Attentional shifts alter pain perception in the chicken. Animal Welfare,
10:187-194
Gentle M.J. (2011). Pain issues in poultry. Applied Animal Behaviour Science 135:252– 258
doi:10.1016/j.applanim.2011.10.023
Gerritzen M. A., Lambooij E, Hillebrand S.J., Lankhaar, J.A. and Pieterse C. (2000). Behavioral
responses of broilers to different gaseous atmospheres. Poultry Science 79(6): 928-933.
http://www.ncbi.nlm.nih.gov/pubmed/10875779.
Gerritzen M. A, Lambooij B., Reimert H., Stegeman A., Spruijt B. (2004). On-farm euthanasia
of broiler chickens: effects of different gas mixtures on behavior and brain activity.
Poultry Science, 83(8): 1294–1301. doi: 10.1093/ps/83.8.1294.
Gerritzen, M., B. Lambooij, H. Reimert, A. Stegeman, and B. Spruijt (2007). A note on
behaviour of poultry exposed to increasing carbon dioxide concentrations. Applied
Animal Behaviour Science 108(1–2): 179–185. doi: 10.1016/j.applanim.2006.11.014.
Gerstenbrand F. and Stepan C. A (2001). Mild traumatic brain injury. Brain injury, 15(2): 95–97.
doi: 10.3171/2010.11.FOCUS.Intro.
Getty, R. (1975). Sisson and Grossman’s the Anatomy of the Domestic Animals.W.B. Saunders
Company, Philadelphia
Gibson T. J., Johnson C. B., Stafford K. J., Mitchinson S. L. and Mellor D. J. (2007). Validation
of the acute electroencephalographic responses of calves to noxious stimulus with scoop
dehorning. New Zealand veterinary journal, 55(4): 152–7. doi:
10.1080/00480169.2007.36760.
Gibson T. J., Johnson C. B., Murrell J. C., Hulls C. M., Mitchinson S. L., Stafford K. J.,
Johnstone A. C. and Mellor D. J. (2009).Electroencephalographic responses of halothane
- anaesthetised calves to slaughter by ventral-neck incision without prior stunning. New
Zealand veterinary journal, 57(2): 77–83. doi: 10.1080/00480169.2009.36882.
Gibson T. J., Ridler A. L., Lamb C. R., Williams A., Giles S., and Gregory, N. G. (2012).
Preliminary evaluation of the effectiveness of captive-bolt guns as a killing method
without exsanguination for horned and unhorned sheep. Animal Welfare, 21, 35–42
Page 208
195
Gibson T. J., Dadios N., Gregory N. G. (2015a). Effect of neck cut position on time to collapse
in halal slaughtered cattle without stunning. Meat Science. Elsevier B.V., 110: 310–314.
doi: 10.1016/j.meatsci.2015.03.026.
Gibson T. J., Mason C. W., Spence J. Y., Barker H., Gregory N. G. (2015b). Factors Affecting
Penetrating Captive Bolt Gun Performance. Journal of Applied Animal Welfare Science,
18 (3):222-238
Gibson T.J., Rebelo C.B., Gowers T.A., Chancellor N.M. (2018). Electroencephalographic
assessment of concussive non-penetrative captive bolt stunning of turkeys. British
Poultry Science 59 (1):13-20. DOI: 10.1080/00071668.2017.1401215
Gleed R. D., and Ludders J.W. (eds). (2001). Recent Advances in Veterinary Anesthesia and
Analgesia: Companion Animals. Ithaca, NY, International Veterinary Information
Service
Goldberg M.E. (2014). Pain Management for Veterinary Technicians and Nurses. John Wiley
and Sons
Grandin T. (2002). Return-to-sensibility problems after penetrating captive bolt stunning of
cattle in commercial beef slaughter plants. Journal of the American Veterinary Medical
Association, 221:1258–1261
Grandin T. (2013). Making Slaughterhouses More Humane for Cattle, Pigs, and Sheep. Annual
Review of Animal Biosciences, 1(1): 491–512. doi: 10.1146/annurev-animal-031412-
103713.
Grandin, T., S.E. Curtis, T.M. Widowski, and J.C. Thurmon (1986). Electro-immobilization
versus mechanical restraint in an avoid-avoid choice test for ewes. J Animal Science
62(6):1469-1480.
Gregory N. G., Fielding H. R., von Wenzlawowicz M. and von Holleben K. (2010).Time to
collapse following slaughter without stunning in cattle. Meat Science. Elsevier Ltd,
85(1): 66–69. doi: 10.1016/j.meatsci.2009.12.005.
Gregory N. G., Lee C. J. and Widdicombe J. P. (2007). Depth of concussion in cattle shot by
penetrating captive bolt. Meat Science, 77:499–503
Gregory N.G. and Wotton S.B. (1990a). Comparison of neck dislocation and percussion of the
head on visual evoked responses in the chicken's brain. Veterinary Records, 126:570–572
Gregory N.G. and Wotton S.B. (1990b). Effect of stunning on spontaneous physical activity and
evoked activity in the brain. British Poultry Science 31:215-220
Guais A., Brand G., Jacquot L., Karrer M., Dukan S., Grévillot G., Molina T. J., Bonte J.,
Regnier M., Schwartz L. (2011). Toxicity of carbon dioxide: A review, Chemical
Research in Toxicology, 24(12): 2061–2070. doi: 10.1021/tx200220r.
Page 209
196
Haas D.A. and Harper D.G. (1992). Ketamine: A review of its pharmacologic properties and
use in ambulatory anesthesia. Anesthesia Progress 39:61-68
Hall J. B. and McShane P.J. (2013). Ventilatory Failure. in The Merck Manual. Sharp and
Dohme Corp, Whitehouse Station, NJ.
Hameroff S. (2001). Anesthesia: The “Other Side” of Consciousness’. Consciousness and
Cognition, 10(2): 217–229. doi: 10.1006/ccog.2001.0512.
Harrop J., Sharan A., Vaccaro A.R., Przybylski G.J. (2001). The cause of neurologic
deterioration after acute cervical spinal cord injury. Spine 26:340–346.
Hari R., Portin K., Kettenmann B., Jousmaki V., Kobal G., (1997). Right-hemisphere
preponderance of responses to painful CO stimulation of the human nasal mucosa. Pain
72, 145–151.
Haselsberger K., Pucher R., Auer L. M. (1988). Prognosis after acute subdural or epidural
haemorrhage. Acta Neurochirurgica, 90(3–4): 111–116. doi: 10.1007/BF01560563.
Heard D. (2000). Perioperative supportive care and monitoring. Veterinary Clinics of North
America: Exotic Animal Practice 3(3): 587-615. https://doi.org/10.1016/S1094-
9194(17)30064-6
Hernandez E. (2018). Electroencephalographic and behavioral evaluation of physical methods
for on-farm euthanasia of poultry. Doctoral thesis, University of Guelph, Canada
Hiestand W.A. and Randall W.C. (1941). Species differenciation in the respiration of birds
following carbon dioxide administration and the location of inhibitory receptors in the
upper respiratory tract. J Cell Comp Physiol17:333.
Hogg D. A. (1982). Fusions occurring in the postcranial skeleton of the domestic fowl. J Anat.
,135(3):501-512
Hogg D.A. (1984). The development of pneumatisation in the postcranial skeleton of the
domestic fowl. J. Anatomy. 139(1): 105-113
HSA (2004). Practical Slaughter of Poultry: A Guide for the Small Producer. Humane Slaughter
Association.
Hughes B.O (1983). Head shaking in fowls: the effect of environmental stimuli. Applied Animal
Ethology 11:45–53.
Husband S. and Shimizu T. (1999). Evolution of the avian visual system.
http://luna.cas.usf.edu/~husband/evolve/default.htm.
International Association for the study of pain (IASP). Pain terms: A current list with definitions
and notes on usage. Pain Suppl. 3, S217. https://www.iasp-pain.org/
Jacobs L., Bourassa D.V., Harris C.E., Buhr R.J. (2019). Euthanasia: Manual versus mechanical
cervical dislocation for broilers. Animals 9:47. doi:10.3390/ani9020047
Page 210
197
Jaksch W. (1981). Euthanasia of day-old male chicks in the poultry industry. Int. J. Stud. Anim.
Prob. 2:203–213.
Jarvis E.D., Gu¨ ntu¨ rku¨ n, O., Bruce, L., Csillag, A., Karten, H., Kuenzel, W., Medina, L.,
Paxinos, G., Perkel, D.J., Shimizu, T., Striedter, G., Wild, J.M., Ball, G.F., Dugas- Ford,
J., Durand, S.E., Hough, G.E., Husband, S., Kubikova, L., Lee, D.W., Mello, C.V.,
Powers, A., Siang, C., Smulders, T.V., Wada, K., White, S.A., Yamamoto, K., Yu, J.,
Reiner, A., Butler, A.B. (2005). Avian Brain Nomenclature Consortium. Avian brains
and a new understanding of vertebrate brain evolution. Nature Reviews Neuroscience 6,
151–159
Jaynes J. (1985). Sensory pain and conscious pain. Behavioural and brain sciences 8:61-63
Jiang C., Rojas A., Wang R. and Wang X. (2005). CO2 central chemosensitivity: Why are there
so many sensing molecules?. Respiratory Physiology and Neurobiology, 145(2–3): 115–
126. doi: 10.1016/j.resp.2004.07.005.
KC L., A. K. Johnson, T. A. Shepherd, J. P. Stinn, H. Xin, K. J. Stalder, L. A. Karriker, M. A.
Sutherland, D. C. Lay, Jr., and S. T. Millman. (2016). Aversion to carbon dioxide gas in
pigs using approach–avoidance and conditioned place avoidance paradigms. Journal of
Animal Science, Volume 94, Issue suppl 2: 9-10. doi.org/10.2527/msasas2016-020
KC L. (2018). Investigation of Aversion Associated with Gas Inhalants in Pigs Using Cognitive
Paradigms. Graduate Theses and Dissertations, Iowa State University Digital Repository,
16726. Accessed online: https://lib.dr.iastate.edu/etd/16726.
KC L., A. K. Johnson, T. A. Shepherd, J. P. Stinn, H. Xin, K. J. Stalder, L. A. Karriker, M. A.
Sutherland, D. C. Lay, Jr., and S. T. Millman. (2016). Aversion to carbon dioxide gas in
pigs using approach–avoidance and conditioned place avoidance paradigms. Journal of
Animal Science, Volume 94, Issue suppl 2: 9-10. doi.org/10.2527/msasas2016-020
Keegan R.D., Greene S.A., Bagley R.S., Moore M.P., Weil A.B., Short C.E. (1995). Effects of
medetomidine administration on intracranial pressure and cardiovascular variables of
isoflurane-anesthetized dogs. American Journal of Veterinary Research 56:193–198.
Kinney H., Korein J., Panigrahy A., Dikkes P., Goode R. (1994). Neuropathological findings in
the brain of Karen Ann Quinlan. The New England Journal of Medicine, 330: 1469-1475.
Kirkden R. D. and Pajor E. A. (2006). Using preference, motivation and aversion tests to ask
scientific questions about animals’ feelings, Applied Animal Behaviour Science, 100(1–
2): 29–47. doi: 10.1016/j.applanim.2006.04.009.
Kirkden R.D., Niel L., Lee G., Makowska I. J., Pfaffinger M.J., Weary D.M. (2008). The validity
of using an approach-avoidance test to measure the strength of aversion to carbon dioxide
in rats. Applied Animal Behaviour Science 114: 216–234
Koechner M.F.G. Co., Inc. Tipton, MO (2016) Available online at:
http://www.turkeycoops.com/euthanizingdevice.html.
Page 211
198
Kohler I., Meier R., Busato A., Neiger-Aeschbacher G., Schatzmann U. (1999). Is carbon
dioxide (CO2) a useful short acting anaesthetic for small laboratory animals? Laboratory
Animals Ltd. Laboratory Animals 33: 155-161
Kristensen H. H., Burgess L. R., Demmers T. G. H., Wathes C. M. (2000). The preferences of
laying hens for different concentrations of atmospheric ammonia. Applied Animal
Behaviour Science 68:307-318
Label L. (1997). Injuries and disorders of the head and brain, Mosby, New York.
Lambooij E., Gerritzen M. A., Engel B., Hillebrand S. J. W., Lankhaar J., Pieterse C. (1999).
Behavioural responses during exposure of broiler chickens to different gas mixtures. Applied Animal Behaviour Science 62:255–265
Lambooij E., Reimert H., Van De Vis J. W., Gerritzen M. A. (2008). Head-to-Cloaca Electrical
Stunning of Broilers. Poultry Science, 87: 2160–2165. doi: 10.3382/ps.2007-00488.
Lancet (1976). Conference of the Royal Colleges and Faculties of the United Kingdom:
Diagnosis of brain death. Lancet 1976; 2: 1069-1070.
Lansing R.W., Gracely R.H., Banzett R.B. (2009). The multiple dimensions of dyspnea: Review
and hypotheses. Respiratory Physiology and Neurobiology 167: 53–60, 2009
Leach M. C., Bowell V. A, Allan T. F., Morton D. B. (2002). Aversion to gaseous euthanasia
agents in rats and mice. Comparative medicine, 52(3): 249–257.
Lee J., Taira T., Pihlaja P., Ransom B. R., Kaila K. (1996). Effects of CO2 on excitatory
transmission apparently caused by changes in intracellular pH in the rat hippocampal
slice. Brain Research, 706(2): 210–216. doi: 10.1016/0006-8993(95)01214-1.
Lierz M. and Korbel R. (2012). Anesthesia and Analgesia in Birds. Journal of Exotic Pet
Medicine. Elsevier Inc., 21(1): 44–58. doi: 10.1053/j.jepm.2011.11.008.
Llonch P., Rodríguez P., Jospin M., Dalmau A., Manteca X., Velarde A. (2012). Assessment of
unconsciousness in pigs during exposure to nitrogen and carbon dioxide mixtures.
Animal, pp. 1–7. doi: 10.1017/S1751731112001966.
Magnussen H., Wilmer H., Scheid P. (1979). Gas exchange in air sacs: contribution to
respiratory gas exchange in duck lungs. Respiratory Physiology. 27: 129–146.
Maina J.N. (2003). Developmental dynamics of the bronchial (airway) and air sac systems
of the avian respiratory system from day 3 to day 26 of life:a scanning electron
microscopic study of the domestic fowl, Gallus gallus variant domesticus. Anatomy and
Embryology 207 (2):119–134. DOI 10.1007/s00429-003-0333-6
Martin J. E., McKeegan D. E. F., Sparrey J., Sandilands V. (2016). Comparison of novel
mechanical cervical dislocation and a modified captive bolt for on-farm killing of poultry
on behavioural reflex responses and anatomical pathology. Animal Welfare, 25(2): 227–
241. doi: 10.7120/09627286.25.2.227.
Page 212
199
Martin J.E., McKeegan D.E.F., Sparrey J., Sandilands V. (2017). Evaluation of potential killing
performance of novel percussive and cervical dislocation tools in chicken cadavers. Br.
Poult. Sci. 58:216–223.
Martin J.E., Sandilands V., Sparrey J., Baker L., McKeegan D.E. (2018a). On farm evaluation of
a novel mechanical cervical dislocation device for poultry. Animals 8: 10.
doi:10.3390/ani8010010
Martin J.E., Sandercock D.E., Sandilands V., Sparrey J., Baker L., Sparks N.H.C., McKeegan
D.E.F. (2018b). Welfare risks of repeated application of on-farm killing methods for
poultry. Animals 8(3): 39; doi:10.3390/ani8030039
Martin J.E., Sandilands V., Sparrey J., Baker L., Dixon L.M., McKeegan D.E.F. (2019) Welfare
assessment of novel on-farm killing methods for poultry. PLoS ONE 14(2): e0212872.
https://doi.org/10.1371/journal.pone.0212872
Martoft L., Stødkilde-Jørgensen H., Forslid A., Pedersen H. D. and Jørgensen P. F. (2003). CO2
induced acute respiratory acidosis and brain tissue intracellular pH: a 31P NMR study in
swine. Laboratory animals, 37: 241–248. doi: 10.1258/002367703766453092.
Mason C., Spence J., Bilbe L., Hughes T., Kirkwood J. (2009). Methods for dispatching
backyard poultry. Veterinary Record. 164: 220
Mason J.R. and Clark L. (2000). The chemical senses in birds, in G.C. Whittow (Ed.), Sturkies
Avian Physiology, Academic Press, London
McKeegan D.E.F. (2004). Mechano-chemical nociceptors in the avian trigeminal mucosa. Brain
Research Reviews, 46: 146-154
McKeegan D.E.F., Smith F. S., Demmers T.G.M, Wathes C.M. and Jones R. B. (2005).
Behavioural correlates of olfactory and trigeminal gaseous stimulation, Gallus
domesticus. Physiology and Behavior, 84: 761-776
McKeegan D.E.F., Mclntyre J., Demmers T.J.M, Wathes C.M., Jones R.B. (2006). Behavioural
responses of broiler chickens during acute exposure to gaseous stimulation. Applied
Animal Behaviour Science, 99:271-286.
Mclelland J., (1990). A Colour Atlas of Avian Anatomy, Edinburgh, Scotland
McLeod W.M., Trotter, D.M., Lumb, J.W. (1964). Avian Anatomy. Burgess Publishing
Company, United States of America
Mellor D.J. (2016). Updating Animal Welfare Thinking: Moving beyond the “Five Freedoms”
towards “A Life Worth Living”. Animals (Basel) 6 (3): 21. doi: 10.3390/ani6030021
Mellema M.S (2008). The neurophysiology of dyspnea. Journal of Veterinary Emergency
and Critical Care 18: 561–71
Page 213
200
Melzack R. and Wall P. (1965). Pain Mechanisms: A New Theory. Science: 971–979. doi:
10.1126/science.150.3699.971.
Menon D. K., Schwab K., David W. Wright D.W., Maas A.I. (2010). Position Statement:
Definition of Traumatic Brain Injury. Arch Phys Med Rehabil Vol 91,
doi:10.1016/j.apmr.2010.05.017. Accessed online
https://www.archives-pmr.org/article/S0003-9993(10)00650-7/pdf
Merck M. Veterinary Forensics: Animal Cruelty Investigations. 2nd ed. New York, NY: Wiley;
2012.
Moberg G.P. (2000). Biological responses to stress: implications of animal welfare. In: Moberg,
G.P., Mench J.A. (eds), Biology of Animal Stress. CABI Publishing. New York, pp 1-21.
Mohandas C. (1971). Brain Death: a clinical and pathological study. J Neurosurg. :35-211
Mollaret P., Bertrand I., Mollaret H. (1959). Comadepasse et necroses nerveuses centrales
massives. Revue Neurologique, 101, 116-139.
Nasr M. A. F., Nicol C. J., Murrell J. C. (2012). Do laying hens with keel bone fractures
experience pain? PLoS ONE, 7(8). doi: 10.1371/journal.pone.0042420.
National Farm Animal Care Council Code of Practice for the Care and Handling of Pullets and
Laying Hens (2016). Available online:
http://www.nfacc.ca/pdfs/pullets_and_laying_hens_code_of_practice.pdf
National Farm Animal Care Council Code of Practice for the Care and Handling of Hatching
Eggs, Breeders, Chickens, and Turkeys. (2017).http://www.nfacc.ca/codes-of-
practice/chickens-turkeys-and-breeders
Nevarez J. G. (2005). Monitoring during avian and exotic pet anesthesia. Seminars in Avian and
Exotic Pet Medicine, 14(4): 277–283. doi: 10.1053/j.saep.2005.09.011.
Nickel R. et al. In: Anatomy of Domestic Birds. Verlag Paul Parey. Berlin; pp 118-121. (1977)
Orosz S. E. (1996). Principles of avian clinical neuroanatomy. Seminars in Avian and Exotic Pet
Medicine, 5(3): 127–139. doi: 10.1016/S1055-937X(96)80002-8.
Parshall M.B., Schwartzstein R.M., Adams L., Banzett R.B., Manning H.L.,Bourbeau J.,
Calverley P.M., Gift A.G., Harver A.., Lareau S. et al. An official American thoracic
society statement: Update on the mechanisms, assessment, and management of dyspnea.
American Journal of Respiratory and Critical Care Medicine 185, 435–52, 2012
Patterson J.L., Mullinax P.F., Bain T., Kreuger J.J. Richardson D.W. (1962) Carbon dioxide-
induced dyspnea in a patient with respiratory muscle paralysis. American Journal of
Medicine, 32:811-816.
Page 214
201
Paul-Murphy J., Fialkowski J. (2001). Injectable anesthesia and analgesia of birds. In: Gleed RD,
Ludders JW, editors. Recent Advances in Veterinary Anesthesia and Analgesia:
Companion Animals. Ithaca, NY: International Veterinary Information Service (2001).
Pavlov I.P. (1927). Conditioned Reflexes: An Investigation of the Physiological Activity of the
Cerebral Cortex. London: Oxford Univ. Press.
Pearson R., (1972), The Avian Brain, Academic Press, London.
Prinz S., Van Oijen G., Ehinger F., Bessei W., Coenen A. (2010). Effects of water bath stunning
on the electroencephalograms and physical reflexes of broilers using a pulsed direct
current. Poultry Science, 89(6): 1275–1284. doi: 10.3382/ps.2009-00136.
Prys-Roberts C. (1987). Anaesthesia- A practical or impractical construct. Br J Anaesth.
59:1341– 45.
Raj A. B. M. and N.G. Gregory. (1991). Preferential feeding behaviour of hens in different
gaseous atmospheres. British Poultry Science, 32(1): 57-65.
doi.org/10.1080/00071669108417327
Raj A. B. M., Wotton S. B. and Gregory N. G. (1992). Changes in the somatosensory evoked
potentials and spontaneous electroencephalogram of hens during stunning with a carbon
dioxide and argon mixture. British Veterinary Journal, 148(2):147–156. doi:
10.1016/0007-1935(92)90106-B.
Raj A.B.M., and Gregory N.G. (1993). Time to loss of somatosensory evoked potentials and
onset of changes in the spontaneous electroencephalogram of turkeys during gas
stunning. Veterinary Record. 133: 318–320.
Raj A.B.M., and Gregory N.G. (1994). An evaluation of humane gas stunning methods for
turkeys. Veterinary Records 135:222-223.
Raj A. B. M. and Gregory N. G. (1995). Welfare implications of the gas stunning of pigs 1.
Determination of aversion to the initial inhalation of carbon dioxide or argon. Anim.
Welfare 4:273280.
Raj B. M. (1996). Aversive reactions of turkeys to argon, carbon dioxide and a mixture of carbon
dioxide and argon. Veterinary Record 138(24): 592-593. doi: 10.1136/vr.138.24.592
Raj B. M. and O’Callaghan M. (2001). Evaluation of a pneumatically operated captive bolt for
stunning/killing broiler chickens. British poultry science, 42(3): 295–9. doi:
10.1080/00071660120055232.
Ramsaya D. S., Watson C. H., Leroux B. G., Prall C. W., Karl J., Kaiyala K. J. (2003).
Conditioned place aversion and self-administration of nitrous oxide in rats.
Pharmacology Biochemistry and Behavior, Volume 74, (3): 623-
633.https://doi.org/10.1016/S0091-3057(02)01048-1
Page 215
202
Rattenborg N. C., Martinez-Gonzalez D. and Lesku J. A. (2009). Avian sleep homeostasis:
Convergent evolution of complex brains, cognition and sleep functions in mammals and
birds, Neuroscience and Biobehavioral Reviews, 33(3), pp. 253–270. doi:
10.1016/j.neubiorev.2008.08.010.
Revzin A. M. (1965). Characteristics of the spontaneous electrical activity in the neostriatum of
the pigeon. Fed. Proc. 24: 338.
Ries C. R. and Puil E. (1999). Mechanism of anesthesia revealed by shunting actions of
isoflurane on thalamocortical neurons. Journal of neurophysiology, 81(4):pp. 1795–1801.
Rosenthal M. (1993). Mild traumatic brain injury syndrome. Annals of Emergency Medicine,
22, (6):1048-1051. DOI: https://doi.org/10.1016/S0196-0644 (05)82749-0
Sandercock D. A., Auckburally A., Flaherty D., Sandilands V. and McKeegan D. E. F. (2014)
Avian reflex and electroencephalogram responses in different states of consciousness.
Physiology and Behavior. doi: 10.1016/j.physbeh.2014.05.030.
Sandilands V., Abm R., Baker L., Nhc S. (2011). Aversion of chickens to various lethal gas
mixtures. Animal Welfare, 20(2): 253–262.
Schepens B. (2004). Independent and Convergent Signals From the Pontomedullary Reticular
Formation Contribute to the Control of Posture and Movement During Reaching in the
Cat. Journal of Neurophysiology, 92(4): 2217–2238. doi: 10.1152/jn.01189.2003.
Serpa L., Widowski. T. M., Schwean-Lardner K., Torrey S. Under pressure: determining
optimum air pressure for humane euthanasia of mature broiler breeders with an air
powered non-penetrating captive bolt device. Proceedings of the X th European
symposium on poultry welfare. Ploufragan, France. (2017).77
Seth A. K., B. J. Baars, and D. B. Edelman. 2005. Criteria for consciousness in humans and other
animals. Conscious. Cogn. 14:119–139
Sharp T. M., McLeod S. R. , Leggett K. E. A., Gibson T. J. (2014). Evaluation of a spring-
powered captive bolt gun for killing kangaroo pouch young. Wildlife Research 41(7)
623-632 https://doi.org/10.1071/WR14094
Shaw N.A. (2002). The neurophysiology of concussion. Prog. Neurobiology 67: 281-344
Siegal A., and Sapru H. N. (2006). Essential neuroscience. Philadelphia, PA: Lippincott,
Williams & Wilkins
Silver W.L., Maruniak, J.A., (1981). Trigeminal chemoreception in the nasal and oral cavities.
Chem. Senses 6, 295–305.
Sinclair M.D. (2003). A review of the physiological effects of α2-agonists related to the clinical
use of medetomidine in small animal practice. Canadian Veterinary Journal 44(11): 885–
897.
Page 216
203
Sommerhoff G. and MacDorman K. (1994). An account of consciousness in physical and
functional terms: A target for research in the neurosciences. Integrative Physiological and
Behavioral Science, 29(2): 151–181. doi: 10.1007/BF02691012.
Sparrey J, Sandercock D.A., Sparks N.H.C., Sandilands V. (2014). Current and novel methods
for killing poultry individually on-farm. Worlds Poult Sci J. 70:737–58. doi:
10.1017/S0043933914000816
Starzl T. E., C. W. Taylor, and H. W. Magoun. 1951. Ascending conduction in reticular
activating system, with special reference to the diencephalon. J. Neurophysiol. 14:461–
477.
Stinson J. M., and Mattsson J. L. (1970). Tolerance of rhesus monkeys to graded increase in
environmental CO2- Serial changes in heart rate and cardia rhythm. Aerosp. Med. 42:78 –
80.
St-John W. M., Paton J. F. R. and Leiter J. C. (2004). Uncoupling of rhythmic hypoglossal from
phrenic activity in the rat. Experimental Physiology, 89(6): pp. 727–37. doi:
10.1113/expphysiol.2004.028829.
St-John W. M. (2009). Noeud vital for breathing in the brainstem: gasping--yes, eupnoea--
doubtful. Philosophical transactions of the Royal Society of London. Series B, Biological
sciences, 364(1529): 2625–33. doi: 10.1098/rstb.2009.0080.
Sun M., Sun C., Yang Y. (1996). Effect of low-concentration CO2 on stereoacuity and energy
expenditure Aviation, Space, and Environmental Medicine, 67(1):34-39
Taneichi H., Suda K., Kajino T., Kaneda K. (2005). Traumatically Induced Vertebral Artery
Occlusion Associated with Cervical Spine Injuries: Prospective Study Using Magnetic
resonance angiography. Spine (Phila Pa 1976), 30(17):1955-62.
Terlouw C., Bourguet C. Deiss V. (2016a). Consciousness, unconsciousness and death in the
context of slaughter. Part I. Neurobiological mechanisms underlying stunning and killing.
Meat Science. Elsevier Ltd, 118: 133–146. doi:
http://dx.doi.org/10.1016/j.meatsci.2016.03.010
Terlouw C., Bourguet C. Deiss V. (2016b). Consciousness, unconsciousness and death in the
context of slaughter. Part II. Evaluation methods. Meat Science. Elsevier Ltd, 118: 147–
156. doi: 10.1016/j.meatsci.2016.03.010.
Thomas P. D. (2000). The differential diagnosis of fixed dilated pupils: a case report and review
[online]. Critical Care and Resuscitation: journal of the Australasian Academy of Critical
Care Medicine, 2(1): 34–37.
<https://search.informit.com.au/documentSummary;dn=524263947124870;res=IELHEA
> ISSN: 1441-2772.
Page 217
204
Tidswell S., Blackmore D., Newhook J. (1987). Slaughter methods: electro- encephalographs
(EEG) studies on spinal cord section, decapitation and gross trauma of the brain in lambs.
New Zealand Veterinary Journal, 35: 46–49.
Tseng W.C., Shih H.M., Su Y.C., Chen H.W., Hsiao K.Y., Chen I.C. (2011). The association
between skull bone fractures and outcomes in patients with severe traumatic brain injury.
J. Trauma 71:1611–1614. doi: 10.1097/TA.0b013e31823a8a60
Turner B. H. and Knapp M. E. (1995). Consciousness: A neurobiological approach’, Integrative
Physiological and Behavioral Science, 30(2):151–156. doi: 10.1007/BF02691683.
Turner P.V., Kloeze H., Dam A., Ward D., Leung N., Brown E.E.L., Whiteman A., Ciappetta
M.E., Hunter D.B. (2012). Mass depopulation of laying hens in whole barns with liquid
carbon dioxide: Evaluation of welfare impact. Poult. Sci. (2012) 91:1558–1568. doi:
10.3382/ps.2012-02139
Vander A. J., Sherman J. H., Luciano D. S. (2001). Human physiology the mechanisms of body
function, 8th ed. Boston: McGraw-Hill.
Veras L., Pedraza-Gutiérrez, S., Castellanos, J., Capellades, J., Casamitjana J., Rovira- Cañellas
A. (2000). Vertebral Artery Occlusiion After Acute Cervical Spine Trauma. Spine 25:
1171-1177.
Verhoeven M. T. W., Gerritzen M. A, Hellebrekers L. J. and Kemp, B. (2015) ‘Indicators used in
livestock to assess unconsciousness after stunning: a review.’, Animal: an international
journal of animal bioscience, 9(2), pp. 320–30. doi: 10.1017/S1751731114002596.
Von-Leupoldt A., Sommer T., Kegat S., Baumann H.J., Klose H., Dahme B., Buchel C. (2008).
The unpleasantness of perceived dyspnea is processed in the anterior insula and
amygdala. American Journal of Respiratory and Critical Care Medicine 177,1026–32,
2008
Walsh J., Percial A., Turner P.V. (2017). Efficacy of blunt force trauma, mechanical cervical
dislocation, and a non-penetrating captive bolt device for on-farm euthanasia of pre-
weaned kits, growers, and adult commercial meat rabbits. Animals 7:100
Waran N., Williams V. M., Clarke N., Bridge I. S. (2010). Recognition of pain and use of
analgesia in horses by veterinarians in New Zealand. New Zealand veterinary journal, 58:
274–280. doi: 10.1080/00480169.2010.69402.
Webster, A. B., and D.L. Fletcher. (2001). Reactions of laying hens and broilers to different
gases used for stunning poultry. Poultry Science 80(9): 1371-1377. doi:
10.1093/ps/80.9.1371
Webster A. B. and Fletcher D. L. (2004). Assessment of the aversion of hens to different gas
atmospheres using an approach-avoidance test. Applied Animal Behaviour Science,
88(3–4): 275–287. doi: 10.1016/j.applanim.2004.04.002.
Page 218
205
White B.C. and Krause G.S. (1993). Brain injury and repair mechanisms: the potential for
pharmacologic therapy in closed-head trauma. Annals of Emergency Medicine 22: 970-
979. doi.org/10.1016/S0196-0644 (05)82734-4
Whiting T. L., Steele G. G., Wamnes S., Green C. (2011). Evaluation of methods of rapid mass
killing of segregated early weaned piglets. Can Vet J. 52:753–758
Whittow G.C. (ed): Sturkie’s Avian Physiology (ed 5). (2000). San Diego, CA, Academic Press
Widowski T., Woolcott C., Turner P., Schwean-Lardner K., Caston L., Torrey S. (2018).
Evaluation of a mechanical cervical dislocation tool for growing and mature turkeys
(Abstract). Proceedings of Poultry Science Association 107th Annual Meeting, pg 97
Widowski T. M., Elgie R. H., Lawlis P. (2008). Assessing the effectiveness of a non-penetrating
captive bolt for euthanasia of newborn piglets. Differences, 107-111.
Wilberger J. E., Harris M. and Diamond D. L. (1991). Acute subdural hematoma: morbidity,
mortality, and operative timing. Journal of Neurosurgery, 74(2): 212–218. doi:
10.3171/jns.1991.74.2.0212.
Winslow C., Rozovsky J. (2003). Effect of spinal cord injury on the respiratory system. Am J
Phys Med Rehabil, 82:803–814. DOI: 10.1097/01.PHM.0000078184.08835.01
Withrock I.C. (2015). The Use of Carbon Dioxide (CO2) as an Alternative Euthanasia Method
for Goat Kids. Graduate Theses and Dissertations, Iowa State University Digital
Repository, 14718. Accessed online: https://lib.dr.iastate.edu/etd/14718
Woolcott C.R., Torrey S., Turner P.V., Serpa L., Schwean-Lardner K., Widowski T.M.
Evaluation of two models of non-penetrating captive bolt devices for on-farm euthanasia
of turkeys. Animals (2018a) 8:42-58. doi:10.3390/ani8030042
Woolcott C.R., Torrey S., Turner P.V., Chalmers H., Levison L., Schwean-Lardner K.,
Widowski T.M. (2018b). Assessing a method of mechanical cervical dislocation as a
humane option for on-farm killing using anesthetized poults and young turkeys. Front.
Vet. Sci. 5:275 doi.org/10.3389/fvets.2018.00275
Yang Y., Sun C., Sun M. (1987). The effect of moderately increased CO2 concentration on
perception of coherent motion. Aviation, space and environment medicine, 68:187-191
Yew D. T. W., Yu M. C., Kwong W. H. (1996). Basic Neuroanatomy. World Scientific
Publishing Co. Pte. Ltd. Singapore.
Zar J.H. (1998). Biostatistical Analysis. 4th ed. Prentice Hall, Upper Saddle River, NJ
Zeman A. (2005). What in the world is consciousness?. Progress in Brain Research, 150:1–10.
doi: 10.1016/S0079-6123(05)50001-3.
Zerari-Mailly F., Dauvergne C., Buisseret P., Buisseret-Delmas C. (2003). Localization of
Trigeminal, Spinal, and Reticular Neurons Involved in the Rat Blink Reflex. Journal of
Comparative Neurology, 467(2): 173–184. doi: 10.1002/cne.10917.
Page 219
206
Zornow M.H., Fleischer J.E., Scheller M.S. (1990). Dexmedetomidine, and α2-adrenergic
agonist, decreases cerebral blood flow in the isoflurane-anesthetized dog. Anesthesia and
Analgesia 70:624–630
Zulkifli I., Goh Y., Norbaiyah B., Sazili A., Lotfi M. and Small A. H. (2014). Changes in blood
parameters and electroencephalogram as affected by different stunning methods in cattle.