64
Atomic Resolution Structures of Amyloid Fibrils ------- Sedimented Protein DNP Dynamic Nuclear Polarization and Dipolar Recoupling Winter School Stowe, VT January 11, 2013 MAS MAS+CryoEM

Atomic Resolution Structures of Amyloid Fibrils-------
Sedimented Protein DNPDynamic Nuclear Polarization and Dipolar Recoupling
Winter SchoolStowe, VT
January 11, 2013
MAS MAS+CryoEM

CollaboratorsCyro-EM
Anthony FitzpatrickHelen Saibil
Transthyretin105-115Galia Delbouchina
Marvin BayroChris Jaroniec
Vik BajajMarc Caporini
Patrick van der WelAlexander Barnes
Cait MacPheeMichele Vendruscuolo
Chris Dobson
β-2-MicroglobulinGalia Delbouchina
Geoffrey PlattMarvin Bayro
Sheena Radford
PI3-SH3Marvin BayroNeil BirkettMatt Eddy
Cait MacPheeChris Dobson
National Institute of Biomedical Imaging and Bioengineering
Sedimented Protein DNPEnrico Ravera
Bjoern CorziliusVladimir MichaelisClaudio Luchinat
Ivano Bertini
EB00315, EB002804, EB002026

Amyloid fibrils
Sup35 fibril strains in S. cerevisiae lead to different phenotypes.
Tessier & Lindquist, NSMB, 2009
Func%onal amyloid
β2-‐microglobulin amyloid deposits
(dialysis-‐related amyloidosis)
Floege & Ehlerding Nephron,1996
Pathogenic amyloid
Non-‐crystalline and insolubleDipolar recoupling & DNP

TTR 105-11 Amyloid Fibrils
• Binds the dye congo red --> green birefringence
• “Cross-β” powder X-ray diffraction pattern
• Twisted fibrillar structures shown above
• Insoluble and do not diffract to high resolution MAS NMR
TTR L111M: YTIAALMSPYSTTR 105-115: YTIAALLSPYS

Amyloid protofilament structure -‐ MAS
4.7 Å
~10 Å
characterisGc cross-‐β structure fib
ril axis
secondary structure
intra-‐sheetorganizaGon
protofilamentinter-‐sheetorganizaGon
protofilamentinteracGons
Levels of structural organiza%on
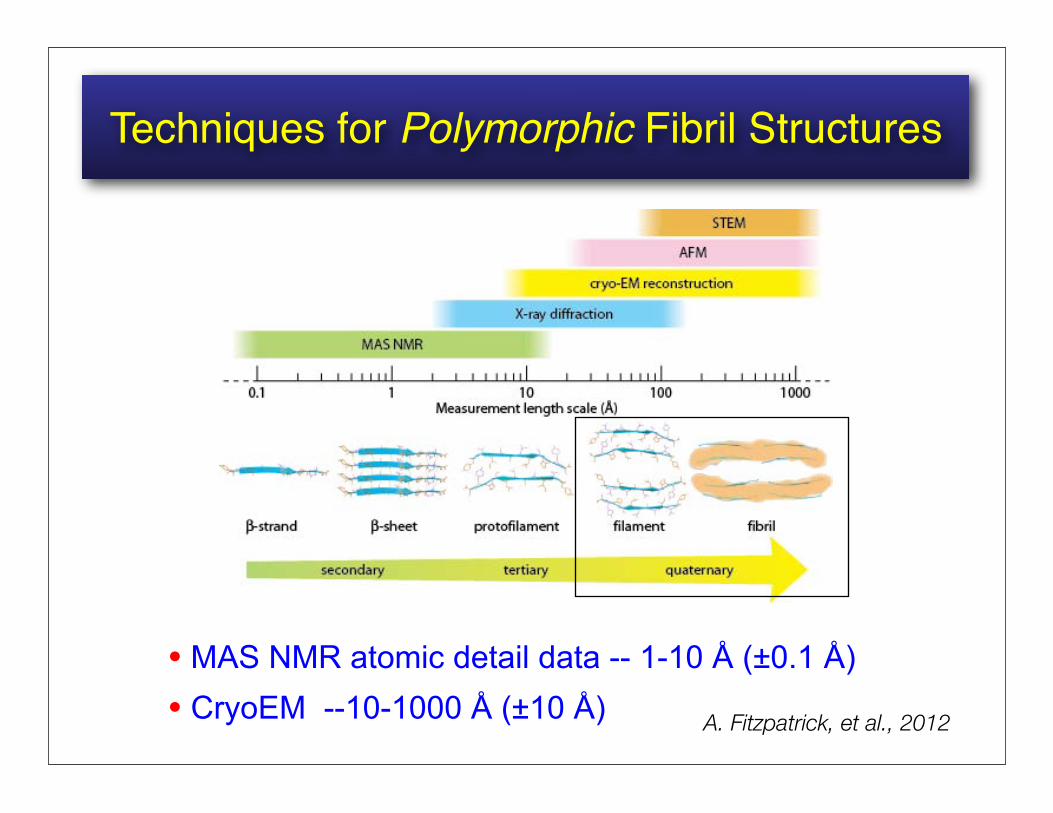
Techniques for Polymorphic Fibril Structures
• MAS NMR atomic detail data -- 1-10 Å (±0.1 Å)• CryoEM --10-1000 Å (±10 Å) A. Fitzpatrick, et al., 2012

13C-15N and 13C-13C Distance Measurements
Rotational Resonance (R2) Width3D experiment yielding multiple
13C-13C dipolar couplings
Raleigh, Levitt and Griffin(1988) Chem. Phys. Lett. 146, 71-76
Ramachandran, Ladizhansky, Bajaj, Griffin(2003) JACS 125:15623-9
Ramachandran, van der Wel, Lewandowski, Griffin(2006) J. Chem. Phys. 124, 214107
EvolutionMixing/2 Mixing/2
3D ZF-TEDORObtaining multiple 15N-13C
couplings per 3DJaroniec, Filip, Griffin
(2002) JACS 124, 10728-10742
Hing, Vega, Schaefer(1992) JMR 96, 205-209

13C-15N and 13C-13C Distance Measurements
Rotational Resonance (R2) Width3D experiment yielding multiple
13C-13C dipolar couplings
Raleigh, Levitt and Griffin(1988) Chem. Phys. Lett. 146, 71-76
Ramachandran, Ladizhansky, Bajaj, Griffin(2003) JACS 125:15623-9
Ramachandran, van der Wel, Lewandowski, Griffin(2006) J. Chem. Phys. 124, 214107
EvolutionMixing/2 Mixing/2
3D ZF-TEDORObtaining multiple 15N-13C
couplings per 3DJaroniec, Filip, Griffin
(2002) JACS 124, 10728-10742
Hing, Vega, Schaefer(1992) JMR 96, 205-209

Summary of SSNMR Restraints
• About 70 structural restraints in the form of backbone torsion angles, 13C-15N and 13C-13C distances
• Precision of restraints: torsion angles ~ ±20o, distances ~ ±0.2-0.5 Å
• Low density of restraints for Tyr side-chains and C-terminus
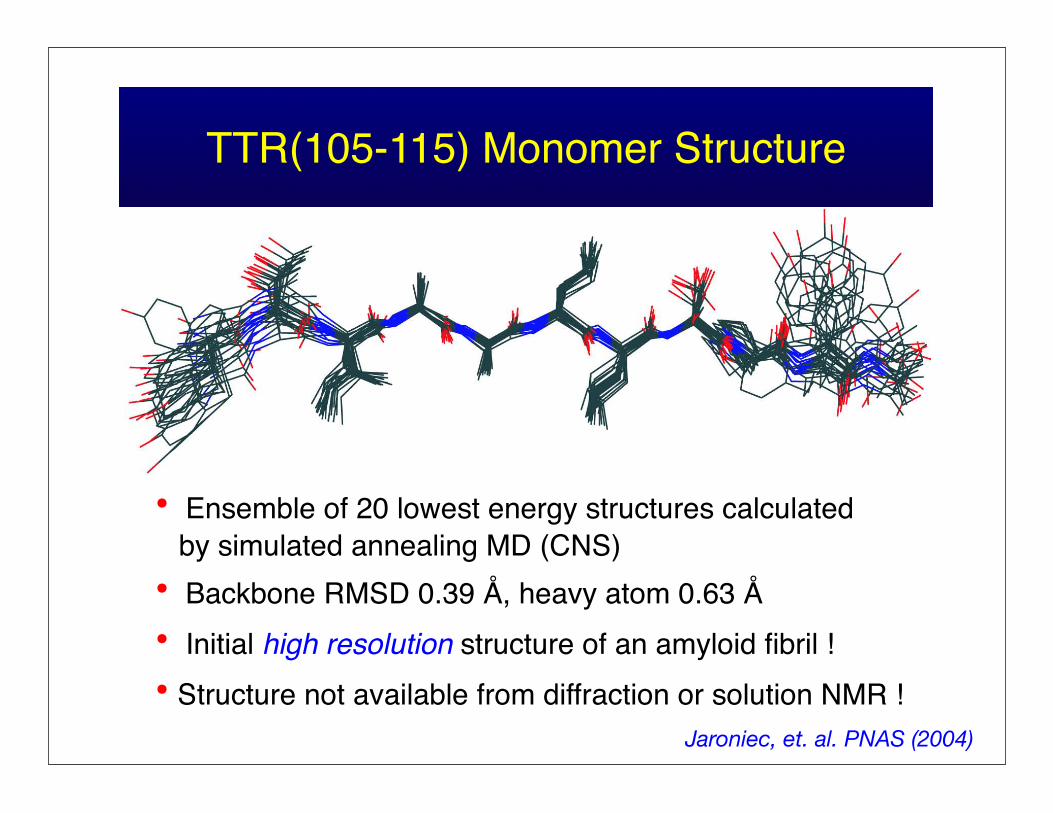
TTR(105-115) Monomer Structure
• Ensemble of 20 lowest energy structures calculated by simulated annealing MD (CNS)• Backbone RMSD 0.39 Å, heavy atom 0.63 Å• Initial high resolution structure of an amyloid fibril !• Structure not available from diffraction or solution NMR !
Jaroniec, et. al. PNAS (2004)

Structural models by MAS NMRAβ(1-‐40)
Alzheimer’s disease
Wasmer et al., Science, 2008
HET-‐s(218-‐289)P. anserina
β-‐helix
Paravastu et al. PNAS, 2008Petkova, Biochemistry, 2006
parallel, in-‐register

Amyloid fibril structure with DNP
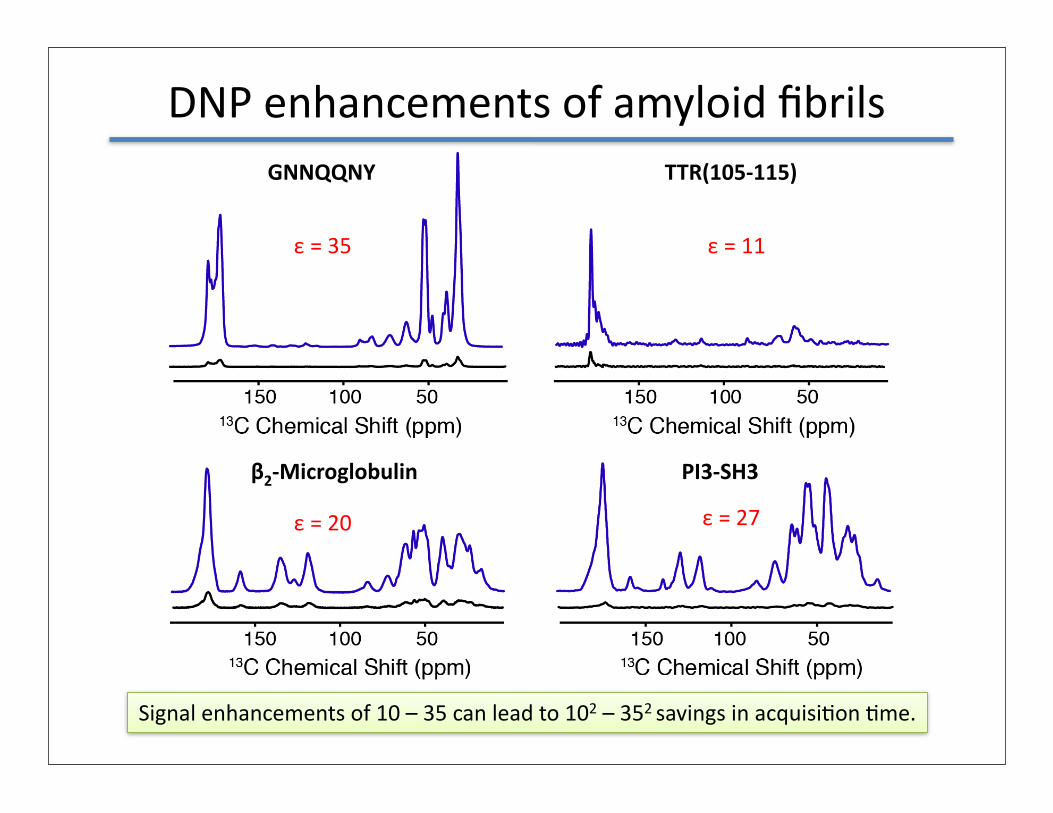
DNP enhancements of amyloid fibrilsGNNQQNY TTR(105-‐115)
β2-‐Microglobulin PI3-‐SH3
ε = 35 ε = 11
ε = 20 ε = 27
Signal enhancements of 10 – 35 can lead to 102 – 352 savings in acquisiGon Gme.

Distance constraints with DNP
TTR(105-‐115) fibrils
DQ-‐DRAWS experiment to measure a 4.3 Å 13C-‐13C distance.
avec DNP45 min
sans DNP3.5 days
PI3-‐SH3 fibrils
TEDOR experiment used to obtain inter-‐molecular constraints.
Bayro MJ, Debelouchina GT et al., JACS, 2011, 13967Debelouchina GT, Bayro MJ, Fitzpatrick A et al., in prepara,on
sans DNP16 days
avec DNP1.5 days
15N Chemical shid (ppm)

The atomic resoluGon structure of TTR(105-‐115) fibrils

Fibril Structure
secondary structure
intra-‐sheetorganizaGon
inter-‐sheetorganizaGon
protofilamentinteracGons
MAS NMR (and DNP)
Cryo-‐EMand AFM
+

Structure of the TTR(105-‐115) monomer
• One of the first structures obtained by biomolecular MAS NMR.
• 70 structural restraints including backbone torsion angles, 13C-‐15N and 13C-‐13C distances.
Jaroniec CP, MacPhee CE, Bajaj VS, McMahon MT, Dobson CM, Griffin RG, PNAS, 2004

13C=O-13C=O Distance MeasurementsParallel β-strands
• DQF DRAWS is more sensitive to distance than dephasing experimentsInfinite linear chain of spins
M. Caporini , V. Bajaj (2010)
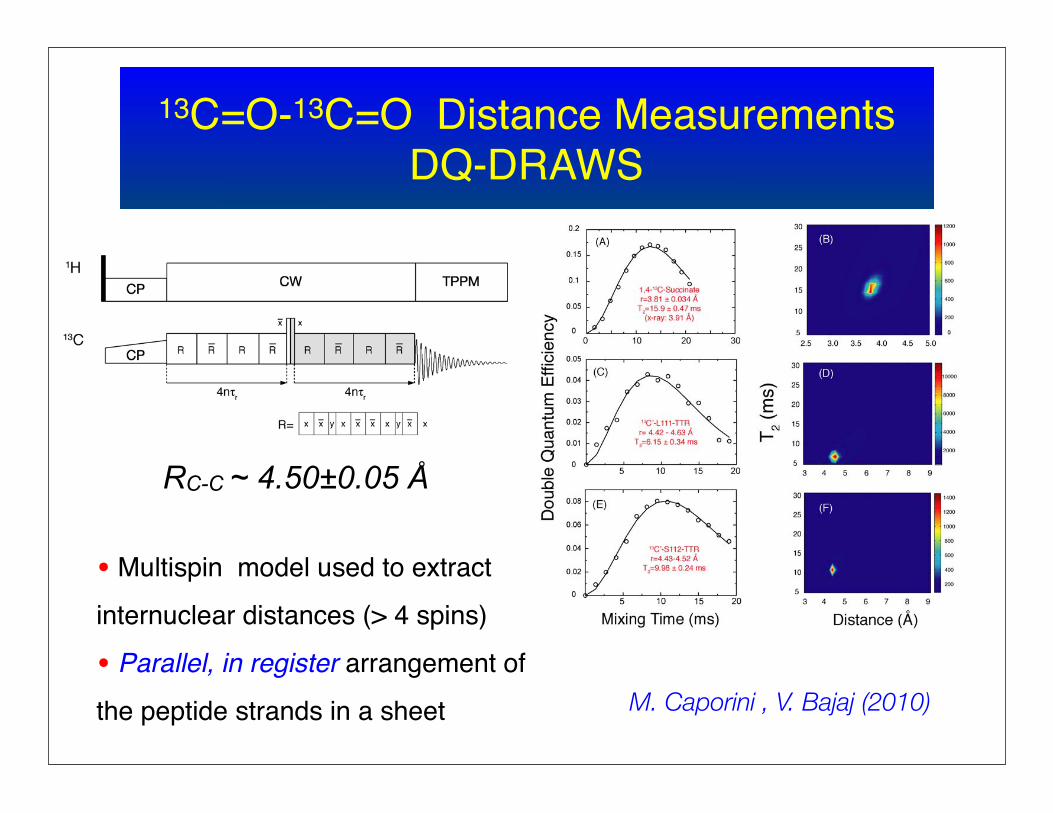
13C=O-13C=O Distance MeasurementsDQ-DRAWS
• Multispin model used to extract
internuclear distances (> 4 spins)
• Parallel, in register arrangement of
the peptide strands in a sheet
RC-C ~ 4.50±0.05 Å
M. Caporini , V. Bajaj (2010)

Intermolecular Distance Measurements
TTR
• Label a single 13C=O site; use DQ DRAWS experiment
Typical fit-curveFor Ser-112
• All 7 distances consistent with a parallel in register β-sheet ! Botto (DRAWS dephasing); Tycko (RFDR) (2001)

Intra-‐sheet arrangement
Caporini et al., J. Phys. Chem. B, 2010Debelouchina, Bayro et al, in prepara,on
DNP
S115 CO4.26 ± 0.03 Å
45 min
fibril axis
DQ-‐DRAWS experiment
• Prepare 8 different samples.• Each has a different CO labeled.• Measure inter-‐molecular CO-‐CO distances.
parallel, in-‐register β-‐strands

Fibril Structure
secondary structure
intra-‐sheetorganizaGon
inter-‐sheetorganizaGon
protofilamentinteracGons
MAS NMR (and DNP)
Cryo-‐EMand AFM
+

TTR 105-115 YTIAallSPYs13C-15N TEDOR Spectra
• ODD-EVEN x-peaks -- i.e., P113-A108
• U-[13C/15N]-YTIA/SPY TTR fibrils
• 10.24 ms TEDOR, 750 MHz, ωr/2π=12.5 kHz

TTR 105-115 YTIAallSPYs13C-13C PDSD Spectra
• U-[13C/15N]-YTIA/SPY TTR fibrils
• 200 ms PDSD, 900 MHz, ωr/2π=11 kHz
•ODD-EVEN x-peaks ....I107-S112A108-P113

Inter-‐sheet arrangement
10 ms TEDOR
-‐ 23 quanQtaQve and qualitaQve constraints-‐ anQparallel β-‐sheets-‐ even-‐odd-‐even-‐odd interface-‐ defines protofilament
fibril axis

Protofilament arrangement
head-‐to-‐tail head-‐to-‐head

3 -‐ 4 Å > 7 Å
[15N-‐Y]TIAALLSPY[13C1-‐S]
15N -‐13C experiment performed
Head-‐to-‐tail protofilament arrangement.
3.5 ± 0.2 Å
DNP
Protofilament arrangement

Different chemical environments
Data is consistent with four different chemical environments for the termini.

Atomic resolution structure of TTR(105-115) amyloid fibrils
fibril axis
fibril axis
protofilament
-‐ 10 constraints/residue-‐ several constraints obtained with DNP-‐ protofilament-‐to-‐protofilament contacts observed for the first Qme

Protofibrils to FibrilsCryoEM + MAS
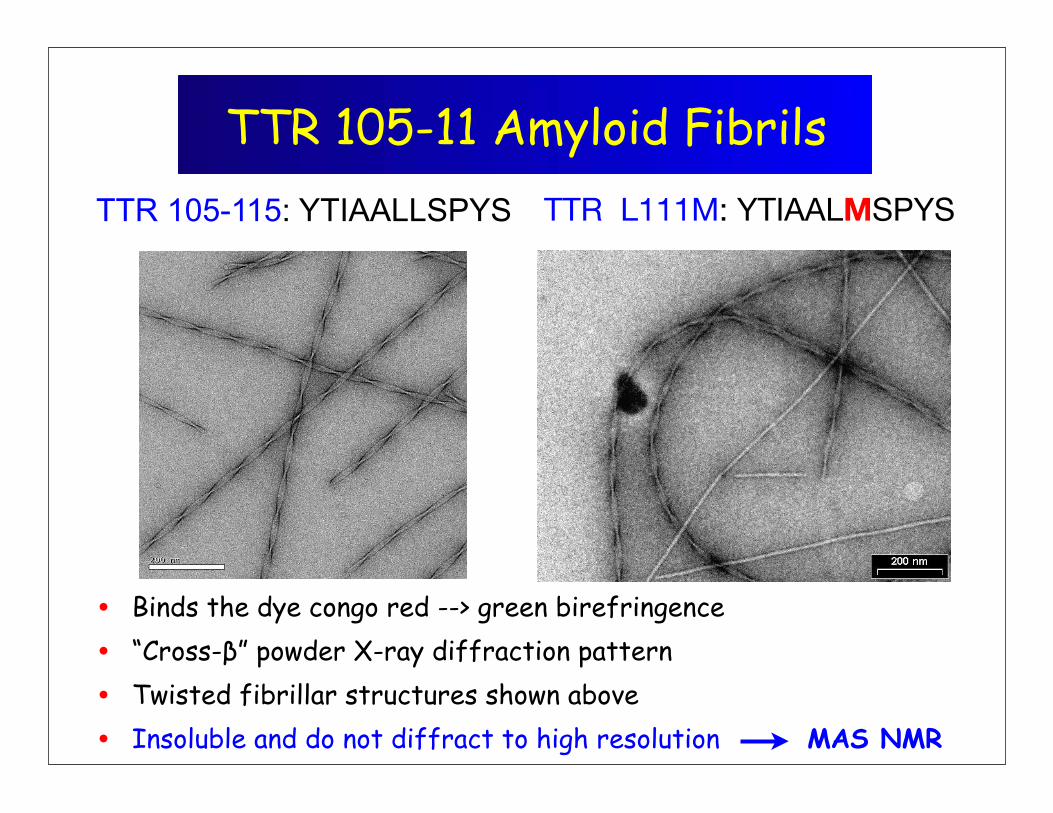
TTR 105-11 Amyloid Fibrils
• Binds the dye congo red --> green birefringence
• “Cross-β” powder X-ray diffraction pattern
• Twisted fibrillar structures shown above
• Insoluble and do not diffract to high resolution MAS NMR
TTR L111M: YTIAALMSPYSTTR 105-115: YTIAALLSPYS

• Based on the high resolution structure of the monomer ~70 constraints (±0.3-0.5 Å)
• Intermolecular distances and packing constraints ~ 40 constraints (±0.05 Å)
• Cryo-EM and STEM data to refine supramolecular structure
• “Wet” interface and “ steric zipper”
A. Fitzpatrick, Helen Saibil, et al.
Structure of the TTR amyloid fibrilSTEM Mass/Length
8 sheets
16 sheets
12 sheets
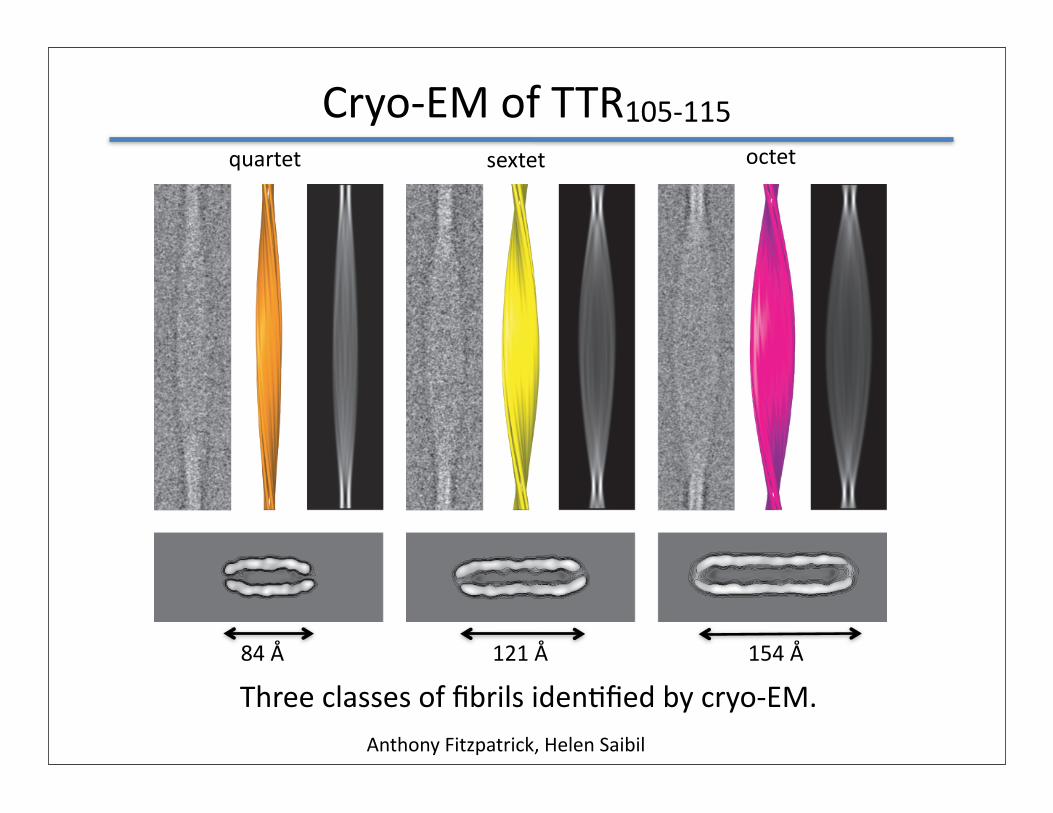
Cryo-‐EM of TTR105-‐115
Anthony Fitzpatrick, Helen Saibil
Three classes of fibrils idenQfied by cryo-‐EM.
quartet sextet octet
84 Å 121 Å 154 Å

Quartet cross-‐secGon
solvent solvent
A solvent cavity is accommodated in the fibril interior.
10 Å

Polymorphic Fibril Cross Sec,ons
35
84 ÅQuartet
121 ÅSextet
154 ÅOctet
• 3 different polymorphs -‐-‐ quartet, sextet and octet

Sextet -- MAS NMR and cryoEM
data from NMR only
NMR+
cryoEMelectrondensity
• Use MAS and cryoEM to determine atomic level structure of fibrils !

Polymorphic TTR105-115 Fibrils
77 ÅQuartet
116 ÅSextet
149 ÅOctet
•Three different periods
•Three different widths
•Note H2O layer

Amyloid fibrils formed by full-‐length proteins

~600
Å
42 Å
PI3-SH3 Cryo-EM Architecture
Four ProtofibrilsJimenez et al. EMBO J. 18 (1999) 815
14Å
2 β−sheets
Phosphatidylinositol 3-kinaseP85α subunit SH3 domain
64 x 38 ÅH2O ?
pH=2

PI3-SH3 protein fibrils-- EM and NMR
PI3-SH3
Cryo-EM: Jiménez et al. (1999) EMBO J. 18: 815
PI3-SH3 fibrils
• Dimensions -- consistent with cryoEM from Jiminez, et. al.(1999)
• Parallel, in-register alignment, contacts between strands
•Additional constraints to refine structure calculation....Bayro et. al. Biochemistry (2010)

PI3-SH3 protein fibrils- assignments
PI3-SH3
Sequential assignments RFDR, 750 MHz, ωr/2π= 20.161 kHz 0.8 ms
mixing, no 1H decoupling
• Excellent resolution -- 0.5 ppm linewidths
• 75/86 residues sequentially assigned
• 87% assigned !
Bayro, et al. Biochemistry (2010)

Intra-‐sheet arrangement
Mixed sample
50% 15N50% 13C (2-‐glycerol)
fibril axis
Target intermolecular N-‐Ca correlaQons.Distances ~ 4.5 Å.
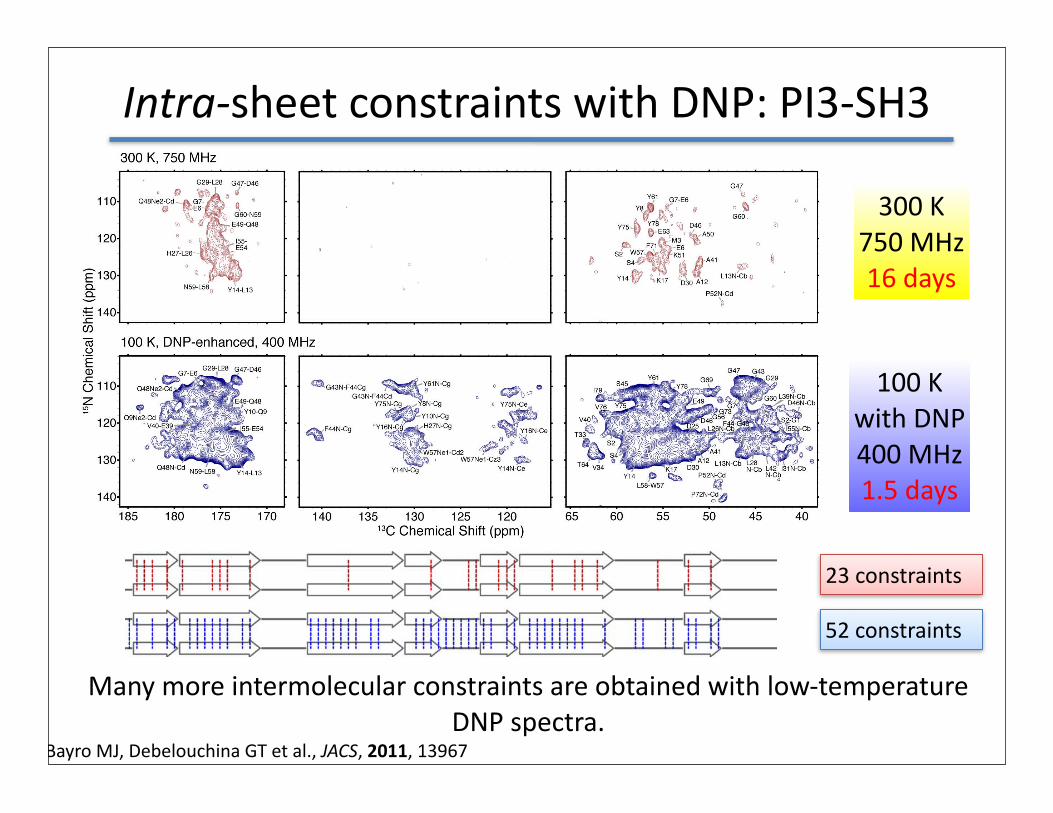
Intra-‐sheet constraints with DNP: PI3-‐SH3
b2m
MHC-‐I 300 K750 MHz16 days
100 Kwith DNP400 MHz1.5 days
Many more intermolecular constraints are obtained with low-‐temperature DNP spectra.
Bayro MJ, Debelouchina GT et al., JACS, 2011, 13967
23 constraints
52 constraints

Protofilament Structural Constraints
• Needed: Intersheet constraints• Contacts between protofilaments also possible
Model of the fibrilcross-section

15N-13C Intersheet ContactsZF-TEDOR
• Backbone 15N – sidechain 13C contacts• Also sidechain 15N – sidechain 13C• Distance estimates from build-up curve
45
15 ms mixing period12.5 kHz MAS1,3-PI3-SH3
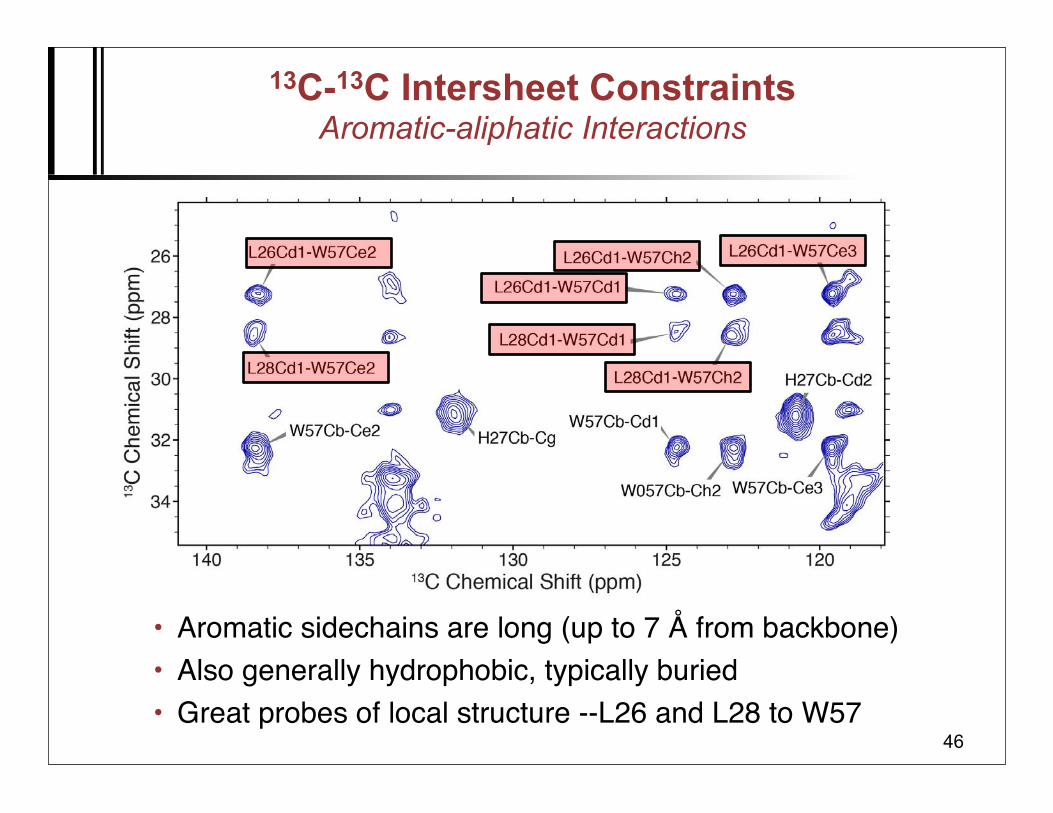
13C-13C Intersheet ConstraintsAromatic-aliphatic Interactions
• Aromatic sidechains are long (up to 7 Å from backbone)• Also generally hydrophobic, typically buried• Great probes of local structure --L26 and L28 to W57
46

Current structural constraints/model
• Data agrees with the proposed model• Also defines positions of sidechains• Ongoing work

Preliminary Structure -- PI3-SH324-61
• Contacts agree with our proposed model• Defines positions of sidechains• Additional constraints in the 1-26 and 61-86
W57L26
L28L39
W57
L26L28
Q48
I31 ?
I55 ?
N35 ?
E53 ?
RMSDBackbone: 1.66 ÅHeavy-atom: 2.41 Å

~600
Å
42 Å
PI3-SH3 Cryo-EM Architecture
Four ProtofibrilsJimenez et al. EMBO J. 18 (1999) 815
14Å
2 β−sheets
Phosphatidylinositol 3-kinaseP85α subunit SH3 domain
64 x 38 ÅH2O ?
pH=2

Sedimented Protein NMR and DNP
Ivano BertiniDecember 1940 - July 2012

Motivation – NMR of large biolecules
liquid-state solid-state
intermediatetumbling
deuterationTROSY
micro-crystallization
immobilization by sedimentationDNP

Protein sedimentation & cryoprotection?
• Glycerol prevents sedimentation (density and viscosity)
supernatantwater
proteinsediment
glycerol
removesupernatant seal
removesuper-natant
addglycerol
mix &seal
~700 mg/mL
~280 mg/mL

SedNMR In situ sedimentation during MAS
homogeneous solution of protein60 mg/mL
sedimented protein at rotor walls(thin, but high local concentration)
≤700 mg/mL
distance from rotor axis (mm)
12 kHz7 kHz4 kHz
2 kHz
1 kHz
static
1.36 1.38 1.481.461.441.421.40 1.50
1.5
1.0
0.5lo
cal p
rote
in c
onc.
(mM
)magic angle spinning
Enrico Ravera, Claudio Luchniat & Ivano Bertini
Bertini et al, PNAS (2011)
apoferritinmonomer: 20 kDa24-mer: 480 kDa
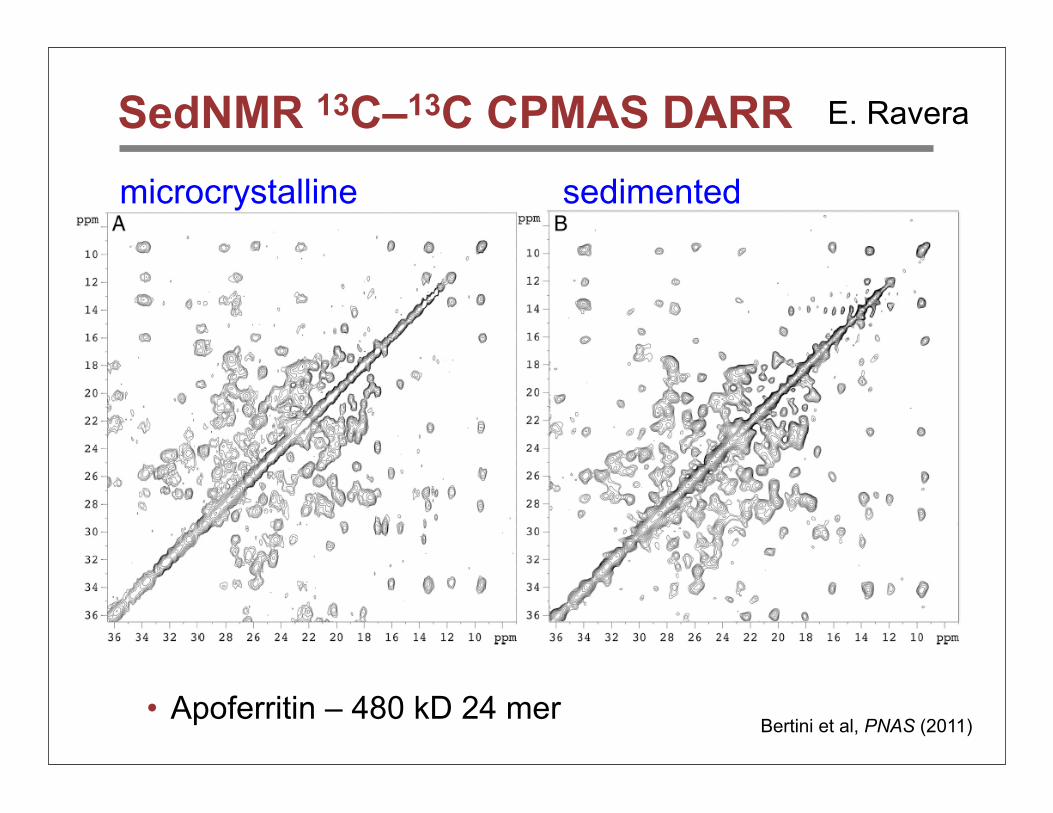
SedNMR 13C–13C CPMAS DARR
• Apoferritin – 480 kD 24 merBertini et al, PNAS (2011)
microcrystalline sedimented
E. Ravera

Sedimented DnaB helicase (12 × 57 kDa)se
dim
ente
dm
icro
crys
talli
ne
Gardiennet et al., ACIEE 2012

SedDNP -- can sedimented proteins be polarized ?
homogeneous solution of protein(low concentration)
magic angle spinning
sedimented protein at rotor walls(thin but high concentration)
frozen solution of proteincrystalline!
frozen protein sediment at rotor wallsamorphous/glassy?
80 K
80 K
DNP ?
µw
µw

In situ SedDNP – apoferritin
• Sedimented solution -- 1H and direct 13C DNP via cross effect• Frozen solution -- no significant DNP, phase separation
Sedimented solution
Frozen solution

2D: 24 × 20 kDa protein @ 212 MHz
• 13C–13C correlation spectra (PDSD) can be obtained within few hours• 6 times larger sensititvity of sedimented solution• Resolution difficult to assess due to low field (5 T)
Sedimented solution 60/40 glycerol/water solution
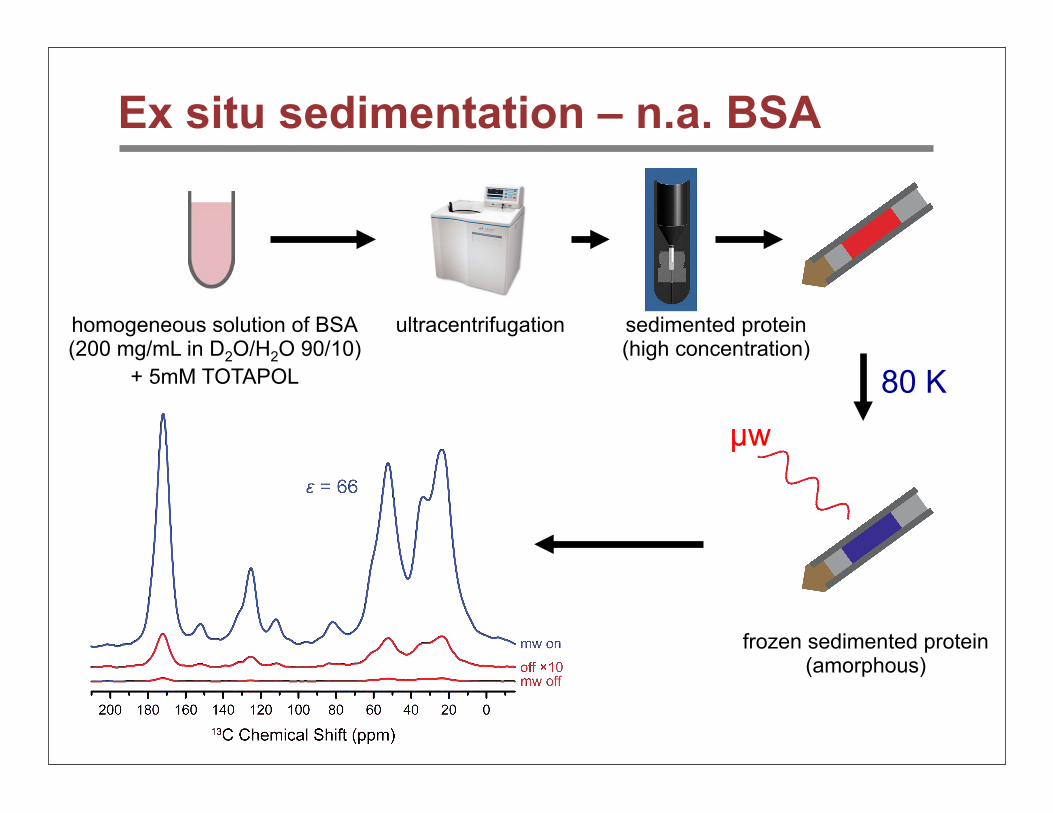
Ex situ sedimentation – n.a. BSA
µw
homogeneous solution of BSA(200 mg/mL in D2O/H2O 90/10)
+ 5mM TOTAPOL
ultracentrifugation sedimented protein(high concentration)
frozen sedimented protein(amorphous)
80 K

BSA – sensitivity
• Sediments yield superior sensitivity – up to 4.5-fold larger than glycerol solution

Conclusions
Amyloid Protein Structure
• PI3-SH3 assignments • Non-native, parallel in-register
Polymorphic TTR Fibril Structure
• Monomer structure, and strand and sheet alignment
• CyroEM and STEM

Conclusions
Sedimented Protein DNP (SedDNP)
• functionally -- “microcrystalline glass”

CollaboratorsCyro-EM
Anthony FitzpatrickHelen Saibil
Transthyretin105-115Galia Delbouchina
Marvin BayroChris Jaroniec
Vik BajajMarc Caporini
Patrick van der WelAlexander Barnes
Cait MacPheeMichele Vendruscuolo
Chris Dobson
β-2-MicroglobulinGalia Delbouchina
Geoffrey PlattMarvin Bayro
Sheena Radford
PI3-SH3Marvin BayroNeil BirkettMatt Eddy
Cait MacPheeChris Dobson
National Institute of Biomedical Imaging and Bioengineering
Sedimented Protein DNPEnrico Ravera
Bjoern CorziliusVladimir MichaelisClaudio Luchinat
Ivano Bertini

Thank you for
your attention!

















