This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
Acanthocheilonema delicata n. sp. (Nematoda: Filarioidea) from Japanese badgers(Meles anakuma): Description, molecular identification, and Wolbachia screening
Shigehiko Uni a,b,⁎, Odile Bain c, Kazuo Suzuki d, Takeshi Agatsuma e, Masashi Harada f,Masaharu Motokawa g, Coralie Martin c, Emilie Lefoulon c, Masako Fukuda h,i, Hiroyuki Takaoka a,i
a Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysiab Department of Parasitology, Graduate School of Medicine, Osaka City University, Abeno-ku, Osaka 545‐8585, Japanc Parasitologie comparée, UMR 7205, Muséum National d'Histoire Naturelle, 75231 Paris, Franced Hikiiwa Park Center, Tanabe, Wakayama 646‐0051, Japane Department of Environmental Health Science, Faculty of Medicine, Kochi University, Nankoku, Kochi 783‐8505, Japanf Laboratory Animal Center, Graduate School of Medicine, Osaka City University, Osaka 545‐8585, Japang The Kyoto University Museum, Yoshida Honmachi, Sakyo-ku, Kyoto 606‐8501, Japanh Research Promotion Project, Oita University, Oita 879‐5593, Japani Department of Infectious Disease Control, Faculty of Medicine, Oita University, Oita 879‐5593, Japan
a b s t r a c ta r t i c l e i n f o
Article history:Received 30 January 2012Received in revised form 9 August 2012Accepted 15 August 2012Available online 25 August 2012
Keywords:Acanthocheilonema delicata n. sp.Dermal microfilariaEndemicityFilarioidMeles anakumaMolecular identificationWolbachia
Acanthocheilonema delicata n. sp. (Filarioidea: Onchocercidae: Onchocercinae) is described based on adultfilarioids and microfilariae obtained from subcutaneous connective tissues and skin, respectively, of Japanesebadgers (Meles anakuma) inWakayama Prefecture, Japan. No endemic species of the genus had been found inJapan. Recently, some filarioids (e.g., Acanthocheilonema reconditum, Dirofilaria spp., and Onchocerca spp.)have come to light as causative agents of zoonosis worldwide. The new species was readily distinguishedfrom its congeners by morphologic characteristics such as body length, body width, esophagus length, spiculelength, and the length of microfilariae. Based on the molecular data of the mitochondrial cytochrome coxidase subunit 1 (cox1) gene, A. delicata n. sp. was included in the clade of the genus Acanthocheilonemabut differed from two other congeneric species available for study, A. viteae and A. reconditum.Acanthocheilonema delicata n. sp. did not harbor Wolbachia. It is likely that the fauna of filarioids from mam-mals on the Japanese islands is characterized by a high level of endemicity.
Cobbold created the genus Acanthocheilonema with one species,Acanthocheilonema dracunculoides, from specimens collected fromthe aardwolf (Proteles cristatus: Hyaenidae) in South Africa [1]. Thegenus consists of 14 species which have a wide range of mammalianhosts (Carnivora, Macroscelidea, Rodentia, Pholidota, Edentata, andMarsupialia) from the major continents of the world.
Although a high degree of endemicity appears to characterize thespecies among several genera of filarioids obtained from mammalianhosts in Japan [2–7], no endemic species have yet been found in thegenus Acanthocheilonema. The widespread species recorded in Japanare: A. reconditum (Grassi, 1889) from dogs in Okinawa [8]; A. odendhali(Perry, 1967) and A. spirocauda (Leidy, 1858) from Pinnipedia in thecoasts of Hokkaido [9,10].
Acanthocheilonema reconditum may be a zoonotic agent as shownrecently in Australia [11], similar to several filarial species from dogs,
the most common being Dirofilaria immitis (Leidy, 1856) and Dirofilariarepens Railliet and Henry, 1911 [12,13], and Onchocerca lupi Rodonaja,1967 with a single case from Turkey [14]. Onchocerca dewittei japonicaUni, Bain and Takaoka, 2001 fromwild boar was identified as a zoonoticagent in Japan [15–17]. Because of the potential of onchocercid wormsto infest humans, research endeavors on the fauna of filarial parasitesand on their vectors are needed for public health [18].
In this article, we describe a new species of Acanthocheilonemabased on adult filarioids and the microfilariae collected from Japanesebadgers and show its phylogenetic relationships by an analysis of theDNA sequences of its mitochondrial cytochrome c oxidase subunit 1(cox1) gene. We also indicate the absence of the endosymbiont bacte-ria Wolbachia in the new species, because Wolbachia have biologicaland evolutional relationships to the Onchocercidae, as well as patho-logic significance to humans [19].
2. Materials and methods
The host animals examined for filarioids were Japanese badgers(Meles anakuma Temminck, 1844), an endemic animal of Japan.
Contents lists available at SciVerse ScienceDirect
Parasitology International
j ourna l homepage: www.e lsev ie r .com/ locate /par in t
Author's personal copy
Twenty-five animals were found in road-kill between December 2005and May 2010 in the vicinity of Tanabe City, Wakayama Prefecture,Japan. The carcasses were shipped refrigerated to a laboratory forfilarial examination one or two days after being found.
For the detection of microfilariae, skin snips were taken from theface, ears, neck, back, limbs, and tail of each animal by the methodsdescribed by Uni et al. [20]. Blood films were made from each badgerand stained with 3% Giemsa's solution in 0.1 M phosphate butter(pH 6.8). For the detection of adult worms, skin and subcutaneousconnective tissues of the whole animal were inspected under astereomicroscope.
Adult worms detected were placed in 2% formalin in saline solu-tion and the specimens were cleared in lactophenol for morphologicstudy. Drawings of the parasites were done by the use of a cameralucida. For histologic examinations, the midbody of a female parasitewas embedded in paraffin by routine methods and the sectionswere stained with hematoxylin and eosin. Scanning electron micros-copy (SEM) of the male worms was prepared as described elsewhere[4]. The body length of adult worms, the length of ovejector, and thelength of area rugosa were measured in millimeters; the other di-mensions in micrometers. Esophagus ratio (%) was determined asthe ratio of the esophagus length to the body length. In the male,the caudal pairs of papillae were tentatively numbered on the basisof Chabaud and Petter [21]. The authorities of the new species areUni and Bain.
For molecular analysis, three female worms were frozen at −20 °Candwere used to examine the nucleotide sequences of themitochondrialcytochrome c oxidase subunit 1 (cox1) of these females as describedin Agatsuma et al. [22]. DNA extraction and PCR amplification were car-ried out according to the methods in our previous study. The primersused for the cox1 mitochondrial region were COIinfF (5′-TGA TTG GTGGTT TTG GTA A-3′) and COIintR (5′-ATA AGT ACG AGT ATC AATATC-3′) [22].
The cox1 sequences of the following 14 species from the GenBankwere used in this analysis: Acanthocheilonema delicata n. sp. (accessionnumber, tentatively registered);A. reconditum;A. viteae (Krepkogorskaya,1933) [23]; Cercopithifilaria bulboidea Uni and Bain, 2001 [4]; C. crassaUni, Bain and Takaoka, 2002 [20]; C. minuta Uni and Bain, 2001 [4];C. multicauda Uni and Bain, 2001 [4]; C. shohoi Uni, Suzuki and Katsumi,1998 [24]; C. tumidicervicata Uni and Bain, 2001 [4]; C. japonica (Uni,1983) [25]; Mansonella (Cutifilaria) perforata Uni, Bain and Takaoka,2004 [5];O. dewittei japonica;O. skrjabiniRukhlyadev, 1961 [3]; and Filariamartis Gmelin, 1790.
Pairwise distances in amino acid sequences of the cox1 geneamong the 14 species mentioned above were obtained using theJones–Taylor–Thornton (JTT) matrix-based model. The rate of varia-tion among sites was modeled with a gamma distribution (MAGA 5)[26]. All positions containing gaps, missing data, or both gaps andmissing data were eliminated. There were a total of 191 positions inthe final dataset.
Phylogenetic analyses were done using the neighbor-joining (NJ)and maximum-likelihood (ML) methods in MEGA 5 (ver. 5) [26].
Immunologic staining for Wolbachia was done according to themethods described by Kramer et al. [27]. Briefly, one female specimenof A. delicata n. sp. fixed in 4% paraformaldehyde was embeddedin paraffin. Sections (4 μm thick) were made and placed on Silane(3-aminpropyltriethoxysilane)-coated glass slides. A rabbit polyclonalantiserum raised against the Wolbachia surface protein (WSP) ofthe endobacteria from Brugia pahangi (Buckley and Edeson, 1956)was used (1:2000) to stain sections of A. delicata n. sp. Sectionsof Litomosoides sigmodontis Chandler, 1931 were used as a positivecontrol. Negative controls were done by omitting the primary anti-body. PCR screening for Wolbachia of two females of A. delicata n. sp.was conducted following the methods described by Casiraghi et al.[28] and Ferri et al. [29], using general Wolbachia primers for 16SrDNA.
3. Results
3.1. Description of Acanthocheilonema delicata Uni & Bain, n. sp.
Small, thin, and delicate filarioids. Females long, twice as long asmales (Table 1). Anterior extremity slightly bulbous, bearing two setsof four papillae and amphids (Fig. 1A-B). Buccal cavity distinct; buccalcapsule with thick buccal ring (Fig. 1C–E). Esophagus divided intoshort anterior muscular portion and long, broad posterior glandularportion (Fig. 1A). Deirids at level of junction of muscular and glandularportions of esophagus (Fig. 1F–G). Caudal extremity of females andmales equipped with three conical lappets (detailed description infemale, microfilaria, and male sections).
Female (19 complete specimens and 17 fragments; Fig. 1A–N): fe-male worms with body swelling at level posterior to vulva (Fig. 1A, *).Cephalic end trapezoidal in most of specimens in lateral view. Headpapillae (Fig. 1B): anterior set of four external labial papillae andposterior set of four cephalic papillae; en face view, both sets in twosquares; buccal cavity narrow with wall composed of two parts(Fig. 1D): cuticle of anterior part of head invaginated, widened nearposterior buccal ring, and flattened on it; buccal ring as wide asesophagus apex. Length of buccal cavity 6 (holotype; range: 6–11)μmand length of buccal ring 2.5 (2.5)μm. Slits of amphids anterior to papil-lae (Fig. 1E, arrow).
Nerve ring at middle level of anterior esophagus (Fig. 1A). Deiridswith short point 320 and 325 (320–380)μm from head (Fig. 1F–G).Intermediary flat piece at junction of anterior and posterior parts ofesophagus in some worms. Intestine narrower than glandular esoph-agus. Vulva, transverse slit at anterior level of glandular esophagus(Fig. 1A and H). Vagina with complex lumen; length/width: 70/70(70–100/65–80)μm (Fig. 1H). In vagina, transverse flattened shorttube, chamber directed posteriorly and lined with thick epithelialcells, narrow tube directed anteriorly, and beginning of ovejector di-rected posteriorly. Long, thin, and straight ovejector 2.1–3.4 mmlong (Fig. 1I). Uteri narrow, straight, and opisthodelphic with aligneddeveloping embryos.
At midbody (Fig. 1J), lateral cuticular thickening (*) and lateralchord (LC) 40 μm wide. Single, lateral, deiridlike structure near levelof anus, extending outwards from left lateral chord (Fig. 1K, arrow).Ovary apices rounded, 340 and 710 (340–3000)μm from tail end(Fig. 1L, *). Tail dorsally at level of anus. Three cuticular terminal lap-pets ventrally (Fig. 1M–N); each lappet conical with terminal smallnipple; at caudal end, two lateral extensions and shorter dorsal oneof subcuticular tissue of body (Fig. 1M). Phasmids subterminal atbase of lateral lappets (Fig. 1N).
Microfilaria (Fig. 1O–Q): Microfilariae from uterus unsheathed.Relatively short and thick body (Table 1, Fig. 1O); left cephalic hookshort (Fig. 1P); head round with slight neck (Fig. 1Q), distinctly inmedian view; cephalic space 5 μm long; column of body nuclei thin-ner than body width; nerve ring 37 μm and excretory pore 60 μmfrom head; tail 32 μm long. Tail attenuated; terminal end not sharp;anucleated terminal part 12 μm long. Microfilariae in skin snips ofhost animals tightly and spirally coiled in general.
Male (9 complete specimens and 2 fragments; Fig. 2): Anteriorpart with slightly bulbous cervix (Fig. 2A). Lateral thickening and lat-eral chord near nerve ring; width of lateral chords 12 (allotype;range: 12–14)μm (Fig. 2A, *). Length of buccal cavity 8 (6–8)μm;length of buccal ring 2.5 (2.0–2.5)μm (Fig. 2B–C). Deirid posteriorto nerve ring (Fig. 2A, arrow; D, arrows; E); both deirids 320(280–320)μm from head. Esophagointestinal junction in Fig. 2F. Tes-tis apex bent, 240 (215–275) μm posterior to esophagointestinaljunction (Fig. 2G).
Area rugosa extending from anterior border 1.93 (1.2–2.2)mmfrom tail end to posterior border 0.48 (0.25–0.85)mm from tail end,being composed of transverse bands (160–222 bands) with longitudi-nal rods in band (Fig. 2H). Posterior region of body spirally coiled
15S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
(Fig. 2I–J); caudal alae present. Right spicule short with narrowhandle (Fig. 2K–L); posterior half spoon-shaped with membranousedges; delicate dorsal crests in terminal third of spoon (Fig. 2L, *);at tip, membranous triangular small flap. Left spicule thin with han-dle, long twisted intermediary piece (50–60 μm), long lamina withrod-like, round posterior end (Fig. 2I–J; M–N). Ratio of left spiculeto right spicule 2.5–2.9.
Caudal papillae (Fig. 2I; O–R): regular arrangement consisted offour pairs of precloacal papillae numbered 1 to 4; two post-cloacalpairs (pairs 5 and 6) almost on same transverse line posteriorlyclose to cloacal aperture; pair 7 close to this line; pairs 8 and 9 nearcaudal end (pair 8 ventral; pair 9 lateral). Papillae of pairs 1 and 7inconstant: one of papillae of pair 7 rarely at mid-length of tail (Fig. 2I).Phasmids subterminal at base of lateral lappets. Tail end narrow, withthree cuticular lappets (Fig. 2Q–R).
On transverse section of midbody of female, thicker cuticle atlateral fields than that of median field; 6–9 muscular cells perquadrant (Fig. 3). In SEM pictures of male worms, deirid withsmall projection (0.7 μm long, Fig. 4); distance between cuticu-lar striations at midbody 0.5 μm (Fig. 5); area rugosa: longitudi-nal rods with 2 μm high in transverse band and distancebetween bands 3 μm (Fig. 6); and three lappets at posteriorend, with phasmidial pore at base of lateral lappet (Fig. 7,arrow).
Habitat: Adult worms in the subcutaneous connective tissues.Microfilariae in the skin, mainly of the back. No microfilariae of thepresent filarioid in the blood films of the badgers examined.
Type locality: Mountainside, Nakaheji-cho, Tanabe City, WakayamaPrefecture, Japan.
Collection dates: Type specimens on 7–8 May 2009.Specimens deposited: Holotype (female, N37), Collection number
of the Museum National d'Histoire Naturelle (MNHN), Paris, 91YU;allotype (male, N46), Collection number of MNHN, 91YU; paratypesfemales (N40, N43) and males (N22, N30), Collection number ofMNHN, 91YU. Paratypes females (N4, N7) and males (N2, N10) inthe Kyoto University Museum, Kyoto, Japan. Fragments of specimens(AG1-3) used for DNA analysis at Kochi University.
3.3. Prevalence and distribution of adult worms and microfilariae ofA. delicata n. sp. in Japanese badgers
Adult specimens of A. delicata n. sp. (47worms) were collected fromonly one badger (ID no. A15): 36 females and 11 males. Thirty-eightspecimens were found in the subcutaneous connective tissues in the
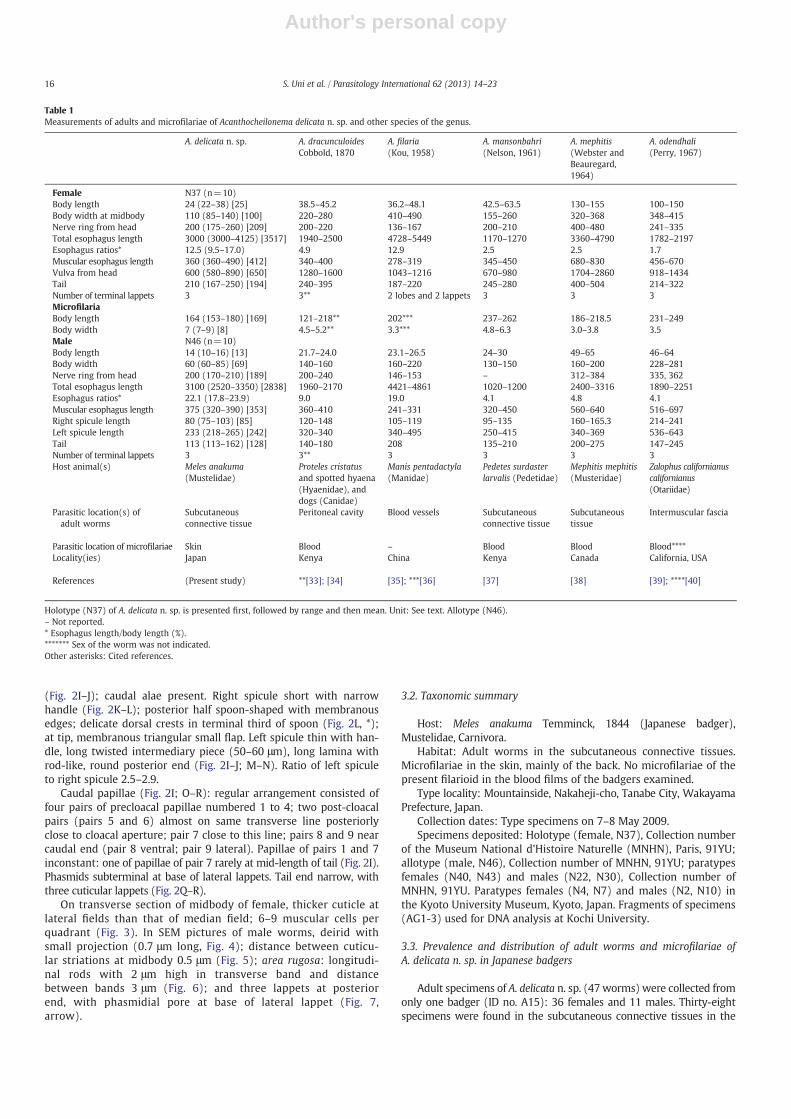
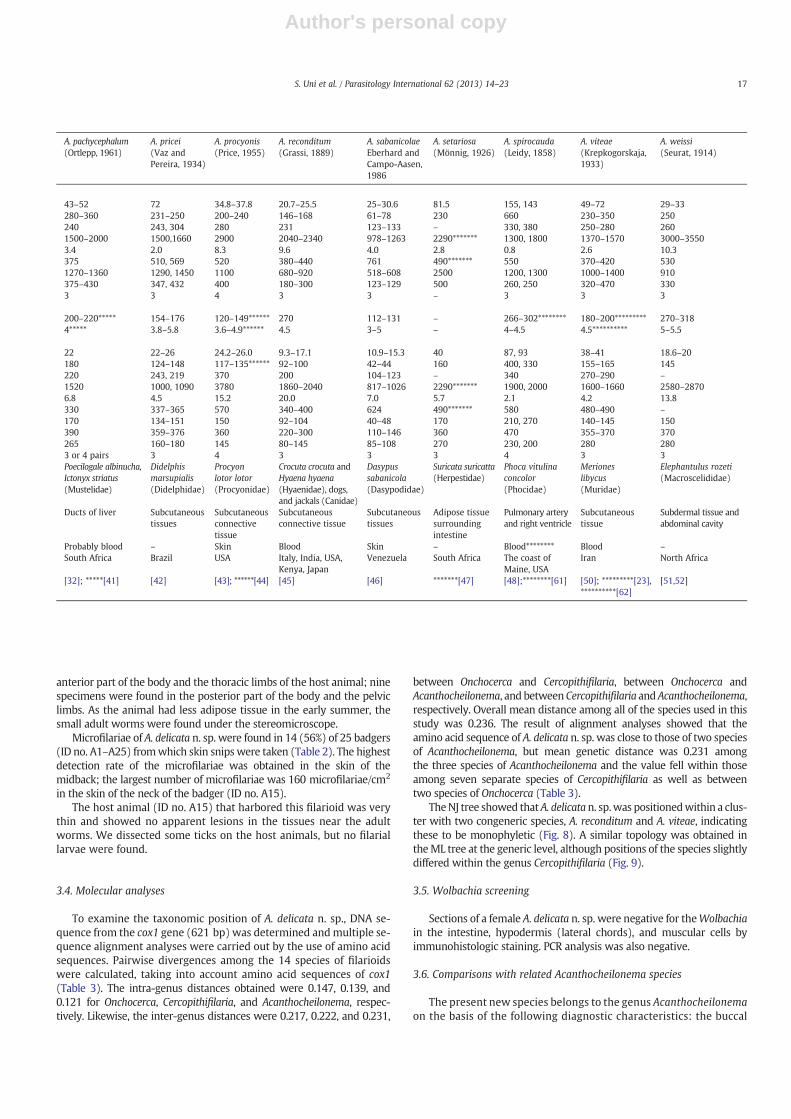
Table 1Measurements of adults and microfilariae of Acanthocheilonema delicata n. sp. and other species of the genus.
Holotype (N37) of A. delicata n. sp. is presented first, followed by range and then mean. Unit: See text. Allotype (N46).– Not reported.* Esophagus length/body length (%).******* Sex of the worm was not indicated.Other asterisks: Cited references.
16 S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
anterior part of the body and the thoracic limbs of the host animal; ninespecimens were found in the posterior part of the body and the pelviclimbs. As the animal had less adipose tissue in the early summer, thesmall adult worms were found under the stereomicroscope.
Microfilariae of A. delicata n. sp. were found in 14 (56%) of 25 badgers(ID no. A1–A25) fromwhich skin snips were taken (Table 2). The highestdetection rate of the microfilariae was obtained in the skin of themidback; the largest number of microfilariae was 160 microfilariae/cm2
in the skin of the neck of the badger (ID no. A15).The host animal (ID no. A15) that harbored this filarioid was very
thin and showed no apparent lesions in the tissues near the adultworms. We dissected some ticks on the host animals, but no filariallarvae were found.
3.4. Molecular analyses
To examine the taxonomic position of A. delicata n. sp., DNA se-quence from the cox1 gene (621 bp) was determined andmultiple se-quence alignment analyses were carried out by the use of amino acidsequences. Pairwise divergences among the 14 species of filarioidswere calculated, taking into account amino acid sequences of cox1(Table 3). The intra-genus distances obtained were 0.147, 0.139, and0.121 for Onchocerca, Cercopithifilaria, and Acanthocheilonema, respec-tively. Likewise, the inter-genus distances were 0.217, 0.222, and 0.231,
between Onchocerca and Cercopithifilaria, between Onchocerca andAcanthocheilonema, and betweenCercopithifilaria andAcanthocheilonema,respectively. Overall mean distance among all of the species used in thisstudy was 0.236. The result of alignment analyses showed that theamino acid sequence of A. delicata n. sp. was close to those of two speciesof Acanthocheilonema, but mean genetic distance was 0.231 amongthe three species of Acanthocheilonema and the value fell within thoseamong seven separate species of Cercopithifilaria as well as betweentwo species of Onchocerca (Table 3).
TheNJ tree showed thatA. delicatan. sp.was positionedwithin a clus-ter with two congeneric species, A. reconditum and A. viteae, indicatingthese to be monophyletic (Fig. 8). A similar topology was obtained intheML tree at the generic level, although positions of the species slightlydiffered within the genus Cercopithifilaria (Fig. 9).
3.5. Wolbachia screening
Sections of a female A. delicata n. sp. were negative for theWolbachiain the intestine, hypodermis (lateral chords), and muscular cells byimmunohistologic staining. PCR analysis was also negative.
3.6. Comparisons with related Acanthocheilonema species
The present new species belongs to the genus Acanthocheilonemaon the basis of the following diagnostic characteristics: the buccal
17S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
capsule with a distinct posterior ring, the esophagus withwell-developed glandular part, and the tail with caudal lappets inboth sexes [30–32].
Acanthocheilonema delicata n. sp. and 14 other congeneric species arelisted (Table 1). The specimens herein described have been first discov-ered from Japanese badgers (M. anakuma) among Mustelidae.
A
100
BC
D
E
F
G
H
P
Q
JLC
LK
M NI
O
2050
5050
50
1010
200
20
20
20
20 20
20
2020
Fig. 1. Acanthocheilonema delicata n. sp., females andmicrofilariae. A. Anterior part, left lateral view. * Body swelling. B. Head, apical view. C. Head,median view. D. Buccal capsule. E. Head,lateral view. Amphid (arrow). F. Deirids (arrows) posterior to the nerve ring,median view. G. Enlargeddeirid. H. Vaginawith body swelling. I. Ovejector. J.Midbody, lateral view. Twouteriwith embryos and microfilariae. LC: lateral chord. * Lateral cuticular thickening. K. Posterior part. Single, lateral, deiridlike structure near level of anus (arrow), ventral view. L. Posteriorpart, left lateral view. * Ovary.M. Posterior end, ventral view. N. Posterior end, right lateral view. O.Microfilaria, right lateral view. P. Anterior part, left lateral view. Q. Anterior part, medianview. Bars, micrometers.
18 S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
7
4567
89
1
2
3
B
A
E
H
IK
ML
J
O
P
Q
R
C
D
50
50
50
20 20
20
F
20
G
20
20 10
20
2010
10
50
N
50
50
10
Fig. 2. Acanthocheilonema delicata n. sp., males. A. Anterior part, lateral view. * Lateral thickening (center) and lateral chord with both dotted areas. Deirid (arrow). B. Head, median view.C. Head, lateral view. D. Deirids (arrows), median view. E. Enlarged deirid. F. Esophagointestinal junction. G. Testis apex. H. Area rugosa, ventral view. I. Posterior part. J. Posterior part, rightlateral view. K. Right spicule, lateral view. L. Posterior part of the right spicule, lateral view. * Ridges. M. Left spicule, ventral view. N. Left spicule, right lateral view. O. Posterior part, ventralview. Numbers indicate the pair of caudal papaillae. P. Cloacal region with different arrangement of the caudal papillae. Q. Posterior end, ventral view. R. Posterior end, left lateral view.Bars, micrometers.
19S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
Comparisons of A. delicata n. sp. with 14 known specieswere done on thebody length, esophagus length, esophagus ratios, body width, spiculelength, and the length ofmicrofilariae. Distinctive characteristics are indi-cated below.
Adult worms of A. dracunculoides Cobbold, 1870 are at least twiceas thick as those of A. delicata n. sp.; the microfilariae with a filamen-tous tail, whereas the microfilariae of A. delicata n. sp. lack a sharp tail[33,34]. Adult worms of A. filaria (Kou, 1958) have the longest esoph-agus among the genus; one pair of ventral subterminal lappets andtwo small lobes in the female tail end [35]. Microfilariae of A. filaria(=D. fausti) have a filamentous tail [36].
Adult worms of A. mansonbahri (Nelson, 1961) have a round headwith neck, a particularly short esophagus, and a larger axial lappet thanthe two lateral ones [37]. Females ofA.mephitis (Webster and Beauregard,1964) have a body five times as large with thick conical, straight tailthan that of A. delicata n. sp. [38]. Adult worms of A. odendhali havegreater body and the left spicule has a bifurcated tip [39,40]; themicrofilariae have long cephalic hook and filamentous tail [40].
In A. pachycephalum (Ortlepp, 1961) three small caudal points arefound in the female; three or four round tubercles on the tail end ofthe male [32,41]. Adult worms of A. pricei (Vaz and Pereira, 1934)have a short esophagus [42].
Acanthcheilonema procyonis (Price, 1955) is similar to our specimenswith a high esophagus ratio, but the tail end is equipped with fourlappets in both sexes [43,44]. Microfilariae of A. procyonis and A. delicatan. sp. have a tiny cephalic hook and a short nonfilamentous tail.Acanthocheilonema reconditum together with A. delicata n. sp. has thesmallest adult worms in the genus, but the microfilariae are longerthan A. delicata n. sp. [45] and have a filamentous tail [8].
Acanthocheilonema sabanicolae Eberhard and Campo-Aasen, 1986distinctly differs from A. delicata n. sp. and other congeners, becausethe anterior esophagus is longer than the posterior esophagus [46].Adult worms of A. setariosa (Mönnig, 1926) have a large and thickbody; the esophagus ratios are low (the sex of these worms was notindicated) [47].
The tail in the male of A. spirocauda terminates in four tubercles[48] and has been examined in detail by SEM [49] vis-à-vis threesalient conical lappets in our specimens. Adult worms of A. viteaehave a short esophagus and the male has a long left spicule [50].Adult worms of A. weissi (Seurat, 1914) have a short and thick body[51]; the microfilariae have a long body with a conspicuous cephalichook and a filamentous tail [52].
Regarding parasitic locations of adult worms and microfilariae ofAcanthocheilonema species in the host animals (Table 1), adult wormsof eight species including A. delicata n. sp. occur in the subcutaneousconnective tissues and the remaining species occur in various othertissues; microfilariae of eight species occur in the blood and onlythree species (A. delicata n. sp., A. procyonis, and A. sabanicolae) occurin the skin.
Therefore, we conclude that A. delicata n. sp. found from the Japanesebadgers can be distinguished from all the 14 Acanthocheilonema specieson the basis of morphologic characteristics and parasitic locations inthe host animals.
DNA sequences of the cox1 gene of A. delicata n. sp. indicated thatthe present new species was included in the clade of the genusAcanthocheilonema but distinguishable from two other congenericspecies, A. viteae and A. reconditum.
4. Discussion
Species of Cercopithifilaria and Onchocerca were also found to bemonophyletic, and clustered together; the cluster of these two generawas grouped with that of Acanthocheilonema (Figs. 8 and 9).Mansonella(C.) perforata was basal to the cluster consisting of three genera(Cercopithifilaria, Onchocerca, and Acanthocheilonema). F. martis wasfound to be basal to all of these filarioids. These findings obtained by
themolecular analysis established the genus Acanthocheilonema definedby morphologic characteristics [53].
Regarding the vectors, the louse-fly, fleas, and lice have beenrecorded to harbor Acanthocheilonema species [34,37,45]. In the pres-ent study, we did not find any potential vectors of this filarioid.Wolbachia endosymbionts were not detected in A. delicata n. sp. byimmunohistologic examination and PCR screening. The result is similarto the previous findings that the endosymbionts are not present inA. viteae and A. reconditum [19]. The loss of Wolbachia is likely to haveoccurred in the evolution of an ancestral species of Acanthocheilonemain Onchocercinae, because Wolbachia-like gene sequences were incor-porated in the filarial nuclear genomes of the endobacteria-free A. viteae[54]. Many species of the other filarioids in the subfamily possess theendosymbionts, with which they have coevolved since one infectionevent [29].
Chabaud and Bain [55] have suggested that the Acanthocheilonemalineage originated in Africa and expanded to the Oriental and Holarcticregions, and later to South America. From the present study, A. delicatan. sp. from Japanese badgers is closely similar to A. procyonis fromraccoons in the North America. The morphologic similarity has beenfound between Mansonella (Mansonella) akitensis (Uni, 1983) frombears in Japan and two congeneric American species,M. (M.) interstitium(Price, 1962) from the gray squirrel and M. (M.) llewellyni (Price, 1962)from the raccoon [25,56]. Dirofilaria ursi Yamaguti, 1941 has been foundfrom bears in Japan and North America [25]. These findings suggestthat the geographic expansion of filarioids has occurred by means ofthe migration of host animals from Eurasia towards North Americathrough the Bering Land Bridge [57].
Eurasian badgers are divided into four species according to theirmitochondrial DNA as well as morphologic characteristics: Meles melesin Europe, an unspecified species in Southwest Asia, M. leucurus inNorth and East Asia, and M. anakuma in Japan [58,59]. Suzuki [60]suggested that half of more than 100 species of mammals recordedfrom the Japanese islands are endemic andmaintain their own lineages.
We have examined 20 species of filarioids in five genera(Acanthocheilonema, Cercopithifilaria, Loxodontofilaria, Mansonella, andOnchocerca) from endemic mammals such as badgers, bears, boars,serows, and sika deer living on the Japanese Archipelago, and have todate found 15 new species (with A. delicata n. sp.) and one new subspe-cies; four other species (A. reconditum, A. spirocauda, A. odendhali, andO. skrjabini) were not endemic [3,4,7]. It is likely that the fauna offilarioids from mammals on the Japanese islands is characterized by ahigh level of endemicity.
Fig. 3. Histologic section of a female midbody of A. delicata n. sp. I: Intestine.Mf: Microfilaria. U: Uterus. * The cuticular thickening at the lateral field. Bar, micrometers.
20 S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
Figs. 4–7. SEM of males of A. delicata n. sp. 4. Deirid. 5. Striations of the midbody. 6. Area rugosa. 7. Lappets of the tail end. Phasmidial pore (arrow). Bars, 1 μm.
Table 2Distribution of microfilariae of Acanthocheilonema delicata n. sp. in skin snips of Japanese badgers with such microfilariae.
Badger ID no. Face Ears Neck Thoracic limbs Midback Abdomen Pelvic limbs Tail
Number of microfilariae: (−) not found; (+) 1–9; (++) 10–19; (+++) 20 or more. ND: not done.⁎ Adult worms were found.
21S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
Acknowledgements
We are grateful to Academician Dr. Yong Hoi Sen, Senior fellowof Academy of Sciences Malaysia, and Professor Emeritus, Universityof Malaya, who has warmly encouraged us in our research efforts.We heartily thank Professors Dr. Mohd Sofian Bin Azirun, Dean, theFaculty of Science, University of Malaya; Dr. Rosli Bin Hashim, Head,
the Institute of Biological Sciences, Faculty of Science, University ofMalaya; and Dr. Rosli Bin Ramli of the same institute, who likewisehave supported our study. We thank Mr. T. Kenko, the Central Labora-tory, Osaka City University Medical School for the histologic prepara-tion and Mr. H. Nakagawa of the same laboratory for SEMexamination of the parasites. We thank Mr. J. L. Yohay for readingthe manuscript.
Table 3Pairwise distance in amino acid sequences of the cox1 gene among 14 species of filarioids.
Species 1 2 3 4 5 6 7 8 9 10 11 12 13
1. A. delicata n. sp. (JQ289993) –
2. A. reconditum JF461456 0.102 –
3. A. viteae AJ272117 0.144 0.118 –
4. O. dewittei japonica AB18872 0.220 0.213 0.249 –
5. O. skrjabini AM74269 0.225 0.177 0.249 0.147 –
6. C. bulboidea AB178834 0.240 0.203 0.244 0.247 0.228 –
7. C. minuta AB178846 0.227 0.196 0.248 0.208 0.205 0.112 –
C. shohoi AB178850 C. bulboidea AB178834 C. crassa AB178840
C. multicauda AB178848 C. japonica AM749262
Cercopithifilaria
O. dewittei japonica AB18872 O. skrjabini AM74269
Onchocerca
A. reconditum JF461456 A. viteae AJ272117
A. delicata n. sp. (JQ289993)Acanthocheilonema
M. (Cutifilaria) perforata AM749265 F. martis AJ544880
82
74
62
66
61
69
0.05
C. bulboidea AB178834 C. crassa AB178840 C. shohoi AB178850
C. minuta AB178846 C. tumidicervicata AB178852
C. multicauda AB178848 C. japonica AM749262
Cercopithifilaria
O. dewittei japonica AB18872 O. skrjabini AM74269
Onchocerca
A. reconditum JF461456 A. viteae AJ272117
A. delicata n. sp. (JQ289993)
Acanthocheilonema
M. (Cutifilaria) perforata AM749265 F. martis AJ544880
70
64
6058
0.05
Figs. 8–9. Taxonomic position of A. delicata n. sp. with two other species of Acanthocheilonema. Trees are constructed by the JTTmatrix-basedmodel on the basis of amino acid sequencesof the cox1 gene among 14 species, usingMEGA5. 8. TheNJ tree (upper). 9. TheML tree (lower).A: Acanthocheilonema, C: Cercopithifilaria, F: Filaria,M:Mansonella, andO:Onchocerca. Bars,the number of changes inferred as having occurred along each branch.
22 S. Uni et al. / Parasitology International 62 (2013) 14–23
Author's personal copy
References
[1] Cobbold TS. Description of a new generic type of entozoon from the aard wolf(Proteles); with remarks on its affinities, especially in reference to the questionof parthenogenesis. Proceedings of the Zoological Society of London 1870;1:9–14.
[2] Yamaguti S. Studies on the helminth fauna of Japan. Part 35. Mammalian nema-todes II. Japanese Journal of Zoology 1941;9:409-39.
[3] Yagi K, Bain O, Shoho C. Onchocerca suzukii n. sp. and O. skrjabini (=O. tarsicola)from a relict bovid, Capricornis crispus, in Japan. Parasite 1994;1:349-56.
[4] Uni S, Suzuki Y, Baba M, Mitani N, Takaoka H, Katsumi A, et al. Coexistence of fiveCercopithifilaria species in the Japanese rupicaprine bovid, Capricornis crispus.Parasite 2001;8:197-213.
[5] Uni S, Bain O, Takaoka H. Affinities between Cutifilaria (Nematoda: Filarioidea),parasites of deer, and Mansonella as seen in a new onchocercid, M. (C.) perforatan. sp., from Japan. Parasite 2004;11:131-40.
[6] Uni S, BainO, AgatsumaT, KatsumiA, BabaM,Yanai T, et al. Newfilarial nematode fromJapanese serows (Naemorhedus crispus: Bovidae) close to parasites from elephants.Parasite 2006;13:193-200.
[7] Uni S, Bain O, Agatsuma T, Harada M, Torii H, Fukuda M, et al. Onchocerca eberhardin. sp. (Nematoda: Filarioidea) from sika deer in Japan; relationships between speciesparasitic in cervids and bovids in the Holarctic region. Parasite 2007;14:199-211.
[8] Pennington NE, Phelps CA. Canine filariasis on Okinawa, Ryukyu Islands. Journalof Medical Entomology 1969;6:59-67.
[9] Kagei N, Oda T. Dipetalonema odendhali Perry, 1967 found from fur seal, Callorhinusursinus (Linnaeus, 1758). Bulletin of the High Institute of Public Health 1975;24:203-5.
[10] Machida M. Two species of Dipetalonema from pinnipeds caught off Northern Japan.Bulletin of the National Science Museum, (Tokyo) Series A (Zoology) 1977;3:67-71.
[11] Huynh T, Thean J, Maini R. Depetalonema reconditum in the human eye. BritishJournal of Ophthalmology 2001;85:1384.
[13] Pampiglione S, Rivasi F. Human dirofilariasis due to Dirofilaria (Nochtiella) repens:an update of world literature from 1995 to 2000. Parassitologia 2000;42:231-54.
[14] Otranto D, Sakru N, Testini G, Gürlü VP, Yakar K, Lia RP, et al. Case report: first evi-dence of human zoonotic infection by Onchocerca lupi (Spirurida, Onchocercidae).The American Journal of Tropical Medicine and Hygiene 2011;81:55-8.
[15] Uni S, Bain O, Takaoka H, Miyashita M, Suzuki Y. Onchocerca dewittei japonican. subsp., a common parasite from wild boar in Kyushu Island, Japan. Parasite2001;8:215-22.
[16] Takaoka H, Bain O, Uni S, Korenaga M, Kozek WJ, Shirasaka C, et al. Zoonotic on-chocerciasis caused by a parasite from wild boar in Oita, Japan. A comprehensiveanalysis of morphological characteristics of the worms for its diagnosis. Parasite2004;11:285-92.
[17] Uni S, Boda T, Daisaku K, Ikura Y, Maruyama H, Hasegawa H, et al. Zoonotic filari-asis caused by Onchocerca dewittei japonica in a resident of Hiroshima Prefecture,Honshu, Japan. Parasitology International 2010;59:477-80.
[18] Fukuda M, Otsuka Y, Uni S, Bain O, Takaoka H. Molecular identification of infectivelarvae of three species of Onchocerca found in wild-caught females of Simuliumbidentatum in Japan. Parasite 2010;17:39-45.
[19] Taylor MJ, Bandi C, Hoerauf A. Wolbachia bacteria endosymbionts of filarial nem-atodes. Advances in Parasitology 2005;60:245-84.
[20] Uni S, Bain O, Takaoka H, Katsumi A, Fujita H, Suzuki Y. Diversification ofCercopithifilaria species (Nematoda: Filarioidea) in Japanese wild ruminantswith description of two new species. Parasite 2002;9:293-304.
[21] Chabaud AG, Petter AJ. Remarques sur l'évolution des papilles cloacales chez lesnématodes phasmidiens, Parasites de Vertébrés. Parassitologia 1961;3:51-70.
[22] Agatsuma T, Iwagami M, Uni S, Takaoka H, Katsumi A, Kimura E, et al. Molecularphylogenetic relationships among seven Japanese species of Cercopithifilaria.Parasitology International 2005;54:195-9.
[23] Krepkogorskaja TA. Beitrag zur Fauna der Nematoden aus Rhombomys opimusLicht. aus Kasakstan. Zoologischer Anzeiger 1933;102:87-91.
[24] Uni S, Suzuki Y, Katsumi A. Cercopithifilaria shohoi n. sp. (Nematoda: Filarioidea)from the relict Bovidae, Capricornis crispus, in Japan. Parasite 1998;5:119-26.
[25] Uni S. Filarial parasites from the black bear of Japan. Annales de ParasitologieHumaine et Comparée 1983;58:71-84.
[26] Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecularevolutionary genetics analysis using maximum likelihood, evolutionary distance,and maximum parsimony methods. Molecular Biology and Evolution 2011;28:2731-9.
[27] Kramer LH, Passeri B, Corona S, Simoncini L, Casiraghi M. Immunohistochemical/immunogold detection and distribution of the endosymbiontWolbachiaofDirofilariaimmitis and Brugia pahangi using a polyclonal antisirum raised against WSP(Wolbachia surface protein). Parasitology Research 2003;89:381-6.
[28] Casiraghi M, Bain O, Guerrero R, Martin C, Pocacqua V, Gardner SL, et al. Mappingthe presence ofWolbachia pipientis on the phylogeny of filarial nematodes: evidencefor symbiont loss during evolution. International Journal of Parasitology 2004;34:191-203.
[29] Ferri E, Bain O, BarbutoM,Martin C, Lo N, Uni S, et al. New insights into the evolutionofWolbachia infections in filarial nematodes inferred from a large range of screenedspecies. PLoS One 2011;6:e20843, http://dx.doi.org/10.1371/journal.pone.0020843.
[30] Chabaud AG, Bain O. La lignée Dipetalonema. Nouvel essai de classification.Annales de Parasitologie Humaine et Comparée 1976;51:365-97.
[31] Anderson RC, Bain O. No. 3. Keys to genera of the Order Spirurida, Part 3.Diplotriaenoidea, Aproctoidea and Filarioidea. In: Anderson RC, Chabaud AG, WillmottS, editors. CIH Keys to the nematode parasites of vertebrates; 1976. p. 59–116.
[32] Bain O, Baker M, Chabaud AG. Nouvelles données sur la lignée Dipetalonema(Filarioidea, Nematoda). Annales de Parasitologie Humaine et Comparée 1982;57:593-620.
[33] Rioche M. Présence de Dipetalonema dracunculoides (Cobbold, 1870) chez le chiendans la région d'Alger. Archives de l'Institut Pasteur d'Algérie 1960;38:386-98.
[34] Nelson GS. Dipetalonema dracunculoides (Cobbold, 1870), from the dog in Kenya:with a note on its development in the louse-fly, Hippobosca longipennis. Journal ofHelminthology 1963;37:235-40.
[35] Kou CC. Studies on parasitic nematodes of mammals from Canton. I. Some newspecies from Paradoxurus minor exitus Schwarz, Paguma larvata larvata (HamiltonSmith) and Manis pentadactyla aurita Hodgson. Acta Zoologica Sinica, Peking1958;10:60-72with 3 plates, (60–67 in Chinese).
[36] Esslinger JH. Dipetalonema fausti sp. n. (Filarioidea: Onchocercidae), a filarial par-asite of the scaly anteater, Manis pentadactyla L. (Pholidota), from China. Journalof Parasitology 1966;52:494-7.
[37] Nelson GS. On Dipetalonema manson-bahri n. sp., from the spring-hare, Pedetessurdaster larvalis, with a note on its development in fleas. Journal of Helminthology1961;35:143-60.
[38] Webster WA, Beauregard M. Dipetalonema mephitis n. comb. (=Microfilariamephitis: Webster and Beauregard, 1964) from the skunk, Mephitis mephitis.Canadian Journal of Zoology 1965;43:325-32.
[39] Perry ML. A new species of Dipetalonema from the California sea lion and a report ofmicrofilaria from a steller sea lion (Nematoda: Filarioidea). Journal of Parasitology1967;53:1076-81.
[40] Perry ML, Forrester DJ. Dipetalonema odendhali (Nematoda: Filarioidea) from thenorthern fur seal, with a description of the microfilaria. Journal of Parasitology1971;57:469-72.
[41] Ortlepp RJ. On two rictularias and a filariid from South African wild carnivores.Journal of Helminthology 1961;35:131-40.
[42] Esslinger JH, Smith JL. Dipetalonema (Acanthocheilonema) didelphis sp. n. (Nematoda:Filarioidea) from opossums, with a redescription of D. (A.) pricei (Vaz and Pereira1934). Journal of Parasitology 1979;65:928-33.
[43] Price DL. Dipetalonema procyonis n. sp. from Procyon lotor lotor (Linnaeus). Pro-ceedings of the Helminthological Society of Washington 1955;22:38-41.
[44] Smith JL. Redescription of Dipetalonema (Acanthocheilonema) procyonis Price 1955(Nematoda: Filarioidea) from the raccoon. Journal of Parasitology 1980;66:333-6.
[45] Nelson GS. Dipetalonema reconditum (Grassi, 1889) from the dog with a note onits development in the flea, Ctenocephalides felis and the louse, Heterodoxusspiniger. Journal of Helminthology 1962;36:297-308.
[46] Eberhard ML, Campo-Aasen I. Acanthocheilonema sabanicolae n. sp. (Filarioidea:Onchocercidae) from the savanna armadillo (Dasypus sabanicola) in Venezuela,with comments on the genus Acanthocheilonema. Journal of Parasitology 1986;72:245-8.
[47] Mönning HO. Helminthological notes. 11th and 12th Reps Dir Vet Ed Res. S. Afr:Dept Agric; 1926. p. 221-8.
[48] Anderson RC. The taxonomy of Dipetalonema spirocauda (Leidy, 1858) n. comb.(=Skrjabinaria spirocauda) and Dirofilaria roemeri (Linstow, 1905) n. comb.(=Dipetalonema roemeri). Canadian Journal of Zoology 1959;37:481-93.
[49] Leidenberger S, Boström S. Characterization of the heartworm Acanthocheilonemaspirocauda (Leidy, 1858) Anderson, 1992 (Nematoda:Onchocercidae) in Scandinavia.Parasitology Research 2008;104:63-7.
[50] Chabaud AG. Le genre Dipetalonema Diesing 1861; Essai de classification. Annalesde Parasitologie Humaine et Comparée 1952;27:250-85.
[51] Seurat LG. Sur une filaire péritonéale du Macroscélide. Comptes Rendus de laSociété de Biologie 1914;77:524-6.
[52] Bain O, Quentin JC. Dévelopment de Dipetalonema (A.) weissi, filaire deMacroscélide,chez un Ornithodore. Annales de Parasitologie Humaine et Comparée 1977;52:569-75.
[53] Bain O, Casiraghi M, Martin C, Uni S. The Nematoda Filarioidea: critical analysislinking molecular and traditional approaches. Parasite 2008;15:342-8.
[54] McNutty SN, Foster JM, Mitreva M, Dunning Hotopp JC, Martin J, Fischer K, et al.Endosymbiont DNA in endobacteria-free filarial nematodes indicates ancienthorizontal genetic transfer. PLoS One 2010;5:e11029.
[55] Chabaud AG, Bain O. The evolutionary expansion of the Spirurida. InternationalJournal of Parasitology 1994;24:1179-201.
[56] Eberhard ML, Orihel TC. The genusMansonella (syn. Tetrapetalonema): a new clas-sification. Annales de Parasitologie Humaine et Comparée 1984;59:483-96.
[57] Bain O. Evolutionary relationships among filarial nematodes. In: Klei TR, Rajan TV,editors. World class parasites. Dordrecht, The Netherlands.: Kluwer AcademicPublishers; 2002. p. 21-9.
[58] Marmi J, López-Giráldez F, Macdonald DW, Calafell F, Zholnerovskaya E,Domingo-Roura X. Mitochondrial DNA reveals a strong phylogeograhic structurein the badger across Eurasia. Molecular Ecology 2006;15:1007-20.
[59] Kaneko Y.Meles anakuma Temminck, 1844. In: Ohdachi SD, Ishibashi Y, Iwasa MA,Saitoh T, editors. The wild mammals of Japan. Kyoto: Shoukadou; 2009. p. 258-60.
[60] SuzukiH. Amolecular phylogenetic viewofmammals in the “three-storymuseum” ofHokkaido, Honshu, and Ryukyu Islands, Japan. In: Ohdachi SD, Ishibashi Y, Iwasa MA,Saitoh T, editors. The wild mammals of Japan. Kyoto: Shoukadou; 2009. p. 261-3.
[61] Anderson RC. Nematode parasites of vertebrates. Their development and transmis-sion. Wallingford Oxon UK: CABI Publishing, CAB International; 2000. p. 494-6.
[62] Sonin MD. Filariata of Animals and man and diseases caused by them, Part 3,Filariidae, Onchocercinae. Fundamentals of Nematology, vol. 24. Moscow: NaukaPublishers; 1975 (Translated from Russian, published for the US Department ofAgriculture, and the National Science Foundation, Washington, D.C., by AmerindPublishing Co., Pvt. Ltd., New Delhi, 1985. p. 273–8).
23S. Uni et al. / Parasitology International 62 (2013) 14–23