268
AUSTRALIA’S BIODIVERSITY – RESPONSES TO FIRE Plants, birds and invertebrates Biodiversity Technical Paper, No. 1 A.M. Gill, J.C.Z. Woinarski, A. York

AUSTRALIA’S
BIODIVERSITY –
RESPONSES
TO FIRE
Plants, birds and invertebrates
Biodiversity Technical Paper, No. 1
A.M. Gill, J.C.Z. Woinarski, A. York

Cover photograph creditsGroup of 3 small photos, front cover:• Cockatiel. The Cockatiel is one of a group of highly mobile birds which
track resource-rich areas. These areas fluctuate across broad landscapes inresponse to local rainfall or fire events. Large flocks may congregate onrecently-burnt areas. /Michael Seyfort © Nature Focus
• Fern regeneration post-fire, Clyde Mountain, NSW, 1988. /A. Malcolm Gill• These bull ants (Myrmecia gulosa) are large ants which generally build small
mounds and prefer open areas in which to forage for food. They are foundon frequently burnt sites. Despite their fierce appearance, they feed mainlyon plant products. /Alan York.
Small photo, lower right, front cover:• Fuel reduction burning in dry forest. This burn is towards the “hotter” end
of the desirable range. /Alan YorkLarge photo on spine:• Forest fire, Kapalga, NT, 1990. /Malcolm GillSmall photo, back cover:• Cycad response after fire near Darwin, NT. /Malcolm Gill
ISBN 0 642 21422 0
Published by the Department of the Environment and Heritage
© Commonwealth of Australia, 1999
Information presented in this document may be copied for personal use or pub-lished for educational purposes, provided that any extracts are acknowledged.
The views expressed in this paper are those of the authors and do not necessarily representthe views of the Department, or of the Commonwealth of Australia.
Biodiversity Convention and Strategy SectionDepartment of the Environment and HeritageGPO Box 636 CANBERRA ACT 2601General enquiries, telephone 1800 803772
Design: Design One Solutions, CanberraPrinting: Goanna Print, CanberraPrinted in Australia on recycled Australian paper

Biodiversity Technical Paper, No. 1
A. Malcolm GillCSIRO Division of Plant Industry
J.C.Z. WoinarskiParks and Wildlife Commission of the Northern Territory
Alan YorkState Forests of New South Wales
AUSTRALIA’SBIODIVERSITY –RESPONSES TO FIREPlants, birds and invertebrates

2

This is the first of the new Biodiversity TechnicalSeries to be published by the CommonwealthDepartment of the Environment and Heritage.The series has been initiated to collate and makeavailable information on the conservation andsustainable use of Australia’s biodiversity to allthose involved or interested in biodiversity inAustralian environments.
The papers in this publication have beenwritten by scientists with expertise in fire and itseffect on Australia’s biodiversity. The papers willadd considerably to the continuing debate on firein Australia. They will increase access toinformation on how major elements of Australia’sbiodiversity respond to fire and will be ofsignificance to land management agencies, landmanagers and policy makers at all levels.
The papers have been prepared for theBiodiversity Conservation Branch of EnvironmentAustralia, Department of the Environment andHeritage, Canberra by:
• Malcolm Gill, CSIRO, Canberra, whoaddresses the effects of fire events on plantbiodiversity;
• John Woinarski, Parks and WildlifeCommission, Northern Territory, who hasprepared a review of the literatureconcerning the impacts of fire on Australianbirds and an annotated bibliography whichcollates references to fire and Australianbirds; and
• Alan York, then of NSW State Forests, whoassesses whether frequent hazard reductionburning is a sustainable long-termmanagement practice with regard to theconservation of terrestrial invertebratebiodiversity. Alan’s work was made possible bythe foresight of NSW State Forest in initiatinga long-term project in which data was collectedon the same sites for some 20 years.The papers were originally prepared for
Environment Australia in 1996.
3
Introduction to Biodiversity Technical Paper No. 1

4

Biodiversity and bushfires: an Australia-wide perspective on plant-species changes after a fire event 9A. Malcolm GillCentre for Plant Biodiversity Research, CSIRO Division of Plant Industry, ACT
Summary 11
1. Preamble 13
2. Measurement of Biodiversity for Studies of ‘Disturbance’ Effects 152.1 What is Biodiversity? 15
2.2 Measures of biodiversity 15
2.3 Functional groups and measures of plant-species diversity 18
2.4 Declared species identities and diversity measures 20
2.5 Discussion 20
3. Models of Changes of Plant Diversity after Fire 233.1 Species richness (S-t models) 23
3.2 Species-area relationships (S-A models) 24
3.3 Rank-abundance and rank-cover relationships (i-mi and i-mc models) 25
3.4 Rank trajectories (i-t models) 25
4. Field Methods and Observed Changes in Plant Diversity after Fire 294.1 Before-and-after fire comparisons 29
4.2 Comparing adjacent burnt and unburnt stands 29
4.3 Chronological ordination using sites with different ages since fire 34
4.4 Plots, in which changes are observed in situ 35
4.5 Inferential methods 36
4.6 Discussion 37
5. Interpretation of data 395.1 Patterns of richness after fire: invasion-caused changes in composition 39
5.2 Patterns of richness after fire: simplification, no invasion 40
5.3 Distribution of patterns of richness change 41
5.4 A plant-community dynamics viewpoint 42
6. Fire Management of Plant Diversity 43
7. Research on Shifts in Plant Diversity after Fire 45
8. Conclusions 47
9. Acknowledgements 48
10. References cited 49
5
CONTENTS

Fire and Australian birds: a review 55J. C. Z. WoinarskiParks and Wildlife Commission of the Northern Territory
Summary 57
1. Introduction 59
2. History of Fire Regimes 61
3. Fire Regimes and their Impacts on Birds: A Review Across Main Environments 63
3.1 Coastal Heaths and Thickets 63
3.2 Mallee 67
3.3 Temperate Eucalypt Open Forests 69
3.4 Temperate Woodlands 73
3.5 Tropical Eucalypt Open Forests and Savanna Woodlands 74
3.6 Tussock Grasslands 76
3.7 Hummock Grasslands 77
3.8 Acacia Shrublands and Woodlands 77
3.9 Other Habitats 78
3.10 Special Case: Islands 78
4. Research Adequacy Methods and Priorities 79
5. Conservation Overview 83
6. References 85
7. Tables 103Table 1. Scientific Names of Birds mentioned in Text 103
Table 2. Summary Table of Incidence of Subects in Fire-Bird References 106
Table 3. Threatened Bird Taxa for which Inappropriate Fire Regime has been listed as a Threatening Process 108
Fire and Australian birds: an annotated bibliography 113J. C. Z. WoinarskiParks and Wildlife Commission of the Northern Territory
1. Introduction 115
2. Acknowledgements 116
3. References 117
6

7
Long-term effects of repeated prescribed burning on forest invertebrates: managment implications for the conservation of biodiversity 181Alan YorkCentre for Biodiversity and Bioresources, School of Biological Sciences, Macquarie University
Acknowledgements 182
Executive Summary 183
1. Introduction 187
2. Methodology 189
2.1 Study Area 189
2.2 Experimental Design 189
2.3 Measurement of Environmental Parameters 192
2.3.1 Understorey Vegetation Structure 192
2.3.2 The Litter Environment 192
2.3.3 Sticks and Logs 192
2.3.4 Insolation 192
2.3.5 The Soil Environment 192
2.4 Terrestrial Invertebrate Communities 193
2.5 Analytical Procedures 193
2.5.1 Treatment, Exposure and Position Effects 193
2.5.2 Inter-relationships Between Environmental Variables 194
2.5.3 Terrestrial Invertebrate Communities 194
3. Results 199
3.1 Environmental Parameters 199
3.1.1 Understorey Vegetation Structure 199
3.1.2 Litter Biomass 202
3.1.3 Sticks & Logs 203
3.1.4 Insolation 204
3.1.5 The Soil Environment 204
3.2 Inter-relationships Between Environmental Variables 204
3.3 Terrestrial Invertebrate Communities 207
3.3.1 Ordinal Diversity 207
3.3.2 Invertebrate Abundance 207
3.3.3 Invertebrate Species Richness 213
3.3.4 Community Composition 217
3.3.5 Community Structure 234
3.3.6 Biodiversity Indicators 240

4. Discussion 243
4.1 Habitat Structure 243
4.2 Terrestrial Invertebrate Communities 245
4.2.1 Invertebrate Abundance 245
4.2.2 Invertebrate Species Richness 246
4.2.3 Community Composition 248
4.2.4 Community Structure 251
4.2.5 Biodiversity Indicators 253
5. Conclusions 257
References 261

A. Malcolm Gill
Centre for Plant Biodiversity Research, CSIRO Division of Plant Industry, GPO Box 1600,
Canberra, ACT 2601, Australia
BIODIVERSITY AND BUSHFIRES:An Australia-wide Perspective on
Plant-species Changes after a Fire Event


Fires can have major effects on plant-speciesdiversity. These effects occur through variationsin fire regimes but are most obvious in thechanges that take place after a fire event, the topicof this review. Attention here is biased towardvascular plant species, the source of most data.Trends observed in these data may differ fromthose observed for non-vascular plants, vertebrateanimals or other life forms.
How we measure ‘diversity’ is important tothe way we perceive the effects of fires. Variablesthat contribute to measures of diversity includethe number of species present in an area, the areaitself, and the numbers of individual plants perspecies. In some cases, plant cover, height orbiomass for each species may be more usefulmeasures than numbers of plants that are oftenvery difficult to determine.
Measurements of diversity can be expressedin the form of relationships - like the numbers ofspecies in areas of increasing size - or as indices.The search for a common theoretical foundationfor indices and relationships has beenunsuccessful. In Australia, the number of species,or ‘species richness’, found in a certain fixed areahas been the most common and simplest measureused. If data on the numbers of plants, or cover,per species is available, the species can be rankedaccording to the strength of these measures andthe relationships between them expressed as rank-abundance or rank-cover curves; the use of suchcurves should be explored further as a way todepict the changes in plant communities that takeplace after fire. Species’ rank through time can beplotted to indicate shifts in relative performance.
Most indices and relationships do notrequire the explicit identity of the species presentbut this is fundamentally important for theconservation of biodiversity. Species attainindividuality when ‘similarity’ indices are used.For any one site, however, the listing of species(with a measure of performance in terms ofnumbers, cover, height or biomass) remains thebasic data set.
Interpretation of relationships and indicesrecorded at various times after fire can beenhanced by the stratification of species accordingto their responses to a single fire of a specified‘biological’ intensity (all foliage killed) at a
specified life stage (mature) through ‘functionalgroups’. Major ‘functional groups’ are ‘sprouters’,‘seeders’, and ‘ephemerals’. Groups of specieswhich can reproduce without fire are the ‘tolerant’or ‘requiring’ species. Using subdivisions of thesegroups can assist our understanding even further.
Models allow one to explore a wider range ofvariability than is currently known. Thus, whilespecies-area, rank-performance and species-richness may all be expected to change with timeafter fire, the way they do so will be affected bythe functional groups present and theircontributions to the total species composition.
Data on the changes of species numbers andother variables with time since fire come from arange of vegetation types in Australia. Mostinformation has come from areas burnt atdifferent times in the past but examined at thesame time; this method assumes that the pre-firespecies’ compositions, fire characteristics andseasonalities of fires were equitable across sites.
Species richness can fall, rise or remainmore-or-less constant with time after the initialpost-fire recovery phase. Declining richness iscommon in the arid and semi-arid regions. More-or-less constant species richness with time afterfire is a feature of open-forests. Increasing speciesrichness some time after fire is associated with theinvasion of eucalypt forests by species-richrainforests. Because rainforest richness increasesfrom cold to warm, and from drier to wetterenvironments, however, richness can increase ordecrease as encroachment occurs.
A post-fire diversity sequence from lower tohigher diversity as species-rich rainforestsencroach upon open-forests involves a change intotal community composition. This situationcontrasts with that in ephemeral-strong sequenceswhere fire events strongly increase apparent plantdiversity soon after the disturbance while changesin total diversity (apparent plus that hidden in thesoil in the form of seeds and other propagules)may be subdued or absent; changes such as thisrepresent a simplification of an initialcomposition.
We have only a rudimentary knowledge oftime sequences of plant diversity with time afterfire. Not all communities have been sampled. Wedo not know to what extent fire types, seasons of
11
Biodiveristy and bushfires
SUMMARY

fire occurrence and post-fire conditions affectsequences. Is there a gradient of patterns of timesequences of species richness from arid to mesicenvironments as postulated here? Is anygeographical gradient influenced by soil type?
Managers set fires under prescribedconditions for a variety of reasons, the mostcommon in Parks, State Forests and Reservesbeing to maintain biodiversity and to protect livesand property. To achieve goals managers need tomanage fire regimes. Management burning can berelated to times since fire or previous fire-intervaldistribution but the way in which this is donecould have important consequences to thedistribution of diversity in a landscape.
A major gap in the management ofbiodiversity by fire would be filled by the adoptionof a practical, effective, monitoring system linkedwith an effective response system. However, theover-arching research challenge that remains is todefine ecologically appropriate fire regimes fordifferent ecosystems in different ecological,economic and geographical contexts whileidentifying the need for particular shapes, sizesand spatial arrangements of burned areas.
12
Australia’s Biodiveristy - Responses to Fire

Fires occur over the majority of the Australianlandscape and in most vegetation types - includingrainforest - but in a wide variety of regimes (Gillet al. 1981). The ‘regime’ may include: theintervals between fires; whether the fires burnpeat or only the fuels above the soil surface; theintensities of the fires; and, the seasons duringwhich the fires occur (Gill 1975, 1981). After anyindividual fire, the numbers of species presentmay remain the same as the years pass but theymay also decrease substantially or increase. Howthe numbers of plant species, and other measuresof diversity, change with time since fire is thesubject of this review.
The Australian landscape is not pristine.Rapid change has taken place since Europeansettlement. There has been an imposition of aplethora of new land uses across the continent.With the new land uses has come a wide variety ofexotic species of plants and animals. Many of theseintroduced species have become naturalized.
Exotic species are a component ofbiodiversity just as native species are. However, ina conservation context, the elimination of exoticspecies while maintaining all native species is agoal. Among vascular plant species there are now1,952 officially recognized naturalized species inAustralia among a total of 17,590 species (Hnatiuk1990). The number of exotic species in Australia isincreasing by naturalization while the number ofknown native species is increasing as the result oftaxonomic research.
Among the many plant species that may befound at any particular location, only a few -sometimes only one - contribute significantly tothe fuel available to carry a fire but the fire mayaffect all species present. The impact of any onefire depends on its characteristics (Gill 1995).Usually fire intensity has been the firecharacteristic to measure in ‘surface fires’ (burningabove the ground surface) in relation toimmediate impact while the extent of combustionmay be used as a measure of the severity of‘ground fires’ (burning peat or other materialconsidered as being part of the ‘ground’). In thisdiscourse, the immediate impact of a fire willrarely be an issue as it is the events proceedingfrom the time of fire that are the focus. Initial
conditions, of course, may be affected markedlyby fire type and severity but these are rarelyrecorded in studies of the changes in speciescomposition that may take place after fire.
It is perhaps obvious that a fire has an effectthat is independent of its ignition source -whether this be human or natural. No distinctionis made here as to the ignition source of the firethat initiated a particular sequence of change.Indeed, no such distinction is made in theliterature usually. However, there is sometimessensitivity among some members of society as tothe effects of fires ignited for managementpurposes – prescribed fires – compared with thoseignited by lightning.
There has been no attempt before now todraw together the many data that are available onthe changes in species’ complements with timeafter fire and attempt to synthesize them. Whathave we observed? What can we learn from acollective view of these studies? Where and whenis diversity of plants markedly affected by fires?Can we draw conclusions for management fromthese data? What should be done next?
Most botanical studies of changes with timesince fire in Australia have concerned changes innumbers of species or changes in populations of asingle species only. While the emphasis here is ontemporal changes, it is being realized increasinglythat the study of temporal changes cannot bedivorced from a consideration of spatial changes.Spatial matters are considered here, albeit briefly,at several scales.
In this study the aims were to:(i) assess measures of biodiversity in relation to
time since fire;(ii) collate all the relevant Australian literature
on changes in plant species composition withtime since fire;
(iii) identify species’, environmental andvegetation characteristics that might helpexplain the changes observed in vascular-plant diversity; and,
(iv) seek national patterns in diversity changes.
13
1. PREAMBLE


The way in which biodiversity is defined andmeasured affects the way that changes after a fireare documented and how the results areinterpreted. In this chapter a simple definition ofbiodiversity is given in the first section and this isfollowed by a technical section describing ways inwhich biodiversity may be measured. In a furthersection, a way in which the species contributing tothe ‘biodiversity’ may be assigned to ‘functionalgroups’ on the basis of their characteristics isdescribed. Finally, the importance of the explicitidentity of species is noted.
2.1 WHAT IS BIODIVERSITY?
‘Biodiversity’ is the variety of life. In commonusage ‘biodiversity’ is often taken to be the varietyof vertebrate animals and vascular plants but inreality it also includes microbes and fungi, insectsand algae, molluscs and bryophytes. In the studiesbeing reviewed the focus is on vascular plants.
Even with vascular plants what is observedand recorded is what the observer can see directly- ‘apparent biodiversity’. There can be a wholecommunity of species that remains hidden in thesoil until appropriate conditions occur - such as anextraordinary rainfall or a fire for example - torelease them to become visible as above-groundplants. Surveys of land for rare species or unusualflora should involve, therefore, an appreciation ofthe effects of rainfalls and fires on ‘hiddenbiodiversity’.
2.2 MEASURES OF BIODIVERSITY
An observer standing in a plant community maynotice that as the radius of his or her gazeincreases so too does the number of plant species.The relationship between the number of speciesand the area of observation is “one of the moststudied relationships in all of ecology” (Palmerand White 1994). Such relationships may be usedas the base from which some of the manymeasures of diversity can be derived (i.e.Kobayashi and Kimura 1994).
The species-area relationship was formalizedfirst by Arrhenius (1921):
S = cAz (1)
or,
log S = log c + z log A (2)
where S is the number of species, A is the areasampled and c and z are fitted parameters. Datafor a set of islands undergoing primary succession(Rydin and Borgegard 1988) give us examples ofthe values for c and z. Numbers of plant specieswere recorded 5 times between 1886 and 1985(Rydin and Borgegard 1988, Loehle 1990). Fittedcurves to the data gave values of c from 6.3 to10.8, z from 0.16 to 0.30 and r2 from 0.4 to 0.81(Rydin and Borgegard 1988, Loehle 1990).
Whittaker (1965) distinguished three formsof diversity: “α-diversity” was the “diversitywithin individual communities”; “β-diversity”represented the “relative extent(s) ofdifferentiation of communities along topographicgradients”; and, “γ-diversity” was due to“diversities of vegetation patterns, resulting fromboth α and β diversities” (Whittaker 1965). Anysuggestion that c and z reflect α and β diversitieshas been dismissed by Connor and McCoy (1979).
There has not been universal satisfactionwith the Arrhenius (1921) equation (1) above.Gleason (1922), expanding the area of species-areadata from Arrhenius’ 300 dm2 to 240 m2,suggested:
S = c + z log A (3)
Rydin and Borgegard (1988) found thatcurves fitted to equation (3) gave values of cbetween -14 and -61, values of z between 13.2 and39.3 and values of r2 from 0.44 to 0.85 for thisform of equation.
The area considered by various authors wasfurther expanded by Preston (1960). His“Arrhenius plots” (equation (2)) for nearctic andneotropical birds were sigmoidal in shape withthree zones where the graph was more-or-lesslinear viz., areas up to about 100 acres (40 ha), areasfrom 100 acres to about 108.5 acres (ca. 1.3 millionkm2), and areas greater than 108.5 acres.
Gitay et al. . (1991) found that neither theArrhenius curve (equation (1)) nor the Gleason
15
2. MEASUREMENT OF BIODIVERSITY FOR STUDIES OF DISTURBANCE EFFECTS

curve (equation (3)) gave good fits to their NewZealand data. They suggested a three-parameterequation rather than the two parameter forms ofprevious authors:
Sx = c + z log A (4)
where x is the additional parameter. They calledthis model the “general root model” which hadconsiderable success in extrapolation from thesmall areas of their plots to the whole of NewZealand with an area of about 269, 000 km2.
Species-area curves are usually calculatedfrom data collected from a series of independentareas of varying size (such as the islands reportedby Rydin and Borgegard 1988) or from a series ofquadrats of increasing size, each larger oneincorporating all smaller ones (i.e. ‘nested’quadrats in which the areas are not independent).Both of these methods have statistical flaws. Thesize of quadrat or sample area may have an affectso using a number of separate quadrats of equalsize (independent samples) seems pertinent to anysampling strategy (Palmer and White 1994).Palmer and White (1994) found that thegeographic distances separating quadrats (see alsoGleason 1922) and the number of samples takenwere important.
So far we have considered only the mostbasic of diversity indexes, the count of speciespresent in an area - ‘species richness’ - and itsdependence on area. If species richness was thesame in two equal areas but the numbers of plantspresent were vastly different, would comparisonsof diversity between the two be valid? Knight(1975) examined species-rich tropical forest wherethe number of tree species in younger and olderforests respectively were virtually the same on anarea basis - 58 and 57 per 1000 m2 - but quitedifferent on a per-tree basis - 115 and 151 speciesper 500 trees. On these sorts of grounds, manyauthors have considered that numbers of plantsshould be explicit in any diversity measure.
Preston (1962 b) proposed that the chancesof extinction were inversely proportional topopulation size and that population size wasdirectly proportional to area so that the chances ofextinction would be greater on smaller islandsthan larger ones. Thus, from this reasoning alone,smaller areas would have fewer species than largerareas. If the areas of concern were sample areas onthe mainland, rather than islands, the chances ofextinction may be expected to be less because asmall population there may be replenished from
an adjacent larger one. Replenishment of an islandpopulation would have to take place from arelatively distant source over an inhospitablebarrier. If so, by this mechanism alone, thenumbers of species occurring in a sample may beexpected to be larger than those on an island ofthe same size - as is the case (Preston 1962 a, b).Such observations formed the basis of islandbiogeographic theory (MacArthur and Wilson1963, 1967) which has had such a major role inthe debates over selection of reserves forconservation of biodiversity.
Measures of biodiversity which includenumbers of plants could be derived from species-area curves if the numbers of plants per area wereknown. At any one site, it may be expected thatthe number of plants present would beproportional to area (e.g. Preston 1962 b,Kobayashi and Kimura 1994):
N = d A (5)
where N is the number of plants in the areaand d is a constant. (Note that d is site and stand-age specific.) Using this relationship, equations (1)to (4) could be modified to reflect numbers ofplants rather than areas of land. For example,equation 1 becomes:
S = cd-z Nz (6)
or,
S = c' Nz' (7)
where c' equals cd-z
Alternatively:
log S = log c' + z log N (8)
where c' is a constant.
At Myall Lakes National Park in NSW, Foxand Fox (1986) found this relationship to holdamong understorey plants in two areas withdifferent fire histories. The value of z was sitedependent.
Menhinick (1964) tried logS/logN as ameasure of diversity of insects. This is the same asz in equation (8) when log c' equals 0. The valuesobtained ideally should be independent of samplesize (N). Menhinick (1964) found that S /√N wasthe most stable of the indexes (in relation tosample size) he tried across a range of samplesizes. Kobayashi and Kimura (1994) compared six
16
Australia’s Biodiveristy - Responses to Fire

diversity indexes and found that the most stableindex was a new one, δ :
δ = √S/ log N (9)
which could be regarded as a simplification ofequation (3) modified by equation (5). Given xequal to 0.5, equation (4) becomes:
S0.5 = c + z log A (10)
If substitution for A is made (equation (5))then:
S0.5 = c + z log (N/d) (11)
If c equals 0 and d equals 1 then:
z = S0.5/log N (12)
or, z (equation (12)) equals δ (equation (9)).
In the data of Gitay et al. (1991) the best-fitexponent of S for vascular plants in New Zealandwas 0.46 while c was 4.5. Thus the assumptionneeded for a ratio to be derived from a species-area curve as the measure of diversity (equation(12)) viz., that c be zero, was not met by this data.
Another variant on equation (12) is givenwhen x of equation (4) equals 1, i.e. z becomesS/log N. McNaughton (1993) used this index inhis studies of the effects of grazing on plantcommunities while Menhinick (1964) consideredit (and rejected it) in relation to the depiction ofdiversity in insect catches.
If numbers of plants per species in an areafitted a particular statistical distribution, such as alog-normal distribution (e.g. Preston 1962 a, b),then the abundances of individuals in each species(mi) could be calculated from equations such asequation (7) and ranked (by rank i) such that a‘rank-abundance’ curve (i-mi), could be drawn(Kobayashi and Kimura 1994) for a site at aparticular time. Such a curve would not predictwhich species would be of which rank, of course,but would indicate the shape of the curve.
Rank-abundance curves indicate the numberof species in the sample (they are truncatedcurves, the last entry indicating the number ofspecies present), the numbers of individuals perspecies and the relative numerical dominance ofthe top-ranking species. If the rank-one specieshas 1000 individuals, its numerical dominanceover the next may be assessed according towhether it has 1000, 100 or only 10 individualspresent for example. Whittaker (1965) displays a
number of theoretical models of this type ofrelationship.
In a plant context, numerical dominance maybe of little significance. Whittaker (1965) defineda dominant plant as one which, if removed, allowsa strong positive response in one or moreremaining species. “In a given community one ora few species, the dominants, overshadow allothers in their mass and biological activity andmay strongly affect conditions of environment forother species” (Whittaker 1965). Dominance alsoaffects plant species richness (Specht and Specht1989). Thus, biomass, cover, height or some other“importance value” (Whittaker 1965) will often bemore appropriate in expressing functionalrelationships than number of individuals. Even so,that a species ranks first does not necessarilyexpress its functional dominance. At very lowcover values soon after fire, for example, thespecies with the highest cover, biomass ornumerical dominance does not necessarily exertany functional dominance at all. The only species-diversity relationship that allows for dominance ofany nature to be expressed is one involvingspecies-rank.
There seems to be no theory for predictingthe frequency distributions of biomass or covervalues among species. Such a theory for biomassseems unlikely in a general sense but cover-valuedistributions (or, more specifically, foliage-coverdistributions) among species and plant sizes maybe worth exploring because of the importance ofleaf-area index to ecosystem function and itsdependence on site conditions. Whittaker (1965)used net annual production as his measure offunctional dominance.
Examination of the changes in biodiversitywith time since fire have mostly involved thesimplest possible diversity index - the number ofspecies in a particular area. Species-area curvesexpress relationship at a particular time. Whilespecies numbers present in a particular area, orspecies-area curves, are valuable in being able todemonstrate some of the changes that may takeplace after fire, an increase in the number ofinputs would provide further insight into the waysin which the ecosystem may be changing.
Changes in numbers of individuals,important to the expression of demographicchange, can be expressed in rank-abundancecurves. Any numerical dominance effects onspecies numbers can be shown in the same curves.In many plant communities, rank-cover curves
17
Biodiveristy and bushfires

(i-ci) will be more appropriate than rank-abundance curves. ‘Cover’ lacks, of course, thegenetic and demographic implications of plantnumber. Lamont et al. (1977) in their studies ofthe plant diversity of a Western Australianheathland used the “importance value index”(Curtis and McIntosh 1950) which included thefrequency of occurrence of species amongquadrats, their cover and the numbers ofindividuals present.
No single attribute seems ideal for the studyof diversity change with time since fire. The useof constant plant numbers as the basis of an indexeliminates the expression of importantdemographic change. The use of constant areawhen numbers of plants are changing throughorders of magnitude may bias estimates of relativespecies numbers present. The use of total cover,height or biomass, only, will exclude theimportance of dominance effects of individualspecies being recognized. Using a variety ofrelationships will help demonstrate the dynamicsof change in plant communities with time afterfire. Explicitly recognizing the species involved isa further step in appreciating the changes that aretaking place while classifying the species into‘functional groups’ (see below) may allow a moregeneral understanding.
It is important that the changes in speciesrichness detected in the field make biological aswell as statistical sense. Biological sense can beginto be made only in the light of knowledge ofspecies identities and functional groups.
2.3 FUNCTIONAL GROUPS ANDMEASURES OF PLANT-SPECIESDIVERSITY
There are many ways of classifying species intogroups that behave similarly. A commonclassification is into life forms. Taxonomicgroupings at a gross level e.g. cryptogams andvascular plants can be instructive. In the context ofthis contribution, however, there are threecommonly recognized functional groups (e.g.Noble and Slatyer 1980, 1981; Gill 1981, 1993):(i) ‘sprouters’ (species which have mature plants
that resprout after a standard fire intensity -100% leaf scorch) which can be divided intotwo convenient groups - those with seedlingreproduction at the time of fire, ‘sprouterswith seedlings’ or SPRwSD, and thoseusually with little or no seed regeneration,‘sprouters without seedlings’ or SPRoSD;
(ii) ‘seeders’ (species which have mature plantsthat die when exposed to the standard fireintensity), or CDR; and,
(iii) ‘ephemerals’ (species which do not have live‘plants’ exposed to fires but have propagulesthat develop into mature plants after firesonly to senesce and die before the next one)or EPH.Subdivision of seeders’ into those with or
without soil seed storage can be important tounderstanding population dynamics.
To examine the sensitivities of variousmeasures of diversity, the ways in which thesemeasures might change after a fire is outlined forthe different functional groups of plants. Plantnumbers and cover, rather than or biomass orheight, is used for convenience.
Sprouters, by definition, have a resproutingresponse from mature plants after fire. Apercentage, usually small, may die at the time offire, the extent of death in the population perhapsdepending on the life stages present and thecharacteristics of the fire. In the example (Table2.1), two cases in which there is no mortality ofprefire plants are considered. In the first case,SPRwSD, all the plants present resprout after thefire but there are more plants of the speciespresent because of substantial seed germination.In the second case, SPRoSD, all the plants presentresprout as in the first case but there is nogermination or other multiplication of thepopulation. Mature plants of the ‘seeders’ all dieat the time of the fire in our example but return inthe form of seedlings. Ephemerals are notapparent at the time of the fire but many specieswith numerous seedlings emerge from soil-storedseed in our example.
The relationships or indexes to beconsidered (Table 2.1) are:(i) S, the number of species in a selected, fixed,
area (i.e. species richness);(ii) S-A, the species-area curve;(iii) S-N, the species richness versus plant
number curve;(iv) δ = √S/ log N(v) i-mi, the rank-abundance curve; and,(vi) i-ci, the rank-cover curve.
Table 2.1 indicates the changes after fire thatmay be expected in the indexes or in mathematicalrelationships for the functional groups concerned.The changes that occur through a fire event maybe expected to reflect the changes that take placeover a longer time period between fires but in thereverse direction.
18
Australia’s Biodiveristy - Responses to Fire

All indexes or relationships shift for theephemerals because these species were not presentbefore the fire (by definition). By way of contrast,no change in any index would occur for sprouterswithout post-fire seedlings. For the two otherfunctional groups no change was expected in S orS-A but could be for δ and would be for i-mi.Only the rank-cover diagrams (i-ci) would recordchanges for all functional groups. Thus, the rank-cover plot seems to be the most appropriate singlerelationship to use. However, any shift in an indexor relationship as a result of fire needs to beconsidered in the light of the functional groupspresent. The same conclusion would follow if theindexes or relationships were to be compared forthe effects of other disturbances.
Fire is a natural variable but is also a majorand often controversial management tool. Twomajor issues in the use of fire are its value in theprotection of life and property and its affects onconservation of species (e.g. Gill and Bradstock1994, 1995). An important role of diversity indexesand relationships, therefore could be to measurethe effects of fires in relation to conservation. InTable 2, the possibility of detection of a localextinction of a species by each index andrelationship is given for each functional group.
Loss in a species of ephemeral would not bedetected because there is no knowledge of whichephemerals were present before the fire. Wherethe number of species can be separated from thenumber of plants, the measure can record theabsence of a species. Where the number of plantsalone is the basis for an open-ended relationship(i.e. S-N and δ) then it is doubtful that the losswould be detected when seedlings arise after thefire. Thus, again, the use of the measure ofdiversity is greatly improved if the nature of thespecies responses is taken into account.
For the purposes of this paper the number ofspecies in a fixed area (i.e. S), the S-A relationshipand the i-mi or i-ci relationships seem to be themost useful to describe the changes taking placeafter fire. When dominance affects are considered,the last two of these relationships become themost appropriate choices. Plant numberrelationships are most useful if all the plants are ofsimilar size but lose their impact as far asdominance is concerned if sizes are varied. Covermeasurements may be essential if the numbers ofplants cannot be measured or if functional-dominance (as opposed to numerical dominance)is to be represented.
19
Biodiveristy and bushfires
Table 2.1. An indication of the likelihood of change in indexes or relationships after fire accordingto plant functional group. SPRwSD refers to a sprouter group of species producing seedlings aswell as resprouts after fire; SPRoSD refers to a sprouter group without seedlings after fire (nomortality); CDR refers to a seeder group; and, EPH refers to species of ephemerals. S is speciesrichness; S-A is the species-area relationship; S-N is the species-plant number relationship; δ = √S/ log N; i-mi is the rank-abundance relationship; and, i-ci is the rank-cover relationship.
Functional group S S-A S-N δ i-mi (number) i-ci(cover)
SPRwSD no change no change change change change change
SPRoSD no change no change no change no change no change change
CDR no change no change change no change? change change
EPH change change change change change change
Table 2.2. The likelihood of detecting an extinction from various indices and relationships whenapplied to particular functional groups of species. Symbols are as for Table 2.1. ‘Extinction’ in thiscontext refers to the loss of the species from the site due to fire occurrence whether or not it is‘hidden’ or ‘apparent’.
Functional group S S-A S-N d i-mi i-ci
SPRwSD yes yes ? ? yes yes
SPRoSD yes yes yes yes yes yes
CDR yes yes ? ? yes yes
EPH no no no no no no

2.4 DECLARED SPECIES IDENTITIESAND DIVERSITY MEASURES
A constant species richness maintained with timeelapsed after fire may hide a species turnoverwhich can only be detected if the identities of thespecies are known and declared. All of the indexesand relationships so far discussed have anindependence from any declared identity ofspecies. Temporal variations in species diversitycan only be properly understood when the taxainvolved are known. Even spatial comparisons aresomewhat hollow unless taxa are identified atsome level - kingdom, family, species.
Methods of comparing species lists havebeen developed over decades. Similarity indexesare based on species lists for two sites or at twotimes. Species common to both lists a, or speciesfound in one, b, or the other list, c, may bedistinguished. The grand total number of species,G, in this example is then (a+b+c). An index ofsimilarity may be a/G or an index of dissimilaritymay be (b+c)/G. When numbers of sites or timesare considered there is a further possibility, theabsence from the two sites or times of a set ofother species. Such species are often designated byd, a term increasing the variety of possibleindexes.
Allan and Baker (1990) considered thespecies’ composition of a series of sites withdifferent times after fire in arid Australia. After aplotting of species richness on a constant areabasis they graphed the dissimilarities (Bray andCurtis 1957) between every pair of sites againstthe differences in times since fire. Dissimilaritybetween sites increased until the differences inages between sites was about 16 years. Thereafterit remained about the same.
One class of changes after fire is that of a setof species which is gradually depleted (byelimination) as time progresses during phase “c”as in Fig. 3.1. Elimination models have beendeveloped in quite another context, that ofevolutionary changes in species numbers onisolated mountaintops. If all mountain tops beganwith the same composition but were depleted dueto the differential reduction in habitat area(‘islands’) then, all species present must be subsetsof a grand list of all species in all areas and wouldrepresent a sequence from relatively rich torelatively poor. A “nested-subset” analysis hasbeen invented to examine such cases (Pattersonand Atmar 1986).
The nested-subset analysis begins byexamining the data for departure from‘nestedness’ (i.e. all shorter species’ lists aresubsets of larger ones) then applying a statisticaltest to assess its statistical significance. Data areexamined for their departure from nestedness by:(i) determining the shortest faunal list (in theircase) in which a particular species occurs; (ii)examining all richer faunas for the same speciesand recording the absences of that particularspecies in the richer faunas; (iii) repeating thesesteps for all species; and, (iv) counting thenumbers of absences in the data set and divide itby the number of sites multiplied by the numbersof species in the richest set. The analysis is animprovement on species-area curves because itrequires species’ identities to be explicit but somestatistical problems remain (see Doak and Mills1994 for example). Nested subset analysis couldbe relevant to analysis of temporal species changesafter fire but, as yet, has not been considered inthis context. It has the valuable property thatnestedness within a time sequence of stands couldbe considered.
2.5 DISCUSSION
This section started with a consideration ofspecies-area curves, a starting point familiar tomost plant ecologists. These S-A curves can beconverted to species-plant number curves andthen, with a knowledge of the frequencydistribution of numbers of plants per species,rank-abundance curves could be derived.Unfortunately, site specific parameters are neededto fit the basic relationships so the models lackuniversality. The reason for the lack of a solidcommon theoretical base seems to be that thenumbers of plants or other organisms per speciesmay conform to a variety of model distributions(Gray 1986). Gitay et al. (1991) found eitherequation (1) or (7) fitted data better for differentgroups of plants (cryptogams versus vascularplants). If the number of species in an area ofconcern was small (e.g. in some arid areas ofAustralia some years after fire), the use of afrequency distribution of numbers of plants perspecies would be limited.
Hubbell (1979) proposed that frequencydistributions of individuals per species (log-normal or geometric) could be expected to changewith scales and commonness of disturbances andthe chances of local extinctions and immigrations.The localized disturbances in rainforests - such as
20
Australia’s Biodiveristy - Responses to Fire

tree fall - could be contrasted with the majordisturbances in higher latitude forests of theNorthern Hemisphere - caused by fires forexample.
Noble (1989) found that ranked biomass andcover values in mallee communities ofsouthwestern NSW conformed to the geometricdistribution soon after fire, a result he attributedto the dominance of Stipa, an ephemeral grass. Ina series of stands of increasing times since fire injarrah (Eucalyptus marginata) forest, WesternAustralia (WA), rank-relative frequency curvesmoved from the near log-normal type to thegeometric type (Bell and Koch 1980). Simplegeneralizations relating the type of curve to timesince fire in a variety of communities seemunlikely at present.
Diversity indices and relationships may bederived and examined without recourse to theidentities of the organisms being known. To make
ecological sense of the relationships revealed it isnecessary to discern ‘functional’ groups of plants,at least, as these may be expected to have differentbehaviours after fire. To make conservation sense,the individual taxa involved need to be known, ifnot finer levels of plant variation.
There are many other indexes (e.g. Lamontet al. 1977) than those considered here and anumber of other approaches to the study ofbiodiversity. Trophic methods (e.g. Pimm 1982)and mechanistic models of functionalrelationships (eg. Noble et al. 1988) may be used.No approach, no index, no relationship catersideally to the measurement of the diversity of lifein its myriad forms and levels of organization.What we can do is tailor our efforts to thedemonstration and understanding of the diversityof nature as it relates to the purposes and scales athand and within the limits imposed by the extentof our data and knowledge.
21
Biodiveristy and bushfires

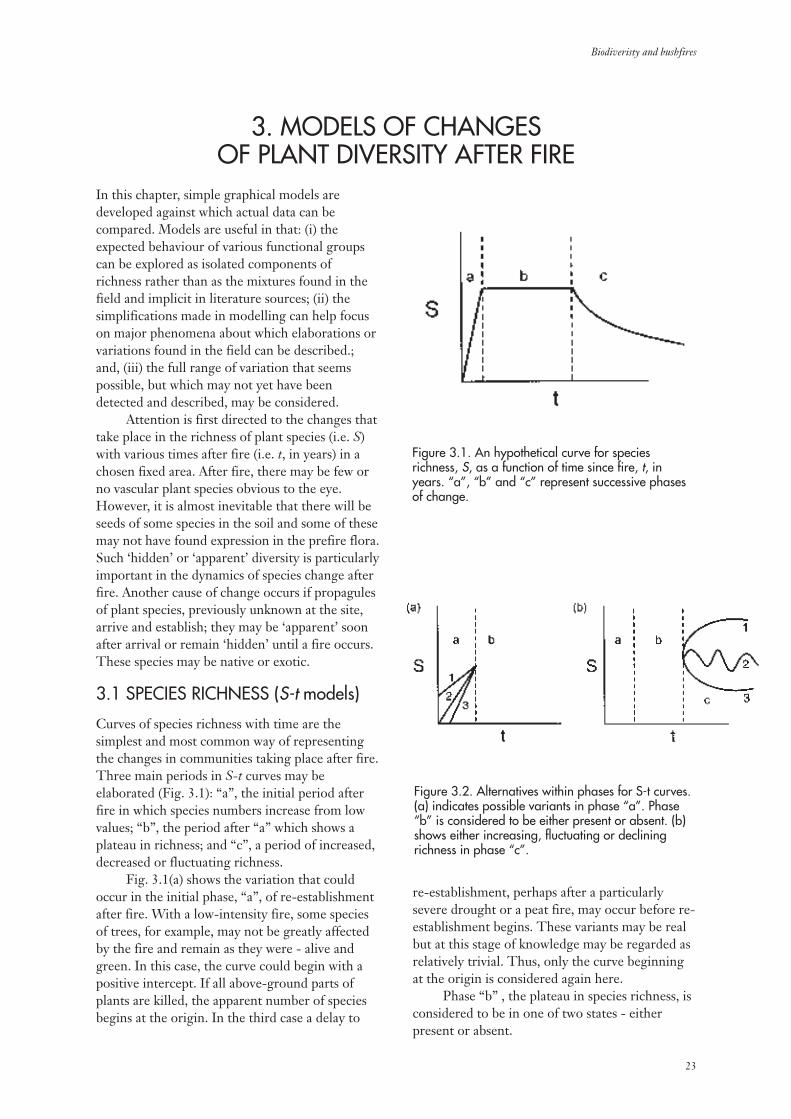
In this chapter, simple graphical models aredeveloped against which actual data can becompared. Models are useful in that: (i) theexpected behaviour of various functional groupscan be explored as isolated components ofrichness rather than as the mixtures found in thefield and implicit in literature sources; (ii) thesimplifications made in modelling can help focuson major phenomena about which elaborations orvariations found in the field can be described.;and, (iii) the full range of variation that seemspossible, but which may not yet have beendetected and described, may be considered.
Attention is first directed to the changes thattake place in the richness of plant species (i.e. S)with various times after fire (i.e. t, in years) in achosen fixed area. After fire, there may be few orno vascular plant species obvious to the eye.However, it is almost inevitable that there will beseeds of some species in the soil and some of thesemay not have found expression in the prefire flora.Such ‘hidden’ or ‘apparent’ diversity is particularlyimportant in the dynamics of species change afterfire. Another cause of change occurs if propagulesof plant species, previously unknown at the site,arrive and establish; they may be ‘apparent’ soonafter arrival or remain ‘hidden’ until a fire occurs.These species may be native or exotic.
3.1 SPECIES RICHNESS (S-t models)
Curves of species richness with time are thesimplest and most common way of representingthe changes in communities taking place after fire.Three main periods in S-t curves may beelaborated (Fig. 3.1): “a”, the initial period afterfire in which species numbers increase from lowvalues; “b”, the period after “a” which shows aplateau in richness; and “c”, a period of increased,decreased or fluctuating richness.
Fig. 3.1(a) shows the variation that couldoccur in the initial phase, “a”, of re-establishmentafter fire. With a low-intensity fire, some speciesof trees, for example, may not be greatly affectedby the fire and remain as they were - alive andgreen. In this case, the curve could begin with apositive intercept. If all above-ground parts ofplants are killed, the apparent number of speciesbegins at the origin. In the third case a delay to
re-establishment, perhaps after a particularlysevere drought or a peat fire, may occur before re-establishment begins. These variants may be realbut at this stage of knowledge may be regarded asrelatively trivial. Thus, only the curve beginningat the origin is considered again here.
Phase “b” , the plateau in species richness, isconsidered to be in one of two states - eitherpresent or absent.
23
Biodiveristy and bushfires
3. MODELS OF CHANGES OF PLANT DIVERSITY AFTER FIRE
Figure 3.1. An hypothetical curve for speciesrichness, S, as a function of time since fire, t, inyears. “a”, “b” and “c” represent successive phasesof change.
Figure 3.2. Alternatives within phases for S-t curves.(a) indicates possible variants in phase “a”. Phase“b” is considered to be either present or absent. (b)shows either increasing, fluctuating or decliningrichness in phase “c”.

Fig. 3.2 illustrates a circumstance in whichthe species richness either increases, fluctuates ordecreases in period “c”. An example of the first ofthese could arise when a species-rich rainforestinvades a eucalypt forest of lesser richness. Adecline in richness during period “c” may occurby an invasion of rainforest but the rainforest inthis case would be one that is species poor.Fluctuating richness may occur in a communitywith many rain-responding ephemeral species.Fox (1990) showed in mallee communities thatcomposition and numbers of species ofephemerals change from year to year.
In Fig. 3.3 the most likely S-t curves areillustrated. Any variation in phase “a” has beenremoved. Phase “b” is present in the firstcircumstance but absent in the second. If “b” waspresent in the second then it would have thegeneral appearance of the first so is not included.In the third example in the Figure, “b” is absentand the richness fluctuates but not necessarily in aregular way; including phase “b” in anothervariant of this would be unlikely biologically. Inthe last two illustrations in Fig. 3.3, “b” is presentand followed by either increasing or decreasingspecies richness. These 5 models therefore involvevarious trends in apparent richness (first 3) orinvolve apparent richness plus that due toprocesses of invasion and depletion of thecommunity (last 2).
In cases where species invade after fire theymust have ‘tolerant’ (T) or ‘requiring’ (R) modesof establishment in that they can tolerate a widerange of establishment circumstances - soon afteror long after fire - or “require some conditionpresent in established communities” (Noble andSlatyer 1980).
Changes in the time-courses of speciesrichness after fire are sometimes linked to the
functional dominance of particular species (e.g.Noble 1989, Specht and Specht 1989). In somearid zone communities, dominance - reflected bycover value - may better be depicted as a functionof cumulative rainfall after fire (Griffin 1992).Thus, rainfall could be a better independentvariable against which to depict changes inrichness after fire. Using rainfall in such cases hasan advantage over the use of calendar years in thatit reflects the influence of a driving variable ratherthan that of a surrogate. Unfortunately, such abasis for the depiction of change does not havenation-wide utility. Many species in manylocations have strongly seasonal rhythms (forflowering in particular) that are a function of thepassage of years rather than related to quantitiesof antecedent rainfall. Driving variables otherthan rainfall are often significant also and arecollectively incorporated into ‘year’.
3.2 SPECIES-AREA RELATIONSHIPS(S-A models)
Above, changes in species richness with time sincefire were depicted for an area of constant size. Inthis section, the area constraint is removed by theconsideration of the changes in species-arearelationships with time after fire. S-A curves arenot uncommon but have not been considered as afunction of time since fire.
Species can enter a fixed area or quadrat anumber of ways. ‘Entry’ can come from sourcesexternal to the community, such as frompropagules of invading species, or from internalsources. In the latter category are species‘appearing’ from the soil soon after fire andspecies that appear in the quadrat as the result ofsampling artefacts. Small plants are less likely tobe sampled in a quadrat than large plants so, as
24
Australia’s Biodiveristy - Responses to Fire
Figure 3.3. The five most significant, but still somewhat hypothetical, trajectories for S-t curves.

plants increase in size after a fire, they may bemore likely to appear in a sampled quadrat. Asmall population of a species is also less likely tobe sampled than a large one so populationsbuilding after a fire may escape sampling earlierbut appear later. These circumstances for a singlefixed quadrat can be overcome if a whole series ofquadrats is used (or, less appropriately statistically,when nested quadrats are used).
Fig. 3.4 represents the way the curves mightappear when a species-poor community (“3”)becomes a species-rich community after fire (“1”)(e.g. Zimmer 1940). With time after fire, then, thecommunity would shift from “1” to “3”.
3.3 RANK-ABUNDANCE AND RANK-COVER RELATIONSHIPS ( i-mi and i-mc models)
At any one time after fire, there is a range ofpossible relationship between the number ofplants of the species present (or their cover,biomass or height) and their rank. Fig. 3.5 depictssome of the range that may be possible. Thecurves shown in Fig. 3.5 are indicative rather thandefinitive, representing a range of possibilities(after Whittaker 1965 and 1972, Hubbell 1979,Bell and Koch 1980, Hughes 1986 and Noble1989). They show increasing richness anddecreasing dominance (numerical or functional)from 1 to 5.
In a species-rich shrubland in WesternAustralia, Lamont et al.’s (1977) plot of“importance values” against rank indicated afunctional dominance of a few species over a
considerable number of other species which thenshowed relatively little difference in theirimportance values, a modified form of curve 3 or4 in Fig. 3.5. Curves from species-rich tropicalrainforests also indicate that the highest rankedspecies are considerably more numerous or‘important’ than all other woody species presenteven though there is a shallow slope to curves ofimportance-value against rank for most of therelationship (Hubbell 1979). Bell and Koch’s(1980) curves for jarrah forest all showed strongdominance for communities at various times afterfire. Even soon after fire, the results of Noble(1989) indicated strong cover dominance of anherbaceous species in a community dominatedstructurally by woody plants when mature viz.,mallee.
Changes in the curves with time after firemay show trends because all components of thecurves may change over time viz., the numbers ofindividuals (or their cover), the numbers ofspecies, and their relationships with each other.
3.4 RANK TRAJECTORIES (i-t models)
If the rank of a species was plotted against timesince fire we would be one step closer toidentifying species performances with time afterfire rather than considering species anonymously.Identifying the positions of any single species inthe curves above (Fig. 3.5) is not possible.
25
Biodiveristy and bushfires
Figure 3.4. Species-area curves for threecommunities representing different stages after fire.In this example, the species-rich curve, “1”, is thatsoon after fire; curve “3” represents a maturecommunity of lower richness; and, “2” is for acommunity at an intermediate stage.
Figure 3.5. Hypothetical rank-abundance or rank-cover curves. Species richness increases, andnumerical or functional ‘dominance’ decreases, fromcurves 1 to 5.

26
Australia’s Biodiveristy - Responses to Fire
Mature spinifex, Rudall River, WA. As the spinifex isin discreet clumps and discontinuous, it will not easilycarry fire and has not been burnt for some time. /A Malcolm Gill
Post fire, Rudall River, WA, showing regeneration ofa variety of plants including ephemerals and aNewcastelia shrub. Immediate post-fire regenerationincludes a flush of annuals which reappears in asecond year at least but is ‘scanty’ in maturecommunities. /A Malcolm Gill
Patchy fire, Rudall River, 1994. Such fires in arid areas result in a mosaic of older and newer growth creating avariety of vegetation age classes, species number and structures. /A Malcolm Gill
Hot fire in spinifex and acacia, Great Sandy Desert, 1986./ James Eldridge

27
Biodiveristy and bushfires
Post-fire, low intensity fire, Bungendore, NSW, 1985.The starting conditions and the nature of the fire canaffect the type of regeneration. /A Malcolm Gill
Nothofagus forest gully, central highlands of Victoria,1996. This area has not been burnt for a longperiod. /A Malcolm Gill
Late dry season fire at Kakadu, 1990, with grassy fuel and pandanus. Fires at this time of year have a highchance of getting away in most areas of Kakadu. Close to wetlands, however, management fires are lit then as itis too wet at other times. /A Malcolm Gill

28
Australia’s Biodiveristy - Responses to Fire
Early post-fire, Nadgee, NSW, 1981, showing weedregeneration. /A Malcolm Gill
Resprouting of Eucalyptus maculata (smooth) andEucalytus globulus (stringy) after the January 1994fire, Mogo State Forest, NSW. /A Malcolm Gill
Post-January 1994 fire, Oxford Falls, NSW. Flannelflowers regenerating. /A Malcolm Gill
Black Mountain, 1984. Litter fuel fire. /A Malcolm Gill
Black Mountain, ACT, 1991. Fire behaviour will affect regeneration. /A Malcolm Gill

The detection of changes in plant diversity afterfire have been made in a number of ways. In thischapter, the observations made have been groupedaccording to the various methods used becauseeach method has its own values and drawbacks.The sites of the studies mentioned are mapped inFig. 4.1 while selected portions of the data areshown in Table 4.1. The Table shows raw datawhich can be misleading in the absence of thetrends of data sequences.
4.1 BEFORE-AND-AFTER FIRECOMPARISONS
Observations made before and after fire provideone of the means of examining changes indiversity. Christensen and Kimber (1975)provided data using this method in south-westernAustralia. Species richness of vascular plantspecies in tall open forests (“wet sclerophyllforests” structurally dominated by eucalypts)showed that up to 3 times the number of specieswere present after fire than before. At the sametime, however, the numbers of species in the‘untreated’, or ‘control’, sites were decliningrapidly. In the drier open forests (“dry sclerophyllforests” - “jarrah forest” in Fig. 4.1), however,numbers showed little change when the ‘unburnt’area was 6 years free from fire but showed a 20%downward change for a 38 year period.
Heaths and heath woodlands insouthwestern Victoria near Anglesea revealed allprefire species within 3 years of being burnt(Wark et al. 1987); the same was true of aheathland near Noosa, southeastern Queensland(Harrold 1979). In the Anglesea case, at least, thespecies richness then declined with elapsed time.
4.2 COMPARING ADJACENT BURNTAND UNBURNT STANDS
Comparing floras in adjacent burnt and ‘unburnt’areas at the same time is perhaps the simplestmethod of recording change with time after fire.Sites often are considered to be identicalphysically and biologically prefire.
A classic case of this method is that ofZimmer (1940) who found that the numbers ofspecies in a northwestern Victorian malleecommunity near Hattah had many more speciesafter the fire than before the fire (Table 4.2). Thedominant mallee plants (i.e. Eucalyptus spp.) andthe main ground cover, of hummock grass (Triodiasp. or ‘spinifex’), had all foliage removed by thefire. Also in north-western Victoria, Cheal et al.(1979) reported 27 instances of greater speciesrichness in burnt cf. unburnt stands of grassland,heath, shrubland, mallee and pine woodland.
In a Western Australian mallee region “620km ESE of Perth” (presumed to be near thesettlement of Salmon Gums) in which there wasnot only mallee vegetation but also woodland andshrubland, richness was greater in all burnt sitesexamined (Van der Moezel and Bell 1984).
In the extensive spinifex-dominatedgrasslands of arid Australia, increases in speciesrichness after fire followed the same trend as thatobserved by Zimmer (1940) in the semi-aridmallee. In northwestern WA (Warralong Station)immediate post-fire regeneration included a flushof annuals which reappeared in a second year atleast but were “scanty” in mature communities(Burbidge 1943). Similar observations were madeby the author and his colleagues P. H. R. Moore,N. D. Burrows and B. Ward, in hummockgrasslands near Rudall River National Park350 km east-south-east of this area.
29
Biodiveristy and bushfires
4. FIELD METHODS AND OBSERVED CHANGES IN PLANT DIVERSITY AFTER FIRE
30
Australia’s Biodiveristy - Responses to Fire
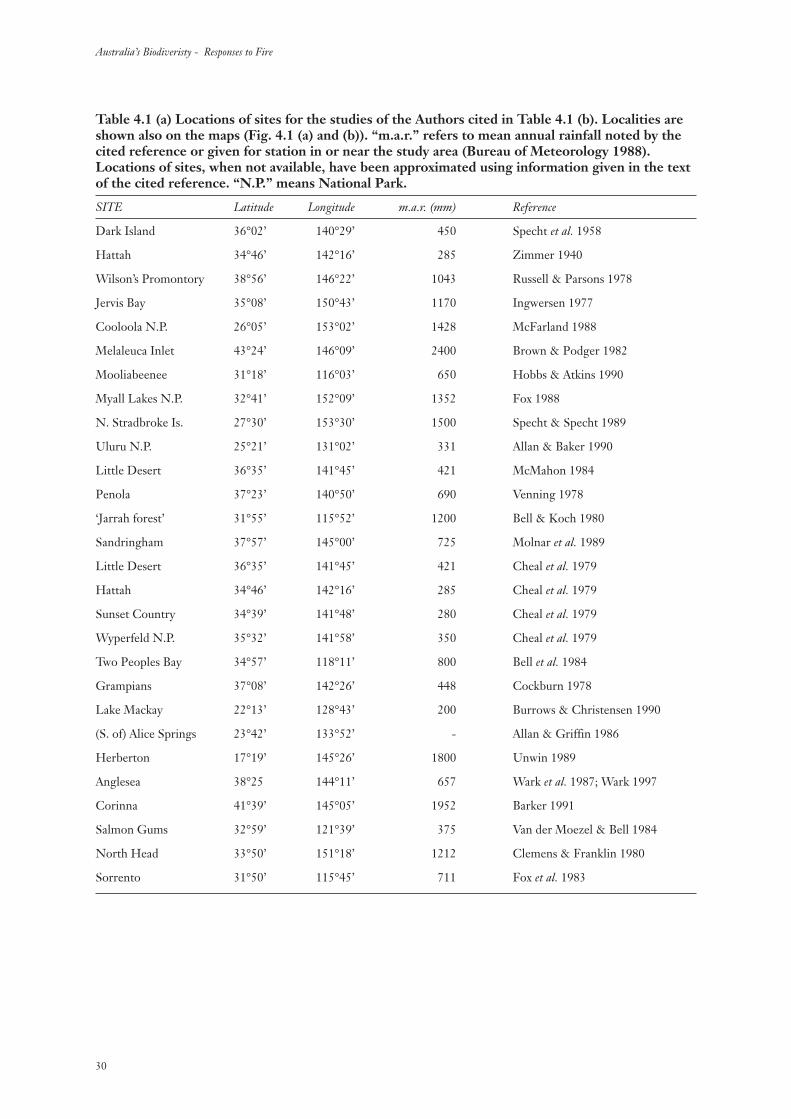
Table 4.1 (a) Locations of sites for the studies of the Authors cited in Table 4.1 (b). Localities areshown also on the maps (Fig. 4.1 (a) and (b)). “m.a.r.” refers to mean annual rainfall noted by thecited reference or given for station in or near the study area (Bureau of Meteorology 1988).Locations of sites, when not available, have been approximated using information given in the textof the cited reference. “N.P.” means National Park.
SITE Latitude Longitude m.a.r. (mm) Reference
Dark Island 36°02’ 140°29’ 450 Specht et al. 1958
Hattah 34°46’ 142°16’ 285 Zimmer 1940
Wilson’s Promontory 38°56’ 146°22’ 1043 Russell & Parsons 1978
Jervis Bay 35°08’ 150°43’ 1170 Ingwersen 1977
Cooloola N.P. 26°05’ 153°02’ 1428 McFarland 1988
Melaleuca Inlet 43°24’ 146°09’ 2400 Brown & Podger 1982
Mooliabeenee 31°18’ 116°03’ 650 Hobbs & Atkins 1990
Myall Lakes N.P. 32°41’ 152°09’ 1352 Fox 1988
N. Stradbroke Is. 27°30’ 153°30’ 1500 Specht & Specht 1989
Uluru N.P. 25°21’ 131°02’ 331 Allan & Baker 1990
Little Desert 36°35’ 141°45’ 421 McMahon 1984
Penola 37°23’ 140°50’ 690 Venning 1978
‘Jarrah forest’ 31°55’ 115°52’ 1200 Bell & Koch 1980
Sandringham 37°57’ 145°00’ 725 Molnar et al. 1989
Little Desert 36°35’ 141°45’ 421 Cheal et al. 1979
Hattah 34°46’ 142°16’ 285 Cheal et al. 1979
Sunset Country 34°39’ 141°48’ 280 Cheal et al. 1979
Wyperfeld N.P. 35°32’ 141°58’ 350 Cheal et al. 1979
Two Peoples Bay 34°57’ 118°11’ 800 Bell et al. 1984
Grampians 37°08’ 142°26’ 448 Cockburn 1978
Lake Mackay 22°13’ 128°43’ 200 Burrows & Christensen 1990
(S. of) Alice Springs 23°42’ 133°52’ - Allan & Griffin 1986
Herberton 17°19’ 145°26’ 1800 Unwin 1989
Anglesea 38°25 144°11’ 657 Wark et al. 1987; Wark 1997
Corinna 41°39’ 145°05’ 1952 Barker 1991
Salmon Gums 32°59’ 121°39’ 375 Van der Moezel & Bell 1984
North Head 33°50’ 151°18’ 1212 Clemens & Franklin 1980
Sorrento 31°50’ 115°45’ 711 Fox et al. 1983
Biodiveristy and bushfires
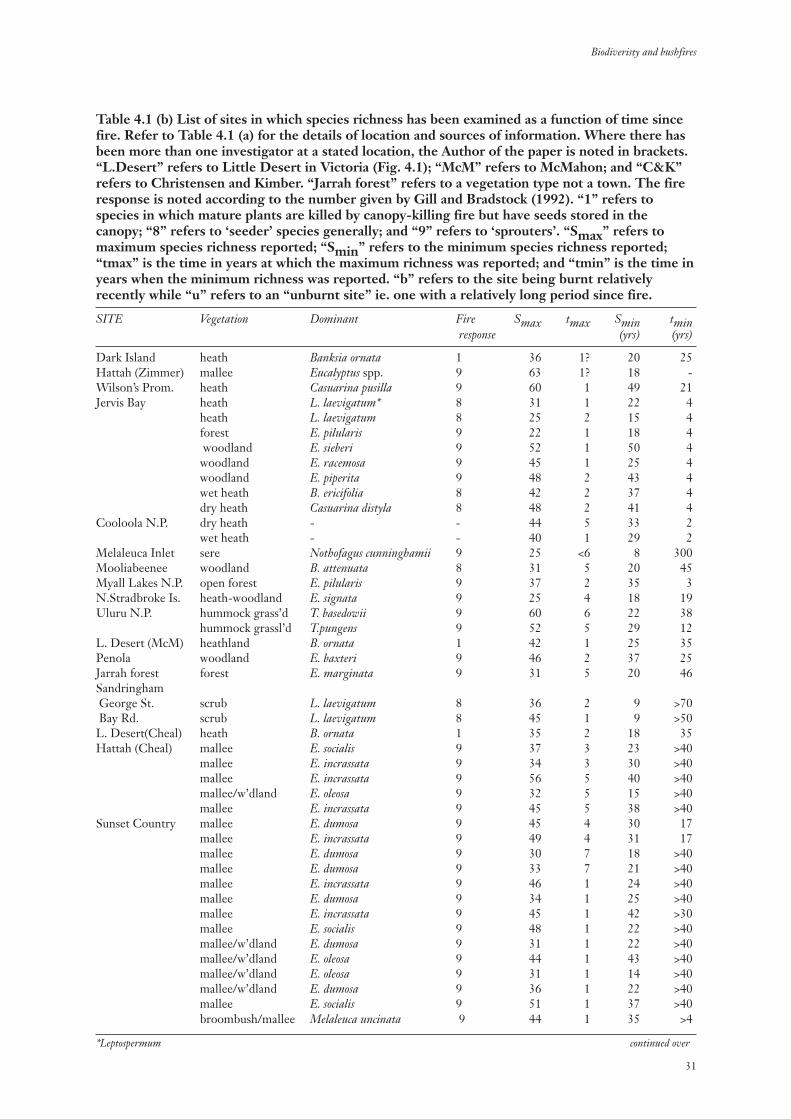
Table 4.1 (b) List of sites in which species richness has been examined as a function of time sincefire. Refer to Table 4.1 (a) for the details of location and sources of information. Where there hasbeen more than one investigator at a stated location, the Author of the paper is noted in brackets.“L.Desert” refers to Little Desert in Victoria (Fig. 4.1); “McM” refers to McMahon; and “C&K”refers to Christensen and Kimber. “Jarrah forest” refers to a vegetation type not a town. The fireresponse is noted according to the number given by Gill and Bradstock (1992). “1” refers tospecies in which mature plants are killed by canopy-killing fire but have seeds stored in thecanopy; “8” refers to ‘seeder’ species generally; and “9” refers to ‘sprouters’. “Smax” refers tomaximum species richness reported; “Smin” refers to the minimum species richness reported;“tmax” is the time in years at which the maximum richness was reported; and “tmin” is the time inyears when the minimum richness was reported. “b” refers to the site being burnt relativelyrecently while “u” refers to an “unburnt site” ie. one with a relatively long period since fire.
SITE Vegetation Dominant Fire Smax tmax Smin tminresponse (yrs) (yrs)
Dark Island heath Banksia ornata 1 36 1? 20 25Hattah (Zimmer) mallee Eucalyptus spp. 9 63 1? 18 -Wilson’s Prom. heath Casuarina pusilla 9 60 1 49 21Jervis Bay heath L. laevigatum* 8 31 1 22 4
heath L. laevigatum 8 25 2 15 4forest E. pilularis 9 22 1 18 4woodland E. sieberi 9 52 1 50 4 woodland E. racemosa 9 45 1 25 4 woodland E. piperita 9 48 2 43 4wet heath B. ericifolia 8 42 2 37 4dry heath Casuarina distyla 8 48 2 41 4
Cooloola N.P. dry heath - - 44 5 33 2wet heath - - 40 1 29 2
Melaleuca Inlet sere Nothofagus cunninghamii 9 25 <6 8 300Mooliabeenee woodland B. attenuata 8 31 5 20 45Myall Lakes N.P. open forest E. pilularis 9 37 2 35 3N.Stradbroke Is. heath-woodland E. signata 9 25 4 18 19Uluru N.P. hummock grass’d T. basedowii 9 60 6 22 38
hummock grassl’d T.pungens 9 52 5 29 12L. Desert (McM) heathland B. ornata 1 42 1 25 35Penola woodland E. baxteri 9 46 2 37 25Jarrah forest forest E. marginata 9 31 5 20 46SandringhamGeorge St. scrub L. laevigatum 8 36 2 9 >70Bay Rd. scrub L. laevigatum 8 45 1 9 >50L. Desert(Cheal) heath B. ornata 1 35 2 18 35Hattah (Cheal) mallee E. socialis 9 37 3 23 >40
mallee E. incrassata 9 34 3 30 >40mallee E. incrassata 9 56 5 40 >40mallee/w’dland E. oleosa 9 32 5 15 >40mallee E. incrassata 9 45 5 38 >40
Sunset Country mallee E. dumosa 9 45 4 30 17mallee E. incrassata 9 49 4 31 17mallee E. dumosa 9 30 7 18 >40mallee E. dumosa 9 33 7 21 >40mallee E. incrassata 9 46 1 24 >40mallee E. dumosa 9 34 1 25 >40mallee E. incrassata 9 45 1 42 >30mallee E. socialis 9 48 1 22 >40mallee/w’dland E. dumosa 9 31 1 22 >40mallee/w’dland E. oleosa 9 44 1 43 >40mallee/w’dland E. oleosa 9 31 1 14 >40mallee/w’dland E. dumosa 9 36 1 22 >40mallee E. socialis 9 51 1 37 >40broombush/mallee Melaleuca uncinata 9 44 1 35 >4
*Leptospermum continued over
31
32
Australia’s Biodiveristy - Responses to Fire
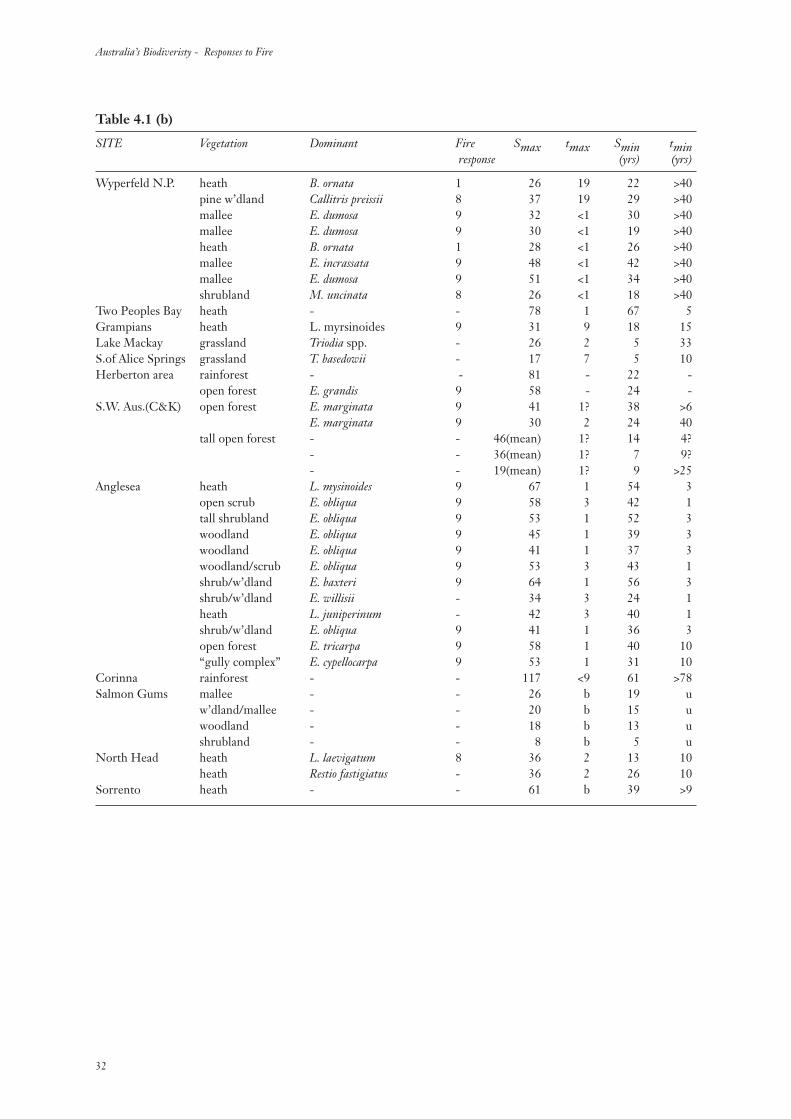
Table 4.1 (b)
SITE Vegetation Dominant Fire Smax tmax Smin tminresponse (yrs) (yrs)
Wyperfeld N.P. heath B. ornata 1 26 19 22 >40pine w’dland Callitris preissii 8 37 19 29 >40mallee E. dumosa 9 32 <1 30 >40mallee E. dumosa 9 30 <1 19 >40heath B. ornata 1 28 <1 26 >40mallee E. incrassata 9 48 <1 42 >40mallee E. dumosa 9 51 <1 34 >40shrubland M. uncinata 8 26 <1 18 >40
Two Peoples Bay heath - - 78 1 67 5Grampians heath L. myrsinoides 9 31 9 18 15Lake Mackay grassland Triodia spp. - 26 2 5 33S.of Alice Springs grassland T. basedowii - 17 7 5 10Herberton area rainforest - - 81 - 22 -
open forest E. grandis 9 58 - 24 -S.W. Aus.(C&K) open forest E. marginata 9 41 1? 38 >6
E. marginata 9 30 2 24 40tall open forest - - 46(mean) 1? 14 4?
- - 36(mean) 1? 7 9?- - 19(mean) 1? 9 >25
Anglesea heath L. mysinoides 9 67 1 54 3open scrub E. obliqua 9 58 3 42 1tall shrubland E. obliqua 9 53 1 52 3woodland E. obliqua 9 45 1 39 3woodland E. obliqua 9 41 1 37 3woodland/scrub E. obliqua 9 53 3 43 1shrub/w’dland E. baxteri 9 64 1 56 3shrub/w’dland E. willisii - 34 3 24 1heath L. juniperinum - 42 3 40 1shrub/w’dland E. obliqua 9 41 1 36 3open forest E. tricarpa 9 58 1 40 10“gully complex” E. cypellocarpa 9 53 1 31 10
Corinna rainforest - - 117 <9 61 >78Salmon Gums mallee - - 26 b 19 u
w’dland/mallee - - 20 b 15 uwoodland - - 18 b 13 ushrubland - - 8 b 5 u
North Head heath L. laevigatum 8 36 2 13 10heath Restio fastigiatus - 36 2 26 10
Sorrento heath - - 61 b 39 >9

33
Biodiveristy and bushfires
Figure 4.1. Locations of sites indicated in Table 4.1 and in the text. (a) Australia; (b) southeastern Australia.

In coastal heathlands just north of Perth,WA, (Sorrento) burning led to a 50% increase inthe number of species recorded (Fox et al. 1983).The burning of scrubland invaded by the nativeshrub Leptospermum laevigatum (a ‘seeder’) duringhistorical times in Melbourne (Sandringham),Victoria (Vic.), “dramatically increased thenumber of native species present” (Molnar et al.1989). A heath dominated by the same species atNorth Head in Sydney, New South Wales(NSW), also revealed more species once burnt(Clemens and Franklin 1980); the same authorsrecorded the same phenomenon for the adjacentRestio-Leucopogon heath.
Burnt temperate rainforest areas innorthwestern Tasmania (near Corinna) showedhigher richness than areas that escaped fire butthere appeared to be an inconsistent effect of fireintensity on richness as well (Barker 1991).
4.3 CHRONOLOGICAL ORDINATIONUSING SITES WITH DIFFERENT AGESAFTER FIRE
An elaboration of the method described in theprevious section occurs when a variety of sites,burnt at different times in the past, are examined.This method has been called “chronologicalcommunity ordination” (Gill 1977). While quickand convenient, it has the disadvantage that it maybe hard to find study sites which vary widely infire history but are uniform in site characteristics.In the best use of the technique, the effects of sitewould be discriminated during statistical analysis.The method implies that there has been no affectof season of fire occurrence or of fire intensity orthat these variables were uniform across all plots.It assumes that all the plots had the same recentfire history (burnt at the same time previously) orthat this feature was unimportant to thecomparisons.
Specht et al. (1958) revealed a decline inspecies richness with time since fire in a Banksiaornata heathland near Keith in South Australia(SA). “Of 36 species recorded after a fire, only 20were found after 25 years” and “Probably only tenof these would persist after 50 years” (Specht et al.1958). In the Little Desert in Victoria Cheal et al.(1979) and McMahon (1984) were able todemonstrate the same trend in similar vegetationtypes. Later Specht and Specht (1989) showedthat species richness was a linear and negativefunction of the foliage cover of the ‘seeder’ species
Banksia ornata. The S-t curves reported by all ofthese authors were similar to that of the seconddiagram in Fig. 3.3 with a short phase “a”. InMcFarland’s (1988) study area in southeasternQueensland (Qld) (Cooloola National Park) therewas a similar short build-up time in apparentspecies richness (1 year) but this was followed byvery little change in richness in sitesrepresentative of the next 10 years. The changesin the first decade were similar to those noted byRussell and Parsons (1978) for a wet heath insouthern Victoria (Wilson’s Promontory) but afterthat period there was a slow decline in richnessfrom near 60 species at age 10 years to 49 speciesat 21 years after fire; the authors noted that thetrend was possibly confounded by site differences.Studies in the heathlands of the Grampians inwestern Victoria suggested a much longer build-up period (perhaps 9 years) followed by the typicaldecline reported by other authors (Cockburn1978).
In the Gibson Desert (in WA near LakeMackay), Burrows and Christensen (1990) foundonly 5 to 7 species in stands more than 20 yearsold but up to 26 species in stands burned 2 yearsbefore examination. Allan and Baker (1990),similarly, recorded a sharp drop in species richnesswith time after fire in arid spinifex grassland butonly after a slow rise that took about 6 years (i.e.phase “a” of Fig. 3.1). A double-humped curvewas published by Allan and Griffin (1986) for aridCentral Australia (south of Alice Springs), thesecond rise in richness being associated with aninvasion by trees and shrubs beginning between 9and 27 years after fire.
In a eucalypt woodland in southeasternSouth Australia (near Penola), Venning’s (1978)data suggested that there was a rapid restorationof species richness to an average of about 43species in the first year after fire followed by aperiod up to year 25 in which there was a slightdecline to an average of about 40 species; such aslight decrease is probably not statisticallysignificant. Presumably there was little or nochange in species composition during this timeeither. In a Banksia woodland north of Perth (atMooliabeenee), WA, species richness rose afterfire to peak at 5 years, with 31 species per 25m2
quadrat, then declined to about 20 species ≥44years after fire (Hobbs and Atkins 1990).
34
Australia’s Biodiveristy - Responses to Fire

A curve for species richness in a “jarrahforest” was constructed in the Darling Range nearPerth by Bell and Koch (1980) using a series offorest sites from different forestry ‘blocks’. There,richness peaked at 30 at 5 years then declined to19 in stands 50 years since last fire. However,species composition varied a great deal betweensites.
That “There is no evidence that fire altersthe floristic composition” of E. obliqua forests inthe Adelaide Hills, SA, region (Wood 1937) maymean that there has been no change detected fromthe range of fire regimes experienced there or thatthere has been no change with time after fire (orboth). Cochrane et al. (1962) leant towards thelatter interpretation when considering pyricsuccession in the same region (near Crafers) inthat within “seven to ten years the sere iscompleted ... the composition of the sclerophyllunderstorey is remarkably similar”. However,there were changes in the dominance of theunderstorey species during this period.
On sandy soils of coastal NSW in MyallLakes National Park, an open forest of E. pilularisshowed a maximum richness 2 years after fire.Richness then declined to 3 years or so whence anincrease occurred again to 16 years, the longesttime after fire in the sequence (Fox 1988). In anearby area, a greater number of species wasrecorded in a younger stand (Fox and Fox 1986).
The longest time sequence examined was inwet southwest Tasmania in the vicinity ofMelaleuca Inlet. There, Brown and Podger (1982)linked changes from sedgeland-heath to woodlandto “mixed forest” and rainforest with time sincefire. After the initial buildup of richness to a peakof 24 species in less than 6 years, the richnessgenerally declined to about 8 species in the ca.300-year sequence. There was a possible outlier inthe data at 19 years which indicated a temporaryincrease in richness.
4.4 PLOTS, IN WHICH CHANGESARE OBSERVED IN SITU
Changes in species richness can be observed eitherin plots being monitored after fire or in thoseintegral to an experiment. The advantage ofexperiments is that there is a known starting pointfrom which changes may be observed. Thedifferences between experimental plots can bedescribed and accounted for in assessing theresults of any change (e.g. Bowman et al. 1988).Effects of seasons of burning may be measured aswell as effects of intensities. Disadvantages may bethat: the range of vegetation types chosen may belimited by the experimenter in order to enable theburning of the plots according to the experimentalplan; fires in plots during critical fire-dangerperiods may not be allowed by authorities; and,experimental fires cannot reflect the full range ofintensities encountered in the forest (Gill 1977,Gill and Moore 1990). There can also beinteractive effects between fires and grazinganimals, native or exotic (Leigh and Holgate1979, Hesp et al. 1983, Leigh et al. 1987, Noble1989). The advantage of plots set up after fire (e.g.Ingwersen 1977, Wark et al. 1987, Wark 1997) isthat there is no question of any apparent changebeing a consequence of site sampling. However, inboth experimental and post-fire plot observation,sampling problems can arise when species growinto plots thereby artificially increasing numbers.
In mallee vegetation in southwestern NSWnear Pooncarie, large numbers of herbaceousspecies, not apparent before the fire, appearedafter it (Noble 1989). Numbers of species wereboosted by post-fire rainfall conditions. A mulgashrubland (dominated by the ‘seeder’ Acaciaaneura) in arid Central Australia (Alice Springsregion) showed increased richness one year afterfire but subsequent declines in the following threeyears (Griffin and Hodgkinson 1986). This trendwas evident in the control as well as the treatedplots but the treated plots showed more species inyears 2 to 4 than the control. A neighbouring“intergrove” community (possibly dominated bytussock grasses) showed parallel trends in richnessto the mulga areas but revealed no differencebetween treatments.
In “sclerophyll woodland” or “heathywoodland” in southeastern Queensland(Stradbroke Island), numbers of species per 10m2
declined from 26 to 18 in a 19-year period afterfire (Specht and Specht 1989 p. 342).
35
Biodiveristy and bushfires

Ingwersen (1977) examined species numbersin plots for 4 years after a large unplanned fire in anumber of heaths, scrubs, woodlands and forestsat Jervis Bay south of Sydney. Peak richness wereattained within 2 years. Numbers of species weremaintained or dwindled in the following two yearsin most cases but the decline in the E.racemosa-E.gummifera woodland was quite marked.‘Control’ areas were unavailable for comparison inIngwersen’s study area.
Richness in a southwestern Australian heathat Two Peoples Bay peaked at 78 species one yearafter fire and then gradually declined to 67 at year5 (Bell et al. 1984).
Near Canberra, Australian Capital Territory(ACT), Purdie (1977) found that the speciescomposition remained the same after burning“dry sclerophyll vegetation” (‘open forest’ ofEucalyptus spp.). This appeared to be the case alsofor eucalypt forest and woodland in the northernNorthern Territory (Munmarlary) where burningtreatments were as frequent as annual inoccurrence (Hoare et al. 1980, Bowman et al.1988). Tolhurst and Oswin (1992) studied theeffects of a range of fire regimes in a Victorianforest (Wombat State Forest) and reported that“No species was gained or lost from anytreatment”.
4.5 INFERENTIAL METHODS
Some changes in richness may take so long thatexperiments seem impracticable and dating ofstands is beyond the reach of photogrammetric orrecorded history. In these sorts of cases inferencemay be used to piece sequences together. Theprime examples are for rainforests originatingfrom eucalypt-forest or eucalypt-woodland sites.The sites in question may lie anywhere on thecoastal and subcoastal areas of eastern andnorthern Australia (as far west as The Kimberley).
In Tasmania, Gilbert (1959) drew attentionto the trend from tall open forest to rainforest inhis studies in the Florentine Valley. Further studyin the same State has reinforced this: “the majorfactor determining the nature of the rainforest-eucalypt forest boundary is fire” (Neyland 1991).This is, or implies, a species-replacementargument for succession. Mount (1979), however,favoured the idea that “the pattern is one of stablefire cycles rather than one of succession”, perhapsa sort of “initial species composition model”(Egler 1954) or “complete initial floristics model”
(Wilson et al. 1992). Along the putative timegradient, the floristics simplify (Gilbert 1959).Jackson (1968) expanded the length of the sereending in rainforest by including a number offurther vegetation types beginning with sedgelandor grassland. Brown and Podger (1982) quantifiedspecies richness of the expanded sere in southwestTasmania (Melaleuca Inlet). Howard (1974)identified a sere from grassland to rainforestthrough Acacia stands in the northwest of theState. Ellis (1985), also in northwestern Tasmania,found evidence for the penetration of rainforestinto eucalypt communities and into shrublandswhich, in turn, were previously grasslands.
The situation in eastern Victoria in MitchellRiver National Park seems similar to thatdescribed for Tasmania to the extent that thereseems to be a trend from wet sclerophyll forest (or‘tall open forest’) to rainforest (Melick and Ashton1991). Melick’s (1990) ordination suggested thatundisturbed rainforest (27 quadrats) had fewerspecies than the fire-disturbed vegetation (30quadrats).
In tropical Australia eucalypt communitiesare being replaced by rainforests in some areas.Harrington and Sanderson (1994) usingphotogrammetry found that wet sclerophyll forestdominated by E. grandis was being replaced byrainforest. Unwin (1989) observed the rate ofadvance of rainforests in such situations to be 1.2m yr-1 over a 10-year study period; speciesrichness of the eucalypt forest was much less thanthat of the rainforest so the richness wasincreasing during succession in his sites nearHerberton.
Across northern Australia are ‘monsoonforests’, a type of rainforest often dominated byone or a few species; it may be regarded as a less-rich type of tropical rainforest. Fires may causethe retreat of its boundaries but, whenundisturbed, re-establishment at patch marginscan occur (Bowman and Fensham 1991) overdecades (Russell-Smith and Bowman 1992).Monsoon forest patches, surrounded by savanna(or, ‘grassy eucalypt woodland’) may contain moretree species (Bowman 1992) but fewer vascularplant species overall (Taylor and Dunlop 1985).
36
Australia’s Biodiveristy - Responses to Fire

4.6 DISCUSSION
The various methods used to discover changestaking place in species richness after fire all yielduseful data. However, the data needs to beinterpreted in the light of the methodology used.In many cases, the actual species present are notenumerated so whether or not the quoted richnessfor a number of sites have taxonomic andecological equivalence cannot be judged. By thechoice of technique, authors may assume thatspecies-richness changes occur in sites with: exactecological equivalence; identical initial speciescomposition; the same fire circumstances andproperties; and, equivalent post-fireenvironmental conditions.
The data reported here concern onlychanges in species richness after fire. They do notconcern extinction (Gill and Bradstock 1995). Inmost cases there is no indication whether or notthe changes identified are a simplification of aninitial complement or whether additions anddepletions have occurred. However, most species-richness depletions in phase “c” of Fig. 3.1 may beassumed to be due, as a first approximation, tospecies deletions without any addition. Wherethere has been an invasion of rainforest into othercommunities, substantial or total speciesreplacement may take place.
The range of plant communities directlyaffected by fire is large. Communities affectedinclude those in arid zones (e.g. Burbidge 1943,Allan and Griffin 1986, Allan and Baker 1990,Burrows and Christensen 1990) through to highrainfall areas dominated by temperate (eg. Barker1991, Melick and Ashton 1991) and tropical (e.g.Ridley and Gardner 1961, Bowman and Fensham1991) rainforests. Most, if not all, terrestrial plantcommunities seem to be affected.
A number of patterns of change in speciesrichness may be detected from the data presented: (i) the numbers of vascular plant species in most
communities soon after fire rise to levelshigher than those occurring immediatelybefore the fire;
(ii) numbers rise to reach a peak often withinone year but sometimes after 2 or more;
(iii) changes appear most conservative in openforests of Eucalyptus (‘dry sclerophyll forests’)(Christensen and Kimber 1975, Purdie 1977,Venning 1978, Hoare et al. 1980, Bowman etal. 1988, Tolhurst and Oswin 1992), possibleexceptions being some of the forests withheathy understoreys (presumed to be thecase in forests studied by Ingwersen 1977,Fox 1988 and Wark 1997);
(iv) heaths showed either no change in richnesswith time after fire (such as those studied byMcFarland 1988 in southeasternQueensland) or decreases as in Specht et al.(1958), Ingwersen (1977), Cheal et al.(1979), Clemens and Franklin (1980) andWark et al. (1987);
(v) rapid decreases soon after fire occurred inarid and semi-arid communities (especiallyhummock grasslands and mallee) (e.g.Zimmer (1940), Cheal et al. (1979), Burrowsand Christensen (1990), Noble (1989) andAllan and Baker (1990); and,
(vi) species-replacement sequences after fire mayinvolve increases (species-rich rainforest -Unwin 1989) or decreases (species poorrainforests - Brown and Podger 1982) inrichness.Observations of an absence of change in
richness may be due to intervals between ‘burnt’and ‘unburnt’ being too short. Thus one ofChristensen and Kimber’s 1975 examplesindicated a significant change over a 38 yearperiod but comparisons over a relatively shortperiod showed little change.
37
Biodiveristy and bushfires


In this chapter, the main aim is to explain the datapresented. The main hypothesis to emerge is thatthere are gradients in time sequences of speciesrichness after fire. Thus, we may hypothesize that:in the arid zone, there is a flush of herbaceousephemeral species after fire (hidden diversity)which increases species richness only temporallyas richness declines quickly as hummock grassesgains cover; in open forests of relatively mesicareas there is either no decline in richness afterfire or else there is a slow one; in the most mesicsites (well drained) there is a species replacementsequence from tall open forest to rainforest inwhich the richness increases or decreasesaccording to the site suitability for rainforests. Anapparent coarse-grained pattern based on rainfallcould be modified according to vegetation type,fire history, functional groups and soil typesperhaps. Formal testing of such ideas would bevaluable.
There are two main causes of the patterns inrichness after fire. The first is one that involves noaddition of species during time after fire, merelyan absence of change or a simplification in species’composition. The second is one that involvesspecies’ change (addition and/or replacement) atsome time during the sequence of changes thattake place with time after fire. The first is a formof “complete initial floristics model” (Egler 1954,Wilson et al. 1992) while the second conforms toClements’ “relay floristics” model (Wilson et al.1992). The first may be seen as havingchronological nestedness while the secondinvolves gains and losses in species with time. Avariant of the first is due to the expression ofherbaceous ephemerals as a consequence ofweather variations from year to year. Examples ofthe first of these two extremes are common inmallee, hummock grasslands and heathlands. Thesecond ‘extreme’ occurs when rainforest invadeseucalypt forest but may also occur when exoticsinvade or native dominants like Leptospermumlaevigatum (Russell and Parsons 1978) spread.
Only broad identification of patterns cancurrently be achieved because declarations ofspecies’ compositions are often not given whenshifts in species richness are described. Thereasons for this may include the lack ofdetermination of names (despite the
identifications of taxa) and interactions betweensites and treatments Thus, the importance of‘appearances’ and ‘disappearances’ of speciescannot be accurately assessed. However, enough isknown to hypothesize which patterns are likely tobe present and where they may occur.
It is likely that most patterns of the form ofFig.3.3 (b) - a rapid rise in numbers after firefollowed by a decline almost immediately - are ofthe chronologically-nested type while patternslike those in Fig. 3.3 (d and e) involve replacementof eucalypt-forest species by rainforests of variousrichness. However, the long sere identified inTasmania from sedgeland to rainforest (Brownand Podger 1982) shows a downward trend inspecies numbers during a long species-replacement series. That such a speciesreplacement series ending in rainforest can startfrom a number of communities was indicated byHoward (1974) and Ellis (1985). Furthermore,these observations do not preclude a self-replacement sequence within these communitieswith time after fire as evidenced by the changes inspecies richness in rainforest after fire describedby Melick (1990) in eastern Victoria.
5.1 PATTERNS OF RICHNESS AFTERFIRE: INVASION-CAUSED CHANGESIN COMPOSITION
Some of the variants in compositional changesthat may take place in relation to environment areillustrated in Fig. 5.1 (after Jackson 1968 andMount 1979). Compositional changes arerepresented by “communities”. The diagramshows four contiguous communities arrangedalong an environmental gradient. “Community 4”may be regarded as rainforest, “community 3” aseucalypt forest and the other two communities asearlier stages in the possible sere or habitatsequence. In the first case, (a), the communitiesare static in species composition but may passthrough a series of phases according to the timesince the passage of fire. In the second case, thecommunities can spread across the environmentalgradient, the only limitations to spread in the longterm being the fire regimes in the communitiespresent. Also, in this case, the rainforest mayspread into all the other communities. There is
39
Biodiveristy and bushfires
5. INTERPRETATION OF DATA
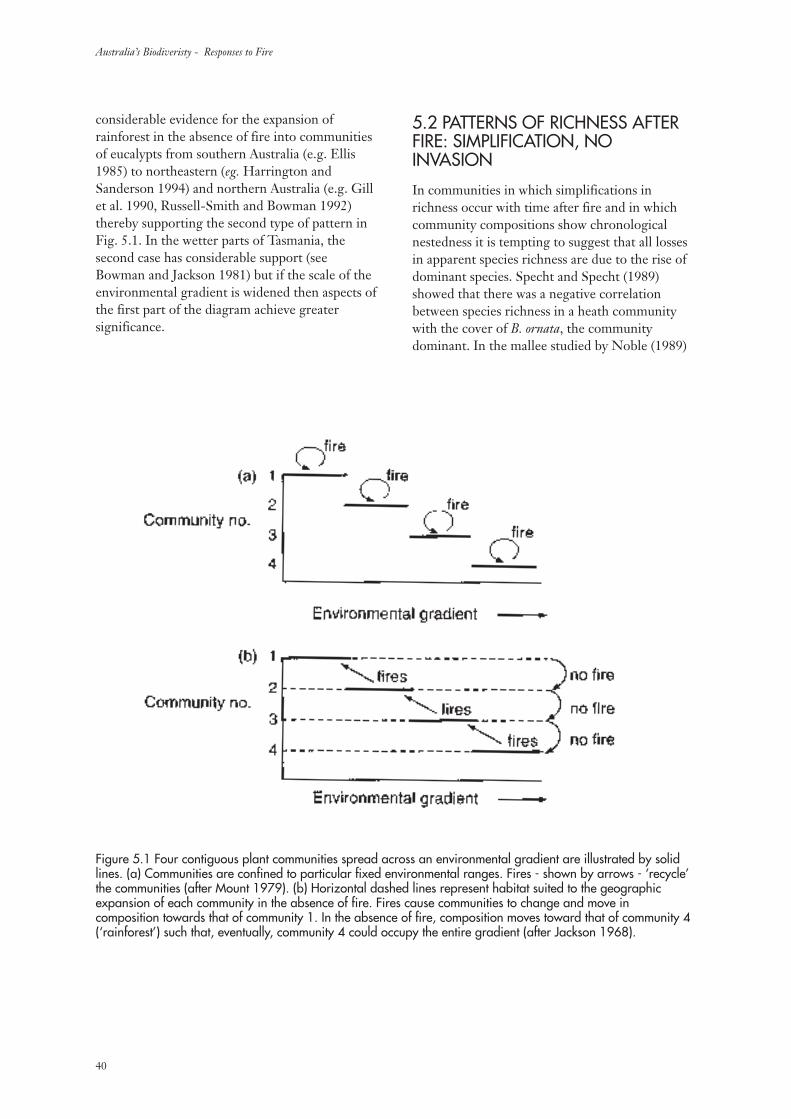
considerable evidence for the expansion ofrainforest in the absence of fire into communitiesof eucalypts from southern Australia (e.g. Ellis1985) to northeastern (eg. Harrington andSanderson 1994) and northern Australia (e.g. Gillet al. 1990, Russell-Smith and Bowman 1992)thereby supporting the second type of pattern inFig. 5.1. In the wetter parts of Tasmania, thesecond case has considerable support (seeBowman and Jackson 1981) but if the scale of theenvironmental gradient is widened then aspects ofthe first part of the diagram achieve greatersignificance.
5.2 PATTERNS OF RICHNESS AFTERFIRE: SIMPLIFICATION, NOINVASION
In communities in which simplifications inrichness occur with time after fire and in whichcommunity compositions show chronologicalnestedness it is tempting to suggest that all lossesin apparent species richness are due to the rise ofdominant species. Specht and Specht (1989)showed that there was a negative correlationbetween species richness in a heath communitywith the cover of B. ornata, the communitydominant. In the mallee studied by Noble (1989)
40
Australia’s Biodiveristy - Responses to Fire
Figure 5.1 Four contiguous plant communities spread across an environmental gradient are illustrated by solidlines. (a) Communities are confined to particular fixed environmental ranges. Fires - shown by arrows - ‘recycle’the communities (after Mount 1979). (b) Horizontal dashed lines represent habitat suited to the geographicexpansion of each community in the absence of fire. Fires cause communities to change and move incomposition towards that of community 1. In the absence of fire, composition moves toward that of community 4(‘rainforest’) such that, eventually, community 4 could occupy the entire gradient (after Jackson 1968).

the post-fire community was dominated by Stipa,a grass; as the community matured one wouldexpect a return to the dominance of eucalypts.This latter example is the simplest case of whatmay be termed ‘relay dominance’ where successivewaves of dominant species occur through timeeven while a simplification of communitycomposition occurs. Such trends would beobvious in rank-trajectory curves (see Section 3.4).
Zimmer (1940) ascribed diminishing speciesrichness during period “c” (Fig. 3.1) in a malleecommunity to increasing water use by thedominants - the mallee eucalypts and thehummock grass Triodia - while Noble (1989)suggested other possibilities for the effect.Functionally removing the dominant species couldremove allelopathic suppression of other specieswhile enhanced post-fire nutrition could allowexpression of some ephemerals. In tall openforests, post-fire environments may have morelight, more nutrients, lower disease potential andreduced herbivory (Ashton 1981).
Rank-abundance and rank-cover curvessuggest that there is a whole hierarchy ofdominance and that the extent of the differencesbetween first and second rankings - and others -may vary. The shallower the slope of the curve(Fig. 3.5) the greater the spread of ‘dominance’and the greater the potential for spatial variationin dominance to be expressed. Dominance isundoubtedly important but its extent andeffectiveness are important issues to resolve. Is aspecies which is most abundant or ‘important’with a high relative density, but at a low absolutedensity (in a post-fire community for example), afunctional dominant? Is a species which iscurrently first ranked but being replaced by thesecond-ranked species behaving as a dominant?
5.3 DISTRIBUTION OF PATTERNS OFRICHNESS CHANGE
The distribution of patterns of species richnesswith time may be examined by first simplifyingthe main patterns (Fig. 3.3) to three: (i) thepattern of general decline in richness in phase “c”,a pattern in which composition through time is‘nested’; (ii) the pattern in which phase “b” ispresent but “c” apparently is not, such that thecomposition of the community does not change;and, (iii) the pattern in which species rise ordecline in phase “c” due to species’ additionand/or replacement. The first pattern seems to bethe pattern of the arid and semi-arid zones in
which there are hummock grasslands, Acaciashrublands, mallee etc.. The second pattern, to alarge extent, seems characteristic of open eucalyptforests (‘dry sclerophyll forest’) where rainfall ishigher and the third pattern seems to be mostcommon in the highest rainfall areas where tall-open forests of eucalypts occur together withrainforests. Rainforest richness changes withdecreasing rainfall, increasing seasonality ofrainfall, decreasing soil fertility, decreasingtemperatures and increasingly impeded drainage(Kikkawa et al. 1981) so a temporal trend toincreasing richness in phase “c” may turn to oneof decreasing richness relative to that of thecommunities invaded along these environmentalgradients.
The geographic model above is a caricatureby definition. It does not reflect the detailperceived in some areas such as the wetter parts ofTasmania, for example, but is an hypothesis. Itreflects what may be expected generally across avast continent with a plethora of species andnumerous vegetation and soil types. There aremany areas still unstudied (Fig. 5.1).
Another way of examining the spatialpatterns in apparent richness after fire is toexamine what is known of the distributions ofrelevant functional groups. In Section 2.3, threefunctional groups were identified in relation tofires. They were the ‘sprouters’, ‘seeders’ and‘ephemerals’. Later (Section 3.1) ‘invader’ specieswere identified as ‘tolerant’ (T) or ‘requiring’ (R)species. These T and R species may be ‘seeders’ or‘sprouters’ (see Bowman 1991 for northernAustralian examples, Barker 1991 for Tasmanianexamples) but presumably such ‘sprouters’ inadvancing rainforests have a relatively lowtolerance to repeated fires.
In Victorian heathlands, invasion byLeptospermum laevigatum, a tall native ‘seeder’shrub, has been studied in relation to its effect onspecies richness (Molnar et al. 1989, Cheal 1996).The invasion of eucalypt forest understoreys bythe bird-dispersed ‘seeder’ shrub Pittosporumundulatum has been noted in Victoria (Gleadowand Ashton 1981). The exotic ‘seeder’ speciesPinus radiata has been invading forests in the ACTand northern Victoria (see Gill and Williams1996). There are other examples cited by Cheal(1996) who also noted the commonness of‘seeders’ among the invaders. As P. radiata and P.undulatum, at least, will invade without firedisturbance, they may be regarded as ‘T’ or ‘R’species in some circumstances.
41
Biodiveristy and bushfires

Ephemerals constitute a large part of theincreased richness observed after fire in arid andsemi-arid communities. A few ephemerals occureven after fire in tall open forests (Ashton 1981)and in rainforests (McMahon 1987). They appearto be rare in the drier eucalypt forests.Ephemerals are usually herbaceous in arid areas,woody in heathlands.
It would be interesting to know whether ornot the richness of ephemeral species was relatedto the extent and duration that bare ground wasexposed in different plant communities during afire rotation. Communities dominated by treesthat develop new shoots throughout their crownssoon after fire, or which retain their canopies aftera fire of low intensity (e.g. open forests ofeucalypts), may represent the circumstance wherefull exposure of bare ground to the sun is least. Inarid areas - where fires can be quite common -and bare ground is always exposed, the duration ofexposure of bare soil to full sun may reach amaximum.
Perhaps ephemeral richness could be relatedto the evolutionary radiation of particular taxa andtheir environmental tolerances? Solanum spp. canbe ephemerals from rainforest (McMahon 1987)to desert but seem more common in the latterenvironments (personal observation). C4 grassesmay be expected in the north, C3 in the south;Aristida spp. seem more frequent in the northwhile Stipa spp. seem more frequent in thesouthern, temperate, half of Australia. HerbaceousAmaranthaceae (e.g. Ptilotus, Gomphrena) seemmore common in the northern parts of Australiaand herbaceous Asteraceae (eg. Helichrysum,Helipterum) in the southern.
A guild of ephemeral species will notnecessarily be expressed in toto at any one time.Indeed, the results of Fox (1990) suggest that thecomposition fluctuates a great deal from year toyear. A number of authors have noted that in aridand semi-arid lands ephemeral forbs appear afterwinter rain while grasses respond to summer rains(Griffin and Hodgkinson 1986). Will ephemeralsin northern arid lands, where rainfalls arepredominantly in summer, be largely grassytherefore?
‘Sprouter’ species are very widespread indistribution. They are found in rainforests(Bowman 1991) and deserts (Griffin 1984).Sprouters are the most common fire-responsetype in the savannas of northern Australia (Gill etal. 1990). Many species of the open eucalypt forestof southeastern Australia, including thedominants, also fall into this category. ‘Sprouters’with no observed seed regeneration are rare buthave been recorded in Victorian forests(McMahon 1987).
‘Seeders’, like ‘sprouters’, are widespread.Many of the tall eucalypt forest dominants areintolerant ‘seeders’ (Ashton 1981) as are many ofthe dominants of heathlands (e.g. Banksia ornata,Specht et al. 1958). Indeed, 50-60% species in tallopen forests are ‘seeders’ (Ashton 1981) andcomprise over 60% of species in mallee, woodlandand shrubland in southwestern Australia (Van derMoezel and Bell 1984). In a Victorian heathland33% species were ‘seeders’ (Wark et al. 1987), apercentage within the 20-40% found for heaths insoutheastern Queensland (C. Sandercoe personalcommunication). Woody ‘seeders’ are notuncommon in the arid zone (Griffin 1984) and areoften dominants e.g. Acacia aneura, Thryptomenemaisonneuvii and Callitris spp. ‘Seeders’ are spreadacross the landscapes of Australia, theirproportions varying from place to place.
5.4 A PLANT-COMMUNITYDYNAMICS VIEWPOINT
The patterns observed in any one communitydepend on the dynamics of the species in thatcommunity, the species within dispersal distancethat can invade, and various aspects of the physicaland biological environments at those sites. Intrying to understand the patterns observed, then,one might turn to the study of the detaileddynamics of the communities of interest. There isno intention to attempt an analysis of such studieshere. Rather, it is to be pointed out that thedynamics of any of the communities studied willinvolve the compositions and life cycles of thesoil- propagule pool, the ‘apparent’ species pool,and, the within-dispersal-distance pool (see Fig.6.1 in the next chapter).
42
Australia’s Biodiveristy - Responses to Fire

From earlier chapters of this Report, it is apparentthat fires are a major factor affecting the plantdiversity of terrestrial Australia. Fires can affectthe expression of diversity and the distribution ofdiversity. Inappropriate fire regimes (Gill 1975,1981) can cause the local extinction of plantspecies (Gill and Bradstock 1995). As fire is amanagement tool, as well as a natural agent ofcommunity dynamics, management fires can beused to manipulate biodiversity. Understandingfires and their effects is important if biodiversity isto be understood and properly managed.
The fire-management aims for any parcel ofland - implicit or explicit - are a subset of generalmanagement aims. General aims often include themaintenance of species diversity and theprotection of lives and property. To effectivelymanage the ecological estate, an assessment ofdiversity, especially richness, is necessary. Howthis can be done and the results used in thedevelopment of a monitoring and decision-support system has been suggested by Gill andNicholls (1989). To assist in the development ofsuch systems, a data base listing the fire-responsesof Australian vascular plant species has beeninitiated (Gill and Bradstock 1992).
Fire management involves both thesuppression of unwanted fires and the ignition ofother fires under specified conditions for specifiedpurposes. It may also involve attempts to controlunwanted ignitions through education campaignsand the closure of various areas at times of‘extreme’ fire danger.
The fuels for management fires arise fromthe extant plant community in most cases. [Insome circumstances, peat derived largely fromcommunities long gone, may be the main sourceof fuel for unplanned, unwanted fires.] The fuelsfor management fires come from a select group ofspecies, not from all species making an equalcontribution. The “fuel species” (Fig. 6.1) areusually dominants but not necessarily so. Majorfuels in Australian plant communities are suppliedby grasses such as Themeda spp., Sorghum spp.,Heteropogon spp. and Triodia spp., by varioussedges and shrubs, and by tree and shrub litters(especially those of eucalypts). Of course,unintentional fires may be fuelled by cereal crops,pastures and plantations as well. In the depictionof an ecosystem being managed by fire, the fuelspecies need to be distinguished (Fig. 6.1). Fuelspecies can be manipulated by fire to increase
43
Biodiveristy and bushfires
6. FIRE MANAGEMENT OF PLANT DIVERSITY
Figure 6.1. A simplified conceptual diagram illustrating inter-relationships between groups of species in a fire-prone environment.

their productivity (e.g. by killing shrubbydominants thereby allowing a grass response) ordecreasing it (e.g. by killing the fuel species as hasbeen done with Sorghum in the NorthernTerritory - Stocker and Sturtz 1966).
Increasingly, exotic species are forming partsof Australian plant communities. Exotic plantspecies may be expected to provide fuel species andmembers for all the functional groups identified inearlier sections. Exotic species may enter the soilseed pool and remain dormant until the appropriateconditions occur for regeneration or they mayinvade the plant community just as other ‘requiring’species (Noble and Slatyer 1980) may do so (Fig.6.1). In conservation management, exotic species arean unwanted part of the diversity of biota.
If, as asserted here, exotic species comprisemembers of the same functional groups as thenative biota, opportunities for the elimination ofsuch species using fires while retaining the nativespecies may seem slim. However, if the exotic isthe only species in a particular functional group ata particular site or has extreme attributes makingit the most vulnerable in a class, then the use offire for its control is a distinct possibility. Anexample of this class of circumstances occurs whenPinus radiata invades perennial grassland oreucalypt open forest in the Australian CapitalTerritory (ACT) or elsewhere (Gill and Williams1996). In some cases fires may be used, perhaps incombination with other potential control agents(like selective herbicides).
Exotic species can be fuel species. Examplesare invasive Pinus radiata (personal observation),the shrub Lantana camara, and mission grass,Pennisetum polystachyon (see Gill et al. 1990).Exotic animals can affect fuel species by eatingthem, by transporting them or by creatingconditions suitable for their spread. In theKimberley it appears that cattle are breakingdown rainforest edges and either transportingseeds of grasses (fuel species) into them ormodifying the environment such that fires canenter more readily (McKenzie and Belbin 1991).Rabbits may exert a marked effect on fuel loads insemi-arid woodlands (Leigh et al. 1989) and nodoubt elsewhere. By affecting fuel characteristicsexotic species can alter fire regimes and affect theability of managers to maintain biodiversity.
An examination of the time for theattainment of maximum species richness after firemay, incorrectly, suggest a suitable timing of firesfor the management of plant-species diversity. Forexample, where maximum diversity is reached at
the end of phase “a”, burning may be wronglyscheduled then in order to keep the intervalsbetween fires short (to obtain maximum control ofunplanned fires) and keep the diversity at amaximum. This procedure, however, would be amistake because certain species of ‘seeders’ may bemade to become locally extinct due toinadequacies in the time necessary for the supplyof seed for subsequent regeneration (Gill andBradstock 1995). On the other hand, if themaintenance of a diverse set of stages within thespectrum of change after fire is seen as theobjective of management then it is possible that allthe species in the system could be maintained.Burning would still have to be within a domain offire regimes commensurate with the survival of allspecies. Ways in which the variety of times sincefire, or post-fire stages, could be achieved include:(i) burning at a fixed, relatively long, interval
but in different places each year;(ii) burning at different fixed rotations in
different burning blocks within anecologically safe range of intervals; and,
(iii) as for (ii) but using a stochastic orprobability method to allocate intervals ineach block.The last of these seems most appropriate for
conservation management as it provides the mostvariation in time and space (see also Gill andMcCarthy 1998). Variations in such a prescriptionwould need to be made in some places because ofthe need to accommodate other management aimssuch as the protection of life and property.Calibration of the probability model used (and itsform) would be essential and may be assisted by arobust monitoring method (see Gill 1998).
Fire management takes place at a landscapeor vegetation-community scale. At such scalesthere are many species present, some of whichmay be ‘special’ in that they are rare or vulnerableto extinction. Because fire management takesplace at a broad scale, any management aimed atthe preservation of ‘special’ species will necessarilytake place at the same broad scale. Thecommunity will be managed for the species.
A major need for fire management today isthe implementation of practical monitoringsystems. Testing the suggestions already made(Gill and Nicholls 1989, Gill 1998) would be astart. Having a suitable response system to theresults of monitoring is important also. Assessingwhere such systems are most appropriate andimplementing them is important if biodiversity isto be managed in an accountable way.
44
Australia’s Biodiveristy - Responses to Fire

Research into the effects of fires on biodiversityhas a wider compass than the focus of this review.However, the overlap is extensive. The problem ishow to assess the ecological effects of fires on thesometimes overwhelming biodiversity ofterrestrial Australia when this includes vascularand non-vascular plants, vertebrate and non-vertebrate animals, bacteria, algae and fungi,native and exotic. Fire effects vary with fireregimes (see the Preamble) so the task is notsimply to examine the consequences of a singlefire on community composition over time - thecontext of this review - but to assess all the effectsof all fire regimes on all components of thebiodiversity across Australia, a formidablechallenge.
Of around 18,000 vascular plant species inAustralia (Hnatiuk 1990), a quarter (23%) are rareor threatened (Briggs and Leigh 1996). There arealso nearly 2,000 officially recognized naturalizedspecies (Hnatiuk 1990). Experimental examinationof the responses of each of the rare and threatenedspecies, let alone the whole flora, to a range of fireregimes - even if the appropriate range of regimeswas known, and experimentally feasible toimplement, and funding and land was available -seems unlikely. A variety of methods to observeeffects - and not just those after a fire - needs tobe adopted.
Data for the determination of plant-specieschanges with time after fire is collected in avariety of ways but the variables of importance atany one stage are the number of species, thenumber of individuals of each species, the areasover which sampling occurs and the cover orbiomass of each species. There is a need forefficient statistically-robust methods for thecollection of data on all of these variables at anyone site. Problems of methodology wereaddressed in Chapter 4. The more measures ofdiversity that are used, and the greater the numberof explanatory variables that are also measured,the greater the potential understanding.
Interpretation of the data collected may bemade in terms of dominance, life cycles andfunctional groups. A knowledge of the attributesof species within dispersal distance but not alreadypresent may also assist. Species attributes -defined along the lines of Noble and Slatyer
(1980) and Gill (1981) - have been used to definefunctional groups; the attributes can often bedetermined from observations made soon afterany fire. In a Register of Plant Species Responses(Gill and Bradstock 1992) about 10% of the florahas been listed but much of it only in the broadestof functional groups. With the addition of timescales for key events for each species (fruiting,longevity), predictions can be made as to species’presence or absence at particular times after a fire(Noble and Slatyer 1980); there are few data ofthis type.
Researchers simplify in order to cope withcomplexity. They will usually tackle one group oforganisms such as vascular plants or vertebrateanimals. They will tend to choose a single area, setup plots and examine the effects of a limited rangeof fire intensities, frequencies and seasonalities fortheir experiments. Safety considerations usuallylimit the range of intensities used (Section 4.4).Metronome-like frequencies are idealexperimentally but unlikely in nature.Seasonalities adopted may be strongly influencedby restrictions imposed by fire authorities. Peatfires are ignored in experimental design becausethe occurrence of peat fires implies prevailingdrought, days, weeks or months for thecombustion process to be completed, and extremefire-suppression difficulties. Site choice may beinfluenced by the likelihood of being able to ignitefires at the designated times. Complications suchas the interactions between burning and grazing(Leigh and Holgate 1979), fires and frosts (Duffand Stocker 1989) and fires and cyclones (Stockerand Mott 1981) are worth further examination.There are financial and practical limits to theconduct of experiments but experiments providecontrolled data.
State and Federal Forestry Departmentshave been the main instigators of fire-regimeexperiments in Australia. Examples are those setup by: the Forest Research Institute of theAustralian Forestry and Timber Bureau (now partof CSIRO) in the Northern Territory and ACT(J. Hoare); the Qld Department of Forestry insouth-eastern Queensland (Byrne 1989); NSWState Forests (A. York and D. Binns); VictorianState government authorities (Tolhurst and Flynn1992); the SA Woods and Forests Department
45
Biodiveristy and bushfires
7. RESEARCH ON SHIFTS IN PLANT DIVERSITY AFTER FIRE

(B. Gepp) and the Department of Forestry inWestern Australia, now Department ofConservation and Land Management (L. McCawand N. D. Burrows). CSIRO Division of Wildlifeand Ecology set up the large Kapalga experimentin the Northern Territory (Duff and Braithwaite1989) while recently CSIRO Plant Industryestablished two experiments, one in the ACT (byJ. Briggs, now with NSW National Parks andWildlife Service), the other in NSW (byS. Prober, now freelance). Experiments to datehave been mainly in forests and grassy woodlands,the exception in the above examples being thoseexperiments in Queensland where heath andshrubby woodlands were chosen. There isconsiderable scope for further experiment.(Experiments in which one fire only has been usedor in which artificial fuel has been used are not aconsideration here.)
Because safety factors exclude the use ofextreme intensities in experiments, carefulobservations of the effects of unplanned severefires are an important source of informationdespite their occurrence being in unpredictablelocations at unpredictable times. For the samereason, theoretical modelling is valuable.
With a knowledge of the way in which fireregimes are distributed across the nationaccording to biophysical and social (includingmanagement) factors, it will be possible to predictthe proportions of landscapes at particular stagesafter fire. Then we will be able to predict thespecies richness to be found in each piece of thelandscape mosaic. How fire regimes aredistributed in landscapes due to chance is justbeginning to receive attention in Australia (Gilland McCarthy 1998). Models being developed inthis context have the potential to greatly expandour understanding of the effects of fire regimes onbiodiversity in Australia.
From a management point of view, aresearch challenge is to define appropriate fireregimes for different ecosystems in differentsocial, economic, ecological, and geographiccontexts and to assess the need for, or theconsequences of, burned areas having particularsizes, shapes and arrangements.
To assist research we can formulatehypotheses, ask questions. For example:• Are the changes in vascular plant diversity
with time since fire simply an outcome of thefrequency of species in different functionalgroups found there as a result of particularsite histories and environmental (non-fire)selection?
• To what extent do the details within afunctional group (such as time to floweringand fruiting, longevity) affect the outcome oftime-since-fire sequences?
• Are patterns of species richness with timeafter fire a function of local and nationalenvironmental gradients?There is still a great deal to be learned about
the changes in diversity that occur after fireshaving various characteristics and occurring atvarious times of year. Establishing a benchmarkmethodology would help.
46
Australia’s Biodiveristy - Responses to Fire

Biodiversity, the variety of life, can be expressedmost simply as a list of species - ‘species richness’.Species richness varies with sample area (andlocations), the relationships being expressed usingsimple equations. Plant species richness can be‘apparent’ (in the form of plants above ground) or‘hidden (only in the form of seeds, or otherpropagules, below ground). Apparent speciesrichness is negatively affected by ecologicallydominant species, the dominance being illustratedby rank-abundance, rank-cover or rank-biomasscurves. Whether or not the first-ranked speciesare functionally dominant may need someclarification.
Fires occur in most of the plant communitiesof Australia. They can have major affects on thebiota through fire regimes. Plant species richnesschanges, sometimes dramatically, with time afterfire. In the absence of species’ identities beingexplicit, changes in richness with time aredepicted as numbers only. Where this occursthere can be no knowledge of species turnover.
Understanding the dynamics of change withtime since fire requires a knowledge of therelevant functional groups of plants, such as
‘ephemerals’, ‘seeders’ and ‘sprouters’. Subdivisionof broad groups into woody or herbaceousephemerals, ‘seeders’ with canopy-seed or soil-seed stores, and sprouters normally with orwithout seedlings after fire, can be worthwhile.Identifying ‘tolerant’ and ‘requiring’ species isimportant especially where invasions occur.Knowledge of the times to produce seeds and thelongevity of species is highly significant.
To conserve native plant diversity, one needsto avoid extinction and prevent invasion ofexotics. Fire regimes affect species’ survival andare an essential tool of management. Bothprescribed fires and unplanned fires can affectlocal diversity. Management for the conservationof biodiversity should aim at achieving suitableproportions of landscape with a variety of times-since-fire stages using fires within appropriateintensity levels at appropriate times of year andwithin an appropriate frequency range. A researchchallenge is to define these ‘suitable proportions’and ‘appropriate regimes’ for different ecosystemsin different management, ecological, andgeographic contexts and to assess any need for anyparticular spatially-explicit arrangement of burnedareas.
47
Biodiveristy and bushfires
8. CONCLUSIONS

This report is the result of a contract with theAustralian Department of the Environment andHeritage. Neal Hardy, from the Department, is tobe thanked for his encouragement and facilitationof the project. Also, I would like to thank JulieMcAlpine for her technical support. PennyHohnen and Peter Moore compiled the map (Fig.4.1) from data supplied by the author. PeterMoore provided logistic and moral supportthroughout the project. Ross Bradstock, GrahamGriffin, Jim Noble, Kevin Tolhurst, Jann Williamsand John Woinarski kindly read draft manuscriptsand offered constructive suggestions forimprovement.
48
Australia’s Biodiveristy - Responses to Fire
9. ACKNOWLEDGEMENTS

Allan, G. and Baker, L. (1990). Uluru (AyersRock-Mt. Olga) national park: an assessmentof a fire management programme. Proc. Ecol.Soc. Aust. 16, 215-220.
Allan, G. E. and Griffin, G. F. (1986). Fireecology of the hummock grasslands ofCentral Australia. Proceedings of the 4thAustralian Rangeland Society Conference.Pp. 126-129.
Arrhenius, O. (1921). Species and area. J. Ecol. 9,95-99.
Ashton, D. H. (1981). Fire in tall open-forests(wet sclerophyll forests). In: A. M. Gill, R.H. Groves and I. R. Noble (eds) Fire and theAustralian Biota. Pp. 339-366. AustralianAcademy of Science, Canberra.
Barker, M. J. (1991). The Effect of Fire on WestCoast Lowland Rainforests. TasmanianNational Rainforest Research ProgramReport No.7.
Bell, D. T. and Koch, J. M. (1980). Post-firesuccession in the northern jarrah forest ofWestern Australia. Aust. J. Ecol. 5, 9-14.
Bell, D.T., Hopkins, A. J. M. and Pate, J. S.(1984). Fire in the Kwongan. In: J. S. Pateand J. S. Beard. (eds) Kwongan. Plant Life ofthe Sandplain. Pp.178-204. University ofWestern Australia Press, Nedlands, Perth.
Bowman, D. M. J. S. (1991). Recovery of somenorthern Australian monsoon forest treespecies following fire. Proc. Roy. Soc. Qld.101, 21-25.
Bowman, D. M. J. S. (1992). Monsoon forests innorth-western Australia. II. Forest-savannatransitions. Aust. J. Bot. 40, 89-102.
Bowman, D. M. J. S. and Fensham, R. J. (1991).Response of a monsoon forest-savannaboundary to fire protection, Weipa, northernAustralia. Aust. J. Ecol. 16, 111-118.
Bowman, D. M. J. S. and Jackson, W. D. (1981).Vegetation succession in southwestTasmania. Search 12, 358-362.
Bowman, D. M. J. S., Wilson, B. A. and Hooper,R. J. (1988). Response of Eucalyptus forestand woodland to four fire regimes atMunmarlary, Northern Territory, Australia.J. Ecol. 76, 215-232.
Bray, J. R. and Curtis, J. T. (1957). An ordinationof the upland forest communities ofsouthern Wisconsin. Ecol. Monog. 27, 325-349.
Briggs, J. D. and Leigh, J. H. (1996). Rare orThreatened Australian Plants. CSIRO,Melbourne.
Brown, M. J. and Podger, F. D. (1982). Floristicsand fire regimes of a vegetation sequencefrom sedgeland-heath to rainforest atBathurst Harbour, Tasmania. Aust. J. Bot.30, 659-676.
Burbidge, N. T. (1943). Ecological successionobserved during regeneration of Triodiapungens after burning. J. Roy. Soc. W.A. 28,149-156.
Burrows, N. D. and Christensen, P. E. S. (1990).A survey of Aboriginal fire patterns in thewestern desert of Australia. USDA For. Serv.Gen. Tech. Rep. SE 69, 297-305.
Bureau of Meteorology (1988). Climatic Averages.Australia. Australian Government PublishingService, Canberra.
Byrne, P. J. (1989). Forest fire research by theDepartment of Forestry in Queensland. In:B. R. Roberts (ed) Fire Research in RuralQueensland. Pp. 229-236. University ofSouthern Queensland, Toowoomba.
Cheal, D. (1996). Fire succession in heathlandsand implications for vegetation management.Australian Department of Environment, Sportand Territories Biodiversity Series, Paper No. 8,67-79.
Cheal, P. D., Day, J. C. and Meredith, C. W.(1979). Fire in the National Parks of NorthWest Victoria. National Parks Service,Melbourne.
Christensen, P. E. and Kimber, P. C. (1975).Effects of prescribed burning on the floraand fauna of south-west Australian forests.Proc. Ecol. Soc. Aust. 9, 85-106.
Clemens, J. and Franklin, M. H. (1980). Adescription of coastal heath at North Head,Sydney Harbour National Park: Impact ofrecreation and other disturbance since 1951.Aust. J. Bot. 28, 463-478.
Cockburn, A. (1978). The distribution ofPseudomys shortridgei (Muridae: Rodentia)and its relevance to that of other heathlandPseudomys. Aust. Wildl. Res. 5, 213-219.
49
Biodiveristy and bushfires
10. REFERENCES CITED

Cochrane, G. R., Burnard, S., and Philpott, J. M.(1962). Land use and forest fires in theMount Lofty Ranges South Australia. Aust.Geographer 8(4), 143-160.
Connor, E. F. and McCoy, E. D. (1979). Thestatistics and biology of the species-arearelationship. Amer. Nat. 113, 791-833.
Curtis, J. T. and McIntosh, R. P. (1950). Theinterrelationships of certain analytic andsynthetic phytosociological characters.Ecology 31, 434-455.
Doak, D. F. and Mills, L. S. (1994). A useful rolefor theory in conservation. Ecology 73, 615-626.
Duff, G. A. and Braithwaite, R. W. (1989). Fireand Top End Forests - Past, Present andFuture Research. In: B.R.Roberts (ed) FireResearch in Rural Queensland. Pp. 84-98.University of Southern Queensland,Toowoomba.
Duff, G. A. and Stocker, G. C. (1989). The effectsof frosts on rainforest/open forest ecotonesin the highlands of North Queensland. Proc.Roy. Soc. Qld. 100, 49-54.
Egler, F. E. (1954). Vegetation science concepts. 1.Initial floristic composition, a factor in old-field vegetation development. Vegetatio 4,412-417.
Ellis, R. C. (1985). The relationships amongeucalypt forest, grassland and rainforest in ahighland area in north-east Tasmania. Aust.J. Ecol. 10, 297-314.
Fox, J. E.D., Pepper, C. and Craig, G. (1983).Vegetation and reaction to fire in coastalheath at Sorrento, Western Australia. MulgaResearch Institute Annual Report No. 6. Pp.97-109.
Fox, M. D. (1988). Understorey changesfollowing fire at Myall Lakes, New SouthWales. Cunninghamia 2(1), 85-95.
Fox, M. D. (1990). Composition and richness ofNew South Wales mallee. In: J. C. Noble, P.J. Joss and G. K. Jones (eds) The MalleeLands. A Conservation Perspective. Pp. 8-11.CSIRO, Melbourne.
Fox, M. D. and Fox, B. J. (1986). The effect of firefrequency on the structure and floristiccomposition of a woodland understorey.Aust. J. Ecol. 11, 77-85.
Gilbert, J. M. (1959). Forest succession in theFlorentine Valley, Tasmania. Pap. Proc. Roy.Soc. Tas. 93, 129-151.
Gill, A. M. (1975). Fire and the Australian flora: areview. Aust. For. 38, 4-25.
Gill, A. M. (1977). Management of fire-pronevegetation for plant species conservation inAustralia. Search (Syd.) 8, 20-26.
Gill, A. M. (1981). Coping with fire. In: J. S. Pateand A. J. McComb (eds) The Biology ofAustralian Plants. Pp. 65-87. University ofWestern Australia Press, Nedlands.
Gill, A. M. (1993). Interplay of Victoria’s florawith fire. In: D. B. Foreman and N. G.Walsh (eds) Flora of Victoria. Volume 1.Introduction. Pp. 212-226. Inkata Press,Melbourne.
Gill, A. M. (1995). Stems and fires. In: B. L.Gartner (ed) Plant Stems. Physiological andFunctional Morphology. Pp. 323-342.Academic Press, San Diego.
Gill, A. M. (1998). Monitoring for biodiversity infire-prone areas. In: R. Melzer (ed) FireManagement: Presented Papers. QueenslandDepartment of the Environment,Rockhampton.
Gill, A. M. and Bradstock, R. A. (1992). A nationalregister for the fire responses of plantspecies. Cunninghamia 2, 653-660.
Gill, A. M. and Bradstock, R. A. (1994). Theprescribed burning debate in temperateAustralian forests: towards a resolution. Pp.703-712. Proceedings of the 2ndInternational Conference on Forest FireResearch, Coimbra, Portugal.
Gill, A. M. and Bradstock, R. A. (1995).Extinctions of biota by fires. In: R. A.Bradstock, T. D. Auld, D. A. Keith, R. T.Kingsford, D. Lunney and D. P. Sivertsen(eds) Conserving Biodiversity: Threats andSolutions. Pp. 309-322. NSW National Parksand Wildlife Service, Sydney.
Gill, A. M. and McCarthy, M. C. (1998). Intervalsbetween prescribed fires in Australia: whatintrinsic variation should apply? Biol.Conserv.85, 161-169.
Gill, A. M. and Moore, P. H. R. (1990). Fireintensities in Eucalyptus forests ofsoutheastern Australia. Proceedings 1stInternational Forest Fire ResearchConference, Coimbra, Portugal. Pp. B.24-1to 12.
Gill, A. M. and Nicholls, A. O. (1989).Monitoring fire-prone flora in reserves fornature conservation. In: N. Burrows, L.McCaw, and G. Friend (eds) FireManagement on Nature Conservation Lands.W.A. Dept. C.A.L.M. Occ. Pap. 1/89, pp137-151.
50
Australia’s Biodiveristy - Responses to Fire

Gill, A. M. and Williams, J. E. (1996). Fireregimes and biodiversity: the effects offragmentation of southeastern eucalyptforests by urbanisation, agriculture and pineplantations. For. Ecol. & Managt 85, 261-278.
Gill, A. M., Hoare, J. R. L and Cheney, N. P.(1990). Fires and their effects in the wet-drytropics of Australia. Ecol. Stud. 84, 159-178.
Gill, A. M., Groves, R. H. and Noble, I. R. (eds)(1981). Fire and the Australian Biota.Australian Academy of Science, Canberra.
Gitay, H., Roxburgh, S. H. and Wilson, J. B.(1991). Species-area relations in NewZealand tussock grassland, with implicationsfor nature reserve design and for communitystructure. J. Veg. Sci. 2, 113-118.
Gleadow, R. M. and Ashton, D. H. (1981).Invasion by Pittosporum undulatum of theforests of Central Victoria. I. Invasionpatterns and plant morphology. Aust. J. Bot.29, 705-720.
Gleason, H. A. (1922). On the relation betweenspecies and area. Ecology 3, 158-162.
Gray, J.S. (1986). Species-abundance patterns. In:J. H. R. Gee and P. S. Giller (eds)Organization of Communities: Past and Present.Symposium of the British Ecological Society.Pp. 53-67. Blackwell Scientific, Oxford.
Griffin, G. F. (1984). Vegetation patterns. In: E.C. Saxon (ed) Anticipating the Inevitable: APatch-burn Strategy for Fire Management atUluru (Ayers Rock - Mt Olga) National Park.Pp. 25-37. CSIRO, Melbourne.
Griffin, G. F. (1992). Will it burn - should itburn?: management of the spinifexgrasslands of inland Australia. In: G.P.Chapman (ed) Desertified Grasslands: TheirBiology and Management. Pp. 63-76.Academic Press, London.
Griffin, G. F. and Hodgkinson, K. C. (1986). Theuse of fire for the management of the mulgaland vegetation in Australia. In: P. Sattler(ed) The Mulga Lands. Pp. 93-97. RoyalSociety of Queensland, Brisbane.
Harrington, G. N. and Sanderson, K. D. (1994).Recent contraction of wet sclerophyll forestin the wet tropics of Queensland due toinvasion by rainforest. Pacific ConservationBiology 1, 319-327.
Harrold, A. (1979). Heathland regeneration afterfire at Noosa. Qld. Natur. 22, 88-96.
Hesp, P. A., Wells, M. R., Ward, B. H. R. andRiches, J. R. H. (1983). Land Resource Surveyof Rottnest Island. Western AustralianDepartment of Agriculture, Perth.
Hnatiuk, R. J. (1990). Census of Australian VascularPlants. Australian Flora and Fauna SeriesNumber 11. Australian GovernmentPublishing Service, Canberra.
Hoare, J. R. L., Hooper, R. J., Cheney, N. P. andJacobsen, K. L. S. (1980). A Report on theEffects of Fire in Tall Open Forest andWoodland with particular Reference to FireManagement in Kakadu National Park in theNorthern Territory. Unpublished Report tothe Australian National Parks and WildlifeService, Canberra.
Hobbs, R. J. and Atkins, L. (1990). Fire relateddynamics of a Banksia woodland in south-western Western Australia. Aust. J. Bot., 38,97-110.
Howard, T. M. (1974). Nothofagus cunninghamiiecotonal stages. Buried viable seed in NorthWest Tasmania. Proc. Roy. Soc. Vic. 86, 137-142.
Hubbell, S. P. (1979). Tree dispersion, abundanceand diversity in a tropical dry forest. Science203, 1299-1309.
Hughes, R. G. (1986). Theories and models ofspecies abundance. The American Naturalist128, 879-899.
Ingwersen, F. (1977). Regeneration of Vegetationafter Fire at Jervis Bay and its Implicationsfor Management. Unpublished M.Sc. thesis,Australian National University. 415 p.
Jackson, W. D. (1968). Fire, air, water, and earth -an elemental ecology of Tasmania. Proc. Ecol.Soc. Aust. 3, 9-16.
Kikkawa, J., Webb, L. J., Dale, M. B., Monteith,G. B., Tracey, J. G. and Williams, W. T.(1981). Gradients and boundaries ofmonsoon forests in Australia. Proc. Ecol. Soc.Aust. 11, 39-52.
Knight, D. H. (1975). A phytosociological analysisof species-rich tropical forest on BarroColorado Island, Panama. Ecol. Monog. 45,259-284.
Kobayashi, S. and Kimura, K. (1994). Thenumber of species occurring in a sample of abiotic community and its connections withspecies-abundance relationship and spatialdistribution. Ecol. Res. 9, 281-294.
51
Biodiveristy and bushfires

Lamont, B. B., Downes, S. and Fox, J. E. D.(1977). Importance-value curves anddiversity indices applied to a species-richheathland in Western Australia. Nature 265,438-441.
Leigh, J. H. and Holgate, M. D. (1979). Theresponses of the understorey of forests andwoodlands of the Southern Tablelands tograzing and burning. Aust. J. Ecol. 4, 25-45.
Leigh, J. H., Wimbush, D. J., Wood, D. H.,Holgate, M. D., Slee, A. V., Stanger, M. G.and Forrester, R. I. (1987). Effects of rabbitgrazing and fire on a subalpine environment.I. Herbaceous and shrubby vegetation. Aust.J. Bot. 35, 433-464.
Leigh, J. H., Wood, D. H., Holgate, M. D., Slee,A. and Stanger, M. G. (1989). Effects ofrabbit and kangaroo grazing on two semi-arid grassland communities in Central-western New South Wales. Aust. J. Bot. 37,375-396.
Loehle, C. (1990). Proper statistical treatment ofspecies-area data. Oikos 57, 143-145.
MacArthur, R. H. and Wilson, E. O. (1963). Anequilibrium theory of insular zoogeography.Evolution 17, 373-387.
MacArthur, R. H. and Wilson, E. O. (1967). TheTheory of Island Biogeography. PrincetonUniversity Press, Princeton, New Jersey.
McFarland, D. C. (1988). Fire and vegetationcomposition and structure of subtropicalheathlands in south-east Queensland. Aust.J. Bot. 36, 533-546.
McKenzie, N. L. and Belbin, L. (1991).Kimberley rainforest communities: reserverecommendations and managementconsiderations. In: N. L. McKenzie, R. B.Johnson and P. G. Kendrick (eds) KimberleyRainforests. Pp. 453-480. Surrey Beatty andSons, Chipping Norton, New South Wales.
McMahon, A. (1984). The effects of time sincefire on heathlands in the Little Desert, N.W.Victoria, Australia. In: B. Dell. (ed)MEDECOS IV, Proceedings 4th InternationalConference on Mediterranean Ecosystems.Pp.99-100.
McMahon, A. R. G. (1987). The Effects of the1982-83 Bushfires on Sites of Significance.Victorian Department of Forests & Lands,Environmental Studies Publication SeriesNo. 411.
McNaughton, S. J. (1993). Biodiversity andfunction of grazing ecosystems. Ecol. Stud.99, 361-383.
Melick, D. R. (1990). Ecology of rainforest andsclerophyllous communities in the MitchellRiver National Park, Gippsland, Victoria.Proc. Roy. Soc. Vic. 102, 71-87.
Melick, D. R. and Ashton, D. H. (1991). Theeffects of natural disturbances on warmtemperate rainforests in south-easternAustralia. Aust. J. Bot. 39, 1-30.
Menhinick, E. F. (1964). A comparison of somespecies-individuals diversity indexes appliedto samples of field insects. Ecol. 45, 859-861.
Molnar, C. D., Fletcher, D. and Parsons, R. F.(1989). Relationships between heath andLeptospermum laevigatum scrub atSandringham, Victoria. Proc. Roy. Soc. Vic.101, 77-87.
Mount, A. B. (1979). Natural regenerationprocesses in Tasmanian forests. Search 10,180-186.
Neyland, M. G. (1991). Relict rainforest ineastern Tasmania. Tasmanian NationalRainforest Conservation Program TechnicalReport No. 6.
Noble, I. R. and Slatyer, R. O. (1980). The use ofvital attributes to predict successionalchanges in plant communities subject torecurrent disturbance. Vegetatio 43, 5-21.
Noble, I. R. and Slatyer, R. O. (1981). Conceptsand models of succession. In: A. M. Gill, R.H. Groves and I. R. Noble (eds) Fire and theAustralian Biota. Pp. 311-335. AustralianAcademy of Science, Canberra.
Noble, I. R., Moore, A. D. and Strasser, M. J.(1988). Stand structure and the prediction ofvegetation dynamics. In: H. J. During, M. J.A. Werger and J. H. Willems (eds) Diversityand Pattern in Plant Communities. Pp. 263-275. SPB Academic Publishing, The Hague,The Netherlands.
Noble, J. C. (1989). Fire studies in mallee(Eucalyptus spp.) communities of westernNew South Wales: The effects of firesapplied in different seasons on herbageproductivity and their implications formanagement. Aust. J. Ecol. 14, 169-187.
Palmer, M. W. and White, P. S. (1994). Scaledependence and the species-arearelationship. Amer. Nat. 144, 717-740.
Patterson, B. D. and Atmar, W. (1986). Nestedsubsets and the structure of insularmammalian faunas and archipelagos. Biol. J.Linn. Soc. 28, 65-82.
Pimm, S. L. (1982). Food Webs. Chapman andHall, London.
52
Australia’s Biodiveristy - Responses to Fire

Preston, F. W. (1960). Time and space and thevariation of species. Ecology 41, 611-627.
Preston, F. W. (1962a,b). The canonicaldistribution of commonness and rarity.Ecology 43, 185-215, 410-432.
Purdie, R. W. (1977). Early stages of regenerationafter burning in dry sclerophyll vegetation. I.Regeneration of the understorey byvegetative means. Aust. J. Bot. 25, 21-34.
Ridley, W. F. and Gardner, A. (1961). Fires inrainforest. Aust. J. Sci. 23, 227-228.
Russell, R. P. and Parsons, R. F. (1978). Effects oftime since fire on heath floristics at Wilson’sPromontory, southern Australia. Aust. J. Bot.26, 53-61.
Russell-Smith, J. and Bowman, D. M. J. S. (1992).Conservation of monsoon rainforest isolatesin the Northern Territory, Australia. Biol.Conserv. 59, 51-63.
Rydin, H. and Borgegard, S. O. (1988). Plantspecies richness on islands over a century ofplant succession: Lake Hjalmaren. Ecol. 69,916-927.
Specht, R. L. and Specht, A. (1989). Speciesrichness of sclerophyll (heathy) plantcommunities in Australia - the influence ofoverstorey cover. Aust. J. Bot. 37, 337-350.
Specht, R. L., Rayson, P., and Jackman, M. E.(1958). Dark Island heath (Ninety MilePlain, South Australia). VI. Pyric succession:changes in composition, coverage, dryweight and mineral nutrient status. Aust. J.Bot. 6 , 59-88.
Stocker, G. C. and Mott, J. J. (1981). Fire in thetropical forests and woodlands of northernAustralia. In: A. M. Gill, R. H.Groves and I.R. Noble (eds) Fire and the Australian Biota.Pp. 425-439. Australian Academy of Science,Canberra.
Stocker, G. C. and Sturtz, J. D. (1966). Use of fireto establish Townsville lucerne in theNorthern Territory. Aust. J. Expt. Agric. &Anim. Husb. 6, 277-279.
Taylor, J.A. and Dunlop, C.R. (1985). Plantcommunities of the wet-dry tropics ofAustralia: the Alligator Rivers region,Northern Territory. Proc. Ecol. Soc. Aust. 13,83-127.
Tolhurst, K. and Flinn, D. (1992). (eds) EcologicalImpacts of Fuel Reduction Burning in DrySclerophyll Forest: First Progress Report.Victorian Department of Conservation andEnvironment, Research Report 349.
Tolhurst, K. G. and Oswin, D. A. (1992). Effectsof spring and autumn low intensity fire onunderstorey vegetation in open eucalyptforest in west-central Victoria. VictorianDepartment of Conservation andEnvironment, Research Report 349. Pp. 3.1to 3.60.
Unwin, G. L. (1989). Structure and compositionof the abrupt rainforest boundary in theHerberton Highland, North Queensland.Aust. J. Bot. 37, 413-428.
Van der Moezel, P. G. and Bell, D. T. (1984). Firein the Western Australian mallee. In: B. Dell(ed) MEDECOS IV, Proceedings 4thInternational Conference on MediterraneanEcosystems. Pp. 151-152.
Venning, J. (1978). Post-fire responses of aEucalyptus baxteri woodland near Penola inSouth Australia. Aust. For., 41(4), 192-206.
Wark, M. C. (1997). Regeneration of some forestand gully communities in the Angahook-Lorne State Park (north-eastern OtwayRanges) 1-10 years after the wildfire ofFebruary 1983. Proc. Roy. Soc. Vict. 109, 7-36.
Wark, M. C., White, M. D., Robertson, D. J. andMarriott, P. F. (1987). Regeneration of heathand heath woodland in the north-easternOtway Ranges following the wildfire ofFebruary 1983. Proc. Roy. Soc. Vict. 99, 51-88.
Whittaker, R. H. (1965). Dominance and diversityin land plant communities. Science 147, 250-260.
Whittaker, R. H. (1972). Evolution andmeasurement of species diversity. Taxon 21,213-251.
Wilson, J. B., Gitay, H., Roxburgh, S. H., King,W. M. and Tangney, R. S. (1992). Egler’sconcept of ‘initial floristic composition’ insuccession - ecologists citing it don’t agreewhat it means. Oikos 64, 591-593.
Wood, J. G. (1937). The Vegetation of SouthAustralia. Government Printer, Adelaide.
Zimmer, W. J. (1940). Plant invasions in themallee. Vic. Natur. 56, 143-147.
53
Biodiveristy and bushfires


Biodiveristy and bushfires
FIRE AND AUSTRALIAN BIRDS:A REVIEW
J.C.Z. Woinarski
Parks and Wildlife Commission of the Northern TerritoryPO Box 496
Palmerston, NT, 0831

56
Australia’s Biodiveristy - Responses to Fire

57
Fire and Australian Brids
The literature concerning the impacts of fire onAustralian birds is reviewed. Fire effects areconsidered for main Australian environments.The methodology of studies is reviewed.
Detrimental fire regimes contributed to theextinction of two of the three bird species, andthree of the four subspecies which havedisappeared from Australia since Europeancolonisation. Inappropriate fire management isnow a factor in the threatened status of at least 51nationally recognised threatened bird taxa (secondin importance only to habitat clearance andfragmentation, which threatens 52 taxa). In manyenvironments (notably heath and mallee),inappropriate fire regime is now the main threatto most declining bird species.
Despite this recognition of the seriousness offire regime as a process threatening Australianbirds, for only a few species has detailed researchon the relationship between birds and fire regimesbeen conducted, and only in exceptional cases hasthis been translated into management. For manyenvironments, there is no information on theimpacts of fire regimes on birds.
Of the threatened species whose relationshipswith fire regime has been comparatively welldocumented, almost all show clear preference formuch less frequent fire than that currentlyprevailing. The long-unburnt vegetation favouredby these species is becoming disappearingly rare,and will require concerted management effort tomaintain or increase. Most fire-sensitivethreatened birds have low reproductive output andlimited dispersal ability. The persistence of thesespecies is further jeopardised by habitatfragmentation, which accentuates the handicap ofthese traits for recolonisation following fire.
In temperate eucalypt forests, controlburning is widely used to reduce the probability ofextensive wildfire. While the impact of a singlecontrol burn upon birds is generally less than thatof wildfire, there are few data on the long-termimpacts of a sustained regime of control burning.The most detailed long-term study suggests thatsuch frequent mild fires will lead to the declineand loss of some species which are now perceivedas common and little affected by mild fires.
SUMMARY

58
Australia’s Biodiveristy - Responses to Fire

59
Fire and Australian Brids
Fire has long been recognised as one of the majorfactors moulding vegetation patterning acrossAustralia (e.g. Mitchell 1838; Jackson 1968).Manipulation of fire, either explicitly or bydefault, is the main landscape management tool inmany environments and most conservationreserves. Inappropriate fire regime is the mainthreat to many vulnerable and endangeredAustralian birds (Garnett 1992a) and other biota(e.g. plants: Leigh et al. 1984). Yet knowledge ofthe history and effects of fire is negligible in manyenvironments, and there have been remarkablyfew long-term studies of the ecological impacts ofa range of fire regimes, such that the long-termconsequences of fire management are usually verypoorly known.
In this essay, I seek to review therelationships between Australian birds and fire.Previous reviews (e.g. Cowley et al. 1969;Christensen & Kimber 1975; Catling & Newsome1981; Recher 1981; Recher & Christensen 1981;Meredith 1983,1988; Suckling & MacFarlane1983; Christensen et al. 1985; Smith 1987a;Christensen & Abbott 1989; McFarland 1993;Silveira 1993; Arnold et al. 1993; Crowley 1994)have summarised and interpreted studies of fireimpacts on birds for broad regions orenvironments within Australia, and this nationaloverview owes much to these predecessors. Thequantity of these reviews is surprisingly out ofproportion to the very limited number of detailedprimary studies, and the attempt here at a nationaloverview is the only justification for adding yetanother review to this unbalanced literature.
Many of the impacts of fire on Australianenvironments indirectly affect birds, for exampleby change in nutrient availability, food resourcesor floristics. References on these subjects arerelevant to birds even though birds may not beexplicitly mentioned. However, a complete reviewof the environmental impacts of fire is beyond thescope of this essay, and would be redundantanyway given the excellent recent comprehensivereviews by Gill et al. (1981), Pyne (1991),Williams & Gill (1995), and Whelan (1995), andtwo recent broad collections of papers (McCaw etal. 1995; DEST 1996). Rather, I attempt here to:
(i) interpret, in terms of consequences forbirds, the literature which describes
environmental impacts of fires, but which doesn’texplicitly mention birds; and
(ii) review, as comprehensively as possible,the literature explicitly concerned with fire andAustralian birds.
An annotated bibliography of the relevantliterature is compiled as a foundation for the lattergoal.
Interpreting the literature is rendereddifficult by the variation in fires within andbetween environments. Individual fires within agiven environment vary in seasonal timing, extent,intensity, patchiness, and the temporal pattern oftheir (re-) occurrence. Their impacts at aparticular site are influenced by these diversefactors (and also by landscape context and pre-and post-fire climatic conditions), and differentbird species may respond very idiosyncratically tothis variation. The search for general pattern inresponse is further hampered by the very variableand limited research effort, with few long-termstudies and little experimentation with a range offire treatments. In this regard, it is notable thatsuccessional responses of Australian mammalshave been much more thoroughly documentedand subject to far more intensive experimentalinvestigation (e.g. Suckling & MacFarlane 1983;Higgs & Fox 1993).
This document contains a general overviewof fire history, an assessment of fire impacts onbirds across all main Australian environments, areview of the methodological approaches used inthe investigation of impacts, and a concludingsection reviewing fire management and birdconservation.
INTRODUCTION

60
Australia’s Biodiveristy - Responses to Fire

61
Fire and Australian Brids
Evidence from pollen cores and plant fossilmaterial has demonstrated substantial re-arrangement of Australian vegetation since at leastthe Tertiary (Hill 1994), notably including ageneral increase in the extent of Eucalyptus openforests and woodlands at the expense of closedforests. Associated with these broad-scaleenvironmental changes, there has been a substantialextinction of Australian fauna, notably rainforestand megafaunal elements, and including manyorders and families of birds (Rich 1991).
Environmental change accelerated during therapid climatic fluctuations of the late Pleistocene(Singh & Geissler 1985; Kershaw 1986), and thiswas accompanied by extinctions and range changesfor much of the fauna. This period saw the end ofthe line for the massive flightless Mihirungs(Dromornithidae), the disappearance of flamingoes(Phoenicopteridae) from Australia, the extinction oflarge species of coucal and megapodes, themainland loss of the flightless Tasmanian NativeHen1 and further range contractions for groupsassociated with closed forests (e.g. the logrunnersOrthonychidae) (Baird 1991). This change mayhave been compounded (or precipitated: Merilees1968; Smith 1977; Flannery 1990, 1994) by theentry of Aboriginal people to Australia, betweenabout 60–100,000 ybp, as Aboriginal use of firealmost certainly led to a fire regime different fromthat previously prevalent (Jones 1969; Singh et al.1981; Nicholson 1981; Braithwaite & Estbergs1985), and consequently re-shaped the Australianlandscape. The extent to which Aboriginal landmanagement was responsible for environmentalmodification remains unclear and contested (e.g.Williams & Gill 1995).
Any sustained change in burning regimeswill benefit some components of the ecosystembut disadvantage others. Evidence from earlyEuropean accounts of firing by Aborigines (e.g.Hallam 1975; Braithwaite 1991; Jones 1995),current accounts of reasonably traditionalAboriginal land management (Haynes 1985, 1991;Lucas & Lucas 1993; Baker et al. 1993; Bradley1995) and interpretation of vegetation patterning(e.g. Jackson 1968; Price & Bowman 1994)
suggests that Aboriginal people generally burnttheir lands frequently and purposefully. Mostresultant fires appear to have been relatively cooland of limited extent. Hence a fine scale mosaic ofvegetation age was maintained, although frequentburning may have favoured early successionalspecies, and savannas and eucalypts generally.Heaths may also have expanded under Aboriginalfire regimes (Jackson 1968; Smith 1977), althoughshrubbiness under forests and woodlands wasprobably much diminished (Gill 1981; Recher et al. 1993; Stanton 1995).
Late successional species (and fire-sensitivecommunities) may have been retained within themanaged landscape by the deliberate exclusion offire from fire-sensitive areas which supportedparticular resources (e.g. Jones 1995; Lucas &Lucas 1993) or which were otherwise of culturalsignificance (Latz 1995). Areas (or environments)which were unsuitable for habitation or supportedrelatively low densities of Aboriginal residents(perhaps such as much of the mallee: Harris 1990)may have avoided the imposition of Aboriginalfire regimes.
Nonetheless, the flammability of much ofAustralian vegetation (and recurrent drought)must have ensured that there were episodes ofcatastrophic and extensive wildfires, even underAboriginal stewardship. The existence of cohortsof Mountain Ash Eucalyptus regnans pre-datingEuropean colonisation (Gill 1981) is onedemonstration of the extensive, if infrequent,occurrence of such wildfire.
The impacts upon birds of Aboriginalburning regimes were probably very disparate.Directly, Aboriginal people probably used fire asan aid in hunting flightless birds, as they did forhunting macropods and other mammals(Braithwaite 1991). It is possible that theextinction or range contraction of several speciesof flightless birds in the late Pleistocene was dueto Aboriginal hunting (aided by fire), thoughmany other factors may have been involved (e.g.Baird 1984). Aboriginal hunters also used smoketo attract and trap raptors, and the remnants ofsuch traps are still visible in parts of northernAustralia (Boekel 1980).
An increase in the frequency andpredictability of fires, following Aboriginal entry
HISTORY OF FIRE REGIMES
1 Scientific names of all birds mentioned in text or tablesare given in Table 1.

to Australia, probably would have led to increasedabundance of birds which forage around fires (e.g.raptors, wood-swallows: Braithwaite & Estbergs1987; Woinarski 1990) or in recently-burnt areas(e.g. granivores). Such fire-associated speciesprobably could not maintain large populationsunder natural fire regimes of infrequent andunpredictable fires.
However, for birds, the most substantialimpacts of Aboriginal fire regimes were probablyexperienced through resultant vegetation change.An increase in fire frequency would have led tochange in floristics, vegetation structure and grainsize of vegetation mosaics. Fire-sensitivevegetation (and its associated bird fauna) wouldhave declined, except where this provideddesirable resources and could be protected.Probable losers from this re-casting of Australianenvironments included species associated withconifer forests, with Casuarina or Allocasuarinawoodlands (e.g. Glossy Black-cockatoo: Clout1989), with closed forests, or with dense shrubbyunderstoreys below Eucalyptus forests (e.g. scrub-birds, bristlebirds). Probable winners includedspecies associated with the relatively sparsevegetation of early seral stages (e.g. Richard’sPipit), and those associated with eucalypt open
forests and savanna woodlands (e.g. manygranivorous finches, doves and parrots;honeyeaters). Nonetheless, the relatively fine scaleof burning patterns for lands under Aboriginalmanagement probably maintained relatively highlevels of heterogeneity of seral stages andenvironments generally, and hence supported highdiversity of bird species.
Rapidly following European usurpation ofland management across much of Australia, thepractice and purpose of burning changed.Destructive and extensive burns were used to aidclearing or to demonstrate proprietorship (Gill1981; Johnson & Purdie 1981). While somechanges of incidence (and impacts) weredeliberate, others involved a complex interplay ofnew factors. For example, the introduction andrapid spread of livestock and exotic plants changedfuel characteristics and understorey floristics,thereby constraining options for fire regimes.Other introductions, notably rabbits, interferedwith post-fire plant recruitment processes.Clearing and fragmentation of native vegetationchanged the scale and extent of fires. Morerecently, fire has been used as a tool forbiodiversity conservation (Good 1981; Garnett &Crowley 1994; Crowley 1995).
62
Australia’s Biodiveristy - Responses to Fire

For all main (loosely-defined) Australianenvironments, I consider (where possible) currentand pre-European fire regimes, bird speciesresponses to single fires and fire regimes,relationships of threatened taxa to fire regimes andfire management for bird conservation. Theimpacts of fire and potential regimes differappreciably between these environments. In someenvironments (notably temperate eucalypt openforests), there may be marked differences betweenmild control fires (which usually affect only theunderstorey) and wildfire. This distinction is lessclearcut in most heaths and mallee communities,where most vegetation occurs near the ground, andthe response of birds to control burns may besimilar to those following wildfire (Meredith 1983).
COASTAL HEATHS AND THICKETS
Fire regimes
In southwestern Australia, where previouslyAboriginal people had burnt coastal heath countrywith small cool fires at intervals of 5-10 years, earlycolonists torched the heaths at least once every 2–3 years until grasslands suitable for pastoralismreplaced the native bush (Hallam 1975; Smith1977,1987a). Carter (1924) provided a graphiceyewitness account of these changes:
“where there had been dense impenetrablescrub, was mostly bare sand drifts caused byfire made to improve the country for cattlegrazing”.By contrast, in southwestern Tasmania,
Jones (1995) noted that dense thickets and heathsoccurred now in areas where observers in 1829 hadrecorded open sedgelands, a change attributed tothe removal of frequent fires lit by Aboriginalpeople. This disparity in change reflects both thedifficulty of generalising about fire regimes andtheir effects, and the limited data on, and frailty ofinterpretations of, pre-European fire regimes.
In coastal heaths of southeastern and easternAustralia, European fire management was initiallyindiscriminate, unless where directed toconversion of heaths to farmlands. More recently(notably since the 1950s: Luke & McArthur1978), most heathlands have been managed byhigh frequency (often with prescribed intervals of
<5 years) control burning. Such frequent burninghas led to the decline and/or local extinction ofseveral heathland-dependent birds, most notablyGround Parrot and Southern Emu-wren. Overthe last two decades, largely in response toresearch on the requirements of such species,control burning in heathlands has been reduced.In some areas (notably on conservation reserves),fire exclusion or suppression has beenrecommended (e.g. Cooper 1974) and practised(Meredith et al. 1984). Even in such protectedheaths, occasional wildfires have proven difficultto exclude, and have burnt some reserved heathsextensively (notably including the 1994 firethrough heathlands at Royal National Park).
Species responses to single fires
Relative to most other Australian environments,the impacts of fire on birds in heaths has been welldocumented (Table 2). Very high mortality rateshave been reported for heathland birds duringwildfires (Recher et al. 1975; Fox 1978; Pescott1983; Wegener 1984a,b), though Main (1981)suggested that many birds were able to escapesuch fire. Hawking insectivores (e.g. swallows) andsome raptors may be attracted to fire fronts (Main1981).
In days to months following fire in heaths,the dead or exposed invertebrate and vertebrateprey attracts many predator visitors from adjacenthabitats (e.g. Straw-necked Ibis, Torresian Crow,Laughing Kookaburra, raptors, egrets, PiedCurrawong, Australian Magpie, Australian Raven,Magpie-lark), and seed shed by plants in responseto fire (e.g. Banksia spp.) attracts parrots andcockatoos (Roberts 1970; McFarland 1988,1993).
While regrowth vegetation remains relativelyopen and herb and grass species are relativelyabundant (up to about 3 years post-fire), the area iscolonised by open-country species such asRichard’s Pipit, Brown Quail, Little Button-quail,Elegant Parrot, swallows and martins (Smith1987a; McFarland 1988; Brooker & Rowley 1991;Hopkins & Smith 1996) (Fig. 1). In some heaths,fire may trigger unusually abundant flowering forsome plants (notably Xanthorrhoea), and somehoneyeater and parrot species may respond withincreased abundance (Specht 1981; McFarland1993). Insect abundance may increase rapidly after
63
Fire and Australian Brids
FIRE REGIMES AND THEIR IMPACTS ON BIRDS: A REVIEW ACROSS MAIN ENVIRONMENTS
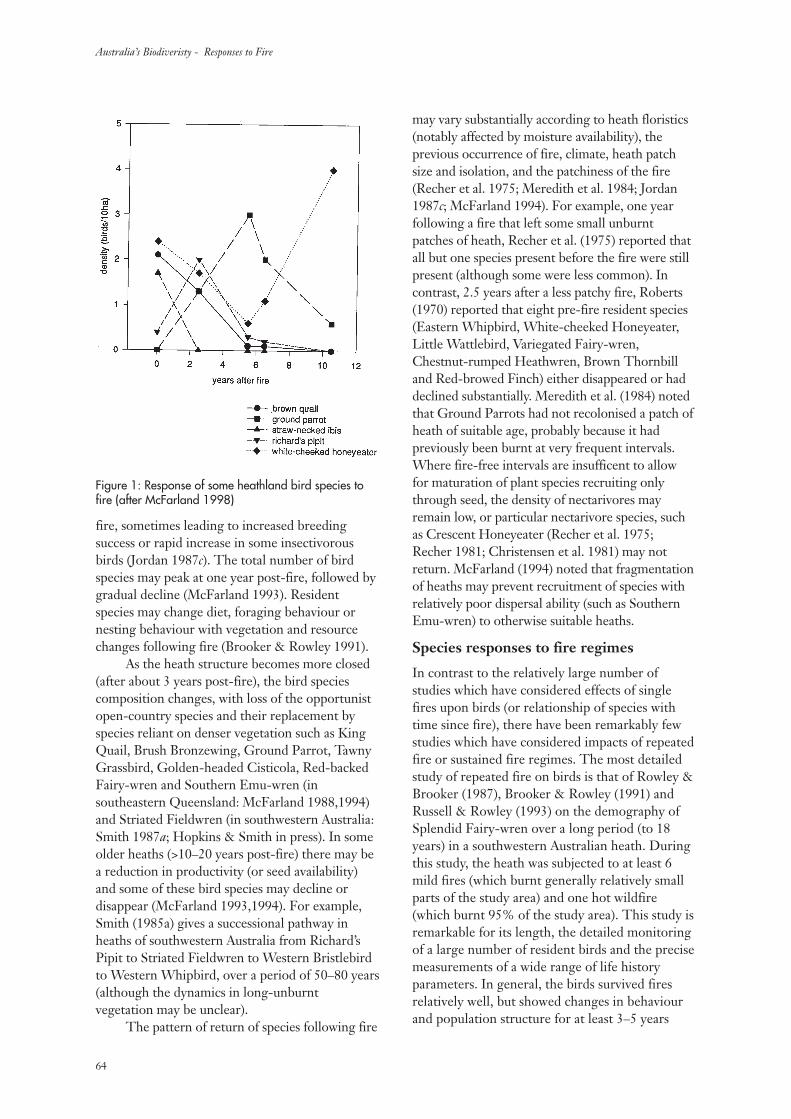
fire, sometimes leading to increased breedingsuccess or rapid increase in some insectivorousbirds (Jordan 1987c). The total number of birdspecies may peak at one year post-fire, followed bygradual decline (McFarland 1993). Residentspecies may change diet, foraging behaviour ornesting behaviour with vegetation and resourcechanges following fire (Brooker & Rowley 1991).
As the heath structure becomes more closed(after about 3 years post-fire), the bird speciescomposition changes, with loss of the opportunistopen-country species and their replacement byspecies reliant on denser vegetation such as KingQuail, Brush Bronzewing, Ground Parrot, TawnyGrassbird, Golden-headed Cisticola, Red-backedFairy-wren and Southern Emu-wren (insoutheastern Queensland: McFarland 1988,1994)and Striated Fieldwren (in southwestern Australia:Smith 1987a; Hopkins & Smith in press). In someolder heaths (>10–20 years post-fire) there may bea reduction in productivity (or seed availability)and some of these bird species may decline ordisappear (McFarland 1993,1994). For example,Smith (1985a) gives a successional pathway inheaths of southwestern Australia from Richard’sPipit to Striated Fieldwren to Western Bristlebirdto Western Whipbird, over a period of 50–80 years(although the dynamics in long-unburntvegetation may be unclear).
The pattern of return of species following fire
may vary substantially according to heath floristics(notably affected by moisture availability), theprevious occurrence of fire, climate, heath patchsize and isolation, and the patchiness of the fire(Recher et al. 1975; Meredith et al. 1984; Jordan1987c; McFarland 1994). For example, one yearfollowing a fire that left some small unburntpatches of heath, Recher et al. (1975) reported thatall but one species present before the fire were stillpresent (although some were less common). Incontrast, 2.5 years after a less patchy fire, Roberts(1970) reported that eight pre-fire resident species(Eastern Whipbird, White-cheeked Honeyeater,Little Wattlebird, Variegated Fairy-wren,Chestnut-rumped Heathwren, Brown Thornbilland Red-browed Finch) either disappeared or haddeclined substantially. Meredith et al. (1984) notedthat Ground Parrots had not recolonised a patch ofheath of suitable age, probably because it hadpreviously been burnt at very frequent intervals.Where fire-free intervals are insufficent to allowfor maturation of plant species recruiting onlythrough seed, the density of nectarivores mayremain low, or particular nectarivore species, suchas Crescent Honeyeater (Recher et al. 1975;Recher 1981; Christensen et al. 1981) may notreturn. McFarland (1994) noted that fragmentationof heaths may prevent recruitment of species withrelatively poor dispersal ability (such as SouthernEmu-wren) to otherwise suitable heaths.
Species responses to fire regimes
In contrast to the relatively large number ofstudies which have considered effects of singlefires upon birds (or relationship of species withtime since fire), there have been remarkably fewstudies which have considered impacts of repeatedfire or sustained fire regimes. The most detailedstudy of repeated fire on birds is that of Rowley &Brooker (1987), Brooker & Rowley (1991) andRussell & Rowley (1993) on the demography ofSplendid Fairy-wren over a long period (to 18years) in a southwestern Australian heath. Duringthis study, the heath was subjected to at least 6mild fires (which burnt generally relatively smallparts of the study area) and one hot wildfire(which burnt 95% of the study area). This study isremarkable for its length, the detailed monitoringof a large number of resident birds and the precisemeasurements of a wide range of life historyparameters. In general, the birds survived firesrelatively well, but showed changes in behaviourand population structure for at least 3–5 years
64
Australia’s Biodiveristy - Responses to Fire
Figure 1: Response of some heathland bird species tofire (after McFarland 1998)
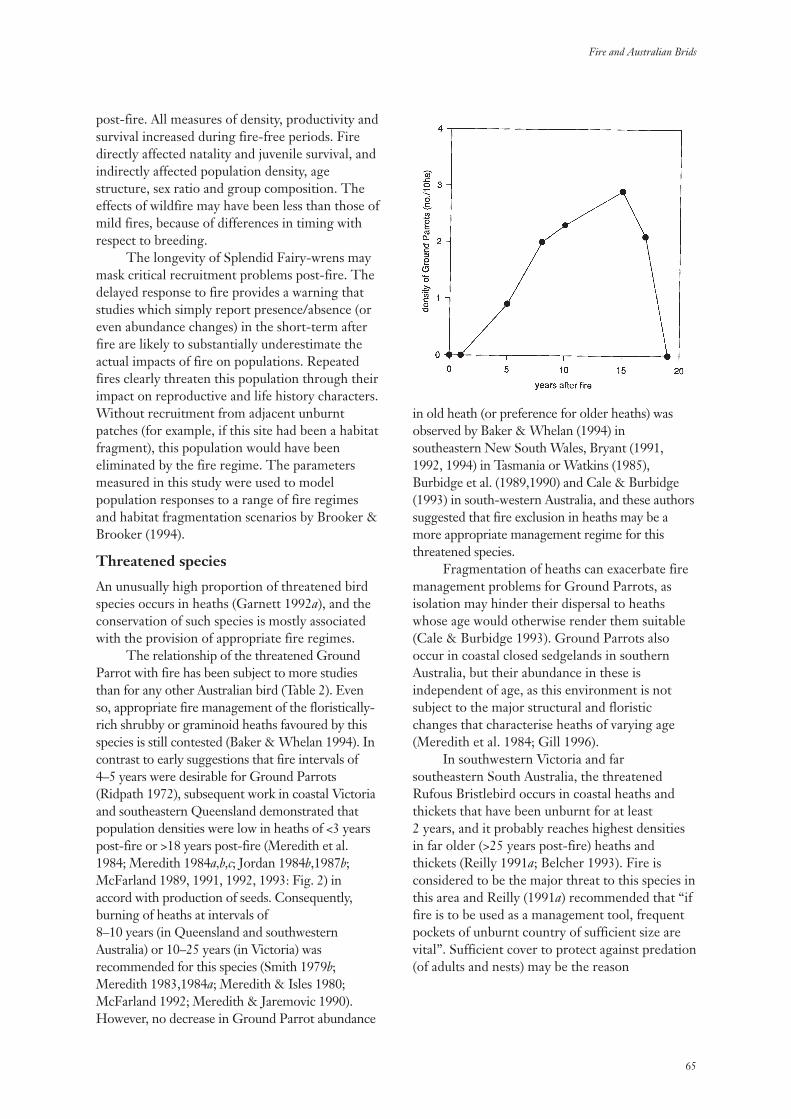
post-fire. All measures of density, productivity andsurvival increased during fire-free periods. Firedirectly affected natality and juvenile survival, andindirectly affected population density, agestructure, sex ratio and group composition. Theeffects of wildfire may have been less than those ofmild fires, because of differences in timing withrespect to breeding.
The longevity of Splendid Fairy-wrens maymask critical recruitment problems post-fire. Thedelayed response to fire provides a warning thatstudies which simply report presence/absence (oreven abundance changes) in the short-term afterfire are likely to substantially underestimate theactual impacts of fire on populations. Repeatedfires clearly threaten this population through theirimpact on reproductive and life history characters.Without recruitment from adjacent unburntpatches (for example, if this site had been a habitatfragment), this population would have beeneliminated by the fire regime. The parametersmeasured in this study were used to modelpopulation responses to a range of fire regimesand habitat fragmentation scenarios by Brooker &Brooker (1994).
Threatened species
An unusually high proportion of threatened birdspecies occurs in heaths (Garnett 1992a), and theconservation of such species is mostly associatedwith the provision of appropriate fire regimes.
The relationship of the threatened GroundParrot with fire has been subject to more studiesthan for any other Australian bird (Table 2). Evenso, appropriate fire management of the floristically-rich shrubby or graminoid heaths favoured by thisspecies is still contested (Baker & Whelan 1994). Incontrast to early suggestions that fire intervals of4–5 years were desirable for Ground Parrots(Ridpath 1972), subsequent work in coastal Victoriaand southeastern Queensland demonstrated thatpopulation densities were low in heaths of <3 yearspost-fire or >18 years post-fire (Meredith et al.1984; Meredith 1984a,b,c; Jordan 1984b,1987b;McFarland 1989, 1991, 1992, 1993: Fig. 2) inaccord with production of seeds. Consequently,burning of heaths at intervals of 8–10 years (in Queensland and southwesternAustralia) or 10–25 years (in Victoria) wasrecommended for this species (Smith 1979b;Meredith 1983,1984a; Meredith & Isles 1980;McFarland 1992; Meredith & Jaremovic 1990).However, no decrease in Ground Parrot abundance
in old heath (or preference for older heaths) wasobserved by Baker & Whelan (1994) insoutheastern New South Wales, Bryant (1991,1992, 1994) in Tasmania or Watkins (1985),Burbidge et al. (1989,1990) and Cale & Burbidge(1993) in south-western Australia, and these authorssuggested that fire exclusion in heaths may be amore appropriate management regime for thisthreatened species.
Fragmentation of heaths can exacerbate firemanagement problems for Ground Parrots, asisolation may hinder their dispersal to heathswhose age would otherwise render them suitable(Cale & Burbidge 1993). Ground Parrots alsooccur in coastal closed sedgelands in southernAustralia, but their abundance in these isindependent of age, as this environment is notsubject to the major structural and floristicchanges that characterise heaths of varying age(Meredith et al. 1984; Gill 1996).
In southwestern Victoria and farsoutheastern South Australia, the threatenedRufous Bristlebird occurs in coastal heaths andthickets that have been unburnt for at least2 years, and it probably reaches highest densitiesin far older (>25 years post-fire) heaths andthickets (Reilly 1991a; Belcher 1993). Fire isconsidered to be the major threat to this species inthis area and Reilly (1991a) recommended that “iffire is to be used as a management tool, frequentpockets of unburnt country of sufficient size arevital”. Sufficient cover to protect against predation(of adults and nests) may be the reason
65
Fire and Australian Brids

for absence from younger heaths for thisinsectivorous species. Predation of nests by FoxesVulpes vulpes is considered a main threat for somepopulations of Rufous Bristlebirds (Garnett1992b).
In heathland populations of the vulnerableEastern Bristlebird (in eastern New South Walesand southeastern Victoria), abundance increaseswith age since fire up to at least 9 years, but maythen plateau (Jordan 1984a; Bramwell et al. 1992;Pyke et al. 1995). Unburnt patches left after firemay be critical for recolonisation (Pyke et al. 1995).
The endangered Orange-bellied Parrot has acomplex relationship to seral stages of heath andbutton-grass at its breeding grounds in south-western Tasmania (Brown & Wilson 1981,1984). Ingeneral, it prefers to feed in relatively young heathsand sedgelands (<10 years post-fire), though itchanges feeding preferences for successional stagesduring different months of the breeding season (inresponse to variation in phenology). As its nestinghollows in adjacent open forests are vulnerable tofire, management of fire is critical, and shouldinvolve the maintenance of a range of fire histories.
In southwestern Australia, changed fireregimes and clearing have led to the precipitousdecline of four species occurring in heathlands,thickets or swamp/forest margins (Ground Parrot,Western Whipbird, Western Bristlebird and NoisyScrub-bird) and to the extinction of the WesternAustralian subspecies of Rufous Bristlebird D.broadbenti litoralis (Milligan 1904; Ashby 1921;Carter 1923a,b, 1924; Whittell 1936; Whitley1971; Smith 1977, 1985a, 1987a; How et al. 1987;Garnett 1992b; Cale & Burbidge 1993). Theendangered Western Bristlebird and Noisy Scrub-bird have persisted only in very localisedpopulations which have escaped frequent burningbecause of topographic protection from fire, orluck (Smith 1979a; Burbidge et al. 1986; Cale &Burbidge 1993). While the habitats of these threespecies differ somewhat (the Noisy Scrub-bird isnot really a heathland species), it is convenient toconsider them together because of their co-occurrence in a small number of sites in coastalsouthwestern Australia, and the detailed researchconducted at these sites on relationships betweenall three species and fire (Smith 1987a; Cale &Burbidge 1993; Hopkins & Smith 1996).
Western Bristlebirds require dry heath of atleast 6–10 years post-fire or wet heath of at least 3 years post-fire before habitat is suitable (Smith1977,1985a,1987b), and density generally declines
in very old heath (though some birds persist inheath >45 years post-fire: Cale & Burbidge 1993;Smith 1994), probably because of loweredproductvity (Smith 1985a,1987a,b).
For Noisy Scrub-birds, vegetation (thicketsand low scrubby eucalypt forests) has to be at least4–10 years post-fire (depending on floristics andlandscape position) before it becomes suitable:vegetation suitability also declines when very old(>30 years post-fire) (Smith & Robinson 1976;Smith 1977, 1979a, 1985a,b,c; Smith & Forrester1981) (Fig. 3).
Where Western Whipbirds occur in heaths,these have to be 4-7 years post-fire beforebecoming suitable (Smith 1985a), and the speciespersists in long-unburnt thickets (up to 50 yearspost-fire: Smith 1991).
For all three species, post-fire recruitment orrecolonisation is hampered by relatively poordispersal ability and/or low reproductive rates(Smith 1985a) and the very fragmented nature ofthe habitat (Smith 1979a; Cale & Burbidge 1993).Hence managers should aim to prevent entirehabitat patches from being burnt (Smith 1987a).Fire intervals of <10 years will lead to the localdecline and possible extinction of these three taxa.Fire intervals of >50 years may also be disadvan-tageous. The fire requirements of these three taxaare slightly different, such that an optimumregime for one species will not coincide exactlywith that of the others (Smith 1987a). Suitabilityof heaths post-fire can be substantially delayed bygrazing pressure by macropods (Smith 1985a;Cale & Burbidge 1993; Hopkins & Smith 1996).
66
Australia’s Biodiveristy - Responses to Fire
Figure 3: Abundance of male noisy scrub-birds afterfire (after Smith 1985c)
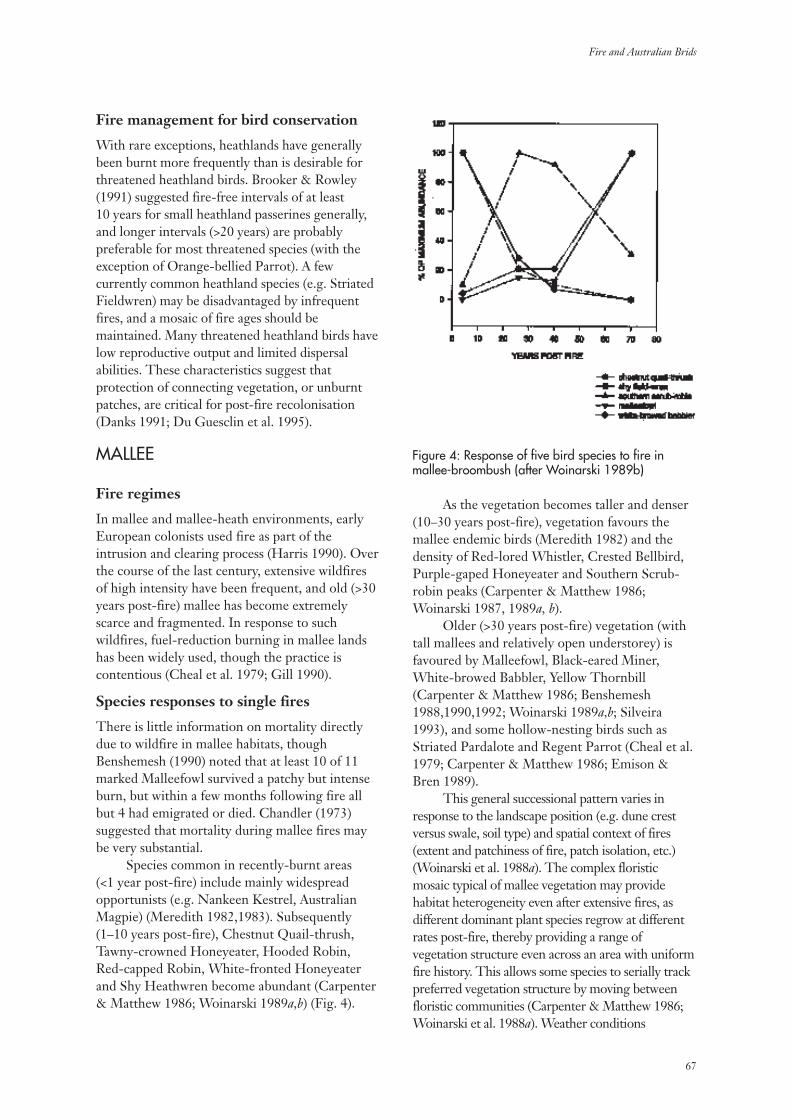
Fire management for bird conservation
With rare exceptions, heathlands have generallybeen burnt more frequently than is desirable forthreatened heathland birds. Brooker & Rowley(1991) suggested fire-free intervals of at least 10 years for small heathland passerines generally,and longer intervals (>20 years) are probablypreferable for most threatened species (with theexception of Orange-bellied Parrot). A fewcurrently common heathland species (e.g. StriatedFieldwren) may be disadvantaged by infrequentfires, and a mosaic of fire ages should bemaintained. Many threatened heathland birds havelow reproductive output and limited dispersalabilities. These characteristics suggest thatprotection of connecting vegetation, or unburntpatches, are critical for post-fire recolonisation(Danks 1991; Du Guesclin et al. 1995).
MALLEE
Fire regimes
In mallee and mallee-heath environments, earlyEuropean colonists used fire as part of theintrusion and clearing process (Harris 1990). Overthe course of the last century, extensive wildfiresof high intensity have been frequent, and old (>30years post-fire) mallee has become extremelyscarce and fragmented. In response to suchwildfires, fuel-reduction burning in mallee landshas been widely used, though the practice iscontentious (Cheal et al. 1979; Gill 1990).
Species responses to single fires
There is little information on mortality directlydue to wildfire in mallee habitats, thoughBenshemesh (1990) noted that at least 10 of 11marked Malleefowl survived a patchy but intenseburn, but within a few months following fire allbut 4 had emigrated or died. Chandler (1973)suggested that mortality during mallee fires maybe very substantial.
Species common in recently-burnt areas (<1 year post-fire) include mainly widespreadopportunists (e.g. Nankeen Kestrel, AustralianMagpie) (Meredith 1982,1983). Subsequently (1–10 years post-fire), Chestnut Quail-thrush,Tawny-crowned Honeyeater, Hooded Robin,Red-capped Robin, White-fronted Honeyeaterand Shy Heathwren become abundant (Carpenter& Matthew 1986; Woinarski 1989a,b) (Fig. 4).
As the vegetation becomes taller and denser(10–30 years post-fire), vegetation favours themallee endemic birds (Meredith 1982) and thedensity of Red-lored Whistler, Crested Bellbird,Purple-gaped Honeyeater and Southern Scrub-robin peaks (Carpenter & Matthew 1986;Woinarski 1987, 1989a, b).
Older (>30 years post-fire) vegetation (withtall mallees and relatively open understorey) isfavoured by Malleefowl, Black-eared Miner,White-browed Babbler, Yellow Thornbill(Carpenter & Matthew 1986; Benshemesh1988,1990,1992; Woinarski 1989a,b; Silveira1993), and some hollow-nesting birds such asStriated Pardalote and Regent Parrot (Cheal et al.1979; Carpenter & Matthew 1986; Emison &Bren 1989).
This general successional pattern varies inresponse to the landscape position (e.g. dune crestversus swale, soil type) and spatial context of fires(extent and patchiness of fire, patch isolation, etc.)(Woinarski et al. 1988a). The complex floristicmosaic typical of mallee vegetation may providehabitat heterogeneity even after extensive fires, asdifferent dominant plant species regrow at differentrates post-fire, thereby providing a range ofvegetation structure even across an area with uniformfire history. This allows some species to serially trackpreferred vegetation structure by moving betweenfloristic communities (Carpenter & Matthew 1986;Woinarski et al. 1988a). Weather conditions
67
Fire and Australian Brids
Figure 4: Response of five bird species to fire inmallee-broombush (after Woinarski 1989b)

following fire may also influence bird responses, withChandler (1973) noting that drought post-fire mayaccentuate the fire impact.
Species responses to fire regimes
There is no information on the responses of birdsto repeated fire, or particular fire regimes inmallee.
Threatened species
A relatively high proportion of threatened birds inAustralia occurs in mallee (Garnett 1992a). Fire isone of the most serious threatening processes forthreatened mallee birds (Stephens 1992; Silveira1993; Smith et al. 1994, 1996).
The vulnerable Malleefowl requires a denseand extensive layer of litter to provide material forincubation. This is not available until at least 10–15 years post-fire (Frith 1962; Tarr 1965), thenincreases in older vegetation. Optimum fire intervalfor this species is at least 60 years (Benshemesh1990). Fire intervals of 20 years have beenestimated to reduce Malleefowl densities to 6% ofthe maximum carrying capacity (Benshemesh1990). Young vegetation (<10 years post-fire) mayhave a greater density of food resources (notablyseeds) for this species, and a mosaic of fire ages maybe desirable (Carpenter & Matthew 1986; Brickhill1987; Priddell 1989,1990). Unburnt patches withinextensively-burnt vegetation appear critical forrecolonisation post-fire or persistence in a mainlyburnt environment (Benshemesh 1988,1990,1992).Another major threatening process, predation, mayhave increased impact in burnt country(Benshemesh 1992). Suitably old mallee hasbecome rare (Priddell 1990), and maintenance ofMalleefowl will require management to protectexisting old vegetation and to provide for additionallong-unburnt areas in the future (Benshemesh1990, 1992, 1994).
The endangered Black-eared Miner foragespreferentially in decorticating bark of very old(>50 year post-fire) mallee eucalypts (McLaughlin1992), though may occur in much younger malleewhere it adjoins older vegetation (Silveira1993,1995). The main threat to this taxon isgenetic swamping from the related Yellow-throated Miner, a process partly brought about byextensive mallee clearing (Garnett 1992b),however the limited area of old mallee is a factorin its threatened status (Starks 1987) andmaintenance of patches of old mallee may be animportant management requirement.
The vulnerable Mallee Emu-wren is closelyassociated with Triodia hummock grasses undermallee. Silveira (1993) noted that it was absent for5-6 years post-fire, and then persisted in long-unburnt vegetation. However, Garnett (1992b)reported the species bred at high densities within5 years of fire, and Emison et al. (1987)considered that it was common in youngregrowth. Low dispersal ability may limit itsrecolonisation to isolated patches or followingvery extensive non-patchy fires. The threatenedStriated Grasswren may also be severelydisadvantaged by intense and extensive fire(Brickhill 1980; Garnett 1992b; Silveira 1993).
The threatened mallee-heath subspecies ofSlender-billed Thornbill Acanthiza iredalei hedleyimay be vulnerable to frequent fire (Matthew 1994).
Limited data on the rare Scarlet-chestedParrot suggest that this species may preferrecently-burnt (3–5 years post-fire) mallee withhummock-grass understorey (Forshaw 1981;Robinson et al. 1990).
The vulnerable eastern subspecies of WesternWhipbird Psophodes nigrogularis leucogaster has beenrecorded from a wide range of post-fire ages, butappears to be most abundant in mallee vegetation10–25 years old in the Murray Mallee (Woinarskiet al. 1988a). The western mallee subspecies,P.n.oberon, has been recorded only from long-unburnt (>25 years) vegetation in southwesternAustralia (Smith 1985a,1991; McNee 1986).McNee (1986) recommended that management forthis species required long fire-free intervals (at least30 years and preferably 50 years).
The vulnerable Red-lored Whistler is mostabundant in mallee 5-30 years post-fire, and maynot persist in very old mallee (Woinarski 1987).
Threatened populations of Major Mitchell’sCockatoo and Regent Parrot may require malleewhich is sufficiently old (probably >50 years post-fire) to form suitable nesting hollows (Cheal et al.1979; Emison & Bren 1989; Silveira 1993).
Fire management for bird conservation
Overviews of the management of mallee birdshave recommended fire regimes which maintain amosaic of vegetation of a range of ages, but with abias towards retention of older ages (Cheal et al.1979; Emison & Bren 1989). Meredith (1982)noted that the goal of maintenance of a mosaic ofvegetation ages was not equivalent to, and unlikelyto be achieved by, unplanned random burns.
Fire management has been complicated by
68
Australia’s Biodiveristy - Responses to Fire

mallee fragmentation, especially in southwesternAustralian and western New South Wales.Individual fragments tend to each have ahomogeneous fire history, leading to reduced birdspecies diversity within fragments (Menkhorst &Bennett 1990; Priddell 1990), though theirisolation may offer some fire protection.
TEMPERATE EUCALYPT OPEN FORESTS
Fire regimes
There has been considerable debate about the fireregimes operating in (and understoreycharacteristics of) temperate eucalypt open forestsbefore European colonisation, and in the earlyyears of settlement (Gill 1981; Nicholson 1981;Williams & Gill 1995). The regimes and theirimpacts probably showed important differencesbetween forest types (Wakefield 1970), renderinggeneralisations difficult to draw.
In response to Aboriginal management, theeucalypt forests of temperate southeastern Australiawere probably generally characterised by low grassyunderstoreys (maintained by either frequent coolfiring or very rare fires: Benson 1981; Recher et al.1993). With exclusion of Aboriginal management,these forests rapidly developed shrubbyunderstoreys, probably markedly increasing fuellevels and hence increasing the probability of hotand extensive wildfire (Smith 1979a). While birdspecies diversity at any given point probablyincreased in response to greater structuralcomplexity of vegetation (Recher 1969), the loss offine-scale management probably led to increasedregional homogenisation of the environment (interms of floristic variation and seral ages) and hencereduced regional bird species diversity. Change inunderstorey structure would also havedisadvantaged birds reliant upon forests with grassyunderstorey (Recher et al. 1993), although to anextent some of these species may have been able tosubstitute the developing mosaic of farmlands andforests.
Frequent wildfires in temperate eucalyptforests were an inevitable consequence of limitedEuropean understanding of fire management inthese systems. More recently, in response to thehuman deaths and destruction of property inwildfire, fire suppression and/or frequent andextensive fuel-reduction (or control) burning havebeen implemented in most temperate eucalyptforests.
Species responses to single fires: wildfire
Bird mortality may be very high in severe wildfiresin temperate eucalypt forests (e.g. Hemsley 1967;Christensen et al. 1981; Suckling & MacFarlane1983; Wegener 1984a,b). Hood (1941) provided avivid eye-witness account of birds in a forestwildfire:
“As the terrific blast of fire swept along,birds could be seen rising from the groundor leaving the trees in front of it only to beoverpowered by the dense smoke and risingscorching heat. In all cases the birds fellexhausted into the flames”.Some bird species, notably aerial insectivores
(e.g. swifts: Lord 1936; McCulloch 1966) andraptors may follow fires, hunting disturbed insectsor vertebrates.
Where wildfire has killed the canopy trees,wholesale recomposition of the bird fauna mayfollow. Recently burnt areas initially have very fewbirds (Dedman 1983a,b,c,d,e), or at least asubstantial decline (20-65%: Ratkowsky1979,1985; Hewish 1983; Braithwaite et al. 1984;Loyn et al. 1992a) in species richness andabundance. However, even very small birds maysurvive hot wildfire (Rowley & Brooker 1987),especially where some patches remain unburnt(Smith 1989). Surviving individuals may adoptunusual behaviours or diet, for example SuperbFairy-wrens have been recorded feeding inscorched canopy (Dedman 1983e), and the diet ofSooty Owls was found to be much less variedpost-fire (Loyn et al. 1986).
Ground-feeding insectivores or carnivoresmay invade or remain in burnt areas, probably inresponse to the availability of dead or injured prey,or its greater accessibility (Hewish 1983). Speciessuch as Richard’s Pipit, Australian Magpie, ravens,Flame Robin, Scarlet Robin, LaughingKookaburra, Grey Butcherbird, Nankeen Kestreland Superb Fairy-wren colonise open burnt areasuntil regrowth becomes too dense (up to 3 years)(Christensen 1974; Stokes 1975; Chambers 1983;Braithwaite et al. 1984; Loyn 1985a,b; Reilly1991a,b). Granivorous birds (e.g. Red-browedFinch, Blue-winged Parrot, Diamond Dove,Common Bronzewing, Brown Quail, PaintedButton-quail) may also invade or become moreabundant in recently burnt areas (Recher et al.1985; Loyn 1985b).
In contrast, nectarivorous, frugivorous andfoliage-gleaning insectivorous birds (e.g. BrownThornbill, Striated Thornbill, Yellow-faced
69
Fire and Australian Brids
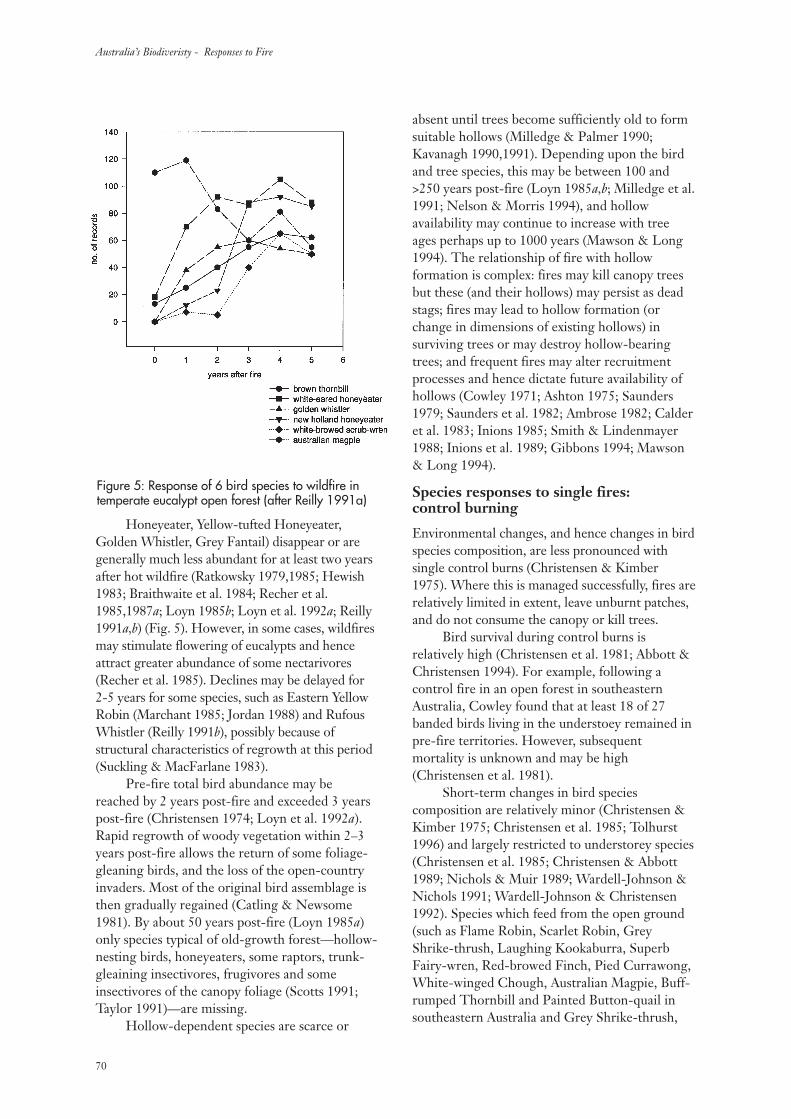
Honeyeater, Yellow-tufted Honeyeater,Golden Whistler, Grey Fantail) disappear or aregenerally much less abundant for at least two yearsafter hot wildfire (Ratkowsky 1979,1985; Hewish1983; Braithwaite et al. 1984; Recher et al.1985,1987a; Loyn 1985b; Loyn et al. 1992a; Reilly1991a,b) (Fig. 5). However, in some cases, wildfiresmay stimulate flowering of eucalypts and henceattract greater abundance of some nectarivores(Recher et al. 1985). Declines may be delayed for2-5 years for some species, such as Eastern YellowRobin (Marchant 1985; Jordan 1988) and RufousWhistler (Reilly 1991b), possibly because ofstructural characteristics of regrowth at this period(Suckling & MacFarlane 1983).
Pre-fire total bird abundance may bereached by 2 years post-fire and exceeded 3 yearspost-fire (Christensen 1974; Loyn et al. 1992a).Rapid regrowth of woody vegetation within 2–3years post-fire allows the return of some foliage-gleaning birds, and the loss of the open-countryinvaders. Most of the original bird assemblage isthen gradually regained (Catling & Newsome1981). By about 50 years post-fire (Loyn 1985a)only species typical of old-growth forest—hollow-nesting birds, honeyeaters, some raptors, trunk-gleaining insectivores, frugivores and someinsectivores of the canopy foliage (Scotts 1991;Taylor 1991)—are missing.
Hollow-dependent species are scarce or
absent until trees become sufficiently old to formsuitable hollows (Milledge & Palmer 1990;Kavanagh 1990,1991). Depending upon the birdand tree species, this may be between 100 and>250 years post-fire (Loyn 1985a,b; Milledge et al.1991; Nelson & Morris 1994), and hollowavailability may continue to increase with treeages perhaps up to 1000 years (Mawson & Long1994). The relationship of fire with hollowformation is complex: fires may kill canopy treesbut these (and their hollows) may persist as deadstags; fires may lead to hollow formation (orchange in dimensions of existing hollows) insurviving trees or may destroy hollow-bearingtrees; and frequent fires may alter recruitmentprocesses and hence dictate future availability ofhollows (Cowley 1971; Ashton 1975; Saunders1979; Saunders et al. 1982; Ambrose 1982; Calderet al. 1983; Inions 1985; Smith & Lindenmayer1988; Inions et al. 1989; Gibbons 1994; Mawson& Long 1994).
Species responses to single fires: control burning
Environmental changes, and hence changes in birdspecies composition, are less pronounced withsingle control burns (Christensen & Kimber1975). Where this is managed successfully, fires arerelatively limited in extent, leave unburnt patches,and do not consume the canopy or kill trees.
Bird survival during control burns isrelatively high (Christensen et al. 1981; Abbott &Christensen 1994). For example, following acontrol fire in an open forest in southeasternAustralia, Cowley found that at least 18 of 27banded birds living in the understoey remained inpre-fire territories. However, subsequentmortality is unknown and may be high(Christensen et al. 1981).
Short-term changes in bird speciescomposition are relatively minor (Christensen &Kimber 1975; Christensen et al. 1985; Tolhurst1996) and largely restricted to understorey species(Christensen et al. 1985; Christensen & Abbott1989; Nichols & Muir 1989; Wardell-Johnson &Nichols 1991; Wardell-Johnson & Christensen1992). Species which feed from the open ground(such as Flame Robin, Scarlet Robin, GreyShrike-thrush, Laughing Kookaburra, SuperbFairy-wren, Red-browed Finch, Pied Currawong,White-winged Chough, Australian Magpie, Buff-rumped Thornbill and Painted Button-quail insoutheastern Australia and Grey Shrike-thrush,
70
Australia’s Biodiveristy - Responses to Fire
Figure 5: Response of 6 bird species to wildfire intemperate eucalypt open forest (after Reilly 1991a)
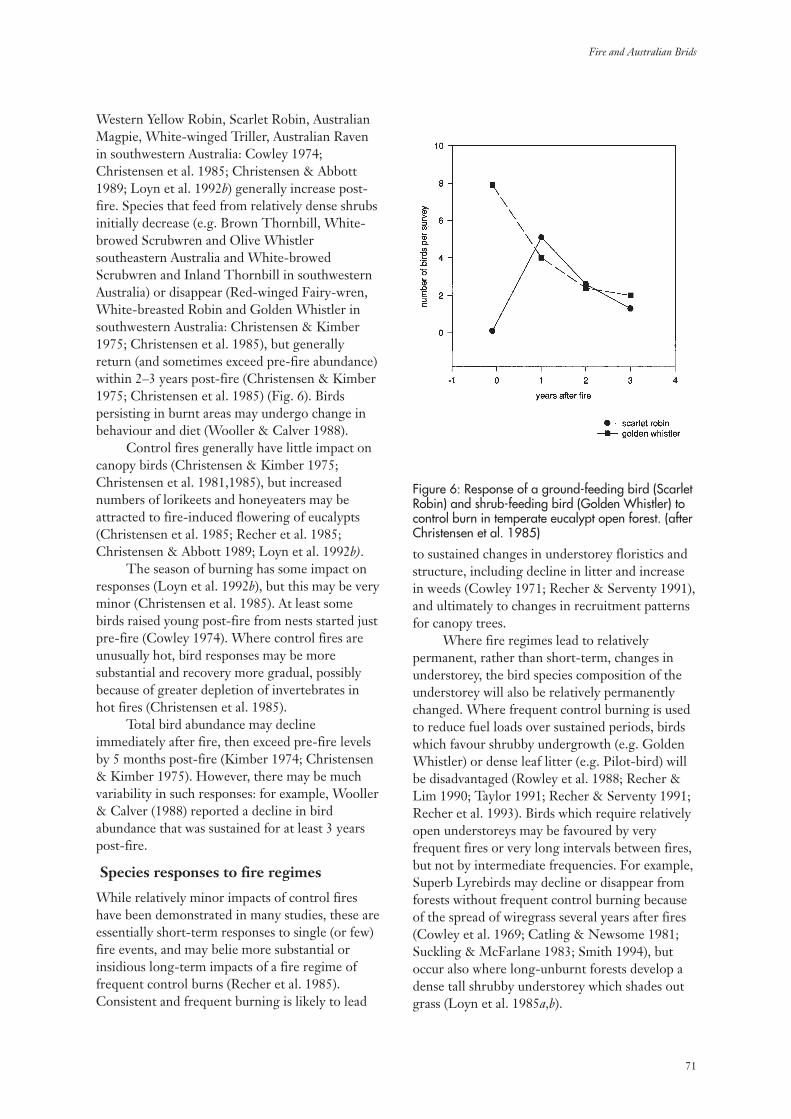
Western Yellow Robin, Scarlet Robin, AustralianMagpie, White-winged Triller, Australian Ravenin southwestern Australia: Cowley 1974;Christensen et al. 1985; Christensen & Abbott1989; Loyn et al. 1992b) generally increase post-fire. Species that feed from relatively dense shrubsinitially decrease (e.g. Brown Thornbill, White-browed Scrubwren and Olive Whistlersoutheastern Australia and White-browedScrubwren and Inland Thornbill in southwesternAustralia) or disappear (Red-winged Fairy-wren,White-breasted Robin and Golden Whistler insouthwestern Australia: Christensen & Kimber1975; Christensen et al. 1985), but generallyreturn (and sometimes exceed pre-fire abundance)within 2–3 years post-fire (Christensen & Kimber1975; Christensen et al. 1985) (Fig. 6). Birdspersisting in burnt areas may undergo change inbehaviour and diet (Wooller & Calver 1988).
Control fires generally have little impact oncanopy birds (Christensen & Kimber 1975;Christensen et al. 1981,1985), but increasednumbers of lorikeets and honeyeaters may beattracted to fire-induced flowering of eucalypts(Christensen et al. 1985; Recher et al. 1985;Christensen & Abbott 1989; Loyn et al. 1992b).
The season of burning has some impact onresponses (Loyn et al. 1992b), but this may be veryminor (Christensen et al. 1985). At least somebirds raised young post-fire from nests started justpre-fire (Cowley 1974). Where control fires areunusually hot, bird responses may be moresubstantial and recovery more gradual, possiblybecause of greater depletion of invertebrates inhot fires (Christensen et al. 1985).
Total bird abundance may declineimmediately after fire, then exceed pre-fire levelsby 5 months post-fire (Kimber 1974; Christensen& Kimber 1975). However, there may be muchvariability in such responses: for example, Wooller& Calver (1988) reported a decline in birdabundance that was sustained for at least 3 yearspost-fire.
Species responses to fire regimes
While relatively minor impacts of control fireshave been demonstrated in many studies, these areessentially short-term responses to single (or few)fire events, and may belie more substantial orinsidious long-term impacts of a fire regime offrequent control burns (Recher et al. 1985).Consistent and frequent burning is likely to lead
to sustained changes in understorey floristics andstructure, including decline in litter and increasein weeds (Cowley 1971; Recher & Serventy 1991),and ultimately to changes in recruitment patternsfor canopy trees.
Where fire regimes lead to relativelypermanent, rather than short-term, changes inunderstorey, the bird species composition of theunderstorey will also be relatively permanentlychanged. Where frequent control burning is usedto reduce fuel loads over sustained periods, birdswhich favour shrubby undergrowth (e.g. GoldenWhistler) or dense leaf litter (e.g. Pilot-bird) willbe disadvantaged (Rowley et al. 1988; Recher &Lim 1990; Taylor 1991; Recher & Serventy 1991;Recher et al. 1993). Birds which require relativelyopen understoreys may be favoured by veryfrequent fires or very long intervals between fires,but not by intermediate frequencies. For example,Superb Lyrebirds may decline or disappear fromforests without frequent control burning becauseof the spread of wiregrass several years after fires(Cowley et al. 1969; Catling & Newsome 1981;Suckling & McFarlane 1983; Smith 1994), butoccur also where long-unburnt forests develop adense tall shrubby understorey which shades outgrass (Loyn et al. 1985a,b).
71
Fire and Australian Brids
Figure 6: Response of a ground-feeding bird (ScarletRobin) and shrub-feeding bird (Golden Whistler) tocontrol burn in temperate eucalypt open forest. (afterChristensen et al. 1985)

Frequent burning may also lead to change inthe spatial patterning of eucalypt open forestsrelative to vegetation dominated by more fire-sensitive plant species (notably rainforest). Suchchange may be to the disadvantage of birds ofrainforests or their margins (Ferrier 1985; Holmes1988,1989).
While milder fires generally have less impactthan hot wildfires on invertebrates (Majer1984,1985; Christensen et al. 1985) and otherresources for birds, the cumulative impacts ofrepeated mild fires upon these resources, andhence upon birds, may be more severe (Friend1995), although the limited evidence is notconsistent (Springettt 1976; Abbott et al. 1984;Friend 1995).
Forestry
Much of the management of temperate eucalyptforests is associated with forestry. Firemanagement is a significant component of forestryactivities. The impacts upon birds of interactionsof fire and forestry practice (or comparisonsbetween the two) have been considered by Cowley(1971), McIlroy (1978), Wilson (1981), Green(1982), Tingay & Tingay (1984), Loyn (1985a),Recher et al. (1985,1987b), Dickinson et al. (1986),Recher (1991), Curry (1991), Milledge et al.(1991), Taylor (1991), Wardell-Johnson & Nichols(1991), Wardell-Johnson & Christensen (1992)and Abbott & Christensen (1994). In someeucalypt forests (notably E.regnans andE.delegatensis), hot fire following harvesting isessential for tree regrowth. Such fires mayexacerbate harvesting effects for species such asSpotted Quail-thrush (Wilson 1981), thoughMcIlroy (1978) claimed that post-harvest fires hadlittle impact on bird species. Piles of slashedvegetation left after harvesting may attract morebirds if left unburnt (Dickinson et al. 1986; Curry1991). Recher et al. (1987b) noted that the effectsof wildfire upon birds were more severe in smallretained (unlogged) strips than in larger strips orin larger unlogged areas. Regrowth forestsfollowing logging showed greater impact ofwildfire upon birds than did burnt unloggedforests (Recher et al. 1985), and the combinedeffects of logging and fire were more serious thaneither individually. Hollow availability may bemore affected by forestry activities than by fireregime (Lindenmayer et al. 1990).
Threatened species
A small proportion of the bird species intemperate open forests is threatened relative tothose in other habitats (Garnett 1992a). Forestryand clearing, rather than fire, are the primarythreatening process for most of these taxa. Mostof the threatened bird species occurring intemperate eucalypt open forests (e.g. Sooty Owl,Masked Owl, Powerful Owl, southern subspeciesof Red-tailed Black-cockatoo and Long-billedBlack-cockatoo) are associated with oldvegetation, usually because of their requirementfor hollows. The endangered Orange-belliedParrot nests in hollows in eucalypt open forestfringing heaths and button-grass plains insouthwestern Tasmania. Fire management iscritical for these species, to ensure protection ofexisting hollows and to allow the development ofsufficient areas of suitable old trees for the future.
The endangered Helmeted Honeyeater isrestricted to riparian eucalypt forests nearMelbourne. Two small isolated populationsdisappeared after recent wildfire (Backhouse 1987).While widlfire is a continuing threat to the onlyremaining population, fire prevention works maycreate more disturbance than fire (Backhouse 1987).
The endangered Forty-spotted Pardaloteoccurs in coastal eucalypt open forests inTasmania. Isolated populations may have beeneliminated by wildfire, but mild fires are probablynot detrimental to this canopy-feeding bird(Rounsevell & Woinarski 1983), although itsrequirement for hollows for breeding suggeststhat fire management may be required for itsmaintenance.
The rare Rufous Scrub-bird and northernpopulations of the vulnerable Eastern Bristlebirdinhabit ecotones between rainforests and eucalyptopen forests, and are threatened by too frequentfires (Ferrier 1985; Holmes 1988,1989), thoughboth may also be disadvantaged by rainforestexpansion associated with fire exclusion.
Fire management for bird conservation
A number of studies have recommended that firemanagement for bird conservation in eucalyptopen forests should involve the flexible use of abroad range of fire regimes, with specific attentionto threatened bird species (e.g. Recher 1981,1991;Wardell-Johnson et al. 1989). Until more isknown about the long-term effects of particularregimes, this is probably the most prudentstrategy.
72
Australia’s Biodiveristy - Responses to Fire

3.4 TEMPERATE WOODLANDS
Fire regimes
Little information is available on the pre-European fire regimes in temperate woodlands, orthe impacts of current regimes on birds of thishabitat. Hopkins (1985a) considered that fireswere probably infrequent in temperate woodlandsof southwestern Australia under Aboriginal landmanagement. He also noted that single fires inwoodlands could have dramatic effects onvegetation structure, that repeated burning couldcause permanent structural and floristic changesand that regeneration rates were generally veryslow. In contrast, Prober & Thiele (1993)considered that woodlands in a slightly higherrainfall area in southeastern Australia were burntevery year under Aboriginal land management,and this maintained a floristically rich grass/herbunderstorey dominated by Themeda.
Subsequent to European settlement, in bothsoutheastern and southwestern Australia,woodlands have been much affected by clearing,fragmentation, grazing, forestry operations andalteration of ecological processes (includingchanged fire regimes) (Saunders 1989; Saunders &Curry 1990; Bennett 1993; Robinson 1994; Yateset al. 1994). The age structure of most woodlandshas probably markedly changed since Europeancolonisation, with removal of most old trees onpublic lands and absence of regrowth on private(grazed) lands (Bennett 1993).
Species responses to single fires
Turner (1987,1992) considered short-term (8 months post-fire) and longer-term (8 yearspost-fire) responses of birds to a single wildfire ina mixed Callitris-Eucalyptus woodland insoutheastern Australia. In the short-term, birdabundance and richness was much reduced.Crested Pigeon, cuckoos, Rainbow Bee-eater,Speckled Warbler, White-browed Babbler,White-plumed Honeyeater, Little Friarbird,Yellow-faced Honeyeater, Chestnut-breastedMannikin, White-browed Wood-swallow, SuperbFairy-wren and Mistletoebird declined, whileEmu, White-winged Triller and Dusky Wood-swallow were more common in burnt vegetation.At 8 years post-fire, richness and abundance ofbirds was greater in the regrowth vegetation.Species more common in the burnt area includedGalah, Cockatiel, Turquoise Parrot, Rainbow
Bee-eater, Black-faced Cuckoo-shrike, White-winged Triller, Superb Fairy-wren, WesternGerygone, Rufous Songlark, Grey Fantail, BrownTreecreeper, Varied Sittella, Grey-frontedHoneyeater, White-naped Honeyeater, NoisyFriarbird and Dusky Wood-swallow. GoldenWhistler and Speckled Warbler were morecommon in the unburnt area.
Open-country bird species invaded Banksiawoodlands in southwestern Australia soon afterfire, but most species present before fire wereagain recorded within a few months after fire(Bamford 1985a,b). No species showed apreference for the long unburnt (22 years post-fire) sites.
Species responses to fire regimes
There is very limited information on response ofbirds to repeated fire, or particular fire regimes, intemperate woodlands. Adam & Robinson (1996)found greatly reduced density of Grey-crownedBabblers in roadside remnants that were annuallyburnt compared to unburnt strips, in associationwith greatly reduced shrub cover.
Threatened species
Woodland birds are declining across much ofsouthern Australia (Recher & Lim 1990;Robinson 1991). In many woodlands, hollow-nesting birds (and other fauna) are threatened bycurrent (and/or projected) limited supply ofhollows (Saunders et al. 1982; Traill 1993; Bennett1993), which may be influenced by fire regimes(Webster & Ahern 1992; Quin & Baker-Gabb1993). Birds which nest or forage on the groundare threatened by introduced predators andfloristic or vegetation structural changesassociated with grazing or changed fire regimes(Bennett 1993). A recent review of threatenedwoodland birds (Robinson 1994) did not listinappropriate fire regimes as a major threateningprocess, but little is known of the fire regimespreferred or required by such species, nor ofimpacts of the interaction between fire andgrazing. Management of fire may be a majorconcern if grazing is excluded from woodlandconservation reserves.
Casuarina or Allocasuarina may be a locallyprominent component of some woodlandcommunities and provides focal resources forsome bird species, notably the endangered GlossyBlack-cockatoo. A high frequency of intense firessince European settlement has led to decline in
73
Fire and Australian Brids

mature Casuarina, and hence of this dependentbird (for example, its extinction from King Islandis associated with extensive hot wildfires around1920: Green & McGarvie 1971). Its persistencerequires management to ensure fire exclusion, orlong intervals between intense fire (Joseph 1982;Clout 1989). For example, Joseph (1982) foundthat woodland areas 22 years post-fire were stillunsuitable for Glossy Black-cockatoos.
The extinction of the Paradise Parrot fromgrassy woodlands of inland northeastern NewSouth Wales and southeastern Queensland hasbeen linked to change in the fire regime followingEuropean settlement (Chisholm 1922,1945),although pastoralism, drought or weeds (singly, orin combination) may also have been involved(Forshaw 1981; Garnett 1992b).
Fire management for bird conservation
There is an urgent need for data to guide firemanagement for the conservation of temperatewoodland birds. Some fire management actionsfor Superb Parrot, Turquoise Parrot, Grey-crowned Babbler and Glossy Black-Cockatoo havebeen described (e.g. Davidson & Chambers 1991;Davidson & Robinson 1992; Webster & Ahern1992; Quin & Baker-Gabb 1993).
3.5 TROPICAL EUCALYPT OPENFORESTS AND SAVANNAWOODLANDS
Fire regimes
Fire management by Europeans has variedsubstantially across the extensive tropical eucalyptopen forests and savanna woodlands of northernAustralia. In the Top End and Kimberley, theincidence of hot extensive late Dry season fires hasincreased (Braithwaite & Estbergs 1985; Lewis1985). This increase has largely been due topastoral goals, but a high frequency of extensivelate Dry season fires occurs across all land tenures,including conservation reserves (Russell-Smith &Bowman 1992; Russell-Smith & Ryan 1994).
Fires in tropical open forests and savannawoodlands are generally relatively mild andimmediate impacts are often restricted to theunderstorey (Braithwaite & Estbergs 1985;Bowman 1988), superficially more similar tocontrol burning than wildfire in temperatesystems. Rapid build up of fuel and long dryseasons almost inevitably lead to a high frequencyof fires, hence generally denying successional
patterns comparable to those in temperate openforests (Bowman 1988).
Species responses to single fires
There may be very marked short-term response bybirds to fire in tropical eucalypt open forests andsavanna woodlands. Typically, the slow mild fireslead to little direct bird mortality (Braithwaite1985). Many hawking insectivores (e.g. wood-swallows, swifts) and raptors are attracted to fires.
From hours to months after fire, a widerange of ground-feeding birds (e.g. TorresianCrow, raptors, Pied Butcherbird, Straw-neckedIbis, Red-tailed Black-cockatoo, Little Corella,Galah, Blue-winged Kookaburra, Red-backedKingfisher, Magpie-lark) is attracted to burntareas to feed on resources made more accessibleby the removal of the dense grass layer (Crawford1972,1979; Beeton 1985; Braithwaite 1985;Braithwaite & Estbergs 1987; Press 1987;Woinarski 1990) (Fig. 7). Many of these speciesmay track fires or recently-burnt areas across thelandscape (Crawford 1972; Woinarski &Tidemann 1991,1992; Woinarski et al. 1992).This attraction may be reduced in intense late Dryseason fires, as the hot fires may destroy a farhigher proportion of seeds than do the coolerburns of the early dry season (Woinarski 1990).Individual bird species may also differ in theirshort-term responses to late and early Dry seasonfires (Woinarski 1990; Tidemann 1993b).
Few bird species of tropical open forests andwoodlands seem to be clearly disadvantaged in theshort-term by fire. Those that do, include theinsectivorous Golden-headed Cisticola and Red-backed Fairy-wren, which feed and shelter indense grass (Crawford 1972,1979; Woinarski 1990)and Purple-crowned Fairy-wren which requiresdense grass and pandanus (Rowley 1987,1993;Rowley & Russell 1993). Early dry season firesmay also destroy the nests and/or young of birdswhich nest on the ground or in grass tussocks(notably Partridge Pigeon and Masked Finch:Woinarski 1990; Lucas & Lucas 1993).
In tropical eucalypt open forests and savannawoodlands, hot fires late in the Dry season maydestroy hollow-bearing trees, to the detriment ofhollow-nesting birds, including Gouldian Finchand Palm Cockatoo (Young 1991; Tidemann1992; Stanton 1992,1995; Crowley 1995). Whilethis may be a serious threat for Palm Cockatoos,the smaller hollows suitable for Gouldian Finchappear to be relatively abundant and not limiting(Tidemann et al. 1992).
74
Australia’s Biodiveristy - Responses to Fire
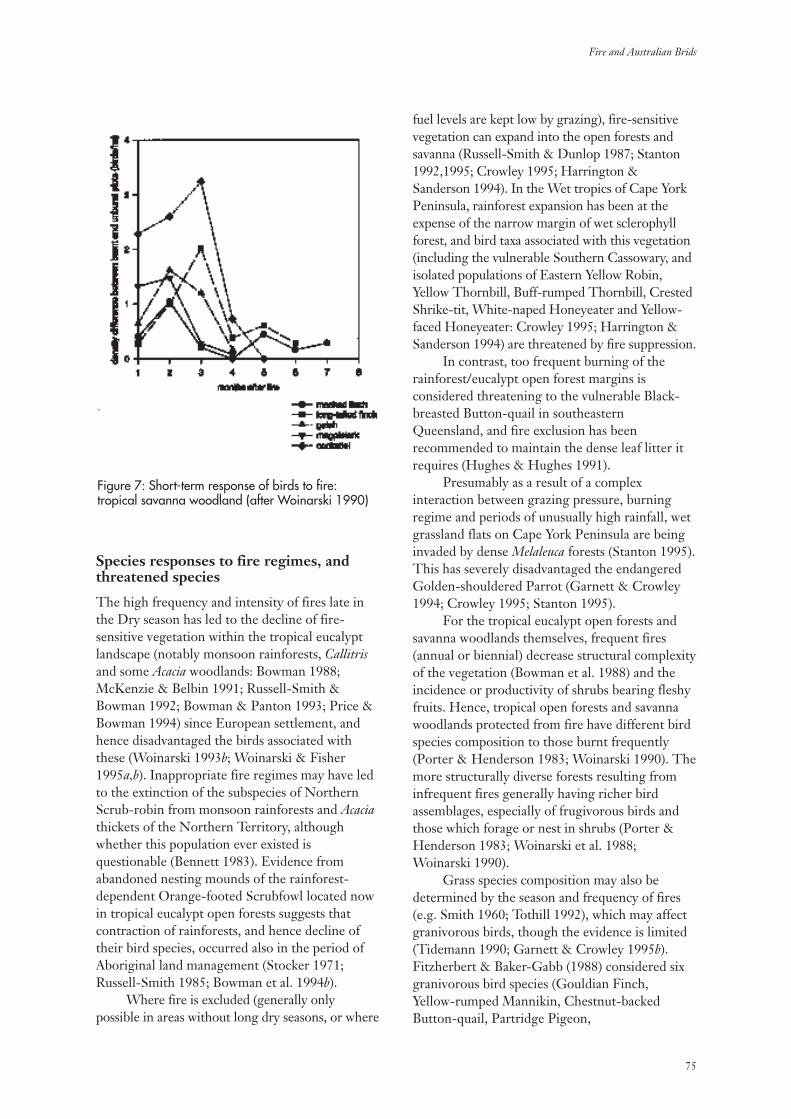
Species responses to fire regimes, andthreatened species
The high frequency and intensity of fires late inthe Dry season has led to the decline of fire-sensitive vegetation within the tropical eucalyptlandscape (notably monsoon rainforests, Callitrisand some Acacia woodlands: Bowman 1988;McKenzie & Belbin 1991; Russell-Smith &Bowman 1992; Bowman & Panton 1993; Price &Bowman 1994) since European settlement, andhence disadvantaged the birds associated withthese (Woinarski 1993b; Woinarski & Fisher1995a,b). Inappropriate fire regimes may have ledto the extinction of the subspecies of NorthernScrub-robin from monsoon rainforests and Acaciathickets of the Northern Territory, althoughwhether this population ever existed isquestionable (Bennett 1983). Evidence fromabandoned nesting mounds of the rainforest-dependent Orange-footed Scrubfowl located nowin tropical eucalypt open forests suggests thatcontraction of rainforests, and hence decline oftheir bird species, occurred also in the period ofAboriginal land management (Stocker 1971;Russell-Smith 1985; Bowman et al. 1994b).
Where fire is excluded (generally onlypossible in areas without long dry seasons, or where
fuel levels are kept low by grazing), fire-sensitivevegetation can expand into the open forests andsavanna (Russell-Smith & Dunlop 1987; Stanton1992,1995; Crowley 1995; Harrington &Sanderson 1994). In the Wet tropics of Cape YorkPeninsula, rainforest expansion has been at theexpense of the narrow margin of wet sclerophyllforest, and bird taxa associated with this vegetation(including the vulnerable Southern Cassowary, andisolated populations of Eastern Yellow Robin,Yellow Thornbill, Buff-rumped Thornbill, CrestedShrike-tit, White-naped Honeyeater and Yellow-faced Honeyeater: Crowley 1995; Harrington &Sanderson 1994) are threatened by fire suppression.
In contrast, too frequent burning of therainforest/eucalypt open forest margins isconsidered threatening to the vulnerable Black-breasted Button-quail in southeasternQueensland, and fire exclusion has beenrecommended to maintain the dense leaf litter itrequires (Hughes & Hughes 1991).
Presumably as a result of a complexinteraction between grazing pressure, burningregime and periods of unusually high rainfall, wetgrassland flats on Cape York Peninsula are beinginvaded by dense Melaleuca forests (Stanton 1995).This has severely disadvantaged the endangeredGolden-shouldered Parrot (Garnett & Crowley1994; Crowley 1995; Stanton 1995).
For the tropical eucalypt open forests andsavanna woodlands themselves, frequent fires(annual or biennial) decrease structural complexityof the vegetation (Bowman et al. 1988) and theincidence or productivity of shrubs bearing fleshyfruits. Hence, tropical open forests and savannawoodlands protected from fire have different birdspecies composition to those burnt frequently(Porter & Henderson 1983; Woinarski 1990). Themore structurally diverse forests resulting frominfrequent fires generally having richer birdassemblages, especially of frugivorous birds andthose which forage or nest in shrubs (Porter &Henderson 1983; Woinarski et al. 1988;Woinarski 1990).
Grass species composition may also bedetermined by the season and frequency of fires(e.g. Smith 1960; Tothill 1992), which may affectgranivorous birds, though the evidence is limited(Tidemann 1990; Garnett & Crowley 1995b).Fitzherbert & Baker-Gabb (1988) considered sixgranivorous bird species (Gouldian Finch,Yellow-rumped Mannikin, Chestnut-backedButton-quail, Partridge Pigeon,
75
Fire and Australian Brids
Figure 7: Short-term response of birds to fire:tropical savanna woodland (after Woinarski 1990)

Golden-shouldered Parrot and Hooded Parrot) tobe threatened by too frequent fires in the tropicalsavannas. Garnett & Bredl (1985) noted that thethreatened Star Finch was largely restricted tolong-unburnt patches of grass on Cape YorkPeninsula. On Cape York Peninsula, pastoralmanagement often involves “storm-burning” afterthe first rains of the Wet season (Crowley 1995).This may have removed much of the annualsorghum, possibly to the detriment of theendangered Gouldian Finch (Tidemann 1993a;Tidemann et al. 1993), though possibly to theadvantage of Golden-shouldered Parrots (Garnett& Crowley 1994).
The decline of two trunk-gleaninginsectivores, the Black Treecreeper of open forestson Cape York Peninsula and the northernsubspecies of Crested Shrike-tit of open forestsacross northern Australia, has been linked tofrequent intense and extensive late dry season fires(Robinson & Woinarski 1992; Garnett & Crowley1995a).
The vulnerable Red Goshawk occurs widelyacross open forests of northern Australia.Although there are reports of nestlings beingkilled by fire and nest trees being destroyed by fire(Aumann & Baker-Gabb 1991) and suggestionsthat high fire frequency is disadvantageous (Debus& Czechura 1988), the evidence to evaluate fireeffects is insufficient.
Exclusion of fire has been recommended forthe riparian habitat of the threatened Purple-crowned Fairy-wren (Rowley & Russell 1993),and less frequent fire (especially of extensive hotfires) for the threatened Black Treecreeper andCrested Shrike-tit (Robinson & Woinarski 1992;Garnett & Crowley 1995a).
Fire management for bird conservation
Fire management for the conservation of birds inthe tropical eucalypt open forests and savannawoodlands has emphasized the need formaintenance of a range of burning practicespreferably creating a fine-scale mosaic of firehistories (Press 1987; Brooker et al. 1990; Stanton1992; Crowley 1995; Garnett & Crowley 1995a),with emphasis on burning early in the dry seasonto prevent destructive late Dry season fires(Tidemann 1992; Woinarski 1993a) or “storm-burning” in the early Wet season (Garnett &Crowley 1994; Crowley 1995) for themanagement of particular bird species.
3.6 TUSSOCK GRASSLANDS
There is surprisingly little information on theimpact upon birds of fire in tussock grasslands(Fitzherbert & Baker-Gabb 1988; Woinarski1993a). Research on effects of fires in savannawoodlands is at least partly transferable tograsslands. Burning patterns in tussock grasslandsacross Australia are very variable, ranging fromattempted exclusion to frequent firing to promote“green pick” (Walker & Tothill 1992; Pressland etal. 1992). Their impact is almost everywhereregulated or confounded by grazing pressure.
Burning patterns in the floodplain grasslandsof northern Australia have now changed from thoseemployed by Aboriginal people, who lit mild earlydry season fires to decrease the likelihood ofsubsequent extensive destructive fire and tomanipulate graminoid species composition tofavour Magpie Goose, a pivotal food resource(Jones 1980; Lewis 1989; Lucas & Lucas 1993;Lucas & Russell-Smith 1993). Burning patternsnow tend to be more erratic, with a higherfrequency of wildfire (Press 1988).
In semi-arid Mitchell grasslands, landmanagement usually attempts fire exclusion (Orr& Holmes 1984; Anderson et al. 1988). This leadsto change in plant species composition andstructure of the grasslands, and, as fire promotesseeding (Scanlan 1980), to decrease in seedavailability. The consequences of such habitat andresource changes to birds are unknown(Woinarski 1993a).
Fire is also used extensively to maintainsemi-arid grasslands under invasion by “woodyweeds” (Walker & Tothill 1992; Sullivan et al.1992), and to aid in the clearance of woody speciesto create grasslands (Johnson & Purdie 1981;Anderson & Back 1992). Given the correlation ofbird species richness with vegetation structuralcomplexity (Recher 1969), this habitat alterationprobably reduces local species richness.
In temperate grasslands, burning regimeshave also changed appreciably from relativelyfrequent firing by Aboriginal people (Nicholson1981) usually to a contemporary regime ofattempted fire exclusion with occasional hotwildfire (Fitzherbert & Baker-Gabb 1988).Occasional wildfires in temperate grasslands ofthe Riverina may have severe impacts on grasslandbirds, notably the threatened Plains-wanderer(Baker-Gabb et al. 1990). Many grasses and othergrassland plant species depend upon specific
76
Australia’s Biodiveristy - Responses to Fire

burning regimes (Lunt 1991). Plant speciesdiversity is increased by regular burning at about 5year intervals (McDougall 1989). While thesefloristic (and vegetation structural) responses tofire regime probably influence grassland birdspecies composition, there is no substantialinformation on bird-fire relationships intemperate grasslands.
Early seral grasslands occurring soon after hot fires in heaths and forests attract a wide rangeof typical grassland birds, such as Richard’s Pipit, Blue-winged Parrot, quail and Australian Magpies(Loyn 1985b; Dickinson et al. 1986; Brooker &Rowley 1991).
3.7 HUMMOCK GRASSLANDS
Hummock grasslands form a distinctive andextensive environment acoss much of arid andsemi-arid Australia and in sandstone ranges ofnorthern Australia. Suijdendorp (1981)summarises prevailing fire regimes in hummockgrasslands and their broad ecological impacts.Current fire regimes generally differ from thepatch mosaic burns used by Aboriginal people tochanged scale of fires with fewer small mild firesbut more frequent extensive wildfire (Kimber1982; Griffin et al. 1983; Griffin 1984; Burbidge1985; Reid & Fleming 1992; Baker et al. 1993; Russell-Smith & Ryan 1994).Hummock grasslands may be expanding undersome current fire regimes at the expense of othervegetation types, such as Acacia woodlands andshrublands (Bowman et al. 1994a; Latz 1995).Environmental responses to fire in hummockgrasslands (and other arid communities) may beparticularly influenced by season of fire andrainfall following fire (Noble et al. 1984; Reid et al. 1993a).
In contrast to some detailed studiesdemonstrating successional patterns in mammalsand reptiles of hummock grasslands (e.g. Masters1993), there has been little research on therelationships of birds to fire regimes in hummockgrasslands. Reid et al. (1993a,b) found that short-term effects of fire in hummock grasslands ofcentral Australia include invasion of recently-burnt areas by nomadic open-country birds(including White-winged Triller, Budgerigar,Black Honeyeater, Crimson Chat, Zebra Finch,Banded Whiteface and Masked Wood-swallow).Pioneer plants may provide a rich seed resourcesoon after fire (Latz 1995), and many granivoresare hence attracted to recently-burnt areas (Bakeret al. 1993).
Longer-term impacts are less clearcut. Theavailability of hummock grass seeds generallyincreases with time since fire up to about 10-20years but may then decline (Westoby et al. 1988).Invertebrate numbers may be low in the first yearafter fire (Masters 1993). Several bird speciesshow preference for, or are restricted to, long-unburnt hummock grasslands. These includeRufous-crowned Emu-wren, Spinifexbird and thethreatened Striated Grasswren, all insectivoreswhich shelter or nest in large clumps of hummockgrass (Pedler 1991; Garnett et al. 1993; Reid et al.1993a,b). The threatened CarpentarianGrasswren, of sandstone ranges in the Gulf ofCarpentaria hinterland, also requires long-unburnt hummock grasslands or, at least, isseverely disadvantaged by the current regime offrequent extensive hot fires (CSIRO 1976;Schodde 1982; McKean & Martin 1985,1989).This may also be the case for the Black Grasswren(in the Kimberley) and White-throatedGrasswren (of the Arnhem Land sandstonemassif) (Fitzherbert & Baker-Gabb 1988),however, although extensive hot fires are nowfrequent in this environment (Russell-Smith &Ryan 1994), the evidence of their impact onWhite-throated Grasswrens is unclear (Noske1988,1992a,b: Woinarski 1992).
The threatened Night Parrot formerlyoccurred in hummock grasslands and other aridhabitats across much of inland Australia. Its rapiddecline may have been due to increased frequencyof extensive fires (Ashby 1924a). Recent reportshave been from an area where small control burnshad led to a mosaic of vegetation ages and hadprevented extensive fire (Garnett et al. 1993).
Fine-scale mosaic burning (largely to reducethe likelihood of extensive hot wildfire) has beenrecommended for the conservation of birdsgenerally in hummock grasslands (Gibson 1986;Pedler 1991; Reid et al. 1993a,b; Baker et al. 1993).
3.8 ACACIA SHRUBLANDS ANDWOODLANDS
There is relatively little information on the birdsof Acacia woodlands and shrublands, fire regimesoperating in them, or the effects of fire regimes ontheir birds. Aboriginal use of fire in semi-arid andarid Acacia communities may have been generallyrestricted to infrequent burning of many smallpatches, creating a fine-scale mosaic ofsuccessional states (Griffin & Hodgkinson 1986).European colonisers in these environments
77
Biodiveristy and bushfires

initially had no fire strategy, or used fire to clearcountry. The incidence of extensive wildfire hasaccordingly increased markedly (Griffin &Hodgkinson 1986).
Some Acacia formations, such as Mulga A. aneura, are notably fire-sensitive, and aredeclining under current management (Reid et al.1993b; Bowman et al. 1994a). Reid et al. (1993b)noted that:
“given the importance of this species to theassociated rich bird community, it wouldseem that the destruction of extensive areasof mulga by fire would decimate birdpopulations, at least until the mulga hadregenerated to a certain age”.Most bird species will recolonise mulga by
about 10–15 years post-fire (Reid et al. 1993a,b),though older mulga may support more mistletoe,and hence the bird species associated with this(e.g. White-fronted Honeyeater: Reid et al.1993a). Recently-burnt mulga contains manygeneralist nomadic species, such as White-wingedTriller and Zebra Finch (Reid et al. 1993a).
The loss, through burning and mechanicalclearing, of Brigalow Acacia harpohylla forests andother trees in inland southeastern Queensland,may have widespread ripple effects, as this area isprobably an important wintering base for manybird species migrating from southeastern Australia(Nix 1993).
Increased incidence of intense wildfire isregarded as the major conservation threat to birdsof Lancewood Acacia shirleyi woodlands(Woinarski & Fisher 1995 a,b).
Conservation managers have recommendedthat extensive wildfires in Acacia communitiesshould be avoided, and hence biodiversitymaintained, by mosaic burning, with small mildfires (Griffin & Hodgkinson 1986).
3.9 OTHER HABITATS
In chenopod shrublands, Brooker et al. (1979)noted that White-winged Fairy-wrensdisappeared from a bluebush site for at least5 years post-fire. Wildfire may be a substantialthreat to the Nullabor Quail-thrush, especially asregrowth of chenopods is often prevented (ordelayed) by rabbit grazing (Burbidge & Pedler1993,1996; Pedler & Burbidge 1995).
There is little information on the impact offire regimes on birds in other Australianenvironments. In some cases (such as mangroves,saltpans, rainforests), this is because fire may notbe a major management issue, at least in the short-term.
3.10 SPECIAL CASE: ISLANDS
Increased use and extent of fire followingEuropean colonisation contributed to theextinction of the Kangaroo Island Emu (Ashby1924a) and the Glossy Black-Cockatoo on KingIsland (Green & McGarvie 1971), although theevidence for its role in the former case is limited.
Birds which nest in colonies on, or close to,the ground may be particularly vulnerable to fire.The most obvious examples of this susceptibilityare seabirds, whose island (or, occasionallymainland) colonies have been frequentlydevastated by fire, often killing many thousands(and/or a large proportion) of the nesting adults,young and/or eggs (MacGillivray 1910; Hull1922; Whitley 1944; Fowler 1945; Abbott 1981;Lane 1976; White 1979a,b; Garnett 1987; Chatto1995), and/or rendering vegetation subsequentlyless suitable for breeding (Pescott 1976; Paton &Paton 1977; Brothers 1983; Brothers & Skira1987,1988; Skira & Brothers 1988a,b; Walker &Hulsman 1993). In many cases, these fires havebeen deliberately lit by fishermen.
78
Australia’s Biodiveristy - Responses to Fire

Recher et al. (1985) noted that:“Considering the frequency with whichfires occur in eucalypt forests and woodlandsand their dramatic impact on thelandscape, there is remarkably littleinformation about the effects of fire onfauna or the long-term consequences ofburning on forest ecosystems”.This is a striking feature of this review of the
effects of fire on Australian birds. In general, thereis insufficient information available to directmanagement or to evaluate a range ofconservation options. Most information isanecdotal or serendipitous. There have been veryfew long-term studies. There have been fewcomparisons of different fire regimes, or oftreatments with controls. There have beenrelatively few studies with individually-markedbirds. For some environments, and manybiogeographic regions, there is effectively noinformation on responses of birds to current, oralternative, fire regimes (Table 2).
The disparate approaches, and limitations, ofresearch on birds has hampered comparisons,synthesis and overview. A similar problem hasrecently been described for research on theimpacts of fire upon invertebrates (Friend 1995).
Sampling sites across a range of fire ages
A relatively high proportion of studies haveconsidered succession (or change in bird speciescomposition with post-fire age) by comparing birdassemblages at separate sites across a range of ages(e.g. Cheal et al. 1979; Meredith et al. 1984; Loyn1985a; Bamford 1985a,b; Carpenter & Matthew1986; McFarland 1988; Woinarski 1989b). Suchresearch provides a broad-brush response to fireages, and is attractive in that sampling of a rangeof ages can be undertaken over a short timeperiod. However, the ages sampled areconstrained by availability, rendering predictionfrom beyond the sampled range uncertain (Baker& Whelan 1994). Because different sites aresampled to represent different post-fire ages, it isalso likely that fire effects are compounded orconfounded by other environmental factors orlocal site effects (Burbidge et al. 1989). Exceptwhere specific searches are made for rare species
(e.g. Meredith et al. 1984; McFarland 1988), thisapproach may also provide too few data to enablestatistical analysis of responses of such species.
In most cases, this approach also fails toconsider fire regime, concentrating instead ontime since last fire. Where more detailedinformation on fire histories of sites are known(e.g. Russell-Smith & Ryan 1994), systematicsampling of sites should be able to indicate theimpacts of a range of fire regime parametersrather than simply period since last occurrence.
Repeated sampling of individual sites
There have been a small number of valuablestudies which have monitored bird assemblages atsingle sites over an extended time followingwildfire (Hewish 1983; Smith 1985a; Reilly1991a,b; Turner 1987,1992; Brooker & Rowley1991; Cale & Burbidge 1993). In some cases thesehave included comparisons with before-fireabundances (Hewish 1983; Recher et al. 1987b),or with nearby unburnt vegetation (Hewish 1983;Recher et al. 1987b; Turner 1992). More studieshave considered short-term (0–2 years post-fire)responses (e.g. Roberts 1970; Ratkowsky1979,1985; Dedman 1983a–e,1984; Recher et al.1985; Loyn et al. 1992a), during which changesmay be most rapid. In general, the interpretation(and extrapolation) of these studies is constrainedby lack of replication, such that it may be difficultto distinguish idiosyncratic site (or fire) factorsand responses from more general responses.
Because control burning is, by definition,more tractable than wildfire, there has beensomewhat more systematic sampling of the short-term responses of birds to control fires (Hodgson& Heislers 1972; Cowley 1974; Kimber 1974;Christensen & Kimber 1975; Ratkowsky1978,1979; Tingay & Tingay 1984; Wardell-Johnson & Christensen 1992; Woinarski 1990;Loyn et al. 1992b), usually with before-aftercomparisons, and occasionally with comparisonswith unburnt controls (Kimber 1974; Christensenet al. 1985; Woinarski 1990; Loyn et al. 1992b;Tolhurst 1996) or between a range of controlburning approaches (notably season of burn:Woinarski 1990; Loyn et al. 1992b; Tolhurst 1996).However, these studies cover a remarkably smallproportion of the environments in which
79
Biodiveristy and bushfires
4. RESEARCH ADEQUACY, METHODS AND PRIORITIES

control burning is regularly and extensivelyimplemented. Most also report changes in themore common species only (for which more dataare compiled), and hence may overlook impacts offire upon rarer birds (Christensen et al. 1985;Meredith 1988). Again, the extrapolation fromthese studies is usually hampered by inadequatereplication; though generally consistent findingsacross a range of studies at separate locationssuggest that the main conclusions are probablyrobust.
Sampling sites of known long-term fire regimes
There have been extremely few studies whichhave examined longer-term effects upon birds ofspecified fire regimes (Meredith 1988).Exceptional have been studies in tropical eucalyptopen forests, where Porter & Henderson (1983)compared bird assemblages in plots of threedifferent fire regimes (unburnt, annually burntand burnt at 2–5 year intervals) maintained overnearly 30 years; and Woinarski (1990) comparedbird assemblages in plots of four regimes(unburnt, burnt annually in the early dry season,burnt annually in the late dry season and burntbiennially) maintained over 14 years. Thedifficulty of maintaining consistent fire treatmentsover such long period (and the unreality ofassuming that such strictly-enforced regimescould occur beyond experimental sites) hasprovided a strong disincentive for such studies.
Fire season and breeding
There have also been very few studies which havecompared the effects of fires occurring at differentseasons. Several authors have advocated that firesshould avoid the breeding season, as fires thenmay destroy a high proportion of the season’sreproductive output for birds nesting on theground or in low vegetation (Bedggood 1980;Reilly 1991a,b; Baker et al. 1993), and this impactmay be compounded when fires are frequent(Lucas & Lucas 1993; Brooker & Brooker 1994).For example, Rowley & Brooker (1987) suggestedthat a single hot wildfire had less impact on thesurvival of Splendid Fairy-wrens than did coolcontrol fires, because the wildfire occurred afterthe breeding season whereas the control fires wereearlier. In contrast, Christensen et al. (1985)considered that occasional fires during thebreeding season probably have limited andinsignificant impact on populations of forest birds.
As noted above, a special case may be birds whichnest in isolated colonies, where a single fire maydestroy many individuals and/or a largeproportion of the population.
An unusual relationship between birds andfire is the consumption of ash and charcoal by arange of bird species (Baldwin 1965; Coate 1985;Pescott 1985; Hutchins 1988), which may berelated to mineral requirements for breeding, butlimited analysis has failed to establish any patternor compelling explanation of this feature.
Autecological studies
Some detailed long-term autecological studieshave revealed important subtleties of fire impacts,such as delayed responses due to reduced breedingsuccess in the years after fire (Marchant 1985;Rowley & Brooker 1987; Brooker & Rowley1991; Russell & Rowley 1993), the fate ofdispersing birds, role of unburnt patches andcomplex patterns of use of mosaics of different fireage (Smith 1979a,1985a,1987a; Brown & Wilson1981,1984; Benshemesh 1992), change inpopulation age structure, sex ratio andsurvivorship (Rowley & Brooker 1987; Brooker &Rowley 1991; Russell & Rowley 1993), andcritical requirements for apparently trivialresources (e.g. cobwebs) which may beparticularly affected by fire (Brooker & Rowley1991; Recher 1991). The subtlety but importanceof these factors suggests that brief monitoringpost-fire may provide misleading assessments oflonger-term impacts. It also demonstrates thatmanagement of fire for the conservation ofindividual bird species must be preceded bydetailed long-term research (Smith 1987a).
Translation of research to management
Such long-term autecological studies areparticularly valuable in that they can providespecific information on which to base carefullytargetted fire management (e.g. Hopkins 1985b;Brooker & Brooker 1994). There has beenremarkably little effective translation of researchresults to evaluated management options - thoughsome exceptions include Meredith (1982), Smith(1987a), Benshemesh (1990), McFarland (1992),Cale & Burbidge (1993) and Hopkins & Smith(1996) - possibly because of the paucity ofquantitative data.
Brooker & Brooker (1994) provide easily themost detailed assessment of a range of firemanagement options for any Australian bird
80
Australia’s Biodiveristy - Responses to Fire

species, using modelling to predict survival/extinction probabilities for a range of fire regimesacross a range of habitat patch sizes. However, thiscase is exceptional, as it is based on data from oneof the very few long-term studies of the responsesof individual species to fire (Rowley & Brooker1987; Russell & Rowley 1993). Comparable dataare available for very few other species (probablybeing limited to Noisy Scrub-bird, WesternBristlebird and possibly Western Whipbird,Rufous Scrub-bird and Ground Parrot). Yet suchmodelling may be vital for management of fire-susceptible threatened species, to decide whetherto translocate populations, how to protectpopulations from fire, whether to provide a rangeof fire ages in different patches, etc. These studiesprovide good examples of how research shouldprecede and guide management.
In the absence of a sufficient researchhistory, fire management (especially inconservation reserves) should conservativelyinclude a broad range of regimes, so as not toforeclose options (Recher 1981,1991; Christensenet al. 1985; Brooker et al. 1990). Fire managementshould be accompanied by ongoing monitoring ofimpacts upon the fauna, and particularly on theknown fire-sensitive species, and assessment ofthese impacts should then be used to refine, ormore precisely target, that management (Wardell-Johnson et al. 1989). Acceptance of theimportance of fire management for conservationof fauna is relatively recent. Only 20 years ago,Newsome et al. (1975) considered that:
“it is too early to think of utilising fire as atool for management of fauna in NationalParks”.
Landscape context
Relatively little research has been directed atspatial analysis of fire effects, such as the role ofunburnt patches, the relationship between fireextent and direct impacts and recolonisation, andthe process of recolonisation of burnt isolates.Many experienced field workers have reportedthat fires which comprehensively burn areas havegreater impacts than patchy burns which leavesome unburnt areas (e.g. Recher 1981; Smith1989; Reilly 1991a); however there is littlequantitative documentation of the role, orrequired size, of unburnt patches. Using radio-telemetry, Benshemesh (1990) demonstrated theimportance of small unburnt patches in the post-fire persistence of Malleefowl. Smith (1989)
reported that a forest bird assemblage provedremarkably resilient to a wildfire largely becauseof small areas (in gullies) that were left unburnt.Rowley & Brooker (1987) noted that SplendidFairy-wrens that retained some unburnt patchesin their territories persisted after fire, whereasthose whose territories were completely burntdisappeared. However, Brooker & Rowley (1991)found no apparent preference for unburnt patchesof heath among a range of bird species after fire.
Recolonisation of burnt areas is generallyquicker if smaller areas are burnt and aresurrounded by similar unburnt vegetation (e.g.Reilly 1991b), but again the evidence is moreanecdotal than analytical. Small burnt areas mayattract more concentrated populations of post-firescavengers than extensively burnt areas(Woinarski 1990).
Small distant isolates which are unlike theirsurrounds are less quickly recolonised post-firethan are large isolates close to unburnt sourceareas (Reilly 1991a,b; McFarland 1991), andunburnt corridors may be important in thisrecolonisation (Danks 1991; DuGuesclin et al.1995). Small fragments may also suffer moresevere fire impacts than large fragments (Recheret al. 1987b), possibly because there is a higherprobability of some unburnt areas (or somesurvivors) in larger fragments. Increasedfragmentation of suitable habitat, either throughclearing or widespread application ofinappropriate fire regimes, may exacerbate localfire effects, especially for species with relativelypoor dispersal ability.
In some cases, fragmentation may also offersome protection from landscape-wide fires (forexample, much of the limited long-unburnt malleeexists as small patches isolated by farmland), andpossibly more flexibility in manipulatingvegetation ages (e.g. because it is relatively easy toimpose contrasting regimes on different isolates).However, in general, fragments are probably moreprone to homogenising fires than are extensivehabitats (Menkhorst & Bennett 1990), supportrelatively small populations and are less likely to berecolonised post-fire (Brooker & Brooker 1994),with the result that they are less likely to retainenvironmental diversity (Williams et al. 1994) andsustain, over the long-term, populations of specieswhich are associated with particular seral stages.Fragments, particularly smaller ones, may also besubjected to fire regimes which are
81
Biodiveristy and bushfires

unlike those occurring in extensive habitats,typically being burnt much more frequently ormuch less frequently (depending upon theirsurrounds). This disruption of ecological processesis recognised as a major management problem infragmented habitats (e.g. Abensperg-Traun &Smith 1993; Yates et al. 1994), and is likely to becritically important to fire-susceptible threatenedbird species in fragmented habitats.
Fire control
There has been little research on the impactsupon birds of fire control mechanisms other thanfuel reduction burning. Adam & Robinson (1996)and Davidson & Robinson (1992) suggested thatland managers who must protect property fromwildfire should preferably use judicious slashingrather than control burning for roadside remnantsused by Grey-crowned Babblers. However, Schulz(1991) noted that slashing also had problems forthe conservation of this species. The provision ofwater sources for fire fighting may locally alterbird community dynamics in arid or semi-aridareas (Cheal et al. 1979; Meredith 1982).Firebreaks and associated road networks are likelyto lead to increased penetration of predators, andincreased predation post-fire (May 1994).
Interactive effects between fire and otherland uses or threatening processes
There has been limited research directed at theimpacts upon birds of fire/forestry interactions(e.g. Recher et al. 1985,1987b; Dickinson et al.1986), but many aspects of fire management inforestry operations have not been assessed interms of impacts upon birds.
Other than preliminary studies by Brooker(1988) and Tidemann (1990), there has been noresearch investigating the impacts upon birds ofinteractions between fire and grazing by livestock.Grazing by stock leads to changes in fuel loads,floristic composition, vegetation structure,recruitment of tree species, successional processand, hence, to different fire regimes to thoseoperating in ungrazed areas. Pastoralists may alsoimpose particular fire regimes with the single aimto provide short-term benefit to stock (Head et al.1992). Removal of stock from woodlandfragments is advocated as a conservation measurefor woodland birds (Robinson 1994), but theimpact of this removal on fire regimes is unclear.Removal of the immediate perceived problem,grazing, may lead to its replacement with anothermanagement issue, fire.
Grazing by other herbivores may delay orstall recovery of vegetation after fire, extending ormagnifying fire impact. The interaction of rabbits,fire and chenopod shrublands is threateningNullabor Quail-thrush (Burbidge & Pedler 1996).Grazing by macropods may stall regrowth ofheath, to the detriment of Western Bristlebirds(Cale & Burbidge 1993).
Fire management, even in conservationreserves, serves many purposes other thanoptimising environments for particular bird species.In order to include bird conservation in a multi-purpose fire management plan, it is critical thatresearch be directed at the assessment theconsequences for birds of fire/land-use interactions.
There has also been little study of theinteraction between fire and predation, althoughthe compounding effects of these two factors hasbeen recognised (e.g. Braithwaite & Estbergs 1987;Benshemesh 1990; Recher 1991). While birds maysurvive fire, lack of vegetation cover in burnt areasmay make them far more susceptible to predation,and predator density may increase in recently-burnt areas.
Multidisciplinary studies
With notable exceptions (e.g. Recher et al. 1985;Tolhurst & Flinn 1992), almost all research on theeffects of fire upon birds has been carried out inan ecological vacuum, without synchronouscomplementary studies of the responses of othercomponents of the ecosystem. Howevermultidisciplinary approaches may provide themechanisms for explaining why individual birdspecies respond in particular ways to a given fireor fire regime, and hence provide powerfulcapability to better guide management.
Representation of biogeographic regionsand environments
The research effort directed at the relationshipsbetween Australian birds and fire has been veryinequitably distributed across habitats andgeographic regions (Table 2). While the knownoccurrence of fire-sensitive threatened birds inheaths and mallee is reasonable grounds for givingthese environments high priority attention, otherenvironments where fire is a major managementissue have been neglected. Critical gaps occur in thelong-term effects of control burning in temperateeucalypt forests, and in the impacts of fire regimesin woodlands (including those dominated by Acacia,Callitris and Casuarina species), tussock grasslands,hummock grasslands and rainforest margins.
82
Australia’s Biodiveristy - Responses to Fire

Of the three species and four subspecies of birdswhich have become extinct since Europeansettlement, inappropriate fire regimes caused, orcontributed to, the loss of two species (KangarooIsland Emu and Paradise Parrot) and threesubspecies (Northern Territory subspecies ofNorthern Scrub-robin, southwestern Australiansubspecies of Rufous Bristlebird and southwesternAustralian subspecies of Lewin’s Rail). Fire is nowrecognised as a main threatening process for manyrare, vulnerable and endangered Australian birds.Brouwer & Garnett (1990) listed inappropriate fireregime as a threat for 22 of 52 threatenedAustralian bird species, and Garnett 1992bconsidered it threatened 51 taxa (second only tohabitat clearance and fragmentation, whichthreatened 52 taxa) (Table 3).
This recognition of the significant impact offire is not recent. For example, in reviewing thestatus of threatened Australian birds, Ashby(1924b) noted:
“The most serious factor of all is thedestruction of both food supply and shelter,and, of course, breeding haunts, by bush-fires. I consider that these … factors,especially …. bush-fires, account for morethan nine-tenths of the disappearance ofcertain forms … I am confident that theindiscriminate burning of bush, which isthe concomitant of all farming and grazingoperations, is by a long way the major causeof the disappearance of many of our rarerbirds”.Despite this long-standing appreciation of the
problem, there has been very limited researchdirected at the impacts of fire regimes on birds andlittle informed management of fire for birdconservation. To a large extent, myth, anecdote andcasual observations substitute for detailedknowledge in management advice (Meredith 1982).
However, recent research has served to focusmore sharply on the intricacies of fire regime, andaway from the perception that occasionalcatastrophic wildfires are inevitably the mostserious fire threats to bird conservation (Recher1981). Rather, minor changes in fire regime maybe critical (Saunders 1985) and lead to almostimperceptibly gradual, but inexorable, birddecline (Brooker & Brooker 1994).
It is striking that, across a broad range ofenvironments, most fire-sensitive threatenedspecies require fire intervals longer than thosewhich have been imposed since Europeansettlement. Too frequent burning has endangeredspecies such as Noisy Scrub-bird, WesternBristlebird, Malleefowl and Ground Parrot. Theold-growth (or mid to late seral) vegetation thatthese species require, or are most abundant in, isnow becoming disappearingly rare. Themaintenance of suitably-aged vegetation isrequired to retain such species. The endangermentof so many species reliant on relatively oldvegetation is a clear indication that land managersare now generally burning far more extensively orfrequently than prior to European settlement, orthat fires now are generally more destructive.
The very low fire frequency, or fireexclusion, required by many of these species (e.g.preferred intervals of at least 20 years for mostthreatened heathland birds (Smith 1985a) or atleast 60 years for Malleefowl (Benshemesh 1990))will pose serious management problems,especially where many of these species somewhatparadoxically live in very fire-prone environments(mallee and heath), where potentially competingfire management goals exist, and where adjacentpopulations of humans lead almost inevitably touncontrollable increased ignition of fires.
The long fire-free periods that many ofthese fire-sensitive birds require, and the very oldtrees (generally centuries to millenia old) requiredby most of the many hollow-nesting birds,suggests that much of the bird fauna has adaptedto an environment which has a long history oflittle or mild disturbance. The association with, orreliance upon, long-unburnt vegetation by asignificant component of the bird fauna is inmarked contrast to that shown by other vertebrategroups, where the successional sequence is oftenplayed out within 10 to 20 years (e.g. Fox 1982;Masters 1993), other than for hollow-dependentspecies. This disparity may be because most of themammal, reptile and frog fauna is terrestrial, andhence responds to the relatively rapid changes inunderstorey, ground and litter characteristicsrather than to the slower changes in taller shrubsand trees. Alternatively, the lesser mobility ofmammals, reptiles and frogs may have precluded
83
Biodiveristy and bushfires
5. CONSERVATION OVERVIEW

the development of species tightly associated withold seral stages.
However, two features generally common tomost of the fire-sensitive threatened bird speciesare low reproductive output and relatively limiteddispersal abilities (Smith 1977,1985a,c,1987a,b;Reilly 1991b): indeed, these features probablypartly define why these species are threatened byfrequent fire. While these are useful adaptivetraits in stable (and often resource-poor)environments, they are profound handicaps wherethat environment is disturbed. Such species areparticularly vulnerable to habitat fragmentationand fire, and their conservation must involvelandscape-scale perspective and management, elserecovery from fire may be fatally compromised.
Friend (1993) considered the responses tofire of mallee mammals, reptiles and frogs, andsought to compare the ecological characteristics ofspecies according to their preferred seral stages.Following this approach, although close totautological, it is clear that most fire-sensitive birdspecies are mainly insectivorous and forage indense shrubs, shrubby understorey or thick leaflitter.
The broad requirements of some of thesethreatened fire-sensitive bird species are nowreasonably obvious. However, many species nowconsidered reasonably abundant may be moresubtly disadvantaged by current fire regimes. Thedetailed study of the common Splendid Fairy-wren by Rowley & Brooker (1987), Brooker &Rowley (1991), Russell & Rowley (1993) andBrooker & Brooker (1994) has illustrated thedanger of small but repeated impacts associatedwith a regime of frequent control burning. Wheresuch regimes are sustained over long periods(decades to centuries), the bird assemblage withinhabitats will be gradually re-sorted and speciesnow considered secure may be lost. Over longerperiods (decades to millenia), sustained fireregimes will re-arrange spatial relationshipsbetween habitats, resulting in decline or extinctionof fire-sensitive habitats (and their associated birdfauna). The long period of such change relative tothat of most studies on fire impacts suggests greatcaution should be exercised in interpretation ofstudies which suggest minor or no changesassociated with fire.
In general, currently threatened bird specieswill benefit from longer intervals between fires.Such change will disadvantage species associatedwith early seral stages. However, these species
(such as Richard’s Pipit, Little Button-quail, Blue-winged Parrot, Scarlet Robin, Flame Robin) aregenerally widespread and common, and typicallyoccur after fire across a broad range ofenvironments. In the first few years post-fire, thepresence of such invading generalist speciesalongside recolonisers may lead to local speciesrichness which is greater than that pre-fire (e.g.Christensen et al. 1985). However, on a landscapescale this is not equivalent to increasedbiodiversity, as habitat characteristics (and hencebird species composition) of early seral stages ofdisparate environments tend to be more similar toeach other than are later stages. Hence, frequentregular fires across a range of environments willresult in a convergence of their bird fauna, and aloss of the late seral species which contributemuch of the distinctiveness to differentenvironments.
Such homogenisation may be avoided byplanned fine-scale mosaic burning with a range offire regimes, but with priority protection ofcurrent long-unburnt areas (and ensuring futureprovision of such areas). Such management hasbeen recommended across a very broad range ofAustralian environments (Recher 1981,1991;Christensen et al. 1985; Braithwaite 1985; Emison& Bren 1989; Brooker et al. 1990). The approachhas the advantage of conservatively covering abroad spectrum of fire regime options, until betterknowledge of the requirements of individualspecies is available. Such knowledge will beacquired only when a much more comprehensive,strategic and concerted research effort isestablished.
84
Australia’s Biodiveristy - Responses to Fire

Abensperg-Traun, M. and Smith, G.T. (1993).Predictable effects of agriculturaldevelopment on the long-term availability ofhollows for animals: observations from theWestern Australian wheatbelt. PacificConservation Biology 1, 78–79.
Abbott, I. (1981). Seabird Islands: No. 106.Mondrain Island, Archipelago of theRecherche, Western Australia. Corella 5,60–61.
Abbott, I., & Christensen, P. (1994). Applicationof ecological and evolutionary principles toforest management in Western Australia.Australian Forestry, 57, 109–122.
Abbott, I., Van Heurck, P. and Wong, L. (1984).Responses to long-term fire exclusion:physical, chemical and faunal features oflitter and soil in a Western Australian forest.Australian Forestry 47, 237–242.
Adam, P., & Robinson, D. (1996). Negative effectsof fuel-reduction burning on the habitat ofthe Grey-crowned Babbler Pomatostomustemporalis. Victorian Naturalist 113, 4–9.
Ambrose, G.J. (1982). An ecological andbehavioural study of vertebrates usinghollows in eucalypt branches. Ph.D. thesis.La Trobe University: Bundoora.
Anderson, E. and Back, P. (1992). Fire in brigalowlands. In B.R. Roberts (Ed.) Fire research inrural Queensland. (pp. 324–339).Toowoomba: University of SouthernQueensland.
Anderson, E.R., Pressland, A.J., McLennan, S.R.,Clem, R.L. and Rickert, K.G. (1988). Therole of fire in native pasture management. InW.H. Burrows, J.C. Scanlan and M.T.Rutherford (Eds.) Native pastures inQueensland - the resources and their management(pp. 112–124). Brisbane: QueenslandDepartment of Primary Industries.
Arnold, G. W., Smith, G. T., Rowley, I. C. R., &Brooker, M. G. (1993). The effects of fire onthe abundance and distribution of animals inAustralian ecosystems, with emphasis onMediterranean ecosystems. In L. Trabaud &R. Prodon (Eds.), Fire in Mediterraneanecosystems (pp. 237–257). Brussels:Commission of the European Communities.
Ashby, E. (1921). Notes on the supposed “extinct”
birds of the south-west corner of WesternAustralia. Emu 20, 123–124.
Ashby, E. (1924a). Notes on extinct or rareAustralian birds, with suggestions as to someof the causes of their disappearance. Part I.Emu 23, 178–183.
Ashby, E. (1924b). Notes on extinct or rareAustralian birds, with suggestions as to someof the causes of their disappearance. Part II.Emu 23, 294–298.
Ashton, D.H. (1975). The root and shootdevelopment of Eucalyptus regnans F.Muell.Australian Journal of Botany 23, 867–887.
Aumann, T., & Baker-Gabb, D. J. (1991). Theecology and status of the Red Goshawk innorthern Australia. RAOU Report No. 75.Royal Australasian Ornithologists Union.
Backhouse, G. N. (1987). Management ofremnant habitat for conservation of theHelmeted Honeyeater Lichenostomusmelanops cassidix. In D. A. Saunders, G. W.Arnold, A. A. Burbidge, & A. J. M. Hopkins(Eds.), Nature conservation: the role ofremnants of native vegetation (pp. 287–294).Chipping Norton: Surrey Beatty.
Baird, R.F. (1984). The Pleistocene distribution ofthe Tasmanian Native-hen Gallinula mortieriimortierii. Emu 84, 119–123.
Baird, R.F. (1991). Avian fossils from theQuaternary of Australia. In P. Vickers-Rich,J.M. Monaghan, R.F. Baird and T.H. Rich(Eds.) Vertebrate plaeontology of Australasiapp.809–870. Lilydale: Pioneers DesignStudios.
Baker, J., & Whelan, R. J. (1994). Ground Parrotsand fire at Barren Grounds, New SouthWales: a long–term study and an assessmentof management implications. Emu 94,300–304.
Baker, L., Woenne–Green, S., & MutitjuluCommunity (1993). Anangu knowledge ofvertebrates and the environment. In J. R. W.Reid, J. A. Kerle, & S. R. Morton (Eds.),Uluru fauna. The distribution and abundance ofvertebrate fauna of Uluru (Ayers Rock-MountOlga) National Park, N.T. (pp. 79–132).Canberra: Australian National Parks andWildlife Service.
Baker-Gabb, D. J., Benshemesh, J. S., & Maher, P.
85
Fire and Australian Brids
REFERENCES

N. (1990). A revision of the distribution,status and management of thePlains-wanderer Pedionomus torquatus. Emu,90 161–168.
Baldwin, M. (1965). Bird eating charcoal. Emu, 64208.
Bamford, M. J. (1985a) The dynamics of smallvertebrates in relation to fire in banksiawoodland near Perth, Western Australia. PhD,Murdoch University.
Bamford, M. J. (1985b). The fire-related dynamicsof small vertebrates in Banksia woodland: asummary of research in progress. In J. R.Ford (Ed.), Fire ecology and management inWestern Australian ecosystems (pp. 107–110).Perth: Western Australian Institute ofTechnology.
Bedggood, G.W. (1980). Birdlife between LakeTyers and Marlo, Victoria. Australian BirdWatcher 8, 147–162
Beeton, R.J.S. (1985). The Little Corella - aseasonally adapted species. Proceedings of theEcological Society of Australia 13, 53–63.
Belcher, C. (1993). Rufous Bristlebird survey andhabitat analysis, Port Campbell National Park1992. Department of Conservation andNatural Resources.
Bennett, A.F. (1993). Fauna conservation in boxand ironbark forests. Victorian Naturalist110, 15–23.
Bennett, S. (1983). The Northern Scrub-robinDrymodes superciliaris in the NorthernTerritory. Emu 83, 105–107.
Benshemesh, J. (1988). Report on a study ofmalleefowl ecology. Department ofConservation, Forests and Lands.
Benshemesh, J. (1990). Management ofMalleefowl - with regard to fire. In J. C.Noble, P. J. Joss, & G. K. Jones (Eds.), Themallee lands: a conservation perspective (pp.206–211). Melbourne: CSIRO.
Benshemesh, J. S. (1992) The conservation biology ofMalleefowl, with particular regard to fire. PhD,Monash University.
Benshemesh, J. S. (1994). Malleefowl Leipoaocellata. Department of Conservation andNatural Resources.
Benson, D.H. (1981). Vegetation of UpperMangrove Creek, Wyong, New SouthWales. Cunninghamia 1, 7–22.
Boekel, C. (1980). Birds of Victoria River DownsStation and of Yarralin, Northern Territory.Part 1. Australian Bird Watcher 8, 171–193.
Bowman, D.M.J.S. (1988). Stability amidturmoil?: towards an ecology of northAustralian eucalypt forests. Proceedings of theEcological Society of Australia 15, 149–158.
Bowman, D.M.J.S. and Panton, W.J. (1993).Decline of Callitris intratropica in theNorthern Territory: implications for pre-and post-European colonisation fire regimes.Journal of Biogeography 20, 373–381.
Bowman, D.M.J.S., Wilson, B.A. and Hooper,R.J. (1988). Response of Eucalyptus forest andwoodland to four fire regimes, Munmarlary,Northern Territory, Australia. Journal ofEcology 76, 215–232.
Bowman, D.M.J.S. Latz, P.K. and Panton, W.J.(1994a). Pattern and change in an Acaciaaneura shrubland and Triodia hummockgrassland mosaic on rolling hills in centralAustralia. Australian Journal of Botany 43,25–37.
Bowman, D. M. J. S., Woinarski, J. C. Z., &Russell-Smith, J. (1994b). Environmentalrelationships of Orange-footed ScrubfowlMegapodius reinwardt nests in the NorthernTerritory. Emu 94, 181–185.
Bradley, J. J. (1995). Fire: emotion and politics. InD.B. Rose (Ed.) Country in flames. Proceedingsof the 1994 symposium on biodiversity and fire inNorth Australia. (pp. 25–32). Canberra:Department of the Environment, Sport andTerritories.
Braithwaite, L. W., Clayton, M., MacLean, L., &Parker, B. S. (1984). Vertebrate fauna of a144-ha water catchment within eucalypt forestbeing harvested for woodpulp at Eden, south-eastern New South Wales. CSIRO Wildlifeand Rangelands Research.
Braithwaite, R. W. (1985). Fire and fauna. In R.W. Braithwaite (Ed.), Kakadu Fauna Survey.Final report to Australian National Parks andWildlife Service. (pp. 634–650). Darwin:CSIRO.
Braithwaite, R.W. (1991). Aboriginal fire regimesof monsoonal Australia in the 19th century.Search 22, 247–249.
Braithwaite, R.W. and Estbergs, J.A. (1985). Firepatterns and woody vegetation trends in theAlligator Rivers region of northern Australia.In J.C. Tothill and J.J. Mott (Eds.) Ecologyand management of the world’s savannas (pp.359–364). Canberra: Australian Academy ofScience.
86
Australia’s Biodiveristy - Responses to Fire

Braithwaite, R., & Estbergs, J. (1987). Fire-birdsof the Top End. Australian Natural History22, 299–302.
Bramwell, M., Pyke, G., Adams, C., & Coontz, P.(1992). Habitat use by Eastern Bristlebirdsin Barren Grounds Nature Reserve. Emu 92,117–121.
Brickhill, J. (1980). Striated Grasswren Amytornisstriatus. In C. Haigh (Ed.), Endangeredanimals of New South Wales (p. 68). Sydney:NSW National Parks and Wildlife Service.
Brickhill, J. (1987) The conservation status ofmalleefowl in New South Wales.M.Nat.Res.Sc., University of New England.
Brooker, L. C., & Brooker, M. G. (1994). A modelfor the effects of fire and fragmentation onthe population viability of the SplendidFairy–wren. Pacific Conservation Biology 1,344–358.
Brooker, M. G. (1988). Some aspects of thebiology and conservation of the Thick-billedGrasswren Amytornis textilis in the Shark Bayarea, Western Australia. Corella 12, 101–108.
Brooker, M. G., & Rowley, I. (1991). Impact ofwildfire on the nesting behaviour of birds inheathland. Wildlife Research 18, 249–263.
Brooker, M. G., Ridpath, M. G., Estbergs, A. J.,Bywater, J., Hart, D. S., & Jones, M. S. (1979).Bird observations on the north-westernNullabor Plain and neighbouring regions,1967–1978. Emu 79, 176–190.
Brooker, M. G., Braithwaite, R. W., & Estbergs, J.A. (1990). Foraging ecology of someinsectivorous and nectarivorous species ofbirds in forests and woodlands of theWet-Dry tropics of Australia. Emu 90,215–230.
Brothers, N. P. (1983). Seabird Islands: No. 136.Actaeon Island, Tasmania. Corella 7, 89–90.
Brothers, N. P., & Skira, I. J. (1987). SeabirdIslands: No. 173. Chappell Island, FurneauxGroup, Tasmania. Corella 11, 81–82.
Brothers, N. P., & Skira, I. J. (1988). SeabirdIslands: No. 185. Little Dog Island, FurneauxGroup, Tasmania. Corella 12, 85–86.
Brouwer, J., and Garnett, S. (Eds) (1990).Threatened birds of Australia: an annotated list.Royal Australasian Ornithologists Union:Melbourne.
Brown, P. B., & Wilson, R. I. (1981). A survey ofthe Orange-bellied Parrot Neophemachrysogaster in Tasmania, Victoria and SouthAustralia. National Parks and Wildlife
Service, Tasmania.Brown, P. B., & Wilson, R. I. (1984). The
Orange-bellied Parrot. In R. H. Groves &W. D. L. Ride (Eds.), Species at risk (pp.106–116). Canberra: Australian Academy ofScience.
Bryant, S. L. (1991). The Ground Parrot, Pezoporuswallicus, in Tasmania: distribution, density andconservation status. Scientific report no. 91/1.Parks, Wildlife & Heritage, Tasmania.
Bryant, S. L. (1992). The Ground Parrot and ageof vegetation in Tasmania. In L. Joseph(Ed.), Issues in the conservation of parrots inAustralasia and Oceania: challenges toconservation biology. (pp. 42–45). Melbourne:Royal Australasian Ornithologists Union.
Bryant, S. L. (1994). Habitat and potential diet ofthe Ground Parrot in Tasmania. Emu 94,166–171.
Burbidge, A. (1985). Fire and mammals inhummock grasslands of the arid zone. In J.R. Ford (Ed.), Fire ecology and management inWestern Australian ecosystems (pp. 91–94).Perth: Western Australian Institute ofTechnology.
Burbidge, A. H., & Pedler, L. (1993). Conservationstatus of the Nullabor Quail-thrush. WorldWide Fund for Nature.
Burbidge, A. H., & Pedler, L. (1996). Habitat ofthe threatened Nullabor Quail-thrush.Wildlife Research, 23.
Burbidge, A. A., Folley, G. L., & Smith, G. T.(1986). The Noisy Scrub-bird. WesternAustralian Wildlife Management Program 2.Department of Conservation and LandManagement.
Burbidge, A. H., Watkins, D., & McNee, S.(1989). Conservation of the Ground Parrot inWestern Australia. Final report (Project 118).World Wildlife Fund.
Burbidge, A. H., McNee, S., Newbey, B., & Rolfe,J. (1990). Conservation of the Ground Parrot inWestern Australia. Supplementary report:project 118. World Wildlife Fund.
Calder, T.G., Golding, B.G. and Manderson, A.D.(1983). Management for arboreal species inthe Wombat State Forest. Graduate Schoolof Environmental Science. EnvironmentalReport no. 16. Clayton: Monash University.
Cale, P. G., & Burbidge, A. H. (1993). Researchplan for the Western Ground Parrot, WesternWhipbird and Western Bristlebird. AustralianNational Parks and Wildlife Service.
87
Biodiveristy and bushfires

Campbell, A. G. (1937). Birds of Wilson’sPromontory. Emu 37, 157.
Carpenter, G., & Matthew, J. (1986). The birds ofBilliatt Conservation Park. South AustralianOrnithologist 30, 29–37.
Carter, T. (1923a). Birds of Broome Hill district.Part II. Emu 23, 223–235.
Carter, T. (1923b). Birds of the Broome Hilldistrict. Part I. Emu 23, 125–142.
Carter, T. (1924). Birds of the Broome Hilldistrict. Part III. Emu 23, 306–318.
Catling, P. C., & Newsome, A. E. (1981).Responses of the Australian vertebrate faunato fire. In A. M. Gill, R. H. Groves, & I. R.Noble (Eds.), Fire and the Australian biota(pp. 273–310). Canberra: AustralianAcademy of Science.
Chambers, W. (1983). Birds at Urquhart’s Bluff:before and after the fire. Geelong Naturalist20, 50–51.
Chandler, L. G. (1973). In the wake of a bushfire.Wildlife in Australia 10, 140–141.
Chatto, R. (1995). The effects of fire on abreeding colony of Australian Pelicans.Corella 19, 70.
Cheal, P. D., Day, J. C., & Meredith, C. W.(1979). Fire in the national parks of north-westVictoria. National Parks Service.
Chisholm, A. H. (1922). The “lost” ParadiseParrot. Emu 22, 4–17.
Chisholm, A. H. (1945). Birds of the Gilbertdiary, Part 2. Emu 44, 183–200.
Christensen, P. (1974). The concept of faunapriority areas. In Third Fire EcologySymposium (pp. 66–73). Melbourne: ForestsCommission, Victoria.
Christensen, P., & Abbott, I. (1989). Impact of firein the eucalypt forest ecosystem of southernWestern Australia: a critical review.Australian Forestry 52, 103–121.
Christensen, P. E., & Kimber, P. C. (1975). Effectof prescribed burning on the flora and faunaof south-west Australian forests. Proceedingsof the Ecological Society of Australia 7, 85–107.
Christensen, P., Recher, H., & Hoare, J. (1981).Responses of open forests (dry sclerophyllforests) to fire regimes. In A. M. Gill, R. H.Groves, & I. R. Noble (Eds.), Fire and theAustralian biota (pp. 367–393). Canberra:Australian Academy of Science.
Christensen, P. E. S., Wardell-Johnson, G., &Kimber, P. (1985). Birds and fire insouthwestern forests. In A. Keast, H. F.Recher, H. Ford, & D. Saunders (Eds.), Birdsof eucalypt forests and woodlands: ecology,conservation, management (pp. 291–299).Chipping Norton: Surrey Beatty.
Christidis, L. and Boles, W.E. (1994), Thetaxonomy and species of birds of Australia and itsterritories., Melbourne: Royal AustralasianOrnithologists Union.
Clout, M. N. (1989). Foraging behaviour ofGlossy Black-cockatoos. Australian WildlifeResearch 16, 467–473.
Coate, K. (1985). Black Honeyeaters feeding onash. Western Australian Naturalist 16, 51–52.
Cooper, R. P. (1974). The avifauna of Wilson’sPromontory. Part 1. Australian Bird Watcher5, 137–174.
Cowley, R.D. (1971). Birds and forestmanagement. Australian Forestry 3, 234–250.
Cowley, R. D. (1974). Effects of prescribedburning on birds of the mixed species forestsof West Central Victoria. In Third FireEcology Symposium (pp. 58–65). Melbourne:Forests Commission, Victoria.
Cowley, R. D., Heislers, A., & Ealey, E. H. H.(1969). Effects of fire on wildlife. Victoria’sResources 11, 18–22.
Crawford, D. N. (1972). Birds of Darwin area,with some records from other parts ofNorthern Territory. Emu 72, 131–148.
Crawford, D. N. (1979). Effects of grass and fireson birds in the Darwin area, NorthernTerritory. Emu 79, 150–152.
Crowley, G. M. (1995). Fire on Cape YorkPeninsula. Cape York Peninsula Land UseStudy.
CSIRO (1976). A survey of the fauna of the lowerMcArthur River Region, Northern Territory.Report to Mimets Pty Ltd. (CSIRO Divisionof Wildlife Research; Canberra)
Curry, G. N. (1991). The influence of proximityto plantation edge on diversity andabundance of bird species in an exotic pineplantation in north-eastern New SouthWales. Wildlife Research 18, 299–314.
Danks, A. (1991). The role of corridors in themanagement of an endangered passerine. InD. A. Saunders & R. J. Hobbs (Eds.), Natureconservation 2: the role of corridors (pp.291–296). Chipping Norton: Surrey Beatty.
88
Australia’s Biodiveristy - Responses to Fire

Davidson, I., & Chambers, L. (1991). Vegetationmanagement for Superb Parrot foraging habitatin Victoria. Victorian Department ofConservation and Environment.
Davidson, I., & Robinson, D. (1992).Grey-crowned Babbler Pomatostomus temporalis.Department of Conservation and NaturalResources.
Debus, S. J. S., & Czechura, G. V. (1988). TheRed Goshawk Erythrotriorchis radiatus: areview. Australian Bird Watcher 12, 175–199.
Dedman, V. (1983a). G. F. N. C. OtwayRegeneration Survey – Progress report.Geelong Naturalist 20, 52–56.
Dedman, V. (1983b). G.F.N.C. Otwaysregeneration survey – 2nd progress report.Geelong Naturalist 20, 98–100.
Dedman, V. (1983c). Mammals and birds after thefires: Forest Road, Anglesea. GeelongNaturalist 20, 27.
Dedman, V. (1983d). Moggs Creek: eight dayslater. Geelong Naturalist 20, 22–25.
Dedman, V. (1983e). Notes on the effects of theAsh Wednesday fire on the Ironbark Basin,Point Addis. Geelong Naturalist 20, 25–26.
Dedman, V. (1984). Otways regeneration survey.Third progress report. Geelong Naturalist 21,89–92.
Department of Environment, Sport and Territories(1996). Fire and biodiversity: the effects andeffectiveness of fire management. Proceedingsof the Conference held 8–9 October 1994,Footscray, Melbourne. Biodiversity Series,Paper No. 8. Canberra: Department ofEnvironment, Sport and Territories
Dickinson, K. J. M., Wall, L. E., & Wilson, R. I.(1986). Birds in a partly clearfelled dryeucalypt forest on dolerite in southeasternTasmania. Papers and Proceedings of the RoyalSociety of Tasmania 120, 39–49.
Du Guesclin, P., Smith, S., O’Shea, B., & Debbis,C. (1995). “Brushing for bristles”: habitatcorridors for the Rufous Bristlebird. In A.Bennett, G. Backhouse, & T. Clark (Eds.),People and nature conservation: perspectives onprivate land use and endangered species recovery(pp. 163–165). Chipping Norton: SurreyBeatty.
Emison, W. B., & Bren, W. M. (1989). Commonbirds of the mallee, northwestern Victoria. InJ. C. Noble & R. A. Bradstock (Eds.),Mediterranean landscapes in Australia: malleeecosystems and their management (pp. 221–242).Melbourne: CSIRO.
Emison, W. B., Beardsell, C. M., Norman, F. I., &Loyn, R. H. (1987). Atlas of Victorian birds.Melbourne: Department of Conservation,Forests and Lands, and Royal AustralasianOrnithologists Union.
Ferrier, S. (1985). Habitat requirements of a rarespecies, the Rufous Scrub-bird. In A. Keast,H. F. Recher, H. Ford, & D. Saunders(Eds.), Birds of eucalypt forests and woodlands:ecology, conservation, management (pp.241–248). Chipping Norton: Surrey Beatty.
Fitzherbert, J. C., & Baker-Gabb, D. J. (1988).Australasian grasslands and their threatenedavifauna. In P. D. Goriup (Ed.), Ecology andconservation of grassland birds (pp. 227–250).Cambridge: International Council for BirdPreservation.
Flannery, T. F. (1990). Pleistocene faunal loss:implications of the aftershock for Australia’spast and future. Archaeology in Oceania 25,45–67.
Flannery, T. F. (1994). The future-eaters: anecological history of the Australasian lands andpeople. Sydney: Reed.
Forshaw, J. M. (1981). Australian Parrots.Melbourne: Lansdowne Press.
Fowler, S. (1945). The fire on Mondrain Island.Emu 44, 334–335.
Fox, A. (1978). The ’72 fire of Nadgee NatureReserve. Parks & Wildlife 2, 5–24.
Fox, B.J. (1982). Fire and mammalian secondarysuccession in an Australian coastal heath.Ecology 63, 1332–1341.
Friend, G.R. (1993). Impact of fire on smallvertebrates in mallee woodlands andheathlands of temperate Australia: a review.Biological Conservation 65, 99–114.
Friend, G.R. (1995). Fire and invertebrates - areview of research methodology and thepredictability of post-fire response patterns.CALMScience Supplement 4, 165–174.
Frith, H. J. (1962). Conservation of theMallee-fowl (Leipoa ocellata, Gould). CSIROWildlife Research 7, 33–49.
Garnett, S. (1987). Seabird Islands: No. 171.Kusamet Island, Torrest Strait, Queensland.Corella 11, 77–78.
Garnett, S. (1992a). The action plan for Australianbirds. Canberra: Australian National Parksand Wildlife Service.
Garnett, S. (1992b). Threatened and extinct birds ofAustralia. Melbourne: Royal AustralasianOrnithologists Union.
89
Biodiveristy and bushfires

Garnett, S., & Bredl, R. (1985). Birds in thevicinity of Edward River Settlement. Part II.Discussion, references, list of passerines.Sunbird 15, 25–40.
Garnett, S. T., & Crowley, G. M. (1994). Theecology and conservation of theGolden-shouldered Parrot. Cape YorkPeninsula Land Use Strategy.
Garnett, S., & Crowley, G. (1995a). The declineof the Black Treecreeper Climacteris picumnusmelanota on Cape York Peninsula. Emu 95,66–68.
Garnett, S., & Crowley, G. (1995b). Feedingecology of Hooded Parrots Psephotusdissimilis during the early wet season. Emu95, 54–61.
Garnett, S., Crowley, G., Duncan, R., Baker, N.,& Doherty, P. (1993). Notes on live NightParrot sightings in north-westernQueensland. Emu 93, 292–296.
Gibbons, P. (1994). Sustaining key old-growthcharacteristics in native forests used forwood production: retention of trees withhollows. In T.W. Norton and S.R. Dovers(Eds.) Ecology and sustainability of southerntemperate ecosystems (pp. 59–84). Canberra:CSIRO.
Gibson, D. F. (1986). A biological survey of theTanami Desert in the Northern Territory.Conservation Commission of the NorthernTerritory.
Gill, A. M. (1981). Post-settlement fire history inVictorian landscapes. In A. M. Gill, R. H.Groves, & I. R. Noble (Eds.), Fire and theAustralian biota (pp. 77–98). Canberra:Australian Academy of Science.
Gill, A. M. (1990). Fire management of malleelands for species conservation. In J. C.Noble, P. J. Joss, & G. K. Jones (Eds.), Themallee lands: a conservation perspective (pp.202–205). Melbourne: CSIRO.
Gill, A. M. (1996). How fires affect biodiversity.In DEST (Eds.), Fire and biodiversity: theeffects and effectiveness of fire management.Proceedings of the conference held 8–9 October1994, Footscray, Melbourne (pp. 47–55(&123–124)). Canberra: Department of theEnvironment, Sport and Territories.
Gill, A. M., Groves, R. H. and Noble, I. R.(1981). Fire and the Australian biota.Canberra: Australian Academy of Science.
Good, R.B. (1981). The role of fire inconservation reserves. In A. M. Gill, R. H.Groves, & I. R. Noble (Eds.), Fire and theAustralian biota (pp. 529–550). Canberra:Australian Academy of Science.
Green, R. H. (1982). The activity and movementof fauna in compartment 2, MaggsMountain, Tasmania, in the first five years offorest regeneration. Records of the QueenVictoria Museum 75, 1–31.
Green, R. H., & McGarvie, A. M. (1971). Thebirds of King Island. Records of the QueenVictoria Museum 40, 1–42.
Griffin, G.F. (1984). Fire, vegetation, and man. InE.C. Saxon (Ed.) Anticipating the inevitable: apatch-burn strategy for fire management atUluru (Ayers Rock - Mt Olga) National Park(pp. 7–12). Melbourne: CSIRO.
Griffin, G.F. and Hodgkinson, K.C. (1986). Theuse of fire for the management of mulga landvegetation in Australia. In P. Sattler (Ed.)The mulga lands. (pp.93–97). Brisbane: RoyalSociety of Queensland.
Griffin, G.F., Price, N.F. and Portlock, H.F.(1983). Wildfires in the central Australianrangelands, 1970–1980. Journal ofEnvironmental Management 17, 311–323.
Hallam, S. J. (1975). Fire and hearth: a study ofAboriginal usage and European usurpation insouth-western Australia. Canberra: AustralianInstitute of Aboriginal Studies.
Harrington, G. N., & Sanderson, K. D. (1994).Recent contraction of wet sclerophyll forestin the wet tropics of Queensland due toinvasion by rainforest. Pacific ConservationBiology 1, 319–327.
Harris, C. R. (1990). The history of mallee landuse: Aboriginal and European. In J. C.Noble, P. J. Joss, & G. K. Jones (Eds.), Themallee lands: a conservation perspective (pp.147–151). Melbourne: CSIRO.
Haynes, C. D. (1985). The pattern and ecology ofmunwag: traditional Aboriginal fire regimesin north-central Arnhemland. Proceedings ofthe Ecological Society of Australia 13, 203–214.
Haynes, C. D. (1991). Use and impact of fire. InC. D. Haynes, M. G. Ridpath and M. A. J.Williams (Eds.), Monsoonal Australia:landscape, ecology and man in the northernlowlands. (pp. 61–71). Balkema: Rotterdam.
Head, L. (1989). Prehistoric Aboriginal impactson Australian vegetation: an assessment ofthe evidence. Australian Geographer 20,37–46.
90
Australia’s Biodiveristy - Responses to Fire

Head, L. (1993). Unearthing prehistoric culturallandscapes: a view from Australia.Transactions of the Institute of BritishGeography 18, 481–499.
Head, L., O’Neill, A.L., Martick, J.K. andFullager, R.L.K. (1992). A comparison ofAboriginal and pastoral fires in the north-west Northern Territory. In I. Moffatt and A.Webb (Eds.), Conservation and developmentissues in northern Australia. Darwin: NorthAustralia Research Unit.
Hemsley, J. H. (1967). Bushfire – S.E. Tasmania 7thFebruary, 1967. Some aspects of the fire inrelation to animal and plant life. TasmanianForestry Commission.
Hewish, M. (1983). The effect of a wildfire onbirdlife in a eucalypt forest: a preliminaryreport on the Lerdederg Gorge seven weeksafter the Wombat State Forest fire. GeelongNaturalist 20, 3–16.
Higgs, P. and Fox, B.J. (1993). Interspecificcompetition: a mechanism for rodentsuccession after fire in wet heathland.Australian Journal of Ecology 18, 193–201.
Hill, R.S. (Ed.) (1994). History of Australianvegetation: Cretaceous to Recent. Cambridge:Cambridge University Press.
Hodgson, A., & Heislers, A. (1972). Some aspects ofthe role of forest fire in South-eastern Australia.Forests Commission, Victoria.
Holmes, G. (1988). Eastern Bristlebird: summaryconservation statement for northern populations.QNPWS.
Holmes, G. (1989). Eastern Bristlebird. Speciesmanagement plan for northern populations.Queensland NPWS and NSW NPWS.
Hood, J. B. (1941). Birds and bushfires. SouthAustralian Ornithologist 15, 125–127.
Hopkins, A.J.M. (1985a). Fire in the woodlandsand associated formations of the semi-aridregion of south-western Australia. In J. R.Ford (Eds.), Fire ecology and management inWestern Australian ecosystems (pp. 83–90).Perth: Western Australian Institute ofTechnology.
Hopkins, A. J. M. (1985b). Planning the use offire on conservation lands in south-westernAustralia. In J. R. Ford (Ed.), Fire ecology andmanagement in Western Australian ecosystems(pp. 203–208). Perth: Western AustralianInstitute of Technology.
Hopkins, A. J. M., & Smith, G. T. (1996). Fire:effects and management implications. In A.J. M. Hopkins & G. T. Smith (Eds.), Thenatural history of Two Peoples Bay NatureReserve Perth: Department of Conservationand Land Management (CALMScienceSupplement no. 5).
How, R. A., Dell, J., & Humphreys, W. F. (1987).The ground vertebrate fauna of coastal areasbetween Busselton and Albany, WesternAustralia. Records of the Western AustralianMuseum 13, 553–574.
Hughes, P., & Hughes, B. (1991). Notes on theBlack-breasted Button-quail at Widgee,Queensland. Australian Bird Watcher 14,113–118.
Hull, A. F. B. (1922). A visit to the Archipelago ofthe Recherche S.W. Australia. Emu 21,277–289.
Hutchins, B. R. (1988). Black Honeyeatersfeeding among charcoal and ash. SouthAustralian Ornithologist 30, 160.
Inions, G. (1985). The interactions betweenpossums, habitat trees and fire. B.Sc. (Hons)thesis. Australian National University,Canberra.
Inions, G.B., Tanton, M.T. and Davey, S.M.(1989). The effect of fire on the availabilityof hollows in trees used by the CommonBrushtail Possum, Trichosurus vulpecula Kerr1872, and Ringtail Possum, Pseudocheirusperegrinus Boddarts 1785. Australian WildlifeResearch 16, 449–458.
Jackson, W.D. (1968). Fire, air, water, earth - anelemental ecology of Tasmania. Proceedings ofthe Ecological Society of Australia 3, 9–16.
Johnson, R.W. and Purdie, R.W. (1981). The roleof fire in the establishment and managementof agricultural systems. In A. M. Gill, R. H.Groves, & I. R. Noble (Eds.), Fire and theAustralian biota (pp. 497–528). Canberra:Australian Academy of Science.
Jones, R. (1969). Fire-stick farming. AustralianNatural History 16, 224–228.
Jones, R. (1980). Hunters in the Australian coastalsavanna. In D. R. Harris (Ed.), HumanEcology in Savanna Environments London:Academic Press.
Jones, R. (1995). Mindjongrok: legacy of thefirestick. In D.B. Rose (Ed.) Country inflames. Proceedings of the 1994 symposium onbiodiversity and fire in North Australia. (pp.11–18). Canberra: Department of theEnvironment, Sport and Territories.
91
Biodiveristy and bushfires

Jordan, R. (1984a). The Eastern Bristlebird.Effects of fire on a population. RAOU Report11, 30.
Jordan, R. (1984b). The Ground Parrot – Effectof fire on a population. RAOU Report 11,28–29.
Jordan, R. (1987a). The Barren Grounds rollingbird survey. RAOU Report 27, 15–18.
Jordan, R. (1987b). The Ground Parrot in BarrenGrounds Nature Reserve. RAOU Report 27,19–23.
Jordan, R. (1987c). The Southern Emu–wren inBarren Grounds. RAOU Report 27, 24.
Jordan, R. (1988). Population changes of somecommon insectivore species in woodlandnear the wardens house. RAOU Report 51,16–18.
Joseph, L. (1982). The Glossy Black–cockatoo onKangaroo Island. Emu 82, 46–49.
Kavanagh, R. P. (1990). Survey of Powerful andSooty Owls in south–eastern New South Wales.Final report (Project 120). World WildlifeFund (Australia).
Kavanagh, R. P. (1991). The target speciesapproach to wildlife management: glidersand owls in the forests of southeastern NewSouth Wales. In D. Lunney (Ed.),Conservation of Australia’s forest fauna (pp.377–383). Mosman: Royal ZoologicalSociety of NSW.
Kershaw, A.P. (1986). Climatic change andAboriginal burning in north-east Australiaduring the last two glacials. Nature 322,47–49.
Kimber, P. C. (1974). Some effects of prescribedburning on Jarrah Forest birds. In Third FireEcology Symposium (pp. 49–57). Melbourne:Forests Commission, Victoria.
Kimber, R.G. (1982). Black lightning: Aboriginesand fire in central Australia and the Westerndesert. Archaeology in Oceania 18, 38–45.
Lane, S. G. (1976). Seabird islands: No. 18.Broughton Island, New South Wales.Australian Bird Bander 14, 10–13.
Latz, P. (1995). Fire in the desert: increasingbiodiversity in the short-term, decreasing itin the long term. In D.B. Rose (Ed.) Countryin flames. Proceedings of the 1994 symposium onbiodiversity and fire in North Australia. (pp.77–86). Canberra: Department of theEnvironment, Sport and Territories.
Leigh, J., Boden, R. and Briggs, J. (1984). Extinctand endangered plants of Australia. Melbourne:MacMillan.
Lewis, H.T. (1985). Burning the “Top End”:kangaroos and cattle. In J.R. Ford (Ed.). Fireecology and management in Western Australianecosystems (pp. 21–31). Perth: WesternAustralian Institute of Technology.
Lewis, H. T. (1989). Ecological and technologicalknowledge of fire: Aborigines versus parkrangers in northern Australia. AmericanAnthropologist 91, 940–961.
Lindenmayer, D.B., Norton, T.W., and TantonM.T. (1990). Differences between wildfireand clearfelling on the structure of montaneash forests of Victoria and their implicationsfor fauna dependant on tree hollows.Australian Forestry 53, 61–68.
Lindenmayer, D.B., Cunningham, R.B., Donnelly,C.F., Tanton, M.T. and Nix, H.A. (1993).The abundance and development of cavitiesin Eucalyptus trees: a case study in themontane forests of Victoria, southeasternAustralia. Forest Ecology and Management 60,77–104.
Lord, E. A. R. (1936). Notes on swifts. Emu 35,216–218.
Loyn, R. H. (1985a). Bird populations insuccessional forests of Mountain AshEucalyptus regnans in central Victoria. Emu85, 213–230.
Loyn, R. H. (1985b). Ecology, distribution anddensity of birds in Victorian forests. In A.Keast, H. F. Recher, H. Ford, & D. Saunders(Eds.), Birds of eucalypt forests and woodlands:ecology, conservation, management (pp. 33–46).Chipping Norton: Surrey Beatty.
Loyn, R. H., Traill, B. J., & Triggs, B. E. (1986).Prey of Sooty Owls in East Gippsland beforeand after fire. Victorian Naturalist 103,147–149.
Loyn, R. H., Cameron, D. G., Traill, B. J., Sloan,J. F., Malone, B. S., Schulz, M., Earl, G. E.,& Triggs, B. E. (1992a). Flora and fauna of theCooaggalah Forest Block, East Gippsland,Victoria. Ecological Survey Report 20.Department of Conservation andEnvironment.
92
Australia’s Biodiveristy - Responses to Fire

Loyn, R. H., Hewish, M. J., & Considine, M.(1992b). Short-term effects of fuel reductionburning on bird populations in WombatState Forest. In K. Tolhurst & D. Flinn(Eds.), Ecological impact of fuel reductionburning in dry sclerophyll forest: first progressreport (pp. 5.1–5.11). Melbourne:Department of Conservation andEnvironment (Research Report no. 349).
Lucas, D., & Russell-Smith, J. (1993). Traditionalresources of the South Alligator floodplain:utilisation and management. Australian NatureConservation Agency.
Lucas, K. and Lucas, D. (1993). Aboriginal firemanagement of the Woolwonga wetlands inKakadu National Park. Report to AustralianNature Conservation Agency.
Luke, R. H. and McArthur, A. G. (1978). Bushfiresin Australia. Canberra: AustralianGovernment Publishing Service.
Lunt, I.D. (1991). Management of remnantlowland grasslands and grassy woodlands fornature conservation: a review. VictorianNaturalist 108, 56–66.
MacGillivray, W. D. K. (1910). Along the GreatBarrier Reef. Emu 10, 216–233.
Mackowski, C. M. (1984). The ontogeny ofhollows in Blackbutt, Eucalyptus nilularis andits relevance to the management of forestsfor possums, gliders and timber. In A.P.Smith and I. D. Hume (Eds ). (pp. 553–567)Chipping Norton: Surrey Beatty.
Main, A. R. (1981). Fire tolerance of heathlandanimals. In R. L. Specht (Ed.), Ecosystems ofthe World. 9B. Heathlands and relatedshrublands. Analytical studies. Amsterdam:Elsevier.
Majer, J. D. (1984). Short-term responses of soiland litter invertebrates to a cool autumnburn in jarrah (Eucalyptus marginata) forestin Western Australia. Pedobiologia 26,299–247.
Majer, J. D. (1985). Fire effects on invertebratefauna of forest and woodland. In J. R. Ford(Ed.), Fire ecology and management in WesternAustralian ecosystems (pp. 103–106). Perth:Western Australian Institute of Technology.
Marchant, S. (1985). Breeding of the EasternYellow Robin. In A. Keast, H. F. Recher, H.Ford, & D. Saunders (Eds.), Birds of eucalyptforests and woodlands: ecology, conservation,management (pp. 231–240). ChippingNorton: Surrey Beatty.
Masters, P. (1993). The effects of fire-drivensuccession and rainfall on small mammals inspinifex grassland at Uluru National Park,Northern Territory. Wildlife Research 20,803–813.
Matthew, J. (1994). The status, distribution andhabitat of the Slender-billed ThornbillAcanthiza iredalei in South Australia. SouthAustralian Ornithologist 32, 1–19.
Mawson, P. R., & Long, J. L. (1994). Size and ageparameters of nest trees used by four speciesof parrot and one species of cockatoo insouth-west Australia. Emu 94, 149–155.
May, S. A. (1994). Sustaining key old-growthcharacteristics in native forests used forwood production: the significance of exoticvertebrate predators. In T.W. Norton and S. R. Dovers (Eds.), Ecology and sustainabilityof southern temperate ecosystems (pp. 51–58).Canberra: CSIRO.
McCaw, W. L., Burrows, N. D., Friend, G. R., &Gill, A. M. (1995). Landscape fires ’93:proceedings of an Australian bushfireconference, Perth, Western Australia, 27–29September 1993. CALMScience Supplement 4
McCulloch, E. M. (1966). Swifts and bushfires.Emu 65, 290.
McDougall, K.L. (1989). The re-establishment ofThemeda triandra (Kangaroo Grass):implications for the restoration of grassland.Melbourne: Department of Conservation,Forests and Lands.
McFarland, D. C. (1988). The composition,microhabitat use and response to fire of theavifauna of subtropical heathlands in ColoolaNational Park, Queensland. Emu 88,249–257.
McFarland, D. C. (1989). The Ground ParrotPezoporus wallicus (Kerr) in Queensland:habitat, biology and conservation. Departmentof Conservation, Parks & Wildlife,Queensland.
McFarland, D. C. (1991). The biology of theGround Parrot, Pezoporus wallicus, inQueensland. III. Distribution andabundance. Wildlife Research 18, 199–213.
McFarland, D. C. (1992). Fire and themanagement of ground parrot habitat. In B. R. Roberts (Ed.), Fire Research in ruralQueensland (pp. 483–495). Toowomba:University of Southern Queensland.
93
Biodiveristy and bushfires

McFarland, D. C. (1993). Fire and birdconservation. In C. P. Catterall, P. V.Driscoll, K. Hulsman, D. Muir, & A. Taplin(Eds.), Birds and their habitats: status andconservation in Queensland (pp. 41–44). St Lucia: Queensland Ornithological SocietyInc.
McFarland, D. C. (1994). Notes on the BrushBronzewing Phaps elegans and SouthernEmu-wren Stipiturus malachurus in CooloolaNational Park. Sunbird 24, 14–17.
McIlroy, J.C. (1978). The effects of forestrypractices on wildlife in Australia: a review.Australian Forestry 41, 78–94.
McKean, J. L., & Martin, K. C. (1985).Distribution and status of the CarpentarianGrasswren Amytornis dorotheae. ConservationCommission of the Northern Territory.
McKean, J. L., & Martin, K. C. (1989).Distribution and status of the CarpentarianGrass-wren Amytornis dorotheae. NorthernTerritory Naturalist 11, 12–19.
McKenzie, N. L. and Belbin, L. (1991).Kimberley rainforest communities: reserverecommendations and managementconsiderations. In N. L. McKenzie, R. B.Johnson and P. G. Kendrick (Eds.) Kimberleyrainforests of Australia. (pp. 453–468).Chipping Norton: Surrey Beatty.
McLaughlin, J. (1992). The floristic and structuralfeatures of Black-eared Miner Manorinamelanotis habitat. RAOU Report No. 84. RoyalAustralasian Ornithologists Union.
McLaughlin, J. (1994). Searches for the Black-earedMiner Manorina melanotis in the VictorianMurray Mallee. RAOU Report No. 93. RoyalAustralasian Ornithologists Union.
McNee, S. (1986). Surveys of the Western Whipbirdand Western Bristlebird in Western Australia.RAOU Report No. 18. Royal AustralasianOrnithologists Union.
Menkhorst, P. W., & Bennett, A. F. (1990).Vertebrate fauna of mallee vegetation insouthern Victoria. In J. C. Noble, P. J. Joss,& G. K. Jones (Eds.), The mallee lands: aconservation perspective (pp. 39–53).Melbourne: CSIRO.
Meredith, C. W. (1982). The research input tofire management in natural areas withparticular reference to wildlife research inthe semi-arid lands of Victoria and NewSouth Wales. In A. Heislers, P. Lynch, & B.Walters (Eds.), Fire ecology in semi-arid lands(pp. 10pp). Deniliquin: CSIRO.
Meredith, C. W. (1983). Fire and birds. Theresult of two studies and their relevance tofuel reduction burning. In E. H. M. Ealey(Eds.), Fighting fire with fire (pp. 193–202).Melbourne: Graduate School ofEnvironmental Science Monash University.
Meredith, C. W. (1984a). The Ground Parrot.RAOU Conservation Statement, 1.
Meredith, C. W. (1984b). Management of theGround Parrot Pezoporus wallicus in Victoria.Fisheries & Wildlife Division, Victoria.
Meredith, C. W. (1984c). Recent records of theGround Parrot Pezoporus wallicus in westernVictoria. Geelong Naturalist 21, 3–4.
Meredith, C. W. (1988). Fire in the Victorianenvironemnt - a discussion paper. ConservationCouncil of Victoria.
Meredith, C. W., & Isles, A. C. (1980). A study ofthe Ground Parrot (Pezoporus wallicus) inVictoria. report No. 304. EnvironmentalStudies Division of the Ministry forConservation, Victoria.
Meredith, C. W., & Jaremovic, R. (1990). Currentstatus and management of the Ground Parrot inVictoria. Arthur Rylah Institute forEnvironmental Research.
Meredith, C. W., Gilmore, A. M., & Isles, A. C.(1984). The Ground Parrot (Pezoporuswallicus Kerr) in south-eastern Australia: afire-adapted species? Australian Journal ofEcology 9, 367–380.
Merilees, D. (1968). Man the destroyer: lateQuaternary changes in the Australianmarsupial fauna. Journal of the Royal Society ofWestern Australia 51, 1–24.
Milledge, D. R., & Palmer, C. L. (1990). The SootyOwl in Mountain Ash forests in the VictorianCentral Highlands. Department ofConservation and Environment.
Milledge, D. R., Palmer, C. L., & Nelson, J. L.(1991). “Barometers of change”: thedistribution of large owls and gliders inMountain Ash forests of teh VictorianCentral Highlands and their potential asmanagement indicators. In D. Lunney (Ed.),Conservation of Australia’s Forest Fauna (pp.53–65). Sydney: Royal Zoological Society ofNSW.
Milligan, A. W. (1904). Notes on a trip to theWongan Hills, Western Australia. Emu 4,2–11.
94
Australia’s Biodiveristy - Responses to Fire

Mitchell, T.M. (1838). Three expeditions into theinterior of eastern Australia, with descriptions ofrecently explored regions of Australia Felix, andthe present colony of New South Wales London:T. & W. Boone.
Nelson, J. L., & Morris, B. J. (1994). Nestingrequirements of the Yellow-tailedBlack-cockatoo, Calyptorhynchus funereus, inEucalyptus regnans forest, and implicationsfor forest management. Wildlife Research 21,267–278.
Newsome, A. E., McIlroy, J., & Catling, P. (1975).The effects of extensive wildfire onpopulations of twenty ground vertebrates insouth-east Australia. Proceedings of theEcological Society of Australia 9, 107–123.
Nichols, O. G., & Muir, B. (1989). Vertebrates ofthe jarrah forest. In B. Dell, J. J. Havel, & N.Malajczuk (Eds.), The Jarrah Forest (pp.133–153). Dordrecht: Kluwer.
Nicholson, P. H. (1981). Fire and the AustralianAboriginal - an enigma. In A. M. Gill, R. H.Groves, & I. R. Noble (Eds.), Fire and theAustralian biota (pp. 55–76). Canberra:Australian Academy of Science.
Nix, H.A. (1993). Bird distributions in relation toimperatives for habitat conservation inQueensland. In C. P. Catterall, P. V. Driscoll,K. Hulsman, D. Muir, & A. Taplin (Eds.),Birds and their habitats: status and conservationin Queensland (pp. 12–21). St Lucia:Queensland Ornithological Society Inc.
Noble, J.C., Harrington, G.N. and Hodgkinson,K.C. (1984). The ecological significance ofirregular fire in Australian rangelands. In P. J. Joss, P. W. Lynch and O. B. Williams(Eds.), Rangelands: a resource under siege.(pp.577–580) Canberra: Australian Academyof Science.
Noske, R. (1988). The status and biology of theWhite-throated Grass-wren. AustralianNational Parks & Wildlife Service.
Noske, R. A. (1992a). Do Grasswrens have thenumbers? Reply to Woinarski (1992).Northern Territory Naturalist 13, 5–8.
Noske, R. A. (1992b). The status and ecology ofthe White-throated Grasswren Amytorniswoodwardi. Emu 92, 39–51.
Orr, D.M. and Holmes, W.E. (1984). Mitchellgrasslands. In G.N. Harrington, A.D.Wilson and M.D. Young (Eds.) Managementof Australia’s rangelands. Melbourne: CSIRO.
Paton, J. B., & Paton, D. C. (1977). SeabirdIslands: No. 52. Wright Island, SouthAustralia. Corella 1, 68-69.
Pedler, L. (1991). Rare bird survey: implications forfire management for Uluru National Park.Australian National Parks and WildlifeService.
Pedler, L., & Burbidge, A. H. (1995). The rangeand status of the Nullabor Quail-thrush.South Australian Ornithologist 32, 45–52.
Pescott, T. W. (1976). Seabird Islands: No. 27.Lady Julia Percy Island, Victoria. AustralianBird Bander 14, 29–31.
Pescott, T. (1983). Beach-washed birds after theAsh Wednesday fire. Geelong Naturalist 20,17–19.
Pescott, T. (1985). Black Honeyeaters andcharcoal. Geelong Naturalist 22, 37–39.
Porter, J. W., & Henderson, R. (1983). Birds andburning histories of open forest at Gundiah,southeastern Queensland. Sunbird 13, 61–69.
Press, A. J. (1987). Fire management in KakaduNational Park: the ecological basis for theactive use of fire. Search 18, 244–248.
Press, A. J. (1988). Comparisons of the extent offire in different land management systems inthe Top End of the Northern Territory.Proceedings of the Ecological Society of Australia15, 167–175.
Pressland, A. J., Scanlan, J. C. and McLennan, S.(1992). The role of fire in the grazing landsof Queensland. In B.R. Roberts (Ed.) Fireresearch in rural Queensland. (pp. 237–250).Toowoomba: University of SouthernQueensland.
Price, O. and Bowman, D.M.J.S. (1994). Fire-stick forestry: a matrix model in support ofskilful fire management of Callitrisintratropica R.T. Baker. Journal ofBiogeography 21, 573–580.
Priddel, D. (1989). Conservation of rare fauna:the Regent Parrot and the Malleefowl. In J.C. Noble & R. A. Bradstock (Eds.),Mediterranean landscapes in Australia: malleeecosystems and their management (pp.243–249). Melbourne: CSIRO.
Priddel, D. (1990). Conservation of theMalleefowl in New South Wales: anexperimental management study. In J. C.Noble, P. J. Joss, & G. K. Jones (Eds.), Themallee lands: a conservation perspective (pp.71–74). Melbourne: CSIRO.
95
Biodiveristy and bushfires

Prober, S.M. and Thiele, K.R. (1993). Theecology and genetics of remnant grassyWhite Box woodlands in relation to theirconservation. Victorian Naturalist 110,30–36.
Pyke, G.H., Saillard, R. and Smith, J. (1995).Abundance of Eastern Bristlebirds inrelation to habitat and fire history. Emu 95,106–110.
Pyne, S.J. (1991). Burning bush: a fire history ofAustralia. New York: Henry Holt &Company.
Quin, B. R., & Baker-Gabb, D. (1993).Conservation and management of the TurquoiseParrot Neophema pulchella in north-eastVictoria. Department of Conservation andEnvironment.
Ratkowsky, A. V. (1978). The effect of a springfire on the number of bird species.Tasmanian Naturalist (53), 11–12.
Ratkowsky, A. V. (1979). The bird species of Mt.Nelson in relation to microhabitat andrecent bushfires. Tasmanian Naturalist (57),12–18.
Ratkowsky, A. V. (1985). The effect of a severe fireon the number of bird species in a wetsclerophyll environment. TasmanianNaturalist, 8–9.
Recher, H.F. (1969). Bird species diversity andhabitat diversity in Australia and NorthAmerica. American Naturalist 103, 75–80.
Recher, H. F. (1981). Death of an Australianmyth: fire and its effects on wildlife. In P. J.Stanbury (Ed.), Bushfires: their effect onAustralian life and landscape. (pp. 39–48).Sydney: Macleay Museum, University ofSydney.
Recher, H. F. (1991). The conservation andmanagement of eucalypt forest birds:resource requirements for nesting andforaging. In D. Lunney (Ed.), Conservation ofAustralia’s forest fauna. (pp. 25–34). Sydney:Royal Zoological Society of NSW.
Recher, H. F., & Christensen, P. E. (1981). Fireand the evolution of the Australian biota. InA. Keast (Ed.), Ecological biogeography inAustralia (pp. 135–162). The Hague: Junk.
Recher, H. F., & Lim, L. (1990). A review ofcurrent ideas of the extinction, conservationand management of Australia’s terrestrialvertebrate fauna. Proceedings of the EcologicalSociety of Australia 16, 287–301.
Recher, H. F., & Serventy, D. L. (1991). Longterm changes in the relative abundance ofbirds in Kings Park, Perth, WesternAustralia. Conservation Biology 5, 90–102.
Recher, H. F., Lunney, D., & Posamentier, H.(1975). A grand natural experiment – theNadgee wildfire. Australian Natural History 18,150–163.
Recher, H. F., Allen, D., & Gowing, G. (1985).The impact of wildfire on birds in anintensively logged forest. In A. Keast, H. F.Recher, H. Ford, & D. Saunders (Eds.), Birdsof eucalypt forests and woodlands: ecology,conservation, management (pp. 283–290).Chipping Norton: Surrey Beatty.
Recher, H. F., Davis, W. E., & Holmes, R. T.(1987a). Ecology of Brown and StriatedThornbills in forests of south-eastern NewSouth Wales, with comments on forestmanagement. Emu 87, 1–13.
Recher, H. F., Shields, J., Kavanagh, R., & Webb,G. (1987b). Retaining remnant mature forestfor nature conservation at Eden, New SouthWales: a review of theory and practice. In D.A. Saunders, G. W. Arnold, A. A. Burbidge,& A. J. M. Hopkins (Eds.), Natureconservation: the role of remnants of nativevegetation (pp. 177–194). Chipping Norton:Surrey Beatty.
Recher, H. F., Hutchings, P. A. and Rosen, S.(1993). The biota of the Hawkesbury-Nepean catchment: reconstruction andrestoration. Australian Zoologist 29, 3–41.
Reid, J., & Fleming, M. (1992). The conservationstatus of birds in arid Australia. RangelandsJournal 14, 65–91.
Reid, J. R. W., Kerle, J. A., & Morton, S. R.(1993a). Birds. In J. R. W. Reid, J. A. Kerle,& S. R. Morton (Eds.), Uluru fauna: thedistribution and abundance of vertebrate faunaof Uluru (Ayers Rock – Mount Olga) NationalPark, N.T. (pp. 36–57). Canberra: AustralianNational Parks and Wildlife Service.
Reid, J. R. W., Kerle, J. A., & Morton, S. R.(1993b). Ecological patterns and processes ofimportance. In J. R. W. Reid, J. A. Kerle, &S. R. Morton (Eds.), Uluru fauna: thedistribution and abundance of vertebrate faunaof Uluru (Ayers Rock – Mount Olga) NationalPark, N.T. (pp. 133–148). Canberra:Australian National Parks and WildlifeService.
96
Australia’s Biodiveristy - Responses to Fire

Reilly, P. (1991a). The effect of wildfire on birdpopulations in a Victorian coastal habitat.Emu 91, 100–106.
Reilly, P. (1991b). The effect of wildfire on bushbird populations in six Victorian coastalhabitats. Corella 15, 134–142.
Ridpath, M. G. (1972). The effects of fire onfauna. In R. J. Hooper & M. Rowell (Eds.),Proceedings of the 1971 Tropical and Arid FireSymposium. (pp. 64–66). Darwin: NTGovernment Printer.
Roberts, P. E. (1970). Some effects of a bushfireon heathland birdlife. Proceedings of the RoyalZoological Society of New South Wales 89,40–43.
Robinson, A. C., Casperson, K. D., & Copley, P.B. (1990). Breeding records of theMalleefowl (Leipoa ocellata) andScarlet-chested Parrot (Neophema splendida)within the Yellabinna Wilderness area, SouthAustralia. South Australian Ornithologist 31,8–12.
Robinson, D. (1991). Threatened birds inVictoria: their distributions, ecology andfuture. Victorian Naturalist 108, 67–77.
Robinson, D. (1994). Research plan forthreatened woodland birds of southeasternAustralia. Melbourne: Department ofConservation and Natural Resources.(Technical Report series no. 133).
Robinson, D., & Woinarski, J. C. Z. (1992). Areview of records of the Northern Shrike-titFalcunculus frontatus whitei in northwesternAustralia. South Australian Ornithologist 31,111–117.
Rounsevell, D. E., & Woinarski, J. C. Z. (1983).Status and conservation of the Forty-spottedPardalote, Pardalotus quadragintus (Aves:Pardalotidae). Australian Wildlife Research 10,343–349.
Rowley, I. (1987). Conservation of the Purple-crownedFairy-wren Malurus coronatus in northernAustralia. World Wildlife Fund (Australia).
Rowley, I. (1993). The Purple-crownedFairy-wren Malurus coronatus. I. History,distribution and present status. Emu 93,220–234.
Rowley, I., & Brooker, M. (1987). The response ofa small insectivorous bird to fire inheathlands. In D. A. Saunders, G. W.Arnold, A. A. Burbidge, & A. J. M. Hopkins(Eds.), Nature Conservation: the role ofremnants of native vegetation (pp. 211–218).
Chipping Norton: Surrey Beatty.Rowley, I., & Russell, E. (1993). The
Purple-crowned Fairy-wren Maluruscoronatus. II. Breeding biology, socialorganisation, demography and management.Emu 93, 235–251.
Rowley, I., Russell, E., Brown, R., & Brown, M.(1988). The ecology and breeding biology ofthe Red-winged Fairy-wren Malurus elegans.Emu 88, 161–176.
Russell, E. M., & Rowley, I. (1993). Demographyof the cooperatively breeding SplendidFairy-wren, Malurus splendens (Maluridae).Australian Journal of Zoology 41, 475–505.
Russell-Smith, J. (1985). A record of change:studies of Holocene vegetation history in theSouth Alligator Region, Northern Territory.Proceedings of the Ecological Society of Australia13, 191–202.
Russell-Smith, J. and Bowman, D.M.J.S. (1992).Conservation of monsoon rainforest isolatesin the Northern Territory, Australia.Biological Conservation 59, 51–63.
Russell-Smith, J. and Dunlop, C. (1987). Thestatus of monsoon vine forests in theNorthern Territory. In The rainforest legacy(pp.227–288). Canberra: AustralianGovernment Publishing Service.
Russell-Smith, J. and Ryan, P. (1994). Long-termmonitoring of the effects of management-imposedfire regimes on old-growth vegetation in KakaduNational Park: fire history 1980–1993. Reportto Department of Environment, Sport andTerritories.
Saunders, D. A. (1985). Human impact: theresponse of forest and woodland birdcommunities. Whither the future? Asynthesis. In A. Keast, H. F. Recher, H.Ford, & D. Saunders (Eds.), Birds of eucalyptforests and woodlands: ecology, conservation,management (pp. 355–357). ChippingNorton: Surrey Beatty.
Saunders, D.A. (1989). Changes in the avifauna ofa region, district and remnant as a result offragmentation of native vegtetation: theWheatbelt of Western Australia. A casestudy. Biological Conservation 50, 99–135.
Saunders, D.A. and Curry, P.J. (1990). The impactof agricultural and pastoral industries onbirds in the southern half of WesternAustralia – past, present and future.Proceedings of the Ecological Society of Australia16, 303–321.
97
Biodiveristy and bushfires

Saunders, D. A., Smith, G. T., & Rowley, I.(1982). The availability and dimensions oftree hollows that provide nest sites forcockatoos (Psittaciformes) in WesternAustralia. Australian Wildlife Research 9,541–556.
Scanlan, J.C. (1980). Effects of spring wildfires onAstrebla (mitchell grass) grasslands in north-west Queensland under varying levels ofgrowing season rainfall. AustralianRangelands Journal 2, 162–168.
Schodde, R. (1982). The Fairy-wrens. A monographof the Maluridae. Melbourne: Lansdowne.
Schulz, M. (1991). The Grey-crowned BabblerPomatostomus temporalis - a cause for concernin Victoria. Australian Bird Watcher 14,37–43.
Scotts, D. J. (1991). Old-growth forests: theirecological characteristics and value toforest-dependent vertebrate fauna ofsouth-east Australia. In D. Lunney (Ed.),Conservation of Australia’s forest fauna (pp.147-159). Sydney: Royal Zoological Societyof NSW.
Silveira, C. E. (1993). Recovery plan for threatenedmallee birds - addressing fire regimes. RoyalAustralasian Ornithologists Union.
Silveira, C. E. (1995). The Black–eared Miner.Australian Bird Watcher 16, 96–109.
Singh, G., Kershaw, A.P. and Clark, R. (1981).Quaternary vegetation and fire history inAustralia. In A. M. Gill, R. H. Groves, & I.R. Noble (Eds.), Fire and the Australian biota(pp. 23–54). Canberra: Australian Academyof Science.
Singh, G. and Geissler, E.A. (1985). LateCainozoic history of vegetation, fire, lakelevels and climate at Lake George, NewSouth Wales, Australia. PhilosophicalTransactions of the Royal Society, London. B311,379–447.
Skira, I. J., & Brothers, N. P. (1988a). SeabirdIslands: No. 183. Little Green Island,Furneaux Group, Tasmania. Corella 12,80–81.
Skira, I. J., & Brothers, N. P. (1988b). SeabirdIslands: No. 184. Great Dog Island, FurneauxGroup, Tasmania. Corella 12, 82–84.
Smith, A. P., and Lindenmayer, D. (1988). Treehollow requirements of Leadbeater’s Possumand other possums and gliders in timberproduction ash forests of the Victoriancentral highlands. Australian Wildlife Research15, 347–362.
Smith, E.L. (1960). Effects of burning andclipping at various times during the wetseason on tropical tall grass range innorthern Australia. Journal of RangelandManagement 13, 197–203.
Smith, G. T. (1977). The effect of environmentalchange on six rare birds. Emu 77, 173–179.
Smith, G. T. (1979a). The Noisy Scrub-bird. InM. J. Tyler (Ed.), The status of endangeredAustralasian wildlife (pp. 117–121). Adelaide:Royal Zoological Society of South Australia.
Smith, G. T. (1979b). The status of Australianparrots. In M. J. Tyler (Ed.), The status ofendangered Australasian wildlife (pp.101–108). Adelaide: Royal ZoologicalSociety of South Australia.
Smith, G. T. (1985a). Fire effects on populationsof the Noisy Scrub–bird (Atrichornisclamosus), Western Bristle-bird (Dasyornislongirostris) and Western Whip-bird(Psophodes nigrogularis). In J. R. Ford (Ed.),Fire ecology and management in WesternAustralian ecosystems (pp. 95–102). Perth:Western Australian Institute of Technology.
Smith, G. T. (1985b). The Noisy Scrub-birdAtrichornis clamosus. Does it’s past suggest afuture? In A. Keast, H. F. Recher, H. Ford, &D. Saunders (Eds.), Birds of eucalypt forests andwoodlands: ecology, conservation, management (pp.301–308). Chipping Norton: Surrey Beatty.
Smith, G. T. (1985c). Population and habitatselection of the Noisy Scrub-bird, Atrichornisclamosus, 1962–83. Australian WildlifeResearch 12, 479–485.
Smith, G. T. (1987a). The changing environmentfor birds in the south-west of WesternAustralia; some management implications. InD. A. Saunders, G. W. Arnold, A. A.Burbidge, & A. J. M. Hopkins (Eds.), NatureConservation: The role of remnants of nativevegetation (pp. 269–277). Chipping Norton:Surrey Beatty.
Smith, G. T. (1987b). Observations on the biologyof the Western Bristlebird Dasyornislongirostris. Emu 87, 111–118.
Smith, G. T. (1991). Ecology of the WesternWhipbird Psophodes nigrogularis in WesternAustralia. Emu 91, 145–157.
Smith, G. T., & Forrester, R. I. (1981). The statusof the Noisy Scrub-bird Atrichornis clamosus.Biological Conservation 19, 239–254.
Smith, G. T., & Robinson, F. N. (1976). TheNoisy Scrub-bird: an interim report. Emu76, 37–42.
98
Australia’s Biodiveristy - Responses to Fire

Smith, L. H. (1994). A critical analysis of thefactors responsible for the decline of theSuperb Lyrebird Menura novaehollandiae inSherbrooke Forest, Victoria. Australian BirdWatcher 15, 238–249.
Smith, P. (1989). Changes in a forest birdcommunity during a period of fire anddrought near Bega, New South Wales.Australian Journal of Ecology 14, 41–54.
Smith, P. J., Pressey, R. L., & Smith, J. E. (1994).Birds of particular conservation concern inthe Western Division of New South Wales.Biological Conservation 69, 315–338.
Smith, P. J., Smith, J. E., & Pressey, R. L. (1996).Birds of particular conservation concern in theWestern Division of New South Wales:distribution, habitats and threats. NSWNational Parks and Wildlife Service.
Specht, R. L. (1981). Responses to fires inheathlands and related shrublands. In A. M.Gill, R. H. Groves, & I. R. Noble (Eds.),Fire and the Australian biota (pp. 395–415).Canberra: Australian Academy of Science.
Springett, J.A. (1976). The effect of prescribedburning on the soil fauna and on litterdecomposition in Western Australian forests.Australian Journal of Ecology 1, 72–82.
Stanton, J. P. (1992). J.P. Thomson oration. Theneglected lands: recent changes in theecosystems of Cape York Peninsula and thechallenge of their management. Journal of theQueensland Geographical Society 7, 1–18.
Stanton, P. (1995). A tropical Queenslandperspective. In D. B. Rose (Ed.), Country inflames. Proceedings of the 1994 symposium onbiodiversity and fire in North Australia (pp.71–76). Canberra: Department of theEnvironment, Sport and Territories, andNorth Australia Research Unit.
Starks, J. (1987). The status and distribution of theBlack-eared Miner (Manorina melanotis) inVictoria. Technical report no.49. Arthur RylahInsitute of Environmental Research.
Stephens, S. (1992). Endangered species andcommunities and threatening processes in theMurray Mallee. Australian National Parksand Wildlife Service.
Stocker, G. C. (1971). The age of charcoal fromold jungle fowl nests and vegetation changeon Melville Island. Search 2, 28–30.
Stokes, T. (1975). The effect of a bushfire on thebanding of Flame Robins in the BrindabellaRanges. Australian Bird Bander 13, 75–76.
Suckling, G. C., & MacFarlane, M. A. (1983).The effects of fire on fauna – a review. In E.Ealey (Ed.), Fighting fire with fire. (pp.107–128). Melbourne: Monash University.
Suijdendorp, H. (1981). Responses of thehummock grasslands of northwesternAustralia to fire. In A. M. Gill, R. H. Groves,& I. R. Noble (Eds.), Fire and the Australianbiota (pp. 417–424). Canberra: AustralianAcademy of Science.
Sullivan, M., Cowan, D. and Pressland, A. (1992).The effects of controlled burns in the mulgalands of southwest Queensland. In B.R.Roberts (Ed.) Fire research in ruralQueensland. (pp. 300–307). Toowoomba:University of Southern Queensland.
Tarr, H. E. (1965). The Mallee-Fowl inWyperfeld National Park. Australian BirdWatcher 2, 140–144.
Taylor, R. J. (1991). Fauna conservation inproduction forests in Tasmania. Hobart:Forestry Commission Tasmania.
Thackway, R. and Creswell, I.D. (1995). Aninterim biogeographic regionalisation forAustralia. Canberra: Australian NatureConservation Agency.
Tidemann, S. C. (1990). Relationships betweenfinches and pastoral practices in northernAustralia. In J. Pinowski & J. D.Summers–Smith (Eds.), Granivorous birds andagriculture (pp. 305–315). Warsaw:PWN-Polish Scientific Publishers.
Tidemann, S. C. (1992). Conservation of theGouldian Finch, N.T. World Wide Fund forNature.
Tidemann, S. C. (1993a). Management of athreatened species: the Gouldian Finchexample. In C. P. Catterall, P. V. Driscoll, K.Hulsman, D. Muir, & A. Taplin (Eds.), Birdsand their habitats: status and conservation inQueensland (pp. 123–131). St Lucia:Queensland Ornithological Society Inc.
Tidemann, S. C. (1993b). Where are GouldianFinches after the breeding season? VictorianNaturalist 110, 238–243.
Tidemann, S.C., Boyden, J., Elvish, R., Elvish, J.and O’Gorman, B. (1992). Characteristics ofnest sites and breeding habitat of theendangered Gouldian Finch Erythruragouldiae and abundant, co-occurring Long-tailed Finch Poephila acuticauda at two sites inthe Northern Territory. Journal of TropicalEcology 8, 373–388.
99
Biodiveristy and bushfires

Tidemann, S. C., McArtney, J., & Smith, I.(1993). Queensland Gouldian FinchesErythrura gouldiae and air-sac miteSternostoma tracheacolum. Sunbird 23, 36–40.
Tingay, A., & Tingay, S. R. (1984). Bird communitiesin the karri forest of Western Australia.Australian Conservation Foundation.
Tolhurst, K. (1996). Effects of fuel reductionburning on fauna in a dry sclerophyll forest.In DEST (Ed.), Fire and biodiversity: theeffects and effectiveness of fire management.Proceedings of the conference held 8–9 October1994, Footscray, Melbourne (pp. 113–121(&129)). Canberra: Department of theEnvironment, Sports and Territories.
Tolhurst, K. and Flinn, D. (eds.) (1992). Ecologicalimpact of fuel reduction burning in drysclerophyll forest: first progress report.Melbourne: Department of Conservationand Environment (Research Report no. 349).
Tothill, J.C. (1992). Fire in black speargrass(Heteropogon contortus) pastures – a ten yearcomparison of burnt and unburnttreatments. In B.R. Roberts (Ed.) Fireresearch in rural Queensland. (pp. 274–279).Toowoomba: University of SouthernQueensland.
Traill, B.J. (1993). Forestry, birds, mammals andmanagement in box and ironbark forests.Victorian Naturalist 110, 11–14.
Turner, R. J. (1987). Effect of fire on birds –Weddin Mountain. In Disappearing islands.Proceedings of a seminar on conservation andco-operation in the Central West (pp. 66–86).Bathurst: National Parks and WildlifeService, NSW.
Turner, R. J. (1992). Effect of wildfire on birds atWeddin Mountain, New South Wales.Corella 16, 65–74.
Vickers-Rich, P. (1991). The Mesozoic andTertiary history of birds on the Australianplate. In (Eds.) P. Vickers-Rich, J.M.Monaghan, R.F. Baird and T.H. RichVertebrate plaeontology of Australasia. (pp.721–808). Lilydale: Pioneers Design Studios.
Wakefield, N.A. (1970). Bushfire frequency andvegetational change in south-easternAustralian forests. Victorian Naturalist 87,152–158.
Walker, J. and Tothill, J.C. (1992). Fires in thegrasslands of Queensland. In B.R. Roberts(Ed.) Fire research in rural Queensland. (pp.251–262). Toowoomba: University ofSouthern Queensland.
Walker, T. A., & Hulsman, K. (1993). SeabirdIslands: No. 221. Wilson Island, Great BarrierReef, Queensland. Corella 17, 155–157.
Wardell-Johnson, G., & Christensen, P. (1992). Areview of the effects of disturbance onwildlife of the karri forest. In Research on theimpact of forest management in south-westWestern Australia (pp. 33–57). Perth:Department of Conservation and LandManagement (Occasional Paper 2/92).
Wardell-Johnson, G., & Nichols, O. (1991).Forest wildlife and habitat management insouthwestern Australia: knowledge, researchand direction. In D. Lunney (Ed.),Conservation of Australia’s forest fauna (pp.161–192). Mosman: Royal ZoologicalSociety of NSW.
Wardell-Johnson, G., McCaw, W. L., & Masey,K. G. (1989). Critical data requirements forthe effective management of fire on natureconservation lands in south WesternAustralia. In N. Burrows, L. McCaw, & G.Friend (Eds.), Fire management on natureconservation lands Perth: Department ofConservation and Land Management.
Watkins, D. (1985). Report of the R.A.O.U. GroundParrot survey in Western Australia. RoyalAustralasian Ornithologists Union.
Webster, R., & Ahern, L. (1992). Management forconservation of the Superb Parrot (Polytelisswainsonii) in New South Wales and Victoria.Department of Conservation and NaturalResources.
Wegener, A. (1984a). Animals killed by bushfiresin coastal Victoria, February 1983. AustralianBird Watcher 10, 248–250.
Wegener, A. (1984b). Survey of animal species killedby the Ash Wednesday fires along the GreatOcean Road. Geelong Naturalist 21, 13–16.
Westoby, M., Rice, B., Griffin, G. and Friedel, M.(1988). The soil seed banks of Triodiabasedowii in relation to time since fire.Australian Journal of Ecology 13, 161–169.
Whelan, R. J. (1995). The ecology of fire.Cambridge: Cambridge University Press.
White, G. (1979a). Seabird islands: No. 75.Louisa Island, Tasmania. Corella 3, 61–62.
White, G. (1979b). Seabird Islands: No. 76. DeWitt Island, Tasmania. Corella 3, 63–65.
Whitley, G. P. (1944). Fire and Petrels: themystery of Mondrain Island. Emu 44, 6–7.
Whitley, G. P. (1971). Field notes on birds byThomas Carter. Western Australian Naturalist12, 41–44.
100
Australia’s Biodiveristy - Responses to Fire

101
Fire and Australian Birds
Whittell, H. M. (1936). The Bristle-birds ofWestern Australia. Emu 35, 197–201.
Williams, J. E., & Gill, A. M. (1995). The impact offire regimes on native forests in eastern NewSouth Wales. NSW National Parks andWildlife Service.
Williams, J.E., Whelan, R.J. and Gill, A.M.(1994). Fire and environmentalheterogeneity in southern temperate forestecosystems: implications for management.Australian Journal of Botany 42, 125–137.
Wilson, R. I. (1981). The woodchip industry andTasmanian birds. Tasmanian Bird Report 11,11–14.
Woinarski, J. C. Z. (1987). Notes on the statusand ecology of the Red–lored WhistlerPachycephala rufogularis. Emu 87, 224–231.
Woinarski, J. C. Z. (1989a). Broombush harvestingin southeastern Australia. In J. C. Noble & R.A. Bradstock (Eds.), Mediterranean landscapes inAustralia: mallee ecosystems and their management(pp. 362–378). Melbourne: CSIRO.
Woinarski, J. C. Z. (1989b). The vertebrate faunaof broombush Melaleuca uncinata vegetationin north-western Victoria, with reference toeffects of broombush harvesting. AustralianWildlife Research 16, 217–238.
Woinarski, J. C. Z. (1990). Effects of fire on thebird communities of tropical woodlands andopen forests in northern Australia.Australian Journal of Ecology 15, 1–22.
Woinarski, J. C. Z. (1992). The conservationstatus of the White-throated GrasswrenAmytornis woodwardi, an example ofproblems in status designation. NorthernTerritory Naturalist 13, 1–5.
Woinarski, J. C. Z. (1993a). Australian tropicalsavannas, their avifauna, conservation statusand threats. In C. P. Catterall, P. V. Driscoll,K. Hulsman, D. Muir, & A. Taplin (Eds.),Birds and their habitats: status and conservationin Queensland (pp. 45–63). St Lucia:Queensland Ornithological Society Inc.
Woinarski, J. C. Z. (1993b). A cut-and-pastecommunity: birds of monsoon rainforests inKakadu National Park, Northern Territory.Emu 93, 100–120.
Woinarski, J. C. Z., & Fisher, A. (1995a). Wildlifeof lancewood (Acacia shirleyi) thickets andwoodlands in northern Australia: 1. variationin vertebrate species composition across theenvironmental range occupied by lancewoodvegetation in the Northern Territory.Wildlife Research 22, 379–411.
Woinarski, J. C. Z., & Fisher, A. (1995b). Wildlifeof lancewood (Acacia shirleyi) thickets andwoodlands in northern Australia: 2.comparisons with other environments of theregion (Acacia woodlands, Eucalyptus savannawoodlands and monsoon rainforests).Wildlife Research 22, 413–443.
Woinarski, J. C. Z., & Tidemann, S. C. (1991).The bird fauna of a deciduous woodland inthe wet–dry tropics of northern Australia.Wildlife Research 18, 479–500.
Woinarski, J. C. Z., & Tidemann, S. C. (1992).Survivorship and some populationparameters for the endangered GouldianFinch Erythrura gouldiae and two other finchspecies at two sites in tropical northernAustralia. Emu 92, 33–38.
Woinarski, J. C. Z., Eckert, H. J., & Menkhorst,P. W. (1988a). A review of the distribution,habitat and conservation status of theWestern Whipbird Psophodes nigrogularisleucogaster in the Murray mallee. SouthAustralian Ornithologist 30, 146–153.
Woinarski, J. C. Z., Tidemann, S. C., & Kerin, S.(1988b). Birds in a tropical mosaic: thedistribution of bird species in relation tovegetation patterns. Australian WildlifeResearch 15, 171–196.
Woinarski, J. C. Z., Whitehead, P. J., Bowman, D.M. J .S. & Russell-Smith, J. (1992).Conservation of mobile species in a variableenvironment: the problem of reserve designin the Northern Territory, Australia. GlobalEcology and Biogeography Letters 2, 1–10.
Wooller, R. D., & Brooker, K. S. (1980). Theeffects of controlled burning on some birdsof the understorey in Karri forest. Emu 80,165–166.
Wooller, R. D., & Calver, M. C. (1988). Changesin an assemblage of small birds in theunderstorey of dry sclerophyll forest insouth-western Australia after fire. AustralianWildlife Research 15, 331–338.
Yates, C. J., Hobbs, R.J. and Bell, R.W. (1994).Landscape-scale disturbance andregeneration in semi-arid woodlands ofsouthwestern Australia. Pacific ConservationBiology 1, 214–221.
Young, J. (1991). Dealing with fire in parks andprotected areas. In Tropics under fire: firemanagement on Cape York Peninsula (pp.43–45). Cairns: Cairns and Far NorthEnvironment Centre.

103
Fire and Australian Birds
Table 1. Scientific names of birds mentioned intext (following Christidis & Boles 1994).
ORDER STRUTHIONIFORMESCasuariidaeCasuarius casuarius Southern CassowaryDromaius novaehollandiaeEmuDromaius baudinianus Kangaroo Island Emu
ORDER GALLIFORMESMegapodiidaeLeipoa ocellata MalleefowlMegapodius reinwardt Orange-footed ScrubfowlPhasianidaeCoturnix ypsilophora Brown QuailCoturnix chinensis King Quail
ORDER ANSERIFORMESAnaseranatidaeAnseranas semipalmata Magpie Goose
ORDER PELECANIFORMESPelecanidaePelecanus conspicillatus Australian Pelican
ORDER CICONIIFORMESThreskionithidaeThreskiornis spinicollis Straw-necked Ibis
ORDER FALCONIFORMESAccipitridaeErythrotriorchis radiatus Red GoshawkAquila audax Wedge-tailed EagleFalconidaeFalco cenchroides Nankeen Kestrel
ORDER GRUIFORMESRallidaeRallus pectoralis Lewin’s RailGallinula mortierii Tasmanian Native-henOtididaeArdeotis australis Australian Bustard
ORDER TURNICIFORMESTurnicidaeTurnix velox Little Button-quailTurnix varia Painted Button-quail Turnix melanogaster Black-breasted Button-
quail
Table 1. continued
ORDER COLUMBIFORMESColumbidaePhaps chalcoptera Common BronzewingPhaps elegans Brush BronzewingOcyphaps lophotes Crested PigeonGeophaps smithii Partridge PigeonGeopelia cuneata Diamond Dove
ORDER PSITTACIFORMESCacatuidaeProbosciger aterrimus Palm CockatooCalyptorhynchus banskii Red-tailed
Black-CockatooCalyptorhynchus lathami Glossy Black-CockatooCalyptorhynchus funereus Yellow-tailed
Black-CockatooCalyptorhynchus baudinii Long-billed Black-Cockatoo
Cacatua roseicapilla GalahCacatua tenuirostris Long-billed CorellaCacatua pastinator Western CorellaCacatua sanguinea Little CorellaCacatua leadbeateri Major Mitchell’s
CockatooNymphicus hollandicus CockatielPsittacidaePolytelis swainsonii Superb ParrotPolytelis anthopeplus Regent ParrotPlatycercus icterotis Western RosellaBarnardius zonarius Australian RingneckPurpureicephalus spurius Red-capped ParrotPsephotus chrysopterygius Golden-shouldered
ParrotPsephotus dissimilis Hooded ParrotPsephotus pulcherrimus Paradise Parrot Melopsittacus undulatus BudgerigarNeophema chrysostoma Blue-winged ParrotNeophema elegans Elegant ParrotNeophema chrysogaster Orange-bellied ParrotNeophema pulchella Turquoise Parrot
7. TABLES
CacatuidaeProbosciger aterrimus Palm CockatooCalyptorhynchus banskii Red-tailed
Black-CockatooCalyptorhynchus lathami Glossy Black-CockatooCalyptorhynchus funereus Yellow-tailed
Black-CockatooCalyptorhynchus baudinii Long-billed Black-
CockatooCacatua roseicapilla GalahCacatua tenuirostris Long-billed CorellaCacatua pastinator Western CorellaCacatua sanguinea Little CorellaCacatua leadbeateri Major Mitchell’sCockatooNymphicus hollandicus CockatielPsittacidaePolytelis anthopeplus Regent ParrotPlatycercus icterotis Western RosellaBarnardius zonarius Australian RingneckPurpureicephalus spurius Red-capped ParrotPsephotus chrysopterygius Golden-shouldered
ParrotPsephotus dissimilis Hooded ParrotPsephotus pulcherrimus Paradise Parrot Melopsittacus undulatus BudgerigarNeophema chrysostoma Blue-winged ParrotNeophema elegans Elegant ParrotNeophema chrysogaster Orange-bellied ParrotNeophema pulchella Turquoise ParrotNeophema splendida Scarlet-chested ParrotPezoporus wallicus Ground ParrotPezoporus occidentalis Night Parrot
ORDER CHARADRIIFORMESPedionomidaePedionomus torquatus Plains-wandererBurhinidaeBurhinus grallarius Bush Stone-curlew
continued over
continued over

Australia’s Biodiveristy - Responses to Fire
Table 1. continued
ORDER STRIGIFORMESStrigidaeNinox strenua Powerful OwlTytonidaeTyto tenebricosa Sooty OwlTyto novaehollandiae Masked Owl
ORDER APODIFORMESApodidaeHirundapus caudacutus White-throated NeedletailApus pacificus Fork-tailed Swift
ORDER CORACIIFORMESHalcyonidaeDacelo novaeguineae Laughing KookaburraDacelo leachii Blue-winged KookaburraTodiramphus pyrrhopygia Red-backed KingfisherMeropidaeMerops ornatus Rainbow Bee-eater
ORDER PASSERIFORMESMenuridaeMenura novaehollandiae Superb LyrebirdAtrichornithidaeAtrichornis rufescens Rufous Scrub-birdAtrichornis clamosus Noisy Scrub-birdClimacteridaeClimacteris picumnus Brown Treecreeper[C.p. melanotaMaluridaeMalurus coronatus Purple-crowned
Fairy-wrenMalurus cyaneus Superb Fairy-wrenMalurus splendens Splendid Fairy-wrenMalurus lamberti Variegated Fairy-wrenMalurus elegans Red-winged Fairy-wrenMalurus leucopterus White-winged Fairy-wrenMalurus melanocephalus Red-backed Fairy-wrenStipiturus malachurus Southern Emu-wrenStipiturus mallee Mallee Emu-wrenStipiturus ruficeps Rufous-crowned
Emu-wrenAmytornis woodwardi White-throated GrasswrenAmytornis dorotheae Carpentarian GrasswrenAmytornis striatus Striated GrasswrenAmytornis textilis Thick-billed GrasswrenPardalotidaePardalotus punctatus Spotted PardalotePardalotus quadragintus Forty-spotted PardalotePardalotus striatus Striated PardaloteDasyornis brachypterus Eastern BristlebirdDasyornis longirostris Western Bristlebird
Table 1. continued
Dasyornis broadbenti Rufous BristlebirdPycnoptilus floccosus PilotbirdSericornis frontalis White-browed
ScrubwrenHylacola pyrrhopygia Chestnut-rumped
HeathwrenHylacola cauta Shy HeathwrenCalamanthus fuliginosus Striated FieldwrenChthonicola sagittata Speckled WarblerGerygone fusca Western GerygoneAcanthiza pusilla Brown ThornbillAcanthiza apicalis Inland ThornbillAcanthiza inornata Western ThornbillAcanthiza reguloides Buff-rumped ThornbillAcanthiza iredalei Slender-billed ThornbillAcanthiza chrysorrhoa Yellow-rumped ThornbillAcanthiza nana Yellow ThornbillAcanthiza lineata Striated ThornbillAphelocephala nigricincta Banded WhitefaceMeliphagidaeAnthochaera chrysoptera Little WattlebirdPhilemon corniculatus Noisy FriarbirdPhilemon citreogularis Little FriarbirdManorina flavigula Yellow-throated MinerManorina melanotis Black-eared MinerLichenostomus chrysops Yellow-faced HoneyeaterLichenostomus melanops Yellow-tufted
HoneyeaterLichenostomus melanops cassidix Helmeted HoneyeaterLichenostomus cratitius Purple-gaped
HoneyeaterLichenostomus plumulus Grey-fronted HoneyeaterLichenostomus penicillatus White-plumed
HoneyeaterMelithreptus lunatus White-naped HoneyeaterPhylidonyris pyrrhoptera Crescent HoneyeaterPhylidonyris nigra White-cheeked
HoneyeaterPhylidonyris albifrons White-fronted
HoneyeaterPhylidonyris melanops Tawny-crowned
HoneyeaterCerthionyx niger Black HoneyeaterEpthianura tricolor Crimson ChatPetroicidaePetroica multicolor Scarlet RobinPetroica goodenovii Red-capped RobinPetroica phoenicea Flame RobinMelanodryas cucullata Hooded RobinEopsaltria australis Eastern Yellow RobinEopsaltria griseogularis Western Yellow Robin
continued over continued over
104

Table 1. continued
Lonchura castaneothorax Chestnut-breastedMannikin
Erythrura gouldiae Gouldian FinchDicaeidaeDicaeum hirundinaceum MistletoebirdHirundinidaeHirundo nigricans Tree Martin SylviidaeMegalurus timoriensis Tawny GrassbirdEremiornis carteri SpinifexbirdCincloramphus mathewsi Rufous SonglarkCisticola exilis Golden-headed Cisticola
105
Fire and Australian Birds
Table 1. continued
Eopsaltria georgiana White-breasted RobinDrymodes superciliaris Northern Scrub-robinDrymodes brunneopygia Southern Scrub-robinPomatostomidaePomatostomus superciliosus White-browed BabblerCinclosomatidaePsophodes olivaceus Eastern WhipbirdPsophodes nigrogularis Western WhipbirdCinclosoma punctatum Spotted Quail-thrushCinclosoma castanotus Chestnut Quail-thrushCinclosoma cinnamomeumCinnamon Quail-thrush
(includes Nullabor Quail-thrush)
NeosittidaeDaphoenositta chrysopteraVaried SittellaPachycephalidaeFalcunculus frontatus Crested Shrike-titOreoica gutturalis Crested BellbirdPachycephala olivacea Olive WhistlerPachycephala rufogularis Red-lored WhistlerPachycephala pectoralis Golden WhistlerPachycephala rufiventris Rufous WhistlerColluricincla harmonica Grey Shrike-thrushDicruridaeGrallina cyanoleuca Magpie-larkRhipidura fuliginosa Grey FantailCampephagidaeCoracina novaehollandiae Black-faced
Cuckoo-shrikeLalage sueurii White-winged TrillerAatamidaeArtamus personatus Masked WoodswallowArtamus superciliosus White-browed
WoodswallowArtamus cyanopterus Dusky WoodswallowCracticus torquatus Grey ButcherbirdCracticus nigrogularis Pied ButcherbirdGymnorhina tibicen Australian MagpieStrepera graculina Pied CurrawongCorvidaeCorvus coronoides Australian RavenCorvus orru Torresian CrowCorcoracidaeCorcorax melanorhamphosWhite-winged ChoughMotacillidaeAnthus novaeseelandiae Richard’s PipitPasseridaeTaeniopygia guttata Zebra FinchPoephila personata Masked FinchNeochmia ruficauda Star FinchNeochmia temporalis Red-browed Finch
continued over
Neosittidae

Australia’s Biodiveristy - Responses to Fire
Table 2. continued
5 Avon Wheatbelt, ChannelCountry, Daly Basin,Furneaux, Gulf Coastal,Naracoorte Coastal Plain
4 Great Sandy Desert, MulgaLands, NSW South westernSlopes, Riverina
3 Broken Hill Complex, CentralKimberley, Mt Isa Inlier,Northern Kimberley,Nullabor, Sydney Basin
2 Carnarvon, D’Entrecasteaux,Lofty Block, Mitchell GrassDowns, Simpson-StrezleckiDunefields, Sturt Plateau, WetTropics, Woolnorth
1 Australian Alps, Ben Lomond,Brigalow Belt North, BrigalowBelt South, Central Arnhem,Central Mackay Coast,Darling Riverine Plains,Desert Uplands, EinasleighUplands, Eyre & YorkeBlocks, Finke, Flinders andOlary Ranges, Gascoyne,Gawler, Geraldton Sandplains,Gibson Desert, Hampton,Little Sandy Desert,MacDonnell Ranges,Murchison, Nandewar,Pilbara, Stony Plains, Yalgoo
ENVIRONMENTSNo. of references habitat116 (temperate) eucalypt openforest103 heath48 tussock grassland47 mallee25 tropical eucalypt open forest24 tropical eucalypt savanna
woodland21 hummock grassland19 rainforest17 (temperate) eucalypt woodland15 thicket10 sedgeland8 Acacia woodland/shrubland6 wetlands5 Callitris woodland, pandanus,
(Allo)Casuarina woodland,chenopod shrubland
2 Banksia woodland1 riparian vegetation
continued over
Table 2. Summary table of incidence ofsubjects in fire-bird references.
This tally is based on references included within theannotated bibliography. Where possible, I have triedto restrict inclusion in this list to subjects which forma substantial component or focus of the paperconsidered. Note that the same material may beincluded in several papers (notably in reviews), suchthat a large tally doesn’t necessarily equate to asubstantial research effort. A single paper mayconsider several subjects, so the tallies do not sum tothe number of papers included in the annotatedbibliography (361). Bioregions are the biogeographicregions defined in Thackway & Cresswell (1995).
BROAD SUBJECTSNo. of references subject141 threatened species127 management78 wildfire60 review46 community41 control burning26 historic change; seabirds24 hollows21 forestry20 mortality17 succession13 isolate10 Aboriginal burning9 old-growth6 charcoal; Aboriginal knowledge2 corridors; slash-burn
BIOREGIONSNo. of references Bioregions37 Murray-Darling Depression33 South East Corner29 South East Coastal Plain28 Jarrah Forest22 South Eastern Highlands19 Esperance Plains,
Top End Coastal, Warren14 Pine Creek-Arnhem
Swan Coastal Plain12 South Eastern Queensland11 Swan Coastal Plain10 Cape York Peninsula9 West and South West8 Ord-Victoria Plains7 Gulf Falls and Uplands
Victoria Bonaparte6 Freycinet, NSW North Coast,
Victorian Midlands
continued over

107
Fire and Australian Birds
Table 2. continued
INDIVIDUAL BIRD SPECIESNo. of references bird species33 Ground Parrot15 Western Whipbird14 Malleefowl13 Noisy Scrub-bird8 Eastern Bristlebird, Western
Bristlebird7 Gouldian Finch6 Rufous Bristlebird,
Black-eared Miner5 Glossy Black-cockatoo,
Golden-shouldered Parrot,Carpentarian Grass-wrenWhite-throated Grass-wren
4 Orange-footed Scrubfowl,Sooty Owl, Superb Lyrebird,Splendid Fairy-wren
3 Cassowary, Hooded Parrot,Nullabor Quail-thrush,Striated Grass-wren, Purple-crowned Fairy-wren, BlackHoneyeater
2 Red Goshawk, Powerful Owl,Red-tailed Black-Cockatoo,Superb Parrot, Regent Parrot,Turquoise Parrot, Scarlet-chested Parrot, Orange-belliedParrot, Paradise Parrot, WesternRosella, Ringneck Parrot, NightParrot, White-throatedNeedletail, Rufous Scrub-bird,Grey-crowned Babbler, FlameRobin, Red-lored Whistler,Thick-billed Grass-wren,Rufous-crowned Emu-wren,Southern Emu-wren, MalleeEmu-wren, Brown Thornbill,Slender-billed Thornbill, Forty-spotted Pardalote
continued over
Table 2. continued
1 Kangaroo Island EmuAustralian Pelican, MagpieGoose, Australian Bustard,Black-breasted Button-quail,Chestnut-backed Button-quail,Little Button-quail, BushStone-curlew, Wedge-tailedEagle, Plains-wanderer, PalmCockatoo, Pink Cockatoo,Yellow-tailed Black-cockatoo,White-tailed Black-cockatoo,Long-billed Corella LittleCorella Red-capped Parrot,Brush Bronzewing, FlockBronzewing, Partridge Pigeon, Fork-tailed Swift, Tree Martin,Spinifex Bird, Richard’s Pipit,Pilot Bird, Spotted Quail-thrush, Northern Scrub-robin,Eastern Yellow Robin, ScarletRobin, Crested Shrike-tit,Black Grass-wren, White-winged Fairy-wren, Red-winged Fairy-wren, WesternThornbill, Striated Thornbill,Brown Thornbill, Yellow-rumped Thornbill, BlackTreecreeper, HelmetedHoneyeater, Mistletoebird,Yellow-rumped Pardalote,Forty-spotted Pardalote, StarFinch, Yellow-rumpedMannikin

108
Australia’s Biodiveristy - Responses to Fire
Table 3. continued
Malurus coronatus Purple-crowned Fairy-coronatus wren (western subsp.) (V)Stipiturus malachurus Southern Emu-wren (Mt intermedius Lofty Ranges subsp.) (E)Stipiturus malachurus Southern Emu-wren parimeda (Eyre Peninsula subsp.)
(V)Stipiturus mallee Mallee Emu-wren (V)Amytornis dorotheae Carpentarian Grasswren
(K)Amytornis striatus Striated Grasswren striatus (sandplain subsp.) (K)Amytornis striatus Striated Grasswren merrotsyi (Flinders Ranges subsp.)
(K)Pardalotus quadragintus Forty-spotted Pardalote
(E)Dasyornis brachypterus Eastern Bristlebird (V)Dasyornis longirostris Western Bristlebird (E)Dasyornis broadbenti Rufous Bristlebird broadbenti (Victorain subsp.) (R)Dasyornis broadbenti Rufous Bristlebird whitei South Australian subsp.)
(K)Acanthiza pusilla Brown Thornbill (King arcibaldi Island subsp.) (K)Acanthiza iredalei Slender-billed Thornbill hedleyi eastern subsp.) (K)Manorina melanotis Black-eared Miner (E)Lichenostomus melanops cassidix Helmeted Honeyeater (E)Psophodes nigrogularis Western Whipbirdnigrogularis (western heath subsp.) (E)Psophodes nigrogularis Western Whipbird oberon (western mallee subsp.)
(R)Psophodes nigrogularis Western Whipbird lashmari (Kangaroo Island subsp.)
(R)Psophodes nigrogularis Western Whipbird leucogaster (eastern subsp.) (V)Falcunculus frontatus Crested Shrike-tit whitei (northern subsp.) (K)Pachycephala rufogularis Red-lored Whistler (V)Erythrura gouldiae Gouldian Finch (E)Cisticola juncidis Zitting Cisticola normani (Normanton subsp.) (K)Zoothera lunulata Bassian Thrush (Southhalmaturina Australian subsp.) (R)
Table 3. Threatened bird taxa for whichinappropriate fire regime has been listed as athreatening process by Garnett (1992b).Extinct taxa are excluded. Conservation status:R=rare, K=insufficiently known, V=vulnerable,E=endangered.
Casuarius casuarius Southern Cassowary (V)Leipoa ocellata Malleefowl (V)Botaurus poiciloptilus Australasian Bittern (K)Erythrotriorchis radiatus Red Goshawk (V)Turnix castanota Chestnut-backed Button-
quail (K)Turnix olivii Buff-breasted Button-
quail (K)Turnix varia scintillans Painted Button-quail
(Houtmans Abrolhossubsp) (V)
Turnix melanogaster Black-breasted Button-quail (V)
Geophaps smithii smithii Partridge Pigeon (easternsubsp.) (V)
Geophaps smithii blaauwi Partridge Pigeon (westernsubsp.) (K)
Probosciger aterrimus Palm Cockatoo (K)Calyptorhynchus Red-tailed Black-
Cockatoobanskii graptogyne (southeastern subsp.) (E)Calyptorhynchus Glossy Black-Cockatoolathami halmaturinus (Kangaroo Island subsp.)
(E)Eclectus roratus Eclectus Parrot (R)Psephotus chrysopterygius Golden-shouldered Parrot
(E)Neophema chrysogaster Orange-bellied Parrot (E)Neophema splendida Scarlet-chested Parrot (R)Pezoporus wallicus Ground Parrot (western flaviventris subsp.) (E)Pezoporus occidentalis Night Parrot (K)Ninox rufa queenslandica Rufous Owl (eastern
subsp.) (R)Ninox rufa meesi Rufous Owl (Cape York
Peninsula subsp.) (K)Tyto tenebricosa Sooty Owl (R)Tyto novaehollandiae Masked Owl melvillensis (Melville Island subsp.)
(K)Atrichornis rufescens Rufous Scrub-bird (R)Atrichornis clamosus Noisy Scrub-bird (E)
continued over

109
Fire and Australian Birds
Mallee Fowl. Fires in the mallee country may create flushes of food resources for the Mallee Fowl, but generallylong-unburnt country is most favourable, especially for breeding. A complex fire mosaic is probably optimal forthis species, with special management effort directed to maintaining the most vulnerable long-unburnt patches./K Thaler © ANBG. Inset photo /Joe Benshemesh
Noisy Scrub-bird. The range and abundance of thisunusual bird has declined catastrophically over thelast 150 years. It is now restricted to a very fewpopulations in south-western Australia. Much of thisdecline has been due to the imposition of aninappropriate fire regime. Despite living in highlyflammable heath and thicket vegetation, the NoisyScrub-bird is dependent upon vegetation which hasremained unburnt for long periods, typically greaterthan 40 years. /Ray Smith
Splendid Fairy-Wren. The Splendid Fairy-wren is asmall bird found in heathlands in southwesternAustralia. If heathland patches are totally burnt byfire, local extinctions may occur, and the Fairy-wren’spoor dispersal ability renders recruitment andrecolonisation difficult. Cooler fires may also affecthabitat suitability through impacts upon preyabundance, nest site availability, nest parasitism andpredation. /Tom & Pam Gardner © Nature Focus

110
Australia’s Biodiveristy - Responses to Fire
Magpie Goose. Aboriginal people have a long tradition of burning the northern floodplains once the watershave receded during the dry season. The purpose of burning is to increase the suitability of the area for MagpieGoose feeding and nesting. Where Aboriginal fire management of the floodplains has been removed, MagpieGeese have become disadvantaged. /G O’Neill © ANPWS
Gouldian Finch. The endangered Gouldian Finchhas complex relationships with fire. In its tropicalsavanna woodland habitat, dry season fires removethe dense grass layer, allowing it easier access tograss seeds on the ground. Hence, it seeks outrecently burnt areas and in the short term is favouredby frequent (annual) fires. However, fires maychange the seed productivity and timing in thesubsequent seeding period, and little is known of thelonger-term consequences of a frequent fire regimeupon grass species composition and hence foodresources. Fire management for this species shouldaim to maximise environmental variability. /Ian Morris
Red-backed Fairy-wren. The Red-backed Fairy-wrenoccurs in dense ground layer vegetation acrossnorthern Australia. Fires remove most of thisvegetation, and may knock out local populations. A consistent regime of frequent fires over extensiveareas will greatly suppress populations of thisspecies. /Tom & Pam Gardner © Nature Focus

111
Fire and Australian Birds
Ground Parrot. The Ground Parrot is generally severely disadvantaged by frequent fires in heathlands. In somehabitats and areas it may also decline in long-unburnt (>15 years) vegetation, but elsewhere fire exclusion isrecommended. The impacts of fire may be compounded by habitat fragmentation, which is probablyappreciably greater now for heathlands than it was before European colonisation. Such fragmentation maydecrease the probability of recolonisation of isolates where fire (and unsuitable post-fire habitat) has destroyedlocal populations. / John Gray © Nature Focus
Eastern Bristlebird. The Eastern Bristlebird is one of agroup of species restricted to heathlands and coastalthickets in temperate southern Australia. Recently-burnt areas are unsuitable, with population densitiesincreasing as vegetation density builds up, for atleast 15 years post-fire. Its persistence in an areamay require careful management, aimed principallytoward fire suppression. However, some fire isalmost inevitable in this habitat, and remainingunburnt patches are then critical for re-establishmentof populations. / Norman Chaffer Estate © NatureFocus
Red-tailed Black Cockatoo. Like many parrots and arange of other birds, Red-tailed Black Cockatoosrequire hollows for nest sites, typically formed in veryold trees. Intense fires may destroy such importanttrees, although fire may have a role in hollowformation. In northern Australia, Red-tailed Black-cockatoos also track fires, as food is most readilyavailable in recently-burnt areas. / Babs & Bert Wells © Nature Focus

FIRE AND AUSTRALIAN BIRDSAn annotated bibliography
J.C.Z. Woinarski
Parks and Wildlife Commission of the Northern TerritoryPO Box 496
Palmerston, NT, 0831


This annotated bibliography compiles all (361)references I could locate which relate to fire andAustralian birds. It is based on systematic searchesof the journals:
Emu (1960–1995)Corella (1977–1995)Australian Bird Watcher (1970–1994)South Australian Ornithologist (1968–1994)Australian Bird Bander (1969–1976)Sunbird (1992–1995)Australian Journal of Ecology (1980–1995)(Australian) Wildlife Research (1984–1995)Proceedings of the Ecological Society of Australia(1980–1995)Northern Territory Naturalist (1978–1994)Australian Forestry (1977–1995)Australian Forest Research (1981–1987)Victorian Naturalist (1975–1992)and less comprehensive searches through
other journals, reports, theses and otherpublications, notably including the generalbibliography of Gill et al. (1994) on fire ecology inAustralia, as well as publications indicated to meby a range of authorities in most States andTerritories.
All entries are stored on the bibliographicdata base Endnote Plus. For each reference, I listkeywords and bioregion(s) in italics. Keywordsinclude the (broad) vegetation formation to whichthe article refers, the main bird taxa considered,whether the article deals with threatened taxa,whether the article describes responses of a whole
bird community, whether it deals withmanagement issues, etc. Bioregions followThackway and Cresswell (1995). No bioregionsare listed for most review articles or where theobservations reported are relatively trivial (e.g.birds seen feeding on charcoal). A map of thelocations of primary sources reported is presentedas an appendix.
The annotations given are my ownsummaries of relevant sections of the articles,except where material is presented in quotationmarks.
Much of the material presented here isdiscussed in the accompanying review.
Although I have made every attempt to becomprehensive, many studies of the relationshipbetween birds and fire are presented in the greyliterature. I would be grateful for notification ofany papers I may have missed, or any comment onmy interpretation of any articles.
Gill, A.M., Moore, P.H.R., and Martin, W.K.(1994). Bibliography of fire ecology inAustralia (including fire science and firemanagement). Edition 4. NSW NationalParks and Wildlife Service, Hurstville.
Thackway, R. and Creswell, I.D. (1995). Aninterim biogeographic regionalisation forAustralia: a framework for setting prioritiesin the National Reserves SystemCooperative Program. Version 4.0.Australian Nature Conservation Agency:Canberra.
115
INTRODUCTION

I am very grateful to Allan Burbidge, GrantWardell-Johnson, Graeme Smith, David Paton,Richard Loyn, Doug Robinson, David Baker-Gabb, Charlie Silveira, David Rounsevell, RobertTaylor, Dan Lunney, Jann Williams, HarryRecher, Hugh Ford, Dick Turner, Mike Fleming,Gay Crowley and Stephen Garnett for commentson an earlier draft and/or pointers to obscurereferences. I thank the Royal AustralasianOrnithologists Union for library access, and TonyNorton for helping arrange library access atANU. Belinda Oliver constructed theaccompanying map. I thank Neal Hardy of theDepartment of the Environment and Heritage forhis patience and assistance in this project.
116
ACKNOWLEDGEMENTS

Abbott, I. (1981). Seabird Islands: No. 106.Mondrain Island, Archipelago of theRecherche, Western Australia. Corella, 5,60–61.seabirds; heath; tussock grassland; mortalityBioregion: Esperance PlainsFires since 1801 have repeatedly consumed muchof the vegetation, destroying many breedingseabirds.
Abbott, I., & Christensen, P. (1994).Application of ecological and evolutionaryprinciples to forest management in WesternAustralia. Australian Forestry, 57, 109–122.management; control burning; eucalypt open forestBioregion: Jarrah ForestReviews some studies of the impacts of fuelreduction burning on birds in Jarrah forests,suggesting limited effect.
Adam, P., & Robinson, D. (1996). Negativeeffects of fuel-reduction burning on thehabitat of the Grey-crowned BabblerPomatostomus temporalis. Victorian Naturalist,113, 4–9.Grey-crowned Babbler; threatened species;management; control burning; eucalypt woodlandBioregions: Victorian Midlands; South EasternHighlandsFrequent (annual) fuel-reduction burning (inremnant vegetation along roadside verges) ishaving a considerable impact on the habitat (andhence numbers) of the threatened Grey-crownedBabbler. Warns that findings of little impact offuel-reduction burning on birds should be treatedwith caution. Alternative mechanisms of fireprotection (e.g. slashing) should be considered inorder to safeguard these populations.
Arnold, G. W., Smith, G. T., Rowley, I. C. R.,& Brooker, M. G. (1993). The effects of fireon the abundance and distribution of animalsin Australian ecosystems, with emphasis onMediterranean ecosystems. In L. Trabaud &R. Prodon (Eds.), Fire in Mediterraneanecosystems (pp. 237–257). Brussels:Commission of the European Communities.review; heath; mallee; eucalypt open forest; isolateReviews studies of effects of fire on birds (andother fauna) in Mediterrean ecosystems (heath,mallee and open forest) in southwestern andsoutheastern Australia. More studies are requiredon the influence of fire mosaic or patchiness.Mammals (at least in eastern Australia) showpronounced seral responses to fire. Lizards andfrogs are generally not greatly affected by fire.Fauna of mediterranean systems shows less clearseral responses to fire than those in temperateecosystems, as the fauna of mediterraneanecosystems has adapted to substantial climaticvariability. There are substantial differencesbetween bird species in response to fire, forexample the Ground Parrot is fire dependentwhereas Noisy Scrub-bird is fire sensitive.Long-term demographic studies are needed tocomprehend and predict responses to fire. Firemanagement is an essential component of themanagement of conservation reserves: this may becomplicated by fragmentation.
Ashby, E. (1921). Notes on the supposed “extinct” birds of the south-west corner ofWestern Australia. Emu, 20, 123–124.Ground Parrot; Western Whipbird; succession; heath;threatened species.Bioregion: Swan Coastal PlainFrequent fires (lit by farmers) had changedheathlands to grasslands at a site in southwesternAustralia eliminating Ground Parrots andWestern Whipbirds.
117
REFERENCES

Ashby, E. (1924a). Notes on extinct or rareAustralian birds, with suggestions as to someof the causes of their disappearance. Part I.Emu, 23, 178–183.review; threatened species; Night Parrot; hummockgrassland; Ground Parrot; heath; wetlands; KangarooIsland Emu.Speculates that Night Parrots persist only wherehummock grasses are “unswept by fires”.Disappearance of Ground Parrot due to frequentfires. Extinction of Kangaroo Island Emu was dueto hunting and excessive burning.
Ashby, E. (1924b). Notes on extinct or rareAustralian birds, with suggestions as to someof the causes of their disappearance. Part II.Emu, 23, 294–298.review; threatened speciesReview of decline of Australian birds. “The mostserious factor of all is the destruction of both foodsupply and shelter, and, of course, breeding haunts,by bush-fires. I consider that these … factors,especially … bush-fires, account for more thannine-tenths of the disappearance of certain forms… I am confident that the indiscriminate burningof bush, which is the concomitant of all farmingand grazing operations in bush country, is by a longway the major cause of the disappearance of manyof our rarer birds.”
Aumann, T., & Baker-Gabb, D. J. (1991). Theecology and status of the Red Goshawk innorthern Australia. RAOU Report No. 75. RoyalAustralasian Ornithologists Union.Red Goshawk; tropical eucalypt open forest; threatenedspecies; management.Fires have been reported to burn out the nests ofRed Goshawk, killing young. In the long term, acombination of grazing and frequent fires mayresult in reduction of prey and loss of nest trees,however short-term results show little clearindication of differences in density or breedingsuccess related to burning history.
Australian Biological Research Group Pty.Ltd. (1987). Management Plan for the GroundParrot in the Orbost Region. AustralianBiological Research Group Pty Ltd.Ground Parrot; heath; sedgelands; threatened species;management.Bioregion: South east CornerFire regimes proposed for heathlands to maintain
suitability for Ground Parrots.
Backhouse, G. N. (1987). Management ofremnant habitat for conservation of theHelmeted Honeyeater Lichenostomus melanopscassidix. In D. A. Saunders, G. W. Arnold,A. A. Burbidge, & A. J. M. Hopkins (Eds.),Nature conservation: the role of remnants ofnative vegetation (pp. 287–294). ChippingNorton: Surrey Beatty.Helmeted Honeyeater; eucalypt open forest; wildfire;management; threatened species; isolateBioregion: South Eastern HighlandsThe sites of two small isolated populations ofHelmeted Honeyeaters were severely burnt bybushfire, destroying much of the habitat. Bothpopulations disappeared after fire. At the site ofthe remaining population (a narrow riparian strip)fire prevention works may create moredisturbance than fire.
Baird, R. F. (1986). Historical records of theGlossy Black Cockatoo Calyptorhynchuslathami and Red-tailed Black CockatooC. magnificus in south-eastern Australia. SouthAustralian Ornithologist, 30, 38–45.Glossy Black-Cockatoo; historic change; threatenedspecies; review; Casuarina woodlandBioregion: WoolnorthConsiders historic change in the distributions ofGlossy Black-Cockatoo and Red-tailedBlack-Cockatoo. Declines initially as a result ofclimate change, more recently exacerbated byclearing and fire (at least for GlossyBlack-Cockatoo on King Island)
Baker, J., & Whelan, R. J. (1994). GroundParrots and fire at Barren Grounds, NewSouth Wales: a long-term study and anassessment of management implications. Emu,94, 300-304.Ground Parrot; threatened species; heath.Bioregion: South East CornerSome previous studies have suggested thatsuitability of heaths for Ground Parrots (and henceGround Parrot abundance) peaks at around 10years post-fire, and hence have recommended fireintervals of 8-10 years. In contrast, this studysuggests that there is little evidence for a decline insuitability, but rather that abundance plateaus (at c0.25birds/ha) after around 5 years. This was basedon a series of censuses of the same locality fromimmediately after fire to 10.7 years post-fire. Fewstudies have examined abundance in heaths >15years. No single fire regime is typical of all areaswhere Ground Parrots occur.
118
Australia’s Biodiveristy - Responses to Fire

Baker, L., Woenne-Green, S., & MutitjuluCommunity (1993). Anangu knowledge ofvertebrates and the environment. In J. R. W.Reid, J. A. Kerle, & S. R. Morton (Eds.),Uluru fauna. The distribution and abundance ofvertebrate fauna of Uluru (Ayers Rock-MountOlga) National Park, N.T. (pp. 79-132).Canberra: Australian National Parks andWildlife Service.management; Aboriginal knowledge; mallee; hummockgrassland; Acacia woodland.Bioregion: Central RangesDescribes Aboriginal knowledge of wildlife. Manygranivores are favoured by fire, but Emu dislikefreshly-burnt country. Burning in spring candisrupt their breeding and cause them to moveaway. Fires lit in strong winds can kill many smallanimals. Discusses traditional Aboriginal burningregimes.
Baker-Gabb, D. J., Benshemesh, J. S., &Maher, P. N. (1990). A revision of thedistribution, status and management of thePlains-wanderer Pedionomus torquatus. Emu,90, 161-168.Plains-wanderer; tussock grassland; wildfire;management; threatened species.Bioregions: Murray Darling Depression; ChannelCountry; Riverina; Victorian Volcanic PlainPlains-wanderers are sedentary in sparsegrasslands unless these are overgrazed, burnt orcultivated. It is likely that most young and manyadults at one site perished when an extensive fireswept through the area, and wildfire may havesevere impact on this species. This was the firstsuch fire recorded from this area for 70 years.
Baldwin, M. (1965). Bird eating charcoal.Emu, 64, 208.charcoalFour species (Fairy Martin, Dusky Wood-swallow,Double-barred Finch and Zebra Finch) recordedeating charcoal.
Bamford, M. J. (1985a) The dynamics of smallvertebrates in relation to fire in banksiawoodland near Perth, Western Australia. PhD,Murdoch University.Banksia woodland; succession; communityBioregion: Swan Coastal PlainBird communities were assessed at six sites acrossa range of ages (0-22 years) since fire. Soon afterfire in Banksia woodland, bird richness decreased,but the site was colonised by open-country birdsfrom surrounding farmlands.
Bamford, M. J. (1985b). The fire-relateddynamics of small vertebrates in Banksiawoodland: a summary of research in progress.In J. R. Ford (Ed.), Fire ecology andmanagement in Western Australian ecosystems(pp. 107-110). Perth: Western AustralianInstitute of Technology.Banksia woodland; community.Bioregion: Swan Coastal PlainSix Banksia sites with a range of fire histories weremonitored for vertebrates. Birds were moreaffected by fire than reptiles but less thanmammals. Most bird species were recorded withina few months of fire. At one site, the number ofbird species was dramatically lower soon after afire, but returned to pre-fire levels within 1 year,although total number of individuals remainedlow for at least 2 years. No species showed apreference for the long unburnt sites. A fewspecies were more common in the most recentlyburnt sites. These were mainly species typical ofthe surrounding farmlands.
Bates, R. (1980). After the fire: someobservations on the effect of the February 20,1980 (Ash Wednesday) bushfires on wildlife inthe Adelaide Hills. South Australian Naturalist,54, 77-79.wildfire; eucalypt open forestBioregion: Lofty Block.
119
Fire and Australian birds

Bedggood, G. W. (1980). Birdlife betweenLake Tyers and Marlo, Victoria. AustralianBird Watcher, 8, 147-162.heath; eucalypt open forest; control burning.Bioregion: South east CornerSuggests that the widespread spring fuel reductionburns result in heavy toll of young birds and eggs,excessive loss of hollows suitable for breeding, lossof protective undergrowth, loss of ground-nestingbirds, and reduced germination forspring-flowering plants.
Beeton, R. J. S. (1985). The little corella: aseasonally adapted species. Proceedings of theEcological Society of Australia, 13, 53-63.Little Corella; tussock grassland; tropical eucalyptsavanna woodlandBioregions: Ord-Victoria Plains;Victoria-BonaparteAt a season when food is generally limited(March-April), Little Corellas flock to small areasrecently burnt, presumably because such firesincrease accessibility to seeds.
Belcher, C. (1993). Rufous Bristlebird surveyand habitat analysis, Port Campbell NationalPark 1992. Department of Conservation andNatural Resources.Rufous Bristlebird; heath; thicket; threatened species;isolateBioregion: South East Coastal PlainAt Port Campbell National Park optimal habitatfor Rufous Bristlebird is climax heathland morethan 25 years post-fire. Isolated populations aresusceptible to elimination by fire, and the poordispersal ability of the species may limitsubsequent recolonisation
Bennett, S. (1983). The Northern Scrub-robinDrymodes superciliaris in the NorthernTerritory. Emu, 83, 105-107.Northern Scrub-robin; rainforest; threatened speciesBioregion: Gulf CoastalIf records from the Roper River earlier thiscentury are valid, the Northern Scrub-robin hasbecome extinct in the NT, most likely because ofchange in fire regimes leading to decline in theextent of rainforest thickets.
Benshemesh, J. (1988). Report on a study ofmalleefowl ecology. Department ofConservation, Forests and Lands.mallee; Malleefowl; threatened species; wildfireBioregion: Murray Darling DepressionA detailed case study of the response ofMalleefowl to an intense but patchy fire. Resultsare summarised in Benshemesh (1990). Smallunburnt patches were vital for the persistence ofthe species. Long fire-free intervals (>60 years) areoptimal.
Benshemesh, J. (1990). Management ofMalleefowl - with regard to fire. InJ. C. Noble, P. J. Joss, & G. K. Jones (Eds.),The mallee lands: a conservation perspective(pp. 206-211). Melbourne: CSIRO.mallee; Malleefowl; threatened species; wildfireBioregion: Murray Darling DepressionOptimal fire frequency for Malleefowls is >60years. Broad-scale fires eliminate Malleefowl inthe short-term, and even 20-30 years post-fire,breeding densities are only about one third ofthose in long-unburnt mallee (based on fourmatched pairs of 20-30 year old and >40 year oldsites). Patchy burns provide some refuges fromwhich recolonisation can occur. Sufficient litterfor nesting is generally unavailable until 10-15years post-fire. Previous authors had suggestedthat food resources were more abundant inyounger mallee and that periodic fire was neededfor their maintenance. Modelled densities were6% of maximum carrying capacity for 20 year fireinterval, 30% at 40 year intervals, and 54% at 60year intervals.
At least 10 of 11 radio-marked and bandedbirds survived a patchy but intense fire, but in themonths following the fire all but 4 had emigratedor died. Dispersing birds used corridors ofunburnt vegetation rather than traversingextensive burnt areas. The remaining birds bredin the season following fire in small unburntpatches. These birds foraged extensively in burntareas, using the rich pulse of herbs growing afterfire.
120
Australia’s Biodiveristy - Responses to Fire

Benshemesh, J. S. (1992) The conservationbiology of Malleefowl, with particular regard tofire. PhD, Monash University.Malleefowl; mallee; Callitris woodland; wildfire;old-growth; threatened species.Bioregion: Murray Darling DepressionDetailed study of habitat requirements andbiology of the Malleefowl, particularly in responseto a wildfire. Malleefowl requires long-unburntmallee to provide the extensive litter needed forbreeding. Such old-growth has become extremelylimited. Malleefowls may survive fires if unburntpatches occur.
Benshemesh, J. S. (1994). Malleefowl Leipoaocellata. Department of Conservation andNatural Resources.Malleefowl; mallee; threatened species; wildfire;managementBioregion: Murray-Darling DepressionReviews information on Malleefowl (in Victoria).“The extent and frequency of fires pose a seriousthreat to the conservation of Malleefowl asremaining populations may be destroyed andhabitat quality reduced for 40 years or more … theeffect of fire is exacerbated by the fragmentationdue to clearing, as isolated reserves that areentirely burnt are unlikely to be recolonised …more effective fire control may be the single mostimportant factor in improving the conservationstatus of the species.” The scale of fire in malleelands is a problem, as it may be comparable to thatof the largest reserves. Birds may survive and breedafter patchy fires.
Bill, M. E. (1932). Lyre-birds and bushfires.Victorian Naturalist, 49, 24.Superb Lyrebird; wildfire
Blakers, M., Davies, S. J. J. F., & Reilly, P. N.(1984). The atlas of Australian birds.Melbourne: Melbourne University Press.reviewDistributional information on all Australian birds,but includes information on threateningprocesses. Notes Orange-bellied Parrot prefersheaths and button-grass plains <15 years post-firefor feeding. Increase in fire frequencies orintensities may have led to decline in GouldianFinch and Pictorella Mannikins. Notes referencesto fire for other species (e.g. Emerald Dove,Partridge Pigeon, Paradise Parrot, Scarlet-chestedParrot, Golden-headed Cisticola, White-throatedGrass-wren, Eastern Bristlebird).
Boekel, C. (1980). Birds of Victoria RiverDowns Station and of Yarralin, NorthernTerritory. Part 1. Australian Bird Watcher, 8,171-193.tropical eucalypt savanna woodland; huntingBioregion: Ord-Victoria PlainsDescribes the use of smoke and traps byAboriginal people for catching eagles and kites.
Bowman, D. M. J. S., Woinarski, J. C. Z., &Russell-Smith, J. (1994). Environmentalrelationships of Orange-footed ScrubfowlMegapodius reinwardt nests in the NorthernTerritory. Emu, 94, 181-185.Orange-footed Scrubfowl; historic change; rainforest.Bioregions: Top End Coastal; Pine CreekArnhem; Central ArnhemEnvironmental changes, probably includinghistoric changes in fire regime, have led to somecontraction of rainforests, as evidenced by theoccurrence of old scrubfowl mounds in areaswhich are now eucalypt open forests.
Braithwaite, L. W., Clayton, M., MacLean, L.,& Parker, B. S. (1984). Vertebrate fauna of a144-ha water catchment within eucalypt forestbeing harvested for woodpulp at Eden,south-eastern New South Wales. CSIROWildlife and Rangelands Research.eucalypt open forest; wildfire; communityBioregion: South East CornerBirds were surveyed at a site for 15 months beforeand 2 months after January wildfire. Parts of thesite were logged during this period. Speciesrichness declined after fire. LaughingKookaburra, Grey Butcherbird, Flame and ScarletRobins increased after fire. Striated Thornbill,Red Wattlebird, Crescent Honeyeater and PiedCurrawong decreased.
121
Fire and Australian birds

Braithwaite, R. W. (1985). Fire and fauna. InR. W. Braithwaite (Ed.), Kakadu Fauna Survey.Final report to Australian National Parks andWildlife Service. (pp. 634-650). Darwin:CSIRO.management; tropical eucalypt open forest; tropicaleucalypt savanna woodland; rainforestBioregions: Pine Creek Arnhem; Top End CoastalA wide range of carnivorous and hawking birds areattracted to fire. Soon after fire, many carnivorousand granivorous birds move into burnt areas totake advantage of greater abundance oraccessibility of resources. Some nests (of BrownHoneyeater, Mistletoebird, White-throatedHoneyeater) were found to be destroyed by fire.
Braithwaite, R. W. (1996). Biodiversity andfire in savanna landscapes. In O. Solbrig,E. Medina, & J. F. Silva (Eds.), Biodiversity andsavanna ecosystem processes: a global perspective(pp. 121-140). Berlin: Springer-Verlag.tussock grasslands; tropical eucalypt savanna woodland;tropical eucalypt open forest; reviewReviews some previous studies from northernAustralia. In general, there is little succession, butincrease in bird abundance soon after fires.
Braithwaite, R. W., & Estbergs, J. (1987).Fire-birds of the Top End. Australian NaturalHistory, 22, 299-302.community; tropical eucalypt open forest; tropicaleucalypt savanna woodland.Bioregions: Top End Coastal; Pine Creek ArnhemSeveral species (notably Brown Falcon, BlackFalcon, Whistling Kite, Black Kite,woodswallows, Tree Martin) are attracted to fires,often in large aggregations. Torresian Crow,raptors, butcherbirds, Straw-necked Ibis,Black-faced Cuckoo-shrike, nightjars, Red-tailedBlack-cockatoo, Little Corella, Galah, quail,Northern Rosella, Blue-winged Kookaburra,Red-backed Kingfisher, Forest Kingfisher,Partridge Pigeon and Magpie-lark forage in theburnt area immediately (to several months) afterfire, consuming carrion, more accessible animalfood or fallen seeds. In the months following fire,vegetation regrowth attracts herbivorous insectsand birds feeding on them (e.g. Straw-neckedIbis). In the wet season, many of these speciesmigrate into more arid areas where fires continueto occur, though most are then no longerassociated with burnt areas but rather open
country around waterholes. The short-termattraction of many mobile species to burnt areas isin contrast to the succession described intemperate areas.
Bramwell, M., Pyke, G., Adams, C., & Coontz,P. (1992). Habitat use by Eastern Bristlebirdsin Barren Grounds Nature Reserve. Emu, 92,117-121.Eastern Bistlebird; threatened species; heath; eucalyptwoodland.Bioregion: South east CornerEastern Bristlebird population density was greaterin heath/woodland 9 years after fire than 6 yearsafter fire.
Brickhill, J. (1980). Striated GrasswrenAmytornis striatus. In C. Haigh (Ed.),Endangered animals of New South Wales(pp. 68). Sydney: NSW National Parks andWildlife Service.threatened species; mallee; hummock grassland;Striated Grass-wrenExtensive wildfires in mallee have severe andlong-term impacts on Striated Grass-wren.
Brickhill, J. (1987) The conservation status ofmalleefowl in New South Wales. M.Nat.Res.Sc.,University of New England.mallee; Malleefowl; threatened speciesBioregion: Murray Darling DepressionPeriodic fire may be needed to maintain the foodresources on which the Malleefowl depends. Afine-scale fire-age mosaic benefits Malleefowlthrough provision of a diversity of food types.
Brooker, L. C., & Brooker, M. G. (1994). Amodel for the effects of fire and fragmentationon the population viability of the SplendidFairy-wren. Pacific Conservation Biology, 1,344-358.Splendid Fairy-wren; isolate; management; wildfire;control burning; heath; long-term studyBioregion: Swan Coastal PlainBased on 17 years breeding, parasitism andsurvival data, a model of the effects of fire (bothwildfire and controlled burning) on populations ofSplendid Fairy-wrens is developed and explored.The frequency and extent of fire can readily driveisolated populations to extinction. This
122
Australia’s Biodiveristy - Responses to Fire

probability increases with smaller population size(=smaller area of isolate), and increase inprobability of fire. In this case, the main effect offire is manifested through increase in subsequentnest predation.
Brooker, M. G. (1988). Some aspects of thebiology and conservation of the Thick-billedGrasswren Amytornis textilis in the Shark Bayarea, Western Australia. Corella, 12, 101-108.Thick-billed Grass-wren; successionBioregion: CarnarvonIncreased tourism and de-stocking could lead tochanges in fire regime. Evidence of past severefires, but current frequency of fires is low.Thick-billed Grasswrens can occur in vegetationwithin a few years post-fire, but fire is likely to bea main management consideration.
Brooker, M. G., & Rowley, I. (1991). Impactof wildfire on the nesting behaviour of birds inheathland. Wildlife Research, 18, 249-263.heath; eucalypt open forest; wildfire; long-term study;Western Thornbill; Splendid Fairy-wren;Yellow-rumped Thornbill; breeding; control burningBioregion: Swan Coastal PlainAn intense wildfire burnt almost all of the studysite half-way through a nine-year study: threewildfires burnt the rest of the study area over thenext 3 years. Birds changed their nest siting afterfires. In contrast to pre-fire locations, SplendidFairy-wrens nested only in resprouter plantspecies in the first year post-fire. By the secondyear post-fire, they used some seeding shrubs.They had difficulty attaching nests to post-firesubstrate, with several nests falling to the ground.Western Thornbills were unable to nest inpreferred Hakea in the year post-fire but nestedinstead in holes of eucalypts and under sheddingbark. They changed their placement of nests inXanthorrhea. Very small unburnt patches werenot used selectively by any of the three main birdspecies considered. The height distribution ofnests generally changed in burnt vegetation.Breeding of Splendid Fairy-wren was delayed by3-5 weeks in the year after fire, and the number ofnests built per group had almost doubled by thesecond year (probably due to high rate of nestfailure). Western Thornbills also delayed breedingby up to 5 weeks in burnt areas (cf unburnt) in theyear after fire, and only 59% of females attemptedto breed. The delay was due to shortage of nesting
material (e.g. cobwebs) and/or inadequate food foregg production. Yellow-rumped Thornbills usednovel nest sites post-fire, but used only 2 speciesof plant as nest sites, compared to 11 pre-fire.
Of 26 species which bred in the site in theyear preceding the fire, 21 nested in burnt areas inthe year following fire. White-browed Scrubwrenvacated the area and did not nest for 2 yearspost-fire (possibly due to limited food resourcesthrough lack of litter). White-cheekedHoneyeater did not nest until 4 years post-fire.Inland Thornbill became rare and had failed tonest by 5 years post-fire. Fantailed Cuckooremained abundant at the site but did not breeduntil its hosts (White-browed Scrubwren andInland Thornbill) returned to breed. LittleButton-quail and Elegant Parrot were recordednesting only after the fire (attracted to thetemporary conversion of heath to grassland), andWhite-winged Triller and Dusky Wood-swallowwere more numerous breeders post-fire.
Although most species appeared to havesome individuals which survived fire and showedadaptable behaviour in subsequent nesting, lessobvious effects (such as increased mortality,lowered productivity and altered age structure)may be more critical in determining the ultimateviability of populations.
“For small heathland passerines, controlburns at any time of year and even five years apartcould make an area uninhabitable.” Suggestsintervals of perhaps as long as 10 years are neededto maintain these populations.
Brooker, M. G., Ridpath, M. G., Estbergs, A. J.,Bywater, J., Hart, D. S., & Jones, M. S. (1979).Bird observations on the north-westernNullabor Plain and neighbouring regions,1967-1978. Emu, 79, 176-190.chenopod shrubland; wildfireBioregion: NullaborA widespread fire followed by drought may haveled to local loss (for at least 5 years) ofWhite-winged Fairy-wren from a bluebush site.
123
Fire and Australian birds

Brooker, M. G., Braithwaite, R. W., &Estbergs, J. A. (1990). Foraging ecology ofsome insectivorous and nectarivorous speciesof birds in forests and woodlands of theWet-Dry tropics of Australia. Emu, 90,215-230.tropical eucalypt open forest; tropical eucalypt savannawoodland; communityBioregions: Pine Creek Arnhem; Top End CoastalStudy of foraging behaviours of insectivorous andnectarivorous birds across a range of habitats. “… controlled burning and the prevention ofpreviously frequent wildfires are but a few examplesof disturbances which may change … the abundanceand diversity of birds.” More than half of theinsectivorous species forage from the ground orfrom shrubs and grass. “The lower strata are themost affected by fire … (and) the needs of this largeground and shrub-foraging guild should thereforebe considered in the formulation of firemanagement plans. A range of fire types throughoutthe year might increase the habitat diversity … witha positive effect on the diversity of bird species.”
Brothers, N. P. (1983). Seabird Islands: No.136. Actaeon Island, Tasmania. Corella, 7,89-90.seabirds; heath; tussock grasslandBioregion: D’Entrecasteaux“The vegetation on the island has been frequentlyburnt for many years and … this may havesignificantly affected the habitat suitable forburrowing” for nesting seabirds.
Brothers, N. P., & Skira, I. J. (1987). SeabirdIslands: No. 173. Chappell Island, FurneauxGroup, Tasmania. Corella, 11, 81-82.seabirds; tussock grasslandBioregion: FurneauxFiring and grazing has substantially modified thevegetation, considerably reducing the distributionand abundance of breeding shearwaters.
Brothers, N. P., & Skira, I. J. (1988). SeabirdIslands: No. 185. Little Dog Island, FurneauxGroup, Tasmania. Corella, 12, 85-86.seabirds; tussock grasslandBioregion: FurneauxRepeated fires have changed vegetationpatterning, probably affecting suitability forshearwaters.
Brouwer, J., & Garnett, S. (Ed.). (1990).Threatened birds of Australia: an annotated list.Melbourne: Royal Australasian OrnithologistsUnion.review; threatened speciesOf 52 threatened bird species, inappropriate fireregime is a factor affecting status for 22 species.
Brown, P. B., & Wilson, R. I. (1981). A surveyof the Orange-bellied Parrot Neophemachrysogaster in Tasmania, Victoria and SouthAustralia. National Parks and Wildlife Service,Tasmania.Orange-bellied Parrot; threatened species;management; eucalypt open forest; sedgelands; hollowsBioregion: West and South WestUncontrolled burning of button-grass plains insouthwestern Tasmania extend to the breeding sitesof Orange-bellied Parrots in fringing open forest,to the extent that, when nesting, they “areconstantly at risk from wildfires”. However, theParrots feed in the button grass plains, sedgelandsand heaths, and prefer relatively young vegetation(albeit in a seasonally rotating order: in Octoberand November they fed mainly in 7-8 year oldregrowth, in December they mainly fed in 1-4 yearold regrowth, in January and February they mainlyfed in regrowth >8 years post-fire). A firemanagement plan is required.
Brown, P. B., & Wilson, R. I. (1984). TheOrange-bellied Parrot. In R. H. Groves & W. D. L. Ride (Eds.), Species at risk (pp.106-116). Canberra: Australian Academy ofScience.Orange-bellied Parrot; heath; sedgelands; threatenedspecies; management.Bioregion: West and South WestFire management is critical for the maintenance ofOrange- bellied Parrot at their breeding grounds.Relatively young regrowth heath (7 years post-fire)is preferred for feeding, and is probably unsuitableby 10-12 years. In its breeding area it roosts in thickregrowth of 7-10 years post-fire.
124
Australia’s Biodiveristy - Responses to Fire

Bryant, S. L. (1991). The Ground Parrot,Pezoporus wallicus, in Tasmania: distribution,density and conservation status. Scientific reportno. 91/1. Parks, Wildlife & Heritage,Tasmania.Ground Parrot; threatened species; heath; sedgelands.Bioregion: West and South WestProvided density estimates of Ground Parrots at185 sites of varying ages post-fire. No clear peakin abundance at a particular age. Did notrecommend prescribed burning on currentinformation.
Bryant, S. L. (1992). The Ground Parrot andage of vegetation in Tasmania. In L. Joseph(Ed.), Issues in the conservation of parrots inAustralasia and Oceania: challenges toconservation biology. (pp. 42-45). Melbourne:Royal Australasian Ornithologists Union.Ground Parrot; threatened species; heath.Bioregion: West and South WestGround Parrots occur in heath across a very broadrange of ages after fire, though are least abundant inearly regrowth.
Bryant, S. L. (1994). Habitat and potential dietof the Ground Parrot in Tasmania. Emu, 94,166-171.threatened species; heath; sedgelands; Ground Parrot.Bioregion: West and South WestRecorded in vegetation from 1 to 90 yearspost-fire. Minimum vegetation cover required isachieved in most situations in Tasmania within12 months post-fire. Growth rates (and phenology)of moorland plant species are very variable, leadingto wide range of vegetation ages used.
Burbidge, A. A., Folley, G. L., & Smith, G. T.(1986). The Noisy Scrub-bird. WesternAustralian Wildlife Management Program 2.Department of Conservation and LandManagement.Noisy Scrub-bird; eucalypt open forest; heath;threatened species; management.Bioregion: Esperance PlainsThe former frequent burning and drainage ofswamps would have led to decline of this species.Management requires fire control.
Burbidge, A. H., & Pedler, L. (1993).Conservation status of the NullaborQuail-thrush. World Wide Fund for Nature.Nullabor Quail-thrush; threatened species; chenopodshrubland; managementBioregion: NullaborHot fires may make areas unsuitable for NullaborQuail-thrush, and this effect may be exacerbatedby grazing preventing regrowth.
Burbidge, A. H., & Pedler, L. (1997). Habitatof the threatened Nullabor Quail-thrush.Wildlife Research, 24.Nullabor Quail-thrush; chenopod shrublands;threatened species; managementBioregion: NullaborInteracting with grazing by rabbits and livestock,and weed invasion, fire is a main threat to theNullabor Quail-thrush. Habitat should bemanaged to decrease risks (or frequency) ofextensive fire (especially hot wildfire).
Burbidge, A. H., Watkins, D., & McNee, S.(1989). Conservation of the Ground Parrot inWestern Australia. Final report (Project 118).World Wildlife Fund.Ground Parrot; threatened species; heath;management; control burningBioregion: Esperance PlainsGround Parrots in WA prefer long-unburntheath, but it is not clear at what age heathsbecome suitable. “The major approach for themanagement of Ground Parrots in these areas atthis time must be one of total fire exclusion”. Iffuel reduction burning must be done (for bufferstrips) this should be done in autumn, to avoiddisruption of breeding). Describes floristics andresults of radio-tracking.
Burbidge, A. H., McNee, S., Newbey, B., &Rolfe, J. (1990). Conservation of the GroundParrot in Western Australia. Supplementaryreport: project 118. World Wildlife Fund.Ground Parrot; heath; management; threatenedspecies.Bioregion: Esperance PlainsResults of some censuses and floristic associationsfor Ground Parrot in southwest Australia. Thewestern subspecies appears to have very differenthabitat and management requirements to theeastern subspecies. Monitoring of sites followingwildfire is outlined.
125
Fire and Australian birds

Cale, P. G., & Burbidge, A. H. (1993). Researchplan for the Western Ground Parrot, WesternWhipbird and Western Bristlebird. AustralianNational Parks and Wildlife Service.Ground Parrot, Western Whipbird, WesternBristlebird; threatened species; management; heath;mallee.Bioregions: Esperance Plains; Jarrah Forest;WarrenReviews conservation status, research prioritiesand responses to fire for three threatened species(Western Whipbird, Western Bristlebird andGround Parrot). Fire management of these threespecies (and also Noisy Scrub-bird) should beintegrated at sites of co-occurrence, although thismay be complicated by somewhat differentresponses to fire. For Ground Parrot, cautionsagainst correlative studies of abundance and timesince fire (other factors may be involved). Limiteddata from WA suggest response may differ to thatrecorded from eastern Australia: birds in WA uselong-unburnt vegetation (>20, and maybe >30years post-fire), and are more abundant in along-unburnt area than an adjacent 6-yearpost-fire area. This difference may reflect slowergrowth rates of heath in WA. “Determining theage after fire at which vegetation becomes suitablefor Ground Parrots and the length of time forwhich it remains suitable is essential for theproper long-term management of the species”:this can only be done by monitoring existingpopulations and/or determining the time after fireat which colonisation occurs. “Fire has beenidentified as the greatest threat to P.n.nigrogularis … (but) little is known about theresponses to fire of P.n. oberon”. Managementand research for the former subspecies shouldconsider whether there is an upper limit on theage of vegetation used; for the latter subspeciesresearch should examine more comprehensivelylong-term responses of known populations tovegetation age and hence derive fire managementplans. For Western Bristlebirds, research hasindicated that heaths older than c45 years becomeless suitable (due to decrease in productivity andfloristic/structural changes). Research andmanagement should continue long-termmonitoring of population size with heaths ofincreasing age, and examination of the impacts offire breaks.
Calver, M. C., Hobbs, R. J., Horwitz, P., &Main, A. R. (1996). Science, principles andforest management: a response to Abbott andChristensen. Australian Forestry, 59, 1-6.management; control burning; eucalypt open forestBioregion: Jarrah ForestDisputes claims (made by Abbott andChristensen) of limited impacts of fuel reductionburning in jarrah forests; no new data arepresented.
Campbell, A. G. (1937). Birds of Wilson’sPromontory. Emu, 37, 157.eucalypt open forestBioregion: South east Coastal PlainBirds in the Wilson’s Promontory area have beenmuch less common because of fire, and are likelyto be further affected. “The forests of the westcoast are gone and the forests of the east aredoomed owing to the ravages of fire … the utterlysenseless waste of the natural assets of thisnational park can only be termed a satire upon themethods of those in control”.
Carpenter, G., & Matthew, J. (1986). Thebirds of Billiatt Conservation Park. SouthAustralian Ornithologist, 30, 29-37.mallee; Callitris woodlands; community; hollowsBioregion: Murray Darling DepressionVegetation of several ages since fire was sampled.“Fire has an important influence on birdpopulations within the Park due to its effect onvegetation. For example, hollows suitable fornesting are occasionally produced by fires.Elsewhere, suitable hollows exist only inlong-unburnt eucalypts and Callitris.” Areas burnt<10 years before were unsuitable for Gilbert’s andRed-lored Whistlers, Southern Scrub-robin andWestern Whipbird, but suitable for Hooded andRed-capped Robins. Relationship of birds withage since fire may be changed with varyingfloristics: Callitris is slower-growing thaneucalypts and maintains shrubbiness longer,therefore species requiring shrubby vegetationmay persist in Callitris longer than in eucalypts.Southern Scrub-robin, Shy Heath-wren, CrestedBellbird and Purple-gaped Honeyeater werecommon in areas burnt 23 years previously, orolder areas that had shrubby understorey.Mallee-fowl mounds were most common in areasolder than 23 years post-fire with openunderstorey, but recently burnt areas may offer
126
Australia’s Biodiveristy - Responses to Fire

rich food supplies for this species. Several speciesshowed no apparent relationship with fire (GreyShrike-thrush, Golden Whistler, Splendid andVariegated Fairy-wrens, White-browed Babbler,Weebill, Inland Thornbill, Yellow-rumpedPardalote, Grey Butcherbird, and Brown-headed,Yellow-plumed, White-eared and Spiny-cheekedHoneyeaters).
Carter, T. (1923a). Birds of the Broome Hilldistrict. Part I. Emu, 23, 125-142.Malleefowl; mallee; heath; threatened speciesBioregion: Avon WheatbeltIn 1902 the Malleefowl was common in coastalvegetation, but its population had diminished by1920 because of burning of the coastal vegetationto improve grazing for cattle.
Carter, T. (1923b). Birds of Broome Hilldistrict. Part II. Emu, 23, 223-235.Western Whipbird; heath; threatened speciesBioregion: Avon WheatbeltRepeated fires have destroyed much of the heathand thickets that the Western Whipbird requires,and it is becoming rarer.
Carter, T. (1924). Birds of the Broome HillDistrict. Part III. Emu, 23, 306-318.Rufous Bristlebird; heath; threatened speciesBioregions: Warren; Avon WheatbeltRepeated burning of heaths had severe impact onRufous Bristlebird.
Catling, P. C., & Newsome, A. E. (1981).Responses of the Australian vertebrate fauna tofire. In A. M. Gill, R. H. Groves, & I. R. Noble(Eds.), Fire and the Australian biota (pp.273-310). Canberra: Australian Academy ofScience.reviewConsiders a series of propositions that theAustralian vertebrate fauna is fire-adapted. Ineucalypt forests, the greatest bird diversity occursin forests 5-6 years post-fire then mature forestsdeclines as habitat. There is little evidence ofdistinct seral stages, but simply gradual changes inabundance. All species in eucalypt forests have thecapacity to survive fires and recover quickly. Thereare very few fire specialists. It is difficult to ascribelife history traits to adaptation to fire as opposed to
other features of the Australian environment.Primitive birds are mainly in least fire-proneenvironments (rainforests). Fire-proneenvironments tend to be species-poor overall.Ground Parrots may be fire-specialists, requiringheath of a certain range of ages post-fire. SuperbLyrebird and Common Bronzewing may requirefire to eliminate wiregrass and enhance productionof Acacia seeds respectively. White-browedScrubwren decreased after fire in a eucalypt forest.
In eucalypt forests, bird numbers in theunderstorey and ground layers decline with longperiods after fire, and more frequent burning maybe required for these lower layers.
Chaffer, N. (1954). The Eastern Bristle-bird.Emu, 54, 153-162.Eastern Bristlebird; threatened species; heath; wildfireBioregion: South East Corner.Repeated bushfires may have reduced thenumbers of Eastern Bristlebirds, although theyhave survived in areas of frequent fire.
Chambers, W. (1983). Birds at Urquhart’sBluff: before and after the fire. GeelongNaturalist, 20, 50-51.eucalypt open forest; wildfireBioregion: South east Coastal PlainAnecdotal notes of species before fire and thengradually recolonising after fire. By six weekspost-fire, Australian Magpie, Grey Shrike-thrush,Crimson Rosella, Pied Currawong, GreyCurrawong, Australian Raven, Welcome Swallow,Masked Lapwing, Sulphur-crested Cockatoo andYellow-tailed Black Cockatoo had been recorded.Within weeks after that, first returns ofWhite-eared Honeyeater, Eastern Spinebill andFlame Robin.
Chandler, L. G. (1973). In the wake of abushfire. Wildlife in Australia, 10, 140-141.wildfire; malleeBioregion: Murray Darling DepressionGeneral notes, suggesting high mortality of smallbirds during mallee wildfire. Malleefowl may beparticularly disadvantaged, with eggs lost, youngkilled and regrowth vegetation unsuitable. Effectsmay be accentuated if drought follows fire.
127
Fire and Australian birds

Chatto, R. (1995). The effects of fire on abreeding colony of Australian Pelicans.Corella, 19, 70.Australian Pelican; mortalityBioregion: Top End CoastalIn two of the last four years, fire burnt through anisland colony of Australian Pelicans killing at least1000 young birds (=most of the population). Fireswere probably deliberately lit.
Cheal, P. D., Day, J. C., & Meredith, C. W.(1979). Fire in the national parks of north-westVictoria. National Parks Service.threatened species; mallee; heath; management; hollowsBioregion: Murray-Darling DepressionBird communities were censused in a range ofvegetation types (mallee and heath) across a rangeof regrowth ages. “After a fire in mallee vegetation,virtually no birds will breed in the burnt area inthe first three to four years”. Some birds (mainlygranivores) will forage in such burnt areas, butgenerally at low abundance. When regrowth treesbegin to emerge, bird diversity and densitiesincrease rapidly (to peak at about 15 yearspost-fire, though richness may continue toincrease). No bird species appears to be restrictedto young regrowth, but several are restricted tomallee of >15-20 years post-fire. These includehollow-nesting species (e.g. Striated Pardalote,parrots), some canopy species and manyground-foraging species. Most of the “typical”mallee species favour old vegetation. Howeververy old mallee may become unsuitable, even forthese species. Malleefowl may prefer vegetationaround 15-25 years post-fire, as food resourcesprobably decline with increasing time since fire.
In heaths, few species occur in very youngregrowth, richness probably peaks at c15-20 yearspost-fire, and older heaths tend to bespecies-poor.
Predation on birds may be high in theimmediate aftermath of fire. In general, old malleehad more stable bird assemblages (greatersimilarity in species composition betweengeographically separated sites compared toyounger regrowth), and their bird assemblagesincluded more insectivores and greaterstratification of foraging zones. The restriction ofmost mallee specialists to old vegetation (and thegeneralist nature of transient species occurring inyoung regrowth) argues for a relative stability ofold mallee and the adaptation of most mallee birdsto long intervals between fire.
Chisholm, A. H. (1922). The “lost” ParadiseParrot. Emu, 22, 4-17.Paradise Parrot; threatened species; eucalypt woodland.The Paradise Parrot may have declined (now toextinction) over the last century because grazingand frequent firing led to loss of its seed resource.
Chisholm, A. H. (1945). Birds of the Gilbertdiary, Part 2. Emu, 44, 183-200.Paradise Parrot; eucalypt woodland; threatened species.Altered fire regimes since European settlementmay have contributed to the extinction of theParadise Parrot.
Christensen, P. (1974). The concept of faunapriority areas. In Third Fire Ecology Symposium(pp. 66-73). Melbourne: Forests Commission,Victoria.eucalypt open forest; wildfireBioregion: WarrenBird abundance and richness increases sharply inKarri forest up to at least 2 years after a very hotburn. Some species of open formations (e.g. ScarletRobin) can invade wetter forests after hot fires.Other species (e.g. Red-winged Fairy-wren) declinebut then increase within 2 years.
Christensen, P., & Abbott, I. (1989). Impact offire in the eucalypt forest ecosystem ofsouthern Western Australia: a critical review.Australian Forestry, 52, 103-121.review; community; management; eucalypt open forest;successionBioregions: Jarrah Forest; Warren.Current periods between fuel-reduction burns are5-6 years for Jarrah forest and 7-9 years for Karriforest, other than in conservation reserves. Limitedevidence suggests that Aboriginal burning in theseforests may have been more frequent (3-5 yearcycles) or much less frequent. Reviews research oneffects of fire on soil nutrients, floristics, structure,invertebrates and vertebrates. Most studies reportan initial short-term decrease in understorey birdsfollowed by an increase above pre-fire levelsfollowing fuel-reduction burns in Jarrah and Karriforests, with little change in canopy species. Somespecies (e.g. White-winged Chough, WesternYellow Robin, Scarlet Robin) invaded or increasedin abundance in recently-burnt areas. Honeyeatersalso became more common in association with
128
Australia’s Biodiveristy - Responses to Fire

fire-induced flowering. Fire intensity is the majorfactor influencing the impact of fire on birdpopulations.
Christensen, P. E., & Kimber, P. C. (1975).Effect of prescribed burning on the flora andfauna of south-west Australian forests.Proceedings of the Ecological Society of Australia,7, 85-107.review; eucalypt open forest; control burning;community; successionBioregions: Jarrah Forest; WarrenBirds showed surprisingly small changes followingcool fire. Birds were sampled at two sites (oneunburnt for 40 years and one subject to an intenseprescribed fire) in dry sclerophyll forest. There waslittle difference in the bird communities before fire.From one month to one year post-fire there was avery slight decline of ground and understoreyspecies. By 2 years post-fire the population ofunderstorey and ground-dwelling species (e.g.Rufous Treecreeper, Inland Thornbill, WesternThornbill, Western Yellow Robin, GoldenWhistler) had surpassed pre-fire levels, and that ofcanopy species was unchanged. Following a fire inwet sclerophyll forest, bird numbers decreasedinitially, but by 5 months post-fire abundance washigher than pre-fire (due especially to moreWestern Yellow Robin, Inland Thornbill, ScarletRobin, Rufous Treecreeper and GreyShrike-thrush). In the second year after fire therewas an increase in birds of the shrub layer(particularly of Inland Thornbill and White-browedScrubwren), and of White-naped Honeyeater(which foraged in epicormic growth). Some species(e.g. Red-winged Fairy-wren and Golden Whistler)disappeared for the first year post-fire, but returnedin 2-3 years post-fire. White-browed Scrubwrenchanged foraging behaviour after fire. Extensive hotfires or frequent cool fires would produce ahomogenisation of the landscape to the detrimentof bird species diversity.
Christensen, P., Recher, H., & Hoare, J.(1981). Responses of open forests (drysclerophyll forests) to fire regimes. In A. M.Gill, R. H. Groves, & I. R. Noble (Eds.), Fireand the Australian biota (pp. 367-393).Canberra: Australian Academy of Science.review; eucalypt open forest; widlfire; mortalityIn very hot wildfires, bird mortality in fires may behigh. With cooler fires, mortality is usually low,however post-fire mortality may be substantial.Noisy Scrub-bird requires a late seral stage andcannot survive frequent fires. The nectarivorousCrescent Honeyeater may disappear from heathsfor several years after fire. Post-fire changes inforests are most pronounced for birds of groundand understorey layers. In general these birdsdecline in abundance after fire, but then increase toat least pre-fire levels within 2-3 years post-fireRecolonisation and increased populations may bedue to greater insect numbers on regrowthvegetation.
Christensen, P. E. S., Wardell-Johnson, G., &Kimber, P. (1985). Birds and fire insouthwestern forests. In A. Keast, H. F. Recher, H. Ford, & D. Saunders (Eds.),Birds of eucalypt forests and woodlands: ecology,conservation, management (pp. 291-299).Chipping Norton: Surrey Beatty.eucalypt open forest; management; community; controlburning; wildfire; successionBioregion: Jarrah ForestReports two studies of birds and fire in Jarrahforests. A hot prescribed fire burnt the site in thethird year of a 5 year study (a control site remainedunburnt). Bird species richness increased in theburnt site. 13 species (including White-wingedTriller, Australian Magpie, Australian Raven)appeared after fire that had not been recordedbefore, and others (e.g. Dusky Wood-swallow, TreeMartin, Scarlet Robin, Western Yellow Robin,Inland Thornbill, Western Thornbill) became moreabundant (for varying periods post-fire). Somenectarivorous species (Purple-crowned Lorikeet,Silvereye, Red Wattlebird, Western Spinebill)increased post-fire, because of fire-inducedflowering in the eucalypt overstorey.Canopy-feeding insectivores showed no or littleeffect of fire. Golden Whistler, White-breastedRobin and White-browed Scrub-wren declinedafter fire (the latter two did not reappear until2 years post-fire).
129
Fire and Australian birds

A second study considered long-term impactsof fire regimes, comparing bird assemblages in3 areas with different fire regimes (unburnt for14 years, burnt in a severe wildfire 1 year beforestudy, and burnt in a mild fire 6 months beforestudy). The unburnt and mildly burnt sites hadmore species and individuals than the site whichhad been burnt by wildfire. Golden Whistler,Inland Thornbill, Western Yellow Robin andWhite-breasted Robin were most common in theunburnt forest (and least common in the wildfiresite). Scarlet Robin was least common in thewildfire site. Western Thornbill was least commonin the unburnt site. The abundance of smallinsectivores of the canopy showed little variation.
Rapid response of birds to mild firecompared to more substantial impact of hotwildfire may be due to a delayed recovery ofinvertebrates in the latter. As a general rule, theeffect of mild or moderate fires on birds isinversely proportional to their main foragingheight. However responses of species aresomewhat idiosyncratic, and the individualresponses of rare species in particular need to beconsidered.
Fire intensity is the major factor determiningimpact of fire on birds. Season of burn may berelatively unimportant, other than through itsimpact on intensity: this view is contrary topopular belief that spring burning is especiallydetrimental due to its alleged disruption ofbreeding. A diverse burning pattern may be themost appropriate management.
Clout, M. N. (1989). Foraging behaviour ofGlossy Black-cockatoos. Australian WildlifeResearch, 16, 467-473.Glossy Black-cockatoo; Allocasuarina woodland;hollows; eucalypt open forest; threatened species;managementBioregion: South eastern CornerGlossy Black-cockatoos are reliant on seeds ofAllocasuarina. These species are fire-sensitive,requiring heat to open cones, but are easily killedby intense fire. Post-European changes in fireregime have reduced abundance of Allocasuarinaand the large dead trees in which GlossyBlack-cockatoos nest. Conservation of this speciesdepends upon appropriate fire management.
Coate, K. (1985). Black Honeyeaters feedingon ash. Western Australian Naturalist, 16,51-52.charcoal; Black HoneyeaterUp to 4 Black Honeyeaters were feedingsimultaneously in old fireplaces, consuming ash.Analysis showed this had high levels of calcium,and the behaviour may be explained by femalebirds seeking calcium before egg-laying.
Cooper, R. P. (1972). The occurrence of thePilot-bird on Wilson’s Promontory. AustralianBird Watcher, 4, 137-143.Pilot-bird; eucalypt open forest; wildfireBioregion: South east Coastal PlainFrom being common last century in denselyvegetated gullies of the Strzelcki Ranges, thePilot-bird has now been largely wiped out of thearea by clearing and wildfire, particularly thedisastrous fires of 1939.
Cooper, R. P. (1974). The avifauna of Wilson’sPromontory. Part 1. Australian Bird Watcher,5, 137-174.heath; eucalypt open forest; managementBioregion: South east Coastal PlainSubstantial changes to vegetation (increasedscrubbiness) of Wilson’s Promontory over the lastcentury were due to changes in fire regime. Thismust have changed bird species composition. Avery large wildfire in 1951 (during the breedingseason) probably resulted in massive mortality ofbirds. Argues against fuel reduction burning andburning of heathlands. “Fire has a mostdetrimental effect on the avifauna of any area.”
Cooper, R. P. (1975). The avifauna of Wilson’sPromontory. Part 4. Australian Bird-watcher, 6, 17-34.heath; Ground Parrot; threatened speciesBioregion: South east Coastal PlainGround Parrots reported to flee just ahead of firefront. The species disappeared from an area for atleast four years post-fire.
130
Australia’s Biodiveristy - Responses to Fire

Cowley, R. D. (1971). Birds and forestmanagement. Australian Forestry, 35, 234-250.review; eucalypt open forest; management; forestryIn eucalypt forests, birds which feed or nest on theground (e.g. Superb Fairy-wren, White-browedScrubwren, Spotted Quail-thrush, BrownThornbill and White-eared Honeyeater) may bethose most affected by fire. Birds which nest within1.5m of the ground may be vulnerable tofuel-reduction fire during the breeding season.Food (e.g. seed) is generally more available after afire. Cool burns generally leave unburnt patches,typically in gullies, and regrowth of grasses andshrubs is usually rapid after fire. Frequent fires canchange shrubby understorey to grass, to theadvantage of species such as Spotted Quail-thrushand Buff-rumped Thornbill but to the disadvantageof species associated with shrubs (e.g.White-browed Scrubwren, Brown Thornbill,Common Bronzewing). Frequent fires in wetterforest may result in a dense cover of wire grass, tothe detriment of species foraging in litter (such asSuperb Lyrebird, Bassian Thrush, Eastern YellowRobin, Superb Fairy-wren). Some honeyeaters areadvantaged by frequent light fires, because thesepromote profuse flowering.
Cowley, R. D. (1974). Effects of prescribedburning on birds of the mixed species forestsof West Central Victoria. In Third Fire EcologySymposium (pp. 58-65). Melbourne: ForestsCommission, Victoria.eucalypt open forest; control burning; communityBioregion: Victorian MidlandsShort-term effects of a (fairly hot) prescribed burnin open forest were examined, partly throughobservation of changed distribution of bandedbirds. Of 27 banded birds (of 7 species:White-browed Scrubwren, Superb Fairy-wren,Straited Thornbill, Brown Thornbill,White-throated Treecreeper, Eastern YellowRobin, White-eared Honeyeater) regularly usingthe site pre-fire, 18 were known to have survived(all species other than White-eared Honeyeater).Home ranges changed little. Birds appeared morewary after fire (and logs were used as refuge).Several species with nests started before the firesuccessfully raised young after the fire. Only onespecies (Olive Whistler, which prefers denseunderstorey) apparently disappeared. Severalspecies characteristic of open areas (AustralianMagpie, Buff-rumped Thornbill, Painted Quail)invaded after fire.
Cowley, R. D., Heislers, A., & Ealey, E. H. H.(1969). Effects of fire on wildlife. Victoria’sResources, 11, 18-22.mallee; Malleefowl; Superb Lyrebird; eucalypt openforest; reviewMalleefowl require abundant litter for nesting.This may not be available for at least 10 yearspost-fire. In contrast, Superb Lyrebird mayrequire periodic fires to remove wire grass.
Crawford, D. N. (1972). Birds of Darwin area,with some records from other parts ofNorthern Territory. Emu, 72, 131-148.tropical eucalypt open forest; tussock grassland;hummock grassland.Bioregions: Top End Coastal; Pine Creek ArnhemBurning of grasslands during the dry season leadsto local shifts in habitat use by Golden-headedCisticola and Red-backed Fairy-wren (both mayoccur in monsoon rainforest fringes when most ofthe open forests and grasslands have been burnt).Barn Owl concentrates in cleared areas early inthe dry but disperses to extensive burnt areas laterin the season (presumably because hunting overunburnt grasslands is difficult). In contrast, theredoes not appear to be a concentration of PheasantCoucals in unburnt patches of grassland later inthe season (possibly suggesting seasonalmovement). White-throated Grass-wren recordedonly in long-unburnt hummock grassland.
Crawford, D. N. (1979). Effects of grass andfires on birds in the Darwin area, NorthernTerritory. Emu, 79, 150-152.tropical eucalypt open forest.Bioregion: Top End CoastalMonitored bird populations in 24 sites (1.6ha) inwet and dry seasons over 28 months. All sites wereburnt at some stage over the study period.Red-backed Fairy-wren was disadvantaged by fire,and its persistence appears to depend on somecover remaining after fire. Within 2 weeks ofburning, there is an increase in migratory ground-and mid-level foragers, and Pied Butcherbird,Black-faced Cuckoo-shrike, Magpie-lark andRed-tailed Black-cockatoo. Their numbersdeclined slightly at >2 months post-fire.
131
Fire and Australian birds

Crowley, G. M. (1995). Fire on Cape YorkPeninsula. Cape York Peninsula Land UseStudy.review; tropical eucalypt open forest; rainforest;hollows; Cassowary; Golden-shouldered Parrot;threatened species; managementBioregion: Cape York PeninsulaBirds that are advantaged by fire in theshort-term, or by a high frequency regime, tendto be nomadic granivores, carnivores oromnivores. Of particular concern is the effect offires on tree hollows used for nesting by birds.Fires may lead to hollow formation, or todestruction of hollows and nests. Ecotonebetween rainforest and wet eucalypt forest may bethe preferred habitat for Cassowary, and thisspecies may require fire for its maintenance.Widespread late dry season fires have probably ledto the decline of the Brown Treecreeper. Lack ofburning after the first storms of the wet season isconsidered a threat to the Golden-shoulderedParrot. A wide variety of burning histories is likelyto maximise bird diversity.
CSIRO Wildlife Research (1976). A survey ofthe fauna of the Lower McArthur River Region,Northern Territory. Mimets Development PtyLtd.tropical eucalypt open forest; tropical eucalypt savannawoodland; hummock grassland; tussock grassland;Carpentarian Grass-wren; Bush Stone-curlew;Australian Bustard.Bioregions: Gulf Falls and Uplands; Gulf CoastalIncreased frequency of fire, particularly inhummock grasslands, must have a deleteriouseffect on the survival of such terrestrial species asCarpentarian Grass-wren, quail, BushStone-curlew and perhaps Bustard.
Cummings, B., McDonald, B., & Taplin, A.(1993). Knowledge of birds in Queenslandbiogeographic regions and threats to theirconservation. In C. P. Catterall, P. V. Driscoll,K. Hulsman, D. Muir, & A. Taplin (Eds.), Birdsand their habitats: status and conservation inQueensland (pp. 178-186). St Lucia:Queensland Ornithological Society Inc.reviewBioregions: Mitchell Grass Downs, South EastQueensland, Einasleigh Uplands, Cape YorkPeninsula, Wet Tropics, Channel Country, MulgaLands, Brigalow Belt, Mount Isa Inlier, GulfPlains, Desert Uplands, Central Mackay Coast.
The relative importance of fire as a factoraffecting bird conservation is discussed for everyQueensland bioregion. Existing fire regimes maybe a management problem for birds in MitchellGrass Downs, South East Queensland, EinasleighUplands, Cape York Peninsula, Wet Tropics, butare not a problem (or there is insufficientinformation) for other bioregions.
Curry, G. N. (1991). The influence ofproximity to plantation edge on diversity andabundance of bird species in an exotic pineplantation in north-eastern New South Wales.Wildlife Research, 18, 299-314.management; slash-burn; forestryWindrows in plantations allow the entry orpersistence of many bird species. These areusually burnt to reduce fire risk to the plantation,but it would be preferable (for bird conservation)not to burn these windrows, but rather to reducefire hazard by careful positioning of windrows.
Curry, P. J. (1986). Habitat characteristics ofthe Thick-billed Grasswren Amytornis textilisin grazed shrublands in Western Australia. InP. J. Joss, P. W. Lynch, & O. B. Williams(Eds.), Rangelands: a resource under siege (pp.566). Canberra: Australian Academy ofScience.Thick-billed Grasswren; chenopod shrubland; AcaciashrublandBioregion: CarnarvonThe western subspecies of Thick-billedGrasswren had previously been considered to bepossibly on the verge of extinction and reliant onsaltbush and bluebush. Instead, the subspecies wasfound to be widespread in seral shrublands thatreplace Acacia shrublands for at least 40 yearsafter wildfire.
Danks, A. (1991). The role of corridors in themanagement of an endangered passerine. InD. A. Saunders & R. J. Hobbs (Eds.), Natureconservation 2: the role of corridors (pp.291-296). Chipping Norton: Surrey Beatty.Noisy Scrub-bird; eucalypt open forest; thicket;corridors; management; threatened species.Bioregion: Jarrah ForestDispersal of Noisy Scrub-bird is assisted bycorridors which are unburnt (and protected fromother disturbance).
132
Australia’s Biodiveristy - Responses to Fire

Davidson, I., & Chambers, L. (1991).Vegetation management for Superb Parrotforaging habitat in Victoria. VictorianDepartment of Conservation andEnvironment.Superb Parrot; threatened species; managementBioregion: RiverinaSome corridors used by Superb Parrots may beenhanced by the cessation of burning; landownersmay be reluctant to increase or maintain suitablehabitat patches because of their perception thatthis increases the risk of wildfire.
Davidson, I., & Robinson, D. (1992).Grey-crowned Babbler Pomatostomus temporalis.Department of Conservation and NaturalResources.Grey-crowned Babbler; eucalypt woodland; threatenedspecies; management; control burningBioregions: Victorian Midlands; South EasternHighlandsReviews information on this species. Threats todeclining Grey-crowned Babblers include fireprotection works (e.g. firebreaks and fuelreduction burning) and collection of firewood.
Davies, S. J. J. F., Smith, G. T., & Robinson, F.N. (1982). The Noisy Scrubbird in WesternAustralia. In R. H. Groves & W. D. L. Ride(Eds.), Species at risk: research in Australia (pp.117-127). Canberra: Australian Academy ofScience.Noisy Scrub-bird; threatened species; heath; eucalyptopen forest; management.Bioregion: Jarrah ForestThe Noisy Scrub-bird disappeared from areaswhere eucalypt forests fringing swamps had beenburnt. With strict fire control populations havebuilt up.
Debus, S. J. S., & Czechura, G. V. (1988). TheRed Goshawk Erythrotriorchis radiatus: areview. Australian Bird Watcher, 12, 175-199.Red Goshawk; historical change; threatened species;tropical eucalypt open forestSince the Miocene, Australia’s rich raptorcommunity has declined, possibly because ofclimate change and perhaps Aboriginal fireregimes. Currently, the most serious threats to theRed Goshawk are mismanagement of tropicalrangelands (overstocking and too frequentburning), vegetation destruction and wetlanddrainage.
Dedman, V. (1983a). G.F.N.C. OtwayRegeneration Survey - Progress report.Geelong Naturalist, 20, 52-56.eucalypt open forest; heath; wildfireBioregion: South east Coastal PlainBy six months after hot fire in heath and eucalyptopen forest, birds were returning slowly. Largerspecies (Australian Raven, Currawongs, LaughingKookaburra, Sulphur-crested Cockatoo) areconspicuous. Smaller birds were not seenregularly until bracken was high enough toprovide cover, but by 6 months post-fire Brownand Striated Thornbills were being seen regularly.Welcome Swallows were common over the burntheathland.
Dedman, V. (1983b). G.F.N.C. Otwaysregeneration survey - 2nd progress report.Geelong Naturalist, 20, 98-100.eucalypt open forest; wildfireBioregion: South east Coastal PlainAlmost one year after hot fire, bird numbers andspecies were increasing. Honeyeaters wereattracted to flowering Xanthorrhoea. SuperbFairy-wrens were foraging in regrowth bracken.Some species were probably breeding. PiedCurrawongs, Sulphur-crested Cockatoos and RedWattlebirds were noted frequently.
Dedman, V. (1983c). Mammals and birds afterthe fires: Forest Road, Anglesea. GeelongNaturalist, 20, 27.eucalypt open forest; wildfireBioregion: South east Coastal PlainList of 13 bird species observed in severely burnteucalypt open forest two weeks after fire.
Dedman, V. (1983d). Moggs Creek: eight dayslater. Geelong Naturalist, 20, 22-25.eucalypt open forest; wildfireBioregion: South east Coastal PlainEight days following severe fire in eucalypt openforest, few birds were seen. These includedCrimson Rosella, Red Wattlebird,White-throated Treecreeper and Scarlet Robin.
133
Fire and Australian birds

Dedman, V. (1983e). Notes on the effects ofthe Ash Wednesday fire on the IronbarkBasin, Point Addis. Geelong Naturalist, 20,25-26.eucalypt open forest; wildfireBioregion: South east Coastal PlainBird lists from 3 weeks and 6 weeks after hot firein eucalypt open forest. Superb Fairy-wrens wereobserved in scorched tree-tops. Commonest birdsin the burnt area were Eastern Yellow Robin andWhite-throated Treecreeper. Other speciespresent included White-browed Scrubwren,Crimson Rosella, Brown Thornbill and StriatedThornbill.
Dedman, V. (1984). Otways regenerationsurvey. Third progress report. GeelongNaturalist, 21, 89-92.wildfire; eucalypt open forestBioregion: South east Coastal PlainOne year after a hot wildfire, large birds(Yellow-tailed Black-cockatoo, Crimson Rosella,Red Wattlebird, Pied Currawong) appear moreobvious in burnt forest.
Dickinson, K. J. M., Wall, L. E., & Wilson, R.I. (1986). Birds in a partly clearfelled dryeucalypt forest on dolerite in southeasternTasmania. Papers and Proceedings of the RoyalSociety of Tasmania, 120, 39-49.eucalypt open forest; slash-burn; forestry; community;managementBioregion: FreycinetBird assemblages were sampled in unloggedforest, forest clearfelled and slash burnt, andforests clearfelled without subsequent burning.Species typical of open areas (e.g. Blue-wingedParrot, Superb Fairy-wren) invaded the cut andburnt sites. Flame Robin, Dusky Robin, SuperbFairy-wren and Grey Shrike-thrush were morecommon in the slash-burnt area than the unburntlogged forest: Crescent Honeyeater and EasternSpinebill were more common in the unburnt cutforest. Prescribed burning under regeneratingforests may disadvantage some species whichdepend on ground habitats (e.g. SpottedQuail-thrush).
Disney, H. J. d. S. (1968). Bushfires and theireffect on fauna and flora. Australian NaturalHistory, 16, 87-89.reviewBanded birds returned to their pre-burnterritories following fire. Backburns may beparticularly dangerous to small birds.
Du Guesclin, P., Smith, S., O’Shea, B., &Debbis, C. (1995). “Brushing for bristles”:habitat corridors for the Rufous Bristlebird. InA. Bennett, G. Backhouse, & T. Clark (Eds.),People and nature conservation: perspectives onprivate land use and endangered species recovery(pp. 163-165). Chipping Norton: SurreyBeatty.Rufous Bristlebird; heath; corridors; isolateBioregion: South East Coastal PlainRecolonisation after fire is a problem for RufousBristlebirds because of their poor dispersal ability.Corridors may provide effective means forrecolonisation.
Emison, W. B., & Bren, W. M. (1989).Common birds of the mallee, northwesternVictoria. In J. C. Noble & R. A. Bradstock(Eds.), Mediterranean landscapes in Australia:mallee ecosystems and their management (pp.221-242). Melbourne: CSIRO.mallee; heath; eucalypt woodland; Callitris woodland;Casuarina woodland; hollows; managementBioregion: Murray-Darling DepressionMany species are dependent on hollows inwoodlands, and the maintenance of these requiresactive management (including of fire regimes). Inthe previous decade, wildfires have burnt much ofthe mallee shrublands such that long-unburntpatches are now very rare. Species associated withsuch habitat include those nesting in hollows, andthose which require dense litter. The managementof mallee for fauna probably requires maintenanceof a mosaic of vegetation of a range of ages, butwith a bias towards retention of older ages.
134
Australia’s Biodiveristy - Responses to Fire

Emison, W. B., Beardsell, C. M., Norman, F.I., & Loyn, R. H. (1987). Atlas of Victorianbirds. Melbourne: Department ofConservation, Forests and Lands, and RoyalAustralasian Ornithologists Union.reviewDescribes habitat and distribution for Victorianbirds. The Mallee Emu-wren attains highestdensities in early years after fire. The SouthernEmu-wren is “able temporarily to coloniseephemeral habitats such as … scrubs regeneratingafter fire.”
Ferrier, S. (1985). Habitat requirements of arare species, the Rufous Scrub-bird. In A.Keast, H. F. Recher, H. Ford, & D. Saunders(Eds.), Birds of eucalypt forests and woodlands:ecology, conservation, management (pp.241-248). Chipping Norton: Surrey Beatty.Rufous Scrub-bird; management; eucalypt open forest;rainforest; threatened species.Bioregion: NSW North CoastFire has played a major role in the RufousScrub-bird’s current distribution and, inparticular, its close association with rainforest.High intensity wildfires are known to havedestroyed territories. Prescribed burning probablyhas little direct effect (though habitat suitabilitywas low 10 months after a prescribed fire), andmay prevent destruction through wildfire (thoughmost are naturally buffered by their dampness orproximity to rainforests). However long-terminfluence of prescribed burning may be complex.Suitable habitat in eucalypt forests is probablyassociated with a particular post-fire seral stage.With long absence of fire, the eucalypt forestsmay develop into rainforests, to the detriment ofRufous Scrub-birds (due to reduction in groundcover density).
Fitzherbert, J. C., & Baker-Gabb, D. J. (1988).Australasian grasslands and their threatenedavifauna. In P. D. Goriup (Ed.), Ecology andconservation of grassland birds (pp. 227-250).Cambridge: International Council for BirdPreservation.review; tussock grasslands; hummock grasslands;tropical eucalypt savanna woodland; management;threatened species; Gouldian Finch; Yellow-rumpedMannikin; Chestnut-backed Button-quail; PartridgePigeon; Golden-shouldered Parrot; Hooded Parrot;Carpentarian Grass-wren; White-throatedGrass-wren; Black Grass-wrenCarpentarian, White-throated and BlackGrass-wrens are threatened by fire in hummockgrasslands, and require aging (>6 years) Triodia.In tropical grasslands, fire frequency has increasedsince European settlement. “Large areas are burnteach year … The decline of six bird species - theGouldian Finch, Yellow-rumped Mannikin,Chestnut-backed Button-quail, Partridge Pigeon,Golden-shouldered Parrot and Hooded Parrot - isprobably related to the changed fire regime andassociated grazing pressure.” After habitatdestruction (by over-grazing and conversion ofgrasslands to agriculture) “fire is undoubtedly thenext most significant threat”. In tropical savannas,fire effects are poorly understood, but changedphenology or floristics have probably affectedsome birds. In some central and southerngrasslands, a decrease in fire frequency (butincrease in intensity) “has had an equallydestructive effect”.
Fitzherbert, K., McLaughlin, J., &Baker-Gabb, D. (1992). Black-eared MinerManorina melanotis. Department ofConservation and Environment.Black-eared Miner; threatened species; mallee;management; wildfire.Bioregion: Murray-Darling DepressionReviews information on the species. Black-earedMiners inhabit areas that have not been burnt formore than 60 years, a limited and declininghabitat. Suppression of wildfires and reduction infire frequency will assist this species.
135
Fire and Australian birds

Ford, H. A. (1989). Ecology of birds: anAustralian perspective. Chipping Norton:Surrey Beatty.review; eucalypt open forest; Ground Parrot; heathIncludes a brief review of the effect of fires ineucalypt forest. Changes are not dramatic and,except after severe or extensive fire, the birdassemblage will have recovered its originalcomposition within 1-2 years post-fire. GroundParrot cannot survive in habitat where fires occurat intervals of less than every few years or greaterthan 20 years.
Forshaw, J. M. (1981). Australian Parrots.Melbourne: Lansdowne Press.review; Golden-shouldered Parrot; Ground Parrot;Scarlet-chested Parrot; tropical eucalypt open forest;heath; mallee; hummock grassland; threatened speciesFire regime is contributing to the decline ofGolden-shouldered Parrot. The Ground Parrot isdependent on a regime of mosaic burning at 8-10year intervals. Scarlet-chested Parrot is associatedwith recently-burnt mallee with hummock grass.
Fowler, S. (1945). The fire on MondrainIsland. Emu, 44, 334-335.seabirds; tussock grassland; heath; mortalityBioregion: Esperance PlainsDeliberately lit fire burnt 60% of MondrainIsland, leading to seabird deaths.
Fox, A. (1978). The ’72 fire of Nadgee NatureReserve. Parks & Wildlife, 2, 5-24.heath; eucalypt open forest; mortality; wildfireBioregion: South East CornerAfter fire, 609 birds were found dead on beach.Most (89%) came from a backburn area ratherthan from the original fire. 49 bird species wererecorded as killed: the most common dead birdswere Little Wattlebird and New HollandHoneyeater.
Frith, H. J. (1962). Conservation of theMallee-fowl (Leipoa ocellata, Gould). CSIROWildlife Research, 7, 33-49.Malleefowl; threatened species; management; mallee.Bioregion: Murray Darling DepressionMalleefowl requires adequate litter for breeding.This is not available until at least 10-15 yearspost-fire.
Garnett, S. (1987). Seabird Islands: No. 171.Kusamet Island, Torres Strait, Queensland.Corella, 11, 77-78.seabirds; tussock grassland; thicket; mortalityBioregion: Cape York PeninsulaThe grass on the island is often burnt duringOctober and November which is likely to destroynests and chicks of the breeding Bridled andBlack-naped Terns.
Garnett, S. (1992a). The action plan forAustralian birds. Canberra: AustralianNational Parks and Wildlife Service.review; threatened species; managementDescribes the former and current threats to allthreatened Australian birds, and research andmanagement requirements. Altered fire regimes arecurrently threatening 51 taxa (16 confirmed, 35speculative). Only one other threatening process isaffecting more taxa (habitat clearance andfragmentation which affects 54 taxa).
Garnett, S. (1992b). Threatened and extinctbirds of Australia. Melbourne: RoyalAustralasian Ornithologists Union.review; threatened species; managementReviews (including previously unpublishedinformation) the status of Australian threatenedbirds. Altered fire regimes are a current confirmedor speculated threat to 51 Australian bird taxa,second only to clearing and fragmentation(affecting 52 taxa). The most serious fire effectsare increase in frequency of late dry season fires intropical woodlands and all fires in the mallee.Discusses the threat of fire (or relationship withfire) for Kangaroo Island Emu, AustralasianBittern (reports desertion of a swamp for 2 yearsafter burning), Red Goshawk, MalleeFowl(habitat should be protected against too frequentburning), Buff-breasted Button-quail (where latedry season fires during the nesting season areregarded as the major threat), Chestnut-backedButton-quail, Painted Button-quail,Black-breasted Button-quail (control burns onrainforest margins at intervals of less than 4 yearsrender the habitat unsuitable), Lewin’s Rail,Partridge Pigeon (early dry season fires maydestroy the ground nests and eggs), Red-cheekedParrot (main threat is fire in the late dry seasonwhich may destroy nest trees), Eclectus Parrot(nests have been destroyed by management fires),Golden-shouldered Parrot, Partridge Pigeon,
136
Australia’s Biodiveristy - Responses to Fire

Orange-bellied Parrot, Scarlet-chested Parrot,Ground Parrot (fire continues to be the mainthreat), Night Parrot, Palm Cockatoo (the hollowtrees used for nesting are particularly vulnerableto fire, and many have been destroyed by recentfires, including destruction of eggs and young),Red-tailed Black-cockatoo, Masked Owl, SootyOwl, Rufous Owl (the hollow trees used fornesting are particularly vulnerable to fire), RufousScrub-bird, Noisy Scrub-bird, Purple-crownedFairy-wren, Southern Emu-wren (wherefragmentation exacerbates the threat of fire due todecreased possibility of recolonisation), MalleeEmu-wren, Striated Grass-wren (the major threatis extensive fire), Carpentarian Grass-wren,Black-eared Miner, Helmeted Honeyeater,Forty-spotted Pardalote, Rufous Bristlebird,Eastern Bristlebird (the main threat to the speciesappears to be change in fire regime), WesternBristlebird (fire is the main threat), BrownThornbill, Slender-billed Thornbill, NorthernScrub-robin (possible extinction of the NTsubspecies has been attributed to changed fireregime), Western Whipbird (fire appears to be themain threat), Crested Shrike-tit, Red-loredWhistler (in remnant habitat the effects of firemay be critical), Bassian Thrush and ZittingCisticola.
Garnett, S., & Bredl, R. (1985). Birds in thevicinity of Edward River Settlement. Part II.Discussion, references, list of passerines.Sunbird, 15, 25-40.tropical eucalypt open forest; tussock grassland; historicchange; Star Finch; community; threatened speciesBioregion: Cape York Peninsula“Hunting, introduction of plants and animals, andburning are the effects of people most likely to bedetrimental to the birds of the Edward Riverarea.” Grass around the crocodile breeding lagoonis never burnt, and Star Finch is now restricted tothis area, suggesting that the prevailing fireregime in the region has become unsuitable forthis species.
Garnett, S. T., & Crowley, G. M. (1994). Theecology and conservation of theGolden-shouldered Parrot. Cape York PeninsulaLand Use Strategy.Golden-shouldered Parrot; threatened species;management; tropical eucalypt savanna woodlands;tussock grasslands; historic change; control burning
Bioregion: Cape York PeninsulaFire regimes determine boundaries (and relativeextent) of grassland flats and Melaleucawoodlands. Use of fire to control Melaleuca andearly wet season burning (to increase foodresources) are required to conserveGolden-shouldered Parrots.
Garnett, S., & Crowley, G. (1995a). Thedecline of the Black Treecreeper Climacterispicumnus melanota on Cape York Peninsula.Emu, 95, 66-68.historical change; tropical eucalypt open forest; BlackTreecreeper; managementBioregion: Cape York PeninsulaMarked decline in Black Treecreeper on CapeYork Peninsula over the period 1920 to 1995.Principal reason thought to be change in fireregime, with larger hotter fires now. Remnantpopulations are now mainly in areas with naturalfire breaks (rivers etc). Conservation of the speciesdepends upon reintroduction of fine-grainedmosaic burning.
Garnett, S., & Crowley, G. (1995b). Feedingecology of Hooded Parrots Psephotus dissimilisduring the early wet season. Emu, 95, 54-61.Hooded Parrot; tropical eucalypt open forest; tropicaleucalypt savanna woodlandBioregion: Pine Creek ArnhemDetailed foraging study of Hooded Parrots. Fireand grazing may affect plant species compositionand hence food availability for this species.
Garnett, S., Crowley, G., Duncan, R., Baker,N., & Doherty, P. (1993). Notes on live NightParrot sightings in north-westernQueensland. Emu, 93, 292-296.Night Parrot; tussock grassland; hummock grassland;Spinifex-bird; Rufous-crowned Emu-wren; threatenedspeciesBioregion: Mt Isa InlierNight Parrot may be present in this area due tofavourable fire management (small control burnsleading to seral mosaic and preventing extensivewildfire). Rufous-crowned Emu-wrens andSpinifex-birds are also favoured by thismanagement, as both prefer spinifex which hasremained unburnt for long periods.
137
Fire and Australian birds

Gibbons, P. (1994). Sustaining key old-growthcharacteristics in native forests used for woodproduction: retention of trees with hollows. InT. W. Norton & S. R. Dovers (Eds.), Ecologyand sustainability of southern temperateecosystems (pp. 59-84). Canberra: CSIRO.forestry; old-growth; hollows; management; review;eucalypt open forestReviews use of hollows by birds (and other fauna);hollow-formation; relationship between fire (andforestry operations) and hollow formation.
Gibson, D. F. (1986). A biological survey of theTanami Desert in the Northern Territory.Conservation Commission of the NorthernTerritory.hummock grassland; Little Button-quail; Richard’sPipit; Rufous-crowned Emu-wrenBioregion: TanamiLittle Button-quail and Richard’s Pipit occurmostly in recently burnt country. Rufous-crownedEmu-wren is particularly common inlong-unburnt hummock grass.
Gill, A. M. (1990). Fire management of malleelands for species conservation. In J. C. Noble,P. J. Joss, & G. K. Jones (Eds.), The malleelands: a conservation perspective (pp. 202-205).Melbourne: CSIRO.malleeBioregion: Murray Darling DepressionDescribes characteristics of mallee fires. Speciessuch as Malleefowl and Mallee Emu-wren arethreatened by high frequency of fires.
Gill, A. M. (1996). How fires affectbiodiversity. In DEST (Ed.), Fire andbiodiversity: the effects and effectiveness of firemanagement. Proceedings of the conference held8-9 October 1994, Footscray, Melbourne (pp.47-55 (&123-124)). Canberra: Department ofthe Environment, Sports and Territories.Ground Parrot; Mistletoebird; threatened species;heath; eucalypt open forestDescription of some impacts upon biodiversity ofa range of fire regimes. Hot fires in open forestsmay eliminate mistletoe (and hence lead to declinein mistletoebirds). Briefly reviews Queensland andVictorian studies of Ground Parrot response tofire, noting that associations with vegetation ageof graminoid heaths seem to be related to
different responses of shrubs and sedges to fire.Differences between Queensland and Victoria inpreferred ages may be due to faster processes inthe north. Hence, it is misleading to extrapolateresults to different areas or habitats.
Green, R. H. (1982). The activity andmovement of fauna in compartment 2, MaggsMountain, Tasmania, in the first five years offorest regeneration. Records of the QueenVictoria Museum, 75, 1-31.forestry; eucalypt open forest; succession; controlburningBioregion: Ben LomondMainly concerned with recovery of birdpopulations after intensive logging, but considersinteraction with burning.
Green, R. H., & McGarvie, A. M. (1971). Thebirds of King Island. Records of the QueenVictoria Museum, 40, 1-42.historic change; wildfire; Glossy Black-Cockatoo;Forty-spotted Pardalote; eucalypt open forest;Casuarina woodland; threatened speciesBioregion: WoolnorthClearing for agriculture and wildfire (especiallymajor fires around 1920) led to the extinction onKing Island of Glossy Black-Cockatoo andForty-spotted Pardalote, as well as decline inother species.
Hadlington, P., & Hoschke, F. (1959).Observations on the ecology of the phasmatidCtenomorphodes tessulata (Gray). Proceedings ofthe Linnaean Society of New South Wales, 84,146-159.eucalypt open forest; managementBioregion: South Eastern HighlandsWhile some fire regimes may decrease theabundance of phasmatids, others may lead toincreases, presumably because they decrease thenumber of egg parasites or the abundance (orpredation pressure) of predatory birds.
Harrington, G. N., & Sanderson, K. D. (1994).Recent contraction of wet sclerophyll forest inthe wet tropics of Queensland due to invasionby rainforest. Pacific Conservation Biology, 1,319-327.tropical eucalypt open forest; management; rainforest.Bioregion: Wet Tropics
138
Australia’s Biodiveristy - Responses to Fire

Fire suppression has led to contraction of ecotonalwet sclerophyll forests at the expense of expandingrainforest. The eucalypt forests are important forisolated populations of Eastern Yellow Robin,Yellow Thornbill, Buff-rumped Thornbill,Crested Shrike-tit, White-naped Honeyeater andYellow-faced Honeyeater, and these populationsmay require the establishment of a more frequentburning regime.
Haynes, C. D. (1985). The pattern andecology of munwag: traditional Aboriginal fireregimes in north-central Arnhemland.Proceedings of the Ecological Society of Australia,13, 203-214.Hooded Parrot; tropical eucalypt open forestBioregions: Top End Coastal, Pine CreekArnhem.Changes from Aboriginal fire regimes may beimplicated in the decline of Hooded Parrot andsome grass finches.
Hemsley, J. H. (1967). Bushfire - S.E.Tasmania 7th February, 1967. Some aspects ofthe fire in relation to animal and plant life.Tasmanian Forestry Commission.wildfire; mortalityBioregions: Freycinet, D’Entrecasteaux,Tasmanian Midlands931 birds of 60 species were found dead onbeaches adjacent to areas burnt by wildfire. 25species were recorded from burnt area during thefirst two months after the fire.
Hewish, M. (1983). The effect of a wildfire onbirdlife in a eucalypt forest: a preliminaryreport on the Lerdederg Gorge seven weeksafter the Wombat State Forest fire. GeelongNaturalist, 20, 3-16.eucalypt open forest; wildfire; community; successionBioregion: Victorian MidlandsCensuses in open forest before, 3 weeks and 7 weeksafter severe bushfire, including some census pointsthat were unburnt and others that were burnt onlymildly. The fire caused an immediate reduction inbird species diversity (25-26 species pre-fire fallingto 19-20 species post-fire). Unburnt sitesmaintained species richness. Mildly burnt sites wereintermediate in richness. Some species (EasternYellow Robin, Grey Shrike-thrush, SuperbFairy-wren, White-browed Scrubwren,
White-throated Treecreeper) were more common(or more obvious) in burnt areas. Four of thesespecies are insectivorous ground-feeders.Recolonisation of burnt areas occurred very quickly,even before the smoke had cleared. Previouslycommon species which were rarer or absent inburnt areas included Golden Whistler, RufousWhistler, Grey Fantail, Yellow-faced Honeyeater,Yellow-tufted Honeyeater, Spotted Pardalote andStriated Pardalote, though Rufous Whistler andYelow-faced Honeyeater may have migrated fromthe area. Most of these species forage in shrubs orcanopy, most of which was destroyed in the fire.
Hodgson, A., & Heislers, A. (1972). Someaspects of the role of forest fire in South-easternAustralia. Forests Commission, Victoria.eucalypt open forest; reviewFew birds are killed directly in fuel reduction fires.Lyrebirds recolonised forest within 2 years of anextensive wildfire. Frequent burning of forests mayfavour species which forage in open ground (e.g.Spotted Quail-thrush, Buff-rumped Thornbill) butdisadvantage those using shrubs (e.g. White-browedScrubwren, Brown Thornbill) or eating Acaciaseeds (Common Bronzewing).
Holmes, G. (1988). Eastern Bristlebird:summary conservation statement for northernpopulations. QNPWS.Eastern Bristlebird; threatened speciesBioregion: NSW North CoastInappropriate fire regimes threaten EasternBristlebird.
Holmes, G. (1989). Eastern Bristlebird. Speciesmanagement plan for northern populations.Queensland NPWS and NSW NPWS.Eastern Bristlebird; rainforest; eucalypt open forest;threatened species; management.Bioregions: South Eastern Queensland; NSWNorth Coast.Eastern Bristlebird in northern NSW andsoutheastern Queensland uses ecotones betweenrainforest and open forest. Suitability isdetermined by time since fire. Fire management isrequired.
139
Fire and Australian birds

Hood, J. B. (1941). Birds and bushfires. SouthAustralian Ornithologist, 15, 125-127.mortality; wildfire.Bioregion: Naracoorte Coastal PlainEffects of wildfire on birds depend on weatherconditions. In relatively mild fires, some species(e.g. Fork-tailed Swift, Australian Raven) areattracted to fire fronts to feed on disturbed ordead insects. In very hot fires (such as theextensive wildfire of 1939), few birds wereobserved to survive. Species noted to have beenkilled included Emu, Stubble Quail, MaskedLapwing, Bush Stone-curlew, Sulphur-crestedCockatoo, Long-billed Corella, Eastern Rosella,Crimson Rosella, Red-rumped Parrot,Wedge-tailed Eagle, Australian Magpie, ravensand Laughing Kookaburra. “As the terrific blast offire swept along, birds could be seen rising fromthe ground or leaving the trees in front of it onlyto be overpowered by the dense smoke and risingscorching heat. In all cases the birds fell exhaustedinto the flames”.
Hopkins, A. J. M. (1985). Planning the use offire on conservation lands in south-westernAustralia. In J. R. Ford (Ed.), Fire ecology andmanagement in Western Australian ecosystems(pp. 203-208). Perth: Western AustralianInstitute of Technology.review; Noisy Scrub-bird; management; WesternWhipbird; Western Bristlebird; heath; thicket; eucalyptopen forest; threatened speciesBioregion: Jarrah ForestOutlines a fire management plan for Two PeoplesBay Nature Reserve, with major aims to conserveNoisy Scrub-bird, Western Whipbird andWestern Bristlebird
Hopkins, A. J. M., & Smith, G. T. (1996). Fire:effects and management implications. In A. J.M. Hopkins & G. T. Smith (Eds.), The naturalhistory of Two Peoples Bay Nature ReservePerth: Department of Conservation and LandManagement (CALMScience Supplement).heath; threatened species; thicket; eucalypt open forest;successionBioregion: Jarrah ForestFollowing fire in dense closed heath, Richard’sPipit initially colonised (to maximum density at 2years post-fire) then declined to absence at 6 yearspost-fire. Striated Fieldwren increased post-fire toreplace Pipits. As the regrowth heath proceededfrom open to closed the Heath-wren was replaced
by Western Bristlebird (this change may bedelayed by macropod grazing). Three congenerichoneyeaters showed contrasting post-fireresponses. Tawny-crowned Honeyeater wasrecorded 1 year post-fire and peaked at 3 yearspost-fire then declined to reach the unburnt heathabundance at 6 years post-fire. White-cheekedHoneyeater was first recorded 3 years post-fire,and remained at low levels for the next 3 years.New Holland Honeyaeter was not recorded until6 years post-fire.
How, R. A., Dell, J., & Humphreys, W. F.(1987). The ground vertebrate fauna of coastalareas between Busselton and Albany, WesternAustralia. Records of the Western AustralianMuseum, 13, 553-574.heath; historic change; eucalypt open forest; Malleefowl;Western Whipbird; Rufous Bristlebird; NoisyScrub-bird; threatened speciesBioregion: WarrenDecline over this century in several birds in theWarren district was due to changes in fire regime,particularly affecting heath. Species whichdeclined as a result included Malleefowl, WesternWhipbird, Rufous Bristlebird and NoisyScrub-bird.
Hughes, P., & Hughes, B. (1991). Notes onthe Black-breasted Button-quail at Widgee,Queensland. Australian Bird Watcher, 14,113-118.Black-breasted Button-quail; eucalypt open forest;rainforestBioregion: South eastern QueenslandThe Black-breasted Button-quail is dependent ondeep leaf-litter, and occurs in vine thickets andadjacent scrubby eucalypt open forest. Thewidespread practice of frequent burning may bedetrimental to it. It can recover with absence of fire,and fire exclusion is recommended.
Hull, A. F. B. (1922). A visit to the Archipelagoof the Recherche S.W. Australia. Emu, 21,277-289.seabirds; mortality; tussock grassland; heathBioregion: Esperance PlainsNoted repeated intense deliberate burns: “somany times have fires been put through the scrubthat only possibly birds such as the Mutton Birdcould survive”.
140
Australia’s Biodiveristy - Responses to Fire

Hunt, T. J., & Kenyon, R. F. (1970). Therediscovery of the Mallee Whipbird inVictoria. Australian Bird Watcher, 3, 222-226.Western Whipbird; mallee; heath; threatened species.Bioregion: Murray Darling DepressionWestern Whipbirds may have left parts of the BigDesert and Sunset Country this century becausethe vegetation was too old (and hence understoreytoo open). Recorded here in 9 year old regrowth.Fires about a decade previously suggest that manyareas would now be suitable. A previous record ofthis species was in “recently burnt” vegetation,though there are also records from more matureregrowth.
Hutchins, B. R. (1988). Black Honeyeatersfeeding among charcoal and ash. SouthAustralian Ornithologist, 30, 160.charcoal; Black HoneyeaterA number of Black Honeyeaters visited oldcampfires and fed on charcoal and ash. As thebirds included males, the explanation that ash waseaten to provide calcium for egg-laying isinsufficient.
Jones, R. (1980). Hunters in the Australiancoastal savanna. In D. R. Harris (Ed.), HumanEcology in Savanna Environments London:Academic Press.Aboriginal knowledge; Aboriginal burning; tussockgrasslands; management; wetlandsBioregion: Top End CoastalDescribes the traditional use of fire for huntingand other resource management, includingburning of floodplains to maintain waterfowlpopulations.
Jordan, R. (1984a). The Eastern Bristlebird.Effects of fire on a population. RAOU Report,11, 30.Eastern Bristlebird; threatened species; heath; wildfireBioregion: South East CornerBefore a wildfire in coastal heathlands, EasternBristlebirds were common. Following fire, nobirds were detected for 2 months. Populationnumbers then built up to approach pre-fire levelsat 2 years post-fire. All were occupying patches ofrelatively dense regrowth.
Jordan, R. (1984b). The Ground Parrot -Effect of fire on a population. RAOU Report,11, 28-29.Ground Parrot; threatened species; heath; wildfireBioregion: South East CornerPopulations in 5yr. post-fire coastal heath were c.0.2birds/ha. For c10 months after wildfire, noGround Parrots were recorded. Then birdsbecame resident: these were most probablyimmatures.
Jordan, R. (1987a). The Barren Groundsrolling bird survey. RAOU Report, 27, 15-18.heath; eucalypt woodland; wildfire; communityBioregion: South East CornerFour year survey of heathland with somewoodland following fire. Four species (GreyShrike-thrush, Crimson Rosella, BrownThornbill, Rufous Whistler) showed no change.Four species (Golden Whistler, Flame Robin,Australian Magpie, Australian Raven) showedimmediate post-fire increase, followed by decline.Four species (White-browed Scrubwren, BeautifulFiretail, White-eared Honeyeater, EasternSpinebill) showed post-fire decline followed byincrease. Two species (Southern Emu-wren,Tawny-crowned Honeyeater) showed post-firepopulation explosion (1-2 years post-fire) thendecline.
Jordan, R. (1987b). The Ground Parrot inBarren Grounds Nature Reserve. RAOUReport, 27, 19-23.Ground Parrot; heath; threatened speciesBioregion: South East CornerPopulation density of Ground Parrots reaches apeak at 4-8 years post-fire in heathland, thendeclines to virtually absent by 12 years post-fire.
Jordan, R. (1987c). The Southern Emu-wrenin Barren Grounds. RAOU Report, 27, 24.Southern Emu-wren; heath; wildfireBioregion: South East CornerFollowing fire in coastal heathland, a fewemu-wrens moved in during the first year, butrecolonisation became rapid after about12 months. Birds were presumably moving infrom unburnt patches (mostly within 100m.).Then recruitment was rapid, in response to largepost-fire increases in insect numbers.
141
Fire and Australian birds

Jordan, R. (1988). Population changes of somecommon insectivore species in woodland nearthe wardens house. RAOU Report, 51, 16-18.eucalypt woodland; wildfire; communityBioregion: South East CornerRelatively small changes in abundance that couldbe attributable to wildfire (based on bandingresults before and up to five years post-fire) incoastal woodland. Brown Thornbill andWhite-bowed Scrubwren showed short-termincreases, possibly due to increased numbers ofinsects. No change for Striated Thornbill, whichforaged in the canopy (which remained relativelyunaffected by fire). Eastern Yellow Robin declinedfor several years post-fire possibly because of theincreased density of ground-layer vegetation.Eastern Spinebill increased for several yearspost-fire, possibly because of increased nectaravailability.
Joseph, L. (1982). The Glossy Black-cockatooon Kangaroo Island. Emu, 82, 46-49.Glossy Black-cockatoo; Casuarina woodland; eucalyptopen forest; threatened speciesBioregion: Lofty BlockGlossy Black-cockatoo is dependent on Casuarinaseeds and prefers to feed in taller, mature trees.Areas burnt 22 and 11 years prior to this surveywere still unsuitable for Glossy Black-cockatoos.
Joseph, L., Emison, W. B., & Bren, W. M.(1991). Critical assessment of the conservationstatus of Red-tailed Black-Cockatoos insouth-eastern Australia with special referenceto nesting requirements. Emu, 91, 46-50.Red-tailed Black-Cockatoo; hollows; eucalypt openforest; managementBioregions: Murray Darling-Depression;Victorian MidlandsFeeding habitat (Eucalyptus baxteri open forests)of Red-tailed Black-cockatoos in this area isthreatened by fuel-reduction burning. Nestingtrees are being lost by lack of regeneration andclearing (including for firewood).
Kavanagh, R. P. (1990). Survey of Powerful andSooty Owls in south-eastern New South Wales.Final report (Project 120). World WildlifeFund (Australia).Sooty Owl; Powerful Owl; eucalypt open forest;rainforest; old-growth; forestry; hollows; management;threatened speciesBioregions: South East Corner; South EasternHighlands.Powerful Owl requires old-growth forests (acrossa broad floristic range); Sooty Owl requiresold-growth forest, especially where there arerainforest elements in the understorey. Toofrequent fire or logging will seriouslydisadvantage these species.
Kavanagh, R. P. (1991). The target speciesapproach to wildlife management: gliders andowls in the forests of southeastern New SouthWales. In D. Lunney (Ed.), Conservation ofAustralia’s forest fauna (pp. 377-383). Mosman:Royal Zoological Society of NSW.hollows; Sooty Owl; Powerful Owl; eucalypt openforest; forestry; management; old-growth; threatenedspeciesBioregions: South East Corner; South EasternHighlandsSooty Owls and Powerful Owls are associatedwith old-growth forests, and provide goodindicators for conservation planning.
Kimber, P. C. (1974). Some effects ofprescribed burning on Jarrah Forest birds. InThird Fire Ecology Symposium (pp. 49-57).Melbourne: Forests Commission, Victoria.eucalypt open forest; control burning; community;successionBioregion: Jarrah ForestShort-term effects of (hot) control fire were limited,with slight reduction in the number of birdsimmediately following fire and extending to the firstspring after fire. After 2 years numbers increased to25% above pre-fire levels. The greatest increase wasfor species occurring in low to mid canopy levels(Western Gerygone, Brown-headed Honeyeater,White-naped Honeyeater, Grey Shrike-thrush,Inland Thornbill, Western Thornbill, GoldenWhistler). Bird populations in a forest which hadbeen unburnt for 40 years were appreciably lessthan in regularly burnt forests. This is soparticularly for species occurring in understorey andlower canopies (e.g. Rufous Treecreeper, WesternYellow Robin, Western Spinebill, Golden Whistler,Brown Thornbill).
142
Australia’s Biodiveristy - Responses to Fire

King, B., & King, D. (1983). Moggs Creek:Ash Wednesday plus nineteen days. GeelongNaturalist, 20, 19-21.eucalypt open forest; wildfireBioregion: South east Coastal PlainGeneral notes on a brief visit to anintensively-burnt open forest at 19 days followingfire. The only birds seen were a single Black-facedCuckoo-shrike, a woodswallow, Australian Ravenand Australian Magpie.
King, B. R., Limpus, C. J., & Walker, T. A.(1991). Seabird islands: No. 210. Fife Island,Great Barrier Reef, Queensland. Corella, 15,59-61.seabirds; tussock grasslandBioregion: Cape York PeninsulaSeveral fires have been recorded from the island,affecting nesting seabirds.
King, D. F. (1987). Further fauna surveys atMoggs Creek - post Ash Wednesday wildfire.Geelong Naturalist, 23, 81-84.eucalypt open forest; wildfire; communityBioregion: South east Coastal PlainList of bird species recorded during one visit threeyears post-fire. Part of ongoing survey (seeDedman 1984).
Lane, S. G. (1976). Seabird islands: No. 18.Broughton Island, New South Wales.Australian Bird Bander, 14, 10-13.seabirds; tussock grassland; mortalityBioregion: NSW North CoastFishermen frequently set fire to the grasslands onthis island. Burning may have interfered with thebreeding storm-petrels, but apparently not withshearwaters (which nest in deep burrows). Burntareas may assist the shearwaters taking off orlanding.
Lane, S. G. (1982). Seabird Islands: No. 119.Frederick Island, Archipelago of theRecherche, Western Australia. Corella, 6,61-62.seabirds; heath; tussock grasslandBioregion: Esperance PlainsMuch of the vegetation was extensively burnt byfire, probably in the year preceding this report. Ifthis fire occurred during the breeding season itwould probably have destroyed many nesting birds.
Lewis, H. T. (1989). Ecological andtechnological knowledge of fire: Aboriginesversus park rangers in northern Australia.American Anthropologist, 91, 940-961.Aboriginal knowledge; Aboriginal burning;management; tussock grasslands; review; historicchange; Magpie Goose; wetlandsBioregion: Top End CoastalReviews burning practices of Aboriginal peopleand park rangers. Aboriginal people note thatburning is important for Magpie Geese (and otherwaterfowl), as they prefer to feed in burnt areasaround their nest sites. Last fires of the year infloodplains are always set before waterfowl beginto nest.
Lindenmayer, D. B., Norton, T. W., & Tanton,M. T. (1990). Differences between wildfireand clearfelling on the structure of montaneash forests of Victoria and their implicationsfor fauna dependent on tree hollows.Australian Forestry, 53, 61-68.hollows; forestry; reviewBioregion: South Eastern HighlandsFire and clearfelling affect hollow availability verydifferently (age range of trees, spatial patterning,abundance and longevity of hollow-bearing trees),and hollow-nesting birds (such as owls andcockatoos) are more affected by clearfelling thanby fire.
Lord, E. A. R. (1936). Notes on swifts. Emu,35, 216-218.White-throated Needletail; wildfireWhite-throated Needletail (=Spine-tailed Swift)feeds on insects in smoke over bushfires. Flocks ofswifts followed fires for several days.
Loyn, R. H. (1985a). Bird populations insuccessional forests of Mountain AshEucalyptus regnans in central Victoria. Emu,85, 213-230.eucalypt open forest; wildfire; hollows; forestry;succession; managementBioregion: South Eastern HighlandsMountain Ash forests are rarely dry enough toburn, but when they do, trees are killed overextensive areas. Little regeneration occursnaturally in absence of fire. Bird abundance wasestimated at a series of sites of varying age sincefire (including 39, 73, 90 and 223 years post-fire)
143
Fire and Australian birds

or logging. Young regrowth (after clear-felling)had a very different bird fauna to mature forests,including many species typical of open areas (e.g.Nankeen Kestrel, Richard’s Pipit, AustralianMagpie, Blue-winged Parrot, Flame Robin) orlow shrubbery (Superb Fairy-wren, Red-browedFinch). The bird species composition changedlittle between 50 to about 200 years post-fire. Theabundance of some birds that were associated withparticular plants (e.g. Correa lawrenciana, Acaciadealbata) changed with the successional increaseand then decrease of these. Hole-nesting birdswere more abundant in forests >100 years than39-80 years, although dead trees left after the1939 fire offered a reasonable number of hollows.A few species were largely restricted to forests>200 years. These included Powerful Owl, SootyOwl, Australian Owlet-Nightjar andSulphur-crested Cockatoo (all hollow-nesters),Mistletoebird, Australian King-Parrot, Lewin’sHoneyeater and Satin Bowerbird (all at leastpartial fruit-eaters) and Varied Sittella. Theimmediate effects on birds of wildfire in MountainAsh forests must be profound. After the 1939fires, flocks of Gang-gang Cockatoos werereported from many suburban areas from whichthey were normally absent.
Loyn, R. H. (1985b). Ecology, distribution anddensity of birds in Victorian forests. In A. Keast, H. F. Recher, H. Ford, & D.Saunders (Eds.), Birds of eucalypt forests andwoodlands: ecology, conservation, management(pp. 33-46). Chipping Norton: Surrey Beatty.eucalypt open forest; community; hollowsBioregions: South east Coastal Plain; SouthEastern Highlands; Australian Alps; VictorianMidlands; RiverinaDiscusses effects of fire in a range of forests insoutheastern Australia. Compares densities ofhole-nesting birds in a range of mature andregrowth forests. Highest proportion ofhollow-nesters is in mature River Red Gumforests. Mistletoe is more common in older forests(and hence density of Mistletoebirds is greater).Some honeyeaters, foliage-gleaning insectivoresand bark-foraging birds are more common in
mature than in regrowth forests. In mixed foothillforests, birds that feed from bare open ground(e.g. Scarlet Robin, Buff-rumped Thornbill,Spotted Quail-thrush, White-throated Nightjar,Painted Button-quail) are more common onridges than in gullies and in more recently burntareas. In Mountain Ash forests, a fewopen-country birds (Richard’s Pipit, NankeenKestrel, Blue-winged Parrot) occur for about3 years post-fire, but regeneration is rapid andregrowth is soon colonised by bird species typicalof the understorey of mature forests.
Loyn, R. H. (1987). Effects of patch area andhabitat on bird abundances, species numbersand tree health in fragmented Victorianforests. In D. A. Saunders, G. W. Arnold, A. A. Burbidge, & A. J. M. Hopkins (Eds.),Nature conservation: the role of remnants ofnative vegetation (pp. 65-77). ChippingNorton: Surrey Beatty.eucalypt open forest; community; isolateBioregion: South east Coastal PlainBird assemblages were recorded from 56 forestfragments, and the distribution of individualspecies, and assemblage parameters, were relatedto a number of environmental factors (includingfire history). Patch size, grazing history andpresence of Noisy Miners accounted for mostvariation: fire history was not strongly correlatedwith assemblage parameters, but this was possiblybecause fire history showed little variationbetween patches.
Loyn, R. H., Traill, B. J., & Triggs, B. E.(1986). Prey of Sooty Owls in East Gippslandbefore and after fire. Victorian Naturalist, 103,147-149.Sooty Owl; threatened species; rainforest; eucalypt openforestBioregion: South East CornerDiet of Sooty Owl was restricted 4 months afterfire to one terrestrial rodent species, in contrast tovaried diet pre-fire. Owls were observed in burntforests, and contraction in diet may have been dueto greater ease of catching rats in burnt areas.
144
Australia’s Biodiveristy - Responses to Fire

Loyn, R. H., Cameron, D. G., Traill, B. J.,Sloan, J. F., Malone, B. S., Schulz, M., Earl, G.E., & Triggs, B. E. (1992a). Flora and fauna ofthe Cooaggalah Forest Block, East Gippsland,Victoria. Ecological Survey Report 20.Department of Conservation andEnvironment.wildfire; eucalypt open forest; rainforest; community.Bioregion: South East CornerA very intense bushfire burnt the study areaduring survey. The fire had burnt especiallyfiercely in some rainforest gully vegetation.Sampling continued 1-4 months, 1 year and 2years post-fire. In the 1-4 months post-fire, birdnumbers were reduced to 57% of pre-fire levels.Honeyeaters departed, and little food remainedfor them. Numbers of granivorous andfrugivorous birds were also greatly reduced.Bark-foraging and ground-foraging (e.g. SuperbLyrebird, which were observed to forage in ash)birds showed little change, and some carnivorusbirds and Flame and Scarlet Robins increased.Treecreepers and Laughing Kookaburra were theonly conspicuous birds in heavily-burnt stands.Pre-fire abundance was reached 2 years post-fire,and marginally exceeded at 3 years post-fire,though some honeyeaters (e.g. Crescent and NewHolland Honeyeaters) and Beautiful Firetail werestill well below pre-fire levels. Most localisedspecies (e.g. Southern Emu-wren, BeautifulFiretail) had returned to the sites that they hadoccupied pre-fire.
Loyn, R. H., Hewish, M. J., & Considine, M.(1992b). Short-term effects of fuel reductionburning on bird populations in Wombat StateForest. In K. Tolhurst & D. Flinn (Eds.),Ecological impact of fuel reduction burning indry sclerophyll forest: first progress report (pp.5.1-5.11). Melbourne: Department ofConservation and Environment (ResearchReport no. 349).eucalypt open forest; control burning; communityBioregion: Victorian MidlandsOver 3 years, bird populations were assessedbefore and after fuel-reduction burns at 3 areas(each with control, spring burn and autumn burnsites) in eucalypt open forest. Scarlet and FlameRobins were substantially more common inautumn-burnt areas. Birds which fed to someextent on bare ground (Laughing Kookaburra,Grey Shrike-thrush, Superb Fairy-wren) generallyincreased post-fire; those that fed in shrubby
understorey (Brown Thornbill, White-browedScrubwren) generally decreased. Nectarivorousbirds (lorikeets and honeyeaters) were morecommon in burnt areas, attracted to whatappeared to be fire-induced high-qualityflowering in some eucalypts. Three species whichwere rarely recorded before fires, becameabundant post-fire (Red-browed Finch,White-winged Chough, Pied Currawong). Ingeneral, there was little change in total birdrichness or abundance, and most individualspecies showed no significant change. Notes thatthe study areas were small, and no burnt site was>300m from unburnt forest. Also, the area hadrelatively few shrubs before burning, so impactsmay be more profound in forests of differentstructure. The study deals with changes over 2years only, and monitoring is needed to detectlonger-term changes.
Lucas, D., & Russell-Smith, J. (1993).Traditional resources of the South Alligatorfloodplain: utilisation and management.Australian Nature Conservation Agency.wetlands; tussock grasslands; Aboriginal knowledge.Bioregion: Top End CoastalSome traditional burning was aimed at increasingthe abundance of foodplants for people andimportant birds (Magpie Goose, Brolga). Earlydry season burning had to be undertaken carefullyout of consideration for ground-nesting birds,such as Partridge Pigeon.
Lucas, K., & Lucas, D. (1993). Aboriginal firemanagement of the Woolwonga wetlands inKakadu National Park. Australian NatureConservation Agency.wetlands; tussock grasslands; Aboriginal knowledge.Bioregion: Top End CoastalThe grasslands should be burnt annually atappropriate times of year. One aim of burning isto maintain or increase abundance of wild riceOryza because it is an important food for MagpieGoose. Prefer not to burn too early in the dryseason because some birds (notably PartridgePigeon, White-throated Grasswren, quails,finches and whistle-ducks) are nesting then on theground or in grass. Small fires may be relativelysafe then, and may prevent very large destructivefires later.
145
Fire and Australian birds

Lyndon, E. (1977). Aftermath of fire atWaratah Bay - and Lyrebirds. VictorianNaturalist, 94, 18-19.Superb Lyrebird; heath; eucalypt open forestBioregion: South east Coastal PlainAnecdotal report of lyrebirds occurring inregrowth 2-3 years post-fire.
MacGillivray, W. D. K. (1910). Along theGreat Barrier Reef. Emu, 10, 216-233.seabirdsBioregion: Cape York PeninsulaFire reported from Fife Island, killing breedingseabirds.
Main, A. R. (1981). Fire tolerance of heathlandanimals. In R. L. Specht (Ed.), Ecosystems ofthe World. 9B. Heathlands and relatedshrublands. Analytical studies. Amsterdam:Elsevier.review; heathBirds tend to flee in advance of smoke and flamefront, in contrast to mammals, reptiles andamphibians which burrow or re-enter burnt areasthrough gaps in fire front. Ravens, crows, magpiesand currawongs forage in recently burnt areas.Swallows and kestrels may feed in front of the fire.
Marchant, S. (1985). Breeding of the EasternYellow Robin. In A. Keast, H. F. Recher, H.Ford, & D. Saunders (Eds.), Birds of eucalyptforests and woodlands: ecology, conservation,management (pp. 231-240). Chipping Norton:Surrey Beatty.Eastern Yellow Robin; eucalypt open forest; eucalyptwoodland; wildfire.Bioregion: South East CornerAn 8 year study of banded population of EasternYellow Robin at two plots, both of which wereburnt (to various degrees) in the sixth year. Firehad little immediate effect on Robins, with allbirds apparently surviving. In the year after fire,nests were generally higher and clutches smaller,but breeding success was similar to pre-fire. Attwo years post-fire, fewer birds nested and someterritories were deserted. This was assumed to bebecause a dense grass layer had developed whichhindered foraging. Fire effects may have beenmore serious if the fire was during the breedingseason.
Matthew, J. (1994). The status, distributionand habitat of the Slender-billed ThornbillAcanthiza iredalei in South Australia. SouthAustralian Ornithologist, 32, 1-19.Slender-billed Thornbill; heath; mallee.Bioregion: Murray Darling DepressionThe subspecies of Slender-billed Thornbilloccurring in mallee heath (Acanthiza iredaleihedleyi) may be considered vulnerable to fires, butfire is not a threatening process for subspeciesoccurring in samphire.
Mawson, P. R., & Long, J. L. (1994). Size andage parameters of nest trees used by fourspecies of parrot and one species of cockatooin south-west Australia. Emu, 94, 149-155.hollows; eucalypt woodland; eucalypt open forest; RegentParrot; Red-capped Parrot; Western Rosella; PortLincoln Ringneck; Western Long-billed CorellaBioregions: Avon Wheatbelt; Jarrah ForestHot fires may create, enlarge or destroy hollows.Dimensions of hollows (and size and age of trees)used by Regent Parrot, Red-capped Parrot,Western Rosella, Port Lincoln Ringneck andWestern Long-billed Corella are given. Treesused are very old (minimum 73 years, maximum>1300 years). Clearing is main cause of reductionin hollow availability, but frequent burning ofroadside verges prevents seedling establishmentand hence tree recruitment. In some remnantpatches, complete absence of fire may also preventgermination.
McAllan, I. A. W. (1996). The FlockBronzewing Phaps histrionica in New SouthWales, with comments on its biology.Australian Bird Watcher, 16, 175-204.Flock Bronzewing; review; tussock grasslandsBioregions: Mitchell Grass Downs; ChannelCountry; Darling Riverine Plains; Mulga Lands;Broken Hill ComplexAlthough exclusion of fire by pastoralists inMitchell grasslands may have reduced seedavailability, this can “be only a minor problem”for Flock Bronzewings.
146
Australia’s Biodiveristy - Responses to Fire

McCaw, L., Maher, T., & Gillen, K. (1992).Wildfires in the Fitzgerald River National Park,Western Australia, December 1989. Departmentof Conservation and Land Management.wildfire; mortalityBioregion: Esperance PlainsThe remains of a large number of small birds werefound in a woodland burnt by relatively lowintensity wildfire. Following a patchy wildfire,unburnt vegetation was important as refuge forless mobile species. Large flocks of granivorousbirds were observed well within anextensively-burnt area within days followingwildfire.
McCulloch, E. M. (1966). Swifts andbushfires. Emu, 65, 290.White-throated Needletail; Fork-tailed Swift; wildfireBoth White-throated Needletail and Fork-tailedSwifts flocks fed in insects in smoke overbushfires.
McFarland, D. C. (1988). The composition,microhabitat use and response to fire of theavifauna of subtropical heathlands in ColoolaNational Park, Queensland. Emu, 88, 249-257.heath; community; succession; managementBioregion: South Eastern QueenslandBird species composition and richness wasdetermined at sites 0,2.5,5.5,6.5 and 10.5 yearspost-fire. Older heaths had fewer “inconsistent”species (irregular visitors, migrants, etc.). Post-firerecolonisation depends upon extent and intensityof fire and availability of unburnt patches. Speciesrichness was higher (39 spp.) in the first year afterfire than in the oldest site (14 species), althoughmany of the species in the young site weremigrants or visitors from nearby habitats. Theselatter were attracted by post-fire flowering orseeding or exposed and dead invertebrates andvertebrates immediately after fire (e.g. for raptors,Laughing Kookaburra, Torresian Crow,Straw-necked Ibis). Responses to fire age wererelated to food and shelter. Most structuralattributes either peaked or reached a plateau by3-6 years post-fire. Brown Quail used seedinggrasses in open regeneration. King Quail, BrushBronzewing and Ground Parrot preferred thickervegetation of middle-aged heathlands (which alsoheld greatest abundance of their seed resources).Swallows, martins and Richard’s Pipit preferredopen young sites. Tawny Grassbird,
Golden-headed Cisticola, Red-backed Fairy-wrenand Southern Emu-wren were most abundant inmid to late ages. Pheasant Coucal and EasternGrass Owl showed no relationships with age. Pastpractice of burning every c3 years has advantagedopportunistic species, but disadvantaged most ofthe resident species. Longer intervals betweenburns (e.g. 7-8 years) and mosaic burns arerecommended to maintain resident species.
McFarland, D. C. (1989). The Ground ParrotPezoporus wallicus (Kerr) in Queensland:habitat, biology and conservation. Departmentof Conservation, Parks & Wildlife,Queensland.Ground Parrot; threatened species; heath; isolateBioregion: South Eastern QueenslandGround Parrots were most abundant in heathsfrom 5-8 years post-fire, and are threatened byfire intervals of less than this. This threat isexacerbated by fragmentation.
McFarland, D. C. (1991). The biology of theGround Parrot, Pezoporus wallicus, inQueensland. III. Distribution and abundance.Wildlife Research, 18, 199-213.Ground Parrot; heath; threatened species; sedgeland;isolateBioregion: South Eastern QueenslandDescribes distribution and habitat preferences forGround Parrot in subtropical heaths. AlthoughGround Parrots were recorded at a site 2 monthspost-fire, they were not recorded regularly until>9 months post-fire. Densities peaked at 5-8 yearspost-fire, when the number of food plants, andstanding crop of food, is maximum, and whenthese are most stable seasonally. Decline ofnumbers of Ground Parrots in older heath reflectsreduced seed availability. High intensity fires maybe more deleterious than fuel-reduction burns.Ground Parrots recolonise rapidly after eithersummer or winter fires. Increasing heathlandisolation increases the possibility of completeburns with catastrophic fire and reduces thechance of recolonisation.
147
Fire and Australian birds

McFarland, D. C. (1992). Fire and themanagement of ground parrot habitat. In B.R. Roberts (Ed.), Fire Research in ruralQueensland (pp. 483-495). Toowomba:University of Southern Queensland.Ground Parrot; heath; management; threatened speciesBioregion: South Eastern QueenslandDensities of the fire-sensitive and fire-dependentGround Parrots in subtropical heaths peak at 5-8years post-fire, with no birds recorded at >15 yearspost-fire. A management plan for Ground Parrotsin heathlands of southern Queensland is described:it involves moderate to low intensity burning at8-10 years during winter. Impacts of this regimeon other heathland biota are considered. Of 12species of breeding resident birds, 7 reach peakdensities at or >6 years post-fire.
McFarland, D. C. (1993). Fire and birdconservation. In C. P. Catterall, P. V. Driscoll,K. Hulsman, D. Muir, & A. Taplin (Eds.),Birds and their habitats: status and conservationin Queensland (pp. 41-44). St Lucia:Queensland Ornithological Society Inc.review; Ground Parrot; heath; threatened species;community; managementBioregion: South Eastern QueenslandIn southeastern Queensland, Ground Parrots nestonly in dry heaths that are at least 3-4 yearspost-fire. Fire is the main factor affecting density.Ground Parrots can recolonise burnt heathswithin 12 months, densities peak at 5-8 yearspost-fire, and no birds have been recorded inheaths >15 years post-fire (but few such areasoccur or have been searched). This patterncorrelates with seed availability and cover. Otherdeclining species in Queensland affected by fireregime include Golden-shouldered Parrot,Eastern Bristlebird, Rufous Scrub-bird,Carpentarian and Striated Grass-wrens. Inheathland, raptors, egrets and ibis move in duringand just after fires. Within 1 year of fire, there isusually massive flowering of Xanthorrhoea, whichattracts many honeyeaters. Granivores colonisesomewhat later. The total number of species ishighest within 1 year post-fire, but then declines.The number of breeding species peaks between3-8 years post-fire, followed by decline, especiallyfor granivorous species.
Controlling fire to benefit one bird speciesmay have deleterious impact on other species. Inheathlands of southeastern Queensland, mosaicwinter burning at intervals of 8-10 years isrecommended.
McFarland, D. C. (1994). Notes on the BrushBronzewing Phaps elegans and SouthernEmu-wren Stipiturus malachurus in CooloolaNational Park. Sunbird, 24, 14-17.heath; Brush Bronzewing; Southern Emu-wren;isolateBioregion: South Eastern QueenslandHeaths of 0-10.5 years post-fire were searched.No Brush Bronzewings were recorded fromheaths younger than 2 years or older than 10.5years. Southern Emu-wrens occurred in heaths ofage 1.5 to 9 years, with highest density in sites of6-8 years. Fragmentation and generally high pastfire frequency may have eliminated Emu-wrensfrom what now appears suitable heaths.
McIlroy, J. C. (1978). The effects of forestrypractices on wildlife in Australia: a review.Australian Forestry, 41, 78-94.review; eucalypt open forest; management; forestryMost Australian fauna is fire-adapted. Highintensity fires and frequent low intensity fireshomogenise forests, leading to elimination ofspecies dependent on shrubs, litter and logs. Of 48bird species in a native forest, 12 occurredfollowing clear-felling. All of these remained afterthe felled area was burnt.
McKean, J. L., & Martin, K. C. (1985).Distribution and status of the CarpentarianGrasswren Amytornis dorotheae. ConservationCommission of the Northern Territory.Carpentarian Grass-wren; hummock grasslands;management; threatened speciesBioregion: Gulf Fall and UplandsHot extensive wildfires are the most serious threatto the Carpentarian Grass-wren.
McKean, J. L., & Martin, K. C. (1989).Distribution and status of the CarpentarianGrass-wren Amytornis dorotheae. NorthernTerritory Naturalist, 11, 12-19.Carpentarian Grass-wren; hummock grassland;management; threatened speciesBioregion: Gulf Fall and UplandsExtensive wildfires are the most severe threat toCarpentarian Grass-wren, and recent fires haveled to its decline and contraction
148
Australia’s Biodiveristy - Responses to Fire

McLaughlin, J. (1992). The floristic andstructural features of Black-eared MinerManorina melanotis habitat. RAOU Report No.84. Royal Australasian Ornithologists Union.Black-eared Miner; threatened species; old-growth.Bioregion: Murray-Darling Depression.Mallee of at least 55-60 years post-fire is required,with younger mallee possessing few of thestructural features (decorticating bark) preferredby this species.
McLaughlin, J. (1994). Searches for theBlack-eared Miner Manorina melanotis in theVictorian Murray Mallee. RAOU Report No. 93.Royal Australasian Ornithologists Union.Black-eared Miner; mallee; threatened speciesBioregion: Murray-Darling DepressionThe endangered Black-eared Miner is restrictedto long-unburnt mallee remote from edges.
McNamara, E. (1946). Field notes on theEastern Bristlebird. Emu, 45, 260-265.Eastern Bristlebird; heath; threatened speciesBioregion: South East CornerThe causes of the rarity of Eastern Bristlebirds areobscure. Bushfires may be the chief reason, asheaths are vulnerable to fire, and birds would havetrouble surviving in burnt areas. But much of theheath where they occur now has been burnt in thepast. Unburnt patches may be the key to theirsurvival.
McNee, S. (1986). Surveys of the WesternWhipbird and Western Bristlebird in WesternAustralia. RAOU Report No. 18. RoyalAustralasian Ornithologists Union.Western Whipbird; Western Bristlebird; heath; mallee;threatened species; management.Bioregions: Esperance Plains; Warren; JarrahForestThe range and abundance of the WesternWhipbird and Western Bristlebird have beensubstantially reduced by clearing and too frequentfires. Management of these species will requirelong intervals without fire (at least 30 years andpreferably 50 years for Western Whipbird).Western Whipbird is known to survive fire, butthen not persist in burnt areas.
Menkhorst, P. W., & Bennett, A. F. (1990).Vertebrate fauna of mallee vegetation insouthern Victoria. In J. C. Noble, P. J. Joss, &G. K. Jones (Eds.), The mallee lands: a conser-vation perspective (pp. 39-53). Melbourne:CSIRO.malleeBioregion: Murray Darling DepressionMallee vegetation is highly flammable and forsome birds certain seral stages in the post-firesuccession offer a higher quality habitat. Speciesfavouring climax vegetation include Black-earedMiner and Malleefowl. Fire management sinceEuropean settlement has reduced habitatheterogeneity and especially the extent ofold-growth mallee. This is especially so in isolatedpatches which fire can reduce to a single age class.
Meredith, C. W. (1982). The research input tofire management in natural areas withparticular reference to wildlife research in thesemi-arid lands of Victoria and New SouthWales. In A. Heislers, P. Lynch, & B. Walters(Eds.), Fire ecology in semi-arid lands (10pp).Deniliquin: CSIRO.mallee; community; managementBioregion: Murray Darling DepressionComprehensive studies are required for input intomanagement, else casual observations and mythsubstitute for knowledge. Notes that fire regimeincludes season, intensity, pattern and areaaffected, as well as frequency; that fire frequencyshould be a statistical distribution rather than aninflexible repetition of a mid-point; that“optimum” fire regimes vary between species andeven between short- and long-term within species;and that unplanned random burning is not thesame as mosaic burning. Mallee woodlandendemic birds are most common in areas >20-25years post-fire. Specialist species with restricteddistributions tend to be found in older vegetationwhile widespread generalists are typical of earlyregeneration. However some specialists mayfavour early regrowth. While Malleefowl requirelong period between fires, it may be that very oldvegetation has a declining food supply. ShyHeath-wren, Red-lored Whistler, WesternWhipbird and Striated Grass-wren may beassociated with early stages of regeneration.Research needs include study of fire relationshipsof other mallee endemic birds, and the effect offire on hollow formation and seed and nectarproduction.
149
Fire and Australian birds

Meredith, C. W. (1983). Fire and birds. Theresult of two studies and their relevance tofuel reduction burning. In E. H. M. Ealey(Eds.), Fighting fire with fire (pp. 193-202).Melbourne: Graduate School ofEnvironmental Science Monash University.mallee; community; Ground Parrot; heath; sedgelands;control burning; threatened speciesConsiders two examples: bird communities ofmallee and Ground Parrot in coastal heathlands.In both habitats, there is little functionaldifference between wildfire and fuel reductionburns. Burning frequency in mallee of <20-30years would result in 25% or more reduction inbird species richness. Habitat endemic specieswould be those most likely to be lost. Theseinclude those requiring abundant litter (e.g.Malleefowl), hollows (Chestnut-rumpedThornbill, parrots) or dense large spinifex(Striated Grass-wren, Mallee Emu-wren). Birdnumbers may be greatest at about 15 yearspost-fire. Species common in early regrowthinclude mainly widespread opportunists (Kestrel,Australian Magpie, White-fronted Honeyeater,some thornbills). Management should concentrateon localised burning at borders. Ground Parrotcan survive in sedgelands for indefinite period, butin heathlands it requires fire every 20-25 years.Currently, fuel-reduction burning is too frequent,and may eliminate this species from graminoidheathlands.
Meredith, C. W. (1984a). The Ground Parrot.RAOU Conservation Statement, 1.Ground Parrot; heath; sedgeland; threatened speciesInappropriate fire regimes, compounded byfragmentation of populations, are the main threatsto the Ground Parrot
Meredith, C. W. (1984b). Management of theGround Parrot Pezoporus wallicus in Victoria.Fisheries & Wildlife Division, Victoria.Ground Parrot; heath; sedgelands; threatened species;managementDetailed recommendations for fire managementat sites of varying floristics in Victoria, in order toconserve Ground Parrot populations.
Meredith, C. W. (1984c). Recent records ofthe Ground Parrot Pezoporus wallicus inwestern Victoria. Geelong Naturalist, 21, 3-4.Ground Parrot; heath; threatened species; controlburningBioregion: South east Coastal PlainThe disappearance of Ground Parrot from somesites was related to old age of the heaths. The lownumbers in another site were a result of extensiveand frequent fuel reduction burns.
Meredith, C. W. (1988). Fire in the Victorianenvironment - a discussion paper. ConservationCouncil of Victoria.review; managementShort-term effects of fire on bird communitiesvary with fire intensity (and the extent ofvegetation change). However if fire regimes leadto long-term change in vegetation characteristics,there will be long-term change in birdcommunities. If fire frequency is too great, birdsdependent on older stages will be lost. These areoften specialist species with restricteddistributions. Notes that no long-term studies ofthe relationships between birds and fire have beenconducted in any Australian habitat; and thatprevious studies (e.g. Kimber 1974, Christensen &Kimber 1975) have restricted analysis to commonspecies in the communities they studied.
Meredith, C. W., & Isles, A. C. (1980). A studyof the Ground Parrot (Pezoporus wallicus) inVictoria. report No. 304. EnvironmentalStudies Division of the Ministry forConservation, Victoria.Ground Parrot; heath; threatened species.Bioregions: South East Corner; South eastCoastal Plain; Naracoorte Coastal PlainGround Parrot abundance peaks at 5-10 yearspost-fire; heaths then eventually becomeunsuitable.
150
Australia’s Biodiveristy - Responses to Fire

Meredith, C. W., & Jaremovic, R. (1990).Current status and management of the GroundParrot in Victoria. Arthur Rylah Institute forEnvironmental Research.Ground Parrot; heath; wildfire; control burning;threatened species; managementBioregions: South East Corner; South eastCoastal PlainHot fire in a coastal heathland reduced theGround Parrot population at this site by 75%.Elsewhere, widespread and frequentfuel-reduction burns in heathland led tosubstantial and lasting reductions in GroundParrot densities. A model is developed to predictpopulation changes post fire, and hence to designmanagement burning regimes.
Meredith, C. W., Gilmore, A. M., & Isles, A. C.(1984). The Ground Parrot (Pezoporus wallicusKerr) in south-eastern Australia: afire-adapted species? Australian Journal ofEcology, 9, 367-380.Ground Parrot; heath; sedgelandBioregions: South east Corner; South east CoastalPlain; Naracoorte Coastal Plain.Examined Ground Parrot abundance at a series ofsites of known age since fire. Fire age is the majordeterminant of abundance in heathlands, mainlythrough its influence on food resources. Theproduction of seeds eaten by Ground Parrotremains relatively constant over time insedgelands but varies in heathland with time sinceburning. Long unburnt (>20 years) and veryfrequently burnt (<6-8 years) heathlands areunsuitable for Ground Parrot. Fires areuncommon in sedgelands, and Ground Parrotsshow no relationship with sedgleland age. GroundParrots may disperse to unusual habitats after fire,and immatures may move long distances. GroundParrots are not fire-adapted, but rather they areadapted to a diet dominated by the seeds ofgraminoid sedges. In habitats where theavailability of these seeds is unrelated to fire (e.g.sedgelands) Ground Parrot abundance is alsounrelated to fire, but in heathlands, the GroundParrot is a fire-requiring species because of itsadaptation to a particular diet.
Milledge, D. R., & Palmer, C. L. (1990). TheSooty Owl in Mountain Ash forests in theVictorian Central Highlands. Department ofConservation and Environment.Sooty Owl; hollows; eucalypt open forest; old-growth;management; threatened speciesBioregion: South Eastern HighlandsSooty Owls are virtually confined to forests thatare at least 150 years old, because of requirementsfor high density of hollows (for their own nestingand that of their prey).
Milledge, D. R., Palmer, C. L., & Nelson, J. L.(1991). “Barometers of change”: thedistribution of large owls and gliders inMountain Ash forests of the Victorian CentralHighlands and their potential as managementindicators. In D. Lunney (Ed.), Conservation ofAustralia’s Forest Fauna (pp. 53-65). Sydney:Royal Zoological Society of NSW.eucalypt open forest; old-growth; hollows; threatenedspeciesBioregion: South Eastern HighlandsSurvey of owls at four age classes (50 to >250years). Sooty Owl was associated with large areasof old-growth (>250 years) forest, though didoccur in 50-80 year forests (where fire-killed stagsremained).
Milligan, A. W. (1904). Notes on a trip to theWongan Hills, Western Australia. Emu, 4,2-11.Western Whipbird; threatened species; heath.Bioregion: Avon WheatbeltRepeated fires probably led to the disappearanceof Western Whipbirds from the Wongan Hillsarea.
Mollison, B. C., & Green, R. H. (1962).Mist-netting Tree-Martins on charcoalpatches. Emu, 61, 277-280.Tree Martin; charcoalTree Martins observed to collect and maybeconsume charcoal and ash.
151
Fire and Australian birds

Morton, S. R., & Brennan, K. G. (1991).Birds. In C. D. Haynes, M. G. Ridpath, &M. A. J. Williams (Eds.), Monsoonal Australia:landscape, ecology and man in the northernlowlands. (pp. 133-149). Rotterdam: Balkema.review; tropical eucalypt open forest; managementLittle doubt that changed fire regimes have affectedbirds. If current burning regimes have higherfrequency of late dry season hot fires (as is likely)then birds associated with shrubby understorey mayhave declined considerably. Research into themanagement of fire for birds needs high priority.
Nelson, J. L., & Morris, B. J. (1994). Nestingrequirements of the Yellow-tailedBlack-cockatoo, Calyptorhynchus funereus, inEucalyptus regnans forest, and implications forforest management. Wildlife Research, 21,267-278.Yellow-tailed Black-cockatoo; hollows; managementBioregion: South eastern HighlandsThe mean estimated age of trees used for nestingby Yellow-tailed Black-cockatoos was 221 years(with youngest at 162 years). Live trees are betternesting sites than dead stags as they are better ableto withstand fire.
Newsome, A. E., McIlroy, J., & Catling, P.(1975). The effects of extensive wildfire onpopulations of twenty ground vertebrates insouth-east Australia. Proceedings of theEcological Society of Australia, 9, 107-123.wildfire; heath; eucalypt open forestBioregion: South East CornerTracks of large vertebrates were counted before andafter an extensive hot wildfire. No changes weredetected for Superb Lyrebird (the only birdconsidered), though there were few records tocompare. Fire may reset fauna succession in drysclerophyll forests, but “it is too early to think ofutilising fire as a tool for management of fauna inNational Parks”. Wildfire is not necessarily thebogey widely imagined.
Nichols, O. G., & Muir, B. (1989). Vertebratesof the jarrah forest. In B. Dell, J. J. Havel, &N. Malajczuk (Eds.), The Jarrah Forest(pp. 133-153). Dordrecht: Kluwer.eucalypt open forest; reviewBioregion: Jarrah ForestUnderstorey birds are affected for 1-2 years bycool burns in Jarrah forests; canopy species showlittle response. The Jarrah forest fauna is adaptedto a great variety of fire regimes, many irregularand unpredictable.
Noske, R. (1988). The status and biology of theWhite-throated Grass-wren. AustralianNational Parks & Wildlife Service.White-throated Grasswren; hummock grasslandBioregion: Pine Creek ArnhemFire may decrease some populations ofWhite-throated Grass-wrens, but the status of thespecies is probably secure. Fires affect populationsfor two reasons: Grass-wrens rely on livingvegetation both directly and indirectly for plantand insect food; and they may be killed by firebecause of their ground-dwelling habits and poorflying ability. Fire frequency may be moreimportant than intensity.
Noske, R. A. (1992a). Do Grasswrens have thenumbers? Reply to Woinarski (1992).Northern Territory Naturalist, 13, 5-8.White-throated Grasswren; hummock grasslandBioregion: Pine Creek ArnhemMost sites where White-throated Grasswrenswere recorded had been burnt in the previous fewyears. “Indeed I was impressed by the ability ofGrasswrens to survive in areas of recent severefires.” Long-term absence of fire may bedisadvantageous.
Noske, R. A. (1992b). The status and ecologyof the White-throated Grasswren Amytorniswoodwardi. Emu, 92, 39-51.White-throated Grasswren; hummock grasslandBioregion: Pine Creek ArnhemFire is important in the ecology ofWhite-throated Grasswrens, but information onits effects is inconclusive. Regular burning maybenefit this species through maintenance ofhummock grasslands at the expense of morefire-sensitive plants. The species was recordedacross the range of fire ages studied (6 months
152
Australia’s Biodiveristy - Responses to Fire

post-fire to “many years” post-fire). Fire may haveimmediate impacts of reduction in food resourcesand nest sites. Absence from some sites may bedue to high frequency of fires. Recent changes infire regime may suggest that its secure statuscannot be assumed.
Paton, J. B., & Paton, D. C. (1977). SeabirdIslands: No. 52. Wright Island, SouthAustralia. Corella, 1, 68-69.seabirdsBioregion: Eyre and Yorke BlocksA fire lit to control exotic Boxthorn spreadthrough all vegetation. The resultant vegetationloss caused erosion and damage to penguinburrows, and discouraged Silver Gulls frombreeding, but probably had little impact on FairyTerns.
Pattemore, V. (1980). Effects of the pulpwoodindustry on wildlife in Tasmania. NationalParks and Wildlife Service, Tasmania.forestry; management; eucalypt open forestMainly concerned with effects of forestrypractices, but considers interaction with fireregimes.
Pedler, L. (1991). Rare bird survey: implicationsfor fire management for Uluru National Park.Australian National Parks and WildlifeService.hummock grassland; mallee; Striated Grasswren;threatened species; managementBioregion: Great Sandy DesertExamines distribution of rare bird species aroundUluru. Striated Grasswren requires maturespinifex. Recommends a fire management strategywhich produces a fine-grained diversity of fireages in hummock grassland and mallee, and theretention of long-unburnt patches.
Pedler, L., & Burbidge, A. H. (1995). Therange and status of the Nullabor Quail-thrush.South Australian Ornithologist, 32, 45-52.Nullabor Quail-thrush; chenopod shrubland;management; threatened speciesBioregion: NullaborFire is a threat to Nullabor Quail-thrush as itspreferred bluebush is killed in severe fires, andsubsequent regeneration may be affected byherbivore grazing.
Pescott, T. (1983). Beach-washed birds afterthe Ash Wednesday fire. Geelong Naturalist,20, 17-19.eucalypt open forest; wildfire; mortalityBioregion: South east Coastal PlainLarge numbers of bush birds were washed up onthe beach following hot fire. The birds generallydid not show any signs of having been burnt. Atotal of 2183 individuals of 66 species wererecorded. The most numerous dead birds wereCrimson Rosella (984 individuals), New HollandHoneyeater, Red Wattlebird, Currawongs,Sulphur-crested Cockatoo, White-earedHoneyeater, Ravens, Australian Magpies andWhite-naped Honeyeater.
Pescott, T. (1985). Black Honeyeaters andcharcoal. Geelong Naturalist, 22, 37-39.charcoal; Black HoneyeaterBlack Honeyeaters observed to repeatedly collectand consume charcoal.
Pescott, T. W. (1976). Seabird Islands: No. 27.Lady Julia Percy Island, Victoria. AustralianBird Bander, 14, 29-31.seabirds; tussock grasslandBioregion: South east Coastal PlainOccasional fires have had considerable impact onvegetation, at times leading to rapid degenerationof petrel and penguin burrows.
Porter, J. W., & Henderson, R. (1983). Birdsand burning histories of open forest atGundiah, southeastern Queensland. Sunbird,13, 61-69.eucalypt open forest; historic change; managementBioregion: South Eastern QueenslandBird communities were sampled in forests whichhad been subjected to three fire regimes over29 years: burnt annually, burnt periodically at2-5 year intervals, and unburnt. The annuallyburnt forest had developed a grassy understoreyand the unburnt forest had a shrubby understorey.Four species (Pheasant Coucal, Willie Wagtail,Brown Treecreeper and Australian Magpie) weremost abundant in the annually burnt forest. Twospecies (Forest Kingfisher, White-throatedHoneyeater) were most abundant in the forestburnt at longer intervals. Six species (EasternYellow Robin, Golden Whistler, VariegatedFairy-wren, White-throated Treecreeper, Little
153
Fire and Australian birds

Wattlebird and Yellow-faced Honeyeater) weremost abundant in the unburnt forest. 15 of 37common species showed no apparent preference.The total abundance and richness of birds did notdiffer appreciably between treatments. Thecurrent periodic burning of these forests isprobably restricting encroachment of rainforestsand their associated birds.
Press, A. J. (1987). Fire management inKakadu National Park: the ecological basis forthe active use of fire. Search, 18, 244-248.managementBioregions: Pine Creek Arnhem; Top End CoastalIn the absence of management, up to 80% of thearea can be burnt annually. Fires attract falcons,kites and Tree Martin, which hawk in front of thefire. Immediately after fire, Grey Butcherbird,Magpie-lark, Straw-necked Ibis, Torresian Crow,Red-tailed Black-cockatoo, owls and nightjarsfeed in the burnt area.
Priddel, D. (1989). Conservation of rare fauna:the Regent Parrot and the Malleefowl. In J. C.Noble & R. A. Bradstock (Eds.),Mediterranean landscapes in Australia: malleeecosystems and their management (pp. 243-249).Melbourne: CSIRO.Malleefowl; mallee; threatened species; managementBioregion: Murray Darling DepressionActive management of mallee habitat is requiredfor the maintenance of its biota. Important foodsources for the Mallee-fowl are available only afterthe plants reach a certain age. Homogeneoushabitat (such as following extensive fire) may bedetrimental to it.
Priddel, D. (1990). Conservation of theMalleefowl in New South Wales: anexperimental management study. In J. C. Noble, P. J. Joss, & G. K. Jones (Eds.),The mallee lands: a conservation perspective(pp. 71-74). Melbourne: CSIRO.Malleefowl; mallee; threatened species; isolate;managementBioregion: Murray Darling DepressionOne of four given threats to the long-termsurvival of Malleefowl is fire. There is acorrelation between breeding density and agesince fire. Almost all extensive stands of mallee inNSW have been burnt within the last 30 years,
many repeatedly. Extensive areas of long-unburntmallee are now rare in NSW. Fire threatensmallee remnants in a different way to extensivestands, with the isolation of remnants offeringsome protection, but when fires occur in them theyare likely to homogenise the remnant completely.
Priddel, D., & Wheeler, R. (1990). Survival ofMalleefowl Leipoa ocellata chicks in theabsence of ground-dwelling predators. Emu,90, 81-87.Malleefowl; mallee; threatened speciesBioregion: Murray Darling DepressionHabitat quality may be the most critical elementin the recruitment and survival of Malleefowl.This may be related to rainfall. “Alternatively, theage of the mallee community (i.e. time since lastfire) may be the critical determinant of habitatsuitability”.
Pyke, G. H., Saillard, R., & Smith, J. (1995).Abundance of Eastern Bristlebirds in relationto habitat and fire history. Emu, 95, 106-110.Eastern Bristlebird; eucalypt open forest; heath;managementBioregion: South East CornerSampled Eastern Bristlebirds in vegetation at sites(forest, woodland, “mallee-heath” and heath) from0-14 years post-fire (all fires were control burns).Bristlebird numbers increase with increasing timesince fire up to about 9 years and then remainrelatively constant for at least 4 years. Bristlebirdsoccurred in recently burnt areas, but these weremostly small and close to unburnt areas, Firemanagement is essential for the conservation ofBristlebirds. Numbers can be maintained if theinterval between fires is at least 10 years, and ifunburnt patches are retained during controlburning.
Pyne, S. J. (1991). Burning bush: a fire historyof Australia. New York: Henry Holt andCompany.reviewProvides a broad review of the history andcharacter of fire in Australia, with a few reviewexamples of responses of birds to fire.
154
Australia’s Biodiveristy - Responses to Fire

Quin, B. R. (1990) Conservation and status ofthe Turquoise Parrot (Neophema pulchella,Platycercidae) in Chiltern State Park andadjacent areas. M.Sc., Department of Zoology,LaTrobe University.Turquoise Parrot; threatened species; management.Bioregion: South Eastern HighlandsFires (wildfire and control burns) destroy thestumps often used by nesting Turquoise Parrots.
Quin, B. R., & Baker-Gabb, D. (1993).Conservation and management of the TurquoiseParrot Neophema pulchella in north-eastVictoria. Department of Conservation andEnvironment.Turquoise Parrot; eucalypt woodland; threatenedspecies; management; control burning; hollowsBioregions: South Eastern Highlands; NSWSouthwestern SlopesDead trees and stumps, favoured for nesting byTurquoise Parrots, are susceptible to destruction byfire. These should be protected from controlburning (or if burning must occur, the vicinity of allnest sites should be first slashed). Management maybe able to impose a fire regime which increases thedominance of native perennial grasses.
Ratkowsky, A. V. (1978). The effect of a springfire on the number of bird species. TasmanianNaturalist (53), 11-12.eucalypt open forest; wildfire; communityBioregion: FreycinetBirds were counted in burnt and unburnt dryeucalypt forest (either side of a road) from 5 daysto 13 weeks after a moderate intensity Octoberwildfire (foliage up to about 8m was destroyed). Inthe first 11 weeks post-fire, there was an averageof 2.7 species. Then this suddenly increased to 7.0species, equal to that in the unburnt area. Duringthe first week post-fire, Fantailed Cuckoo,Black-headed Honeyeater and Black-facedCuckoo-shrike remained in the burnt area, butthey then disappeared from it. After 11 weeks,Black-headed Honeyeater, Brown Thornbill,Superb Fairy-wren and, to a lesser extent, DuskyWood-swallow, Black-faced Cuckoo-shrike, andStriated Pardalote returned to the burnt area. Atthis period there was considerable regrowth at thebase and trunks of eucalypts and recovery of otherplants. There was no change observed for ForestRaven, Yellow-throated Honeyeater, GreyShrike-thrush, Grey Currawong, Blackbird orDusky Robin.
Ratkowsky, A. V. (1979). The bird species ofMt. Nelson in relation to microhabitat andrecent bushfires. Tasmanian Naturalist (57),12-18.eucalypt open forest; wildfire; community; controlburningBioregion: FreycinetNo. of bird species declined from 11.6 before aNovember controlled burn in dry eucalypt forestto an average of 6.9 in the 3 months after fire.
For 19 weeks after a hot October fire, birdrichness in burnt areas was far less (5.7) than inadjacent unburnt areas (16.2 species). Speciesabsent from, or rarer in, the burnt areas includedSwift Parrot, Green Rosella, Fantailed Cuckoo,Shining Bronze-cuckoo, Olive Whistler, GoldenWhistler, Satin Flycatcher, Superb Fairy-wren,Brown Thornbill, Yellow Wattlebird,Yellow-throated Honeyeater, Strong-billedHoneyeater, Black-headed Honeyeater, CrescentHoneyeater, New Holland Honeyeater, SpottedPardalote, Silvereye, Grey Butcherbird and ForestRaven. Species more common in the burnt areawere Common Bronzewing and DuskyWood-swallow. Common species showing littledifference included Black-faced Cuckoo-shrike,Blackbird, Grey Shrike-thrush and EuropeanGoldfinch. More prolonged impact of this fire,compared to the milder fire considered byRatkowsky (1978), were because this severe firehad consumed much more of the vegetation,including tree canopies. This “indicates clearlythat intensity of fire is a very important factor indetermining the rate at which avifauna willrepopulate a burnt area”.
Ratkowsky, A. V. (1985). The effect of a severefire on the number of bird species in a wetsclerophyll environment. TasmanianNaturalist, 8-9.eucalypt open forest; community; wildfireBioregion: FreycinetFollowing hot fire in January, more bird specieswere observed in unburnt area (17.5 spp.) than inburnt area (6.5 spp) in October-December.Species absent from the burnt area includedShining Bronze-cuckoo, Olive Whistler, GoldenWhistler, White-browed Scrubwren andStrong-billed Honeyeater. Species less common inburnt area included Peregrine Falcon,Yellow-tailed Black-cockatoo, Black-facedCuckoo-shrike, Black-headed Honeyeater,Silvereye, Swift Parrot, Green Rosella, Blackbird,
155
Fire and Australian birds

Yellow-throated Honeyeater, CrescentHoneyeater and Spotted Pardalote. The StriatedPardalote was the only species common in theburnt area (though not more common than in theunburnt area). Results generally similar to thosereported following severe fire in drier eucalyptopen forest (Ratkowsky 1979).
Recher, H. F. (1981). Death of an Australianmyth: fire and its effects on wildlife. In P. J. Stanbury (Ed.), Bushfires: their effect onAustralian life and landscape. (pp. 39-48).Sydney: Macleay Museum, University ofSydney.review; wildfire“The effects of fire on animals … have beenneglected by Australian scientists … There arefew long term studies.” In mild fires, birds easilyavoid flames. Some species are attracted to the firefront. In a mild burn, birds were observed in thestill smoking forest within moments of the flamepassing. In extensive wildfires, immediate effect ismore severe. But birds respond rapidly because oftheir high mobility and high reproductivepotential. Within a year of burning, birds arenumerous in forest. Many will have survived insmall unburnt patches but others probably movein from distant areas unaffected by fire. Birds mayrespond to flush of insects on regrowthvegetation. Forest canopy birds are generally lessaffected than ground-dwelling birds or birds ofthe shrub layer. Nectarivorous birds may beaffected if fire frequency leads to decline innectariferous plants. A diversity of fires (ratherthan regular prescriptive burns) is needed tomaintain the greatest range of bird species.
Recher, H. F. (1991). The conservation andmanagement of eucalypt forest birds: resourcerequirements for nesting and foraging. InD. Lunney (Ed.), Conservation of Australia’sforest fauna. (pp. 25-34). Sydney: RoyalZoological Society of NSW.eucalypt open forest; management; forestryMany bird species have very idiosyncraticrequirements for nesting or foraging (e.g. spiderswebs, dead limbs). Fire (and logging) simplifiesforest environments to the detriment of many birdspecies. Loss of shrubby understorey may result ingreater nest predation or removal of species whichforage in particular shrub species. Effects of
logging may be compounded if they are associatedwith fire. Relatively inflexible imposed fireregimes may be particularly detrimental to birds.
Recher, H. F., & Christensen, P. E. (1981).Fire and the evolution of the Australian biota.In A. Keast (Ed.), Ecological biogeography inAustralia (pp. 135-162). The Hague: Junk.reviewBirds easily survive mild fires: in a controlled firein shrub woodland, birds avoided the flames andmoved into an adjacent unburnt heath, thenreturned to their original sites immediately afterthe fire had passed. However, there may beappreciable mortality in extensive wildfire.Because of their greater mobility and higherreproductive potential, birds respond to theeffects of fire more rapidly than do mammals.Changes are most pronounced for birds of theground and understorey vegetation. After mildfires, bird numbers decline initially but then (1-3years post-fire) quickly return to or exceedpre-fire levels. Reproductive output of birds maybe increased in the year following fire (probablydue to greater insect availability).
Recher, H. F., & Lim, L. (1990). A review ofcurrent ideas of the extinction, conservationand management of Australia’s terrestrialvertebrate fauna. Proceedings of the EcologicalSociety of Australia, 16, 287-301.review; managementSome birds (such as grass-wrens and scrub-birds)which were restricted to specialised habitats, havebeen affected by habitat change through alterationof fire regimes or grazing. Of 13 bird specieswhich declined in King’s Park over the previous60 years, 10 were ground-foragers. This changewas due to alteration of ground vegetation andlitter caused by spread of weeds and changed fireregimes. Frequent fuel-reduction burns in forestsare a concern, as they change understorey andlitter characteristics and lead to reduction inavailable energy. These fire-induced changes willhave long-term effects on many insectivorousbirds.
156
Australia’s Biodiveristy - Responses to Fire

Recher, H. F., & Serventy, D. L. (1991). Longterm changes in the relative abundance ofbirds in Kings Park, Perth, Western Australia.Conservation Biology, 5, 90-102.historic change; eucalypt open forest; managementBioregion: Swan Coastal PlainOver nearly 60 years of intermittent survey,change in ground cover (development of denseunderstorey, loss of eucalypt canopy andproliferation of weeds), associated with fireregime, has led to marked change in the birdassemblage of Kings Park. Nine of 16 specieswhich declined (Western Spinebill, Pallid Cuckoo,Grey Butcherbird, Laughing Kookaburra, GoldenBronze-cuckoo, Scarlet Robin, Indian Turtledove,Western Yellow Robin, Senegal Dove,White-tailed Black-cockatoo, Golden Whistler,Sacred Kingfisher, Tree Martin, Purple-crownedLorikeet, Whistling Kite, Willie Wagtail) wereground-foraging birds. Prescription burning fromthe 1930s to 1960s has exacerbated problems(spread of weeds and debilitation of remainingeucalypts).
Recher, H. F., Lunney, D., & Posamentier, H.(1975). A grand natural experiment - theNadgee wildfire. Australian Natural History,18, 150-163.wildfire; heath; eucalypt open forest; mortalityBioregion: South East CornerAround 70,000 ha (including almost all of Nadgeereserve) of heath and open forest were burnt in awildfire, the first major fire in the area for 40 years.The beach was littered with dead small birds. Oneyear after fire, all pre-fire bird species were presentexcept for Crescent Honeyeater, although somewere less common than previously. Multiplenesting was evident for some insectivorous birds.Management should ensure that fires do not burnthe whole reserve, and that a diversity of firesoccurs.
Recher, H. F., Allen, D., & Gowing, G. (1985).The impact of wildfire on birds in anintensively logged forest. In A. Keast, H. F. Recher, H. Ford, & D. Saunders (Eds.),Birds of eucalypt forests and woodlands: ecology,conservation, management (pp. 283-290).Chipping Norton: Surrey Beatty.eucalypt open forest; community; wildfire;management; forestryBioregion: South East CornerOne year after an intense and extensive wildfire,bird populations (and invertebrate numbers) werecensused in 12 burnt and unburnt sites, includingsome which had been logged, across a range offorest types. Invertebrates were generally moreabundant in foliage in burnt forests. Burnt andlogged forests had fewer bird species andindividuals than unburnt and unlogged forests.The combined effect of these two disturbanceswas greater than either alone. Brown Thornbill,Striated Thornbill, White-throated Treecreeper,Golden Whistler and Grey Fantail were lessabundant in burnt forests. Nectarivores (MuskLorikeet, Yellow-faced Honeyeater, Yellow-tuftedHoneyeater, White-naped Honeyeater),granivores (Diamond Dove, CommonBronzewing, Forest Bronzewing, Brown Quail,Painted Quail, Red-browed Finch, BeautifulFiretail) and Spotted Pardalote were generallymore abundant in burnt forest.
“Considering the frequency with which firesoccur in eucalypt forests and woodlands and theirdramatic impact on the landscape, there isremarkably little information about the effects offire on fauna or the long-term consequences ofburning on forest ecosystems." Generally fireeffects are most substantial on species occurring inlower substrates, and the timing of their responsesis related to vegetation recovery. Proposes amodel that nutrients released after fire promotegrowth of nutrient-rich foliage and hence anincrease in invertebrates (and thus insectivorousbirds). However, following this intense fire, therelatively sparse foliage in burnt areas may haveled to increased predation and to lower density ofinvertebrates per land area (cf. per area of foliage),and hence birds were relatively less common.
157
Fire and Australian birds

Recher, H. F., Davis, W. E., & Holmes, R. T.(1987a). Ecology of Brown and StriatedThornbills in forests of south-eastern NewSouth Wales, with comments on forestmanagement. Emu, 87, 1-13.Brown Thornbill; Striated Thornbill; eucalypt openforest; wildfire; forestry; managementBioregions: South east Corner; South easternHighlandsBrown and Striated Thornbills were moreabundant in unburnt open forest than in forestburnt 1 year previously, regardless of logginghistory.
Recher, H. F., Shields, J., Kavanagh, R., &Webb, G. (1987b). Retaining remnant matureforest for nature conservation at Eden, NewSouth Wales: a review of theory and practice.In D. A. Saunders, G. W. Arnold, A. A.Burbidge, & A. J. M. Hopkins (Eds.), Natureconservation: the role of remnants of nativevegetation (pp. 177-194). Chipping Norton:Surrey Beatty.eucalypt open forest; community; isolate; management;forestryBioregion: South Eastern CornerSix creek reserves (retained strips post-logging)and two sites in unlogged areas were censused forbirds over 8 years. The widest reserve was burntin the fourth year and four others were burnt inthe seventh year. The fire had little effect on birdnumbers in the widest strip, but total birdpopulation and number of forest birds declinedprecipitously in the year post-fire for the foursmaller burnt strips. The decline was only partlydue to fire, but may have been compounded bydrought. Open country birds increased in thestrips post-fire.
Recher, H. F., Hutchings, P. A., & Rosen, S.(1993). The biota of the Hawkesbury-Nepeancatchment: reconstruction and restoration.Australian Zoologist, 29, 3-41.historic changeBioregion: Sydney BasinDescribes environmental change since Europeancolonisation. Many bird species have declined,associated with a complex mix of factors includingchange in fire regimes. Increased fire frequenciesand more frequent wildfires of human origin areprobably the major causes of decline of RockWarbler and Ground Parrot.
Reid, J., & Fleming, M. (1992). Theconservation status of birds in arid Australia.Rangelands Journal, 14, 65-91.review; management; historic change; hummockgrasslands; malleeSuggests that fire regime and extent oflong-unburnt patches have changed in aridenvironments generally since Europeansettlement, though evidence is equivocal. There ismore convincing evidence of changed fire regimein hummock grassland deserts, to the possibledetriment of species such as Striated Grass-wren.However, pastoralism is regarded as a moreserious threat for most birds and mostenvironments (e.g. riparian habitats, tussockgrasslands and chenopod shrublands).
Reid, J. R. W., Kerle, J. A., & Morton, S. R.(1993a). Birds. In J. R. W. Reid, J. A. Kerle, &S. R. Morton (Eds.), Uluru fauna: thedistribution and abundance of vertebrate faunaof Uluru (Ayers Rock - Mount Olga) NationalPark, N.T. (pp. 36-57). Canberra: AustralianNational Parks and Wildlife Service.mallee; hummock grassland; Acacia woodland;community; wildfireBioregion: Great Sandy DesertRecently burnt areas are favoured by nomadicbirds (e.g. Budgerigar, White-winged Triller,Black Honeyeater, Crimson Chat, Zebra Finch,Masked Wood-swallow) and the rarer BandedWhiteface. A few species (e.g. StriatedGrass-wren, Rufous-crowned Emu-wren) arelargely restricted to mature hummock grasslands.By 10 years post-fire, mulga woodlands wererecolonised by most sedentary mulga birds,though much older woodlands support moremistletoe and hence greater numbers ofWhite-fronted Honeyeaters. Recommends apatch-burn strategy as advocated by Aboriginalresidents. Extensive fires probably have a morehomogenising effect. Rainfall after fires maysubstantially influence the rate of regrowth andbird responses.
158
Australia’s Biodiveristy - Responses to Fire

Reid, J. R. W., Kerle, J. A., & Morton, S. R.(1993b). Ecological patterns and processes ofimportance. In J. R. W. Reid, J. A. Kerle, & S.R. Morton (Eds.), Uluru fauna: the distributionand abundance of vertebrate fauna of Uluru(Ayers Rock - Mount Olga) National Park, N.T.(pp. 133-148). Canberra: Australian NationalParks and Wildlife Service.hummock grasslands; mallee; Acacia woodland;managementBioregion: Great Sandy Desert“Mulga is killed by fire, and given the importanceof this species to the associated rich birdcommunity, it would seem that the destruction ofextensive areas of mulga by fire would decimatebird populations, at least until the mulga hadregenerated to a certain age.” However, a mulgastand 11-14 years post-fire supported birds typicalof tall old mulga, as well as many nomadic speciescharacteristic of early regrowth. This may bebecause the site was close to old mulga.Management should aim to avoid very extensivefires, and fires in regrowth mulga not old enoughto have produced seed (c15 years). In spinifex,Striated Grasswren is confined to maturevegetation, while Banded Whiteface (and a rangeof nomadic species) are associated with recentlyburnt areas. Management require a fine-scalepatch-burn strategy, but with special attempt tomaintain old vegetation.
Reilly, P. (1991a). The effect of wildfire onbird populations in a Victorian coastal habitat.Emu, 91, 100-106.eucalypt open forest; wildfire; community;management; isolate; successionBioregion: South east Coastal PlainBird censuses were conducted in coastalvegetation from several months before a hot andextensive wildfire (with very few unburnt patches)to nearly six years post-fire. There were markeddifferences between species in responses. A groupof species were not or little affected (includingCrimson Rosella, Rufous Whistler, GreyShrike-thrush, Grey Fantail, Striated Pardaloteand Pied Currawong). Another group showed aninitial decline with recovery after one or moreyears (including Golden Whistler, SuperbFairy-wren, Striated Thornbill, CrescentHoneyeater, New Holland Honeyeater, SpottedPardalote for recovery after one year, and EasternYellow Robin, White-throated Treecreeper andEastern Spinebill for recovery after 2-3 years).
Seed-eating Bronzewings did not appear until 2years post-fire. Another group showed an initialincrease, generally followed by decrease (FlameRobin, Scarlet Robin, Australian Magpie, ravens).Breeding was recorded within the first yearpost-fire for Flame Robin, Buff-rumpedThornbill, Grey Currawong and AustralianMagpie. By two years post-fire all species presentpre-fire had been recorded, though abundancewas generally still slightly less. Some specieschanged foraging behaviours in burnt areas.Rufous Bristlebirds began returning 2 yearspost-fire, possibly from refuges in town gardens(where they had been unreported before the fire).Fire probably poses the greatest threat to thisspecies. “If fire is to be used as a managementtool, frequent pockets of unburnt country ofsufficient size are vital … The time of burningought also to be considered so that its impact onbreeding populations is minimal.”
Reilly, P. (1991b). The effect of wildfire onbush bird populations in six Victorian coastalhabitats. Corella, 15, 134-142.heath; thicket; eucalypt open forest; community;wildfire; management; successionBioregions: South eastern Highlands, South eastCoastal PlainSix sites across a range of habitats were monitoredat 6-monthly intervals from 14 months after asevere bushfire to 56 months post-fire. Nounburnt controls were available in the area. Forsome species, return showed considerablevariation between sites. Rufous Whistlers initiallydid not appear to be greatly affected by the fire,but became extremely rare or absent 32-56months post-fire. Southern Emu-wrensrecolonised heath 38 months and 56 monthspost-fire, possibly from a nucleus in a small patchthat was unburnt. Rufous Bristlebirds returned at14 months post-fire at one site (near an unburntpatch), but had not returned to another site (offormer occurrence) 7.5 years post-fire.White-throated Treecreepers recolonised sites at15, 27 and 39 months post-fire. Tawny-crownedHoneyeaters were absent until 56 monthspost-fire. For some species, colonisation was notfrom unburnt refuges to burnt margins to interiorof burnt areas, but rather they first appeared deepin the burnt forests. Fire effects are greatest forground-dwelling birds, especially those withrestricted distributions, narrow habitat ranges andlimited dispersal ability (e.g. Rufous Bristlebird,
159
Fire and Australian birds

Southern Emu-wren). Too frequent fires altervegetation and change bird assemblages. If fire isto be used as a management tool, pockets ofunburnt country are vital. Burning during thebreeding season should be avoided.
Ridpath, M. G. (1972). The effects of fire onfauna. In R. J. Hooper & M. Rowell (Eds.),Proceedings of the 1971 Tropical and Arid FireSymposium. (pp. 64-66). Darwin: NTGovernment Printer.historical change; review; Orange-footed Scrub-fowl;Wedge-tailed Eagle; Ground ParrotFire regimes change habitat configurations andrelative extent. For example, abandoned moundsof Orange-footed Scrubfowl indicate thatrainforests have declined over the last 8,000 yearsbecause of Aboriginal burning and/or climatechange. In arid environments, burning mayreduce the number of rabbits, leading to localdecline in Wedge-tailed Eagle population.Ground Parrots need heaths burnt every 4-5 years.
Ridpath, M. G. (1974). The ecologicalconsequences of fire for animal communities.In R. E. Fox (Ed.), Report on the use of fire innational parks and reserves. (pp. 48-53).Darwin: Department of the NorthernTerritory.review
Roberts, P. E. (1970). Some effects of abushfire on heathland birdlife. Proceedings ofthe Royal Zoological Society of New South Wales,89, 40-43.heath; wildfireBioregion: Sydney BasinUp to 2.5 years after fire, eight previously residentbird species disappeared or declined: EasternWhipbird, White-cheeked Honeyeater(occasional visits to flowering Xanthorrhoea sincefire), Little Wattlebird, Eastern Spinebill(occasionally feeding in regrowth heath),Variegated Fairy-wren, Chestnut-rumpedHeath-wren (one pair returned to regrowth heath1 year after fire), Brown Thornbill (declined inabundance in heath), Red-browed Finch(completely disappeared from the locality).Several birds from adjacent woodlandstemporarily moved into the heath immediately(and up to 3 months) after fire: Laughing
Kookaburra, Pied Currawong, Australian Magpie,Australian Raven, Magpie-lark, Eastern Rosella,Crimson Rosella. These fed on the seeds shedafter fire (the rosellas) or invertebrate andvertebrate prey killed or exposed by the fire.White-eared Honeyeaters increased in the burntheath from occasional visitors to nesting residentsabout 1 year post-fire.
Robertson, B. I. (1981). Seabird Islands: No.102. Chalky Island, Furneaux Group,Tasmania. Corella, 5, 49-52.seabirds; tussock grasslands; heathBioregion: FurneauxThe island was burnt by quail-shooters in 1968,affecting breeding seabirds.
Robinson, A. C., Casperson, K. D., & Copley,P. B. (1990). Breeding records of theMalleefowl (Leipoa ocellata) andScarlet-chested Parrot (Neophema splendida)within the Yellabinna Wilderness area, SouthAustralia. South Australian Ornithologist, 31,8-12.Scarlet-chested Parrot; mallee; hummock grassland.Bioregion: Great Victoria DesertScarlet-chested Parrot may show some associationwith young regrowth (3-5 years post-fire).
Robinson, D. (1992). Habitat use and foragingbehaviour of the Scarlet Robin and the FlameRobin at a site of breeding-season sympatry.Wildlife Research, 19, 377-395.Flame Robin; Scarlet RobinBioregion: South eastern HighlandsWildfire or fuel reduction burning may changethe relative competitive ability of these two similarspecies.
Robinson, D., & Woinarski, J. C. Z. (1992). Areview of records of the Northern Shrike-titFalcunculus frontatus whitei in northwesternAustralia. South Australian Ornithologist, 31,111-117.Crested Shrike-tit; tropical eucalypt open forest;threatened speciesCrested Shrike-tits favour old forests in temperateAustralia. Frequent burning of tropical eucalyptforests may be one reason for the rarity of thissubspecies, through loss of large trees. More data
160
Australia’s Biodiveristy - Responses to Fire

are needed on the long-terms effects of fire onarthropods and hence bark-foraging birds.
Rounsevell, D. E., & Woinarski, J. C. Z. (1983).Status and conservation of the Forty-spottedPardalote, Pardalotus quadragintus (Aves:Pardalotidae). Australian Wildlife Research, 10,343-349.Forty-spotted Pardalote; threatened species; eucalyptopen forest; isolateBioregion: FreycinetWildfire may threaten all populations ofForty-spotted Pardalote. One small isolatedpopulation may have disappeared after fire.However, population numbers at another sitewere similar 2 years post-fire to pre-fire levels(possibly because of proximity of unburnt areaswith relatively large populations). Fuel-reductionburns are probably not detrimental.
Rowley, I. (1987). Conservation of thePurple-crowned Fairy-wren Malurus coronatusin northern Australia. World Wildlife Fund(Australia).Purple-crowned Fairy-wren; riparian vegetation;threatened species; pandanus; tussock grassland; tropicaleucalypt savanna woodlandBioregions: Victoria Bonaparte; Ord-VictoriaPlains; Central Kimberley; North Kimberley;Gulf Fall and Uplands; Gulf CoastalFire may destroy or degrade some of the riparianstrips on which this species depends, though themain threat is degradation of this habitat by cattle.
Rowley, I. (1993). The Purple-crownedFairy-wren Malurus coronatus. I. History,distribution and present status. Emu, 93,220-234.Purple-crowned Fairy-wren; pandanus; tussockgrassland; threatened species; tropical eucalypt savannawoodlandBioregions: Victoria Bonaparte; Ord-VictoriaPlains; Central Kimberley; North Kimberley;Gulf Fall and Uplands; Gulf CoastalThe Purple-crowned Fairy-wren is found inriparian vegetation (Pandanus and cane-grasses) innorthern Australia. Heavy grazing by cattle andburning by pastoralists removes both shelter andforaging substrate for this species, forcing them toabandon this habitat at critical times of the year,leading to local declines and extinction.
Rowley, I., & Brooker, M. (1987). Theresponse of a small insectivorous bird to firein heathlands. In D. A. Saunders, G. W.Arnold, A. A. Burbidge, & A. J. M. Hopkins(Eds.), Nature Conservation: the role ofremnants of native vegetation (pp. 211-218).Chipping Norton: Surrey Beatty.Splendid Fairy-wren; heath; wildfire; isolate;managementBioregion: Jarrah ForestSix fires (one of which was intense) burnt parts ofthe study area over a 12 year study ofcolour-banded Splendid Fairy-wrens. In mostcases, unburnt patches were left. But in bothterritories that were completely burnt out, most ofthe previous occupiers disappeared (and werepresumed to have perished). The incidence ofparasitism in the years following fire was low.With the exception of adult survival, all measuresof density, productivity and survival increasedduring fire-free periods. The major changeoccurred in the fourth year post-fire and wassustained thereafter. High adult survival allowspopulation maintenance despite several years oflow productivity post-fire. However, frequent fireseliminated some populations at the site, withsubsequent restoration of territories arising fromrecruitment from beyond the site.
This study also reports short-term (9months) response to an intense wildfire whichconsumed 95% of the vegetation. The populationsurvived the very hot (>600oC at 10cm aboveground) fire well, showed remarkable site tenacity,and showed similar or even reduced mortality inthe months post-fire than in previous years. Thebetter short-term survival after a hot fire thanafter cooler burns may have been due to thetiming of the hot fire, which was several monthsafter breeding season.
Frequent burns had a deleterious effect onthis population. If the site had been a remnant,the Fairy-wrens may well have been eliminated bythis fire regime. Optimum density may bemaintained with at least 12 year intervals betweenfires, which should not be during the breedingseason.
161
Fire and Australian birds

Rowley, I., & Russell, E. (1993). ThePurple-crowned Fairy-wren Maluruscoronatus. II. Breeding biology, socialorganisation, demography and management.Emu, 93, 235-251.Purple-crowned Fairy-wren; tussock grassland;pandanus; management; threatened speciesBioregions: Victoria Bonaparte; Ord-VictoriaPlains; Central Kimberley; North Kimberley;Gulf Fall and Uplands; Gulf CoastalPurple-crowned Fairy-wrens require riparianhabitat which is easily damaged by stock or fire.This habitat should be protected from fire at alltimes.
Rowley, I., & Russell, E. (1995). The breedingbiology of the White-winged Fairy-wrenMalurus leucopterus leuconotus in a WesternAustralian coastal heathland. Emu, 95,175-184.White-winged Fairy-wren; heath; wildfireBioregion: Swan Coastal PlainAll White-winged Fairy-wrens from a 32ha studysite (77 adults banded in the 4 years before fire)could not be relocated shortly after a wildfire, andonly one wren was seen there 3 months after thefire.
Rowley, I., Russell, E., Brown, R., & Brown,M. (1988). The ecology and breeding biologyof the Red-winged Fairy-wren Maluruselegans. Emu, 88, 161-176.Red-winged Fairy-wren; control burning; eucalyptopen forestBioregion: WarrenIn the long term, fuel-reduction burns must havesevere effects on populations of Red-wingedFairy-wren, because nearly half of the nest sites inthis study were in “dead brush”, the fuel that suchburns seek to reduce. This species occurs in a verystable environment and has very low reproductiveoutput: it may be a very poor recoloniser.
Russell, E. M., & Rowley, I. (1993).Demography of the cooperatively breedingSplendid Fairy-wren, Malurus splendens(Maluridae). Australian Journal of Zoology, 41,475-505.Splendid Fairy-wren; wildfireBioregion: Swan Coastal PlainStudied a population for 18 years, which included
a number of minor wildfires and one which burnt95% of the study area. Annual survival decreasedin the two years after fire. Females laid moreclutches when predation or brood-parasitism washigh, particularly in the 2 years post-fire, butproductivity was low for several years post-fire.Territories were not vacated initially after fire, butby 3-5 years post-fire some territories were vacantand group sizes decreased. Population declineoccurred 3-5 years post-fire due to decreasedreproductive output and replacement of olderbreeding birds by less productive novices. Firedirectly affected natality and juvenile survival andindirectly affected population density, agestructure, sex ratio and group composition.
Russell-Smith, J. (1985). A record of change:studies of Holocene vegetation history in theSouth Alligator Region, Northern Territory.Proceedings of the Ecological Society ofAustralia., 13, 191-202.Orange-footed Scrubfowl; historic change; rainforestBioregion: Top End CoastalHistoric changes in fire regime have led tocontraction of monsoon rainforests, as evidencedby old scrubfowl mounds occurring in eucalyptopen forests.
Russell-Smith, J. (1995). Fire management. InT. Press, D. Lea, A. Webb, & A. Graham(Eds.), Kakadu: natural and cultural heritageand management (pp. 217-237). Darwin:Australian Nature Conservation Agency andNorth Australia Research Unit.management; review; tropical eucalypt open forest;tussock grassland; Aboriginal knowledge; AboriginalburningBioregions: Pine Creek - Arnhem; Top EndCoastalReviews burning regimes used traditionally byAboriginal people and currently by parkmanagers; and some studies of the effects of fireon birds in this region.
Saunders, D. A. (1979). The availability of treehollows for use as nest sites by White-tailedBlack Cockatoos. Australian Wildlife Research,6, 202-216.hollows; White-tailed Black-cockatoo
162
Australia’s Biodiveristy - Responses to Fire

Saunders, D. A. (1985). Human impact: theresponse of forest and woodland birdcommunities. Whither the future? Asynthesis. In A. Keast, H. F. Recher, H. Ford,& D. Saunders (Eds.), Birds of eucalypt forestsand woodlands: ecology, conservation,management (pp. 355-357). Chipping Norton:Surrey Beatty.review; eucalypt open forest; managementMuch more research is needed on the role of firein bird conservation and management. “Minorchanges in the fire regime may be as damaging asthe widespread clearing for agriculture.”
Saunders, D. A., Smith, G. T., & Rowley, I.(1982). The availability and dimensions of treehollows that provide nest sites for cockatoos(Psittaciformes) in Western Australia.Australian Wildlife Research, 9, 541-556.hollowsThe smallest trees suitable for nesting by any of 5species of cockatoos considered was at least 130years old. Such trees are becoming scarcer due tovegetation clearing and prevention ofregeneration (due to stock trampling or grazing).
Schodde, R. (1982). The Fairy-wrens. Amonograph of the Maluridae. Melbourne:Lansdowne.Mallee Emu-wren; Carpentarian Grass-wren;threatened species; management; hummock grasslands;review.The Mallee Emu-wren and CarpentarianGrass-wren are probably threatened by fire. Bothmay recover very slowly after fire has consumedtheir highly flammable hummock grasslandhabitat. This may be especially threatening wherepopulations are highly fragmented. SouthernEmu-wren and Striated Grass-wren may also beadversely affected by wildfire, though this doesnot yet threaten the species existence. Both maytake many years to recolonise. The SouthernEmu-wren is reported to fly well before the firefront.
Schodde, R., Mason, I. J., & Wood, J. T.(1993). Geographical differentiation in theGlossy Black-Cockatoo Calyptorhynchuslathami (Temminck) and its history. Emu, 93,156-166.Glossy Black-Cockatoo; threatened species;management; hollowsManagement for the Glossy Black-Cockatooshould include developing a fire regime thatprevents severe wildfires (and hence eliminationof food sources), and maintaining availability ofhollows.
Scotts, D. (1994). Sustaining sensitive wildlifewithin temperate forest landscapes: regionalsystems of retained habitat as a planningframework. In T. W. Norton & S. R. Dovers(Eds.), Ecology and sustainability of southerntemperate ecosystems (pp. 85-106). Canberra:CSIRO.old-growth; forestry; management; eucalypt open forestMainly concerned with management of forestry,but provides a review of old-growth characteristicsand the association of birds with these.
Scotts, D. J. (1991). Old-growth forests: theirecological characteristics and value toforest-dependent vertebrate fauna ofsouth-east Australia. In D. Lunney (Ed.),Conservation of Australia’s forest fauna (pp.147-159). Sydney: Royal Zoological Society ofNSW.eucalypt open forest; old-growth; forestry; hollowsBioregions: South Eastern Highlands; South EastCornerOld-growth forests are characterised by abundantlitter, logs and hollows, and distinctive floristiccomposition. 17 bird species are listed as findingoptimum habitat in old-growth forests. Theseinclude hollow-nesting species, honeyeaters,trunk-gleaning insectivores, frugivores and someinsectivores of the canopy foliage.
163
Fire and Australian birds

Silveira, C. E. (1993). Recovery plan forthreatened mallee birds - addressing fire regimes.Royal Australasian Ornithologists Union.mallee; heath; threatened species; management; review;Malleefowl; Pink Cockatoo; Regent Parrot; MalleeEmu-wren; Striated Grass-wren; Black-eared Miner;Slender-billed Thornbill; Western Whipbird; Red-loredWhistler.Bioregions: Murray-Darling Depression; MalleeProvides a review of the observed association ofthreatened mallee birds (Malleefowl, PinkCockatoo, Regent Parrot, Mallee Emu-wren,Striated Grass-wren, Black-eared Miner,Slender-billed Thornbill, Western Whipbird andRed-lored Whistler) with fire, and research andmanagement requirements. For the Malleefowl, ahigh frequency of fire is a confirmed threat inmallee of eastern Australia, but has not beenestablished in western mallee. For the PinkCockatoo, primary threats are the loss of suitablehollows through natural attrition and theprevention of formation of new hollows due toincreased frequency of fire. The MalleeEmu-wren and Striated Grass-wren are absent inmallee with hummock grassland understorey for5-6 years post-fire, but persist then until thevegetation is long-unburnt. Recolonisation maybe a problem for these species with poordispersive ability following large-scale fires whicheliminate populations. Black-eared Minersapparently prefer long-unburnt mallee forbreeding, though may feed in younger regrowth.In southwestern Australia, Western Whipbirdsprefer mallee-heath older than 14 years, and mayprefer much older vegetation.
Silveira, C. E. (1995). The Black-eared Miner.Australian Bird Watcher, 16, 96-109.Black-eared Miner; threatened species; malleeBioregion: Murray-Darling DepressionIn contrast to previous claims (McLaughlin 1992)that the Black-eared Miner is restricted to malleeat least 55 years post-fire, this paper reportsforaging by Black-eared Miners in mallee of 3-11years post-fire (though these birds apparentlynested in adjacent long-unburnt mallee).
Skira, I. J., & Brothers, N. P. (1988a). SeabirdIslands: No. 183. Little Green Island,Furneaux Group, Tasmania. Corella, 12,80-81.seabirds; tussock grasslandBioregion: FurneauxThe island is regularly fired, and burning andgrazing by sheep have caused much deteriorationof vegetation and soil, affecting breedingshearwaters.
Skira, I. J., & Brothers, N. P. (1988b). SeabirdIslands: No. 184. Great Dog Island, FurneauxGroup, Tasmania. Corella, 12, 82-84.seabirds; tussock grasslandBioregion: FurneauxFrequent burning has caused fewer areas to beused by breeding shearwaters, however burning isnow less frequent due to management control bystate conservation agency.
Smith, G. T. (1977). The effect ofenvironmental change on six rare birds. Emu,77, 173-179.review; historic change; Aboriginal burning; NoisyScrub-bird; Rufous Scrub-bird; Western Bristlebird,Eastern Bristlebird, Rufous Bristlebird; WesternWhipbird; threatened species; managementExamines status, habitat requirements and historyof Noisy Scrub-bird, Rufous Scrub-bird, WesternBristlebird, Eastern Bristlebird, Rufous Bristlebirdand Western Whipbird. The Scrub-birds andBristlebirds are Tertiary relicts, more common inclosed forests of the Tertiary, and now largelyrestricted to very narrow specialised habitats.Aboriginal entry to Australia had a major impacton the fauna during the Pleistocene and Recent by“prolonged, constant and widespread use of fire”.One consequence was a possible increase in thearea of heath (to the advantage of severalheath-favouring birds, including WesternBristlebird and Eastern Bristlebird). However,burning regime of heaths became much morefrequent with European colonisation and led tothe decline of bristlebirds and scrub-birds.Western Bristlebirds require heath of at least8-10 years post-fire before habitat is suitable. ForNoisy Scrub-birds fire intervals of <5 years or>30 years render heath/thicket unsuitable.
164
Australia’s Biodiveristy - Responses to Fire

Smith, G. T. (1979a). The Noisy Scrub-bird.In M. J. Tyler (Ed.), The status of endangeredAustralasian wildlife (pp. 117-121). Adelaide:Royal Zoological Society of South Australia.Noisy Scrub-bird; heath; thicket; isolate; threatenedspecies; eucalypt open forest; management; historicchange; Aboriginal burningBioregion: Jarrah ForestAboriginal use of fire probably had little impacton Noisy Scrub-bird habitat (as such fires wereprobably small and lit after the breeding season).After European colonisation, heaths were burntevery 2-3 years to provide grass for cattle, whileattempts made to exclude fire in forests probablyresulted in higher frequency of occasional hotwildfires. These changes would have led to thedecline and local extinction of Noisy Scrub-birds,and these losses would have further fragmentedpopulations and made recolonisations after firesless likely. Recent increases are due to absence offire. However, lack of fire may allow successionalchanges that render gullies unsuitable forScrub-birds (this has already happened at twosmall sites).
Smith, G. T. (1979b). The status of Australianparrots. In M. J. Tyler (Ed.), The status ofendangered Australasian wildlife (pp. 101-108).Adelaide: Royal Zoological Society of SouthAustralia.Ground Parrot; review; heath; threatened species;managementToo frequent burning renders heath unsuitable forGround Parrot, and heath over-protected fromfire also becomes unsuitable as it becomes too talland dense. Management needs to mosaic burn at8-10 year intervals.
Smith, G. T. (1985a). Fire effects onpopulations of the Noisy Scrub-bird(Atrichornis clamosus), Western Bristle-bird(Dasyornis longirostris) and WesternWhip-bird (Psophodes nigrogularis). In J. R.Ford (Ed.), Fire ecology and management inWestern Australian ecosystems (pp. 95-102).Perth: Western Australian Institute ofTechnology.Noisy Scrub-bird; Western Bristlebird; WesternWhipbird; heath; thicket; eucalypt open forest;management; historic change; threatened species;Aboriginal burningBioregions: Esperance Plains; Warren; SwanCoastal Plain; Jarrah ForestThere have been few studies of the effects of fireon birds, and most are opportunistic andshort-term. Western Whipbird, WesternBristlebird and Noisy Scrub-birds are all poordispersers. Change in fire regime from that usedby Aborigines to that used by settlers resulted insevere contraction of range. These three speciessurvive at Two Peoples Bay, largely throughtopographic protection from fire. Recent firecontrol in this area has led to the expansion ofpopulations of all three species. Vegetation has tobe 4-10 years post-fire before being suitable forNoisy Scrub-birds (being faster in wet gullieswhere dominant eucalypts have not beendestroyed). Reproductive output is low, hencerecruitment after fire may be very gradual. Wetheath may be suitable for Western Bristlebirds 3years post-fire and dry heath by 6-10 yearspost-fire, though grazing pressure from GreyKangaroos (especially if burnt areas are limited)may extend these periods. For WesternWhipbirds, minimum periods before vegetationwas suitable were 4-6 and 7 years post-fire. Inheath, there is a bird succession from Richard’sPipit to Striated Field-wren to WesternBristlebird to Western Whipbird. Thelong-unburnt part of this succession is fairlyspeculative as there are few very old heaths. Some>45 year post-fire heaths retain Bristlebirds, but atdensities less than those in 20 year old heaths.Less is known of later succession in the loweucalypt forests used by Noisy Scrub-birds. A fireinterval of at least 20 years (and possibly of theorder of 50 years) should be maintained, andaccompanied by considered monitoring.
165
Fire and Australian birds

Smith, G. T. (1985b). The Noisy Scrub-birdAtrichornis clamosus. Does it’s past suggest afuture? In A. Keast, H. F. Recher, H. Ford, &D. Saunders (Eds.), Birds of eucalypt forests andwoodlands: ecology, conservation, management(pp. 301-308). Chipping Norton: SurreyBeatty.Noisy Scrub-bird; threatened species; eucalypt openforest; wetlands; heath; management; Aboriginalburning; historic changeBioregions: Jarrah Forest; WarrenCompares Aboriginal fire regimes (small, patchy)with settler fire regimes (frequent in wet heathsand swamp margins; hot summer fires and morefrequent occurrence of bushfires in open forest) inthe restricted habitat of Noisy Scrub-bird.Topographic protection has resulted in lowfrequency of fire at one site and hence the survivalof one relict population. Minimum age ofvegetation to be suitable is known, but there isalso indication that long-term fire exclusion mayresult in reduction of habitat suitability.
Smith, G. T. (1985c). Population and habitatselection of the Noisy Scrub-bird, Atrichornisclamosus, 1962-83. Australian WildlifeResearch, 12, 479-485.Noisy Scrub-bird; heath; thicket; eucalypt open forest;threatened speciesBioregion: Jarrah ForestExclusion of fire has led to substantial vegetationchange and marked increases in populations ofNoisy Scrub-bird. Six years after a hot fire,regeneration was suitable in some, if not all, areasfor Noisy Scrub-bird. But the first male torecolonise was not recorded until 9 years post-fire,suggesting problems of dispersal from unburntareas. Nothing is known about how longvegetation remains suitable, but it is likely to begreater than 40 years.
Smith, G. T. (1987a). The changingenvironment for birds in the south-west ofWestern Australia; some managementimplications. In D. A. Saunders, G. W. Arnold,A. A. Burbidge, & A. J. M. Hopkins (Eds.),Nature Conservation: The role of remnants ofnative vegetation (pp. 269-277). ChippingNorton: Surrey Beatty.historic change; eucalypt open forest; heath; thicket;Noisy Scrub-bird; Western Whipbird; WesternBristlebird; management; threatened species;Aboriginal burningBioregions: Esperance Plains; Jarrah Forest;WarrenChange from Aboriginal burning regimes to thoseassociated with Europeans led to epidemic ofintense fires in the early years of settlement. Incoastal areas, early pastoralists fired the heaths every2-3 years. Clearing and fragmentation have left theremnants more vulnerable to catastrophic fire.Changes in burning regime are thought to be themain cause of decline for Noisy Scrub-bird,Western Bristlebird, Western Whipbird andpossibly Ground Parrot; and probably for theregional extinction of Rufous Bristlebird. Discussesmanagement of Two Peoples Bay for the threatenedNoisy Scrub-bird, Western Bristlebird and WesternWhipbird. Recent large wildfires in nearby areasseverely reduced or wiped out populations ofWestern Bristlebird. Notes the importance ofnatural firebreaks (rocky hills, dissescted gullies) inproviding protection from fire for these remnantpopulations. Recolonisation period post-firedepends on a number of factors (edaphic, proximityof source populations, refuge areas). Fine scalehabitat manipulation of old areas may maintaintheir suitability for Noisy Scrub-birds. Older heaths(45 years post-fire) may have decreased productivityand hence lowered suitability for WesternWhipbirds and Western Bristlebirds. Fire intervalsof <10 years will lead to local decline and possibleextinction for these 3 species. Fire intervals of >50years may also be deleterious. Fire managementshould maintain firebreaks and prevent one fireburning the whole reserve. Long intervals betweenfires will disadvantage some other early successionalbirds (e.g. Richard’s Pipit, Striated Fieldwren).Long-term fire exclusion may also change therelative extent of heaths and thickets to thedisadvantage of one or other of the threatenedspecies. The longevity of many Australian birdsmeans that many which survive fire may live longenough to recolonise. Describes three requirements
166
Australia’s Biodiveristy - Responses to Fire

for management: detailed information on habitatrequirements, data on type and rate of vegetationchange post-fire, and information on therelationship of birds with fire regime.
Smith, G. T. (1987b). Observations on thebiology of the Western Bristlebird Dasyornislongirostris. Emu, 87, 111-118.Western Bristlebird; heath; thicket; threatened species;historic changeBioregions: Esperance Plains; Jarrah ForestWestern Bristlebirds have disappeared from muchof their range over the last 100 years, due to pastpractice of burning the coastal heaths and thicketsevery two or three years to provide cattle feed.Clearing and draining swamps have alsocontrubuted to the decline. Home ranges wereestablished 9 years post-fire (adjacent to gullies)and 11-14 years post-fire (drier areas) at sitesrelatively distant from existing populations, and4-6 years post-fire at sites adjacent to or within2km of existing populations. The preferred shortclosed heaths are suitable 3 years after fire (in wetareas) or 6-10 years after fire (drier areas). Oldestsuitable age is not known, but birds are present in45 year heath (although less abundant than in 20year old heath). While old heaths are probablystructurally suitable, their productivity may havedeclined. Thickets and more swampy vegetationmay be less vulnerable to fire and provideimportant refuges after fire.
Smith, G. T. (1991). Ecology of the WesternWhipbird Psophodes nigrogularis in WesternAustralia. Emu, 91, 145-157.Western Whipbird; mallee; heath; thicketBioregions: Esperance Plains; Jarrah ForestWestern Whipbirds in southwestern Australia aremost abundant in dense coastal thickets.Population increases at some sites are due toabsence of fire (up to at least 50 years). In generalfire intervals of less than 10 years will lead, or hasled, to extinction of local populations. Notes thatfire effects will differ according to vegetation type(for example between Victorian mallee andsouthwestern Australian heath).
Smith, G. T., & Forrester, R. I. (1981). Thestatus of the Noisy Scrub-bird Atrichornisclamosus. Biological Conservation, 19, 239-254.Noisy Scrub-bird; heath; eucalypt open forest; thicket;threatened species; managementBioregion: Jarrah ForestFire suppression at remaining colony sites hashelped to increase the numbers of NoisyScrub-birds.
Smith, G. T., & Robinson, F. N. (1976). TheNoisy Scrub-bird: an interim report. Emu, 76,37-42.Noisy Scrub-bird; eucalypt open forest; threatenedspecies; thicket; heath; historic changeBioregion: Jarrah ForestThe dramatic disappearance of the NoisyScrub-bird soon after European colonisationcertainly resulted from frequent devastating firesand the clearing and burning of eucalypt forests.However total fire exclusion may eventuallyrender habitat unsuitable.
Smith, L. H. (1994). A critical analysis of thefactors responsible for the decline of theSuperb Lyrebird Menura novaehollandiae inSherbrooke Forest, Victoria. Australian BirdWatcher, 15, 238-249.Superb Lyrebird; eucalypt open forest; control burning;managementBioregion: South eastern HighlandsPopulation of Superb Lyrebirds has declined,partly through fox predation and throughunderstorey change (reduction in open leaf litterand increase in grass cover), during a period whenformer frequent burning was replaced by fireexclusion. A firebreak maintained by burning andslashing was abandoned by Lyrebirds when itbecame overgrown following cessation of burning.
Smith, P. (1989). Changes in a forest birdcommunity during a period of fire anddrought near Bega, New South Wales.Australian Journal of Ecology, 14, 41-54.eucalypt open forest; community; rainforest; wildfire;successionBioregion: South East CornerA forest bird community was monitored for 3years, during which it was partly burnt by wildfireand affected by severe drought. The fire affectedmainly the undergrowth, especially on ridges:
167
Fire and Australian birds

gullies were mainly left unburnt. Changes in thebird community were limited. Differences inspecies were restricted to rare visitors. Thedistribution of insectivorous birds contracted (andthe species declined) in the year following fire;nectarivorous and frugivorous species expandedand became more abundant (due to prolificflowering and fruiting in the second year of thestudy, possibly because of the fire). 10 species(Eastern Yellow Robin, Crested Shrike-tit,Golden Whistler, Rufous Fantail, SuperbFairy-wren, White-browed Scrubwren, BrownGerygone, Brown Thornbill, Lewin’s Honeyeater,Silvereye) declined in burnt areas in the 2 yearsfollowing fire: all, other than Crested Shrike-tit,were birds which feed, nest or shelter in denseshrubby understorey. For seven resident species,colour-banding allowed assessment of changes interritory. For 6 of these species (especially BrownThornbill), the number of territories declinedpost-fire, with loss of colour-banded birds fromthe site and/or contraction to unburnt patches. Inthe year post-fire many species bred at atypicaltimes, possibly related to flush of resources.Recovery of populations had not reached pre-firelevels two years after fire, which is slower thanthat reported in other studies. This may have beenbecause of drought, and suggests that a range ofvariables may affect recolonisation. The resilienceof the community was related to the role ofunburnt areas, particularly wet gullies.
Smith, P. J., Pressey, R. L., & Smith, J. E.(1994). Birds of particular conservationconcern in the Western Division of NewSouth Wales. Biological Conservation, 69,315-338.threatened species; review; mallee; eucalypt woodland;Acacia woodland; managementBioregions: Murray Darling Depression; BrokenHill Complex; Mulga Lands; Channel Country;Simpson-Strzelecki DunefieldsReviews change in abundance and threats for birdspecies in the Western Division of New SouthWales. The most frequently-mentioned threatsare overgrazing, introduced predators, clearingand altered fire regimes (in that order). Fire is athreat particularly to mallee birds, especially thefrequent occurrence of extensive wildfires. Alteredfire regime is listed as a threat for 15 taxa(Black-eared Miner, Malleefowl, Red-loredWhistler, Regent Parrot, Scarlet-chested Parrot,Striated Grass-wren, Glossy Black-cockatoo,
Turquoise Parrot, Southern Scrub-robin, ShyHeath-wren, Chestnut Quail-thrush,Black-winged Currawong, Gilbert’s Whistler,Jacky Winter and Golden Whistler).
Smith, P. J., Smith, J. E., & Pressey, R. L.(1995). Birds of particular conservation concernin the Western Division of New South Wales:distribution, habitats and threats. NSWNational Parks and Wildlife Service.threatened species; review; mallee; eucalypt woodland;Acacia woodland; hummock grassland; tussockgrassland; managementBioregions: Murray-Darling Depression; BrokenHill Complex; Mulga Lands; Channel Country;Simpson-Strzelecki Dunefields.Reviews the threatening processes for threatenedbird taxa in the Western Division of NSW. Fire isone of the major threats, particularly for malleebirds.
Smith, S., & Baker-Gabb, D. (1993). RufousBristlebird Dasyornis broadbenti. Departmentof Conservation and Natural Resources.Rufous Bristlebird; threatened species; thicket; controlburning; wildfire.Bioregions: Naracoorte Coastal Plain; South eastCoastal PlainReviews information on the species (in Victoria).Wildfire or inappropriate (control) burningregimes are a threat throughout its range.Management requires determination andimplementation of a preferred fire regime,however this may conflict with protection ofproperty (or current legislation).
Sonter, C. (1984). The Yellow-rumpedPardalote - a declining bird in Sunraysia?Australian Bird Watcher, 10, 234-235.Yellow-rumped Pardalote; wildfire; mallee; eucalyptwoodlandBioregion: Murray-Darling DepressionCombination of bushfires, clearing and droughtmay have led to the decline of this species.
168
Australia’s Biodiveristy - Responses to Fire

Specht, R. L. (1981). Responses to fires inheathlands and related shrublands. In A. M.Gill, R. H. Groves, & I. R. Noble (Eds.), Fireand the Australian biota (pp. 395-415).Canberra: Australian Academy of Science.heath; reviewYellow-tailed Black-cockatoos visited a heathlandsoon after fire to feed from grubs attracted to thefire-induced flowering of Xanthorrhoea.Honeyeaters recolonised when woody plantsreplaced the more herbaceous short-lived plants.Black cockatoos returned to mature heaths to tearapart Banksia flowers.
Stanton, J. P. (1992). J.P. Thomson oration.The neglected lands: recent changes in theecosystems of Cape York Peninsula and thechallenge of their management. Journal of theQueensland Geographical Society, 7, 1-18.Cassowary; management; threatened species; tropicaleucalypt open forest; rainforest; hollows.Bioregion: Cape York PeninsulaThe maintenance of the preferred habitat of theCassowary (ecotone between rainforest and weteucalypt open forest) requires a regime includingfrequent fire. Preservation of trees bearinghollows suitable for nesting birds may requirecontrol burning at times when fuel moisture ishigh.
Stanton, P. (1995). A tropical Queenslandperspective. In D. B. Rose (Ed.), Country inflames. Proceedings of the 1994 symposium onbiodiversity and fire in North Australia (pp.71-76). Canberra: Department of theEnvironment, Sport and Territories, andNorth Australia Research Unit.tropical eucalypt open forest; rainforest; tussockgrassland; management; threatened species;Golden-shouldered Parrot; Palm Cockatoo; Cassowary.Bioregion: Cape York PeninsulaPresent fire regime is leading to expansion ofrainforest at the expense of wet eucalypt forests, tothe detriment of Palm Cockatoo and Cassowary.It is also leading to invasion of grassy flats byMelaleuca, threatening the Golden-shoulderedParrot.
Starks, J. (1987). The status and distribution ofthe Black-eared Miner (Manorina melanotis) inVictoria. Technical report no.49. Arthur RylahInsitute of Environmental Research.Black-eared Miner; mallee; threatened speciesBioregion: Murray Darling DepressionThe endangered Black-eared Miner is associatedwith long-unburnt mallee, and the limited extentof such old vegetation is a factor in its threatenedstatus.
Stephens, S. (1992). Endangered species andcommunities and threatening processes in theMurray Mallee. Australian National Parks andWildlife Service.mallee; review; management; threatened speciesBioregion: Murray Darling DepressionAltered fire regime has contributed to the declineof 8 (sic) bird species (Beautiful Firetail,Black-eared Miner, Chestnut Quail-thrush,Chestnut-rumped Heath-wren, CrestedShrike-tit, King Quail, Mallee Emu-wren,Mallee-fowl, Night Parrot, Orange-belliedParrot, Red-lored Whistler, Red-tailedBlack-cockatoo, Rufous Bristlebird and RufousField-wren) in the Murray Mallee (thefourth-ranking threat after overgrazing,introduced predators and clearing). Includesdiscussion of management and research optionsand priorities.
Stocker, G. C. (1971). The age of charcoalfrom old jungle fowl nests and vegetationchange on Melville Island. Search, 2, 28-30.Orange-footed Scrubfowl; charcoal; historic change;rainforest; Aboriginal burningBioregion: Top End CoastalRecent changes in fire regimes have led to retreatof rainforests, marked by the occurrence ofabandoned mounds of scrubfowl in areas whichare now eucalypt open forests.
Stokes, T. (1975). The effect of a bushfire onthe banding of Flame Robins in theBrindabella Ranges. Australian Bird Bander,13, 75-76.Flame Robin; wildfire; eucalypt open forestBioregion: South Eastern HighlandsIn 28 months following hot wildfire in montaneeucalypt forest, Flame Robins were far moreabundant in burnt than unburnt areas.
169
Fire and Australian birds

Suckling, G. C., & MacFarlane, M. A. (1983).The effects of fire on fauna - a review. In E.Ealey (Ed.), Fighting fire with fire. (pp.107-128). Melbourne: Monash University.review; eucalypt open forest; wildfire; control burning;managementFew studies of effects of fire on fauna other thanmammals. Summarises existing data for birds, interms of effect of fire intensity, frequency andseason. Few birds are killed by low intensity fires,but many may subsequently die through predationor starvation. In contrast, immediate mortalitymay be high with intense fires. Fire intensityaffects vegetation structure, which has moreimportant long-term implications for birds. Morerefuge areas may be left by low intensity fires.Some birds (e.g. Flame Robin) prefer severelyburnt areas to unburnt forests. Return of birdcommunities is probably quicker afterlow-intensity fires. In open forests, understoreymay require more frequent fires than overstorey,and the number of birds in the understorey maydecline in old forests. Ground-feeding birds (e.g.Superb Lyrebird, Superb Fairy-wren, BassianThrush, Eastern Yellow Robin) may requireperiodic fires. Other species (e.g. NoisyScrub-bird) cannot survive frequent fire. Thereare no data on the time of year at which birds aremost susceptible to fire, though effects may begreatest during the breeding season.
Swanson, N. M. (1976). Seabird Islands: No.32. Mutton Bird Island, New South Wales.Australian Bird Bander, 14, 88-91.seabirds; tussock grasslandBioregion: NSW North CoastFires have led to erosion and spread of exoticgrasses. Nesting birds took about 4 years to fullyre-use a site that had been severely burnt.
Tarr, H. E. (1965). The Mallee-Fowl inWyperfeld National Park. Australian BirdWatcher, 2, 140-144.Malleefowl; threatened species.Bioregion: Murray-Darling DepressionMallee may be unsuitable for Malleefowl for up to10-20 years post-fire.
Taylor, R. J. (1991). Fauna conservation inproduction forests in Tasmania. Hobart:Forestry Commission Tasmania.review; forestry; eucalypt open forest; control burning;managementReviews impacts of control burning and wildfireon birds (and other fauna) in eucalypt forests. Ingeneral, effects of mild control fires areshort-term and minor, though long-term changesin understorey as a result of frequent control firesmay lead to more substantial changes in the birdassemblage. Regeneration of wet sclerophylleucalypt forests may require hot burns. Speciessuch as Grey Goshawk and hollow-nesting speciesrequire, or are more abundant in, old-growthforests.
Tidemann, S. C. (1990). Relationshipsbetween finches and pastoral practices innorthern Australia. In J. Pinowski & J. D.Summers-Smith (Eds.), Granivorous birds andagriculture (pp. 305-315). Warsaw:PWN-Polish Scientific Publishers.tropical eucalypt savanna woodland; tussock grassland;managementBioregions: Victoria Basin; Ord-Victoria Plains;Sturt PlateauAcross 21 trapping sites, there was no relationshipbetween number of finches and a general measureof fire extent on pastoral properties (although thismeasure was not related closely to the fire historyof the actual site); in contrast, there was asignificant negative relationship with cattledensity.
Tidemann, S. C. (1992). Conservation of theGouldian Finch, N.T. World Wide Fund forNature.Gouldian Finch; threatened species; tropical eucalyptsavanna woodland; management; control burningBioregions: Daly Basin; Victoria-Bonaparte;Ord-Victoria PlainsManagement of the Gouldian Finch shouldrequire controlled burning of the breeding sitesearly in the dry season, to prevent late dry seasondestructive fires.
170
Australia’s Biodiveristy - Responses to Fire

Tidemann, S. C. (1993a). Management of athreatened species: the Gouldian Finchexample. In C. P. Catterall, P. V. Driscoll, K.Hulsman, D. Muir, & A. Taplin (Eds.), Birdsand their habitats: status and conservation inQueensland (pp. 123-131). St Lucia:Queensland Ornithological Socity Inc.Gouldian Finch; tropical eucalypt savanna woodland;tropical eucalypt open forest; threatened species;management; control burningSorghum seeds are important in the diet of thethreatened Gouldian Finch, but their abundanceis reduced by early dry season burning, and earlywet season burning can eliminate annualSorghum.
Tidemann, S. C. (1993b). Where are GouldianFinches after the breeding season? VictorianNaturalist, 110, 238-243.Gouldian Finch; tropical eucalypt savanna woodland;threatened species; control burning; wildfireBioregion: Daly BasinWithin weeks of a hot fire, Gouldian Finchactivity was monitored in a cool burnt (April) anda hot burnt (October) area. Early in the morning,birds tended to feed in the hot burnt area (wherefallen seeds were less hidden by regrowth), butwould fly to the cool area (which had more foliagein canopies) when disturbed. Later in themorning, birds moved to feed in the cool burntarea. Temperatures on the ground were hotter inthe hot burnt area (to 56°C). Cool fires occurduring the nesting season, but the hollow-nestingGouldian Finches are unaffected.
Tidemann, S. C., McArtney, J., & Smith, I.(1993). Queensland Gouldian FinchesErythrura gouldiae and air-sac miteSternostoma tracheacolum. Sunbird, 23, 36-40.Gouldian Finch; tropical eucalypt savanna woodland;threatened species; management; control burningBioregion: Mount Isa InlierPastoralists burning during the early Wet seasonmay have reduced the availability of Sorghum,reducing the survival of Gouldian Finch.
Tingay, A., & Tingay, S. R. (1982a). SeabirdIslands: No. 113. Middle Island, Archipelagoof the Recherche, Western Australia. Corella,6, 49-50.seabirds; tussock grassland; heath; eucalypt open forestBioregion: Esperance PlainsA fire burnt uncontrolled on the Island for nearly4 weeks in January-February 1977, destroyingmost vegetation. “The effect of the holocaust onbreeding seabirds is not known.”
Tingay, A., & Tingay, S. R. (1982b). SeabirdIslands: No. 118. Hood Island, Archipelago ofthe Recherche, Western Australia. Corella, 6,59-60.seabirds; heathBioregion: Esperance Plains“Fire is an everpresent threat” to seabirdsbreeding on Hood Island.
Tingay, A., & Tingay, S. R. (1982c). SeabirdIslands: No. 120. Sandy Hook Island,Archipelago of the Recherche, WesternAustralia. Corella, 6, 63-64.seabirds; tussock grassland; heathBioregion: Esperance PlainsThe main threat to the breeding seabirds is therisk of fire caused by human visitors.
Tingay, A., & Tingay, S. R. (1984). Birdcommunities in the karri forest of WesternAustralia. Australian ConservationFoundation.eucalypt open forest; community; forestry; controlburning; managementBioregion: WarrenBird distributions were monitored in a series ofplots of varying age in Karri forests, especiallyrelating to forestry. Cool burns in mature forestinitially cause a short-term decrease in totalabundance of birds, followed by an increase ofsmall insectivores occurring in low vegetation.These then decline as the shrubs regenerate.Species richness and abundance peaked at 6 yearspost-fire. However some species were morecommon in long-unburnt forests. The birdcommunities in mature forests showed moreseasonal stability than those in 50-year oldregeneration.
171
Fire and Australian birds

Tolhurst, K. (1996). Effects of fuel reductionburning on fauna in a dry sclerophyll forest. InDEST (Ed.), Fire and biodiversity: the effectsand effectiveness of fire management.Proceedings of the conference held 8-9 October1994, Footscray, Melbourne (pp. 113-121(&129)). Canberra: Department of theEnvironment, Sports and Territories.control burning; eucalypt open forestDescribes experiment with 15ha treatments andreplicates, with range of controlled fire regimes.Only results following 1 year of treatment arepresented. “Bird abundance and speciescomposition remained remarkably stable on burntand unburnt areas. The main changes observedafter burning were influxes of some speciesinhabiting understorey, and influx of some species(e.g. scarlet robin, flame robin) which fed oneucalypt nectar. The latter is of some interest, assuch influxes have not been reported in previousstudies of prescribed burning”. Notes the caveatthat the study area was small, and no part of anyburnt site was more than 300m from unburntforest. Also notes that the pre-fire understorey wasrelatively open - hence the responses may be morepronounced in forests with denser understorey.
Trickett, T. (1983). Geelong garden birds onthe days of dust-storm and bushfire. GeelongNaturalist, 20, 28-29.wildfireBioregion: South east Coastal PlainLorikeets and some other birds were behavingstrangely in a garden on the day of extreme winds,smoke and nearby fire.
Turner, R. J. (1987). Effect of fire on birds -Weddin Mountain. In Disappearing islands.Proceedings of a seminar on conservation andco-operation in the Central West (pp. 66-86).Bathurst: National Parks and Wildlife Service,NSW.Callitris woodland; eucalypt woodland; wildfire;community; successionBioregion: NSW South western SlopesBurnt and unburnt vegetation was surveyed 8months, and 8 years 8 months, after an intensewildfire. At 8 months post-fire, the abundance andrichness of bird species was significantly less inburnt than in unburnt vegetation. Species whichwere more abundant in unburnt vegetationincluded: Crested Pigeon, cuckoos, Rainbow
Bee-eater, Speckled Warbler, White-browedBabbler, White-plumed Honeyeater, LittleFriarbird, Yellow-faced Honeyeater,Chestnut-breasted Mannikin, White-browedWood-swallow, Superb Fairy-wren andMistletoebird. Species more common in the burntareas included Emu, White-winged Triller andDusky Wood-swallow. At 8 years 8 monthspost-fire, the abundance and richness of birds wasgreater in burnt area (7.3 birds per observer-hour)than unburnt (3.8 birds per observer-hour). Speciesmore common in burnt area included: Galah,Cockatiel, Turquoise Parrot, Rainbow Bee-eater,Black-faced Cuckoo-shrike, White-winged Triller,Superb Fairy-wren, Western Gerygone, RufousSonglark, Grey Fantail, Brown Treecreeper, VariedSittella, Grey-fronted Honeyeater, White-napedHoneyeater, Noisy Friarbird and DuskyWood-swallow. Golden Whistler and SpeckledWarbler were more abundant in the unburnt forest.These differences were considered to be due torelease of nutrients after fire, which allowedvigorous regrowth of wattles, other shrubs andeucalypts and which probably supported greaterabundance of insects.
Turner, R. J. (1992). Effect of wildfire on birdsat Weddin Mountain, New South Wales.Corella, 16, 65-74.eucalypt woodland; Callitris woodland; wildfire;community; successionBioregion: NSW South western SlopesBirds were surveyed 8 months and 8.7 years afterwildfire, including comparison with adjacentunburnt areas. At 8 months post-fire there werefewer species and individuals in burnt (50 spp, 225individuals) than unburnt (59 spp, 400 individuals)areas. At 8.7 years post-fire, there were morespecies and individuals in the burnt area (87 spp,660 individuals) than unburnt (63 spp, 340individuals) area. Ground-foraging,foliage-gleaning, aerial, nectarivorous,granivorous, trunk-gleaning and frugivorous birdsincreased significantly from 8 months to 8.7 yearspost-fire. Recruitment may have been limitedbecause the site was largely surrounded by clearedfarmlands. The intense fire at this site led toincrease in the dominance of eucalypts at theexpense of Callitris. By 8 years post-fire, RufousSonglark, parrots, Black-faced Cuckoo-shrike,Superb Fairy-wren, Grey Fantail, Varied Sittellaand Apostlebird were more common in the burntarea than unburnt sites.
172
Australia’s Biodiveristy - Responses to Fire

Venn, D. R., & Fisher, J. (1993). Red-tailedBlack-Cockatoo Calyptorhynchus banksiigraptogyne. Department of Conservation andNatural Resources.Red-tailed Black-Cockatoo; eucalypt woodland;wildfire; management; threatened species; hollows.Bioregions: Naracoorte Coastal Plain;Murray-Darling Depression.Reviews information on this species. Wildfire canhave a devestating effect on this species, leading todeath of River Red Gum and Yellow Gum, andhence removing food sources and nesting trees.Impacts of fuel reduction burning are not known,but hot fires may reduce the availability of fruits(of Brown Stringybark) for up to 3 years. Thesefactors should be further researched. Firewoodcollection is likely to be detrimental to this species.
Verbeek, N. A. M., Braithwaite, R. W., &Boasson, R. (1993). The importance ofPandanus spiralis to birds. Emu, 93, 53-58.pandanus; communityBioregion: Top End CoastalDisturbance due to increasingly intense fires mayhave reduced pandanus thickets and theirassociated birds, though little evidence is available.
Walker, T. A., & Hulsman, K. (1993). SeabirdIslands: No. 221. Wilson Island, Great BarrierReef, Queensland. Corella, 17, 155-157.seabirds; pandanus; tussock grasslandBioregion: South Eastern QueenslandPrevious dominance by Pandanus has been muchreduced by burning. This reduction has led todecline of Wedge-tailed Shearwaters. The islandwas set on fire in the 1960s and 1970s, whichwould have caused mortality to breeding seabirds.Bridled Terns were probably most affected.
Wall, C. A. (1989) Post-fire dynamics of thevegetation, habitat and population of the GroundParrot at Barren Grounds Nature Reserve, NewSouth Wales. B.Sc. (Hons.), University of NewSouth Wales.Ground Parrot; heath; threatened speciesBioregion: South East Corner
Wardell-Johnson, G., & Christensen, P.(1992). A review of the effects of disturbanceon wildlife of the karri forest. In Research onthe impact of forest management in south-westWestern Australia (pp. 33-57). Perth:Department of Conservation and LandManagement (Occasional Paper 2/92).eucalypt open forest; management; review; controlburningBioregion: WarrenHistoric changes in the bird community insouth-western Australia were mainly from heathsas a result of frequent burning. Reviews studies onthe effects upon birds of fuel-reduction burning inKarri forests. Generally, small decrease inabundance of understorey birds for a few monthsafter burning, followed by increase to abovepre-burn levels within 1-2 years post-fire. Manyindividuals remain in the same locality after fire.
Wardell-Johnson, G., & Nichols, O. (1991).Forest wildlife and habitat management insouthwestern Australia: knowledge, researchand direction. In D. Lunney (Ed.),Conservation of Australia’s forest fauna (pp.161-192). Mosman: Royal Zoological Societyof NSW.eucalypt open forest; review; control burning; wildfire;managementBioregions: Jarrah Forest; Swan Coastal Plain;WarrenReviews studies dealing with fire, birdcommunities and their relationship in forests ofsouthwestern Australia. Describes an ongoingstudy of birds in 67 plots, of which nearly halfhave been burnt.
Wardell-Johnson, G., McCaw, W. L., & Masey,K. G. (1989). Critical data requirements forthe effective management of fire on natureconservation lands in south Western Australia.In N. Burrows, L. McCaw, & G. Friend (Eds.),Fire management on nature conservation landsPerth: Department of Conservation and LandManagement.eucalypt open forest; heath; review; management;threatened species; control burningBioregions: Jarrah Forest; Warren; Swan CoastalPlainFire management plans need to be associated withmonitoring of threatened and key species.
173
Fire and Australian birds

Watkins, D. (1985). Report of the R.A.O.U.Ground Parrot survey in Western Australia.Royal Australasian Ornithologists Union.Ground Parrot; heath; threatened speciesBioregions: Esperance Plains; Jarrah Forest;WarrenGround Parrots were recorded only in diverseshrub-heathlands unburnt for at least 16 years.
Watkins, D., & Burbidge, A. H. (1992).Conservation of the Ground Parrot inWestern Australia. In L. Joseph (Ed.), Issues inthe Conservation of Parrots in Australasia andOceania: Callenges to Conservation Biology.Proceedings of the RAOU/WWF Scientific Dayand Workshop, Sydney, 22-23 September 1990.(pp. 46-49). Melbourne: RAOU.Ground Parrot; threatened species; management;heath; wildfireBioregion: Esperance PlainsIn WA, the Ground Parrot has been recorded fromheaths of 15-35 years after fire, though individualsmay sometimes forage in the edge of adjacent areasthat have been burnt more recently. Thesepreferred ages are appreciably greater than thosereported for mainland eastern Australia. Wildfire isthe major threat to this subspecies. Recentextensive wildfires have probably greatly reducedpopulations, including in national parks.
Webster, R., & Ahern, L. (1992). Managementfor conservation of the Superb Parrot (Polytelisswainsonii) in New South Wales and Victoria.Department of Conservation and NaturalResources.Superb Parrot; threatened species; eucalypt woodland;control burning; managementBioregion: RiverinaFor the Superb parrot, the impacts of controlburning are unclear, but may include reduction infood availability during the breeding orpost-breeding period (and hence reduced breedingsuccess), change in vegetation structure orcomposition (possibly including invasion by exoticplants), and damage or destruction of nest sites.However, control burning may provide a tool forincreasing food resources or other habitatcharacteristics. Recommends that nest sites begiven priority protection during fire suppression,that control burns not occur around nest sites(unless slashing is done first), that extensivecontrol burns not occur in major feeding sites, that
planned burning within the breeding range shouldoccur between May and July, and that moreresearch be directed at the effects of fire.
Wegener, A. (1984a). Animals killed bybushfires in coastal Victoria, February 1983.Australian Bird Watcher, 10, 248-250.mortality; wildfireBioregions: South east Coastal Plain; SouthEastern HighlandsDead birds found on the beach following the AshWednesday bushfires were counted. 55 specieswere recorded, the most common being CrimsonRosella, Red Wattlebird and New HollandHoneyeater. Many were badly burnt, but othersshowed no signs of burns. The northerly windsblew the fire towards the coastline and many birdshad tried to escape by flying out to sea.
Wegener, A. (1984b). Survey of animal specieskilled by the Ash Wednesday fires along theGreat Ocean Road. Geelong Naturalist, 21,13-16.mortality; eucalypt open forestBioregion: South east Coastal PlainCount of 55 bird species washed up along thebeach immediately after hot wildfire. The mostcommon dead birds were Crimson Rosella, NewHolland Honeyeater and Red Wattlebird. Manywere badly burnt, but some appeared undamagedand were presumably asphyxiated.
Whelan, R. J. (1995). The ecology of fire.Cambridge: Cambridge University Press.reviewReviews many aspects of fire ecology. For birds,recommends more long-term studies with bandedbirds and research into possible ecologicaldisadvantages of control burning.
White, G. (1979a). Seabird islands: No. 75.Louisa Island, Tasmania. Corella, 3, 61-62.seabirds; mortality; tussock grasslandBioregion: West and South West“A very severe fire was lit on the island in recentyears and burnt for several weeks extending overmost parts and reportedly killing countlessthousands of breeding shearwaters.”
174
Australia’s Biodiveristy - Responses to Fire

White, G. (1979b). Seabird Islands: No. 76.De Witt Island, Tasmania. Corella, 3, 63-65.seabirds; eucalypt open forest; heath; tussock grassland;mortalityBioregion: West and South WestFishermen frequently set fire to the vegetationcausing heavy casualties among the penguincolonies. Three fires in 1975 and 1976 reducedthe population of breeding penguins by almostone thousand, with even greater toll of chicks.
White, G. (1979c). Seabird Islands: No. 77.Flat Witch Island, Tasmania. Corella, 3, 66-67.seabirds; mortality; heath; tussock grasslandBioregion: West and South WestFires are occasionally lit during the seabirdnesting season and undoubtedly kill manyseabirds.
White, G. (1979d). Seabird Islands: No. 79.Flat Island, Tasmania. Corella, 3, 70-72.seabirds; tussock grassland; mortalityBioregion: West and South WestFires are uncommonly lit during the seabirdbreeding season, leading to some losses ofbreeding birds.
Whitley, G. P. (1944). Fire and Petrels: themystery of Mondrain Island. Emu, 44, 6-7.seabirds; tussock grassland; heath; mortalityBioregion: Esperance PlainsMany breeding shearwaters were killed onMondrain Island in an extensive fire.
Whitley, G. P. (1971). Field notes on birds byThomas Carter. Western Australian Naturalist,12, 41-44.Western Whipbird; heath; threatened species; historicchange.Bioregion: WarrenFrequent firing of heaths probably led to the localextinctions of Western Whipbirds.
Whittell, H. M. (1936). The Bristle-birds ofWestern Australia. Emu, 35, 197-201.Western Bristlebird; Rufous Bristlebird; heath;wildfire; threatened speciesBioregion: WarrenReports from Whitlock that Western Bristlebirddisappeared from an area after severe fire.
Williams, J. E., & Gill, A. M. (1995). Theimpact of fire regimes on native forests in easternNew South Wales. NSW National Parks andWildlife Service.review; eucalypt open forest; management; controlburning; wildfire; forestry; historic change; AboriginalburningBioregions: South East Corner; South EasternHighlands; Sydney Basin; NSW North Coast;New England TablelandBroad review of ecological impacts of fire ineastern NSW. For birds, responses “depend tosome extent on fire intensity … Unburnt patchesprovide important resources … Species occupyingthe lower vegetation strata are … likely to be mostaffected by the changes in vegetation structurecaused by fire … Following fire of low tomoderate intensity, a rapid recovery in bothnumbers and species composition is generallyobserved, whereas the recovery process after moreintense fires is usually slower.”
Wilson, B. A. (1996). Fire effects on vertebratefauna and implications for fire managementand conservation. In DEST (Ed.), Fire andbiodiversity: the effects and effectiveness of firemanagement. Proceedings of the conference held8-9 October 1994, Footscray, Melbourne (pp.131-147). Canberra: Department of theEnvironment, Sports and Territories.reviewReviews studies on responses of birds (and othervertebrates) to fire. Recommends increasedsynthesis and modelling (across a range of scales);longer-term studies; effects of fire interacting withother factors (e.g. predation); and improvedcommunication between researchers andmanagers. Also, notes far more studies for plantsthan for animals.
Wilson, R. I. (1981). The woodchip industryand Tasmanian birds. Tasmanian Bird Report,11, 11-14.forestry; eucalypt open forest; Spotted Quail-thrush;management; control burningFire regimes may exacerbate or ameliorateforestry effects. For example, the SpottedQuail-thrush may better tolerate logging if theforest is allowed to regenerate without use of fire.
175
Fire and Australian birds

Wilson, S. J. (1995). Survival of Brown andStriated Thornbills in the Brindabella Range,Australian Capital Territory. Corella, 19,138-146.Brown Thornbill; Striated Thornbill; eucalypt openforest; wildfire.Bioregion: South Eastern HighlandsWildfire burnt about half of a study site in themiddle of a 20-year study. Banding and survivaldata are presented on a year-by-year basis, thoughthe effects of fire are not analysed.
Woinarski, J. C. Z. (1987). Notes on the statusand ecology of the Red-lored WhistlerPachycephala rufogularis. Emu, 87, 224-231.Red-lored Whistler; mallee; heath; threatened speciesBioregion: Murray Darling DepressionRed-lored Whistlers prefer mallee vegetation of5-30 years post-fire, probably because theyrequire dense low cover under relatively openmallee canopies. Consequently, long intervals offire exclusion may lead to the loss of this species.
Woinarski, J. C. Z. (1989a). Broombushharvesting in southeastern Australia. In J. C.Noble & R. A. Bradstock (Eds.),Mediterranean landscapes in Australia: malleeecosystems and their management (pp. 362-378).Melbourne: CSIRO.mallee; heath; management; community; successionBioregion: Murray Darling DepressionAlthough few bird species were restricted toparticular age classes of vegetation, for 18 of 26commonly-recorded species, there was significantvariation in density between sites with differentpost-disturbance age. Shy Heath-wren,Tawny-crowned Honeyeater and ChestnutQuail-thrush favoured young (<10 years)regrowth; Yellow Thornbill, White-browedBabbler and Malleefowl favoured old (>30 years)vegetation.
Woinarski, J. C. Z. (1989b). The vertebratefauna of broombush Melaleuca uncinatavegetation in north-western Victoria, withreference to effects of broombush harvesting.Australian Wildlife Research, 16, 217-238.mallee; heath; community; succession; managementBioregion: Murray Darling DepressionBird assemblages were sampled in malleebroombush stands 4,8,26,40 and 60-80 yearspost-fire, 3 years after a severe frost, and 1-2,3,4-5and 6-10 years after vegetation harvesting.Invertebrates were least abundant in oldvegetation. There was substantial change in birdspecies composition from young to old vegetation.Species characteristic of areas burnt recently (<10years) included Chestnut Quail-thrush and ShyHeath-wren. Species most common inintermediate ages (10-30 years post-fire) includedSouthern Scrub-robin and Red-lored Whistler.Species most common in older vegetationincluded Malleefowl and Gilbert’s Whistler.There were some differences in bird speciescomposition between sites regrowing forcomparable periods after fire, frost and cutting(with burnt sites having fewest species andindividuals, possibly because more trees wereretained with frost and cutting disturbances).
Woinarski, J. C. Z. (1990). Effects of fire onthe bird communities of tropical woodlandsand open forests in northern Australia.Australian Journal of Ecology, 15, 1-22.tropical eucalypt open forest; tropical eucalypt savannawoodland; community; successionBioregions: Top End Coast; Daly Basin.Two studies are reported: a short-term response tofires at 3 times during the dry season, andlonger-term responses to four fire regimesmaintained over 14 years. Of 11 common species,6 showed significant differences betweenlong-term fire treatments (fire exclusion, annualearly dry season burn, annual late dry season burn,biennial early burn): White-throated Honeyeater,Weebill and Lemon-bellied Flycatcher were allsignificantly more common in unburnt plots andthen in plots burnt biennially, and PiedButcherbird, Blue-winged Kookaburra andMagpie-lark were absent in unburnt plots. Ingeneral, granivores were most common in theearly burnt plots, carnivores were least common inunburnt plots, and foliage-gleaners,branch-gleaners and nectarivores were mostcommon in unburnt plots. These differences were
176
Australia’s Biodiveristy - Responses to Fire

related to variation in extent and composition ofunderstorey. With short-term responses, early dryseason burns were relatively cool and localised,and a wide range of bird species (Gouldian Finch,Long-tailed Finch, Masked Finch, Galah,Cockatiel, Magpie-lark, Grey-crowned Babbler,Pied Butcherbird, Black-faced Wood-swallow andBlack-tailed Treecreeper) were more common inburnt areas from 3 days to 4 months after fire.This attraction was less pronounced beyond 4months post-fire, possibly because of greaterextent then of burnt areas outside the study site.Several other species (Tree Martin,White-breasted Wood-swallow, Black Kite,Torresian Crow and Brown Falcon) moved intothe study site when fires were burning. Only onespecies (Red-backed Fairy-wren) showedshort-term decline in burnt areas. There was littleshort-term response to the late fire, either becauseit destroyed more of the seed resources, orbecause of the greater regional extent of burntareas. Many species in this region probably trackfires to take advantage of the consequent rich foodresources or greater access to food (throughclearing of the dense grass layer). In comparisonto temperate forests, there appears to be a greaterpositive response of birds to cool burns, and a lessobvious long-term successional response. Thelatter may be because the prevalence of fireprevents any substantial vegetation succession intropical eucalypt open forests.
Woinarski, J. C. Z. (1992). The conservationstatus of the White-throated GrasswrenAmytornis woodwardi, an example of problemsin status designation. Northern TerritoryNaturalist, 13, 1-5.White-throated Grasswren; hummock grasslandBioregion: Pine Creek ArnhemWhite-throated Grasswren densities were less inrecently burnt (<1yr) sites, and some populationsmay have been eliminated by repeated hot fires.The fire regime of the Arnhem Land massif isundergoing substantial change, probably to thedetriment of this species, and its existence may bethreatened by the current regime.
Woinarski, J. C. Z. (1993a). Australian tropicalsavannas, their avifauna, conservation statusand threats. In C. P. Catterall, P. V. Driscoll,K. Hulsman, D. Muir, & A. Taplin (Eds.),Birds and their habitats: status and conservationin Queensland (pp. 45-63). St Lucia:Queensland Ornithological Socity Inc.tropical eucalypt savanna woodland; topical eucalyptopen forest; tussock grassland; hummock grassland;review; managementFire regimes in monsoonal savannas have changedmarkedly since European colonisation. Limitedresearch in savanna woodlands suggests that earlydry season fires attract many birds, those later inthe dry season are more detrimental. Fireexclusion may benefit species which feed or nest inshrubby understorey. No research has beenundertaken on fire and birds in floodplains. Fire isusually excluded from Mitchell grasslands, whichmay be disadvantageous for granivorous birds.Recommends more research on fire and birds,particularly in Mitchell and floodplain grasslands.
Woinarski, J. C. Z. (1993b). A cut-and-pastecommunity: birds of monsoon rainforests inKakadu National Park, Northern Territory.Emu, 93, 100-120.rainforest; communityBioregion: Pine Creek ArnhemRelative to larger patches, small rainforest patchestend to be more disturbed by fire (and otherfactors), and have fewer obligate rainforest birdspecies. The abundance of several open forest orrainforest margin species (e.g. Pied Butcherbird,Bar-shouldered Dove) within monsoon rainforestswas positively correlated with recent extent of firein rainforest patches.
Woinarski, J. C. Z., & Fisher, A. (1995a).Wildlife of lancewood (Acacia shirleyi) thicketsand woodlands in northern Australia: 1.variation in vertebrate species compositionacross the environmental range occupied bylancewood vegetation in the NorthernTerritory. Wildlife Research, 22, 379-411.Acacia woodlands; community; wildfireBioregions: Sturt Plateau; Gulf Fall and Uplands;Ord-Victoria PlainsMany lancewood patches showed impact of fire,and wildfire is the major threat to lancewood. Theabundance of 11 bird species (including PeacefulDove, Singing Honeyeater, Double-barred Finch
177
Fire and Australian birds

and Pied Butcherbird) was significantly negativelyassociated with fire impact, whereas only 2 speciesshowed positive correlations.
Woinarski, J. C. Z., & Fisher, A. (1995b).Wildlife of lancewood (Acacia shirleyi) thicketsand woodlands in northern Australia: 2.comparisons with other environments of theregion (Acacia woodlands, Eucalyptus savannawoodlands and monsoon rainforests). WildlifeResearch, 22, 413-443.Acacia woodland; rainforest; tropical eucalypt savannawoodland; historic changesThe distributions of bird species offers somesupport to a formerly extensive continuum frommonsoon ranforests to Acacia thickets. Climatechange and/or changes in fire regime may havefragmented this vegetation, with consequentexpansion of Eucalyptus savanna woodlands.
Woinarski, J. C. Z., & Tidemann, S. C. (1991).The bird fauna of a deciduous woodland in thewet-dry tropics of northern Australia. WildlifeResearch, 18, 479-500.tropical eucalypt savanna woodland; communityBioregion: Daly BasinFire, rain, flowering and seeding produce adynamic patchwork of environments, to whichbird species respond by mobility and shiftingcomposition at any site.
Woinarski, J. C. Z., & Tidemann, S. C. (1992).Survivorship and some population parametersfor the endangered Gouldian Finch Erythruragouldiae and two other finch species at twosites in tropical northern Australia. Emu, 92,33-38.Gouldian Finch; tropical eucalypt savanna woodland;threatened speciesBioregions: Daly Basin; Victoria-BonaparteExtensive nomadism in Gouldian Finch may be aresponse to environmental heterogeneity causedby fire, rainstorms, floristic and substratepatterning.
Woinarski, J. C. Z., Eckert, H. J., &Menkhorst, P. W. (1988a). A review of thedistribution, habitat and conservation status ofthe Western Whipbird Psophodes nigrogularisleucogaster in the Murray mallee. SouthAustralian Ornithologist, 30, 146-153.Western Whipbird; mallee; heath; threatened species;managementBioregion: Murray Darling DepressionPrevious studies have differed in characterisingthe preferred age of vegetation used by theWestern Whipbird. This study also shows noconsistent pattern, with records from 2 yearspost-fire to at least 40 years post-fire. However,intermediate (10-25 years) ages seem mostsuitable, but this may change with differentfloristic associations, or with localisedidiosyncracies (such as insect outbreaks). Otherterrestrial bird species showing association withseral stages in mallee include Shy Heath-wren andChestnut Quail-thrush (<10 years post-fire),Southern Scrub-robin (10-30 years), Malleefowland White-browed Babbler (>30 years). Habitatfragmentation may exacerbate fire effects,reducing chances of refuge and recolonisation.Small-area mosaic burning to maintain a diversityof fire ages is recommended.
Woinarski, J. C. Z., Tidemann, S. C., & Kerin,S. (1988b). Birds in a tropical mosaic: thedistribution of bird species in relation tovegetation patterns. Australian WildlifeResearch, 15, 171-196.tropical eucalypt open forest; communityBioregion: Top End CoastalTropical eucalypt open forests with a denseshrubby understorey have far richer birdassemblages than those without. This structuralcomplexity is probably related to fire regime,though may also be influenced by moistureavailability.
Woinarski, J. C. Z., Whitehead, P. J., Bowman,D. M. J. S., & Russell-Smith, J. (1992).Conservation of mobile species in a variableenvironment: the problem of reserve design inthe Northern Territory, Australia. GlobalEcology and Biogeography Letters, 2, 1-10.Gouldian Finch; threatened species; management;tropical eucalypt savanna woodland; tussock grasslandThe endangered Gouldian Finch (and other morecommon species) may exploit landscape patchiness
178
Australia’s Biodiveristy - Responses to Fire

by following episodic fires or occasional Dryseason rainstorms. Changes in distribution relatedto fire constrain reservation options for thisspecies and conservation planning generally.
Wooller, R. D., & Brooker, K. S. (1980). Theeffects of controlled burning on some birds ofthe understorey in Karri forest. Emu, 80,165-166.eucalypt open forest; control burning; communityBioregion: WarrenBirds were mist-netted and banded in Karri forestin the year before and after a fuel-reduction burn.The species and number of birds caught were verysimilar before and after fire. Some species showedquantitative changes. Fairy-wrens were lesscommon, possibly because of changed foodresources or less cover. Black-faced Cuckoo-shrikeand Rufous Treecreeper increased, possiblybecause of more open understorey. Six individuals(of 5 spp) were retrapped at the same locationafter fire.
Wooller, R. D., & Calver, M. C. (1988).Changes in an assemblage of small birds in theunderstorey of dry sclerophyll forest insouth-western Australia after fire. AustralianWildlife Research, 15, 331-338.eucalypt open forest; control burningBioregion: Jarrah ForestMist-netting and banding of understorey birdswas conducted 2 months before a cool burn, then2,7,10,22 and 34 months post-fire in a Jarrahforest. Although the fire was mild, limited inextent and left much surrounding vegetationunburnt, the total number of birds caught afterthe fire was about half that of before fire. 22% ofmarked birds were recaptured after fire. After thefire, birds ate relatively more ants but fewerbeetles, than before fire. Prey size was smallerafter the fire than before. Dietary diversity ofinsectivorous birds declined post-fire. Abundanceof all prey types (including ants) decreased afterfire. One common species (Splendid Fairy-wren)pre-fire disappeared for 10 months post-fire. Onespecies absent before fire (Scarlet Robin) becamecommon within 2 months of the fire. Otherwisemost changes were of relative abundance ratherthan change in species presence.
Wouters, M. (1996). Developing firemanagement planning and monitoring. InDEST (Ed.), Fire and biodiversity: the effectsand effectiveness of fire management.Proceedings of the conference held 8-9 October1994, Footscray, Melbourne (pp. 235-239).Canberra: Department of the Environment,Sports and Territories.control burning; heath; Ground Parrot; management;threatened speciesBioregion: South East CornerOutlines a fire management plan for far EastGippsland, which sets out the fire managementrequired to maintain the ecology of coastalheathland, in particular Ground Parrot habitat,and how this can be accommodated within highprotection priority zones (where ecological valuesmay need to be compromised). Also notes thatsome mallee communities should have a fireexclusion management for conservation ofMalleefowl, but such straightforward cases arerare - in most cases, management generally has tobe conservative, adopt a range of fire managementregimes and monitor consequences.
Young, J. (1991). Dealing with fire in parksand protected areas. In Tropics under fire: firemanagement on Cape York Peninsula (pp.43-45). Cairns: Cairns and Far NorthEnvironment Centre.hollows; tropical eucalypt open forest; rainforest;managementBioregion: Cape York PeninsulaHot fires (including control burns) can destroy thehollows important for nesting birds (especiallyowls and parrots), and have been reporteddestroying nests and young.
179
Fire and Australian birds
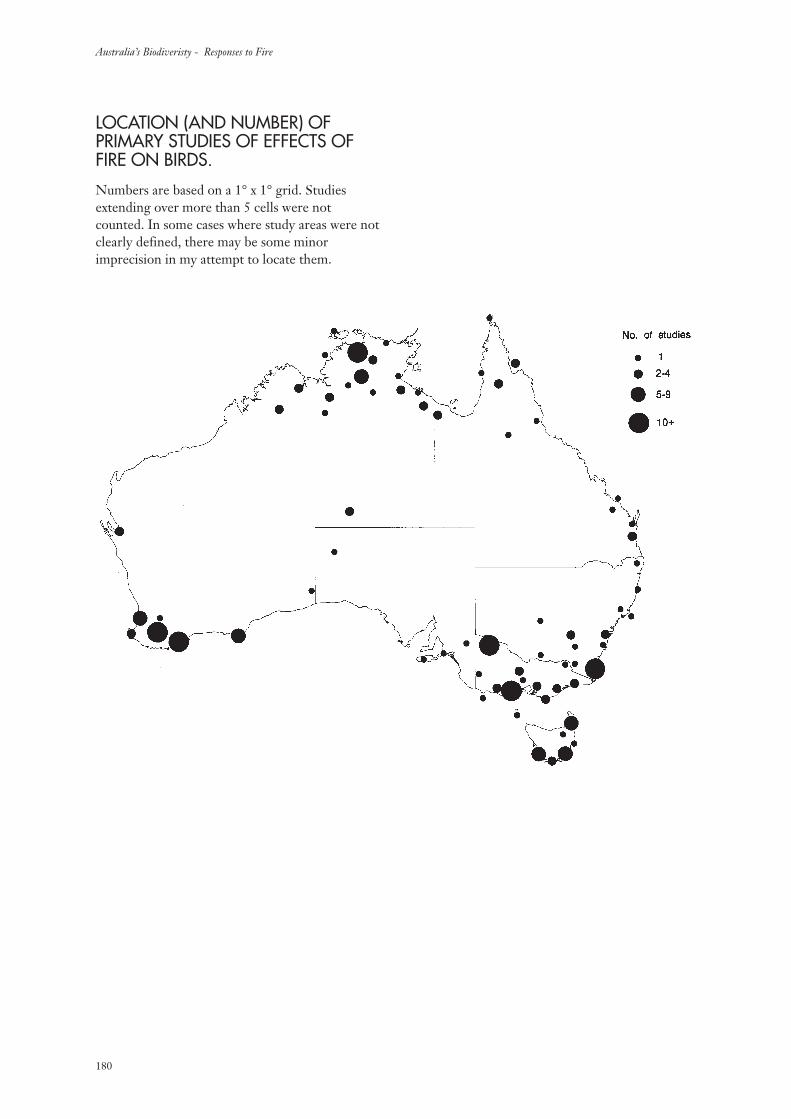
LOCATION (AND NUMBER) OFPRIMARY STUDIES OF EFFECTS OFFIRE ON BIRDS.
Numbers are based on a 1° x 1° grid. Studiesextending over more than 5 cells were notcounted. In some cases where study areas were notclearly defined, there may be some minorimprecision in my attempt to locate them.
180
Australia’s Biodiveristy - Responses to Fire

Forest Research and Development DivisionState Forests of NSW
LONG-TERM EFFECTS OF REPEATED PRESCRIBED BURNING ON
FOREST INVERTEBRATES: Management Implications for the
Conservation of Biodiversity
Consultancy Report to the Department of the Environment and Heritage
Alan York
Final Report April 1996

Full credit must be given to Adrian van Loonwho, in the 1950s, conceived and established afield trial which was sufficiently well designed androbust to not only be addressing questions stillrelevant 40 years later, but also sufficiently flexibleto now facilitate questions probably not evenconceived at that time. Similarly, much is owed tothe Forestry Commission of New South Wales(State Forests) and its staff for the long-termmaintenance of the trial, in particular Bill Bucklerwho methodically and reliably undertook routinemeasurements for over 20 years, and to HughDowden, Bob Bridges and others who maintainedand verified the enormous database.
For my part in this project, which began in1991, I wish to thank Bill Buckler, Bill Chapman,Patrick Murphy and Traecey Brassil for theirassistance with fieldwork, and Rita Holland, RuthBurton, Graeme Price and Darren Waterson forthe tremendous effort and dedication involved insorting in excess of 55,000 specimens. Myappreciation also to Debbie Kent and Chris-Ann
Urquart who assisted in the laboratory work andwith reference material, and to Traecey Brassiland Alison Towerton for their assistance with thedata analyses and presentation. Taxonomicsupport was provided by specialists Mike Gray,Dan Bickel, John Lawrence, Robert Taylor andGerry Cassis who also provided enthusiasticguidance and advice. Thanks also to Andy Beattieand Ian Oliver from the Key Centre forBiodiversity & Bioresources at MacquarieUniversity for their ongoing support and counsel,particularly in regard to the development and useof methodologies for Rapid BiodiversityAssessment.
I also wish to sincerely thank Neal Hardy forhis patience and perseverance, and theDepartment of the Environment and Heritage fortheir financial support.
182
ACKNOWLEDGEMENTS

BACKGROUND
Infrequent, periodic forest fires (bushfires) are anintegral part of the modern physical environment ofAustralian sclerophyll forests. Low-intensity firesare extensively used in managed sclerophyll foreststo stimulate regeneration, manipulate wildlifehabitat and in particular, to reduce fuel levels withthe intention of minimising the extent and severityof wildfires. In Australia, the use of deliberate fire toprevent high-intensity wildfires has becomeprobably the most extensive use of fire in landmanagement. The inherent variability in natural fireregimes generally results in a mosaic of habitatswith vegetation at different stages of floristic andstructural post-fire succession, each potentiallysupporting particular animal communities. Changesto the components of the fire regime (fire intensity,frequency and season of occurrence), as aconsequence of forest management practices, havethe potential to alter the composition and structureof natural communities. The research reported heredeals with the impact of frequent low-intensity fire(“hazard-” or “fuel-reduction burning”) on theabundance, richness, composition and structure ofterrestrial invertebrate communities.
WHY INVERTEBRATES?
Invertebrates (insects, spiders, mites, worms, snails,centipedes etc.) are the most diverse and abundantanimals in most natural systems, but theirimportance in sustaining those systems iscommonly not appreciated. This multitude oforganisms constitutes the bulk of the biodiversitywithin forests and plays an essential role in primaryproduction, nutrient cycling and uptake, population& community level interactions and energy storage& transfer. Through their contribution toecosystem function, these organisms also enableforest ecosystems to provide benefits to humanity.These benefits include amenity values in the formof aesthetics, recreation and education; heritagevalues as forests contribute to long-term securityfor catchment protection, air and water quality andnature conservation; and economic values includingtimber production, grazing and ecotourism. Themaintenance of biodiversity is a fundamentalprinciple underlying the ecologically sustainablemanagement (ESM) of these environments.
THIS REPORT
Little is known about the effects of repeatedhazard-reduction burning over long time scales.The research reported here was conducted as partof the Bulls Ground Frequent Burning Study(Experiment F8/2.9), located on the mid-northcoast of New South Wales and established in1969. The terrestrial invertebrate component ofthe project commenced in 1991 following 20 yearsof repeated low-intensity fire, and was undertakenwith the following aims:• to identify the effects of long-term repeated
burning on terrestrial invertebratebiodiversity,
• to identify species and/or species groupsmost affected by this management practiceand to devise strategies to ensure theircontinued conservation,
• to identify species which, due to the natureof their response, may be useful “indicators”of environmental disturbance anddegradation.
The primary outcome was therefore to assesswhether frequent hazard reduction burning is asustainable long-term management practice withregard to the conservation of our forestbiodiversity.
FREQUENT FIRE & HABITAT STRUCTURE
If frequent fire reduces the diversity of post-fireenvironments, then it has the potential to impactupon animal communities dependent upon thishabitat mosaic. This research indicated thatfrequent burning resulted in a simplification oflarge-scale spatial patterning in the litter (fine-fuel)environment. The components (leaves, twigs, barketc) that give the leaf litter its physical structurechanged with regard to their relative abundanceand spatial distribution. There were markedchanges to the amount (cover) of vegetation in theunderstorey and its spatial patterning. While thequantity of vegetation closest to the ground(ground herbs & small shrubs) was not affected byfrequent burning, there was a decrease in thespatial heterogeneity (patchiness) of these layers.Conversely, the cover of tall and very tall shrubs
183
EXECUTIVE SUMMARY

was substantially reduced and showed an increasein spatial heterogeneity. Top-soil moisture levelswere, on average, 18% lower following 20 years offrequent burning, whereas the amount of lightreaching ground level had increased (on average)by 125% and become more spatially homogeneous(less patchy). A number of habitat components (eg.top-soil hardness, the distribution of large sticks &logs) however showed no significant response tofrequent burning.
TERRESTRIAL INVERTEBRATECOMMUNITIES
This study revealed a rich terrestrial invertebratefauna with representatives from the Chelicerata(spiders, ticks & mites, pseudoscorpions,harvestmen), Crustacea (landhoppers, slaters),Chilopoda (centipedes), Diplopoda (millipedes),and a diverse array of Insect Orders and Families.Numerically, the most abundant groups overallwere the springtails (33%), ticks & mites (24%)and ants (23%), with these three groupsrepresenting 80% of all individuals caught. For 10broad taxonomic groups there were sufficient datato statistically test the effects of frequent burning.The results indicated a variety of responses withstatistically significant decreases in abundance forticks & mites (↓31%), insect larvae (↓35%), flies(↓58%) and beetles (↓31%). Many of these groupsare associated with leaf litter and it is likely thattheir numbers have been influenced by theepisodic removal of this resource. Three groupsshowed substantial increases in abundancefollowing frequent burning; bugs (↑77%), ants(↑250%) and spiders (↑33%), probably as aresponse to both changes in habitat suitability andincreased ease of capture in a simplifiedenvironment.
Biodiversity
Using ants, beetles, flies, spiders & bugs asrepresentative groups and potential indicators ofenvironmental degradation, this researchdemonstrated that although overall speciesrichness at specific sites (α-diversity) did notchange with frequent burning, all groups showedsubstantial changes in the composition of speciesassemblages. There was a loss of taxa dependentupon a substantial litter layer and stable moistconditions, and these species were frequentlyhabitat or dietary specialists and often uncommonor “rare”. The overall diversity of frequently burntareas was maintained by the addition of species
with broad tolerances, or adaptations, to drier andmore open environments.
These shifts in community composition weresubstantial and suggested that the extensive andfrequent application of fuel-reduction burningcould result in a reduction in terrestrialinvertebrate biodiversity at a regional scale, withthis decrease potentially as high as 50%. Currentfuel management strategies which limit the extentof frequent burning will ameliorate these impacts,however there remains a need to establish securerefuges for species with specialist requirements andlimited dispersal abilities, and provide links (iecorridors) between habitat patches to facilitaterecolonisation. The effectiveness of similarstrategies developed to conserve vascular plantsand vertebrates remains untested however for thegroups which actually constitute the bulk of ourforest biodiversity. Realistically, the conservationof biodiversity cannot be achieved withoutconsideration of the important role thatinvertebrates play, both through their involvementin ecological processes, and as a significantcomponent of the overall richness of bioticcommunities.
Community Structure & Ecosystem Function
The biological structure of a community involvesspecies composition (diversity and relativeabundance) and the relationships between species -their ecological role. It was demonstrated here thatconsiderable additional detail concerning, andinsight into, the nature of invertebrate communitychanges could be provided by the inclusion of fairlygeneral information concerning habitat and dietarypreferences. It was apparent that frequent burningleads to a change in the structure of theinvertebrate community. Within speciesassemblages there were shifts based on feedingstrategy and habitat preference. While the impactof these changes on ecosystem function was beyondthe scope of this study, substantial measuredchanges in the structure of invertebrateassemblages and the loss of species associated withthe decomposer cycle implies frequent burningmay be impacting upon nutrient cycling andtransfer within these forests. If this is the case, itwould have serious implications with regard to themaintenance of ecological sustainability.
184
Australia’s Biodiveristy - Responses to Fire

Biodiversity Indicators
Indices used to gauge the success of ecologicallysustainable management practices need to beinterpretable, significant and cost efficient. Theyalso need to account for variability in space andtime, and be appropriate for the scale ofmanagement. The research reported here identifiedthe limited usefulness of data obtained using coarse-scale taxonomic classification (eg. Family or Order),with the cost-effectiveness of abundance data aloneshown to be low. This research also identifiedsubstantial limitations with regard to the use of asingle index, species richness, as a measure ofchange and/or environmental impact. Speciesrichness (α-diversity) is frequently used to describeand compare communities, however in this case itwas found to provide a deceptive summary ofcommunity characteristics and severely restrict thelevel of interpretation that could be derived forimpact assessment purposes. The application ofRapid Biodiversity Assessment (RBA) methodologyhere demonstrated that the study of thecomposition and structure of communities is likelyto prove more rewarding in this regard. Theidentification of individuals to distinct“morphospecies” facilitated the incorporation ofbroad-level ecological information into theassessment, and interpretation, of environmentalimpact. This in turn enabled the development ofmanagement recommendations consistent with theconservation of biological diversity.
Note:
Following the preparation of this report, therehave been some taxonomic revisions andassociated morphospecies corrections of the antdata. These have been independently published,however they were of a minor nature and do notalter the outcomes of the analyses or theconculusion drawn in this report.
185
Bushfire and forest invertebrates


187
Bushfire and forest invertebrates
The concept of Ecologically SustainableDevelopment (ESD) was defined by the UnitedNations in 1987 as “… development that meets theneeds of the present without compromising the ability offuture generations to meet their own needs”(“Bruntland Report” - WCED 1987). Thisconcept has been developed and refined regularlysince that time, most recently at the “EarthSummit”, the United Nations Conference onEnvironment and Development held in Rio deJaneiro in 1992. ESD forms part of the WorldConservation Strategy (IUCN 1980) and is thebasis for the National Conservation Strategy forAustralia (Commonwealth of Australia 1983).
The conservation of biological diversity is afoundation of ESD and is one of the three coreobjectives of the Australian National Strategy forEcologically Sustainable Development. Biologicaldiversity refers to the variety of all life forms - thedifferent plants, animals and micro-organisms, thegenes they contain and the ecosystems of whichthey form part. Australia has ratified theConvention on Biological Diversity arising fromthe Earth Summit, and is now developing strategiesto assess and protect its biodiversity. Theconservation of biological diversity is a majorobjective of the National Forest Policy Statement(NFPS 1992), to be achieved through theprotection of ecosystems (reserve strategies) andcomplementary off-reserve management(Ecologically Sustainable Management — ESM).In New South Wales, State Forests has put forwardESM as a major objective in its 1992–5 CorporatePlan (Forestry Commission of NSW 1992). Thisconcept has been widely adopted by other landmanagement agencies throughout Australia andforms part of the National Strategy for theConservation of Australia’s Biological Diversity.
Low-intensity fires are extensively used inmanaged sclerophyll forests to stimulateregeneration, manipulate wildlife habitat and inparticular, to reduce fuel levels with the intentionof minimising the extent and severity of wildfires.In Australia, the use of deliberate fire to preventhigh-intensity wildfires has become probably themost extensive use of fire in land management(Whelan 1995). While infrequent, periodic fires(bushfires) are an integral part of the modernenvironment of Australian sclerophyll forests,
there is growing concern that repeated low-intensity burning, as a management prescription,may have a negative influence on plant and animalcommunities. Frequent firing may removevegetation species that rely on seed production fortheir persistence (Gill 1981; Bradstock andMyerscough 1981; Benson 1985; Fox and Fox1986), often leading to dominance by herbaceousfire-tolerant species (Cary and Morrison 1995).Fire frequency becomes a significant factor forplant species requiring a long period of time(relative to the interval between fires) to reachreproductive maturity (Zedler et al. 1983;Nieuwenhuis 1987). Changes in habitat structureas a consequence of frequent burning are likely todisadvantage many native mammal and birdspecies (Catling 1991; Whelan 1995).
While sclerophyll forests, woodlands andheaths are dominated by plant species withadaptive responses to fire that enable them tosurvive exposure to periodic burning (see forexample Gill 1981; Noble and Slatyer 1981), theimpact of such fires on terrestrial invertebrates ispoorly understood. The consumption of some orall of the leaf litter by flame, short-lived butsubstantial rises in soil temperature during fire,and post-fire changes in the surface radiationbudget, mean that soil and litter fauna aresubstantially affected by fire in the short-term(Bornemissza 1969; Springett 1979; Moulton1982; Coy 1996). Recovery from a single fire maytake up to 3-5 years (Metz and Farrier 1973;Seastedt 1984; Neumann and Tolhurst 1991),however the timing and intensity of burning isimportant, as is the mobility and recolonisingability of particular species (Morris 1975). Giventhe patchy nature of low-intensity fuel-reductionburns, and the protection afforded by smallhabitat refuges and within the soil, it has beensuggested that periodic fires used for fuelmanagement purposes have few long-term effectson most soil and litter invertebrates (Majer 1980;Campbell and Tanton 1981; Abbott et al. 1984).
There is little information on the effects offire frequency on forest invertebrates, but Abbottet al. (1984) suggest that periodic low intensityfires have few permanent effects on most of theinvertebrate taxa present in the litter and soil ofthe Jarrah forest. Long-term studies of spiders
1. INTRODUCTION

thereby making a substantial contribution to ourNational biodiversity (New 1984; CONCOM1989). Realistically, the conservation ofbiodiversity cannot be achieved withoutconsideration of the important role thatinvertebrates play, both through their involvementin ecological processes, and as a significantcomponent of the overall richness of bioticcommunities.
The research reported here was thereforeundertaken with the following aims:• to identify the effects of long-term repeated
burning on terrestrial invertebratebiodiversity,
• to identify species and/or species groupsmost affected by this management practiceand to devise strategies to ensure theircontinued conservation,
• to identify species which, due to the natureof their response, may be useful “indicators”of environmental disturbance anddegradation.The primary outcome is therefore to assess
whether frequent hazard reduction burning is asustainable long-term management practice withregard to the conservation of our forestbiodiversity.
188
Australia’s Biodiveristy - Responses to Fire
(Huhta 1971; Merrett 1976) and ants (York 1994,1996) suggest that, in the years following fire,there is a replacement series of groups of speciesrelated to their particular habitat requirementsbeing met as the habitat changes in structure overtime. A number of species persist throughout thisperiod, but show changes in relative dominancewithin the community. York (1996) suggested that,for ants, the use of regular widespread fires forfuel reduction was likely to result in a truncationof these successional patterns and an associatedloss of regional biodiversity.
Periodic low-intensity fire (hazard-reductionburning) is a conspicuous management strategy invirtually all of Australia’s dry forest communities.While it is primarily used to reduce fuel levels,little is known about the effects of its repeated useon natural ecosystems over long time-scales. Onthe east coast of NSW, extensive wildfires inJanuary 1994 have led to calls for increased use ofhazard-reduction burning, however the impacts ofthe resulting increase in fire frequency are poorlyunderstood in the very forest environments thismanagement strategy seeks to protect. Thepaucity of information available on the effects ofincreased fire frequency on forest invertebrates isof considerable concern. Invertebrates constitute95% of known species of fauna in Australia,

189
Bushfire and forest invertebrates
2.1 STUDY AREA
As part of the F8 series of “fire effects” studies,State Forests has an ongoing experimental projectwhich is particularly suitable for addressingquestions relating to repeated disturbance andbiodiversity conservation. The F8/2.9 FrequentBurning Study is located in even-aged coastalblackbutt Eucalyptus pilularis regeneration inCompartment 70, Bulls Ground State Forest,Kendall Management Area on the mid-northcoast of New South Wales (31°33'S, 152°38'E,240m ASL.). The stand was logged andsilviculturally treated in 1958–9, with seed treesretained singly and in groups, andunmerchantable trees culled in line with TimberStand Improvement (T.S.I.) techniques. The areahas experienced no further managementtreatment (except experimental fuel-reductionburning) since that time.
In 1969 twenty-one 0.225 acre (0.1 ha.)temporary plots were established in openingscreated by the logging treatment which carried agood stocking of young blackbutt regrowth (11 years old). These areas were found to supporta mean number of 339 stems per hectare (VanLoon 1970), consisting mainly of blackbutt (48%)and bloodwood E. gummifera (31%). Theremainder (21%) consisted most commonly ofturpentine Syncarpia glomolifera, red mahoganyE. resinifera, white mahogany E. acmeniodes andgrey gum E. punctata. Following an assessment ofstand parameters, a number of these plots wereselected on the basis of their similarity for a long-term fire study (F8/2.9), which was formallyinitiated in March 1970.
Fourteen 0.1 ha. permanent research plotswere established, 7 randomly allocated as burningtreatments (burnt), the remaining 7 as control(unburnt) plots from which fire was excluded (7×2 randomised block design). These study plotswere located within similarly treated forest blocksof approximately 1 ha. and separated by clearedbuffer areas to protect them from wildfire (seeFigure 2.1). For the remainder of this report theterm “plot” refers to the 1 ha. treated forest areas,while “research plot” refers to the 0.1ha studyplots defined in 1970 (see Figure 2.2). Fuelreduction burning was implemented in Autumn
whenever fuel build-up permitted, generally every3 years (1970, 1973, 1977, 1980, 1983, 1986,1989, 1992). This burning regime is ongoing.
A program was instituted to monitor aspectsof the response of this forest to repeated low-intensity fire. A number of parameters wereregularly measured on each research plot: treegrowth, major and minor understorey vegetation,litterfall, and fine and heavy fuel. Thesemeasurements were made systematically between1970 and 1987, when the project was reviewed;and then less frequently until 1992.
2.2 EXPERIMENTAL DESIGN
From an inspection (by the author) of the area in1990 it was apparent that twenty years of repeatedburning had resulted in substantial changes inmacro- and micro-habitat parameters. It washypothesised that these changes would have had asignificant effect on terrestrial invertebratecommunities. In 1991, two years after the last fire,a project was initiated to assess the impact oflong-term fuel reduction burning on terrestrialinvertebrates, and to investigate the possibility ofusing this faunal group as monitoring agents inthe assessment of ecologically sustainablemanagement. The overall approach was to viewthis single sample period as a “snapshot” of theeffects of 20 years of prescribed burning bycomparing burnt and unburnt replicates. Whilethis does not enable a description of changes overtime, it does provide a unique opportunity toassess the long-term impact of this managementpractice.
Twelve of the fourteen plots were selected assuitable, six within each treatment (unburnt &burnt). Plots 7A and 7B were excluded as theycontained rocky outcrops and were subjectivelyassessed to be different to other plots.Randomised assignment of treatments toexperimental units ensured “true” replication oftreatment effects (see Hurlbert 1984). In order toincrease the sensitivity of the experiment byincreasing the “precision” with which propertiesof each experimental unit (plot) and hence eachtreatment were estimated, it was necessary to takemultiple samples from each plot. Four 20mtransects were therefore established within each
2. METHODOLOGY
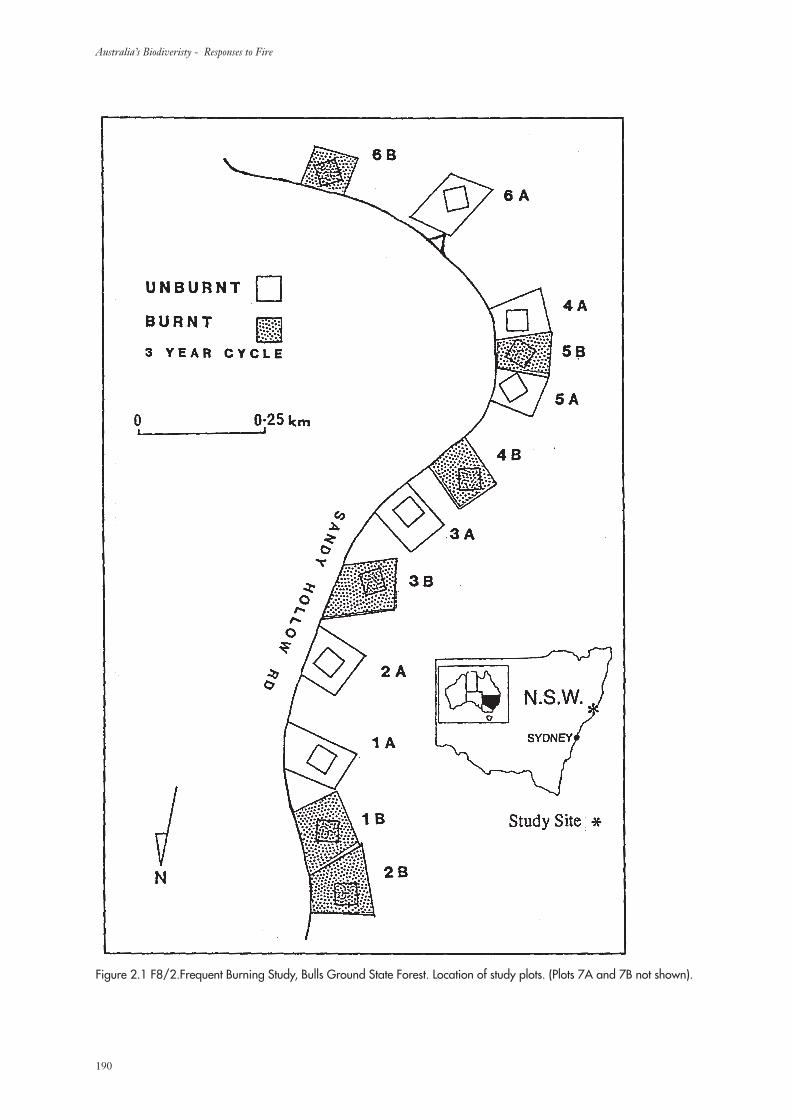
190
Australia’s Biodiveristy - Responses to Fire
Figure 2.1 F8/2.Frequent Burning Study, Bulls Ground State Forest. Location of study plots. (Plots 7A and 7B not shown).
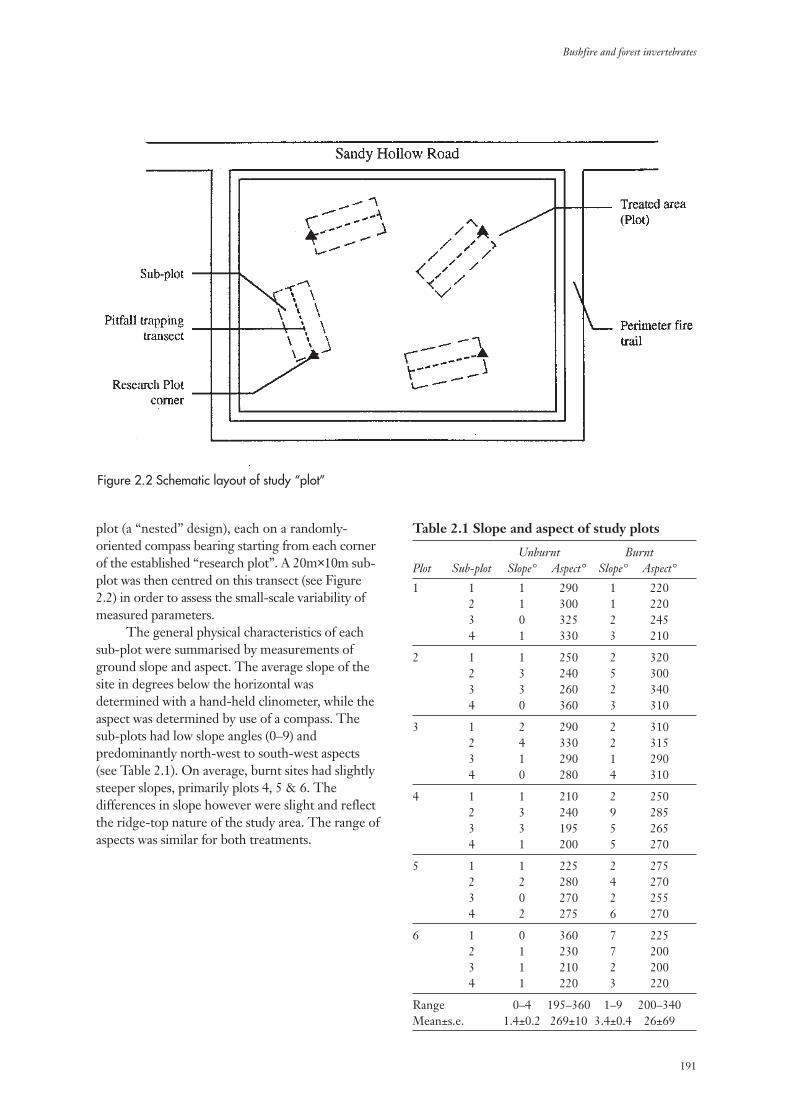
191
Bushfire and forest invertebrates
plot (a “nested” design), each on a randomly-oriented compass bearing starting from each cornerof the established “research plot”. A 20m×10m sub-plot was then centred on this transect (see Figure2.2) in order to assess the small-scale variability ofmeasured parameters.
The general physical characteristics of eachsub-plot were summarised by measurements ofground slope and aspect. The average slope of thesite in degrees below the horizontal wasdetermined with a hand-held clinometer, while theaspect was determined by use of a compass. Thesub-plots had low slope angles (0–9) andpredominantly north-west to south-west aspects(see Table 2.1). On average, burnt sites had slightlysteeper slopes, primarily plots 4, 5 & 6. Thedifferences in slope however were slight and reflectthe ridge-top nature of the study area. The range ofaspects was similar for both treatments.
Table 2.1 Slope and aspect of study plots
Unburnt BurntPlot Sub-plot Slope° Aspect° Slope° Aspect°
1 1 1 290 1 2202 1 300 1 2203 0 325 2 2454 1 330 3 210
2 1 1 250 2 3202 3 240 5 3003 3 260 2 3404 0 360 3 310
3 1 2 290 2 3102 4 330 2 3153 1 290 1 2904 0 280 4 310
4 1 1 210 2 2502 3 240 9 2853 3 195 5 2654 1 200 5 270
5 1 1 225 2 2752 2 280 4 2703 0 270 2 2554 2 275 6 270
6 1 0 360 7 2252 1 230 7 2003 1 210 2 2004 1 220 3 220
Range 0–4 195–360 1–9 200–340Mean±s.e. 1.4±0.2 269±10 3.4±0.4 26±69
Figure 2.2 Schematic layout of study “plot”

Table 2.2 Structual vegetation components
Height class Structural component0–20 cm Ground herbs
20–50 cm Small shrubs50–100 cm Mid-sized shrubs
100–150 cm Tall shrubs150–200 cm Very tall shrubs
2.3.2 The Litter Environment
Ground-dwelling invertebrates have been shownto be sensitive to levels of forest litter, particularlyduring post-fire recovery (Bornemissza 1969;Springett 1976; Seastedt 1984). Five randomlyplaced samples (0.1 m2) of litter (including sticksup to 2.5cm diameter) were collected from eachsub-plot, sieved with 1mm soil sieves to dislodgesoil material, and then dried in an oven at 105° for72 hours. Material was then sorted into 5components and weighed: twigs 0–5mm & twigs6–25mm diameter, bark, leaves, and very fine fuel(miscellaneous decomposing matter). Thisapproach was consistent with that used to estimatethe “fine fuel” fraction over the previous 20 years.
2.3.3 Sticks and Logs
The incidence and diameter of all sticks & logs(“heavy fuel”, >2.5cm) was recorded along 2orthogonal 20m transects centred on each sub-plot. Following an examination of the frequencydistribution of values, data were grouped forsubsequent analyses into the following fivediameter categories: 2.5–9.9, 10–24.9, 25–49.9,50–74.9, 75+cm.
2.3.4 Insolation
Levels of insolation have been shown to be criticalfactors determining the abundance anddistribution of certain terrestrial invertebrates.The amount of light reaching the forest floor wasused as an index of insolation levels, and measuredusing a Lunasix 3 Gossen exposure meter, fittedwith incident light cone. Twenty measurementswere taken systematically within each sub-plotand expressed as a percentage of available light asmeasured outside the forest at that time of day.
2.3.5 The Soil Environment
The underlying geology of the site consistsprimarily of conglomerate, sandstones and shales.These have weathered to form shallow soils(yellow earths & brown podzolics) which arerelatively low in nutrients. Two aspects of the soil
192
Australia’s Biodiveristy - Responses to Fire
2.3 MEASUREMENT OFENVIRONMENTAL PARAMETERS
The environmental framework within whichterrestrial invertebrate communities functionprimarily involves elements of the vegetationunderstorey, the top-soil and litter components. Anumber of parameters were quantitatively assessedto evaluate their possible influence on speciesrichness and community structure.
The distribution of data for most variablessuggested that the sample mean was the bestestimate of average conditions at each sub-plot.Because individual samples were randomly drawnfrom within replicates, a measure of variabilityabout the mean also provided information aboutthe spatial variability (“patchiness”) of the variablesconcerned. The coefficient of variation (CV =standard deviation/mean × 100%) was selected asthe most appropriate measure here due to thelarge fluctuations in mean values and the observeddependence of the standard deviation on the mean.In order to satisfy the assumptions underlyingparticular statistical procedures, variables wereappropriately transformed as required.
2.3.1 Understorey Vegetation Structure
The physical structure of the vegetationunderstorey in this forest environment consistsprimarily of a shrub stratum and a herb stratumwhose heights and spatial distribution are a functionof fire history. A structural classification of thevegetation was chosen because it allowed a relativelyquick and consistent assessment of the sites (48 sub-plots in total) to be made in an environment whichis floristically diverse (Doug Binns pers. comm.).Vegetation structure is of direct significance inecological studies of soil and soil-surfaceinvertebrates because the amount and distributionof vegetation determines both the physicalframework within which activity takes place, and thefood availability and hence carrying capacity of theenvironment (Greenslade and Thompson 1981).
Vegetation structure was quantitativelyassessed using the “cover-board” technique (seeMacArthur & MacArthur 1961; Fox 1979).Percentage cover was measured at 20 pointssystematically located along each transect for fivestructural components of the understorey (seeTable 2.2). The mean of the 20 measurements wasused as an estimate of percentage cover for eachvegetation layer at each sub-plot, and thecoefficient of variation (CV) as an estimate ofspatial variability.

193
Bushfire and forest invertebrates
physical environment were assessed: top-soilmoisture and top-soil hardness.
2.3.5.1 Top-soil MoistureFive samples (10cm diameter 3cm deep) per sub-plot were collected and kept in sealed containers.Samples for each sub-plot were pooled in thelaboratory, weighed and dried in an oven at 105°for 72 hours. An estimate of “field moisturecontent” for each sub-plot was calculated in thefollowing manner (see Lambert 1982):
% Moisture Content =
air-dried weight - oven dried weight air-dried weight
× 100
2.3.5.2 Top-soil HardnessAn index of top-soil hardness (0–5cm) wasobtained using a Geonor inspection vane, whichmeasures soil shear strength. Twentymeasurements were taken within each sub-plot,the mean value representing the average shearstrength and the coefficient of variation (CV) anindication of spatial variability.
2.4 TERRESTRIAL INVERTEBRATECOMMUNITIES
Epigaeic (surface active) invertebrate communitieswere assessed by a single summer pitfall trappingprogram in February 1991. Nine points wereestablished and marked along the 20m transectwithin each sub-plot. At each point a 6.5cmdiameter 9cm deep plastic cup was sunk flush withthe ground surface and half-filled with a non-attractive preservative solution. Pitfall traps wereleft open for a period of 7 days (5–12th February1991), reducing the effect of temporal changes inabundance and activity on estimates of speciesrichness and community composition (York 1989).Weather during this period was typical for thattime of year; temperatures ranged from 17–35°Cand 27mm of rain fell between the 7th and 8th.
Samples were returned to the laboratory andexamined with a binocular microscope wherematerial was sorted to the taxonomic level ofOrder. A number of groups were chosen for moredetailed investigation based on the criteria ofsufficient numbers for statistical analysis, theability to recognise and define species, and thelikely appropriateness of the samplingmethodology. Ants (Hymenoptera: Formicidae),
beetles (Coleoptera), spiders (Araneae), bugs(Hemiptera) and flies (Diptera) were subsequentlysorted to “morphospecies” using the protocolsdescribed in Oliver & Beattie (1993), with finaltaxonomic verifications being performed by MikeGray (spiders), Gerry Cassis (bugs) and DanBickel (flies) of the Australian Museum, JohnLawrence (beetles) and Robert Taylor (ants) of theCSIRO.
Oliver and Beattie (1996a) have shown thatmorphospecies can provide a robust estimate ofspecies richness across a variety of habitats. Thisstudy provides a substantial test of the hypothesisthat the lack of knowledge concerning so manyAustralian invertebrates, the so called “taxonomicimpediment” (Taylor 1983), no longer preventsthe inclusion of invertebrates in biodiversityassessment and studies of management impacts.
2.5 ANALYTICAL PROCEDURES
All initial analytical procedures were performedusing the SPSS statistical package on a 486PC atSFNSW’s Research Division. Data distributionswere examined using exploratory data analysistechniques (EXAMINE) and transformed (asrequired) for subsequent analyses (MANOVA andREGRESSION). Canonical CorrespondenceAnalyses (CCA - Ter Braak 1986) were performedusing programs written in Splus on a SunWorkstation.
2.5.1 Treatment, Exposure and Position Effects
Plots had been allocated to one of two treatments:burnt (1B-6B) or unburnt (1A-6A). Aspect valueswere coded from 1-6 to reflect the relative“exposure” of sub-plots to solar radiation; with300–330° = 1 (highest), 270–300° = 2, 330–360° = 3, 240–270° = 4, 210–240° = 5, and180–210° = 6 (lowest). Sites intermediate betweencategories were allocated an average (mean) code.
To evaluate any large-scale spatial trend inhabitat variables (and species’ responses) a newvariable (position) was generated to reflected thenorth-south location of plots along Sandy Hollow Road (see Figure 2.1). The value of aspectand position for each plot ranged from 1-6 (seeTable 2.3).

component delineates the largest pattern ofrelationships in the data (defines the greatestamount of variation in the data); the seconddelineates the next largest pattern and so on.
2.5.3 Terrestrial InvertebrateCommunities
Samples were sorted to Order usingmorphological characteristics and generaltaxonomic keys. Relative abundance of individualswithin these groups at plots differing in treatmentand position were examined using Analysis ofVariance (ANOVA) procedures.
2.5.3.1 BiodiversitySelected taxa (see 2.4) were described in terms ofthe relative abundance of individuals withinconstituent groups (families, sub-families, generaetc. as appropriate), and their species richness (asdefined by morphospecies). Patterns in thesecommunity descriptors at plots differing intreatment and position were examined graphically,and using Analysis of Variance (ANOVA)procedures.
2.5.3.2 Community CompositionPatterns of species’ responses to treatments areillustrated in tables of relative abundance. Thisenabled broad “assemblages” of species, withsimilar responses to disturbance, to be identified.
2.5.3.3 Environmental Determinants ofCommunity CompositionThe relative importance or ability of the measuredhabitat variables to explain the composition ofinvertebrate assemblages was assessed usingCanonical Correspondence Analysis (CCA, TerBraak 1986, 1991). This method arranges speciesalong environmental gradients by constructinglinear combinations of environmental factorswhich result in maximal separation of species’distributions in ordination species-space. Theseanalyses were performed to determine whetherany differences in turnover or spatial similarity ofassemblages among taxa might be explained by thedifferent taxa responding to environmentalgradients. Ter Braak (1986) fully describes theunderlying assumptions and strengths of thismethod. The main assumption is that individualspecies response models are all similar and ofunimodal, Gaussian form. Although it is doubtfulwhether this assumption is reasonable for allspecies, CCA has been shown to be robust tomoderate violations of assumptions (Palmer 1993)and offers the potentially most powerful method
194
Australia’s Biodiveristy - Responses to Fire
Table 2.3 Values of exposure and position foreach sub-plot. Aspect was coded from 1-6 toreflect the relative exposure of sub-plots to solarradiation (see text). Position was coded from 1(north) to 6 (south) to reflect location along SandyHollow Road (see Figure 2.1) so as to detectpossible spatial patterns.
Exposure (sub-plots)Treatment Plot Position 1 2 3 4
1A 1 2 1.5 1 22A 2 4 4.5 4 3
Unburnt 3A 3 2 2 2 24A 5 5.5 4.5 6 65A 4 5 2 3 26A 6 3 5 5.5 5
1B 2 5 5 4 5.52B 1 1 1.5 3 1
Burnt 3B 3 1 1 2 14B 4 4 2 4 35B 5 2 3 4 36B 6 5 6 6 5
Patterns in environmental parameters atplots differing in treatment and position wereexamined graphically, and using Analysis ofVariance (ANOVA) procedures. For frequencydata, the degree of association between variableswas examined using contingency tables(crosstabulation), with significant associationstested using the χ2 statistic.
2.5.2 Inter-relationships BetweenEnvironmental Variables
The environmental (habitat) variables weresubsequently analysed using an ordinationprocedure (Principal Components Analysis -PCA) in order to untangle linear relationshipsbetween variables, and reflect inherent structuralpatterns. In this analysis, each pattern appears as acomponent delineating a distinct cluster ofinterrelated data. Components are rotatedorthogonally (VARIMAX procedure) to clarify thedefinition of these clusters by maximising orminimising correlations between variables andcomponents. The projection (the loadings) ofeach variable on the component axes defines theclusters of variables. Kaiser’s criterion (only thecomponents with eigenvalues greater than one)was used to determine the number of componentsto be extracted. Eigenvalues measure the amountof variation accounted for by a pattern, while theloadings measure the degree to which variablesare involved in the component pattern. The first

195
Bushfire and forest invertebrates
available in revealing patterns of communitycomposition in relation to environmental factors.It also has the advantage that the results areunaffected by correlations among environmentalvariables.
Results of the CCA ordination weredisplayed as “bi-plots” which show theconfiguration of the variables, the scatter of sub-plots, and the relationship between the two. Thisgives an overview of how community compositionvaries with the environment (Ter Braak 1986).The interpretation of the results of the bi-plotswas simplified by using a sub-set of theenvironmental variables in the analyses. This sub-set was composed of representative variables fromeach of the eight independent patterns identifiedby the Principle Components Analysis (see 2.5.2),with the additional inclusion of two largelyindependent variables: aspect and insolation.
2.5.3.4 Community StructureAnalyses of species’ “assemblages” often fails toadequately account for rare species, which arefrequently represented by too few records to allowany meaningful patterns to be determined (seeYork 1994). One common means to overcomingthis problem, at least to some extent, is to groupspecies according to some ecologicalcharacteristic, so that the collective behaviour ofthe group can be assessed. At the species levelthere is insufficient ecological information formost groups to do this with confidence, howeverbroad grouping may be identified at highertaxonomic levels, such as sub-family or family.
Broad groups of this kind (functional groups) wereidentified in this analysis by reference to theliterature (eg. see Andersen 1990) and followingdiscussions with relevant taxonomists. Thenumbers of morphospecies within these groupswas graphically presented and examined in orderto detect those which may be sensitive tomicrohabitat features associated with structuralcharacteristics of the environment.
Additionally, the relative abundance ofspecies recorded in one treatment only wasdisplayed in tabular form and discussed in relationto their likely ecological roles.
2.5.3.5 Biodiversity IndicatorsDecisions regarding conservation evaluation oftenare based upon the diversity (species richness) ofthe area under concern (see Margules et al. 1988).Similarly, species richness is a common“performance indicator” used for monitoringdisturbance impacts (see Kremen 1992). In orderto simplify these processes, it is oftenhypothesised that one taxonomic or functionalgroup may reflect the response of other taxa, andhence function as “indicator” or “umbrella”species.
To test whether the richness of particularinvertebrate taxa could be useful in predictingoverall invertebrate biodiversity, the relationshipbetween species richness of selected taxa wasinvestigated using correlation analyses (Pearson’sProduct-Moment and Spearman’s Rank).

196
Australia’s Biodiveristy - Responses to Fire
Forest area one day after a low-intensity fuel-reduction burn. The small areas of leaf litter and unburnt understoreyvegetation remaining indicate the patchy nature of such burns. These represent potential refuges for terrestrialinvertebrates; refuges that are reduced in number and extent by frequent fire. /Alan York
Dry eucalypt forest that has remained unburnt for over 25 years. These forests are characterised by deep leaf litter andlow light levels. /Alan York

197
Bushfire and forest invertebrates
Dry eucalypt forest that has been frequently burnt for the past 25 years. These forests are very open and characterisedby low leaf litter levels and high light levels. /Alan York
A spider from the Family Zodariidae. These spiderstypically live under stones, logs and in leaf litter. Therewere four times as many species from this Family onfrequently burnt plots, suggesting that they prefer thesemore open habitats in which to hunt. /Alan York
A spider from the Family Salticidae. These “jumpingspiders” hunt for their food on understorey vegetation,trees and logs. The two species from this Family wereonly found on unburnt plots, suggesting that they preferhabitats with more structurally complex vegetation inwhich to hunt. /Alan York

198
Australia’s Biodiveristy - Responses to Fire
This is a species of ant known as Rhytidoponerametallica which is an “opportunist” commonly found indisturbed habitats. In this study it was 500 time moreabundant on frequently burnt sites, potentially indicatingthat frequent burning is having a negative impact on theenvironment. /Alan York
An ant from the genus Orectognathus. These ants arespecialist predators who use their long mandibles tocatch soft-bodied insects such as Springtails(Collembola). Because of their specialist habitatrequirements, they were not caught in pitfall traps butonly in leaf litter samples collected from near large logson unburnt sites. They were not found on frequentlyburnt sites. In these forests they could be considered anuncommon species with high conservation status. /Alan York
An ant from the genus Probolomyrmex. It is a rare“cryptic” species that was only found in leaf littersamples collected from one site. Little is known about itshabitat preferences, although it is not thought to bedisadvantaged by frequent burning at this stage. /Alan York
A spider from the Family Lycosidae. These “wolfspiders” are ground hunters. All three species from thisFamily were only found on frequently burnt plots,suggesting that they prefer these more open habitats inwhich to hunt. /Alan York

199
Bushfire and forest invertebrates
The results for the various components of theproject are reported separately, and then discussedin terms of their relevance to the existingexperimental fire regime.
3.1 ENVIRONMENTAL PARAMETERS
The following sections summarise the results ofan investigation into the effects of frequentburning on environmental variables as they relateto components of terrestrial invertebrate habitat.Where differences are described as “significant”,this refers to statistical significance at a probabilitylevel of 0.05. Where results were not consideredto be statistically significant, the general nature ofany observed patterns is described. “Box andWhisker” plots are utilised to graphicallyrepresent variation in environmental variables.The box represents the interquartile range(25th–75th percentile) with the median shown.The whiskers indicate the range of values whichlie within 1.5 box lengths of the upper and lowerquartile (75th and 25th percentile respectively).
3.1.1 Understorey Vegetation Structure
3.1.1.1 Ground Herb LayerThe cover of Ground Herbs (0–20cm) on sub-plots showed considerable variation overall(means ranged from 29–98%) with average(mean±s.e.) values for unburnt and burnt plots72.5±1.5 and 80.1±1.3 respectively. Cover ofGround Herbs was less variable (“patchy”) onburnt plots.
Mean values of Ground Herb cover were notsignificantly different between burnt andunburnt plots, however there were significantspatial trends, with patterns quite variable andindependent of treatment and spatial location(position) within the study area (see Figure 3.1A).
3.1.1.2 Small Shrub LayerThe cover of Small Shrubs (20–50cm) on sub-plots showed considerable variation overall(means ranged from 12–91%) with average(mean±s.e.) values for unburnt and burnt plots49.5±1.7 and 57.7±1.6 respectively. Cover of SmallShrubs was less variable (“patchy”) on burnt plots.
Mean values of Small Shrub cover were notsignificantly different between burnt and unburntplots, however there were significant spatial trends,
with patterns quite variable and independent oftreatment and but not spatial location (position)within the study area (see Figure 3.1B).
3.1.1.3 Mid-sized Shrub LayerThe cover of Mid-sized Shrubs (50–100cm) onsub-plots showed considerable variation overall(means ranged from 2–47%) with average(mean±s.e.) values for unburnt and burnt plots14.3±0.8 and 14.5±0.8 respectively. Cover of Mid-sized Shrubs was similarly variable (“patchy”) onburnt and unburnt plots.
Mean values of Mid-sized Shrub cover werenot significantly different between burnt andunburnt plots, however there were significantspatial trends, with patterns quite variable andindependent of treatment and but not spatiallocation (position) within the study area (see Figure3.1C).
3.1.1.4 Tall Shrub LayerThe cover of Tall Shrubs (100–150cm) on sub-plots showed moderate variation overall (meansranged from 2–12%) with average (mean±s.e.)values for unburnt and burnt plots 5.5±0.4 and1.9±0.2 respectively. Cover of Tall Shrubs was lessvariable (“patchy”) on unburnt plots.
Mean values of Tall Shrub cover weresignificantly different between burnt andunburnt plots, with substantially lower cover onfrequently burnt plots. There were howeversignificant spatial trends, with patterns quitevariable and independent of treatment and but notspatial location (position) within the study area (seeFigure 3.1D).
3.1.1.5 Very Tall Shrub LayerThe cover of Very Tall Shrubs (150–200cm) onsub-plots showed moderate variation overall(means ranged from 0–10%) with average(mean±s.e.) values for unburnt and burnt plots2.8±0.3 and 0.2±0.6 respectively. Cover of VeryTall Shrubs was less variable (“patchy”) onunburnt plots.
Mean values of Very Tall Shrub cover weresignificantly different between burnt and unburntplots, with substantially lower cover on frequentlyburnt plots. There were however significant spatialtrends, with patterns quite variable andindependent of treatment and spatial location(position) within the study area (see Figure 3.1E).
3. RESULTS
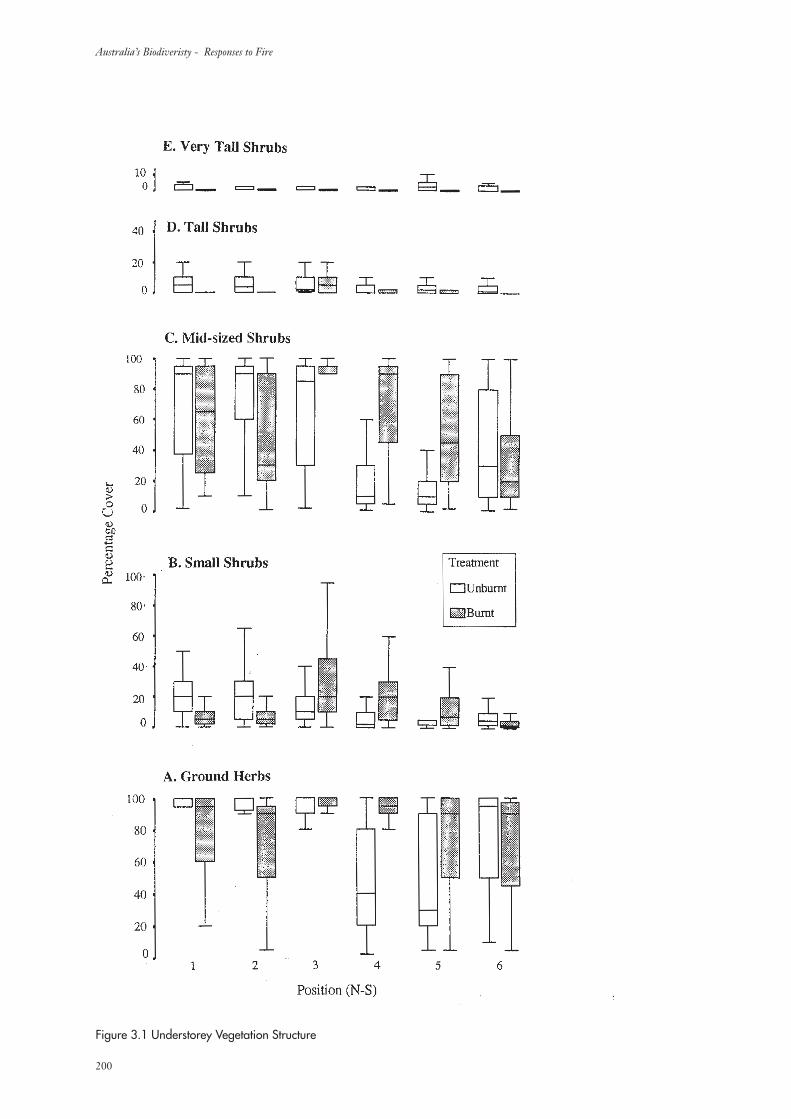
200
Australia’s Biodiveristy - Responses to Fire
Figure 3.1 Understorey Vegetation Structure
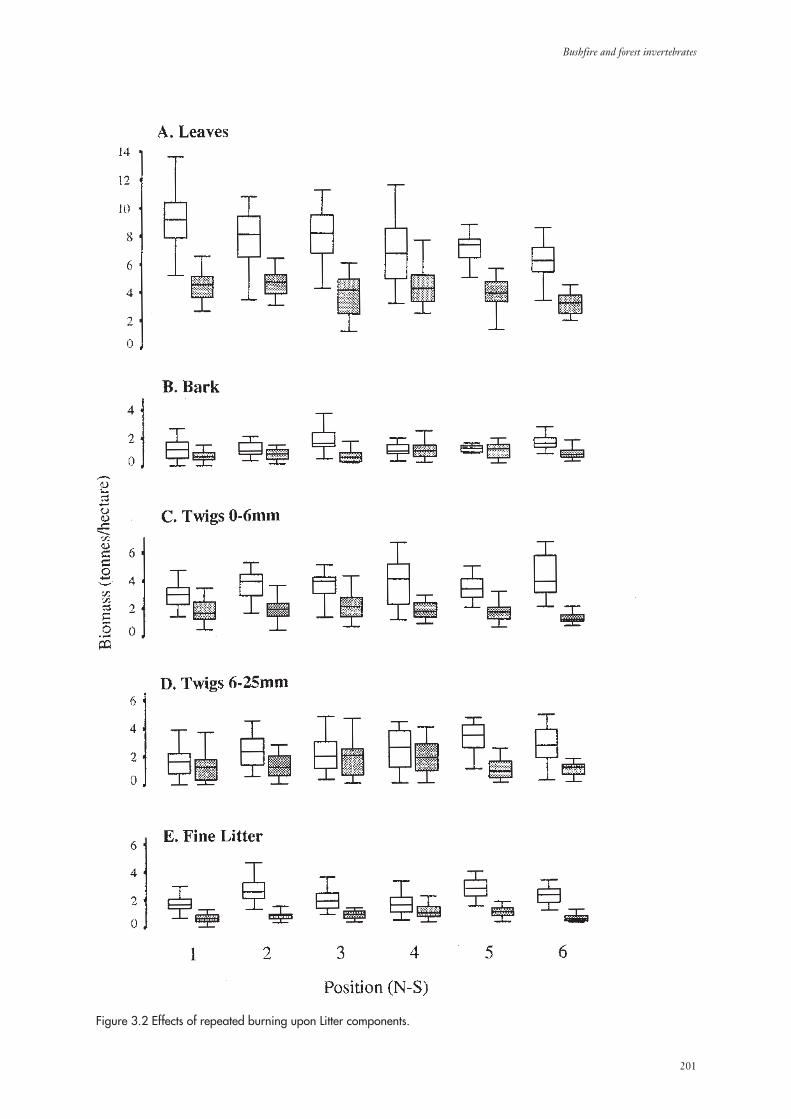
201
Bushfire and forest invertebrates
Figure 3.2 Effects of repeated burning upon Litter components.

202
Australia’s Biodiveristy - Responses to Fire
3.1.2.2 BarkMean biomass of Bark on sub-plots varied from0.50–2.52 t.ha.-1, with the mean (±s.e.) values forunburnt and burnt plots 1.35 (±0.06) and 0.86(±0.06) t.ha.-1 respectively. Bark biomass was quitevariable by nature and considerably more variable(“patchy”) on burnt plots.
Mean values of Bark biomass weresignificantly different between burnt andunburnt plots, with unburnt plots having (onaverage) higher Bark biomass. There were noobvious broad spatial trends with this variable (seeFigure 3.2B), however there was substantialwithin-plot variation, irrespective of treatment, withprimarily a greater level of spatial variabilitywithin burnt plots compared to unburnt plots.
3.1.2.3 Twigs 0-6mmValues of mean Twig biomass (0–6mm diam.) onsub-plots varied from 0.99–5.22 t.ha.-1, with themean (±s.e.) values for unburnt and burnt plots3.65 (±0.01) and 1.84 (±0.08) t.ha.-1 respectively.Twig biomass (0–6mm) was quite variable bynature and slightly more variable (“patchy“) onburnt plots (see Figure 3.2C).
Mean values of Twig biomass weresignificantly different between burnt andunburnt plots, with unburnt plots having (onaverage) higher Twig biomass. There was howeversubstantial within-plot variation, irrespective oftreatment, reflecting the substantial spatial“patchiness” of this variable.
3.1.2.4 Twigs 6–25mmValues of mean Twig biomass (6–25mm diam.) onsub-plots varied from 0.35–6.19 t.ha.-1, with themean (±s.e.) values for unburnt and burnt plots2.72 (±0.21) and 1.53 (±0.16) t.ha.-1 respectively.Twig biomass (0-6mm) was quite variable by natureand slightly more variable (“patchy”) on burntplots (see Figure 3.2D).
Mean values of Twig biomass weresignificantly different between burnt andunburnt plots, with unburnt plots having (onaverage) higher Twig biomass. There was howeversubstantial within-plot variation, particularlywithin burnt plots.
3.1.2.5 Very Fine (Miscellaneous) MaterialMean biomass of Miscellaneous Material on sub-plots varied from 0.33–3.94 t.ha.-1, with the mean(±s.e.) values for unburnt and burnt plots 2.28(±0.11) and 0.85 (±0.04) t.ha.-1 respectively.Miscellaneous Material biomass was quite variableby nature but similarly variable (“patchy”) onburnt and unburnt plots.
3.1.1.6 SummaryThe amount (cover) of vegetation in the firstmetre above the ground (Ground Herbs, SmallShrubs & Mid-sized Shrubs) was not significantlydifferent between burnt and unburnt plots.Significant interactions between the three factors(treatment, position & sub-plot) however reflects thespatially variable nature of these understoreyvegetation components, irrespective of fire history(treatment) and large- (position) & small-scale (sub-plot) location, within the study area. Cover ofGround Herbs and Small Shrubs was however lessspatially variable (“patchy”) on burnt plots, whilethe cover of Mid-sized Shrubs was similarlyspatially variable on burnt and unburnt plots.
In contrast, the amount (cover) of vegetationin the second metre above the ground (Tall &Very-tall Shrubs) was significantly differentbetween burnt and unburnt plots. There wassubstantially lower cover on frequently burntplots, and it was more spatially variable. Therewere however significant interactions between thethree factors (treatment, position & sub-plot) againreflecting the spatially variable nature of theseunderstorey vegetation components, irrespectiveof fire history (treatment) and large- (position) &small-scale (sub-plot) location, within the studyarea.
3.1.2 Litter Biomass
For purposes of analysis, Dry Litter Biomass wasconsidered in 5 categories compatible with otherstudies of “fine fuel”. These categories were:Leaves, Bark, Twigs 0–6mm & 6–25mm diameter,and Miscellaneous (very fine) Material. Green(live) and cured (dead) vegetation data collected aspart of the fuel studies were not used in thisanalysis.
3.1.2.1 LeavesMean biomass of Leaves on sub-plots varied from2.02–9.84 t.ha.-1, with the mean (±s.e.) values forunburnt and burnt plots 7.54 (±0.19) and 4.23(±0.14) t.ha.-1 respectively. Biomass of Leaves wassimilarly variable (“patchy”) on burnt and unburntplots.
Mean values of biomass of Leaves weresignificantly different between burnt andunburnt plots, with unburnt plots having (onaverage) higher leaf biomass. There was anobvious spatial (N-S) trend with regard to thisvariable, which was more apparent for unburntplots (see Figure 3.2A).
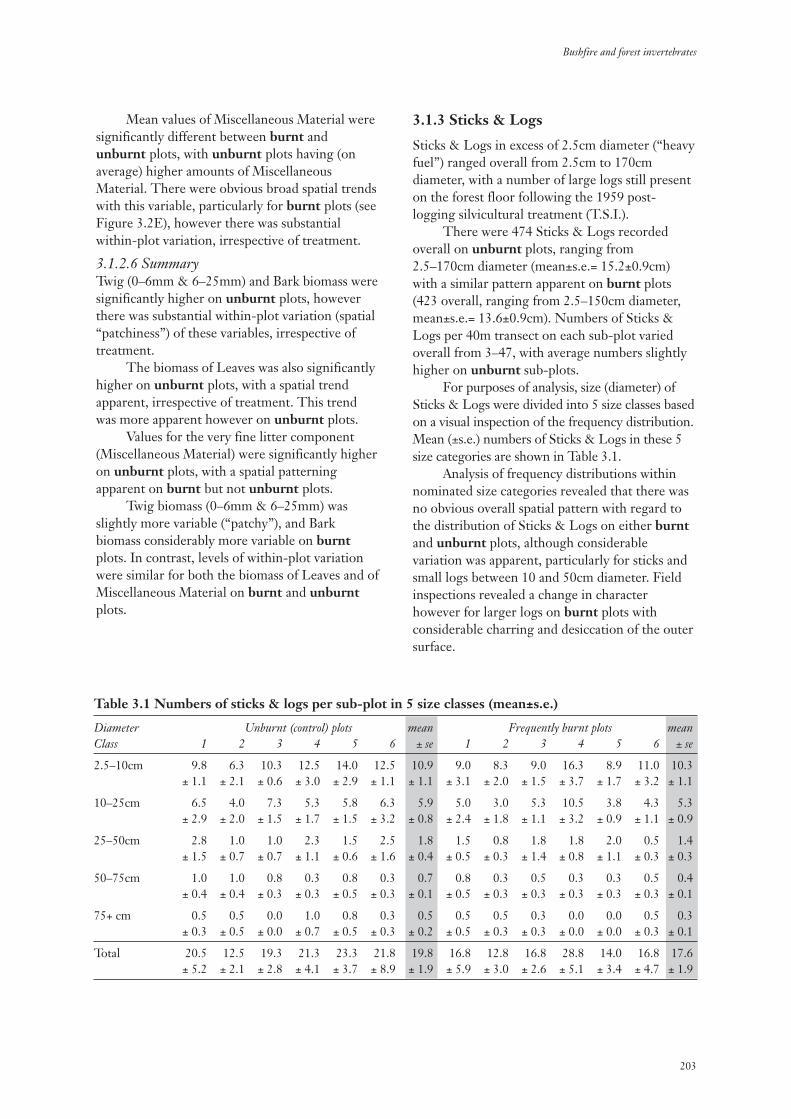
203
Bushfire and forest invertebrates
Mean values of Miscellaneous Material weresignificantly different between burnt andunburnt plots, with unburnt plots having (onaverage) higher amounts of MiscellaneousMaterial. There were obvious broad spatial trendswith this variable, particularly for burnt plots (seeFigure 3.2E), however there was substantialwithin-plot variation, irrespective of treatment.
3.1.2.6 SummaryTwig (0–6mm & 6–25mm) and Bark biomass weresignificantly higher on unburnt plots, howeverthere was substantial within-plot variation (spatial“patchiness”) of these variables, irrespective oftreatment.
The biomass of Leaves was also significantlyhigher on unburnt plots, with a spatial trendapparent, irrespective of treatment. This trendwas more apparent however on unburnt plots.
Values for the very fine litter component(Miscellaneous Material) were significantly higheron unburnt plots, with a spatial patterningapparent on burnt but not unburnt plots.
Twig biomass (0–6mm & 6–25mm) wasslightly more variable (“patchy”), and Barkbiomass considerably more variable on burntplots. In contrast, levels of within-plot variationwere similar for both the biomass of Leaves and ofMiscellaneous Material on burnt and unburntplots.
3.1.3 Sticks & Logs
Sticks & Logs in excess of 2.5cm diameter (“heavyfuel”) ranged overall from 2.5cm to 170cmdiameter, with a number of large logs still presenton the forest floor following the 1959 post-logging silvicultural treatment (T.S.I.).
There were 474 Sticks & Logs recordedoverall on unburnt plots, ranging from2.5–170cm diameter (mean±s.e.= 15.2±0.9cm)with a similar pattern apparent on burnt plots(423 overall, ranging from 2.5–150cm diameter,mean±s.e.= 13.6±0.9cm). Numbers of Sticks &Logs per 40m transect on each sub-plot variedoverall from 3–47, with average numbers slightlyhigher on unburnt sub-plots.
For purposes of analysis, size (diameter) ofSticks & Logs were divided into 5 size classes basedon a visual inspection of the frequency distribution.Mean (±s.e.) numbers of Sticks & Logs in these 5size categories are shown in Table 3.1.
Analysis of frequency distributions withinnominated size categories revealed that there wasno obvious overall spatial pattern with regard tothe distribution of Sticks & Logs on either burntand unburnt plots, although considerablevariation was apparent, particularly for sticks andsmall logs between 10 and 50cm diameter. Fieldinspections revealed a change in characterhowever for larger logs on burnt plots withconsiderable charring and desiccation of the outersurface.
Table 3.1 Numbers of sticks & logs per sub-plot in 5 size classes (mean±s.e.)
Diameter Unburnt (control) plots mean Frequently burnt plots meanClass 1 2 3 4 5 6 ± se 1 2 3 4 5 6 ± se
2.5–10cm 9.8 6.3 10.3 12.5 14.0 12.5 10.9 9.0 8.3 9.0 16.3 8.9 11.0 10.3± 1.1 ± 2.1 ± 0.6 ± 3.0 ± 2.9 ± 1.1 ± 1.1 ± 3.1 ± 2.0 ± 1.5 ± 3.7 ± 1.7 ± 3.2 ± 1.1
10–25cm 6.5 4.0 7.3 5.3 5.8 6.3 5.9 5.0 3.0 5.3 10.5 3.8 4.3 5.3± 2.9 ± 2.0 ± 1.5 ± 1.7 ± 1.5 ± 3.2 ± 0.8 ± 2.4 ± 1.8 ± 1.1 ± 3.2 ± 0.9 ± 1.1 ± 0.9
25–50cm 2.8 1.0 1.0 2.3 1.5 2.5 1.8 1.5 0.8 1.8 1.8 2.0 0.5 1.4± 1.5 ± 0.7 ± 0.7 ± 1.1 ± 0.6 ± 1.6 ± 0.4 ± 0.5 ± 0.3 ± 1.4 ± 0.8 ± 1.1 ± 0.3 ± 0.3
50–75cm 1.0 1.0 0.8 0.3 0.8 0.3 0.7 0.8 0.3 0.5 0.3 0.3 0.5 0.4± 0.4 ± 0.4 ± 0.3 ± 0.3 ± 0.5 ± 0.3 ± 0.1 ± 0.5 ± 0.3 ± 0.3 ± 0.3 ± 0.3 ± 0.3 ± 0.1
75+ cm 0.5 0.5 0.0 1.0 0.8 0.3 0.5 0.5 0.5 0.3 0.0 0.0 0.5 0.3± 0.3 ± 0.5 ± 0.0 ± 0.7 ± 0.5 ± 0.3 ± 0.2 ± 0.5 ± 0.3 ± 0.3 ± 0.0 ± 0.0 ± 0.3 ± 0.1
Total 20.5 12.5 19.3 21.3 23.3 21.8 19.8 16.8 12.8 16.8 28.8 14.0 16.8 17.6± 5.2 ± 2.1 ± 2.8 ± 4.1 ± 3.7 ± 8.9 ± 1.9 ± 5.9 ± 3.0 ± 2.6 ± 5.1 ± 3.4 ± 4.7 ± 1.9
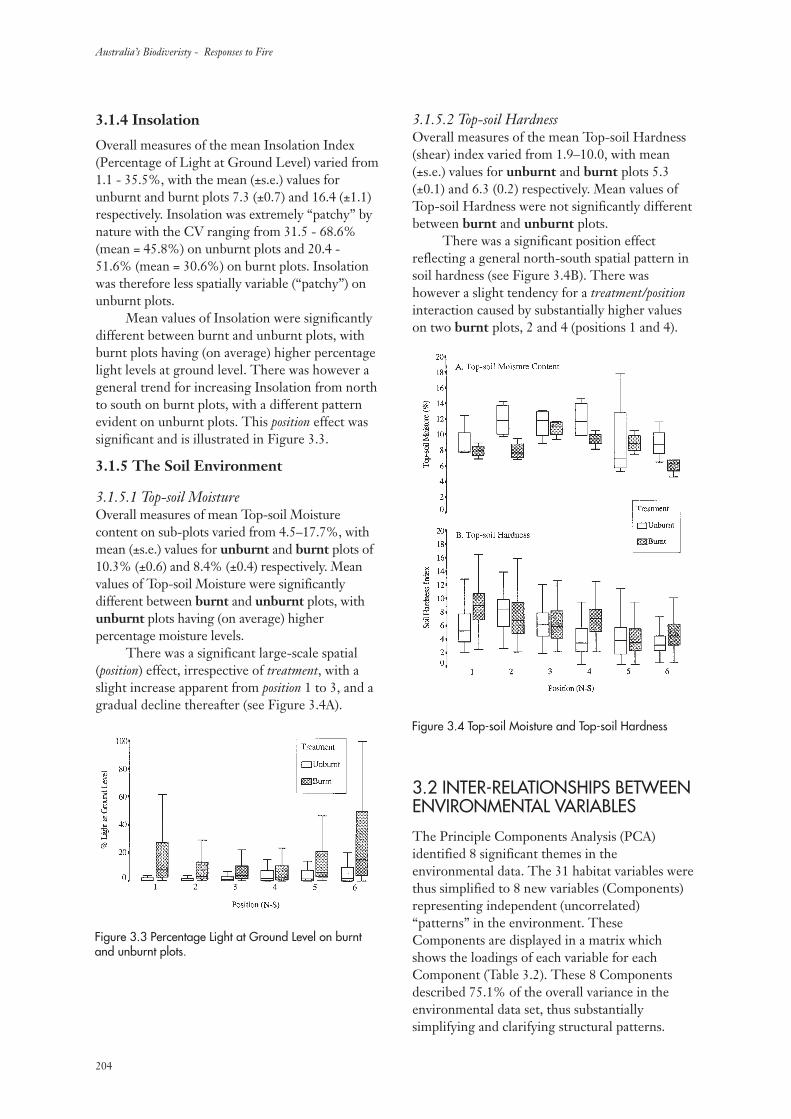
204
Australia’s Biodiveristy - Responses to Fire
3.1.4 Insolation
Overall measures of the mean Insolation Index(Percentage of Light at Ground Level) varied from1.1 - 35.5%, with the mean (±s.e.) values forunburnt and burnt plots 7.3 (±0.7) and 16.4 (±1.1)respectively. Insolation was extremely “patchy” bynature with the CV ranging from 31.5 - 68.6%(mean = 45.8%) on unburnt plots and 20.4 -51.6% (mean = 30.6%) on burnt plots. Insolationwas therefore less spatially variable (“patchy”) onunburnt plots.
Mean values of Insolation were significantlydifferent between burnt and unburnt plots, withburnt plots having (on average) higher percentagelight levels at ground level. There was however ageneral trend for increasing Insolation from northto south on burnt plots, with a different patternevident on unburnt plots. This position effect wassignificant and is illustrated in Figure 3.3.
3.1.5 The Soil Environment
3.1.5.1 Top-soil MoistureOverall measures of mean Top-soil Moisturecontent on sub-plots varied from 4.5–17.7%, withmean (±s.e.) values for unburnt and burnt plots of10.3% (±0.6) and 8.4% (±0.4) respectively. Meanvalues of Top-soil Moisture were significantlydifferent between burnt and unburnt plots, withunburnt plots having (on average) higherpercentage moisture levels.
There was a significant large-scale spatial(position) effect, irrespective of treatment, with aslight increase apparent from position 1 to 3, and agradual decline thereafter (see Figure 3.4A).
3.1.5.2 Top-soil HardnessOverall measures of the mean Top-soil Hardness(shear) index varied from 1.9–10.0, with mean(±s.e.) values for unburnt and burnt plots 5.3(±0.1) and 6.3 (0.2) respectively. Mean values ofTop-soil Hardness were not significantly differentbetween burnt and unburnt plots.
There was a significant position effectreflecting a general north-south spatial pattern insoil hardness (see Figure 3.4B). There washowever a slight tendency for a treatment/positioninteraction caused by substantially higher valueson two burnt plots, 2 and 4 (positions 1 and 4).
3.2 INTER-RELATIONSHIPS BETWEENENVIRONMENTAL VARIABLES
The Principle Components Analysis (PCA)identified 8 significant themes in theenvironmental data. The 31 habitat variables werethus simplified to 8 new variables (Components)representing independent (uncorrelated)“patterns” in the environment. TheseComponents are displayed in a matrix whichshows the loadings of each variable for eachComponent (Table 3.2). These 8 Componentsdescribed 75.1% of the overall variance in theenvironmental data set, thus substantiallysimplifying and clarifying structural patterns.
Figure 3.3 Percentage Light at Ground Level on burntand unburnt plots.
Figure 3.4 Top-soil Moisture and Top-soil Hardness

205
Bushfire and forest invertebrates
Tab
le 3
.2 S
igni
fican
t P
rinc
iple
Com
pone
nts
(eig
enva
lues
> 1
) sho
win
g va
riab
le lo
adin
gs >
0.3
Vari
able
Com
pone
nt 1
Com
pone
nt 2
Com
pone
nt 3
Com
pone
nt 4
Com
pone
nt 5
Com
pone
nt 6
Com
pone
nt 7
Com
pone
nt 8
Gro
und
herb
s–.
949
CV
Gro
und
herb
s.9
46Sm
all s
hrub
s–.
938
CV
Smal
l shr
ubs
.940
Mid
-siz
ed s
hrub
s–.
779
.367
CV
Mid
-siz
ed s
hrub
s.7
53A
spec
t.5
96–.
341
Twig
s 0–
6mm
.901
Twig
s 6–
10m
m.3
11.6
14L
eave
s.8
32B
ark
.686
CV
Bar
k–.
438
–.55
0.3
77Fi
ne li
tter
.777
.352
Tops
oil m
oist
ure
.567
PL
AG
–.68
4.3
39C
V P
LA
G.6
14.3
78
Tall
shru
bs–.
349
.357
.722
CV
Tal
l shr
ubs
–.73
4V
ery
tall
shru
bs.3
54.6
96
Stic
ks 0
.5–1
0cm
.638
–.43
7L
ogs
10–2
5cm
.861
Log
s 25
–50c
m.8
17
Tops
oil h
ardn
ess
–.31
6–.
818
CV
Top
soil
hard
ness
.851
CV
Lea
ves
.853
CV
Fin
e lit
ter
.847
CV
twig
s 0–
6mm
.721
CV
Tw
igs
6–10
mm
–.31
9.3
73.5
49C
V V
ery
tall
shru
bs–.
376
.529
Log
s 50
+cm
.645
Slop
e–.
447
–.59
1
Eig
enva
lue
7.12
76.
570
2.17
41.
903
1.66
51.
482
1.24
41.
120
% V
aria
nce
expl
aine
d23
.021
.27.
06.
15.
44.
84.
03.
6C
umul
ativ
e V
aria
nce
23.0
44.2
51.2
57.3
62.7
67.5
71.5
75.1
%

Table 3.3 Interpretation of environmentalPrincipal Components (based on identificationof variables with high loadings - see Table 3.2).
Component Interpretation
1 Absence and spatial patchiness ofGround Herbs, Small Shrubs andMid-sized Shrubs.
2 Abundance of litter (Twigs, Leaves,Bark & Miscellaneous Material [veryfine litter]); high Top-soil Moisture;low and spatially variable amounts ofInsolation at ground level.
3 Presence and spatial homogeneity ofTall Shrubs, presence of Very TallShrubs.
4 Presence of Sticks & Logs(0.5–10cm, 10–25cm & 25–50cmdiameter).
5 Low and spatially variable Top-soilHardness.
6 Spatial patchiness of Leaves andMiscellaneous Material [very finelitter].
7 Spatial patchiness of Twigs (0-6,6–10mm) and Very Tall Shrubs.
8 Presence of large logs (50+cmdiameter) and low slope angles.
As the first 3 Components together explainover half (51.2%) of the variance in theenvironmental data set (Table 3.3), theirComponent scores were plotted to see how theindividual sub-plots were responding (Figure 3.5).The complexity of 3-dimensions is displayed intwo plots for clarity. The “elevation” viewillustrates the projection of Components 1 & 2,representing 44.2% of the variance inenvironmental variables. The “plan” viewillustrates the projection of Components 1 & 3,representing 23% and 7% of the variancerespectively.
A number of features are apparent. Firstly, inthe elevation view, there is a distinct separation ofburnt and unburnt sub-plots along Component2. This reflects the distinct treatment effectdetected for Litter biomass (see 3.1.2). Thisseparation is not apparent along Components 1and 3, where there are no significant treatmenteffects. That is, the amount of vegetation in thefirst metre above ground is independent oftreatment (see 3.1.1.6). Secondly, sub-plots withineach plot are only loosely grouped together,
206
Australia’s Biodiveristy - Responses to Fire
The first Component concerns the absenceand spatial patchiness of Ground Herbs, SmallShrubs and Mid-sized Shrubs, and explains 23%of the variance in the environmental data set (seeTable 3.3). This Component describes a pattern inthe data whereby the first three structuralvegetation layers (0–1m) are positively correlated(ie. vary together). As the amount of vegetation inthese layers increases, it becomes more spatiallyhomogenous. Conversely, low amounts of thesestructural vegetation Components imply spatialpatchiness of the vegetation. There is a weaktendency for there to be greater amounts ofvegetation in the first metre above the ground onsub-plots with more exposed (north-westerly)aspects.
The second Component concerns theabundance of litter (Twigs, Bark, Leaves &Miscellaneous Material [very fine litter]), highTop-soil Moisture, and low and spatially patchyamounts of Insolation at ground level. Thisdescribes a pattern that with increasing litterlevels (all Components), top-soil moisture levelsincrease and the amount of insolation reachingground level decreases and becomes morespatially variable. This Component explains anadditional 21.2% of the variance in theenvironmental data set.
The third Component concerns the presenceand spatial homogeneity of Tall Shrubs, andpresence of Very Tall Shrubs, and explains anadditional 7% of the variance in the environmentaldata set. This Component describes a patternwhereby when vegetation occurs in the upperunderstorey layers (1–2m) it tends to occur in bothelements of these layers, however it is spatiallypatchy.
Subsequent Components do not explainsubstantial additional variance in theenvironmental data set, however they do illustratethat many of the patterns in the habitat data areindependent (ie. uncorrelated). For example, thenumber of Sticks & Logs in certain size categories(Component 4) are inter-correlated and unrelatedto the amount of Litter biomass at sub-plots(Component 2). Top-soil Hardness (Component5) is site-specific and independent of otherenvironmental variables. The presence of largelogs on sites (Component 8) is also site-specific,although there is a tendency for a greater numberon sub-plots with lower slope angles.
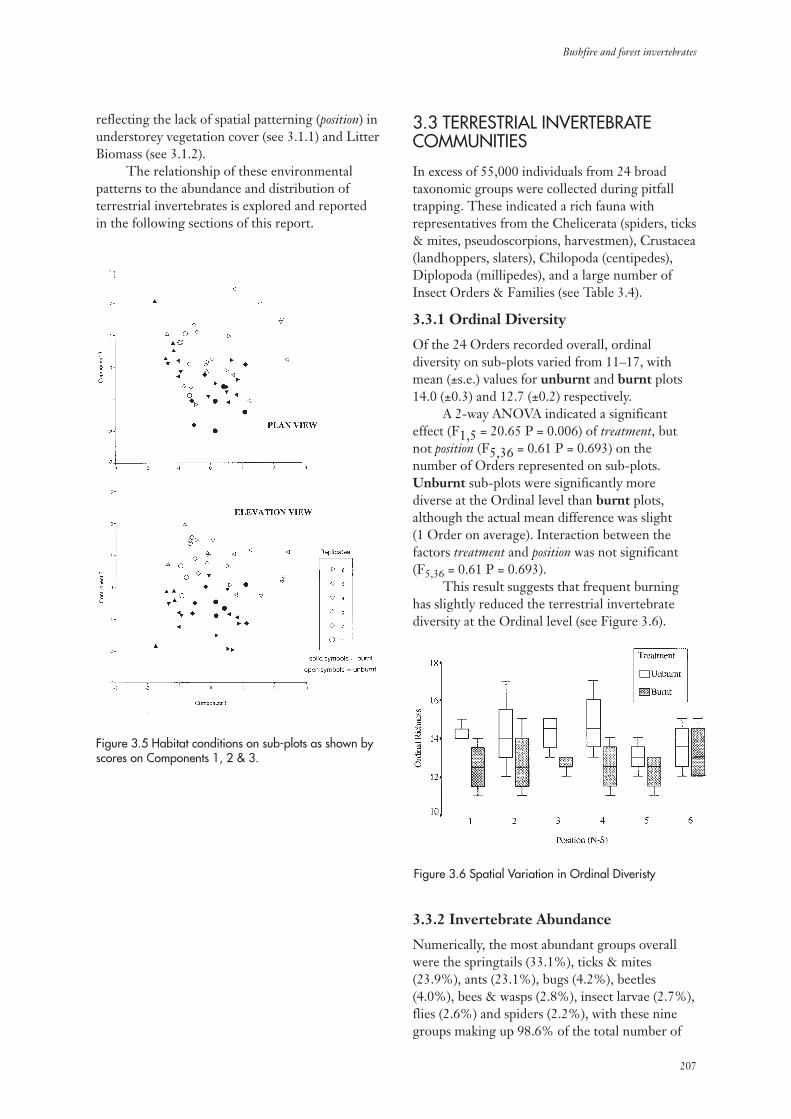
207
Bushfire and forest invertebrates
reflecting the lack of spatial patterning (position) inunderstorey vegetation cover (see 3.1.1) and LitterBiomass (see 3.1.2).
The relationship of these environmentalpatterns to the abundance and distribution ofterrestrial invertebrates is explored and reportedin the following sections of this report.
3.3 TERRESTRIAL INVERTEBRATECOMMUNITIES
In excess of 55,000 individuals from 24 broadtaxonomic groups were collected during pitfalltrapping. These indicated a rich fauna withrepresentatives from the Chelicerata (spiders, ticks& mites, pseudoscorpions, harvestmen), Crustacea(landhoppers, slaters), Chilopoda (centipedes),Diplopoda (millipedes), and a large number ofInsect Orders & Families (see Table 3.4).
3.3.1 Ordinal Diversity
Of the 24 Orders recorded overall, ordinaldiversity on sub-plots varied from 11–17, withmean (±s.e.) values for unburnt and burnt plots14.0 (±0.3) and 12.7 (±0.2) respectively.
A 2-way ANOVA indicated a significanteffect (F1,5 = 20.65 P = 0.006) of treatment, butnot position (F5,36 = 0.61 P = 0.693) on thenumber of Orders represented on sub-plots.Unburnt sub-plots were significantly morediverse at the Ordinal level than burnt plots,although the actual mean difference was slight (1 Order on average). Interaction between thefactors treatment and position was not significant(F5,36 = 0.61 P = 0.693).
This result suggests that frequent burninghas slightly reduced the terrestrial invertebratediversity at the Ordinal level (see Figure 3.6).
3.3.2 Invertebrate Abundance
Numerically, the most abundant groups overallwere the springtails (33.1%), ticks & mites(23.9%), ants (23.1%), bugs (4.2%), beetles(4.0%), bees & wasps (2.8%), insect larvae (2.7%),flies (2.6%) and spiders (2.2%), with these ninegroups making up 98.6% of the total number of
Figure 3.5 Habitat conditions on sub-plots as shown byscores on Components 1, 2 & 3.
Figure 3.6 Spatial Variation in Ordinal Diveristy
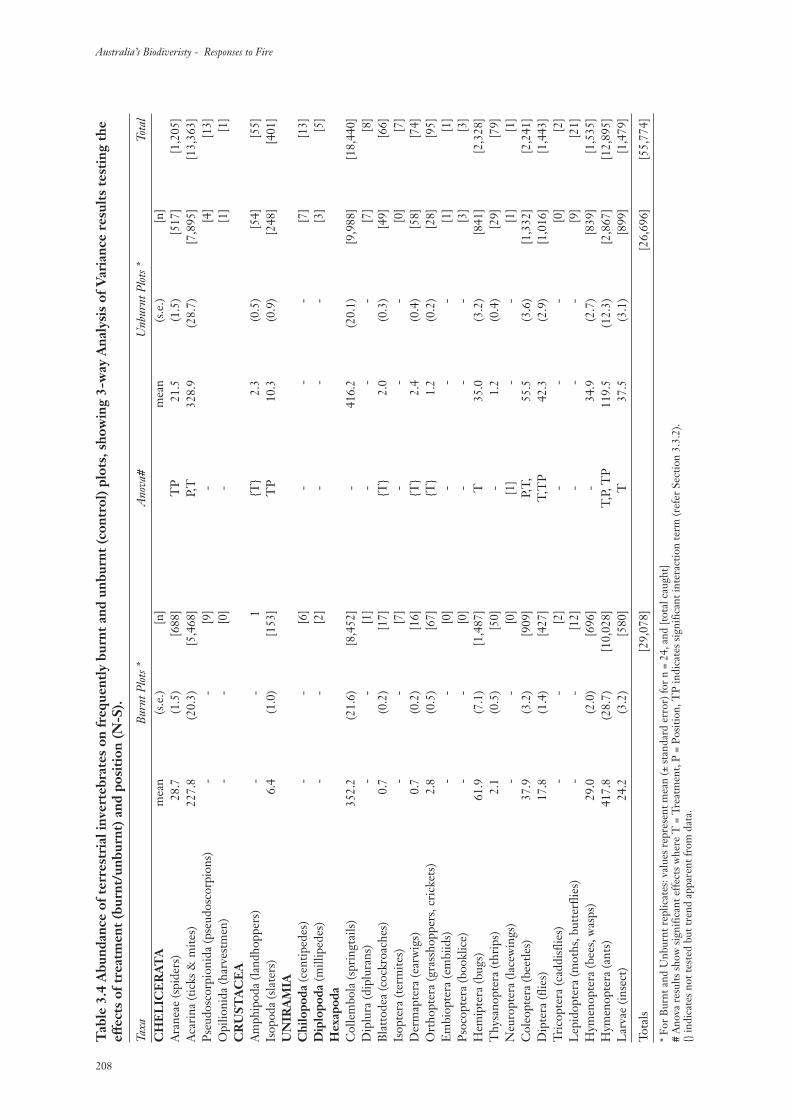
208
Australia’s Biodiveristy - Responses to Fire
Tab
le 3
.4 A
bund
ance
of t
erre
stri
al in
vert
ebra
tes
on fr
eque
ntly
bur
nt a
nd u
nbur
nt (c
ontr
ol) p
lots
, sho
win
g 3-
way
Ana
lysi
s of
Var
ianc
e re
sult
s te
stin
g th
eef
fect
s of
tre
atm
ent
(bur
nt/u
nbur
nt) a
nd p
osit
ion
(N-S
).
Taxa
Burn
t Plo
ts *
Ano
va#
Unb
urnt
Plo
ts *
Tota
lC
HE
LIC
ER
ATA
mea
n(s
.e.)
[n]
mea
n(s
.e.)
[n]
Ara
neae
(spi
ders
)28
.7(1
.5)
[688
]T
P21
.5(1
.5)
[517
][1
,205
]A
cari
na (t
icks
& m
ites)
227.
8(2
0.3)
[5,4
68]
P,T
328.
9(2
8.7)
[7,8
95]
[13,
363]
Pse
udos
corp
ioni
da (p
seud
osco
rpio
ns)
--
[9]
-[4
][1
3]O
pilio
nida
(har
vest
men
)-
-[0
]-
[1]
[1]
CR
UST
AC
EA
Am
phip
oda
(land
hopp
ers)
--
1{T
}2.
3(0
.5)
[54]
[55]
Isop
oda
(sla
ters
)6.
4(1
.0)
[153
]T
P10
.3(0
.9)
[248
][4
01]
UN
IRA
MIA
Chi
lopo
da(c
entip
edes
)-
-[6
]-
--
[7]
[13]
Dip
lopo
da (m
illip
edes
)-
-[2
]-
--
[3]
[5]
Hex
apod
aC
olle
mbo
la (s
prin
gtai
ls)
352.
2(2
1.6)
[8,4
52]
-41
6.2
(20.
1)[9
,988
][1
8,44
0]D
iplu
ra (d
iplu
rans
)-
-[1
]-
--
[7]
[8]
Bla
ttod
ea (c
ockr
oach
es)
0.7
(0.2
)[1
7]{T
}2.
0(0
.3)
[49]
[66]
Isop
tera
(ter
mite
s)-
-[7
]-
--
[0]
[7]
Der
map
tera
(ear
wig
s)0.
7(0
.2)
[16]
{T}
2.4
(0.4
)[5
8][7
4]O
rtho
pter
a (g
rass
hopp
ers,
cri
cket
s)2.
8(0
.5)
[67]
{T}
1.2
(0.2
)[2
8][9
5]E
mbi
opte
ra (e
mbi
ids)
--
[0]
--
-[1
][1
]P
soco
pter
a (b
ookl
ice)
--
[0]
--
-[3
][3
]H
emip
tera
(bug
s)61
.9(7
.1)
[1,4
87]
T35
.0(3
.2)
[841
][2
,328
]T
hysa
nopt
era
(thr
ips)
2.1
(0.5
)[5
0]-
1.2
(0.4
)[2
9][7
9]N
euro
pter
a (la
cew
ings
)-
-[0
][1
]-
-[1
][1
]C
oleo
pter
a (b
eetle
s)37
.9(3
.2)
[909
]P,
T,
55.5
(3.6
)[1
,332
][2
,241
]D
ipte
ra (f
lies)
17.8
(1.4
)[4
27]
T,T
P42
.3(2
.9)
[1,0
16]
[1,4
43]
Tri
copt
era
(cad
disf
lies)
--
[2]
--
-[0
][2
]L
epid
opte
ra (m
oths
, but
terf
lies)
--
[12]
--
-[9
][2
1]H
ymen
opte
ra (b
ees,
was
ps)
29.0
(2.0
)[6
96]
-34
.9(2
.7)
[839
][1
,535
]H
ymen
opte
ra (a
nts)
417.
8(2
8.7)
[10,
028]
T,P
, TP
119.
5(1
2.3)
[2,8
67]
[12,
895]
Lar
vae
(inse
ct)
24.2
(3.2
)[5
80]
T37
.5(3
.1)
[899
][1
,479
]
Tota
ls[2
9,07
8][2
6,69
6][5
5,77
4]
* Fo
r B
urnt
and
Unb
urnt
rep
licat
es: v
alue
s re
pres
ent m
ean
(±st
anda
rd e
rror
) for
n =
24,
and
[tot
al c
augh
t]#
Ano
va r
esul
ts s
how
sig
nific
ant e
ffec
ts w
here
T =
Tre
atm
ent,
P =
Pos
ition
, TP
indi
cate
s si
gnifi
cant
inte
ract
ion
term
(ref
er S
ectio
n 3.
3.2)
. {}
indi
cate
s no
t tes
ted
but t
rend
app
aren
t fro
m d
ata.
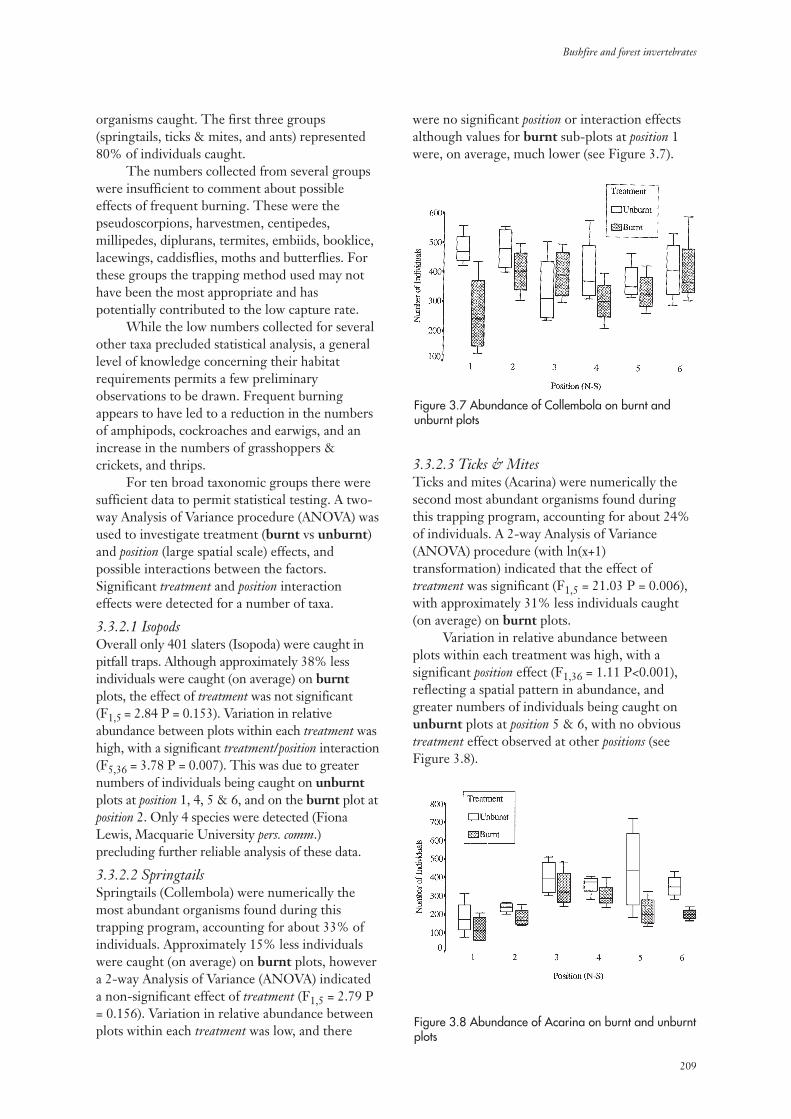
209
Bushfire and forest invertebrates
organisms caught. The first three groups(springtails, ticks & mites, and ants) represented80% of individuals caught.
The numbers collected from several groupswere insufficient to comment about possibleeffects of frequent burning. These were thepseudoscorpions, harvestmen, centipedes,millipedes, diplurans, termites, embiids, booklice,lacewings, caddisflies, moths and butterflies. Forthese groups the trapping method used may nothave been the most appropriate and haspotentially contributed to the low capture rate.
While the low numbers collected for severalother taxa precluded statistical analysis, a generallevel of knowledge concerning their habitatrequirements permits a few preliminaryobservations to be drawn. Frequent burningappears to have led to a reduction in the numbersof amphipods, cockroaches and earwigs, and anincrease in the numbers of grasshoppers &crickets, and thrips.
For ten broad taxonomic groups there weresufficient data to permit statistical testing. A two-way Analysis of Variance procedure (ANOVA) wasused to investigate treatment (burnt vs unburnt)and position (large spatial scale) effects, andpossible interactions between the factors.Significant treatment and position interactioneffects were detected for a number of taxa.
3.3.2.1 IsopodsOverall only 401 slaters (Isopoda) were caught inpitfall traps. Although approximately 38% lessindividuals were caught (on average) on burntplots, the effect of treatment was not significant(F1,5 = 2.84 P = 0.153). Variation in relativeabundance between plots within each treatment washigh, with a significant treatment/position interaction(F5,36 = 3.78 P = 0.007). This was due to greaternumbers of individuals being caught on unburntplots at position 1, 4, 5 & 6, and on the burnt plot atposition 2. Only 4 species were detected (FionaLewis, Macquarie University pers. comm.)precluding further reliable analysis of these data.
3.3.2.2 SpringtailsSpringtails (Collembola) were numerically themost abundant organisms found during thistrapping program, accounting for about 33% ofindividuals. Approximately 15% less individualswere caught (on average) on burnt plots, howevera 2-way Analysis of Variance (ANOVA) indicateda non-significant effect of treatment (F1,5 = 2.79 P= 0.156). Variation in relative abundance betweenplots within each treatment was low, and there
were no significant position or interaction effectsalthough values for burnt sub-plots at position 1were, on average, much lower (see Figure 3.7).
3.3.2.3 Ticks & MitesTicks and mites (Acarina) were numerically thesecond most abundant organisms found duringthis trapping program, accounting for about 24%of individuals. A 2-way Analysis of Variance(ANOVA) procedure (with ln(x+1)transformation) indicated that the effect oftreatment was significant (F1,5 = 21.03 P = 0.006),with approximately 31% less individuals caught(on average) on burnt plots.
Variation in relative abundance betweenplots within each treatment was high, with asignificant position effect (F1,36 = 1.11 P<0.001),reflecting a spatial pattern in abundance, andgreater numbers of individuals being caught onunburnt plots at position 5 & 6, with no obvioustreatment effect observed at other positions (seeFigure 3.8).
Figure 3.7 Abundance of Collembola on burnt andunburnt plots
Figure 3.8 Abundance of Acarina on burnt and unburntplots
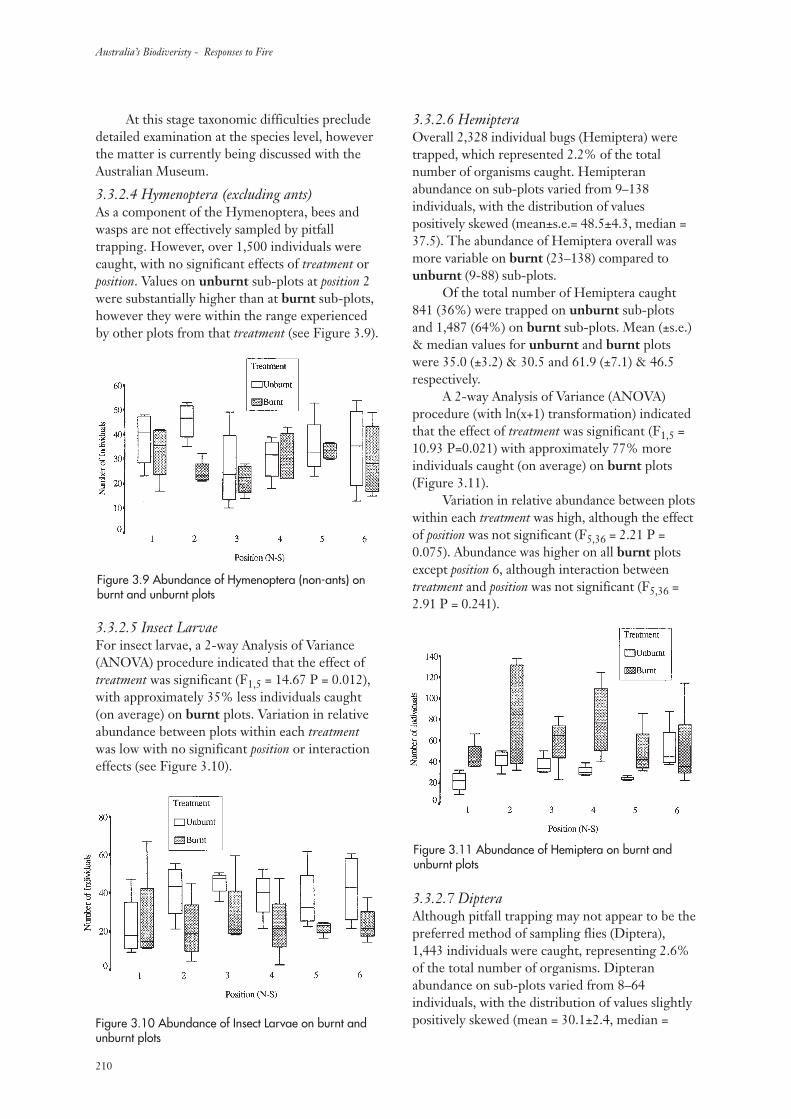
3.3.2.6 HemipteraOverall 2,328 individual bugs (Hemiptera) weretrapped, which represented 2.2% of the totalnumber of organisms caught. Hemipteranabundance on sub-plots varied from 9–138individuals, with the distribution of valuespositively skewed (mean±s.e.= 48.5±4.3, median =37.5). The abundance of Hemiptera overall wasmore variable on burnt (23–138) compared tounburnt (9-88) sub-plots.
Of the total number of Hemiptera caught841 (36%) were trapped on unburnt sub-plotsand 1,487 (64%) on burnt sub-plots. Mean (±s.e.)& median values for unburnt and burnt plotswere 35.0 (±3.2) & 30.5 and 61.9 (±7.1) & 46.5respectively.
A 2-way Analysis of Variance (ANOVA)procedure (with ln(x+1) transformation) indicatedthat the effect of treatment was significant (F1,5 =10.93 P=0.021) with approximately 77% moreindividuals caught (on average) on burnt plots(Figure 3.11).
Variation in relative abundance between plotswithin each treatment was high, although the effectof position was not significant (F5,36 = 2.21 P =0.075). Abundance was higher on all burnt plotsexcept position 6, although interaction betweentreatment and position was not significant (F5,36 =2.91 P = 0.241).
3.3.2.7 DipteraAlthough pitfall trapping may not appear to be thepreferred method of sampling flies (Diptera),1,443 individuals were caught, representing 2.6%of the total number of organisms. Dipteranabundance on sub-plots varied from 8–64individuals, with the distribution of values slightlypositively skewed (mean = 30.1±2.4, median =
210
Australia’s Biodiveristy - Responses to Fire
At this stage taxonomic difficulties precludedetailed examination at the species level, howeverthe matter is currently being discussed with theAustralian Museum.
3.3.2.4 Hymenoptera (excluding ants)As a component of the Hymenoptera, bees andwasps are not effectively sampled by pitfalltrapping. However, over 1,500 individuals werecaught, with no significant effects of treatment orposition. Values on unburnt sub-plots at position 2were substantially higher than at burnt sub-plots,however they were within the range experiencedby other plots from that treatment (see Figure 3.9).
3.3.2.5 Insect LarvaeFor insect larvae, a 2-way Analysis of Variance(ANOVA) procedure indicated that the effect oftreatment was significant (F1,5 = 14.67 P = 0.012),with approximately 35% less individuals caught(on average) on burnt plots. Variation in relativeabundance between plots within each treatmentwas low with no significant position or interactioneffects (see Figure 3.10).
Figure 3.9 Abundance of Hymenoptera (non-ants) onburnt and unburnt plots
Figure 3.10 Abundance of Insect Larvae on burnt andunburnt plots
Figure 3.11 Abundance of Hemiptera on burnt andunburnt plots

211
Bushfire and forest invertebrates
28.0). The abundance of Diptera overall was morevariable on unburnt (9–64) compared with burnt(8–34) sub-plots.
Of the total number of Diptera caught 1,016(70%) were trapped on unburnt sub-plots and427 (30%) on burnt sub-plots. Mean (±s.e.) &median values for unburnt and burnt plots were42.3 (±2.9) & 43.0 and 17.8 (±1.4) & 16.0respectively.
A 2-way Analysis of Variance (ANOVA)procedure indicated that the effect of treatmentwas significant (F1,5 = 24.30 P = 0.004), withapproximately 58% less individuals caught (onaverage) on burnt plots.
Variation in relative abundance betweenplots within each treatment was high, with amarginally non-significant position effect (F5,36 =2.22 P = 0.073) and a significant treatment/positioninteraction effect (F5,36 = 3.49 P = 0.011). Thiswas due to greater numbers of individuals beingcaught on unburnt plots at position 2, 3, 4, 5 & 6,and on the burnt plot at position 1 (see Figure3.12).
3.3.2.8 SpidersOverall 1,205 individual spiders (Araneae) weretrapped, which represented 2.2% of the totalnumber of organisms caught. Spider abundance onsub-plots varied from 8–42 individuals, with thedistribution of values slightly positively skewed(mean ±s.e= 25.1±1.1, median = 23.0). Theabundance of spiders overall was slightly morevariable on unburnt (8-40) compared with burnt(16–42) sub-plots.
Of the total number of spiders caught 515(43%) were trapped on unburnt sub-plots and688 (57%) on burnt sub-plots. Mean (±s.e.) &median values for unburnt and burnt plots were
21.5 (±1.5) & 22.0 and 28.7±1.5 & 28.5respectively. There were (on average) 33% morespiders on frequently burnt sub-plots.
A 2-way ANOVA indicated a non-significanteffect of treatment (F1,5 = 1.48 P = 0.278) andposition (F5,36 = 1.45 P = 0.229), with a significantinteraction detected between these two factors(F5,36 = 3.24 P = 0.016).
Mean values of spider abundance weretherefore not significantly different betweenburnt and unburnt plots, although burnt plotshave (on average) higher numbers of individuals(see Figure 3.13). The significant interactionbetween treatment and position reflects the reversalin the pattern of the treatment effect at position 3and 4, where burnt plots had slightly lower spiderabundance than unburnt, and at position 6 wherethe mean value on burnt plots (11.3±1.7) issubstantially higher than on unburnt plots(4.8±0.5).
3.3.2.9 BeetlesOverall 2,145 individual beetles were trapped,which represented 4.0% of the total number oforganisms caught. Beetle abundance on sub-plotsvaried from 13–88 individuals, with the with thedistribution of values slightly positively skewed(mean = 44.7, median = 43.5). The abundance ofbeetles overall was slightly less variable on burnt(13–67) compared with unburnt (29–88) sub-plots.
Of the total number of beetles caught, 1,269(59%) were trapped on unburnt sub-plots and876 (41%) on burnt sub-plots. Mean (±s.e.) &median values for unburnt and burnt plots were52.9 (±3.2) & 47.5 and 36.5 (±2.9) & 34.0respectively. There were (on average) 31% lessbeetles on frequently burnt sub-plots.
Figure 3.13 Abundance of Spiders on burnt andunburnt plots
Figure 3.12 Abundance of Diptera on burnt andunburnt plots
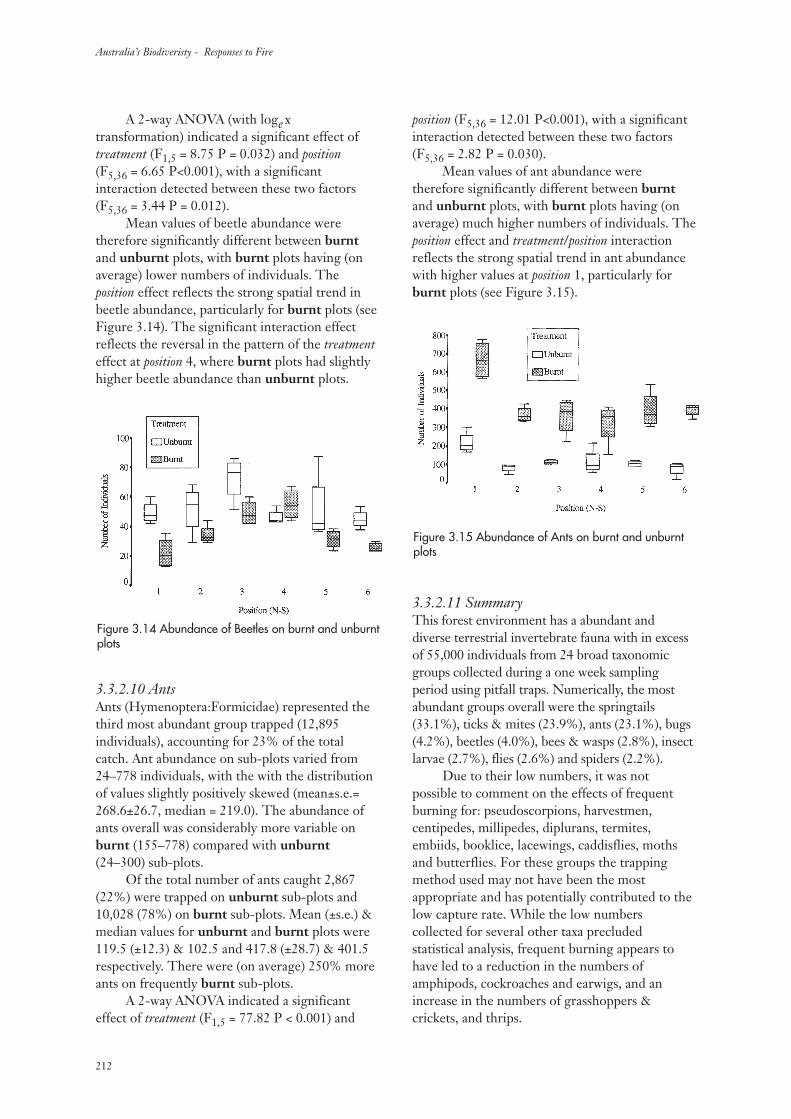
position (F5,36 = 12.01 P<0.001), with a significantinteraction detected between these two factors(F5,36 = 2.82 P = 0.030).
Mean values of ant abundance weretherefore significantly different between burntand unburnt plots, with burnt plots having (onaverage) much higher numbers of individuals. Theposition effect and treatment/position interactionreflects the strong spatial trend in ant abundancewith higher values at position 1, particularly forburnt plots (see Figure 3.15).
3.3.2.11 SummaryThis forest environment has a abundant anddiverse terrestrial invertebrate fauna with in excessof 55,000 individuals from 24 broad taxonomicgroups collected during a one week samplingperiod using pitfall traps. Numerically, the mostabundant groups overall were the springtails(33.1%), ticks & mites (23.9%), ants (23.1%), bugs(4.2%), beetles (4.0%), bees & wasps (2.8%), insectlarvae (2.7%), flies (2.6%) and spiders (2.2%).
Due to their low numbers, it was notpossible to comment on the effects of frequentburning for: pseudoscorpions, harvestmen,centipedes, millipedes, diplurans, termites,embiids, booklice, lacewings, caddisflies, mothsand butterflies. For these groups the trappingmethod used may not have been the mostappropriate and has potentially contributed to thelow capture rate. While the low numberscollected for several other taxa precludedstatistical analysis, frequent burning appears tohave led to a reduction in the numbers ofamphipods, cockroaches and earwigs, and anincrease in the numbers of grasshoppers &crickets, and thrips.
212
Australia’s Biodiveristy - Responses to Fire
A 2-way ANOVA (with logextransformation) indicated a significant effect oftreatment (F1,5 = 8.75 P = 0.032) and position(F5,36 = 6.65 P<0.001), with a significantinteraction detected between these two factors(F5,36 = 3.44 P = 0.012).
Mean values of beetle abundance weretherefore significantly different between burntand unburnt plots, with burnt plots having (onaverage) lower numbers of individuals. Theposition effect reflects the strong spatial trend inbeetle abundance, particularly for burnt plots (seeFigure 3.14). The significant interaction effectreflects the reversal in the pattern of the treatmenteffect at position 4, where burnt plots had slightlyhigher beetle abundance than unburnt plots.
3.3.2.10 AntsAnts (Hymenoptera:Formicidae) represented thethird most abundant group trapped (12,895individuals), accounting for 23% of the totalcatch. Ant abundance on sub-plots varied from24–778 individuals, with the with the distributionof values slightly positively skewed (mean±s.e.=268.6±26.7, median = 219.0). The abundance ofants overall was considerably more variable onburnt (155–778) compared with unburnt(24–300) sub-plots.
Of the total number of ants caught 2,867(22%) were trapped on unburnt sub-plots and10,028 (78%) on burnt sub-plots. Mean (±s.e.) &median values for unburnt and burnt plots were119.5 (±12.3) & 102.5 and 417.8 (±28.7) & 401.5respectively. There were (on average) 250% moreants on frequently burnt sub-plots.
A 2-way ANOVA indicated a significanteffect of treatment (F1,5 = 77.82 P < 0.001) and
Figure 3.15 Abundance of Ants on burnt and unburntplots
Figure 3.14 Abundance of Beetles on burnt and unburntplots

213
Bushfire and forest invertebrates
Overall, the number of invertebrate Ordershad been significantly reduced on sub-plotsexperiencing frequent burning, although themagnitude of this decrease was small (average ≈ 1Order).
For ten broad taxonomic groups there weresufficient data to permit statistical testing usingthe Analysis of Variance (ANOVA) procedure toinvestigate the effects of frequent burning(treatment) and patterns due to large-scale spatialeffects (position). These results are summarised inTable 3.5 and indicate a variety of responses tofrequent burning. Seven groups (isopods,springtails, ticks & mites, bees & wasps, insectlarvae, flies and beetles) showed substantialdecreases in abundance following frequentburning. These decreases ranged from 15 to 58%,but were only statistically significant for ticks &mites, insect larvae, flies and beetles. High spatialvariability in abundance for isopods, springtails,and bees & wasps possibly contributed to the lackof statistical significance.
Three groups showed substantial increasesin abundance following frequent burning. Thesewere statistically significant for bugs (77%) and ants(250%), but not for spiders (33%). Both spidersand ants showed considerable spatial variability intheir numbers.
3.3.3 Invertebrate Species Richness
Five invertebrate groups were identified tomorphospecies (see Section 2.4) in order tofurther investigate the impact of repeated burningon species richness, and the related aspects ofcommunity composition and structure. Thesegroups utilise a diversity of micro-habitats andniches and are representative of the range ofterrestrial invertebrates found in these forestenvironments.
The groups studied at morphospecies levelwere the:Hemiptera (bugs):
as most are terrestrial and phytophagous(plant-feeding), they are a group which havea close association with plant communities.
Diptera (flies):although highly mobile as adults, the flieshave particular requirements with regard tolarval food sources; usually moist, decayingplant and animal material. Many species areparasitic on the larvae of other insect orders.
Araneae (spiders):spiders are a major group of predators inforest ecosystems exploiting a variety ofhabitats. They live in burrows or crevices inthe ground, amongst leaf litter or invegetation.
Coleoptera (beetles):beetles utilise a diverse range of habitats µ- habitats, with a variety of feedingstrategies (adults include herbivores,predators & scavengers, while larval formsfeed either internally or externally on plantsand fungal products).
Formicidae (ants):ants are one of the most numerous andwidespread groups in Australian ecosystems.They have a diverse diet, and utilise a varietyof feeding strategies from predators andscavengers, to plant eaters and fungusfeeders, with frequent and variedinteractions with other invertebrate groups.
These groups are discussed separately withrelated issues considered in Sections 3.3.3.6 & 7.
Table 3.5 Changes in mean abundance following frequent burning for selected terrestrialinvertebrate taxa.
Change with Statistically Large-scale Taxa Common name frequent burning significant ? (P<0.05) spatial patterns ?
Isopoda slaters, pill-bugs ⇓ 38% no yesCollembola springtails ⇓ 15% no noAcarina ticks & mites ⇓ 31% yes yesHymenoptera (excl. ants) bees & wasps ⇓ 17% no yesInsecta insect larvae ⇓ 35% yes noHemiptera bugs ⇑ 77% yes noDiptera flies ⇓ 58% yes no*Araneae spiders ⇑ 33% no yes*Coleoptera beetles ⇓ 31% yes yes*Formicidae ants ⇑ 250% yes yes*
* indicates a statistically significant interaction between treatment and position effects.
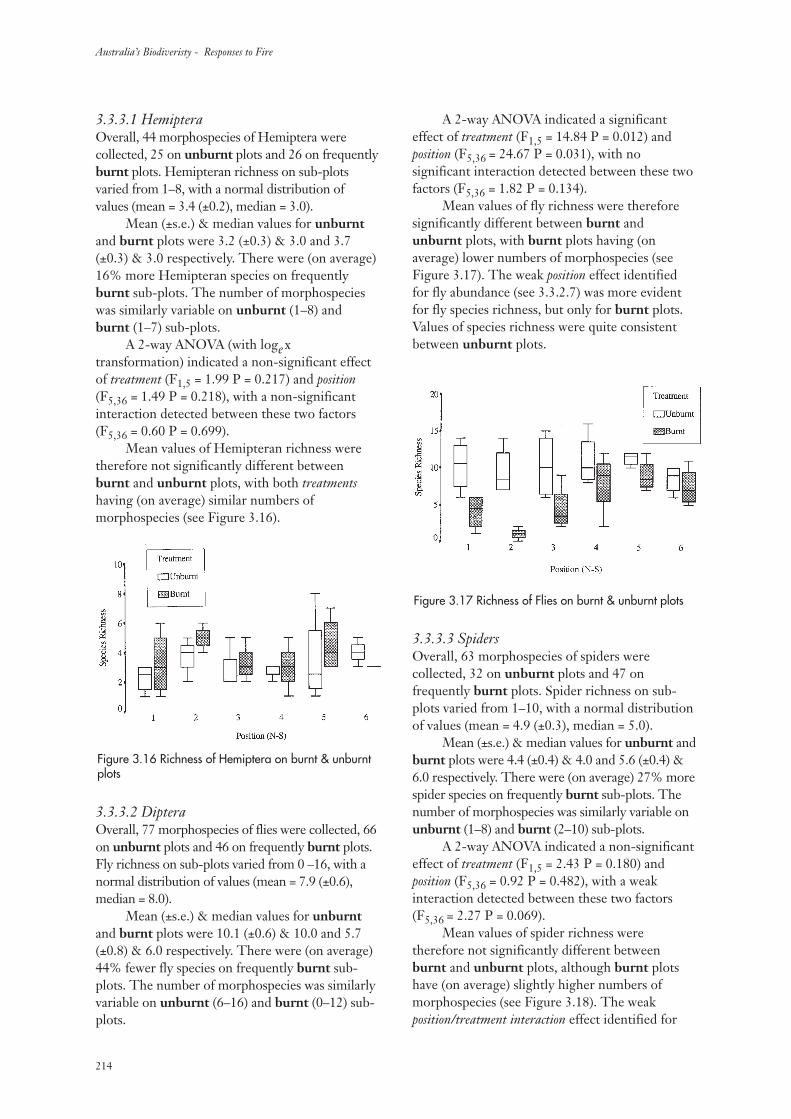
A 2-way ANOVA indicated a significanteffect of treatment (F1,5 = 14.84 P = 0.012) andposition (F5,36 = 24.67 P = 0.031), with nosignificant interaction detected between these twofactors (F5,36 = 1.82 P = 0.134).
Mean values of fly richness were thereforesignificantly different between burnt andunburnt plots, with burnt plots having (onaverage) lower numbers of morphospecies (seeFigure 3.17). The weak position effect identifiedfor fly abundance (see 3.3.2.7) was more evidentfor fly species richness, but only for burnt plots.Values of species richness were quite consistentbetween unburnt plots.
3.3.3.3 SpidersOverall, 63 morphospecies of spiders werecollected, 32 on unburnt plots and 47 onfrequently burnt plots. Spider richness on sub-plots varied from 1–10, with a normal distributionof values (mean = 4.9 (±0.3), median = 5.0).
Mean (±s.e.) & median values for unburnt andburnt plots were 4.4 (±0.4) & 4.0 and 5.6 (±0.4) &6.0 respectively. There were (on average) 27% morespider species on frequently burnt sub-plots. Thenumber of morphospecies was similarly variable onunburnt (1–8) and burnt (2–10) sub-plots.
A 2-way ANOVA indicated a non-significanteffect of treatment (F1,5 = 2.43 P = 0.180) andposition (F5,36 = 0.92 P = 0.482), with a weakinteraction detected between these two factors(F5,36 = 2.27 P = 0.069).
Mean values of spider richness weretherefore not significantly different betweenburnt and unburnt plots, although burnt plotshave (on average) slightly higher numbers ofmorphospecies (see Figure 3.18). The weakposition/treatment interaction effect identified for
214
Australia’s Biodiveristy - Responses to Fire
3.3.3.1 HemipteraOverall, 44 morphospecies of Hemiptera werecollected, 25 on unburnt plots and 26 on frequentlyburnt plots. Hemipteran richness on sub-plotsvaried from 1–8, with a normal distribution ofvalues (mean = 3.4 (±0.2), median = 3.0).
Mean (±s.e.) & median values for unburntand burnt plots were 3.2 (±0.3) & 3.0 and 3.7(±0.3) & 3.0 respectively. There were (on average)16% more Hemipteran species on frequentlyburnt sub-plots. The number of morphospecieswas similarly variable on unburnt (1–8) andburnt (1–7) sub-plots.
A 2-way ANOVA (with logextransformation) indicated a non-significant effectof treatment (F1,5 = 1.99 P = 0.217) and position(F5,36 = 1.49 P = 0.218), with a non-significantinteraction detected between these two factors(F5,36 = 0.60 P = 0.699).
Mean values of Hemipteran richness weretherefore not significantly different betweenburnt and unburnt plots, with both treatmentshaving (on average) similar numbers ofmorphospecies (see Figure 3.16).
3.3.3.2 DipteraOverall, 77 morphospecies of flies were collected, 66on unburnt plots and 46 on frequently burnt plots.Fly richness on sub-plots varied from 0 –16, with anormal distribution of values (mean = 7.9 (±0.6),median = 8.0).
Mean (±s.e.) & median values for unburntand burnt plots were 10.1 (±0.6) & 10.0 and 5.7(±0.8) & 6.0 respectively. There were (on average)44% fewer fly species on frequently burnt sub-plots. The number of morphospecies was similarlyvariable on unburnt (6–16) and burnt (0–12) sub-plots.
Figure 3.17 Richness of Flies on burnt & unburnt plots
Figure 3.16 Richness of Hemiptera on burnt & unburntplots
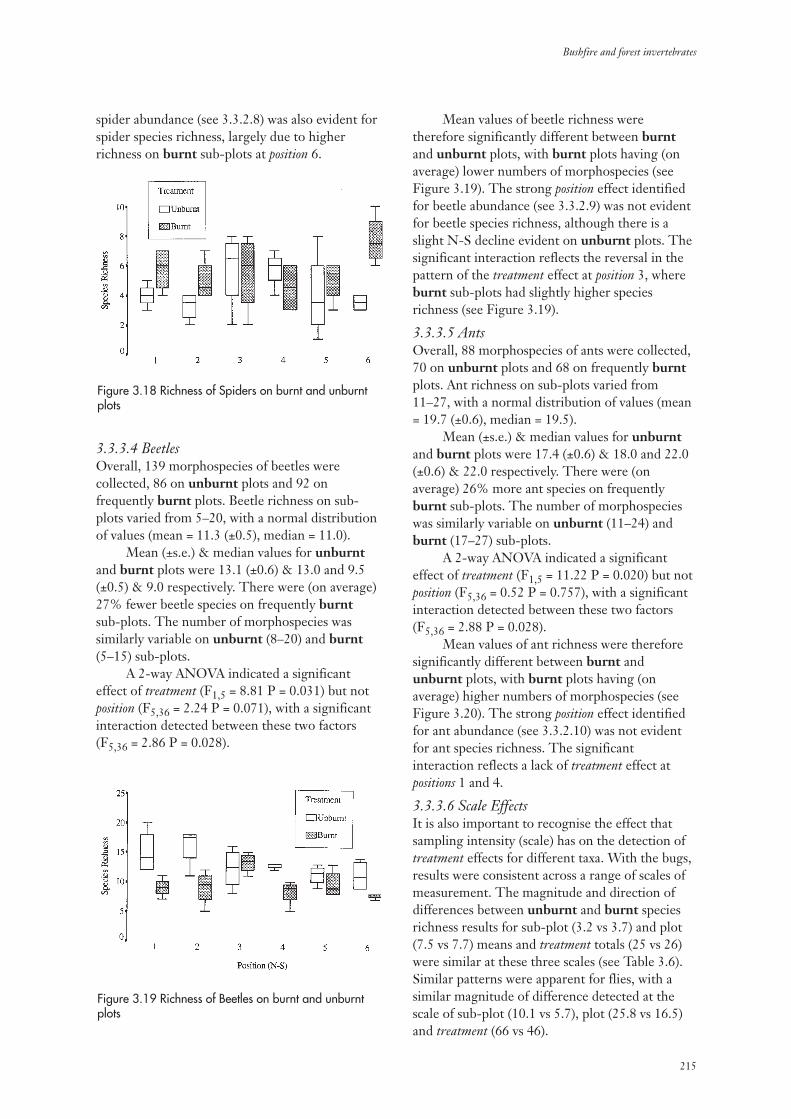
215
Bushfire and forest invertebrates
spider abundance (see 3.3.2.8) was also evident forspider species richness, largely due to higherrichness on burnt sub-plots at position 6.
3.3.3.4 BeetlesOverall, 139 morphospecies of beetles werecollected, 86 on unburnt plots and 92 onfrequently burnt plots. Beetle richness on sub-plots varied from 5–20, with a normal distributionof values (mean = 11.3 (±0.5), median = 11.0).
Mean (±s.e.) & median values for unburntand burnt plots were 13.1 (±0.6) & 13.0 and 9.5(±0.5) & 9.0 respectively. There were (on average)27% fewer beetle species on frequently burntsub-plots. The number of morphospecies wassimilarly variable on unburnt (8–20) and burnt(5–15) sub-plots.
A 2-way ANOVA indicated a significanteffect of treatment (F1,5 = 8.81 P = 0.031) but notposition (F5,36 = 2.24 P = 0.071), with a significantinteraction detected between these two factors(F5,36 = 2.86 P = 0.028).
Mean values of beetle richness weretherefore significantly different between burntand unburnt plots, with burnt plots having (onaverage) lower numbers of morphospecies (seeFigure 3.19). The strong position effect identifiedfor beetle abundance (see 3.3.2.9) was not evidentfor beetle species richness, although there is aslight N-S decline evident on unburnt plots. Thesignificant interaction reflects the reversal in thepattern of the treatment effect at position 3, whereburnt sub-plots had slightly higher speciesrichness (see Figure 3.19).
3.3.3.5 AntsOverall, 88 morphospecies of ants were collected, 70 on unburnt plots and 68 on frequently burntplots. Ant richness on sub-plots varied from11–27, with a normal distribution of values (mean= 19.7 (±0.6), median = 19.5).
Mean (±s.e.) & median values for unburntand burnt plots were 17.4 (±0.6) & 18.0 and 22.0(±0.6) & 22.0 respectively. There were (onaverage) 26% more ant species on frequentlyburnt sub-plots. The number of morphospecieswas similarly variable on unburnt (11–24) andburnt (17–27) sub-plots.
A 2-way ANOVA indicated a significanteffect of treatment (F1,5 = 11.22 P = 0.020) but notposition (F5,36 = 0.52 P = 0.757), with a significantinteraction detected between these two factors(F5,36 = 2.88 P = 0.028).
Mean values of ant richness were thereforesignificantly different between burnt andunburnt plots, with burnt plots having (onaverage) higher numbers of morphospecies (seeFigure 3.20). The strong position effect identifiedfor ant abundance (see 3.3.2.10) was not evidentfor ant species richness. The significantinteraction reflects a lack of treatment effect atpositions 1 and 4.
3.3.3.6 Scale EffectsIt is also important to recognise the effect thatsampling intensity (scale) has on the detection oftreatment effects for different taxa. With the bugs,results were consistent across a range of scales ofmeasurement. The magnitude and direction ofdifferences between unburnt and burnt speciesrichness results for sub-plot (3.2 vs 3.7) and plot(7.5 vs 7.7) means and treatment totals (25 vs 26)were similar at these three scales (see Table 3.6).Similar patterns were apparent for flies, with asimilar magnitude of difference detected at thescale of sub-plot (10.1 vs 5.7), plot (25.8 vs 16.5)and treatment (66 vs 46).
Figure 3.18 Richness of Spiders on burnt and unburntplots
Figure 3.19 Richness of Beetles on burnt and unburntplots
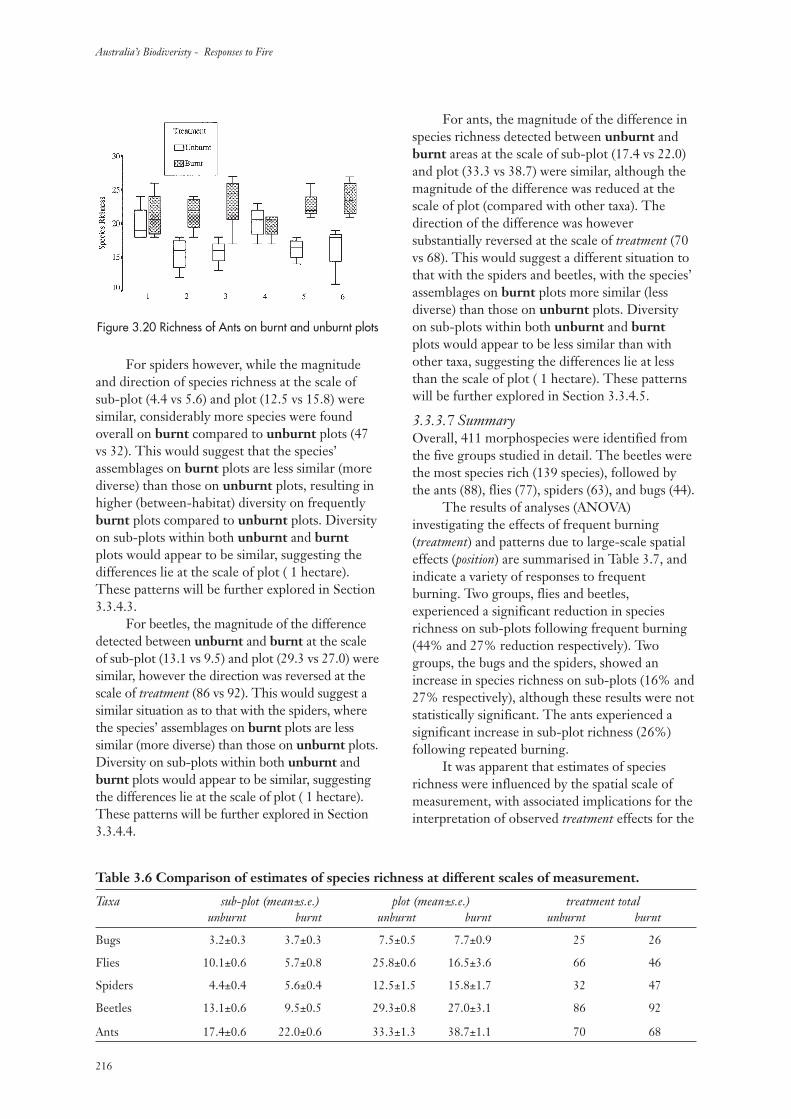
For ants, the magnitude of the difference inspecies richness detected between unburnt andburnt areas at the scale of sub-plot (17.4 vs 22.0)and plot (33.3 vs 38.7) were similar, although themagnitude of the difference was reduced at thescale of plot (compared with other taxa). Thedirection of the difference was howeversubstantially reversed at the scale of treatment (70vs 68). This would suggest a different situation tothat with the spiders and beetles, with the species’assemblages on burnt plots more similar (lessdiverse) than those on unburnt plots. Diversityon sub-plots within both unburnt and burntplots would appear to be less similar than withother taxa, suggesting the differences lie at lessthan the scale of plot ( 1 hectare). These patternswill be further explored in Section 3.3.4.5.
3.3.3.7 SummaryOverall, 411 morphospecies were identified fromthe five groups studied in detail. The beetles werethe most species rich (139 species), followed bythe ants (88), flies (77), spiders (63), and bugs (44).
The results of analyses (ANOVA)investigating the effects of frequent burning(treatment) and patterns due to large-scale spatialeffects (position) are summarised in Table 3.7, andindicate a variety of responses to frequentburning. Two groups, flies and beetles,experienced a significant reduction in speciesrichness on sub-plots following frequent burning(44% and 27% reduction respectively). Twogroups, the bugs and the spiders, showed anincrease in species richness on sub-plots (16% and27% respectively), although these results were notstatistically significant. The ants experienced asignificant increase in sub-plot richness (26%)following repeated burning.
It was apparent that estimates of speciesrichness were influenced by the spatial scale ofmeasurement, with associated implications for theinterpretation of observed treatment effects for the
216
Australia’s Biodiveristy - Responses to Fire
For spiders however, while the magnitudeand direction of species richness at the scale ofsub-plot (4.4 vs 5.6) and plot (12.5 vs 15.8) weresimilar, considerably more species were foundoverall on burnt compared to unburnt plots (47vs 32). This would suggest that the species’assemblages on burnt plots are less similar (morediverse) than those on unburnt plots, resulting inhigher (between-habitat) diversity on frequentlyburnt plots compared to unburnt plots. Diversityon sub-plots within both unburnt and burntplots would appear to be similar, suggesting thedifferences lie at the scale of plot ( 1 hectare).These patterns will be further explored in Section3.3.4.3.
For beetles, the magnitude of the differencedetected between unburnt and burnt at the scaleof sub-plot (13.1 vs 9.5) and plot (29.3 vs 27.0) weresimilar, however the direction was reversed at thescale of treatment (86 vs 92). This would suggest asimilar situation as to that with the spiders, wherethe species’ assemblages on burnt plots are lesssimilar (more diverse) than those on unburnt plots.Diversity on sub-plots within both unburnt andburnt plots would appear to be similar, suggestingthe differences lie at the scale of plot ( 1 hectare).These patterns will be further explored in Section3.3.4.4.
Figure 3.20 Richness of Ants on burnt and unburnt plots
Table 3.6 Comparison of estimates of species richness at different scales of measurement.
Taxa sub-plot (mean±s.e.) plot (mean±s.e.) treatment totalunburnt burnt unburnt burnt unburnt burnt
Bugs 3.2±0.3 3.7±0.3 7.5±0.5 7.7±0.9 25 26
Flies 10.1±0.6 5.7±0.8 25.8±0.6 16.5±3.6 66 46
Spiders 4.4±0.4 5.6±0.4 12.5±1.5 15.8±1.7 32 47
Beetles 13.1±0.6 9.5±0.5 29.3±0.8 27.0±3.1 86 92
Ants 17.4±0.6 22.0±0.6 33.3±1.3 38.7±1.1 70 68

217
Bushfire and forest invertebrates
different taxa. For bugs and flies results wereconsistent across a range of scales ofmeasurement, with the magnitude and directionof differences between unburnt and burnt resultsfor sub-plot, plot and treatment similar.
For spiders, while the magnitude anddirection of species richness at the scale of sub-plot and plot were similar, considerably morespecies were found overall on burnt compared tounburnt plots. This suggested that species’assemblages on burnt plots were more diversethan those on unburnt plots, resulting in higher(between-habitat) diversity. Diversity on sub-plotswithin both unburnt and burnt plots wouldappear to be similar, suggesting the differences lieat the scale of plot ( 1 hectare).
For beetles, the magnitude of the differencedetected between unburnt and burnt at the scaleof sub-plot and plot were similar, however thedirection was reversed at the scale of treatment.This would suggest a similar situation as to thatwith the spiders, where the species’ assemblageson burnt plots are more diverse than those onunburnt plots. Diversity on sub-plots within bothunburnt and burnt plots would appear to besimilar, suggesting the differences lie at the scaleof plot (1 hectare).
For ants, the magnitude of the difference inspecies richness detected between unburnt andburnt areas at the scale of sub-plot and plot weresimilar, although the magnitude of the differencewas reduced at the scale of plot (compared withother taxa). The direction of the difference washowever substantially reversed at the scale oftreatment, suggesting a different situation to thatwith the spiders and beetles, with the species’assemblages on burnt plots less diverse. Diversityon sub-plots within both unburnt and burnt plotswould appear to be less similar than with othertaxa, suggesting the differences lie at less than thescale of plot ( 1 hectare).
These spatial patterns in estimates of speciesrichness are a consequence of the composition ofinvertebrate assemblages (communities) at thevarying scales of investigation. The nature ofthese patterns, and their interaction withenvironmental variables, will be further exploredin Section 3.3.4.
3.3.4 Community Composition
3.3.4.1 HemipteraThe 44 Hemipteran morphospecies wererepresentative of 14 families, the infra-orderDipsocoromorpha (not readily discernible tofamily) and an unidentifiable Homopteran (seeTable 3.8). The most diverse groups overall werethe family Cicadellidae and the infra-orderDipsocoromorpha, containing 27% and 25% ofoverall Hemipteran morphospecies respectively.
All three sub-orders were represented: theSternorrhyncha (5 morphospecies), theAuchenorrhyncha (13 morphospecies) and theHeteroptera (26 morphospecies). TheSternorrhyncha are mainly sedentary, often livingunder waxy secretions or inside galls induced in thehost plants. Four morphospecies from this sub-order were found on unburnt plots and three onburnt plots (see Table 3.8). The Auchenorrhyncha(leaf- and plant-hoppers) are all plant-sap feeders asadults and generally spend the bulk of their time onplant foliage. Seven morphospecies from this sub-order were found on unburnt plots and nine onburnt plots. Most of the “true bugs”, theHeteroptera, are plant-sap feeders, although somegroups, such as the Reduviidae, are predatory.Thirteen morphospecies from this sub-order werefound on unburnt plots and fourteen on burntplots. Overall, a similar number of morphospecieswere found on unburnt and burnt plots (24 and 26respectively) with similar proportions from thethree sub-orders.
Although the two treatments are similarlydiverse, both at the sub-ordinal and
Table 3.7 Changes in mean species richness following frequent burning for selected terrestrialinvertebrate taxa.
Taxa Common name Change with Statistically Large-scalefrequent burning significant ? (P<0.05) spatial patterns ?
Hemiptera bugs ⇑ 16% no noDiptera flies ⇓ 44% yes yesAraneae spiders ⇑ 27% no no*Coleoptera beetles ⇓ 27% yes no*Formicidae ants ⇑ 26% yes no*
* indicates a statistically significant interaction between treatment and position effects.

218
Australia’s Biodiveristy - Responses to Fire
morphospecies level (see Section 3.3.3.1), only 7morphospecies (16%) were common to bothtreatments (Group A - Table 3.9). When themorphospecies are arranged to reflect theirdistribution across plots and treatments, it is clearthat there is a group of species (18) found only onunburnt sub-plots (41% - Group B) and adifferenct group of species (19) found only onburnt sub-plots (43% - Group C). Frequentburning would therefore appear to have resultedin the loss of up to 18 species of Hemipteran,however the changed environment supports upto19 new species not found in unburnt areas.
Morphospecies found on both treatmentsare representative of the families Cicadellidae(3 species), Coccidae (2 species), Fulgoroidea(1 species) and the infra-order Dipsocoromorpha(1 species). Morphospecies found only onunburnt sub-plots are representative of thefamilies Cicadellidae (4 species), Coccidae(1 species), Nabidae (1 species), Enicocephalidae(1 species), Lygaeidae (1 species), Pentatomidae(1 species), Tingidae (1 species), the infra-orderDipsocoromorpha (7 species) and an unnamedHomopteran species. Morphospecies found onlyon burnt sub-plots are representative of thefamilies Cicadellidae (5 species), Coccidae(1 species), Miridae (1 species), Reduviidae(5 species), Lygaeidae (1 species), Plataspidae(1 species), Thaumastocoridae (1 species),Anthocoridae (1 species) and the infra-orderDipsocoromorpha (3 species).
As many of these families are representedonly by single individuals or found only on one
sub-plot of a treatment, it is difficult to identifyclear patterns from these tabulated data (at thespecies level). At this stage it is apparent howeverthat unburnt plots have more species from theinfra-order Dipsocoromorpha (7 vs 3), whileburnt plots have greater numbers of species fromthe family Reduviidae (5 vs 0).
Although this table of relative abundanceenables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed in the formof a “bi-plot” derived from the CCA ordination(see 2.5.3.3). This graphical display (Figure 3.21)shows the configuration of the environmentalvariables, the scatter of sub-plots, and therelationship between the two, giving an overviewof how community composition varies with theenvironment (Ter Braak 1986).
A number of features are apparent from this bi-plot. Firstly, the minimal overlap of unburnt andburnt sub-plots in ordination space reflects thelargely dissimilar species assemblages of these twotreatments. Secondly, the tighter clustering of burntsub-plots indicates a lower within-treatmentdiversity compared to unburnt sub-plots (ie. alower β-diversity). Richness values on burnt sub-plots were on average 16% higher than on unburntsub-plots (see Section 3.3.3.1), however the highsimilarity of sub-plots for this treatment mean thatthe overall richness of unburnt and burnt plots wassimilar (25 vs 26 species). Unburnt sub-plots havelower richness (α-diversity) but are less similar,resulting in higher “turnover” between sub-plots(higher β-diversity).
Table 3.8 Breakdown of Hemipteran morphospecies by sub-order and family.
# MorphospeciesSub-order Family Common name unburnt burnt total
Sternorrhyncha Coccidae scale insects 3 3 4Homoptera* - 1 0 1
Auchenorrhyncha Cicadellidae leaf-hoppers 6 8 12Fulgoroidea plant-hoppers 1 1 1
Heteroptera Dipsocoromorpha* - 8 4 11Enicocephalidae - 1 0 1Nabidae - 1 0 1Tingidae lace bugs 1 0 1Pentatomidae shield bugs 1 0 1Lygaeidae seed bugs 1 1 2Anthocoridae flower bugs 0 1 1Miridae - 0 1 1Thaumastocoridae - 0 1 1Plataspidae - 0 1 1Reduviidae assassin bugs 0 5 5
Totals 24 26 44
* Not readily discernible to Family level
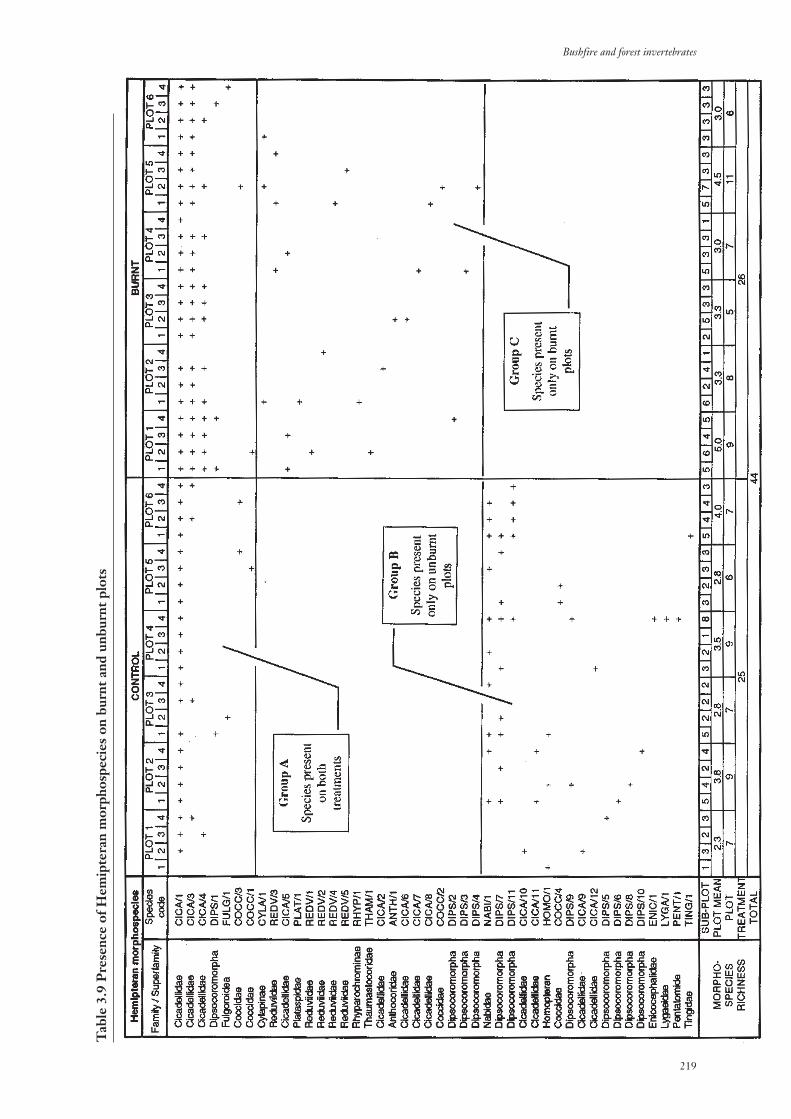
219
Bushfire and forest invertebrates
Tab
le 3
.9 P
rese
nce
of H
emip
tera
n m
orph
ospe
cies
on
burn
t an
d un
burn
t pl
ots

Brachycera (59 morphospecies). Nematocera adultsare generally slender with long legs, and haveaquatic larvae (eg. mosquitoes, midges & sandflies)or are gall makers. Twenty-five morphospecies fromthis sub-order were found on unburnt plots andeighteen on burnt plots (see Table 3.10).Brachycera adults are heavier set with relativelyshort legs (eg. House flies and March flies) and havemainly terrestrial larvae (often found in damp soiland rotting vegetation). Forty-one morphospeciesfrom this sub-order (Brachycera) were found onunburnt plots and twenty-eight on burnt plots.Overall, 30% less morphospecies were found onburnt plots compared with unburnt plots (46 and66 respectively) with similar trends for both sub-orders.
The unburnt treatment was considerablymore diverse than the burnt treatment at themorphospecies level (see Section 3.3.3.2) however 35morphospecies (45%) were common to bothtreatments (Group A - Table 3.11). When themorphospecies are arranged to reflect theirdistribution across plots and treatments, it is clearthat there is a group of species (31) found only onunburnt sub-plots (40% - Group B) and adifferent group of species (11) found only onburnt sub-plots (14% - Group C). Frequentburning would therefore appear to have resultedin the loss of up to 31 species of Diptera, howeverthe changed environment supports up to 11 newspecies not found in unburnt areas.
Morphospecies found on both treatments arerepresentative of the families Phoridae (12 species),Cecidomyiidae and Sciaridae (5 species each),Chloropidae (3 species), Tachydromiinae,Ceratopogonidae, Chironomidae, Muscidae(2 species each), Dolichopodidae and Scatopsidae(1 species each). Morphospecies found only onunburnt sub-plots are representative of the familiesTachydromiinae and Phoridae (4 species each),Ceratopogonidae and Sciaridae (3 species each),Chloropidae, Drosophilidae, Muscidae,Sphaeroceridae and Tachinidae (2 species each),Syrphidaea, Therevidae, Micropezidae, Tipulidae,Cecodomyiidae, Mycetophilidae and Chironomidae(1 species each). Morphospecies found only onburnt sub-plots are representative of the familiesCeratopogonidae (2 species), Tachinidae,Piophilidae, Calliphoridae, Cecidomyiidae,Chloropidae, Muscidae, Tachydromiinae andSphaeroceridae (1 species each).
As many of these families are representedonly by single individuals or found only on one
220
Australia’s Biodiveristy - Responses to Fire
The third feature concerns the contributionof environmental variables to the differences inspecies composition for the two treatments. In thebi-plot (Figure 3.21) the length of the arrowsignifies the relative contribution of that variableto species composition, and the direction signifiesits contribution to the differences betweentreatments. Unburnt sub-plots are characterisedby high levels of litter and high cover of tall andvery tall shrubs. Due to the correlation betweenvariables (see Section 3.2) these plots are alsocharacterised by high top-soil moisture levels andlow and spatially variable amounts of insolation atground level. These environmental variables makethe greatest contribution to the differences inspecies composition between unburnt and burntplots. Burnt sub-plots are characterised by highlevels of insolation at ground level, and to a lesserextent, steeper slopes, greater top-soil hardnessand greater cover of the herb & shrub componentof the understorey vegetation. Otherenvironmental variables make only a minorcontribution to the observed differences inHemipteran species composition between burntand unburnt treatments.
3.3.4.2 DipteraThe 77 Dipteran (fly) morphospecies wererepresentative of 2 sub-orders and 20 families (seeTable 3.10). The most diverse groups overall werethe families Phoridae and Sciaridae, containing22% and 10% of morphospecies respectively. Thefamilies Cecodomyiidae, Ceratopogonidae andEmpididae each contained 9% of overallmorphospecies.
Both Australian sub-orders were represented:the Nematocera (28 morphospecies) and the
Figure 3.21 Bi-plot from CCA ordination of Hemipteranpresence/absence data. (Ellipses represent 1 standarddeviation unit around treatment centroids)

221
Bushfire and forest invertebrates
Table 3.11 Presence of Dipteran morphospecies on burnt and unburnt plots.

222
Australia’s Biodiveristy - Responses to Fire
sub-plot of a treatment, it is difficult to identifyclear patterns from these tabulated data (at thespecies level). At this stage it is apparent howeverthat unburnt plots have more species from thefamilies Sciaridae (8 vs 5), Phoridae (16 vs 13) andEmpididae (6 vs 3).
Although this table of relative abundanceenables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed in the formof a “bi-plot” derived from the CCA ordination(see 2.5.3.3). This graphical display (Figure 3.22)shows the configuration of the environmentalvariables, the scatter of sub-plots, and therelationship between the two, giving an overviewof how community composition varies with theenvironment (Ter Braak 1986).
A number of features are apparent from thisbi-plot. Firstly, the substantial overlap of unburntand burnt sub-plots in ordination space reflectsthe relatively large number of morphospecies (35)shared by the two treatments. Secondly, the looseclustering of both unburnt and burnt sub-plotsindicates similar within-treatment diversity. Burntsub-plots are slightly more diverse (spread-out)indicating a higher turnover (β-diversity) for thistreatment. Richness values on burnt sub-plots
were on average 44% lower than on unburnt sub-plots (see Section 3.3.3.2), however the slightlyhigher similarity of unburnt sub-plots means that,overall, burnt plots had 30% less species thanunburnt plots (46 vs 66). Unburnt sub-plots havehigher richness (α-diversity) and are more similar,resulting in lower “turnover” between sub-plots(lower β-diversity).
The third feature concerns the contributionof environmental variables to the differences in
Figure 3.22 Bi-plot from CCA ordination of Dipteranpresence/absence data. (Ellipses represent 1 standarddeviation unit around treatment centroids)
Table 3.10 Breakdown of Dipteran morphospecies by sub-order and family.
# MorphospeciesSub-order Family Common name unburnt burnt total
Nematocera Sciaridae - 8 5 8Cecidomyiidae gall midges 6 6 7Ceratopogonidae sand flies 5 4 7Chironomidae midges 3 2 3Scatopsidae - 1 1 1Tipulidae crane flies 1 0 1Mycetophilidae fungus gnats 1 0 1
Brachycera Phoridae - 16 13 17Empididae* - 6 3 7Chloropidae - 5 4 6Muscidae bush flies 4 3 5Sphaeroceridae - 2 1 3Tachinidae - 2 1 3Dolichopodidae - 1 1 1Drosophilidae vinegar flies 2 0 2Therevidae - 1 0 1Syrphidae hover flies 1 0 1Micropezidae stilt-legged flies 1 0 1Piophilidae - 0 1 1Calliphoridae blowflies 0 1 1
Totals 66 46 77
* all morphospecies from sub-family Tachydromiinae

223
Bushfire and forest invertebrates
species composition for the two treatments. In thebi-plot (Figure 3.22) the length of the arrowsignifies the relative contribution of that variableto species composition, and the direction signifiesits contribution to the differences betweentreatments. Unburnt sub-plots are characterisedby high levels of litter and high cover of tall andvery tall shrubs. Due to the correlation betweenvariables (see Section 3.2) these plots are alsocharacterised by high top-soil moisture levels andlow and spatially variable amounts of insolation atground level. Burnt sub-plots are characterised byhigh levels of insolation at ground level, and to alesser extent, steeper slopes and greater exposure(more north-westerly aspects). The amount oflitter and the level of insolation at ground levelmake the greatest contribution to the differencesin species composition between unburnt andburnt plots. Other environmental variables makeonly a minor contribution to the observeddifferences in Dipteran species compositionbetween burnt and unburnt treatments.
3.3.4.3 SpidersThe 63 spider morphospecies were representativeof 21 families, with the most diverse groupsoverall being the families Zodariidae,
Gnaphosidae and Corinnidae, containing 14, 13and 13% of overall morphospecies respectively(see Table 3.12). Most (62%) of the families wererepresented by only 1 or 2 species.
Overall, burnt plots had a greater number ofmorphospecies than unburnt plots (48 and 31respectively), with 16 morphospecies (25%)common to both treatments (Group A - Table3.13). When the morphospecies are arranged toreflect their distribution across plots andtreatments, it is clear that there is a group ofspecies (15) found only on unburnt sub-plots(24% - Group B) and a different group of species(32) found only on burnt sub-plots (51% - GroupC). Frequent burning would therefore appear tohave resulted in the loss of up to 15 species ofspider, however the changed environment supportsup to 32 new species not found in unburnt areas.
Morphospecies found on both treatmentswere representative of the families Corinnidae(3 species), Textricellidae and Zodariidae (2 specieseach), Hahniidae, Linyphiidae, Gnaphosidae,Ctenidae, Micropholcommatidae, Oonopidae,Thomisidae, Toxopidae and Theridiidae (1 specieseach). Morphospecies found only on unburnt sub-plots are representative of the families Theridiidae,
Table 3.12 Breakdown of spider morphospecies by family.
# MorphospeciesFamily Common name Ecological Information unburnt burnt total
Zodariidae - terrestrial; under stones, logs & litter 2 9 9Gnaphosidae - terrestrial; under stones, logs & litter 3 6 8Corinnidae - generalised hunters & ant specialists 4 7 8Salticidae jumping spiders terrestrial hunters on foliage, trees & logs. 3 3 6Linyphiidae tent spiders build webs in foliage & near ground 2 4 5Theridiidae - web builders; near ground level 3 2 4Textricellidae - moist habitats; litter dwellers 2 3 3Clubionidae ant-mimicking spiders nocturnal hunters; live in rolled-up
leaves & in litter 1 1 2Tekellidae - moist habitats; litter dwellers 1 1 2Oonopidae - cryptic; occur under stones, logs & litter. 2 1 2Ctenidae - terrestrial; vagrant hunters on ground & in litter 1 1 1Hahniidae - build sheet webs in litter & foliage 1 1 1Micropholcommatidae - moist habitats; litter dwellers 1 1 1Thomisidae crab/flower spiders occur on foliage & bark 1 1 1Toxopidae - moist habitats 1 1 1Lycosidae wolf spiders terrestrial; ground hunters 0 3 3Prododomidae - dry habitats 0 1 1Heteropodidae - bark, foliage & litter dwellers 0 1 1Dictynoidea - sheet web builders on ground, litter & bark 0 1 1Malkaridae - moist habitats; litter dwellers 2 0 2Stiphidiidae - sheet web builders 1 0 1
TOTAL 31 48 63

224
Australia’s Biodiveristy - Responses to FireT
able
3.1
3 P
rese
nce
of S
pide
r m
orph
ospe
cies
on
burn
t an
d un
burn
t pl
ots.

225
Bushfire and forest invertebrates
Salticidae, Gnaphosidae and Malkaridae (2 specieseach), Oonopidae, Tekellidae, Stiphidiidae,Linyphiidae, Clubionidae and Corinnidae(1 species each). Morphospecies found only onburnt sub-plots are representative of the familiesZodariidae (7 species), Gnaphosidae (5 species),Corinnidae (4 species), Salticidae and Lycosidae(3 species each), Linyphiidae (2 species) andTekellidae, Textricellidae, Theridiidae,Clubionidae, Dictynoidea, Heteropodidae andPrododomidae (1 species each).
As many of these families are representedonly by single individuals or found only on onesub-plot of a treatment, it is difficult to identifyclear patterns from these tabulated data (at thespecies level). At this stage it is apparent howeverthat unburnt plots have more species from thefamily Malkaridae (2 vs 0), while burnt plots havegreater numbers of species from the familiesZodariidae (9 vs 2), Gnaphosidae (6 vs 3),Corinnidae (7 vs 4), Linyphiidae (4 vs 2) andLycosidae (3 vs 0).
Although this table of relative abundanceenables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed in the formof a “bi-plot” derived from the CCA ordination(see 2.5.3.3). This graphical display (Figure 3.23)shows the configuration of the environmentalvariables, the scatter of sub-plots, and therelationship between the two, giving an overviewof how community composition varies with theenvironment (Ter Braak 1986).
A number of features are apparent from thisbi-plot. Firstly, the minimal overlap of unburnt andburnt sub-plots in ordination space reflects thelargely dissimilar species assemblages of these twotreatments. Secondly, the tighter clustering ofunburnt sub-plots indicates a lower within-treatment diversity compared to burnt sub-plots.Burnt sub-plots are slightly more diverse (spread-out) indicating a higher turnover (β-diversity) forthis treatment. Richness values on burnt sub-plotswere on average were 27% higher than on unburntsub-plots (see Section 3.3.3.3), however the greaterdissimilarity of burnt sub-plots means that, overall,burnt plots had 55% more species than unburntplots. Burnt sub-plots have higher richness (α-diversity) and are less similar, resulting in higher“turnover” between sub-plots (higher β-diversity).
The third feature concerns the contributionof environmental variables to the differences inspecies composition for the two treatments. In thebi-plot (Figure 3.23) the length of the arrowsignifies the relative contribution of that variable tospecies composition, and the direction signifies itscontribution to the differences between treatments.Unburnt sub-plots are characterised by high levelsof litter and high cover of tall and very tall shrubs.Due to the correlation between variables (seeSection 3.2) these plots are also characterised byhigh top-soil moisture levels and low and spatiallyvariable amounts of insolation at ground level.Burnt sub-plots are characterised by high levels ofinsolation at ground level, and to a lesser extent,steeper slopes and greater cover of the herb &shrub component of the understorey vegetation.The amounts of insolation and litter make thegreatest contribution to the differences in speciescomposition between unburnt and burnt plots.Other environmental variables make only a minorcontribution to the observed differences in spiderspecies composition.
3.3.4.4 BeetlesThe 139 beetle morphospecies wererepresentative of nine super-families and 25families (see Table 3.14). The most diverse groupswere the families Staphylinidae andCurculionidae, containing 22% and 17% ofoverall morphospecies respectively.
Nine super-families were represented: theStaphylinoidea (56 morphospecies), theCurculionoidea (23 morphospecies), Caraboidea,Scarabaeoidea, Cucujoidea (12 morphospecieseach), Chrysomeloidea (10 morphospecies),Tenebrionoidea (9 morphospecies), Bostrichoidea
Figure 3.23 Bi-plot from CCA ordination of spiderpresence/absence data. (Ellipses represent 1 standarddeviation unit around treatment centroids)

on unburnt plots and nine on burnt plots. ManyTenebrionoidea are scavengers inhabiting the litterlayer. Seven morphospecies were found onunburnt plots and two on burnt plots. TheBostrichoidea were represented by one family, theAnobiidae, which feed on a variety of plant &animal products. Three morphospecies from thisfamily were each found, two on unburnt and twoon burnt plots. The Elateroidea were representedby a single family, the Elateridae (Click Beetles),which are mainly predatory. Two morphospeciesfrom this family were found, one on unburnt andone on burnt plots (see Table 3.14).
Overall, slightly more morphospecies werefound on burnt plots compared with unburntplots (92 and 86 respectively), although on average,burnt sub-plots had 27% fewer species (see Section3.3.3.4). Thirty-nine morphospecies (28%) werecommon to both treatments (Group A - Table3.15). When the morphospecies are arranged toreflect their distribution across plots andtreatments, it is clear that there is a group of species(47) found only on unburnt sub-plots (34% -Group B) and a different group of species (53)
226
Australia’s Biodiveristy - Responses to Fire
(3 morphospecies), and the Elateroidea(2 morphospecies).
Most Staphylinoidea are general predators,with some feeding on decomposing fruits. Thirty-eight morphospecies from this super-family werefound on unburnt plots and thirty-six on burntplots (see Table 3.14). The Curculionoidea wererepresented by one family, Curculionidae (theWeevils), with nine morphospecies found onunburnt plots and eighteen on burnt plots. TheCaraboidea were represented by one family,Caraboidea (the Ground Beetles), which are mainlypredatory on plant-inhabiting insects. Elevenmorphospecies from this family were found onunburnt plots and eight on burnt plots. TheScarabaeoidea commonly feed on eucalypt leaves asadults. Eight morphospecies from this super-familywere found on unburnt plots and nine on burntplots. Many Cucujoidea are known to feed on fungigrowing on leaf surfaces. Six morphospecies fromthis super-family were found on unburnt plots andseven on burnt plots. The Chrysomeloidea feed onleaves and other vegetative parts of plants, both aslarvae and adults. Four morphospecies were found
Table 3.14 Breakdown of Beetle morphospecies by super-family and family.
# MorphospeciesSuper-family Family Common name unburnt burnt total
Staphylinoidea Staphylinidae Rove beetles 21 20 31Pselaphidae - 6 7 11Scydmaenidae - 5 7 8Ptiliidae - 3 1 3Leiodidae - 3 1 3
Scarabaeoidea Scarabaeidae - 6 7 9Trogidae - 1 0 1Hybosoridae - 1 1 1Passalidae - 0 1 1
Elateroidea Elateridae Click Beetles 1 1 2Cucujoidea Nitidulidae - 2 1 3
Corylophidae - 1 2 3Endomychidae - 1 1 2Lathridiidae - 2 1 2Silvanidae - 0 1 1Phalacridae - 0 1 1
Chrysomeloidea Chrysomelidae Leaf Beetles 4 8 9Cerambycidae Longicorn Beetles 0 1 1
Curculionoidea Curculionidae Weevils 9 18 23Caraboidea Carabidae Ground Beetles 11 8 12Bostrichoidea Anobiidae - 2 2 3Tenebrionoidea Tenebrionidae Darkling Beetles 3 2 5
Aderidae - 2 0 2Oedemeridae - 1 0 1Anthicidae - 1 0 1
Totals 86 92 139

227
Bushfire and forest invertebrates
found only on burnt sub-plots (38% - Group C).Frequent burning would therefore appear to haveresulted in the loss of up to 47 species of Coleoptera,however the changed environment supports up to 53new species not found in unburnt areas.
Morphospecies found on both treatments arerepresentative of the families Staphylinidae(10 species), Carabidae (7 species), Scarabaeidae,Scydmaenidae, Curculionidae (4 species each),Chrysomelidae (3 species), Pselaphidae (2 species),Ptiliidae, Hybosoridae, Lathridiidae, Anobiidae andLeiodidae (1 species each). Morphospecies foundonly on unburnt sub-plots are representative of thefamilies Staphylinidae (11 species), Curculionidae(5 species), Carabidae, Pselaphidae (4 species each),Tenebrionidae (3 species), Aderidae, Scarabaeidae,Ptiliidae and Nitidulidae (2 species each),Scydmaenidae, Chrysomelidae, Lathridiidae,Anobiidae, Leiodidae, Trogidae, Elateridae,Corylophidae, Endomychidae, Oedemeridae andAnthicidae (1 species each). Morphospecies foundonly on burnt sub-plots are representative of thefamilies Curculionidae (14 species), Staphylinidae(10 species) Chrysomelidae, Pselaphidae (5 specieseach), Scarabaeidae, Scydmaenidae (3 species each),Corylophidae, Tenebrionidae, (2 species each),Passalidae, Elateridae, Nitidulidae, Endomychidae,Silvanidae, Phalacridae, Cerambycidae, Carabidaeand Anobiidae (1 species each).
As many of these families are representedonly by single individuals or found only on onesub-plot of a treatment, it is difficult to identifyclear patterns from these tabulated data (at thespecies level). At this stage it is apparent howeverthat unburnt plots have more species from thefamily Carabidae (11 vs 8), while burnt plots havegreater numbers of species from the familiesCurculionidae (18 vs 9) and Chrysomelidae (8 vs 4).
Although this table of relative abundanceenables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed in the formof a “bi-plot” derived from the CCA ordination(see 2.5.3.3). This graphical display (Figure 3.24)shows the configuration of the environmentalvariables, the scatter of sub-plots, and therelationship between the two, giving an overviewof how community composition varies with theenvironment (Ter Braak 1986).
A number of features are apparent from thisbi-plot. Firstly, the small overlap of unburnt andburnt sub-plots in ordination space reflects thelargely dissimilar species assemblages of these twotreatments. Secondly, the tighter clustering of burnt
sub-plots suggests a lower within-treatmentdiversity compared to unburnt sub-plots (ie. alower β-diversity). Richness values on burnt sub-plots were, on average, 27% lower than on unburntsub-plots (see Section 3.3.3.4), with the apparentlyhigh similarity of sub-plots for this treatmentmeaning that the overall richness of burnt plotsshould be much lower than for unburnt plots. Infact, overall, burnt plots had slightly more speciesthan unburnt plots (92 vs 86). This clustering istherefore an artefact of the two-dimensionalrepresentation of the data, with an inspection ofTable 3.15 revealing a greater number of speciesunique to burnt plots (53 vs 47). While unburntsub-plots have higher richness (α-diversity), burntplots are less similar, resulting in higher “turnover”between sub-plots (higher β-diversity).
The third feature concerns the contributionof environmental variables to the differences inspecies composition for the two treatments. In thebi-plot (Figure 3.24) the length of the arrowsignifies the relative contribution of that variable tospecies composition, and the direction signifies itscontribution to the differences between treatments.Unburnt sub-plots are characterised by high levelsof litter and high cover of tall and very tall shrubs.Due to the correlation between variables (seeSection 3.2) these plots are also characterised byhigh top-soil moisture levels and low and spatiallyvariable amounts of insolation at ground level.Burnt sub-plots are characterised by high levels ofinsolation at ground level, and to a lesser extent,steeper slopes. These environmental variables makethe greatest contribution to the differences inspecies composition between unburnt and burntplots, with other environmental variables makeonly a minor contribution to the observeddifferences in beetle species composition.
Figure 3.24 Bi-plot from CCA ordination of Beetlepresence/absence data. (Ellipses represent 1 standarddeviation unit around treatment centroids)

228
Australia’s Biodiveristy - Responses to Fire
Tab
le 3
.15.
Pre
senc
e of
Bee
tle
mor
phos
peci
es o
n bu
rnt
and
unbu
rnt
plot
s.
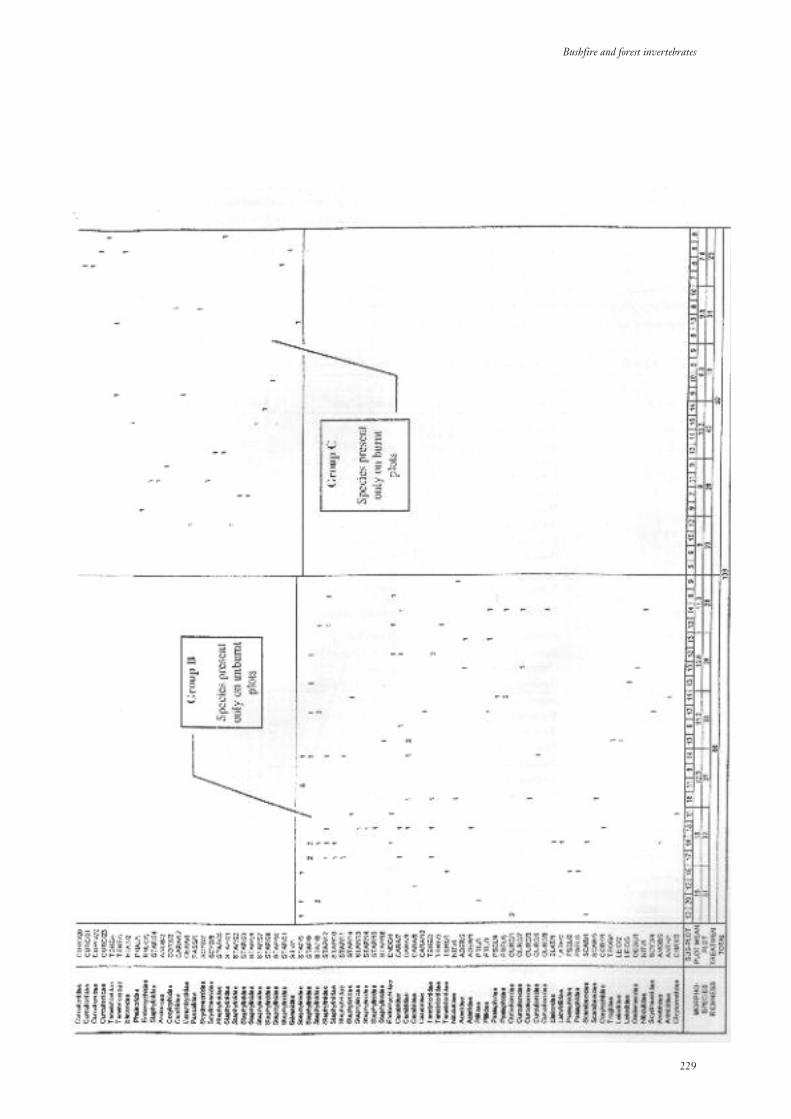
229
Bushfire and forest invertebrates
to be scanned
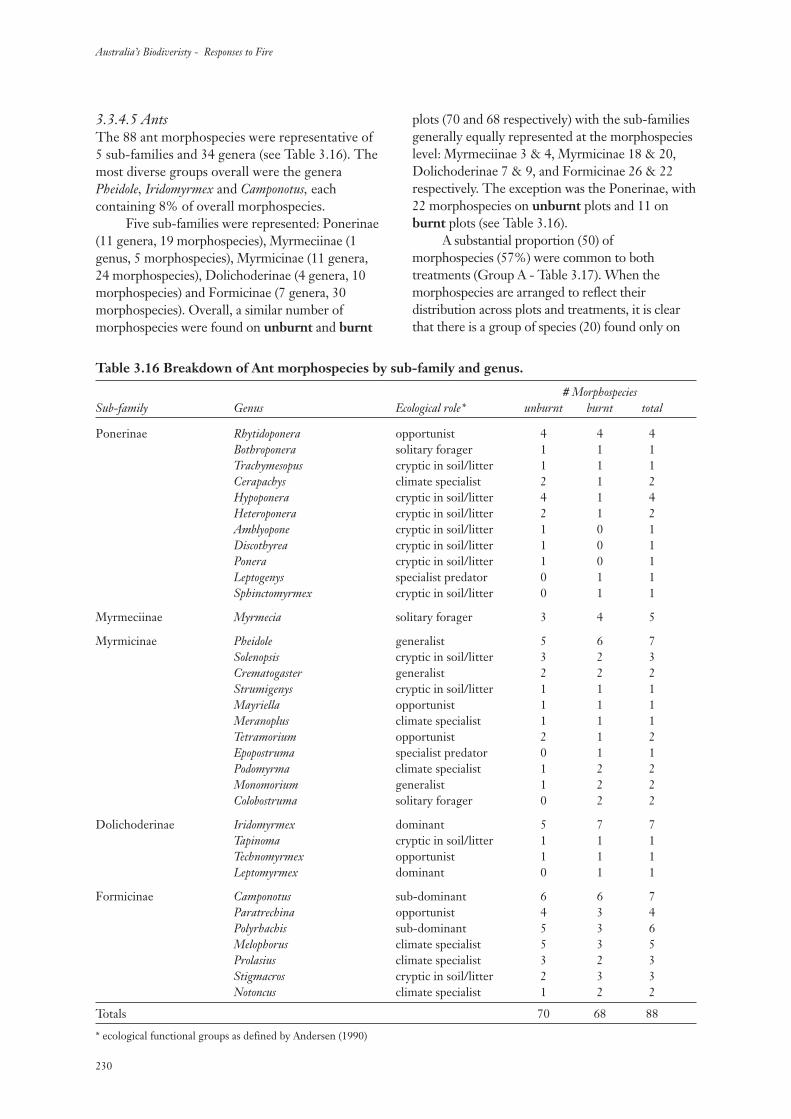
plots (70 and 68 respectively) with the sub-familiesgenerally equally represented at the morphospecieslevel: Myrmeciinae 3 & 4, Myrmicinae 18 & 20,Dolichoderinae 7 & 9, and Formicinae 26 & 22respectively. The exception was the Ponerinae, with22 morphospecies on unburnt plots and 11 onburnt plots (see Table 3.16).
A substantial proportion (50) ofmorphospecies (57%) were common to bothtreatments (Group A - Table 3.17). When themorphospecies are arranged to reflect theirdistribution across plots and treatments, it is clearthat there is a group of species (20) found only on
230
Australia’s Biodiveristy - Responses to Fire
3.3.4.5 AntsThe 88 ant morphospecies were representative of5 sub-families and 34 genera (see Table 3.16). Themost diverse groups overall were the generaPheidole, Iridomyrmex and Camponotus, eachcontaining 8% of overall morphospecies.
Five sub-families were represented: Ponerinae(11 genera, 19 morphospecies), Myrmeciinae (1genus, 5 morphospecies), Myrmicinae (11 genera,24 morphospecies), Dolichoderinae (4 genera, 10morphospecies) and Formicinae (7 genera, 30morphospecies). Overall, a similar number ofmorphospecies were found on unburnt and burnt
Table 3.16 Breakdown of Ant morphospecies by sub-family and genus.
# MorphospeciesSub-family Genus Ecological role* unburnt burnt total
Ponerinae Rhytidoponera opportunist 4 4 4Bothroponera solitary forager 1 1 1Trachymesopus cryptic in soil/litter 1 1 1Cerapachys climate specialist 2 1 2Hypoponera cryptic in soil/litter 4 1 4Heteroponera cryptic in soil/litter 2 1 2Amblyopone cryptic in soil/litter 1 0 1Discothyrea cryptic in soil/litter 1 0 1Ponera cryptic in soil/litter 1 0 1Leptogenys specialist predator 0 1 1Sphinctomyrmex cryptic in soil/litter 0 1 1
Myrmeciinae Myrmecia solitary forager 3 4 5
Myrmicinae Pheidole generalist 5 6 7Solenopsis cryptic in soil/litter 3 2 3Crematogaster generalist 2 2 2Strumigenys cryptic in soil/litter 1 1 1Mayriella opportunist 1 1 1Meranoplus climate specialist 1 1 1Tetramorium opportunist 2 1 2Epopostruma specialist predator 0 1 1Podomyrma climate specialist 1 2 2Monomorium generalist 1 2 2Colobostruma solitary forager 0 2 2
Dolichoderinae Iridomyrmex dominant 5 7 7Tapinoma cryptic in soil/litter 1 1 1Technomyrmex opportunist 1 1 1Leptomyrmex dominant 0 1 1
Formicinae Camponotus sub-dominant 6 6 7Paratrechina opportunist 4 3 4Polyrhachis sub-dominant 5 3 6Melophorus climate specialist 5 3 5Prolasius climate specialist 3 2 3Stigmacros cryptic in soil/litter 2 3 3Notoncus climate specialist 1 2 2
Totals 70 68 88
* ecological functional groups as defined by Andersen (1990)
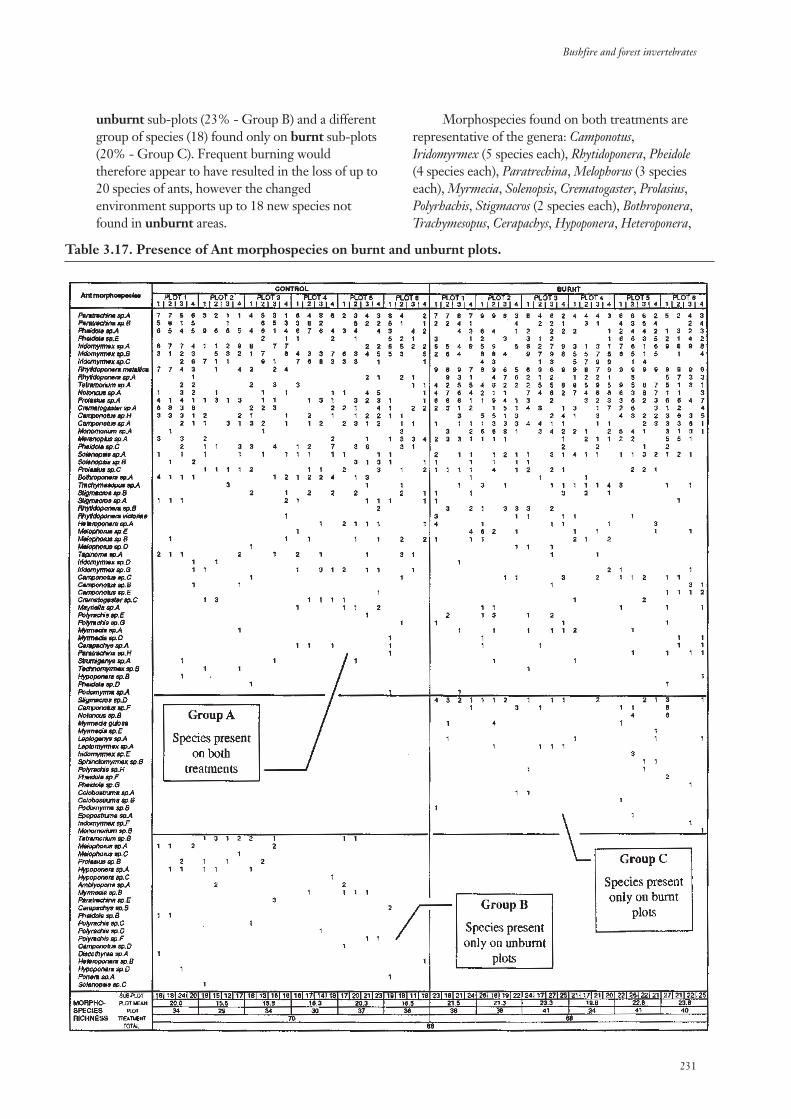
231
Bushfire and forest invertebrates
unburnt sub-plots (23% - Group B) and a differentgroup of species (18) found only on burnt sub-plots(20% - Group C). Frequent burning wouldtherefore appear to have resulted in the loss of up to20 species of ants, however the changedenvironment supports up to 18 new species notfound in unburnt areas.
Morphospecies found on both treatments arerepresentative of the genera: Camponotus,Iridomyrmex (5 species each), Rhytidoponera, Pheidole(4 species each), Paratrechina, Melophorus (3 specieseach), Myrmecia, Solenopsis, Crematogaster, Prolasius,Polyrhachis, Stigmacros (2 species each), Bothroponera,Trachymesopus, Cerapachys, Hypoponera, Heteroponera,
Table 3.17. Presence of Ant morphospecies on burnt and unburnt plots.

232
Australia’s Biodiveristy - Responses to Fire
Strumigenys, Mayriella, Meranoplus, Tetramorium,Podomyrma, Monomorium, Tapinoma, Technomyrmexand Notoncus (1 species each). Morphospecies foundonly on unburnt sub-plots are representative of thegenera: Polyrhachis, Hypoponera (3 species each),Melophorus (2 species), Cerapachys, Heteroponera,Amblyopone, Discothyrea, Ponera, Myrmecia, Pheidole,Solenopsis, Tetramorium, Camponotus, Paratrechinaand Prolasius (1 species each). Morphospecies foundonly on burnt sub-plots are representative of thegenera: Iridomyrmex, Myrmecia, Pheidole,Colobostruma (2 species each), Rhytidoponera,Leptogenys, Sphinctomyrmex, Epopostruma,Podomyrma, Monomorium, Leptomyrmex, Camponotus,Polyrhachis, Stigmacros and Notoncus (1 species each).
As many of these families are representedonly by single individuals or found only on onesub-plot of a treatment, it is difficult to identifyclear patterns from these tabulated data (at theindividual species level). At this stage it is apparenthowever that unburnt plots have more speciesfrom the genera Cerapachys (7 vs 1) and Hypoponera(4 vs 1), while burnt plots have greater numbersof species from the genus Colobostruma (2 vs 0).
Although this table of relative abundanceenables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed in the formof a “bi-plot” derived from the CCA ordination(see 2.5.3.3). This graphical display (Figure 3.25)shows the configuration of the environmentalvariables, the scatter of sub-plots, and therelationship between the two, giving an overviewof how community composition varies with theenvironment (Ter Braak 1986).
A number of features are apparent from thisbi-plot. Firstly, the lack of any overlap of unburntand burnt sub-plots in ordination space reflects thelargely dissimilar species assemblages of these twotreatments. Although 57% of morphospecies werefound on both treatments, their relative abundanceon each treatment differs substantially, leading tofundamentally different species assemblages.Secondly, the tighter clustering of burnt sub-plots
indicates a lower within-treatment diversitycompared to unburnt sub-plots (ie. a lower β-diversity). Richness values on burnt sub-plots wereon average 26% higher than on unburnt sub-plots(see Section 3.3.3.5), however the high similarity ofsub-plots for this treatment means that the overallrichness of unburnt and burnt plots was similar (70vs 68 species). Unburnt sub-plots have lowerrichness (α-diversity) but are less similar, resultingin higher “turnover” between sub-plots (higher β-diversity).
The third feature concerns the contributionof environmental variables to the differences inspecies composition for the two treatments. In thebi-plot (Figure 3.25) the length of the arrowsignifies the relative contribution of that variable tospecies composition, and the direction signifies itscontribution to the differences between treatments.
Unburnt sub-plots are characterised by highlevels of litter and high cover of tall and very tallshrubs. Due to the correlation between variables(see Section 3.2) these plots are also characterisedby high top-soil moisture levels and low andspatially variable amounts of insolation at groundlevel. Burnt sub-plots are characterised by highlevels of insolation at ground level, and to a lesserextent, steeper slopes, greater cover of the herb &shrub component of the understorey vegetation,
Table 3.18 Distribution of invertebrate morphospecies richness by treatment
Invertebrate both unburnt only burnt only totalTaxa # % # % # % #
Hemiptera (bugs) 7 16 18 41 19 43 44Diptera (flies) 35 45 31 40 11 15 77Araneae (spiders) 16 25 15 24 32 51 63Coleoptera (beetles) 39 28 47 34 53 38 139Formicidae (ants) 50 57 20 23 18 20 88
Total 147 - 131 - 133 - 411
Figure 3.25 Bi-plot from CCA ordination of Ant relativeabundance data. (Ellipses represent 1 standarddeviation unit around treatment centroids)

233
Bushfire and forest invertebrates
greater top-soil hardness and greater spatialpatchiness of twigs (0–10mm) and very tall shrubs.
The level of insolation at ground level, theamount of litter and the cover of tall shrubs make thegreatest contribution to the differences in speciescomposition between unburnt and burnt plots.Other environmental variables make only a minorcontribution to the observed differences in ant species.
3.3.4.6. SummaryThe five groups studies in detail proved to beextremely diverse. Beetles had the richness faunaoverall with 139 beetle morphospeciesrepresentative of nine super-families and 25families. The ants were the second richness groupwith 88 morphospecies representative of 5 sub-families and 34 genera. They were followed by theflies with 77 morphospecies representative of 2sub-orders and 20 families, the spiders with 63morphospecies representative of 21 families, andthe bugs with 44 morphospecies representative of16 family (or similar) groups.
Overall, the same number of morphospecies(279) were collected from unburnt and burntplots. Richness on unburnt sub-plots was, onaverage, 48.2 morphospecies, which was similar tothe average value on burnt sub-plots (46.5morphospecies). This initially suggests thatfrequent burning had not reduced biodiversity inthis forest environment. An analysis of therichness of individual groups (see Section 3.3.3)has shown this not to be the case, with groupsresponding differently to frequent burning. ThisSection (3.3.4) examined the nature of thatresponse by looking at the composition of faunalassemblages (communities).
An examination of the distribution ofmorphospecies across sub-plots for eachtreatment, detected a consistent pattern.Morphospecies fell into one of three groups: foundon both treatments (Group A), found only onunburnt plots (Group B), or found only on burntplots (Group C). The relative proportions ofmorphospecies in each category however variedsubstantially between taxonomic groups. ForHemiptera (bugs) the proportions were 16, 41 and43% for both, unburnt and burnt respectively; forDiptera (flies) 45, 40 and 15%; for spiders 25, 24and 51%; for beetles 28, 34 and 38%; and for ants57, 23 and 20% (see Table 3.18).
These results also suggest that frequentburning had led to the loss of up to 131 species(18 bugs, 31 flies, 15 spiders, 47 beetles and 20ants), which represents 47% of the morphospeciesknown from the unburnt areas. Overall
biodiversity of frequently burnt areas wasmaintained by the addition of species notrecorded on unburnt plots. The changedenvironment was supporting an additional 133morphospecies (19 bugs, 11 flies, 32 spiders, 53beetles and 18 ants).
Many of the morphospecies apparently lostfrom frequently burnt sites were however onlydetected on a single sub-plot or represented by asingle individual on unburnt plots. These could begenuinely rare or uncommon species which weremissed purely by chance when sampling burntplots. For this reason it is difficult to identify clearpatterns (at the species level) from the relativeabundance data alone. Some general trends wereapparent however when morphospecies data weregrouped into a higher taxonomic level. For bugs,unburnt plots had more species from the infra-order Dipsocoromorpha (7 vs 3), while burnt plotshave greater numbers of species from the familyReduviidae (5 vs 0). For flies, unburnt plots hadmore species from the families Sciaridae (8 vs 5),Phoridae (16 vs 13) and Empididae (6 vs 3). Forspiders, unburnt plots had more species from thefamily Malkaridae (2 vs 0), while burnt plots hadgreater numbers of species from the familiesZodariidae (9 vs 2), Gnaphosidae (6 vs 3),Corinnidae (7 vs 4), Linyphiidae (4 vs 2) andLycosidae (3 vs 0). For beetles, unburnt plots hadmore species from the family Carabidae (11 vs 8),while burnt plots had greater numbers of speciesfrom the families Curculionidae (18 vs 9) andChrysomelidae (8 vs 4). For ants, unburnt plotshad more species from the genera Cerapachys (7 vs1) and Hypoponera (4 vs 1), while burnt plots hadgreater numbers of species from the genusColobostruma (2 vs 0). The implication of thesechanges for community organisation andecosystem function are considered in Section 3.4.
Although an examination of relative abundancepatterns enables broad “assemblages” of species withsimilar responses to disturbance to be identified,these data are more clearly displayed and interpretedin the form of a “bi-plot” derived from the CCAordination. These graphical displays show theconfiguration of the environmental variables, thescatter of sub-plots, and the relationship between thetwo, giving an overview of how communitycomposition varies with the environment. Anexamination of the bi-plots revealed a number ofconsistent features. Firstly, the degree of overlap ofunburnt and burnt sub-plots in ordination spacereflected the similarity (or dissimilarity) of thespecies’ assemblages of the two treatments. For bugs,

234
Australia’s Biodiveristy - Responses to Fire
spiders, beetles and ants there was little or nooverlap, indicating low similarity of the twoassemblages. For flies however there was asubstantial overlap, reflecting the relatively largenumber of morphospecies shared by the twotreatments.
Secondly, the degree of clustering of the sub-plots from each treatment indicates the relativesimilarity of species’ assemblages on sub-plots andplots within each treatment. The tighter clusteringof burnt sub-plots for bugs, beetles and antsindicated a lower within-treatment diversitycompared to unburnt sub-plots (ie. a lower β-diversity). The converse applied for spiders, withthe tighter clustering of unburnt sub-plotsindicating a lower β-diversity compared to burntsub-plots. The situation for flies indicated similarwithin-treatment diversity for both treatments, withloose clustering of both unburnt and burnt sub-plots. This interaction between point richness (α-diversity) and spatial “turnover” of species (β-diversity) has substantial implications for theinterpretation of the apparent effect of repeatedburning and were previously identified as “scaleeffects” in Section 3.3.3.6. Similar patterns wereapparent for bugs and ants where richness values (α-diversity) on burnt sub-plots were on averagehigher than on unburnt sub-plots, however thehigh similarity of assemblages on burnt sub-plots(low β-diversity) meant that the overall richness ofboth treatments were similar. Unburnt sub-plotshad lower richness (α-diversity) but are less similar,resulting in higher “turnover” between sub-plots(higher β-diversity), increasing overall speciesrichness for that treatment. For flies and beetlesburnt plots had lower richness (α-diversity) butwere less similar, resulting in higher β-diversity.Spiders exhibited a different pattern with bothhigher α- and β-diversity for the burnt treatment,resulting in a large number of species (32) unique tofrequently burnt sites.
The third feature of the bi-plots concerns thecontribution of environmental variables to thedifferences in species composition for the twotreatments. In the bi-plots (Figures 3.21-5) the lengthof the arrows signify the relative contribution of thatvariable to species composition, and the directionsignifies their contribution to the differences betweentreatments. For all taxa unburnt sub-plots werecharacterised by high levels of litter, high cover of talland very tall shrubs, high top-soil moisture levels andlow and spatially variable amounts of insolation atground level. Similarly, burnt sub-plots werecharacterised by high levels of insolation at ground
level and, to a lesser extent, steeper slopes. Theseenvironmental variables make the greatestcontribution to the differences in species compositionbetween unburnt and burnt plots. For bugs, burntsub-plots were also characterised by greater top-soilhardness and greater cover of the herb & shrubcomponent of the understorey vegetation, with herband shrub cover also important for spiders and ants.For flies, burnt sites were also characterised bygreater exposure (more north-westerly aspects).Other environmental variables made only a minorcontribution to the observed differences in speciescomposition between burnt and unburnt treatmentsfor all taxa. Communities were therefore influencedby a combination of site-dependent (slope and aspect)and treatment-dependent (litter, insolation, herb &shrub cover, top-soil moisture & hardness)environmental variables.
3.3.5 Community Structure
While it is possible to describe and assesscommunities using indices such as speciesrichness, or to compare the relative abundance ofspecies using similarity indices, multi-variateapproaches and/or through graphicalrepresentation, these contribute little to anunderstanding of the processes underlying theirdifferences. In order to simplify and interpret thecomplexity of ecological systems, one approachhas been to group species into “guilds” or“functional groups”. These groups recognise theecological rather than the taxonomic affinity ofspecies. In this Section morphospecies wereallocated to guilds following reference to therelevant literature and discussions with taxonomicexperts. To maintain comparability between broadtaxonomic groups, the number of species fromeach group was averaged across the 24 sub-plotsfor each treatment. This method does not takeinto account the relative abundance of individualsof species and therefore is not unfairly biased by afew very abundant species.
3.3.5.1 HemipteraMorphospecies were classified into one of 5 groupsbased on the known habits and requirements at thefamily level (see Table 3.19). These groups wereprimarily based on feeding strategy, and secondarilyhabitat preferences. The groups were; primarilyphytophagous (Cicadellidae, Coccidae, Fulgoridaeand the unidentified Homopteran), moist habitatspecialists (Dipsocoromorpha), primarily predacious(Nabidae, Reduvidae) and others. This last categoryincluded morphospecies from the familiesEnicocephalidae, Anthocoridae, Miridae, Tingidae,

235
Bushfire and forest invertebrates
Thaumastocoridae, Lygaeidae, Plataspididae andPentatomidae which were only found on one sub-plot and therefore offering limited information tothis analysis.
On average, 64% of morphospecies onunburnt plots were primarily phytophagous (planteating) and 12% predacious on other invertebrates.Of the remaining 24%, approximately 23% areknown to be moist habitat specialists from the sub-order Dipsocoromorpha utilising a variety offeeding strategies. On average, 79% ofmorphospecies on burnt plots were primarilyphytophagous and 10% predacious on otherinvertebrates. Of the remaining 11%, approximately4% are known to be moist habitat specialists fromthe sub-order Dipsocoromorpha. The remaining7% of morphospecies on burnt plots were from thefamilies Anthocoridae, Miridae, Thaumastocoridae,Lygaeidae, Plataspidae and Pentatomidae (1 specieseach).
A graphical comparison of these data (Figure3.26) indicates that frequent burning has resultedin a fundamental shift in Hemipteran communitystructure. With regard to feeding strategy, therehas been, on average, a 15% increase in thenumber of phytophagous species. While thenumber of predacious species has remainedlargely unchanged, there has been a total shiftfrom species from the family Nabidae to the
family Reduviidae. The greatest change concernshabitat specialists from the sub-orderDipsocoromorpha where there has been, onaverage, an 83% reduction in the number ofmorphospecies.
3.3.5.2 DipteraMorphospecies were classified into one of 7 groupsbased on the known habits and requirements at thefamily level (see Table 3.20). These groups wereprimarily based on feeding strategy, andsecondarily habitat preferences. The groups were;primarily phytophagous (Cecidomyiidae),primarily predacious (Tachydromiinae), fungalfeeders (Sciaridae, Drosophilidae, Mycetophilidae& Scatopsidae), generalists and scavengers(Phoridae & Chloropidae), moist habitat specialists(Ceratopogonidae & Chironomidae), litterdwellers (Sphaeicieiidae & Tipilidae) and wide-ranging “tourists” (Piophilidae, Micropezidae,Dolichopodidae, Calliphoridae, Syrphidae,Thereuidae, Muscidae & Tachinidae).
On average, 29% of morphospecies onunburnt plots were primarily generalists andscavengers, 23% feeders on fungal products, 17%moist habitat specialists, and 11% phytophagous(plant eating). Of the remaining 20%,approximately 13% are known to be wide-ranging“tourists”, 6% predators, and 2% litter dwellers.On average, 26% of morphospecies on unburntplots were primarily generalists and scavengers,13% feeders on fungal products, 18% moisthabitat specialists, and 26% phytophagous (planteating). Of the remaining morphospecies,approximately 7% are known to be wide-ranging“tourists”, 6% predators, and 1% litter dwellers.
A graphical comparison of these data (Figure3.27) indicates that frequent burning has resultedin a fundamental shift in Dipteran communitystructure. With regard to feeding strategy, therehas been, on average, a 44% decrease in thenumber of morphospecies feeding on fungalproducts. This change was most marked in thefamilies Scaridae and Scatopsidae. The proportion
Table 3.19 Comparison of Hemipteran community structure on burnt and unburnt plots. Datarepresent mean percentage of morphospecies in each category.
Ecological role Taxa Unburnt Burnt
Primarily phytophagous Cicadellidae 48.3 75.7Coccidae, Fulgoridae, & Homoptera 15.4 3.3
Moist habitat specialists Dipsocoromorpha 22.9 3.9Primarily predacious Nabidae 11.7 0.0
Reduvidae 0.0 10.1Others (see text) 1.7 7.0
Total 100 100
Figure 3.26 Comparison of Hemipteran communitystructure on burnt and unburnt plots

With regard to micro-habitat preferences,the number of morphospecies specifically utilisingthe litter layer has, on average, decreased by 60%.This was primarily due to the absence of the familyTipulidae on frequently burnt sub-plots. Thenumber of moist habitat specialists remained similar,although the family Ceratopogonidae was morecommonly represented on burnt sub-plots andthe family Chironomidae on unburnt sub-plots.
3.3.5.3 SpidersMorphospecies were classified into one of 5 groupsbased on the known habits and requirements at thefamily level (see Table 3.21). As all spiders arefundamentally predacious, these groups wereprimarily based on habitat preferences. The groupswere; moist habitat specialists (Theridiidae,Toxopidae, Oonopidae, Malkaridae & Tekellidae),those with a known preference for dry habitats(Gnaphosidae & Corinnidae), litter dwellers(Hahniidae & Textricellidae), open & disturbedhabitat specialists (Linyphiidae & Zodariidae) andothers. This last category included generalisedhunters from the families Ctenidae, Dictynoidea,Heteropodidae, Micropholcommatidae,
236
Australia’s Biodiveristy - Responses to Fire
of predacious species on sub-plots has remainedsimilar, however the number of phytophagousspecies has, on average, increased by 140%. Theseadditional species were from the familyCecidomyiidae. The number of generalists andscavengers has decreased slightly (11%). Thenumber of morphospecies regarded as wide-ranging “tourists” was, on average, reduced by43% on frequently burnt plots.
Table 3.21 Comparison of Spider community structure on burnt and unburnt plots. Data representmean percentage of morphospecies in each category.
Ecological role Family Unburnt Burnt
Theridiidae 22.2 1.8Moist habitat Toxopidae 11.4 1.2specialists Oonopidae 8.1 0.4
Malkaridae 4.0 0.0Tekellidae 4.3 2.2
Dry habitat Gnaphosidae 5.8 8.1preference Corinnidae 14.6 18.4Litter Hahniidae 4.5 10.4dwellers Textricellidae 5.5 10.3Open/disturbed Linyphiidae 3.1 15.7habitat specialists Zodariidae 2.1 20.1Others (see text) 14.3 11.3
Total 100 100
Table 3.20 Comparison of Fly community structure on burnt and unburnt plots. Data representmean percentage of morphospecies in each category.
Ecological role Taxa Unburnt Burnt
Fungal feeders Sciaridae, Drosophilidae, Mycetophilidae & Scatopsidae 23.2 12.9Litter dwellers Sphaeicieiidae & Tipilidae 2.0 0.8Tourists Piophilidae, Micropezidae, Dolichopodidae,
Calliphoridae, Syrphidae, Thereuidae, Muscidae & Tachinidae 12.6 7.2Moist habitat specialists Ceratopogonidae & Chironomidae 17.0 17.8Generalists/scavengers Phoridae & Chloropidae 29.1 25.9Predators Tachydromiinae 5.8 5.8Phytophagous Cecidomyiidae 10.3 25.8
Total 100 100
Figure 3.27 Comparison of Fly community structure onburnt and unburnt plots
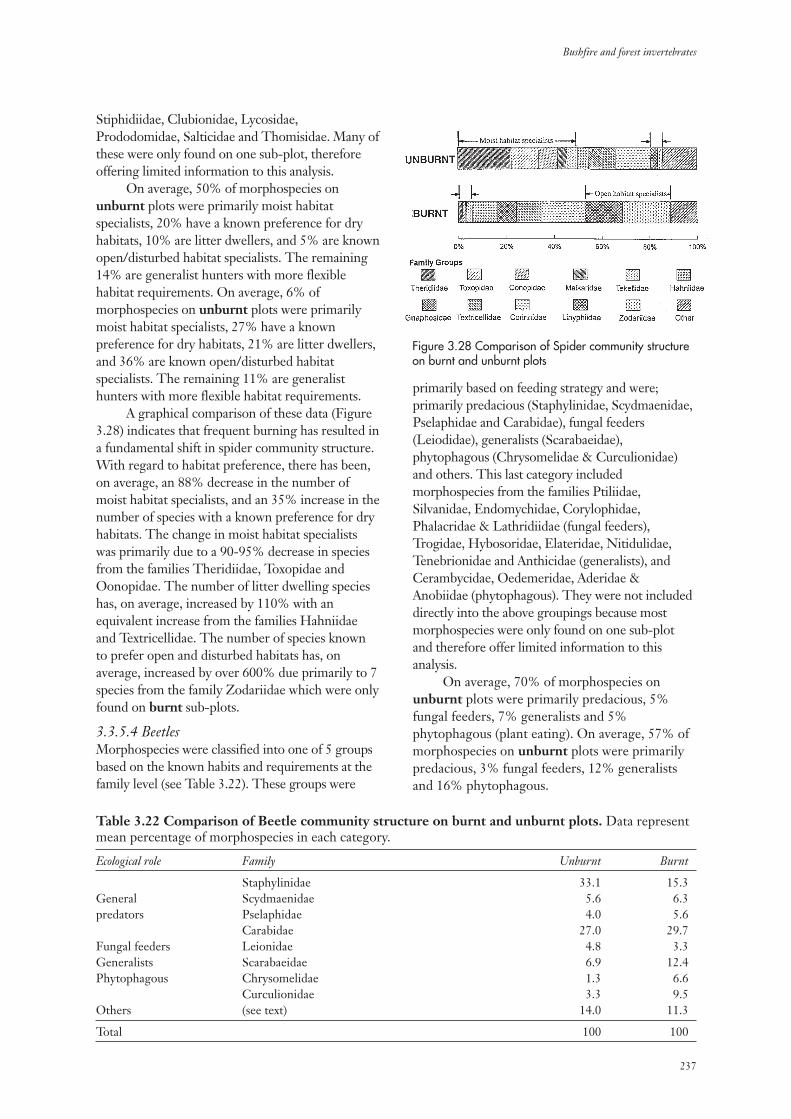
237
Bushfire and forest invertebrates
Stiphidiidae, Clubionidae, Lycosidae,Prododomidae, Salticidae and Thomisidae. Many ofthese were only found on one sub-plot, thereforeoffering limited information to this analysis.
On average, 50% of morphospecies onunburnt plots were primarily moist habitatspecialists, 20% have a known preference for dryhabitats, 10% are litter dwellers, and 5% are knownopen/disturbed habitat specialists. The remaining14% are generalist hunters with more flexiblehabitat requirements. On average, 6% ofmorphospecies on unburnt plots were primarilymoist habitat specialists, 27% have a knownpreference for dry habitats, 21% are litter dwellers,and 36% are known open/disturbed habitatspecialists. The remaining 11% are generalisthunters with more flexible habitat requirements.
A graphical comparison of these data (Figure3.28) indicates that frequent burning has resulted ina fundamental shift in spider community structure.With regard to habitat preference, there has been,on average, an 88% decrease in the number ofmoist habitat specialists, and an 35% increase in thenumber of species with a known preference for dryhabitats. The change in moist habitat specialistswas primarily due to a 90-95% decrease in speciesfrom the families Theridiidae, Toxopidae andOonopidae. The number of litter dwelling specieshas, on average, increased by 110% with anequivalent increase from the families Hahniidaeand Textricellidae. The number of species knownto prefer open and disturbed habitats has, onaverage, increased by over 600% due primarily to 7species from the family Zodariidae which were onlyfound on burnt sub-plots.
3.3.5.4 BeetlesMorphospecies were classified into one of 5 groupsbased on the known habits and requirements at thefamily level (see Table 3.22). These groups were
primarily based on feeding strategy and were;primarily predacious (Staphylinidae, Scydmaenidae,Pselaphidae and Carabidae), fungal feeders(Leiodidae), generalists (Scarabaeidae),phytophagous (Chrysomelidae & Curculionidae)and others. This last category includedmorphospecies from the families Ptiliidae,Silvanidae, Endomychidae, Corylophidae,Phalacridae & Lathridiidae (fungal feeders),Trogidae, Hybosoridae, Elateridae, Nitidulidae,Tenebrionidae and Anthicidae (generalists), andCerambycidae, Oedemeridae, Aderidae &Anobiidae (phytophagous). They were not includeddirectly into the above groupings because mostmorphospecies were only found on one sub-plotand therefore offer limited information to thisanalysis.
On average, 70% of morphospecies onunburnt plots were primarily predacious, 5%fungal feeders, 7% generalists and 5%phytophagous (plant eating). On average, 57% ofmorphospecies on unburnt plots were primarilypredacious, 3% fungal feeders, 12% generalistsand 16% phytophagous.
Table 3.22 Comparison of Beetle community structure on burnt and unburnt plots. Data representmean percentage of morphospecies in each category.
Ecological role Family Unburnt Burnt
Staphylinidae 33.1 15.3General Scydmaenidae 5.6 6.3predators Pselaphidae 4.0 5.6
Carabidae 27.0 29.7Fungal feeders Leionidae 4.8 3.3Generalists Scarabaeidae 6.9 12.4Phytophagous Chrysomelidae 1.3 6.6
Curculionidae 3.3 9.5Others (see text) 14.0 11.3
Total 100 100
Figure 3.28 Comparison of Spider community structureon burnt and unburnt plots

238
Australia’s Biodiveristy - Responses to Fire
A graphical comparison of these data (Figure3.29) indicates that frequent burning has resulted ina fundamental shift in beetle community structure.With regard to feeding strategy, there has been, onaverage, an 18% decrease in the number of predatorspecies, due primarily to a 54% reduction in thenumber of morphospecies from the familyStaphylinidae. The average number ofmorphospecies feeding on fungal products (asrepresented by the family Leiodidae) has declinedby 31%, with some other fungal feeders (familiesPtiliidae & Endomychidae) decreasing by over200%. The generalists, as represented by the familyScarabaeidae, have increased on average by 79%.Phytophagous species have increased dramatically(up by 250%), particularly morphospecies from thefamily Chrysomelidae. For this family, 56% of itsoverall morphospecies were only found onfrequently burnt sub-plots.
3.3.5.5 AntsMorphospecies were classified into one of 7“functional groups” groups based on the knownhabits and requirements at the generic level (seeAndersen 1990). These groups were; dominants(Iridomyrmex & Leptomyrmex), sub-dominants(Camponotus & Polyrhachis), climate specialists(Cerapachys, Melophorus, Meranoplus, Notoncus,Podomyrma & Prolasius), cryptic species(Amblyopone, Discothyrea, Heteroponera, Hypoponera,Ponera, Solenopsis, Sphinctomyrmex, Stigmacros,Tapinoma and Trachymesopus), opportunists(Mayriella, Paratrechina, Rhytidoponera,Technomyrmex, Tetramorium, Crematogaster,Monomorium & Pheidole) and solitary/specialistspecies (Bothroponera, Colobostruma, Epopostruma,Leptogenys & Myrmecia). The average number ofmorphospecies from these genera on eachtreatment are shown in Table 3.23.
On average, 16% of morphospecies onunburnt plots were dominants, 10% sub-dominants, 18% climate specialists, 16% crypticspecies, 19% opportunists, 17% generalists, and 4%solitary/ specialist species. On average, 16% ofmorphospecies on unburnt plots were dominants,10% sub-dominants, 18% climate specialists, 16%cryptic species, 19% opportunists, 17% generalists,and 4% solitary/specialist species.
A graphical comparison of these data (Figure3.30) indicates that frequent burning has resulted ina slight shift in ant community structure. Thenumber of morphospecies within the “dominant”functional group has, on average, decreased by 23%,primarily through a reduction in the occurrence ofIridomyrmex species. The number of “sub-dominants” has increased slightly (25%), largelybecause of an increase in the occurrence of
Figure 3.29 Comparison of Beetle community structureon burnt and unburnt plots
Table 3.23 Comparison of Ant communitystructure on burnt and unburnt plots. Datarepresent mean percentage of morphospecies ineach category.
Ecological role Taxa Unburnt Burnt
Dominants Iridomyrmex 15.6 12.0Leptomyrmex 0.0 0.7
Sub-dominants Camponotus 8.7 10.7Polyrhachis 1.3 1.8Cerapachys 1.2 0.8Melophorus 3.3 3.3
Climate Meranoplus 2.1 3.1Specialists Notoncus 2.1 4.5
Podomyrma 0.2 0.4Prolasius 8.2 6.2Amblyopone 0.5 0.0Discothyrea 0.2 0.0Heteroponera 1.6 1.1Hypoponera 1.7 0.2
Cryptic Ponera 0.2 0.0Species Solenopsis 4.6 4.4
Sphinctomyrmex 0.0 0.4Stigmacros 3.6 4.1Tapinoma 2.5 0.4Trachymesopus 0.8 2.3Mayriella 0.9 0.8Paratrechina 9.9 8.4
Opportunists Rhytidoponera 3.7 10.5Technomyrmex 0.5 0.2Tetramorium 3.8 4.6Crematogaster 4.9 3.9
Generalists Monomorium 0.8 3.8Pheidole 11.5 7.4Bothroponera 2.8 0.7Colobostruma 0.0 0.6
Solitary/ Epopostruma 0.0 0.2Specialists Leptogenys 0.0 0.8
Myrmecia 1.4 2.5
Total 100 100

239
Bushfire and forest invertebrates
Camponotus species. With regard to “climatespecialists”, they have remained largely stable(increased by 4%) as a group, although individualgenera did vary in their response (see Table 3.23).There was a decrease (19%) in the average numberof “cryptic” morphospecies, particularly with regardto the genera Hypoponera (90%) and Tapinoma(86%). A number of morphospecies from this grouphowever increased in their average occurrence;Stigmacros (13%) and Trachymesopus (196%),although the actual number of species involved issmall (overall 3 and 1 respectively).
There was a substantial increase overall inthe average number of “opportunist”morphospecies (30%), primarily due to one genus,Rhytidoponera, which increased by 180%. In factthis was attributable to a single species:Rhytidoponera metallica, which was extremelynumerous on burnt sub-plots and contributed tothe high ant abundance detected for this treatment(see Section 3.3.2.10). The number of “generalist”morphospecies, on average, decreased by 11%following frequent burning. Results were notconsistent within the group with the generaPheidole and Crematogaster decreasing by 35 and19% respectively, and the genus Monomoriumincreasing by 388%. The number of larger“solitary/specialist” morphospecies increasedslightly (13%), although the results were quitevariable within the group. Three new genera werefound on burnt plots (Colobostruma, Epopostruma &Leptogenys) while the numbers of species ofMyrmecia increased, on average, by 87%. Theoccurrence of the solitary forager Bothroponera sp.A decreased substantially (76%).
3.3.5.6 SummaryThis Section examined community structure bygrouping morphospecies into ecological groupsbased upon their feeding strategies and habitatpreferences. By classifying large numbers ofspecies into smaller, more manageable groups, it ispossible to substantially reduce the apparentcomplexity of ecological systems and provide abasis for evaluating environmental change(Andersen 1990).
For all groups studied, frequent burningresulted in a change in the structure of thecommunity. With regard to feeding strategy, therewas on average, a 15, 140 and 250% increase(respectively) in the number of phytophagous(plant feeding) species of bugs, flies and beetles.With regard to the proportion of predator species,it remained unchanged for flies and bugs, howeverfor bugs there was a total shift from the familyNabidae to the family Reduviidae. With beetlesthere was, on average, an 18% decrease in thenumber of predator species, due primarily to a 54%reduction in the number of morphospecies fromthe family Staphylinidae. For groups feedingprimarily on fungal products there was, on average,a 44% (flies) and 31% (beetles) decrease in thenumber of morphospecies. For flies this was mostmarked in the families Scaridae and Scatopsidae,and for beetles, in the families Leiodidae, Ptiliidae& Endomychidae. For flies, the number ofgeneralists and scavengers has decreased slightly(11%), while for beetles the proportion ofgeneralists has increased, on average, by 79%,primarily through an increase in morphospeciesfrom the family Scarabaeidae. With ants, theproportion of generalist morphospecies decreased,on average, by 11% following frequent burning.Results were not consistent within the group withthe genera Pheidole and Crematogaster decreasing by35 and 19% respectively, and the genusMonomorium increasing by 388%. For flies, thenumber of morphospecies regarded as wide-ranging “tourists” was, on average, reduced by 43%on frequently burnt plots.
When groups were compared with regard tohabitat preferences, it was apparent that therewere substantial changes in community structurefor some taxa. For bugs, numbers of moist habitatspecialists from the sub-order Dipsocoromorphawere, on average, reduced by 83%. Amongstspiders, moist habitat specialists were reduced by88%, primarily due to a 90-95% decrease innumbers of morphospecies from the families
Figure 3.30 Comparison of Ant community structure onburnt and unburnt plots
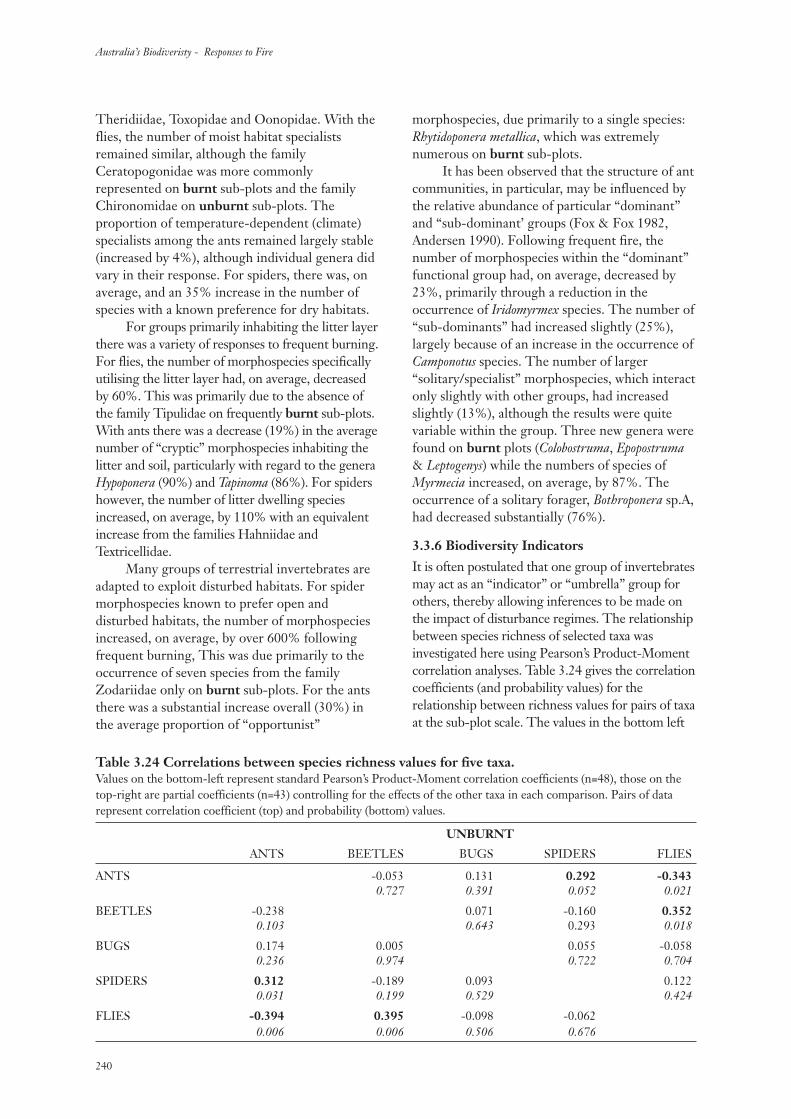
morphospecies, due primarily to a single species:Rhytidoponera metallica, which was extremelynumerous on burnt sub-plots.
It has been observed that the structure of antcommunities, in particular, may be influenced bythe relative abundance of particular “dominant”and “sub-dominant’ groups (Fox & Fox 1982,Andersen 1990). Following frequent fire, thenumber of morphospecies within the “dominant”functional group had, on average, decreased by23%, primarily through a reduction in theoccurrence of Iridomyrmex species. The number of“sub-dominants” had increased slightly (25%),largely because of an increase in the occurrence ofCamponotus species. The number of larger“solitary/specialist” morphospecies, which interactonly slightly with other groups, had increasedslightly (13%), although the results were quitevariable within the group. Three new genera werefound on burnt plots (Colobostruma, Epopostruma& Leptogenys) while the numbers of species ofMyrmecia increased, on average, by 87%. Theoccurrence of a solitary forager, Bothroponera sp.A,had decreased substantially (76%).
3.3.6 Biodiversity Indicators
It is often postulated that one group of invertebratesmay act as an “indicator” or “umbrella” group forothers, thereby allowing inferences to be made onthe impact of disturbance regimes. The relationshipbetween species richness of selected taxa wasinvestigated here using Pearson’s Product-Momentcorrelation analyses. Table 3.24 gives the correlationcoefficients (and probability values) for therelationship between richness values for pairs of taxaat the sub-plot scale. The values in the bottom left
240
Australia’s Biodiveristy - Responses to Fire
Theridiidae, Toxopidae and Oonopidae. With theflies, the number of moist habitat specialistsremained similar, although the familyCeratopogonidae was more commonlyrepresented on burnt sub-plots and the familyChironomidae on unburnt sub-plots. Theproportion of temperature-dependent (climate)specialists among the ants remained largely stable(increased by 4%), although individual genera didvary in their response. For spiders, there was, onaverage, and an 35% increase in the number ofspecies with a known preference for dry habitats.
For groups primarily inhabiting the litter layerthere was a variety of responses to frequent burning.For flies, the number of morphospecies specificallyutilising the litter layer had, on average, decreasedby 60%. This was primarily due to the absence ofthe family Tipulidae on frequently burnt sub-plots.With ants there was a decrease (19%) in the averagenumber of “cryptic” morphospecies inhabiting thelitter and soil, particularly with regard to the generaHypoponera (90%) and Tapinoma (86%). For spidershowever, the number of litter dwelling speciesincreased, on average, by 110% with an equivalentincrease from the families Hahniidae andTextricellidae.
Many groups of terrestrial invertebrates areadapted to exploit disturbed habitats. For spidermorphospecies known to prefer open anddisturbed habitats, the number of morphospeciesincreased, on average, by over 600% followingfrequent burning, This was due primarily to theoccurrence of seven species from the familyZodariidae only on burnt sub-plots. For the antsthere was a substantial increase overall (30%) inthe average proportion of “opportunist”
Table 3.24 Correlations between species richness values for five taxa.Values on the bottom-left represent standard Pearson’s Product-Moment correlation coefficients (n=48), those on thetop-right are partial coefficients (n=43) controlling for the effects of the other taxa in each comparison. Pairs of datarepresent correlation coefficient (top) and probability (bottom) values.
UNBURNT
ANTS BEETLES BUGS SPIDERS FLIES
ANTS -0.053 0.131 0.292 -0.3430.727 0.391 0.052 0.021
BEETLES -0.238 0.071 -0.160 0.3520.103 0.643 0.293 0.018
BUGS 0.174 0.005 0.055 -0.0580.236 0.974 0.722 0.704
SPIDERS 0.312 -0.189 0.093 0.1220.031 0.199 0.529 0.424
FLIES -0.394 0.395 -0.098 -0.0620.006 0.006 0.506 0.676
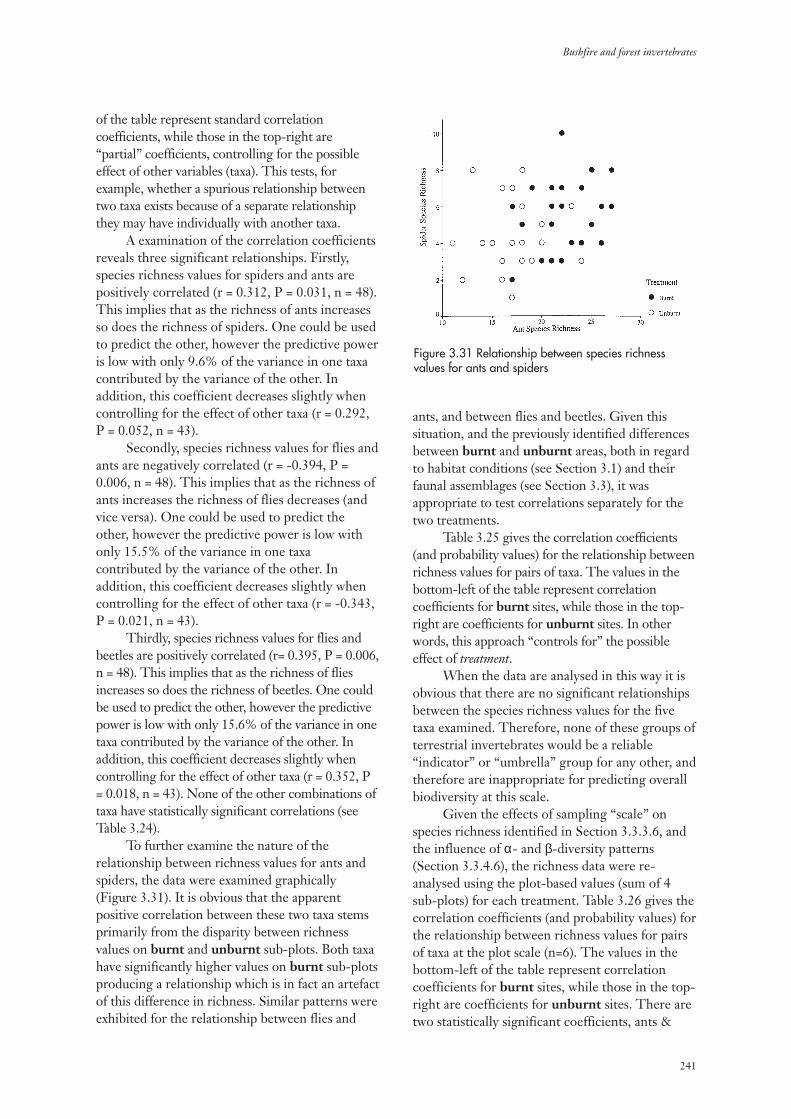
241
Bushfire and forest invertebrates
of the table represent standard correlationcoefficients, while those in the top-right are“partial” coefficients, controlling for the possibleeffect of other variables (taxa). This tests, forexample, whether a spurious relationship betweentwo taxa exists because of a separate relationshipthey may have individually with another taxa.
A examination of the correlation coefficientsreveals three significant relationships. Firstly,species richness values for spiders and ants arepositively correlated (r = 0.312, P = 0.031, n = 48).This implies that as the richness of ants increasesso does the richness of spiders. One could be usedto predict the other, however the predictive poweris low with only 9.6% of the variance in one taxacontributed by the variance of the other. Inaddition, this coefficient decreases slightly whencontrolling for the effect of other taxa (r = 0.292,P = 0.052, n = 43).
Secondly, species richness values for flies andants are negatively correlated (r = -0.394, P =0.006, n = 48). This implies that as the richness ofants increases the richness of flies decreases (andvice versa). One could be used to predict theother, however the predictive power is low withonly 15.5% of the variance in one taxacontributed by the variance of the other. Inaddition, this coefficient decreases slightly whencontrolling for the effect of other taxa (r = -0.343,P = 0.021, n = 43).
Thirdly, species richness values for flies andbeetles are positively correlated (r= 0.395, P = 0.006, n = 48). This implies that as the richness of fliesincreases so does the richness of beetles. One couldbe used to predict the other, however the predictivepower is low with only 15.6% of the variance in onetaxa contributed by the variance of the other. Inaddition, this coefficient decreases slightly whencontrolling for the effect of other taxa (r = 0.352, P= 0.018, n = 43). None of the other combinations oftaxa have statistically significant correlations (seeTable 3.24).
To further examine the nature of therelationship between richness values for ants andspiders, the data were examined graphically(Figure 3.31). It is obvious that the apparentpositive correlation between these two taxa stemsprimarily from the disparity between richnessvalues on burnt and unburnt sub-plots. Both taxahave significantly higher values on burnt sub-plotsproducing a relationship which is in fact an artefactof this difference in richness. Similar patterns wereexhibited for the relationship between flies and
ants, and between flies and beetles. Given thissituation, and the previously identified differencesbetween burnt and unburnt areas, both in regardto habitat conditions (see Section 3.1) and theirfaunal assemblages (see Section 3.3), it wasappropriate to test correlations separately for thetwo treatments.
Table 3.25 gives the correlation coefficients(and probability values) for the relationship betweenrichness values for pairs of taxa. The values in thebottom-left of the table represent correlationcoefficients for burnt sites, while those in the top-right are coefficients for unburnt sites. In otherwords, this approach “controls for” the possibleeffect of treatment.
When the data are analysed in this way it isobvious that there are no significant relationshipsbetween the species richness values for the fivetaxa examined. Therefore, none of these groups ofterrestrial invertebrates would be a reliable“indicator” or “umbrella” group for any other, andtherefore are inappropriate for predicting overallbiodiversity at this scale.
Given the effects of sampling “scale” onspecies richness identified in Section 3.3.3.6, andthe influence of α- and β-diversity patterns(Section 3.3.4.6), the richness data were re-analysed using the plot-based values (sum of 4sub-plots) for each treatment. Table 3.26 gives thecorrelation coefficients (and probability values) forthe relationship between richness values for pairsof taxa at the plot scale (n=6). The values in thebottom-left of the table represent correlationcoefficients for burnt sites, while those in the top-right are coefficients for unburnt sites. There aretwo statistically significant coefficients, ants &
Figure 3.31 Relationship between species richnessvalues for ants and spiders
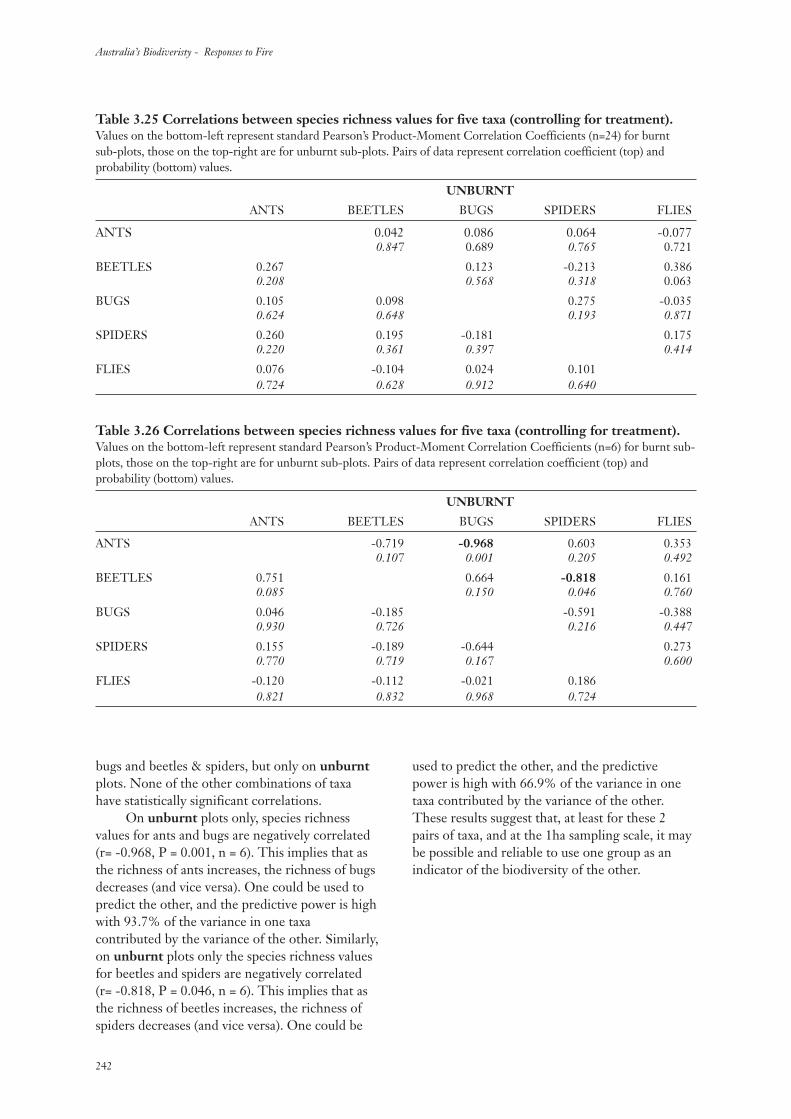
used to predict the other, and the predictivepower is high with 66.9% of the variance in onetaxa contributed by the variance of the other.These results suggest that, at least for these 2pairs of taxa, and at the 1ha sampling scale, it maybe possible and reliable to use one group as anindicator of the biodiversity of the other.
242
Australia’s Biodiveristy - Responses to Fire
bugs and beetles & spiders, but only on unburntplots. None of the other combinations of taxahave statistically significant correlations.
On unburnt plots only, species richnessvalues for ants and bugs are negatively correlated(r= -0.968, P = 0.001, n = 6). This implies that asthe richness of ants increases, the richness of bugsdecreases (and vice versa). One could be used topredict the other, and the predictive power is highwith 93.7% of the variance in one taxacontributed by the variance of the other. Similarly,on unburnt plots only the species richness valuesfor beetles and spiders are negatively correlated(r= -0.818, P = 0.046, n = 6). This implies that asthe richness of beetles increases, the richness ofspiders decreases (and vice versa). One could be
Table 3.25 Correlations between species richness values for five taxa (controlling for treatment).Values on the bottom-left represent standard Pearson’s Product-Moment Correlation Coefficients (n=24) for burnt sub-plots, those on the top-right are for unburnt sub-plots. Pairs of data represent correlation coefficient (top) andprobability (bottom) values.
UNBURNT
ANTS BEETLES BUGS SPIDERS FLIES
ANTS 0.042 0.086 0.064 -0.0770.847 0.689 0.765 0.721
BEETLES 0.267 0.123 -0.213 0.3860.208 0.568 0.318 0.063
BUGS 0.105 0.098 0.275 -0.0350.624 0.648 0.193 0.871
SPIDERS 0.260 0.195 -0.181 0.1750.220 0.361 0.397 0.414
FLIES 0.076 -0.104 0.024 0.1010.724 0.628 0.912 0.640
Table 3.26 Correlations between species richness values for five taxa (controlling for treatment).Values on the bottom-left represent standard Pearson’s Product-Moment Correlation Coefficients (n=6) for burnt sub-plots, those on the top-right are for unburnt sub-plots. Pairs of data represent correlation coefficient (top) andprobability (bottom) values.
UNBURNT
ANTS BEETLES BUGS SPIDERS FLIES
ANTS -0.719 -0.968 0.603 0.3530.107 0.001 0.205 0.492
BEETLES 0.751 0.664 -0.818 0.1610.085 0.150 0.046 0.760
BUGS 0.046 -0.185 -0.591 -0.3880.930 0.726 0.216 0.447
SPIDERS 0.155 -0.189 -0.644 0.2730.770 0.719 0.167 0.600
FLIES -0.120 -0.112 -0.021 0.1860.821 0.832 0.968 0.724

243
There is good theoretical and growing empiricalevidence to support the role of biodiversity in themaintenance of ecological processes withinforests. The multitude of organisms thatconstitute biodiversity play an essential role inprimary production, nutrient cycling and uptake,population and community level interactions andenergy storage and transfer (see Majer 1992b;Woodward 1993; Beattie 1995). Through theircontribution to ecosystem function, theseorganisms also enable forest ecosystems toprovide benefits to humanity. These includeamenity values in the form of aesthetics, recreationand education; heritage values as forests contributeto long-term security for catchment protection,air & water quality and nature conservation; andeconomic values including timber production,grazing and ecotourism (see Hobbs 1992; York1993; New 1995).
The maintenance of biodiversity is afundamental principle underlying ecologicallysustainable management (NSESD 1992). StateForests of New South Wales, through its CorporatePlan (1992), has stated that it will manage itsforests on an ecologically sustainable basis usingbest forest practices. Biodiversity conservation,and hence ecological sustainability, cannot beachieved without consideration of the importantrole that invertebrates play, both through theirinvolvement in ecological processes, and by theirsubstantial contribution to the overall richness ofbiological communities. Invertebrates are themost diverse and abundant animals in mostnatural ecosystems, but their importance insustaining those systems is commonly notappreciated (New 1995). Periodic low-intensityfire (hazard-reduction burning) is a conspicuousmanagement strategy in virtually all of Australia’sdry forest communities. While it is primarily usedto reduce fuel levels, little is known about theeffects of its repeated use on natural ecosystemsover long time-scales. The primary objective ofthis study was therefore to assess whether thefrequent use of this forest management practicewas compatible with the conservation of a majorcomponent of biodiversity: the terrestrialinvertebrates.
4.1 HABITAT STRUCTURE
Low intensity fires used for fuel control generallyresult in incomplete combustion of surface litterand understorey vegetation (Tolhurst et al. 1992;Williams and Gill 1995). A mosaic of habitatpatches results at a small scale, with these patchesinfluencing the spatial distribution of survivingterrestrial invertebrates and their ability torecolonise burnt areas. If repeated low-intensityfires reduce this spatial heterogeneity (see Fox andFox 1986; Nieuwenhuis 1987) then this practicemay have long-term consequences for the survivalof invertebrate populations. In this study, 31environmental variables were measured in orderto firstly, assess the long-term effect of frequentburning on the habitat, and secondly, toinvestigate the nature of relationships betweenhabitat components and invertebrate biodiversity.
Four general trends with regard toenvironmental variables were detected: large-scalespatial patterns, site-dependent patterns,treatment-dependent patterns, and generalpatterns independent of site and treatment. At thelargest scale (the study area), there was a gradualN-S decrease in the mean biomass of leaves as acomponent of the leaf litter. This pattern was onlyevident however on unburnt plots anddisappeared with frequent burning, resulting in asimplification of the large-scale spatial patterningof this habitat component. Top-soil moisturecontent tended to rise to a peak towards to centreof the N-S road transect and then decline towardsthe southern end, with frequent burning havingno effect on this spatial trend.
Frequent low-intensity fire had resulted in anumber of treatment-related changes in measuredhabitat parameters. Within the “fine-fuel”component, there was (on average) a 44%reduction in leaf biomass, and a reduction inlarge-scale spatial trends (see above). There was(on average) a 63% reduction in the very finelitter component, with the appearance of spatialpatterning not evident on unburnt plots. With thetwig component, there had been (on average) a50% reduction in the biomass of twigs 0-6mm,and a 44% reduction in the biomass of twigs 6-25mm. Bark biomass had decreased (on average)
4. DISCUSSION

sized shrubs, the spatial homogeneity of certainlitter components (biomass of leaves and very finematerial) and top-soil hardness (but see site-dependent effects mentioned above). There wasalso no obvious change with regard to thedistribution of sticks & logs (>2.5cm) withinnominated size categories, however some changesto the external nature of logs (charring) wasapparent.
There were a number of “general patterns”with regard to the response of environmentalvariables to frequent fire. As the amount ofvegetation in the first metre above the groundincreased, it became more spatially homogeneous(less “patchy”). The amount of leaf litter (twigs,bark, leaves and very fine material) at study siteswas independent of the amount of understoreyvegetation, but was correlated (positively) withtop-soil moisture levels and (negatively) with theamount and spatial variability of light reaching theground. This suggests that frequent fire has anindependent influence on vegetation understoreyand leaf-litter habitats, and that (in this forest) theleaf-litter environment exerts the primary controlon top-soil moisture and surface insolation levels.Christensen (1985) found that the removal of thelitter layer by fire caused increased surfaceheating, leading to greater evaporation and lowermoisture in the upper soil. This situation wouldbe expected to change with the time-relatedaccumulation of litter after fire.
While the changes to the amount of leaflitter, understorey vegetation and top-soilmoisture may only reflect a time-since-last-firephenomenon, changes to the spatial variabilityfollowing frequent burning may reflect a decreasein habitat heterogeneity which, in turn, couldimpact upon terrestrial invertebrate communities.There were measured reductions in the large-scale spatial patterning of the leaf litter, andchanges to its physical structure with an increasein the patchiness of bark and very fine littercomponents. The increased patchiness of groundinsolation levels would appear primarily to be aresponse to these changes. There was a decreasein the spatial heterogeneity of ground herbs &small shrubs but an increase for tall & very tallshrubs. This potentially reflects a change in thecomposition of the understorey vegetationfollowing frequent burning (see Fox and Fox1986; Nieuwenhuis 1987) and a removal of taller(older) shrubs due to the short interval betweenfires. The relationship of these environmental
244
Australia’s Biodiveristy - Responses to Fire
by 36%, and increased in spatial heterogeneity.These reductions in litter biomass largely reflect aresponse to the most recent fuel-reduction burn (2 years previously) and would be expected tochange with time as fuel continues to accumulate(see Birk and Bridges 1989). Over the 20 yearperiod of frequent hazard-reduction burning,litter biomass (fine fuel) had fluctuated between 15 and 23 tonnes ha-1 on unburnt plots, andbetween 4 and 20 tonnes ha-1 on burnt plots(York 1996). Burning removed between 46 and73% of litter (by weight), but by 3 years post-firelitter levels had usually reached and oftenexceeded that achieved before the previous fire(range = 72–136%). At the time of this study meanlitter biomass was 9.3 tonnes ha-1 on burnt plots,representing about 50% of levels on unburntplots.
Other treatment-related effects involved theamount (cover) of vegetation in the understoreyand its spatial distribution. While the quantity ofvegetation in the layers closest to the ground(ground herbs and small shrubs) was not affectedby frequent burning (see below), there was andecrease in the spatial heterogeneity(“patchiness”) of these layers. Conversely, therewas a substantial reduction in the cover of tall andvery tall shrubs (on average, 65% and 93%respectively). Both these layers showed an increasein spatial heterogeneity with frequent burning.Top-soil moisture levels were, on average, 18%lower following 20 years of frequent burning,whereas the amount of light reaching ground levelhad increased (on average) by 125%, andincreased in its spatial heterogeneity.
Spatial patterning of a number ofenvironmental variables was largely site-dependent, with no overall large-scale ortreatment-related features. Top-soil hardness wassite-specific and independent of other variables.There was a weak tendency for there to be greateramounts of vegetation in the first metre aboveground on sub-plots with more exposed (north-westerly) aspects, and there was a slight tendencyfor there to be a greater number of large logs onsub-plots with lower slope angles.
A number of habitat components showed nosignificant response to frequent burning and didnot appear to exhibit patterns that could beattributed to large-scale spatial influences. Thesewere: the amount of vegetation in the first metreabove the ground (ground herbs, small and mid-sized shrubs), the spatial homogeneity of mid-

245
Bushfire and forest invertebrates
patterns to the abundance and distribution ofterrestrial invertebrates is explored and discussedin the following sections.
4.2 TERRESTRIAL INVERTEBRATECOMMUNITIES
This study revealed a rich terrestrial invertebratefauna with representatives from the Chelicerata(spiders, ticks & mites, pseudoscorpions,harvestmen), Crustacea (landhoppers, slaters),Chilopoda (centipedes), Diplopoda (millipedes),and a large number of Insect Orders & Families.
4.2.1 Invertebrate Abundance
Numerically, the most abundant groups overallwere the springtails (33.1%), ticks & mites(23.9%), ants (23.1%), bugs (4.2%), beetles(4.0%), bees & wasps (2.8%), insect larvae (2.7%),flies (2.6%) and spiders (2.2%), with these ninegroups making up 98.6% of the total number oforganisms caught. The first three groups(springtails, ticks & mites, and ants) represented80% of individuals caught.
Due to their low numbers, it was notpossible to comment on the effects of frequentburning for: pseudoscorpions, harvestmen,centipedes, millipedes, diplurans, termites,embiids, booklice, lacewings, caddisflies, mothsand butterflies. For these groups the trappingmethod used may not have been the mostappropriate and has potentially contributed to thelow capture rate. While the low numberscollected for several other taxa precludedstatistical analysis, frequent burning appears tohave led to a reduction in the numbers ofamphipods, cockroaches and earwigs, and anincrease in the numbers of grasshoppers &crickets, and thrips. Terrestrial amphipods live inand feed on decaying litter of the forest floor,requiring a relatively moist environment becausethey are susceptible to desiccation (Friend andRichardson 1986). Densities of some species havebeen correlated with litter thickness (eg., Duncan1969) suggesting that the reduction in leaf litterassociated with hazard-reduction burning is likelyto result in lower population sizes for this group.Similarly, cockroaches feed mainly on the detritusassociated with leaf litter while the majority ofearwigs feed on live or decaying plant matter(Zborowski and Storey 1995). Both groups wouldbe impacted upon by a reduction in the amount ofthis resource. Surface active groups such asgrasshoppers and crickets tend to prefer more
open habitats (CSIRO 1991) such as those typicalof frequently burnt areas, although Dolva (1993)found that wood crickets (Gryllidae) were moreabundant in unburnt areas, probably in responseto the structure of the litter layer. Thrips aregenerally associated with vegetation and may beresponding to the marked changes in the structureand cover of the understorey.
For the ten broad taxonomic groups wherethere were sufficient data to permit statisticaltesting, the results indicated a variety of responsesto frequent burning. Seven groups (isopods,springtails, ticks & mites, bees & wasps, insectlarvae, flies and beetles) showed substantialdecreases in abundance following frequentburning. These decreases ranged from 15 to 58%(see Section 3.3.2), but were only statisticallysignificant for ticks & mites (31%), insect larvae(35%), flies (58%) and beetles (31%). Many ofthese groups are associated with leaf litter and it islikely that their numbers have been influenced bythe episodic removal of this resource, and the factthat litter levels on frequently burnt plots were, onaverage, 50% of that on unburnt plots. Newmanand Tolhurst (1991) considered that reductions inabundance of Collembola and Diptera following asingle fire event were in response to reduced litter(fuel) levels. Terrestrial mites are exceedinglycommon in soil and leaf litter and are generallypredatory, feeding on small invertebrates (Harveyand Yen 1989). Collembola communities havebeen shown to be initially particularly sensitive tofire (Campbell 1973), and an increase in firefrequency may reduce population sizes and altercommunity structure (Metz and Dindal 1975;Dindal and Metz 1977). It was interesting to notethat the large-scale patterns in abundanceexhibited by ticks & mites and the beetlesparalleled patterns in top-soil moisture content(see Figures 3.4, 3.8 and 3.14). The observedsignificant reduction in abundance of these groups(both down by 31%) may be in response to the18% decrease (on average) in top-soil moisturelevels apparent on sites experiencing frequent fire.High spatial variability in abundance for isopods,springtails, and bees & wasps possibly contributedto the lack of statistical significance in this study.
Three groups showed substantial increasesin abundance following frequent burning. Thesewere statistically significant for bugs (77%) andants (250%), but not for spiders (33%). This maybe due to increased ease of movement (increased“trapability” - see Majer 1980, Andersen 1988) for

some lacewings feeding on honeydew or pollen,while others prey on aphids and scale-insects(Zborowski and Storey 1995). This group may bemore abundant in the more structurally complexvegetation of unburnt sites, however a samplingregime concentrating on understorey vegetationwould be required to more accurately assess this.
In order to test the utility of RBAmethodology, and further investigate the impactof repeated burning on species richness and therelated aspects of community composition andstructure, five taxa were investigated in detail byanalysis to morphospecies level. These groupsutilise a diversity of micro-habitats and niches andare representative of the range of terrestrialinvertebrates found in these forest environments.These were: firstly, the Hemiptera (bugs), amostly terrestrial and phytophagous (plant-feeding) group which have a close association withplant communities. Secondly, the Diptera (flies),which although highly mobile as adults, haveparticular requirements with regard to larval foodsources; usually moist, decaying plant and animalmaterial. Many species are parasitic on the larvaeof other insect orders with specialist habitatrequirements for oviposition. Thirdly, theAraneae (spiders), a major group of predators inforest ecosystems exploiting a variety of habitats.They live in burrows or crevices in the ground,amongst leaf litter or in vegetation, and are agroup with many habitat specialists. Fourthly, theColeoptera (beetles), which utilise a diverse rangeof habitats & micro-habitats, with a variety offeeding strategies (adults include herbivores,predators & scavengers, while larval forms feedeither internally or externally on plants and fungalproducts). Beetles are a rich and diverse groupwhich are active in the litter layer. Lastly, theFormicidae (ants), which are one of the mostnumerous and widespread groups in Australianecosystems. They have a diverse diet, and utilise avariety of feeding strategies from predators andscavengers, to plant eaters and fungus feeders,with frequent and varied interactions with otherinvertebrate groups. Ants nest in the soil and litterand therefore are response to disturbance of thesehabitats, and they are functionally importantwithin the forest ecosystem.
Overall, 411 morphospecies were identifiedfrom the five groups studied in detail. The beetleswere the most species rich (139 morphospecies),followed by the ants (88), flies (77), spiders (63),and bugs (44). The results of analyses (ANOVA)
246
Australia’s Biodiveristy - Responses to Fire
surface active groups such as spiders and ants, aswell as changes in habitat suitability. The relativeabundance of many groups was influenced notonly by fire history, but also by site-specifichabitat conditions (see below), with both spidersand ants showing considerable spatial variability intheir numbers. This suggested that large samplesizes would be required to detect managementeffects when using coarse-scale taxonomicclassification (eg. Family or Order). While theuncertain taxonomy of many groups precludesfiner scale resolution, recent developments inRapid Biodiversity Assessment (RBA) may permitsubsets of the fauna to be investigated more fully(Oliver and Beattie 1993; Beattie and Oliver1994). This would allow the use of smaller samplesizes and produce a more cost-effective outcome(see below).
4.2.2 Invertebrate Species Richness
Diversity (richness) at the Ordinal level variedfrom 11–17 broad taxa on individual study plots,with frequent burning significantly reducingdiversity at this scale. While on average thisdecrease was slight (≈ 1 Order per sub-plot), 4taxa were missing overall from frequently burntplots. These were the Opilionida (harvestmen),Embioptera (embiids), Psocoptera (booklice) andNeuroptera (lacewings). The Opilionida(harvestmen) are small to medium Arachnids(<10mm body length) which are usually found inmoist leaf litter, or under rocks, logs and bark.Most feed on smaller invertebrates but some alsoconsume plant material (Harvey and Yen 1989).Given their habitat requirements, it is expectedthat they would be disadvantaged by the drierconditions found on frequently burnt areas.Embiids (web-spinners) are small to medium(4–15mm body length) insects usually living underrocks, bark or leaf litter. They feed on leaves,bark, mosses and lichens (Zborowski and Storey1995). The Psocoptera (booklice, psocids) areminute to small (<1–10mm body length) insectswhich live on vegetation, or under bark or stones.They feed on minute organic items such as plantspores, algae, lichen and fungi. For both groups,these food resources are likely to be moreabundant in the moister litter and soil conditionsprevalent in infrequently burnt forest. TheNeuroptera (lacewings) are small to large(wingspan 5–150mm) insects with generallyactive, long-legged predacious larvae (antlions).Adults may be predacious or omnivorous, with

247
Bushfire and forest invertebrates
investigating the effects of frequent burning andpatterns due to large-scale spatial effects indicateda variety of responses to frequent burning. Twogroups, flies and beetles, experienced a significantreduction in species richness on sub-plotsfollowing frequent burning (44% and 27%reduction respectively). A further two groups, thebugs and the spiders, showed an increase inspecies richness on sub-plots (16% and 27%respectively), although these results were notstatistically significant. The ants experienced asignificant increase in sub-plot richness (26%)following repeated burning. Few studies havedealt with the impact of fire on invertebratespecies richness. Leonard (1972) found that thespecies richness of leaf litter fauna may drop by50% immediately after fire. Recovery after firemay be rapid (Leonard 1972), or take several years(Moulton 1982), depending upon the season ofburn and the meteorological conditions followingthe fire and the influence these factors have onlitter accumulation, and the mobility andrecolonising ability of particular species (Morris1975). Long-term studies of spiders (Huhta 1971;Merrett 1976) and ants (Brian et al. 1976; York1994) have shown a replacement of species in theyears after fire which is related to their particularhabitat requirements being met as the vegetationstructure changes over time. Species richness maystay largely unchanged (Merrett 1976) or decline(York 1994).
At the scale at which richness (α-diversity)was estimated in this study there were large-scalespatial patterns exhibited by some groups. Forbugs, spiders, beetles and ants these were notstatistically significant overall, but neverthelesshave implications for future projects attempting tomeasure disturbance impacts. For ants, observedspatial variation in richness appeared to reflectrandom variation expected within a samplingprogram such as this. For other groups observedvariation was distinctly non-random and appearedto reflect underlying environmental patterns. Forbugs similar trends in richness were apparent atpaired sites along the road transect suggesting thata smaller number of replicates would have beensufficient to detect impact (or lack of impact). Forflies, spiders and beetles this was not the case.Exhibited spatial patterns were often quitedifferent between the two treatments for a singletaxa (eg. Flies - Section 3.3.3.2), reinforcing theneed for sufficient experimental replication inorder to detect real differences. A most important
observation is the lack of concordance in spatialpatterns of richness between taxonomic groups.This suggests that groups are respondingdifferently to environmental factors and to theagent of disturbance (frequent fire). This hasimplications not only for invertebrate samplingstrategies, but also for the use of a single, orlimited group of taxa as a surrogate in biodiversityassessment. This supports the findings of Oliver(1995) who found that different taxa responded tothe disturbance of forest logging in distinct ways,and that it was not appropriate to use any onetaxon as a surrogate for the richness of any othersin conservation evaluation, environmentalmonitoring or impact assessment.
It was apparent that estimates of speciesrichness were also influenced by the spatial scaleof measurement, with associated implications forthe interpretation of observed treatment effectsfor the different taxa. For bugs and flies resultswere consistent across a range of scales ofmeasurement, with the magnitude and directionof differences between unburnt and burnt resultsfor sub-plot, plot and treatment similar. Forspiders, while the magnitude and direction ofspecies richness at the scale of sub-plot and plotwere similar, considerably more species werefound overall on burnt compared to unburntplots. This suggested that species assemblages onburnt plots were more diverse than those onunburnt plots, resulting in higher β- (between-habitat) diversity. Diversity on sub-plots withinboth unburnt and burnt plots would appear to besimilar, suggesting the differences lie at, or above,the scale of plot ( 1 hectare). For beetles, themagnitude of the difference detected betweenunburnt and burnt at the scale of sub-plot andplot were similar, however the direction wasreversed at the scale of treatment. This wouldsuggest a similar situation as to that with thespiders, where the species assemblages on burntplots are more diverse than those on unburntplots. Diversity on sub-plots within both unburntand burnt plots would appear to be similar,suggesting the differences lie at, or above, thescale of plot ( 1 hectare). For ants, the magnitudeof the difference in species richness detectedbetween unburnt and burnt areas at the scale ofsub-plot and plot were similar, although themagnitude of the difference was reduced at thescale of plot (compared with other taxa). Thedirection of the difference was howeversubstantially reversed at the scale of treatment,

proportions of morphospecies in each categoryhowever varied substantially between taxonomicgroups. For Hemiptera (bugs) the proportionswere 16, 41 and 43% for both, unburnt and burntrespectively; for Diptera (flies) 45, 40 and 15%;for spiders 25, 24 and 51%; for beetles 28, 34 and38%; and for ants 57, 23 and 20%. The overallbiodiversity of frequently burnt areas wasmaintained by the addition of species notrecorded on unburnt plots. The changedenvironment was supporting an additional 133morphospecies (19 bug, 11 fly, 32 spider, 53 beetleand 18 ant species). It is notable that a largeproportion of species (16-47%) are apparentlyindifferent to disturbance history and habitatstructure within the limits sampled in this survey,although changes to relative abundance need to betaken into consideration.
Although an examination of relativeabundance patterns enables broad “assemblages”of species with similar responses to disturbance tobe identified, these different patterns were moreclearly apparent from a comparison of bi-plotsderived from the CCA ordination procedure. Forbugs, spiders, beetles and ants there was little orno overlap of unburnt and burnt sub-plots inordination space, indicating low similarity of thespecies assemblages of the two treatments. Forflies however there was a substantial overlap,reflecting the relatively large number ofmorphospecies shared by the two treatments.Morphospecies found in both burnt and unburntareas can be regarded as habitat “generalists”,largely resilient to frequent disturbance andtherefore of lesser concern with regard tobiodiversity conservation. Of greater importanceare those species absent from frequently burntsites (potential habitat “specialists”). The resultsfrom this study suggest that frequent burning hadled to the loss of up to 131 species (18 bugs, 31flies, 15 spiders, 47 beetles and 20 ants), whichrepresents 47% of the morphospecies known fromthe unburnt areas. Many of the morphospeciesapparently lost from frequently burnt sites werehowever only detected on a single sub-plot orrepresented by a single individual on unburntplots. These could be genuinely rare oruncommon species which were missed purely bychance when sampling burnt plots. For this reasonit is difficult to identify clear patterns (at thespecies level) from the relative abundance dataalone. Some general trends were apparenthowever when morphospecies data were arranged
248
Australia’s Biodiveristy - Responses to Fire
suggesting a different situation to that with thespiders and beetles, with the species assemblageson burnt plots less diverse. Diversity on sub-plotswithin both unburnt and burnt plots would appearto be less similar than with other taxa, suggestingthe differences lie at less than the scale of plot ( 1hectare).
These spatial patterns in estimates of speciesrichness are a consequence of the composition ofinvertebrate assemblages (communities), and theirresponse to habitat conditions, at the varyingscales of investigation. The nature of thesepatterns, and their interaction with environmentalvariables, will be further explored in the followingsection.
4.2.3 Community Composition
The five groups studies in detail proved to beextremely diverse. Beetles had the richness faunaoverall with 139 beetle morphospeciesrepresentative of nine super-families and 25families. The ants were the second richness groupwith 88 morphospecies representative of 5 sub-families and 34 genera. They were followed by theflies with 77 morphospecies representative of 2sub-orders and 20 families, the spiders with 63morphospecies representative of 21 families, andthe bugs with 44 morphospecies representative of16 family (or similar) groups.
Overall, the same number of morphospecies(279) were collected from unburnt and burntplots, with average (mean) richness values similaron both treatments (48.2 and 46.5 morphospeciesrespectively). This initially suggests that frequentburning had not reduced biodiversity in this forestenvironment. An analysis of the richness (α- andβ-diversity) of individual taxonomic groups hasshown this not to be the case, with groupsresponding differently to frequent burning. Thenature and, potentially, the mechanisms behindthis difference can only be elucidated by anexamination of the species composition of faunalassemblages (communities) and would not beapparent from an examination of data at a highertaxonomic level (for example: Neumann andTolhurst 1991; Neumann 1992; Coy 1996).
An inspection of the distribution ofmorphospecies across sub-plots for each treatmentdetected a consistent pattern, irrespective offaunal group. Morphospecies fell into one of threegroups: found on both treatments (Group A),found only on unburnt plots (Group B), or foundonly on burnt plots (Group C). The relative

249
Bushfire and forest invertebrates
by genus (ants) or family (bugs, flies, spiders andbeetles) utilising general information available ontheir biology and ecology at these taxonomiclevels (“guild” / “functional group” approach).
For bugs, unburnt plots had more speciesfrom the infra-order Dipsocoromorpha (7 vs 3),while burnt plots have greater numbers of speciesfrom the family Reduviidae (5 vs 0). TheDipsocoromorpha include species (described &undescribed) primarily known from leaf litter andother moist environments (CSIRO 1991). Theobserved substantially lower (50%) amounts ofleaf litter and changes to its spatial distributionfollowing frequent burning would appear to offeran explanation to the reduction inDipsocoromorpha species. Results of the CCAordination support these conclusions withunburnt sub-plots characterised by high levels oflitter, high cover of tall and very tall shrubs, hightop-soil moisture levels and low and spatiallyvariable amounts of insolation at ground level.Burnt sub-plots were characterised by high levelsof insolation at ground level, greater top-soilhardness and greater cover of the herb & shrubcomponent of the understorey vegetation. TheReduviidae (Assassin Bugs) are predacious onother invertebrates and found generally onvegetation and on the ground. There is no clearexplanation as to their absence from unburntforest and they may be useful disturbance“indicators” (see 4.2.5).
For flies, unburnt plots had more speciesfrom the families Sciaridae (8 vs 5), Phoridae (16vs 13) and Empididae (6 vs 3). The Sciaridae(black fungus gnats) are often associated withdecaying material, with their larvae are oftenfound in rotting vegetable matter or highlyorganic soils (CSIRO 1991). The Phoridae(humpbacked flies) are active scavengers onfoliage and litter, with the larvae generallyscavengers in carrion and other decomposingmatter. Adults generally oviposit in carrion andorganic material on the ground (D.Bickel pers.com.) All the morphospecies of Empididae foundin this study were from the sub-familyTachydromiinae, which are mainly terrestrial andrarely fly. As adults they are generally predaciouson smaller arthropods and frequent moist places,commonly amongst vegetation. The larvae areprobably predacious, living in the soil and withindecaying vegetation and the leaf litter (CSIRO1991). The observed substantially lower amountsof leaf litter and changes to its spatial distribution
following frequent burning, and lower top-soilmoisture levels, would appear to offer anexplanation to the reduction in the number ofspecies from the families Sciaridae, Phoridae andEmpididae. Results of the CCA ordinationsupport these conclusions with unburnt sub-plotscharacterised by high levels of litter, high cover oftall and very tall shrubs, high top-soil moisturelevels and low and spatially variable amounts ofinsolation at ground level. Burnt sub-plots werecharacterised by high levels of insolation atground level, greater exposure (more north-westerly aspects), and to a lesser extent, steeperslopes. These drier, more exposed conditionswould be less favourable for species from these flyfamilies. This supports the findings of Delettre(1994) who, in a study of a heathland chironomid(midge) community, found that speciescomposition was best explained by fire-relatedchanges to the vegetation structure and soilmoisture levels.
For spiders, unburnt plots had more speciesfrom the family Malkaridae (2 vs 0), while burntplots had greater numbers of species from thefamilies Zodariidae (9 vs 2), Gnaphosidae (6 vs 3),Corinnidae (7 vs 4), Linyphiidae (4 vs 2) andLycosidae (3 vs 0). The Malkaridae are moisthabitat specialists, commonly dwelling in the leaflitter (M.Gray pers. com.). The Lycosidae (wolfspiders) and Zorariidae are small to large, groundliving, hunting spiders. Species from theZorariidae are vagrant hunters, frequenting areaswith an open vegetation structure and low litterlevels. The Gnaphosidae and Corinnidae are bothgeneralised hunters, tolerant of drier conditions.The Linyphiidae (tent spiders) are goodcolonisers of disturbed habitats and can “balloon-in” from shrubs some 50m distant (M.Gray pers.com.). The observed substantially lower amountsof leaf litter and changes to its spatial distributionfollowing frequent burning, and lower top-soilmoisture levels, would appear to offer anexplanation to the reduction in the number ofspecies from the family Malkaridae, and theincrease in species from the families Zodariidae,Gnaphosidae, Corinnidae, Linyphiidae andLycosidae. Results of the CCA ordination supportthese conclusions with unburnt sub-plotscharacterised by high levels of litter, high cover oftall and very tall shrubs, high top-soil moisturelevels and low and spatially variable amounts ofinsolation at ground level. Similarly, burnt sub-plots were characterised by high levels of

following frequent burning, and lower top-soilmoisture levels, would appear to offer anexplanation to the reduction in the number ofspecies from the genera Cerapachys andHypoponera. Similarly, the structurally simplifiedenvironment found on frequently burnt areaswould provide suitable habitats for large, solitaryforagers such as Colobostruma spp. Results of theCCA ordination support these conclusions withunburnt sub-plots characterised by high levels oflitter, high cover of tall and very tall shrubs, hightop-soil moisture levels and low and spatiallyvariable amounts of insolation at ground level.Similarly, burnt sub-plots were characterised byhigh levels of insolation at ground level andgreater cover of the herb & shrub component ofthe understorey vegetation.
Across the five groups studied in detail; bugs,flies, spiders beetles and ants, there was aconsistent pattern with regard to the changes thatoccur to community composition followingfrequent burning. Although species richness (α-diversity) decreased by 44% and 27% for flies andbeetles, and increased by 16%, 27% and 26% forbugs, spiders and ants (respectively), all groupsexperienced a loss of species with frequentburning. The results from this study suggest thatfrequent burning had led to the loss of up to 131species (18 bugs, 31 flies, 15 spiders, 47 beetlesand 20 ants), which represents 47% of themorphospecies known from the unburnt areas.The losses were disproportionate across thegroups with percentage reductions ranging from41% (bugs), to 40% (flies), 34% (beetles), 24%(spiders) and 23% (ants). The species lost wouldappear to be from those groups dependent upon asubstantial litter layer and stable moist conditions.The overall biodiversity of frequently burnt areaswas maintained by the addition of species notrecorded on unburnt plots. The changedenvironment was supporting an additional 133morphospecies (19 bugs, 11 flies, 32 spiders, 53beetles and 18 ants). These species would appearto have broad tolerances, or adaptations, to drierand more open environments. Overall, thecomposition of terrestrial invertebratecommunities was therefore influenced by acombination of site-dependent (slope and aspect)and treatment-dependent (litter, insolation, herb& shrub cover, top-soil moisture & hardness)environmental variables.
This research has also shown that patterns ofrelative abundance and species richness are not
250
Australia’s Biodiveristy - Responses to Fire
insolation at ground level and greater cover of theherb & shrub component of the understoreyvegetation. As a number of Corinnidae species areant specialists, the substantial increase in antabundance on frequently burnt sites may have alsocontributed to the increased richness of thisfamily through increased prey availability.
For beetles, unburnt plots had more speciesfrom the family Carabidae (11 vs 8), while burntplots had greater numbers of species from thefamilies Curculionidae (18 vs 9) andChrysomelidae (8 vs 4). The Carabidae (GroundBeetles) are mainly predatory, both as adults andlarvae, on plant-inhabiting insects. TheCurculionidae (Weevils), as adults, feed on thestems, roots, seeds and fruits of plants, with larvaeusually feeding on wood and other plant parts.The Chrysomelidae (Leaf Beetles) feed on leavesand other (living) vegetative parts of plants, bothas larvae and adults. The richness of the beetlefauna here would appear to reduce the ability togeneralise at the level of family. Results of theCCA ordination suggest a reasonable separationof assemblages on burnt and unburnt plots, withunburnt sub-plots characterised by high levels oflitter, high cover of tall and very tall shrubs, hightop-soil moisture levels and low and spatiallyvariable amounts of insolation at ground level.Similarly, burnt sub-plots were characterised byhigh levels of insolation at ground level. It isreasonable to suggest that ground-dwellingspecies from the Carabidae are influenced bysubstantially lower amounts of leaf litter andchanges to its spatial distribution followingfrequent burning. Changes to the structure of thevegetation community with frequent burningappear to have provided additional habitats forplant-dependent species from the familiesCurculionidae and Chrysomelidae.
For ants, unburnt plots had more speciesfrom the genera Cerapachys (7 vs 1) and Hypoponera(4 vs 1), while burnt plots had greater numbers ofspecies from the genus Colobostruma (2 vs 0).Species of Cerapachys are considered by Andersen(1990) to be “climate specialists” and are known tobe specialist predators, often on other ants(Holldobler and Wilson 1990). Species from thegenus Hypoponera are cryptic, nesting and foragingwithin the soil and leaf litter. Members of thegenus Colobostruma are large solitaryforagers/specialist predators (Andersen 1990).The observed substantially lower amounts of leaflitter and changes to its spatial distribution

251
Bushfire and forest invertebrates
concordant between broad taxa. Analysis ofcommunity composition, as illustrated by theCCA ordination, also illustrates that patterns ofinvertebrate community organisation show variedresponses to environmental disturbance. In the bi-plots derived from the CCA ordination, thedegree of clustering of the sub-plots from eachtreatment indicates the relative similarity ofspecies assemblages on sub-plots and plots withineach treatment. The tighter clustering of burntsub-plots for bugs, beetles and ants indicated alower within-treatment diversity compared tounburnt sub-plots (ie. a lower β-diversity). Theconverse applied for spiders, with the tighterclustering of unburnt sub-plots indicating a lowerβ-diversity compared to burnt sub-plots. Thesituation for flies indicated similar within-treatment diversity for both treatments, with looseclustering of both unburnt and burnt sub-plots.This interaction between point richness (α-diversity) and spatial “turnover” of species (β-diversity) has substantial implications for theinterpretation of the apparent effect of repeatedburning (see “scale effects” in Section 3.3.3.6).Similar patterns were apparent for bugs and antswhere richness values (α-diversity) on burnt sub-plots were on average higher than on unburntsub-plots, however the high similarity ofassemblages on burnt sub-plots (low β-diversity)meant that the overall richness of both treatmentswere similar. Unburnt sub-plots had lowerrichness (α-diversity) but are less similar, resultingin higher “turnover” between sub-plots (higher β-diversity), increasing overall species richness forthat treatment. For flies and beetles burnt plotshad lower richness (α-diversity) but were lesssimilar, resulting in higher β-diversity. Spidersexhibited a different pattern with both higher α-and β-diversity for the burnt treatment, resultingin a large number of species (32) unique tofrequently burnt sites.
An explanation for the different levels ofspecies turnover (β-diversity) for the differentgroups is likely to be found in patterns ofenvironmental heterogeneity and its effect onmicro-habitat diversity. Frequently burnt areaswere shown to have more spatially homogeneousground herb, small shrub, tall and very tall shrublayers, more spatially heterogeneous levels ofinsolation at ground level, and more spatiallyheterogeneous bark and twig components of thelitter layer. The β-diversity of bug, beetle and antcommunities was reduced with frequent burning,
increased for spiders, but appeared unchanged forflies. For bugs, beetles and ants it would appearthat the degree of heterogeneity of the vegetationnear the ground plays a role in maintainingbiodiversity, probably by providing additionalhabitats and an associated increase in food andother resources. Structural heterogeneity may alsoimply a greater diversity of plant species, or ofgrowth stages of existing species. Increased levelsof shading and changes in surface and nesttemperature with increased vegetation cover hasbeen shown to reduce ant species richness(Goldstein 1975, Greenslade and Mott 1979), andalter community composition as theenvironmental conditions become sub-optimal forcertain species (Welch 1978, Elmes and Wardlaw1982, York 1994). Moisture, light and temperaturehave been identified as important factors in thedetermination of the composition of invertebrateassemblages (Huhta et al. 1967, Punttila et al.1991, McIver et al. 1992). Frequent burning led togreater spatial heterogeneity in the bark and twigcomponents of the litter layer, with a likely impacton its structural complexity. The composition ofspider communities has previously been shown tobe influenced by the structure of the litter layerfollowing fire (Huhta 1971). The implication ofthese changes for community organisation andecosystem function are considered in thefollowing section.
4.2.4 Community Structure
The biological structure of a community involvesspecies composition and abundance, temporalchanges in communities, and the relationshipsbetween species in communities. This in turnexerts strong influences on the functioning of thecommunity, in other words, how the communityworks as a processor of energy and nutrients(Krebs 1985). The continued functioning ofcommunities and their ecological processes is aprimary goal of ecologically sustainablemanagement (ESDWG-Forest Use 1991).
While it is possible to describe and assesscommunities using indices such as species richness,or to compare the relative abundance of speciesusing similarity indices, multi-variate approachesand/or through graphical representation, theseprovide little information concerning the processesunderlying these differences or any indication as tothe relative sensitivity of species to landscapechange (Samways 1994). In order to simplify andinterpret the complexity of ecological systems, one

dung. Among the beetles, the Leiodidae areabundant in decaying organic matter and alsooccur in carrion and fungal fruiting bodies. Manyare general scavengers, but certain groups areassociated with particular fungi (Lawrence andBritton 1994). The Ptiliidae are minute beetleswhich are relatively abundant in decaying organicmatter, including leaf litter, where their majorfood source appears to be fungal spores andhyphae. The Endomychidae feed on a variety offungi, with many occurring in leaf litter in moisthabitats.
For flies, the number of generalists andscavengers decreased slightly (11%), while forbeetles the proportion of generalists increased, onaverage, by 79%, primarily through an increase inmorphospecies from the family Scarabaeidae.Scarab beetles always live in concealed habitats,feeding on roots, dung or decaying vegetablematter (McQuillan 1985). The reason for theirdramatic increase here is not apparent, althoughwith their generally large size, their mobility maybe enhanced in the more open environment offrequently burnt areas. With ants, the proportionof generalist morphospecies decreased, onaverage, by 11% following frequent burning.Results were not consistent within the group withthe genera Pheidole and Crematogaster decreasingby 35 and 19% respectively, and the genusMonomorium increasing by 388%. In these dryforests, species from the genus Monomorium aremajor seed predators (Andersen 1985, Andersenand Ashton 1985), and may be responding tochanges in the vegetation composition thataccompany a frequent fire regime (Zedler et al.1983, Nieuwenhuis 1987, Cary and Morrison1995). For flies, the number of morphospeciesregarded as wide-ranging “tourists” was, onaverage, reduced by 43% on frequently burntplots.
When groups were compared with regard tohabitat preferences, it was apparent that therewere substantial changes in community structurefor some taxa. For bugs, numbers of moist habitatspecialists from the sub-order Dipsocoromorphawere, on average, reduced by 83%. Amongstspiders, moist habitat specialists were reduced by88%, primarily due to a 90-95% decrease innumbers of morphospecies from the familiesTheridiidae, Toxopidae and Oonopidae. TheTheridiidae build their webs in leaf litter, theToxopidae are a moist-adapted group, and theOonopidae mostly inhabit the litter in moist
252
Australia’s Biodiveristy - Responses to Fire
approach has been to group species into “guilds”or “functional groups”. These groups recognisethe ecological rather than the taxonomic affinity ofspecies. To test the applicability of this approach,morphospecies (see Oliver and Beattie 1993) wereallocated to guilds based upon their feedingstrategies and habitat preferences, followingreference to the relevant literature and discussionswith taxonomic experts. By classifying largenumbers of species into smaller, more manageablegroups, it is possible to substantially reduce theapparent complexity of ecological systems andprovide a basis for evaluating environmentalchange (Andersen 1990).
For all groups studied, frequent burningresulted in a change in the structure of thecommunity. With regard to feeding strategy, therewas on average, a 15, 140 and 250% increase(respectively) in the number of phytophagous(plant feeding) species of bugs, flies and beetles.With regard to the proportion of predator species,it remained unchanged for flies and bugs, howeverfor bugs there was a total shift from the familyNabidae to the family Reduviidae. The Nabidaeare a family of predacious bugs whose eggs areoviposited into grass stems (CSIRO 1991). Thissuggests that a change in this component of thevegetation may be influencing the suitability ofthe habitat for these species, which are thenreplaced by another group of predators from adifferent family. With beetles there was, onaverage, an 18% decrease in the number ofpredator species, due primarily to a 54%reduction in the number of morphospecies fromthe family Staphylinidae. Most species from thisfamily are small, often cryptic and live aspredators hidden in soil and leaf litter. Others areassociated with dung, carrion or fungi (Zborowskiand Storey 1995). It is anticipated that thereduction in litter levels and decrease in soilmoisture associated with frequent burning woulddisadvantage this group.
For groups feeding primarily on fungalproducts there was, on average, a 44% (flies) and31% (beetles) decrease in the number ofmorphospecies. For flies this was most marked inthe families Scaridae and Scatopsidae, and forbeetles, in the families Leiodidae, Ptiliidae &Endomychidae. Species from the family Scaridaeoviposit in fungi in the soil and also feed on fungalproducts, while the Scatopsidae are generallyfound in moist forest environments with theirlarvae occurring in rotting vegetable matter and

253
Bushfire and forest invertebrates
environments. With the flies, the number of moisthabitat specialists remained similar, although thefamily Ceratopogonidae was more commonlyrepresented on burnt sub-plots and the familyChironomidae on unburnt sub-plots. Theproportion of temperature-dependent (climate)specialists among the ants remained largely stable(increased by 4%), although individual genera didvary in their response. For spiders, there was, onaverage, and an 35% increase in the number ofspecies with a known preference for dry habitats.
For groups primarily inhabiting the litterlayer there was a variety of responses to frequentburning. For flies, the number of morphospeciesspecifically utilising the litter layer had, onaverage, decreased by 60%. This was primarilydue to the absence of the family Tipulidae onfrequently burnt sub-plots. Tipulidae (crane flies)use moist soil for breeding, with their larvaecommon in decaying vegetation, and thereforewould be disadvantaged by frequent burning.With ants there was a 19% decrease in theaverage number of “cryptic” morphospeciesinhabiting the litter and soil, particularly withregard to the genera Hypoponera (90%) andTapinoma (86%). Species from the genusHypoponera are specialist predators, feeding largelyon Collembola which were approximately 15%less abundant on frequently burnt sites. Tapinomaspecies are cryptic omnivores in the litter, withsome arboreal nesters. Habitat availability for thisgenus would be reduced by frequent burning. Forspiders however, the number of litter dwellingspecies increased, on average, by 110% with anequivalent increase from the families Hahniidaeand Textricellidae. Species from the familyHahniidae are small spiders that construct smallsheet webs in litter and foliage. It would appearthat they are not disadvantaged by lower litterlevels (M.Gray pers. com.). Spiders in the familyTextricellidae are also very small and live deep inthe litter layer near the litter/soil interface. Asonly 46-73% of litter is removed in each fire eventin these forests (York 1996), this group may not bedisadvantaged by frequent fire.
Many groups of terrestrial invertebrates areadapted to exploit disturbed habitats. For spidermorphospecies known to prefer open anddisturbed habitats, the number of morphospeciesincreased, on average, by over 600% followingfrequent burning, This was due primarily to theoccurrence of seven species from the familyZodariidae only on burnt sub-plots. For the ants
there was a substantial increase overall (30%) inthe average proportion of “opportunist”morphospecies, due primarily to a single species:Rhytidoponera metallica, which was extremelynumerous on burnt sub-plots. R. metallica is a wellknown coloniser of disturbed habitats (Yeatmanand Greenslade 1980).
It has been observed that the structure of antcommunities, in particular, may be influenced bythe relative abundance of particular “dominant”and “sub-dominant’ groups (Fox & Fox 1982,Andersen 1990). Following frequent fire, thenumber of morphospecies within the “dominant”functional group had, on average, decreased by23%, primarily through a reduction in theoccurrence of Iridomyrmex species. The number of“sub-dominants” had increased slightly (25%),largely because of an increase in the occurrence ofCamponotus species. The number of larger“solitary/specialist” morphospecies, which interactonly slightly with other groups, had increasedslightly (13%), although the results were quitevariable within the group. Three new genera werefound on burnt plots (Colobostruma, Epopostruma &Leptogenys) while the numbers of species ofMyrmecia increased, on average, by 87%.Myrmecia have been shown to be more commonin recently burnt habitats in these forests (York1994, 1996). The occurrence of a solitary forager,Bothroponera sp.A, had decreased substantially(76%). Bothroponera are predacious, often ontermites (Holldobler and Wilson 1990), a groupof organisms not found on frequently burnt sitesin this study. While a number of termite moundswere observed on these areas, they did not appearto be active.
4.2.5 Biodiversity Indicators
Programs in land appraisal and applied resourcemanagement increasingly utilise "environmentalindicators" to facilitate and simplify assessmentand decision-making procedures. Indicators maytake the form of an index which conciselysummarises some property of the system, such asabundance or species richness (diversity); ordescribes the community via its speciescomposition, the relative abundance of individualswithin constituent species (evenness etc), or itsorganisation or “structure” (eg. number of guildsor functional groups). An alternative (orcomplimentary) approach may be to use“indicator taxa”: an organism (or group oforganisms) that reveals important aspects of the

assessment of the impact of fire (see Campbell andTanton 1981; Majer 1984; Friend 1996). Temporalvariability following single fires has been shown tobe substantial, with taxa responding to seasonal andmeteorological cues (see Neumann 1992; Coy1996). Where sampling is required for comparativepurposes only, for example burnt/unburnt,logged/unlogged, rehabilitated/not rehabilitatedcontrasts, then the influence of temporal variabilitycan be reduced (controlled for) by simultaneoussampling in the various treatment categories underexamination (see Yeatman and Greenslade 1980;Majer et al. 1984; Burbidge et al. 1992). York(1994) showed that estimates of ant species richnessfrom a single summer sample of chronosequencesites provided an accurate representation of long-term changes over time at a single site. Oliver andBeattie (1996b) demonstrated that the richness ofants recorded from a single summer pitfall samplewas significantly correlated with richness valuesassessed by other sampling methods and seasons ofsampling. This research therefore utilised a singlesummer sample to assess the implications of spatialvariation in α- and β-diversity for investigationsinto frequent disturbance.
In this study, estimates of abundance wereshown to be influenced, not only by fire(treatment), but by large-scale spatial effects(position), with frequent interactions betweenthese two factors. This meant that the particularfire effect (or lack thereof) was not alwaysconsistent across the spatial range of sites(replicates). These interactions were significant (ie.not as a consequence of random variation) forspiders, isopods (slaters), flies, beetles and ants. Attimes spatial patterns were evident, potentially inresponse to underlying environmental trends. Thiswas the case for ticks & mites and beetles.Similarly, estimates of richness (α-diversity) wereshown to be influenced by large-scale spatialeffects (position) for flies and spiders, andinteractions between treatment and position effects(spiders, beetles and ants). These patterns,combined with the often considerable spatialvariability exhibited within a single treatment,suggests that large sample sizes would be requiredto detect disturbance effects when using coarse-scale taxonomic classification (eg Family orOrder). Neumann (1992) recognised that whilesuch a broad taxonomic approach gives acumulative estimate of the responses of the specieswithin each taxon, the behaviour of individualspecies remain unknown, thereby limiting the level
254
Australia’s Biodiveristy - Responses to Fire
structure and function for some part of theecosystem without exhaustive study of that part(Cornaby 1977).
A wide range of terrestrial invertebrates havebeen used as “indicators” in Australia. Examplesinclude spiders (Mawson 1986), springtails(Greenslade 1984, 1985; Greenslade andGreenslade 1987), termites (Nichols and Bunn1980; Greenslade 1985), beetles (Greenslade1985; Yen 1987) and ants (Weir 1978; Majer1980b, 1984, 1985; Whelan et al. 1980; Yeatmanand Greenslade 1980; Majer et al. 1982, 1984;Andersen and McKaige 1987). Arthropods have asignificant role in the forest community, affectingboth primary production by their grazingactivities, and the turnover of nutrients in theirrole as decomposers (Lowman 1982). Forest floorarthropods regulate microfloral decomposerpopulations by their feeding on bacteria andfungal colonies, their transport of spores, and bythe contribution of their faeces and bodies fordecomposition (Van der Drift 1958, Engelmann1961, MacFadyen 1962). Their requirements of asource of cover and food has led arthropods todevelop a sensitivity and responsiveness to systemstructure which has made them useful indicatorsof system status and condition (Mattson 1977).
This research used two complimentaryapproaches to investigate the applicability ofinvertebrate indicators for environmental impactassessment. This was undertaken firstly by lookingat spatial characteristics of commonly usedbiodiversity indices (abundance & richness) andby examining the concordance of these indicesbetween taxa. It is often postulated that one groupof invertebrates may act as an indicator or“umbrella” group for others, thereby allowinginferences to be made on the impact ofdisturbance regimes. This assertion however relieson as yet untested assumptions which flow fromparticular sampling strategies, in particular, spatialpatterns in species richness (α-diversity) andspecies turnover (β-diversity). The secondapproach involved a study of the composition andstructure of these communities in order toidentify species, groups of species, or community“descriptors” which were useful as a means ofassessing potential effects on ecosystem function.This was undertaken to determine theapplicability of these methods for assessingecological sustainability.
High levels of spatial and temporal variabilityin invertebrate populations severely complicate an

255
Bushfire and forest invertebrates
of interpretation that could be applied to theresults. From an analysis of species-level data, thisresearch has shown those limitations to besubstantial when dealing with biodiversity issues.
The concordance between the speciesrichness (α-diversity) of selected taxa wasinvestigated here using correlation analyses.While a number of statistically significantcorrelations were detected, the predictive powerof these relationships was weak. In addition it wasfound that apparently significant correlationsdetected when using the whole dataset (spiders &ants - positive, flies & ants - negative, flies &beetles - positive) did not hold when the twotreatments (burnt and unburnt) were analysedseparately. It was evident that the apparentcorrelations between these taxa were an artefact ofthe differences between average richness values onburnt and unburnt sub-plots. Therefore, none ofthese groups of terrestrial invertebrates would bea reliable “indicator” or “umbrella” group for anyother, and therefore are inappropriate forpredicting overall biodiversity at this scale. Thissupports the findings of Oliver and Beattie(1996b) who demonstrated non-concordancebetween richness estimates for ants, beetles andspiders.
Given the identified influence of sampling“scale” and spatial patterns of α- & β-diversity onestimates of species richness (see 4.2.2),concordance between biodiversity estimates forselected taxa were examined at a larger scale (1hectare). While two statistically significantrelationships were identified (ants & bugs -negative, beetles & spiders - negative), these wereonly apparent on unburnt plots. These resultssuggest that, at least for these 2 pairs of taxa, andat the 1ha sampling scale, it may be possible andreliable to use one group as an indicator of thebiodiversity of the other. The fact that disturbancechanges the nature of these relationships castsdoubt however on the value of this in studies ofimpact assessment.
It was therefore considered that a secondapproach, a study of the composition andstructure of these communities, may prove moreuseful. If species, or groups of species, could befound that typified the response of whole taxa,then these may serve as useful “bio-indicators”.An assessment of the effects of disturbance onecosystem function would then be feasible,providing a valuable tool for monitoring progresstowards, and compliance with, ecologically
sustainable management. When differences incomposition between burnt and unburnt areaswere compared, it was apparent that frequentburning had led to a marked decline in specieswith particular habitat preferences. The groupswhich were indicative of this decline were theinfra-order Dipsocoromorpha and the familiesSciaridae, Phoridae and Empidae (flies), thefamilies Malkaridae (spiders), Carabidae (beetles),and for ants, the genera Cerapachys andHypoponera. Conversely, a number of groups were“disturbance indicators” through marked increasesin their richness on frequently burnt areas. Thesewere the families Reduvidae (bugs), Zorariidae,Gnaphosidae, Corinnidae, Linyphiidae andLycosidae (spiders), Curculionidae andChrysomelidae (beetles), and for ants, the genusColobostruma. The habitat parameters mostconsistently associated with these changes werethe amount and spatial distribution of leaf litter,top-soil moisture levels, and the amount andspatial heterogeneity of insolation levels at theground surface. These environmental factors wereshown to vary primarily as a response to frequentburning, that is, under management control.
Accompanying these habitat-related changesin community composition were a number ofshifts in ecological structure and function. Forsome taxa the guild structure was generallymaintained, but with changes to the suite ofspecies comprising each guild. This was mostmarked with the ants, where the functional groupstructure remained largely unaltered, butcommunity composition changed substantially.This suggests that ant communities are resilient tofire-related disturbances (although individualspecies may be fire sensitive). A similar patternwas evident in the bugs, with a shift within thephytophagous (plant-eating) guild from theCoccidae, Fulgoridae and Homoptera to theCicadellidae; and within the predacious guildfrom Nabidae to Reduvidae. With the flieshowever there was a marked reduction (44%) infungal feeders (Sciaridae, Drosophilidae,Mycetophilidae and Scatopsidae) and a substantialincrease (140%) in phytophagous species(primarily Cecidomyiidae), resulting in aconsiderable change in community structure. Thebeetles were similarly affected with a 31%reduction in the richness of the fungal-feedingguild (primarily Leionidae), and a 250% increasein richness of phytophagous species (primarilyChrysomelidae and Curculionidae). The loss of

given that they cannot be unambiguously defined(Caughley and Gunn 1996). It is recognised that amodern conservation strategy cannot however bebased solely on areas managed purely forconservation purposes. Forests (and otherenvironments) outside the reserve system willcontinue to play an important and complementaryrole in meeting conservation objectives withrespect to biodiversity, even though these forestsmay be available for the production of timber andother commercial uses in an ecologicallysustainable way.
This research has indicated that, through thedevelopment of appropriate managementstrategies, we have the knowledge to ensure thatbiodiversity is adequately conserved. Strategiesrequired to conserve invertebrate biodiversity arefundamentally consistent with those used toprotect other groups, although the emphasis islikely to be placed on the protection of habitatsrather than individual species or assemblages (seeSamways 1994; New 1995). With regard tohazard-reduction burning, the extensive andfrequent use of this management practice has thepotential to substantially reduce regionalbiodiversity. The development of strategies whichset aside (unburnt) refuges, maintain a diversity ofhabitats at various stages in the post-firesuccession, permit variability in other componentsof the fire regime (season of burn and intensity),and allow connectivity between different habitats(corridors), requires urgent attention. Currentdraft strategies in forest zoning (eg. SFNSW1995) are currently addressing some of theseissues, however it is of some concern that otherland management agencies still advocate frequent“broad-acre” burning as a panacea for hazardreduction and protection from wildfire withoutadequate consideration of important biodiversityissues.
256
Australia’s Biodiveristy - Responses to Fire
species associated with the decomposer cycleimplies the frequent burning may be impactingupon nutrient cycling and transfer within theseforests.
While grouping species into guilds orfunctional groups does achieve its intended aim ofsimplifying complex systems, it is at the expense ofconsiderable important detail. Due to the richnessof the communities involved, and the associatedtaxonomic difficulties, this research adopted a“morphospecies” approach where the biology andecology of individual species was not examined.York (1996) has shown that, for ants, a knowledgeof the ecology of the constituent species at thelevel of genus enables considerable insight into themechanisms of change following disturbance.Given the aims of this study, the morphospecies(RBA) approach has however proved to besuccessful. The limitations of this approach areacknowledged, and it is considered that additionalinsight could be gained by a more detailedecological investigation of individual taxa. This isproposed in a series of future studies.
The limitations of grouping species intotaxonomic or ecological units are particularlyevident in the area of conservation biology wherea “species-by-species” management approach maybe required, with the ecological needs of eachspecies addressed separately in Plans ofManagement. The incredible diversity ofterrestrial invertebrates may preclude this strategyin Australian forests, with reserve strategies beingdeveloped based primarily on vascular plants andvertebrates. Whether a conservation strategydeveloped using this approach is appropriate forthe conservation of invertebrates is questionable,given the lack of concordance between therichness and composition of these very disparategroups of organisms (see Oliver 1995). Similarly,the use of “ecosystems” or “communities” asecological units of conservation is problematic

257
Bushfire and forest invertebrates
Infrequent, periodic forest fires (bushfires) are anintegral part of the modern physical environmentof Australian sclerophyll forests. The inherentvariability in natural fire regimes generally resultsin a mosaic of habitats with vegetation at differentstages of floristic and structural post-firesuccession, each potentially supporting particularanimal communities. Changes to the componentsof the fire regime (fire intensity, frequency andseason of occurrence), as a consequence of forestmanagement practices, have the potential to alterthe composition and structure of naturalcommunities. The research reported here dealswith the impact of frequent low-intensity fire(“hazard-reduction burning”) on the abundance,richness, composition and structure of terrestrialinvertebrate communities. This group, whichconstitutes a major component of the overallbiodiversity in these forests, plays a substantialrole in the maintenance of ecosystem processes.The ability of the Australian forest industry toachieve Ecologically Sustainable Management(ESM) depends therefore on a betterunderstanding of the impact of commonly usedmanagement strategies on this importantcomponent of the ecosystem.
While a number of habitat components wereresponding to large-scale environmental patterns,frequent burning was shown to be impacting uponthe amount, structure and spatial distribution ofsurface leaf litter, the structure and spatialheterogeneity of components of the vegetationunderstorey, moisture levels in the top-soil and theamount and patchiness of insolation reaching theground. While some aspects of these changes arelikely to reflect post-fire successional trends, thetruncation of successional patterns by frequent fireand subsequent reduction in environmentalheterogeneity can be expected to impact uponterrestrial invertebrate communities.
Using ants, beetles, flies, spiders & bugs asrepresentative groups and potential indicators ofenvironmental degradation, this researchdemonstrated that although overall speciesrichness (α-diversity) may not change withfrequent disturbance, species turnover (β-diversity) does. There is a lack of concordancehowever between groups in the magnitude anddirection of these responses. All groups showed
substantial changes in the composition of speciesassemblages following frequent disturbancehowever, with a loss of taxa dependent upon asubstantial litter layer and stable moist conditions.The overall diversity of frequently burnt areas wasmaintained by the addition of species with broadtolerances, or adaptations, to drier and more openenvironments. Shifts in community compositionwere best explained by the changes in the amountof leaf litter and insolation at the ground surface,habitat elements shown to be dramaticallymodified by frequent burning. This suggests thatthe extensive application of this managementpractice could result in a reduction in terrestrialinvertebrate biodiversity at a regional scale (γ-diversity), with this decrease potentially as high as50%.
It was demonstrated that considerableadditional detail concerning, and insight into, thenature of these changes could be provided by theinclusion of fairly general information concerningthe habitat and dietary preferences of the groupsunder investigation. It was apparent that frequentburning leads to a change in the structure of theinvertebrate community. Within speciesassemblages there were shifts in feeding strategy,with substantial increases in the proportion ofphytophagous species for bugs, flies and beetles,and a reduction in fly and beetles groups reliantprimarily on fungal products. With regard topredator guilds, there was a substantial decrease inproportional representation for beetles, primarilyin relation to the family Staphylinidae. For bugshowever, the proportion of predators remainedlargely unchanged, however there was a total shiftfrom the family Nabidae to the family Reduviidae.Within one major group of predators, the spiders,there was a substantial reduction in the number ofmoist-habitat and leaf litter specialists, and adramatic increase in the number of species knownto prefer dry and open environments, particularlyfrom the family Zorariidae. Similar patterns wereexhibited by the ants, with changes in functionalgroup representation in response to habitatalteration. While the impact of these changes onecosystem function was beyond the scope of thisstudy, substantial changes in the structure ofinvertebrate assemblages and the loss of speciesassociated with the decomposer cycle implies
5. CONCLUSIONS

The application of Rapid Biodiversity Assessment(RBA) methodology demonstrated that the studyof the composition and structure of communitiesis likely to prove more rewarding in this regard.The identification of individuals to distinct“morphospecies”, while requiring additionallaboratory time and taxonomic expertise,facilitated the incorporation of broad-levelecological information into the assessment andinterpretation of environmental impact.Information currently available at the level ofFamily or Genus, but unavailable at the level ofspecies, was sufficient to enable a meaningfulinterpretation of data in relation to impacts oncommunity structure and ecological function.This in turn enabled the development ofmanagement recommendations consistent withthe conservation of biological diversity.
In studies such as this, the accurateassessment of the impact of long-term frequentdisturbance is potentially confounded by short-term responses to the most recent perturbation. Itis likely that the rate of post-fire successionalchange will be greater on the frequently disturbedplots compared to the unburnt (control) plots, andthe results obtained (and hence assessment ofimpact) will be, to some extent, dependent uponthe current successional stage (see Huhta 1971;Merrett 1976; York 1994, 1996). In this study thedisturbed ares were sampled 2 years after fire andat a time when there was sufficient fuel to supportanother low-intensity fire. With regard to habitatmodification therefore, this study represents(potentially) a “worse-case-scenario” typifying oneend of the spectrum of responses to frequent fire.Nevertheless it typifies many areas of drysclerophyll forest which are regularly burnt forthe purposes of hazard reduction. Of moreconcern is the fact that the experimental designused here substantially “down-plays” the effect ofrecolonisation ability, with potential refuge(unburnt) areas no more than 20m fromfrequently burnt sites. In a managed forestenvironment where fuel-reduction burning isoften spatially extensive, the “habitatfragmentation” effect is likely to be morepronounced. Given that many invertebrate specieswithin a community are of low abundance (seeYork 1994) or are habitat or dietary specialists(York 1996), the risk of local extinction is high.While local extinction of invertebrates is likely tobe a regular occurrence in natural systems(Samways 1994), systems prone to
258
Australia’s Biodiveristy - Responses to Fire
frequent burning may be impacting upon nutrientcycling and transfer within these forests. If this isthe case, it would have serious implications withregard to the maintenance of ecologicalsustainability.
In New South Wales, State Forests has, as astated objective of its Corporate Plan (1992), that itwill achieve ecologically sustainable management(ESM) by refining concepts and developingmeasurable indicators of ecologically sustainableuse. To be useful therefore, ESM indicators need tobe interpretable, significant, cost efficient, and needto account for variability in space and time, and beappropriate for the scale of management (Turner1993, York 1993). The research reported heresupports previous findings concerning the highspatial variability of invertebrate populationnumbers, and confirmed the limited use of dataobtained using coarse-scale taxonomic classification(eg. Family or Order). The cost-effectiveness ofusing abundance data alone was shown to be low,with high spatial variability and spatial patterningrequiring large sample sizes to detect managementeffects. This research also identified substantiallimitations with regard to the use of a single index,species richness, as an measure of change and/orenvironmental impact. Species richness (α-diversity) is frequently used to describe andcompare communities, however in this case it wasfound to provide a deceptive summary ofcommunity characteristics. The lack ofconcordance of richness and abundance patternswithin and between taxa, and the identifiedrelationships between estimates of richness (α-diversity) and turnover (β-diversity) and the spatialscale of measurement, meant that the use of theseindices (alone) for impact assessment willsubstantially restrict the level of interpretation thatcan be derived from the data. Variable spatialpatterns in these indices between disturbed andundisturbed sites, and between taxa, also limitstheir applicability in impact assessment.
It is often postulated that one group ofinvertebrates may act as an “indicator” or“umbrella” group for others, thereby allowinginferences to be made on the impact ofdisturbance regimes. In this project, it was shownthe lack of correlation between taxa with regard torichness indices (α- and β-diversity) restrictedtheir utility in this regard, primarily as aconsequence of the non-concordant spatialpatterning of these community descriptors andassociated implications for sampling effectiveness.

259
Bushfire and forest invertebrates
anthropomorphic disturbance require theestablishment of adequate measures for the in-situprotection of successional stages and theirconstituent invertebrate fauna (eg. refuges &reserves) and the establishment of links (iecorridors) to facilitate recolonisation. This need toprovide undisturbed and secure refuges for specieswith specialist requirements and limited dispersalabilities is the same dilemma facing thoseconcerned with the conservation of floweringplants and vertebrates (see Caughley and Gunn1996), the difference however is that the groupsinvolved may be responding to differentenvironmental cues and strategies developed forone taxon may not adequately conserve others (seeOliver 1995).
The development of a “Forest ZoningSystem” in New South Wales (SFNSW 1995)should be seen as a necessary response to thissituation, whereby a variety of management
strategies, from fire exclusion to frequent burning,can be applied to an area in response tomanagement and conservation needs (see Ridley1993). Given that the various successional statescan provide optimal habitats for certain species(York 1994), and that disturbance is an intrinsicand necessary feature in most natural systems(Pickett and White 1985), broad-scale fireexclusion is not a practical management andconservation option. With the increasingawareness however of the importance ofmaintaining environmental and biologicaldiversity at a range of spatial scales, thedevelopment and implementation of appropriatefire “regimes” which take into consideration thescale (ie. frequency and extent) of disturbance, iswithin the grasp of forest management agencies.


261
Bushfire and forest invertebrates
Abbott, I., Van Heurck, P. and Wong, L. (1984)Responses to long-term fire exclusion :physical, chemical and faunal features oflitter and soil in a Western Australian forest.Aust. For. 47:237-242.
Andersen, A.N. (1985) Seed predation by insects insclerophyllous vegetation at Wilson’s Promontory,Victoria, with particular reference to ants.Unpubl. PhD Thesis, School of Botany,University of Melbourne. 241 pp.
Andersen, A.N. (1988) Immediate and longer-term effects of fire on seed predation by antsin sclerophyllous vegetation in south-easternAustralia. Aust. J. Ecol. 13:285-293.
Andersen, A.N. (1990) The use of antcommunities to evaluate change inAustralian terrestrial ecosystems: a reviewand a recipe. Proc. Ecol. Soc. Aust. 16, 347-357.
Andersen, A.N. and Ashton, D.H. (1985) Rates ofseed removal by ants at heath and woodlandsites in southeastern Australia. Aust.J.Ecol.10:381-390.
Andersen, A.N. and McKaige, M.E. (1987) Antcommunities at Rotamah Island Victoria,with particular reference to disturbance andRhytidoponera tasmaniensis. Proc. Roy. Soc. Vict.99: 141-146.
Beattie, A.J. (1995) (ed.) Biodiversity, Australia’sLiving Wealth. Research Unit forBiodiversity and Bioresources, School ofBiological Sciences, Macquarie University,Australia. 131pp.
Beattie, A.J. and Oliver, I. (1994) Taxonomicminimilisation. Trends in Ecology andEvolution 9:488-490.
Benson, D.H. (1985) Maturation periods for firesensitive shrub species in Hawkesburysandstone vegetation. Cunninghamia 1:339-349.
Birk, E.M. and Bridges, R.G. (1989) Recurrentfires and fuel accumulation in even-agedblackbutt (Eucalyptus pilularis) forests. For.Ecol. and Manage. 29:59-79.
Bornemissza, G.F. (1969) The re-invasion ofburnt woodland areas by insects and mites.Proc. Ecol. Soc. Aust. 4:138.
Bradstock, R.A. and Myerscough, P.J. (1981) Fireeffects on seed release and the emergenceand establishment of seedlings in Banksiaericifolia L.f. Aust. J. Bot. 29:521-532.
Brian, M.V., Mountford, M.D., Abbot, A. andVincent, S. (1976) The changes in antspecies distribution during ten years post-fire regeneration of a heath. J. Anim. Ecol.45:115-133.
Burbidge, A.H., Leicester, K., McDavitt, S. andMajer, J.D. (1992) Ants as indicators ofdisturbance at Yanchep National Park,Western Australia. J.Royal Soc.West.Aust.75:89-95.
Campbell, A.J. (1973) The Effects of PrescribedBurning on Surface Active Invertebrate Faunain Pine and Eucalypt Forest Within theAustralian Capital Territory. BSc. (Hons)Thesis. ANU. Canberra, ACT. Australia.
Campbell, A.J. and Tanton, M.T. (1981) Effect offire on the invertebrate fauna of soil andlitter of a eucalypt forest. In: Fire and theAustralian Biota. (Eds Gill, A.M., Groves,R.H. and Noble, I.R.). Australian Academyof Science, Canberra.
Cary, G.J. and Morrison, D.A. (1995) Effects offire frequency on plant species compositionof sandstone communities in the Sydneyregion: Combinations of inter-fire intervals.Aust.J.Ecol. 20:418-426.
Catling, PC. (1991) Ecological effects ofprescribed burning practices on themammals of southeastern Australia. In:Conservation of Australia’s Forest Fauna. (edLunney, D.), pp. 353-63. Royal ZoologicalSociety of NSW, Mosman.
Caughley, G. and Gunn, A. (1996) ConservationBiology in Theory and Practice. BlackwellScience, Massachusetts USA. 459pp.
Christensen, N. (1985) Shrubland fire regimesand their evolutionary consequences. Pp86-99 in S.T.A. Pickett and P.S.White (Eds) TheEcology of Natural Disturbance and PatchDynamics. Academic Press, Orlando, Florida.
REFERENCES

262
Australia’s Biodiveristy - Responses to Fire
Commonwealth of Australia (1983) NationalConservation Strategy for Australia. LivingResource Conservation for SustainableDevelopment. Department of Home Affairsand Environment. Aust.Govt.Pub.Service,Canberra.
CONCOM (1989) Council of NatureConservation Ministers - Australianstatement on invertebrates. Myrmecia 25,123-125.
Cornaby, B.W. (1977) Saprophagous organismsand problems in applied resourcepartitioning. pp. 96-100 in Mattson, W.J.(ed) (1977) The Role of Arthropods in ForestEcosystems. Springer-Verlag, New York.
Coy, R. (1996) The impact of fire on soilinvertebrates in E.regnans forest atPowelltown, Victoria. In: Fire andBiodiversity. The Effects and Effectiveness of FireManagement. pp. 183-198. BiodiversitySeries, Paper No. 8. Biodiversity Unit,Department of the Environment Sport andTerritories. Canberra, Australia.
CSIRO (1991) The Insects of Australia. SecondEdition. Melbourne University Press.
Dindal, D.L. and Metz, L.J. (1977) Communitystructure of Collembola affected by firefrequency. pp.88-95 in The Role of Arthropodsin Forest Ecosystems. Mattson, W.J. (ed)Springer-Verlag, New York.
Dolva, G. (1993) The effect of fire on the ecology andlife-history of the wood cricket Nambungiabalyarta (Nemobiinae:Gryllidae). UnpublishedMasters Thesis, University of WesternAustralia.
Duncan, K.W. (1969) The ecology of two speciesof terrestrial Amphipoda (Crustaceae:Family Tallitridae) living in waste grassland.Pedobiologia 9:323-341.
Elmes, G.W. and Wardlaw, J.C. (1982) Apopulation study of the ants Myrmicasabuletti and Myrmica scabrinodis, living at twosites in the south of England. II. Effect ofabove-nest vegetation. J.Anim.Ecol. 51:665-680.
Engelmann, M.D. (1961) The role of soilarthropods in the energetics of an old fieldcommunity. Ecol. Monogr. 31:221-238.
ESDWG-Forest Use (1991) EcologicallySustainable Development Working Groups.Final Report - Forest Use. AustralianGovernment Publishing Service, Canberra.227pp.
Forestry Commission of NSW (1992) 1992-1995Corporate Plan. For.Com.NSW, Sydney.26pp.
Fox, B.J. (1979) An objective method ofmeasuring the vegetation structure of animalhabitats. Aust.Wildl.Res. 6:297-303.
Fox, M.D. and Fox, B.J. (1982) Evidence forinterspecific competition influencing antspecies diversity in regenerating heathland.pp.99-110 in Buckley, R.C. (1982) (ed.) Ant-Plant Interactions in Australia. JunkPublishers. The Hague.
Fox, M.D. and Fox, B.J. (1986) The effect of firefrequency on the structure and floristiccomposition of a woodland understorey.Aust. J. Ecol. 11:77-85.
Friend, G. (1996) Fire ecology of invertebrates -implications for nature conservation, firemanagement and future research. In Fire andBiodiversity. The Effects and Effectiveness of FireManagement. Pp. 155-162. BiodiversitySeries, Paper No. 8. Biodiversity Unit,Department of the Environment Sport andTerritories. Canberra, Australia.
Friend, J.A. and Richardson, A.M.M. (1986)Biology of terrestrial amphipods. Ann. Rev.Entomol. 31: 25-48.
Gill, A.M. (1981) Adaptive responses of Australianvascular plant species to fires. In: Fire and theAustralian Biota (eds. A.M. Gill, R.H. Grovesand I.R. Noble), pp.243-272. Aust. Acad.Sci., Canberra.
Goldstein, E.L. (1975) Island biogeography ofants. Evolution 29:750-762.
Greenslade, P. and Greenslade, P.J.M. (1987)Ecological strategies in collembola: a newapproach to the use of terrestrialinvertebrates in environmental assessment.In: B.R. Striganova (Ed.), Soil Fauna and Soilfertility, pp.245-252. Nauka, Moscow.
Greenslade, P.J.M. (1984 ) Invertebrates andenvironmental assessment. Environment andPlanning 3:13-15.
Greenslade, P.J.M. (1985) Pterygote insects andthe soil: their diversity, their effects on soilsand the problem of species identification.Quaest. Ent. 21:571-585.

263
Bushfire and forest invertebrates
Greenslade, P.J.M. and Mott, J.J. (1979) Ants ofnative and sown pastures in the Katherinearea, Northern Territory, Australia.(Hymenoptera:Formicidae). Pp.153-156 inCrosby, T.K. and Pottinger, R.P. (eds)proceedings of The 2nd AustralasianConference on Grassland Invertebrate Ecology.Palmerston North, New Zealand. 22-26thMay, 1978. Wellington Govt. Printer.
Greenslade, P.J.M. and Thompson, C.H. (1981)Ant distribution, vegetation and soilrelationships in the Coolola-Noosa Riverarea, Queensland. In: Vegetation Classificationin Australia (eds A.N. Gillson & D.J.Anderson), pp. 192-207. CSIRO andAustralian National University Press,Canberra.
Harvey, M.S. and Yen, A.L. (1989) Worms toWasps. An Illustrated Guide to Australia’sTerrestrial Invertebrates. Oxford UniversityPress. 203pp.
Hobbs, R.J. (1992) Function of biodiversity inMediterranean ecosystems in Australia:definitions and background. In: Biodiversity ofMediterranean Ecosystems in Australia (ed. R.J.Hobbs), pp. 1-25. Surrey Beatty & Sons.
Holldobler, B. and Wilson, E.O. (1990) The Ants.Springer-Verlag, Berlin. 732pp.
Huhta, V. (1971) Succession in the spidercommunities of the forest floor after clear-cutting and prescribed burning. Ann. Zool.Fennici. 8:483-542.
Huhta, V.E., Karpinnen, E., Nurminen, M. andValpus, A. (1967) Effects of silviculturalpractices upon arthropod annelid andnematode populations in coniferous forestsoil. Anneles Zoologie Fennici 4:87-135.
Hurlbert, S.H. (1984) Pseudoreplication and thedesign of ecological field experiments. Ecol.Monogr. 54:187-211.
IUCN (1980) International Union for theConservation of Nature, United NationsEnvironment Program (UNEP) and WorldWildlife Fund (WWF) - WorldConservation Strategy - Living ResourceConservation for Sustainable Development.IUCN, Gland, Switzerland.
Krebs, C.J. (1985) Ecology. The ExperimentalAnalysis of Distribution and Abundance. ThirdEdition. Harper & Row, New York. 800pp.
Kremen, C. (1992) Assessing the indicatorproperties of species assemblages for naturalareas monitoring. Ecological Applications2:203-217.
Lambert, M.J. (1982) Methods for ChemicalAnalysis. Forestry Commission of NSW.Sydney, Australia. 3rd Edition.
Lawrence, J.F. and Britton, E.B. (1994) AustralianBeetles. Melbourne University Press. 192pp.
Leonard, B.V. (1972) The effect of fire upon selectedsmall mammals and leaf litter fauna insclerophyll forest in southern Australia. Unpubl.MSc. Thesis. Monash University, Victoria,Australia.
Lowman, M.D. (1982) Seasonal variation in insectabundance among three Australian rainforests, with particular reference tophytophagous types. Aust. J. Ecol. 7:353-361.
MacArthur, R.H. and MacArthur, J.W. (1961) Onbird species diversity. Ecology 42:594-598.
MacFadyen, A. (1962) Soil arthropod sampling.Adv. Ecol. Res. 1:1-34.
MacQuillan, P.B. (1985) The identification ofroot-feeding cockchafer larvae (Coleoptera:Scarabaeidae) found in pastures in Tasmania.Aust.J.Zool. 33: 509-46.
Majer, J.D. (1980) Report on a study ofinvertebrates in relation to the KojonupNature Reserve fire management plan. Bull.Dept. Biol. West. Aust. Inst. Tech. 2:1-22.
Majer, J.D. (1984) Ant return in rehabilitationmines - an indication of ecosystem resilience.In : B.Dell (ed.), Medicos IV:Proceedings 4thInternational Conference on MediterraneanEcosystems, pp.105-106, Botany Dept.,University of Western Australia, Nedlands.
Majer, J.D. (1985) Recolonisation by ants ofrehabilitated mineral sand mines on northStradbroke Island, Queensland, withparticular reference to seed removal. Aust. J.Ecol. 10:31-48.
Majer, J.D. (1992) Ant recolonisation ofrehabilitated bauxite mines of Pocos deCaldas, Brazil. J.Trop.Ecol. 8:97-108.
Majer, J.D., Satori, M., Stone, R. and Perriman,W.S. (1982) Recolonisation by ants andother invertebrates in rehabilitated mineralsand mines near Eneabba, Western Australia.Reclaim. Reveg. Res. 1:63-81.
Majer, J.D., Day, J.E., Kabay, E.D. and Perriman,W.S. (1984) Recolonisation by ants inbauxite mines rehabilitated by a number ofdifferent methods. J. Appl. Ecol. 21:355-375.

264
Australia’s Biodiveristy - Responses to Fire
Margules, C.R., Nicholls, A.O. and Pressey, R.L.(1988) Selecting networks of reserves tomaximise biological diversity. BiologicalConservation 43:63-76.
Mattson, W.J. (ed) (1977) The Role of Arthropods inForest Ecosystems. Springer-Verlag, New York.
Mawson, P.R. (1986) A comparative study ofarachnid communities in rehabilitatedbauxite mines. W.A.I.T. School of BiologyBulletin No.14.
McIver, J.D., Parsons, G.L. and Moldenke, A.R.(1992) Litter spider succession after clear-cutting in a western coniferous forest.Canad.J.For.Res. 22:984-992.
Merrett, P. (1976) Changes in the ground-livingspider fauna after heathland fires in Dorset.Bull. Br. Arachnol. Soc. 3:214-221.
Metz, L.J. and Dindal, D.L. (1975) Collembolapopulations and prescribed burning. Environ.Entomol. 4:583-587.
Metz, L.J. and Farrier, M.H. (1973) Prescribedburning and populations of soil mesofauna.Environ. Entomol. 2:433-440.
Morris, M.G. (1975) Preliminary observations onthe effects of burning on the Hemiptera(Homoptera and Auchenorhyncha) oflimestone grassland. Biol. Conserv. 7:312-319.
Moulton, T.P. (1982) The effect of prescribed burningand simulated burning on soil and litterarthropods in open forest at Cordeaux, N.S.W.Australia. Unpubl. PhD. Thesis. MacquarieUniversity.
Neumann, F.G. (1992) Effects of low intensityprescribed fire on invertebrates in mixedeucalypt forest - Wombat State Forest. InTolhurst, K. And Flinn, D. (eds.) EcologicalImpacts of Fuel Reduction Burning in DrySclerophyll Forest: First Progress Report.Conservation and Environment, Victoria,Australia.
Neumann, F.G. and Tolhurst, K. (1991) Effects offuel reduction burning on epigeal arthropodsand earthworms in dry sclerophyll eucalyptforest of west-central Victoria. Aust. J. Ecol.16, 315-330.
New, T.R. (1984) Insect Conservation: An AustralianPerspective. Dr. W.Junk Publishers,Netherlands.
New, T.R. (1995) Introduction to InvertebrateConservation Biology. Oxford UniversityPress, New York. 194pp.
NFPS (1992) National Forest Policy Statement. ANew Focus for Australia’s Forests.Commonwealth of Australia. Canberra.
Nichols, O.G. and Bunn, S. (1980) Termiteutilisation of rehabilitated bauxite minedareas. Alcoa of Australia Limited. Environ.Res. Bull.No.9.
Nieuwenhuis, A. (1987) The effect of firefrequency on the sclerophyll vegetation ofthe West Head, New South Wales.Aust.J.Ecol. 12:373-385.
Noble, I.R. and Slatyer, R.O. (1981) Concepts andmodels of succession in vascular plantcommunities subject to recurrent fire. In Fireand the Australian Biota (Gill, A.M., Groves,R.H. and Noble, I.R. eds.), pp.311-315.Aust.Acad.Sci., Canberra.
NSESD (1992) National Strategy for EcologicallySustainable Development. Commonwealthof Australia. 128pp.
Oliver, I. (1995) Rapid Biodiversity Assessment andits Application to Invertebrate Conservation inProduction Forests. Unpubl. PhD Thesis.School of Biological Sciences, MacquarieUniversity, Australia.
Oliver, I. and Beattie, A. J. (1993) A possiblemethod for the rapid assessment ofbiodiversity. Cons.Biol. 7:562-568.
Oliver, I. and Beattie, A. J. (1996a) Invertebratemorphospecies as surrogates for species: acase study. Cons. Biol. 10:99–109.
Oliver, I. and Beattie, A. J. (1996b) Designing acost-effective invertebrate survey: a test ofmethods for rapid biodiversity assessment.Ecological Applications 6:594-607.
Palmer, M.W. (1993) Putting things in even betterorder: The advantages of canonicalcorrespondence analysis. Ecology 74:2215-2230.
Pickett, S. T. A. and White, P. S. (1985) (Eds) TheEcology of Natural Disturbance and PatchDynamics. Academic Press, Orlando, Florida.
Punttila, P., Haila, Y., Payunen, T and Tukia H.(1991) Colonisation of clearcut forests byants in the southern Finnish taiga: aquantitative survey. Oikos 61, 250-262.
Ridley, D. (1993) Fire fuel management in EdenState Forest District. Pp. 91-101 in Ross, J.(Ed) The Burning Question: Fire Managementin NSW. Proceedings of a Conference heldat the Department of Continuing Education,University of New England, Armidale.

265
Bushfire and forest invertebrates
Samways, M. J. (1994) Insect Conservation Biology.Chapman & Hall, London. 358 pp.
Seastedt, T. R. (1984) Below-groundmacroarthropods of annually burned andunburned tallgrass prairie. Am. Midl. Nat.111:405-408.
Springett, J. A. (1976) The effect of prescribedburning on the soil fauna and litterdecomposition in Western Australian forests.Aust.J.Ecol. 1:77-82.
Springett, J. A. (1979) The effects of a single hotsummer fire on soil fauna and on litterdecomposition in jarrah (Eucalyptusmarginata) forest in Western Australia. Aust.J. Ecol. 4:279-91.
SFNSW (1995) Forest Management Zoning in StateForests (Draft). State Forests of New SouthWales. Sydney, Australia.
Taylor, R. W. (1983) Systematics and ecology:reflections from the interface. Pp50-79 inE.Highley and R.W. Taylor (eds) AustralianSystematic Entomology: A BicentenaryPerspective. CSIRO, Melbourne.
Ter Braak, C. J. F. (1986) CanonicalCorrespondence Analysis: a new eigenvectortechnique for multivariate direct gradientanalysis. Ecology 67:1167-1179.
Ter Braak, C. J. F. (1991) CANOCO version 3.12.Agricultural Mathematics Group, DLO Box100, 6700 AC, Wageningen, TheNetherlands.
Tolhurst, K. G., Kellas, J. D. and Wilson, A. A.(1992) Low intensity fire behaviour and fueldynamics in dry sclerophyll forest, WombatState Forest. In Tolhurst, K. And Flinn, D.(eds.) Ecological Impacts of Fuel ReductionBurning in Dry Sclerophyll Forest: FirstProgress Report. Conservation andEnvironment, Victoria, Australia.
Turner, J. (1993) Research and EcologicallySustainable Forest Management. A DiscussionPaper. Research Division, State Forests ofNew South Wales. 26pp.
Van der Drift, J. (1958) The role of the soil faunain the decomposition of forest litter. pp.357-360 in Proc. 15th Intern. Congr. Zool.
Van Loon, A.P. (1970) Further Investigations inthe Effects of Prescribed Burning on YoungEven-aged Blackbutt. Taree Forestry OfficeResearch Report.
WCED (1987) World Commission onEnvironment and Development. OurCommon Future. Oxford University Press,Oxford, UK. 400pp.
Weir, E.G. (1978) Development of a Fire Policy.Parks and Wildlife, Vol.2. National Parksand Wildlife Service of N.S.W.
Welch, R.C. (1978) Changes in the distribution ofthe nests of Formica rufa L.(Hymenoptera:Formicidae) at Brian WoodsNational Nature Reserve, Kent, during thedecade following coppicing. Insectes Sociaux25:173-186.
Whelan, R.J., Langedyk, W. and Pashby, A.S.(1980) The effects of wildfire on arthropodpopulations in a Jarrah - Banksia woodland.W.A. Naturalist 14:214-220.
Whelan, R.J. (1995) The Ecology of Fire.Cambridge University Press. 346pp.
Williams, J.E. and Gill, A.M. (1995) The Impact ofFire Regimes on Native Forests in Eastern NewSouth Wales. Environmental HeritageMonograph Series No. 2. NSW NationalParks and Wildlife Service. 68pp.
Woodward, F.I. (1993) How many species arerequired for a functioning ecosystem? InEcosystem Function of Biodiversity. Eds. E.D.Schultz and H.A. Mooney. Springer-Verlag,Heidelberg.
Yeatman, E.M. and Greenslade, P.J.M. (1980)Ants as indicators of habitat in threeconservation parks in South Australia.S.Aust.Nat. 55:20-26, 30.
Yen, A.L. (1987) A preliminary assessment of thecorrelation between plant, vertebrate andColeoptera communities in the Victorianmallee. In : Majer, J.D. (ed.), The Role ofInvertebrates in Conservation and BiologicalSurvey pp.73-88. W.A. Dept. ofConservation and Land ManagementReport.
York, A. (1989) The Response of Ant Communities toFire-induced Habitat Disturbance.Unpublished PhD. Thesis, School ofBiological Sciences, University of NewSouth Wales.
York, A. (1993) Towards ecologically sustainableforest use. In Rapid Biodiversity Assessment (edBeattie, A.J.), pp. 51-55. Proceedings of theBiodiversity Workshop MacquarieUniversity - 1993. Research Unit forBiodiversity and Bioresources, School ofBiological Sciences, Macquarie University.

266
Australia’s Biodiveristy - Responses to Fire
York, A. (1994) The long-term effects of fire onforest ant communities: managementimplications for the conservation ofbiodiversity. Memoirs of the QueenslandMuseum 36, 231-239.
York, A. (1996) Long-term effects of fuelreduction burning on invertebrates in a drysclerophyll forest. In Fire and Biodiversity.The Effects and Effectiveness of FireManagement. Pp. 163-181. BiodiversitySeries, Paper No. 8. Biodiversity Unit,Department of the Environment Sport andTerritories. Canberra, Australia.
Zborowski, P. and Storey, R. (1995) A Field Guideto Insects in Australia. Reed Books, Sydney.207pp.
Zedler, P.H., Gautier, C.R. and McMaster, G.S.(1983) Vegetation change in response toextreme events: the effect of a short intervalbetween fires in California chaparral andcoastal scrub. Ecology 64:809-818.


















