Page 1
This article is protected by copyright. All rights reserved.
Autocrine and immune cell derived BDNF in human skeletal muscle:
implications for myogenesis and tissue regeneration
Emanuela Colombo1, Francesco Bedogni2, Isabella Lorenzetti1, Nicoletta Landsberger2,3,
Stefano C. Previtali1 and Cinthia Farina1*.
1Institute of Experimental Neurology (INSpe), Division of Neuroscience, San Raffaele
Scientific Institute, Milan, Italy
2 San Raffaele Rett Research Center, San Raffaele Scientific Institute, Milan, Italy
3 Laboratory of Genetic and Epigenetic Control of Gene Expression, Department of
Theoretical and Applied Sciences, University of Insubria, Busto Arsizio, Italy
∗ Corresponding author: Institute of Experimental Neurology (INSpe), Division of
Neuroscience, San Raffaele Scientific Institute, Via Olgettina, 58, 20132 Milan – Italy.
Phone.: +39 02 2643 6180 E-mail: [email protected]
No conflicts of interest to be declared
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1002/path.4228
Page 2
This article is protected by copyright. All rights reserved.
Abstract
The neurotrophin system has a role in skeletal muscle biology. Conditional depletion of
BDNF in mouse muscle precursor cells alters myogenesis and regeneration in vivo. However,
the expression, localisation and function of BDNF in human skeletal muscle tissue is not
known so the relevance of the rodent findings for human muscle are unknown. Here we
address this by combining ex vivo histological investigations on human biopsies with in vitro
analyses of human primary myocytes. We found that BDNF was expressed by precursor and
differentiated cells both in vitro and in vivo. Differential analysis of BDNF receptors showed
expression of p75NTR and not of TrkB in myocytes, suggesting that the BDNF-p75NTR axis
is predominant in human skeletal muscle cells. Several in vitro functional experiments
demonstrated that BDNF gene silencing or protein blockade in myoblast cultures hampered
myogenesis. Finally, histological investigations of inflammatory myopathy biopsies revealed
that infiltrating immune cells localised preferentially near p75NTR positive regenerating
fibers and that they produced BDNF. In conclusion, BDNF is an autocrine factor for skeletal
muscle cells and may regulate human myogenesis. Furthermore, the preferential localisation
of BDNF producing immune cells near p75NTR positive regenerating myofibers suggests
that immune cell derived BDNF may sustain tissue repair in inflamed muscle.
Keywords: BDNF, inflammation, human skeletal muscle, neurotrophin, myogenesis, immune
cells
Page 3
This article is protected by copyright. All rights reserved.
Introduction
Postnatal growth and repair of skeletal muscle after damage require the activation of satellite
cells, resident muscle precursor cells located between the cell membrane and the basal lamina
of muscle fibers [1]. Several regulatory factors control the myogenic properties of satellite
cells and thereby muscle regeneration. For example, while IGF-1, FGF and VEGF are
positive regulators of myogenesis, TGF-β and myostatin reduce myoblast differentiation
[2,3]. Moreover, TGF-β is a potent regulator of extracellular matrix deposition, thus favoring
fibrosis [2,3].
Recent evidence suggests a role for neurotrophins (NT) in skeletal muscle physiology and
repair. The family of neurotrophins consists of nerve growth factor (NGF), NT-3, NT-4 and
brain derived neurotrophic factor (BDNF). Although initially described as factors specific for
the central nervous system [4], NT have been detected also in skeletal muscle where they
regulate motor neuron survival, enhance presynaptic release of neurotransmitter and promote
maintenance of postsynaptic region on skeletal myofibers [5-9]. Two different types of
receptors mediate neurotrophin signaling: the tropomyosin related kinase (Trk) receptors and
the p75 neurotrophin receptor (p75NTR) [4]. Recently, attention has been devoted to
p75NTR. A study from our group demonstrated that p75NTR is a marker for human skeletal
muscle precursor cells at high differentiation potential, and that it positively regulates
myogenesis and myofiber maturation [10]. Furthermore, blockade of p75NTR signaling in
vivo in an injury mouse model hampers muscle regeneration [11].
Regarding p75NTR ligands, some studies suggest the expression of all neurotrophins in
skeletal muscle [12-15], albeit the sources of neurotrophins in human tissues have not been
clearly characterized. Here we focused our attention on BDNF, as low levels of BDNF are
Page 4
This article is protected by copyright. All rights reserved.
detected in developing and postnatal avian and rodent skeletal muscle [14,16-19], and its
production is maintained by myogenic progenitor cells in adult rodent muscle [20-23].
Further, BDNF transcript levels increase in rat muscle after exercise [24,25] and after
denervation [12,26], indicating that BDNF expression may be regulated in skeletal muscle.
Beyond its function as a retrograde trophic factor supporting motor neuron survival and
neuromuscular transmission [8,19], BDNF may support skeletal muscle regeneration. In fact,
transgenic mice carrying selective depletion of BDNF in skeletal muscle precursor cells
display reduced numbers of regenerating fibers after injury [27]. However, implications of
these findings for the human skeletal muscle physiology are unknown as no studies
performed an accurate characterization of BDNF sources in human skeletal muscle.
Furthermore, it is still not clear whether human pathological conditions are associated with
changes in BDNF levels or distribution. An in situ hybridization study on post-mortem
tissues showing enhanced BDNF mRNA in amyotrophic lateral sclerosis muscle suggests
that expression of the neurotrophin may change in human skeletal muscle under disease [28].
Finally, the evidence of a role for this neurotrophin in human skeletal muscle biology is not
yet available. The aim of our study was therefore to assess BDNF expression and localization
in striated muscle under physiological and pathological conditions and to illustrate its
involvement in human myogenesis.
Methods
Patients and tissues
Muscle biopsies were performed for diagnostic reasons and stored in the institutional tissue
bank. Informed consent for biopsy and its storage for research purposes was obtained in all
cases. Biopsies were taken from quadriceps femoris (n=19), deltoid (n=3), biceps (n=2) or
rectus femoris (n=1) muscles. We selected specimens with clear diagnosis, based on clinical,
electromyographic and histological findings [29,30]. A total of 17 patients suffered from
Page 5
This article is protected by copyright. All rights reserved.
idiopathic inflammatory myopathies (IIM): polymyositis n=8, dermatomyositis n=4 or
inclusion body myositis n=5 (mean age 65±12, 10 male and 7 female subjects). Finally, 8
muscle specimens from adult individuals (mean age 37±13, 6 male and 2 female subjects)
initially suspected of neuromuscular disease whose biopsies were histologically normal were
included as controls.
Myoblast cell culture
Human primary myoblasts were selected for CD56/NCAM expression, cultured and
differentiated as previously described [10,31,32]. Fusion index was calculated as the
percentage of myonuclei within myotubes (with more than two nuclei) on total nuclei
number. Each experiment was performed in triplicates, each figure shows representative data
out of minimum three performed experiments. Data were confirmed in two primary cell lines.
Immunohistochemistry and double immunofluorescence
The following primary antibodies were used: two distinct anti-human BDNF, anti-human
NGFR, anti-human TrkB (all from R&D Systems, Milan, Italy), anti-human CD56 (BD
Biosciences, Buccinasco, Italy), anti-human CD68, anti-human CD4, anti-human CD8, anti-
human desmin (all from Dako, Milan, Italy), anti-human CD3, anti-Ki67 (both from Lab
Vision, Fremont, California), anti-human perforin (BD Biosciences), anti-dystrophin
(provided by Dr. Mora), purified mouse IgG1 isotype (Sigma-Aldrich, Gallarate, Italy),
polyclonal rabbit Ig (Dako). Detection was perfomed as in [10,31].
The Zenon technique was used to perform double staining with multiple mouse monoclonal
antibodies according to the manufacturer’s protocol (Life Technologies, Monza, Italy).
Page 6
This article is protected by copyright. All rights reserved.
Fluorescence images were captured with confocal laser-scanning microscopes (Leica
Microsystems, Milan, Italy). Images from immunohistochemistry sections were acquired
using light microscopes (Leica Microsystems). The ImageProPlus software (Media
Cybernetics, Silver Spring, MD, USA) was used for image analysis. For the quantification of
immune cell distribution, firstly we assessed the total number of regenerating (CD56
positive) or mature (CD56 negative) myofibers in each tissue section. Mature fibers in
contact with regenerating fibers were not included in the quantification. Infiltrating immune
cells were identified as basophilic small nuclei by haematoxylin staining. Nuclei positioned
within myofibers were not considered for the quantification. Then, the number of immune
cells localized in the proximity of regenerating or mature myofibers was calculated as the
ratio between the number of immune cells and the total number of regenerating or mature
myofibers respectively.
In Situ Hybridization
Digoxygenin BDNF exon 5 in situ was performed as previously described [33]. After pre-
hybridization buffer, sections were hybridized overnight at 60°C with a a proper dilution of
riboprobe in hybridization buffer. Sections were then washed four times at 60°C in Washing
Solution, blocked in Blocking Solution at r.t. (Roche, Monza, Italy) then incubated overnight
at 4°C in 1:1000 Anti-DIG (Roche). After several washings and brief incubation in
Levamisole (0,1M, Sigma Aldrich), BCIP/NBT colorimetric reaction (Roche) was then
carried out in the dark at r.t.
Page 7
This article is protected by copyright. All rights reserved.
Flow cytometry
Labelling with mouse monoclonal anti-human TrkB (R&D Systems) or with the
corresponding isotype controls (BD Biosciences) was followed by detection with PE-labeled
F(ab’)2 fragments goat anti-mouse Ig (Dako). Cytofluorimetric analyses were performed on
FACSCalibur (BD Biosciences). CellQuest software (BD Biosciences) was used for
acquisition and FlowJo (Tree Star Inc, Ashland, Oregon, USA) for data analysis. Thresholds
for positivity were set on isotype control.
BDNF RNA interference
Small interfering RNA fragments (siRNA) for BDNF and non-specific control (47% GC
content) were purchased at Eurofins MWG (M-Medical, Cornaredo, Italy). siRNA were
diluted at 20nM in Optimem (Life Technologies). Transfection was obtained by Interferin
(Polyplus, Celbio, Milan, Italy). Three days after siRNA transfection myoblasts were induced
to differentiate and silencing efficiency was monitored at day 3 after differentiation
induction.
Treatment with anti-BDNF blocking antibody
Myoblasts were seeded in 4-well chamber slides and induced to differentiate for 7 days in
medium supplemented with 10μg/ml rabbit polyclonal anti-human BDNF blocking antibody
(Millipore, Vimodrone, Italy) [34] or purified rabbit Ig (Dako). Antibody treatment was
repeated for three consecutive days.
RNA extraction, cDNA synthesis and Real-Time PCR
RNA extraction, cDNA synthesis and Real time PCR were performed as in [34]. To remove
contaminating DNA, RNA was treated with DNase I enzyme (Life Technologies).
Page 8
This article is protected by copyright. All rights reserved.
Amplification sets for BDNF, p75NTR, TrkB were described in [35]. mRNA levels of target
genes were graphically reported as percentage of the housekeeping gene PPIA (Life
Technologies).
Statistical Analysis
Normality of the distribution was assessed by Kolmogorov–Smirnov statistics. Student’s T-
test was used to compare means. All P-values were two-sided and subjected to a significance
level of 0.05.
Results
BDNF is strongly expressed in myoblasts and in differentiating cells in vitro
First, we analyzed BDNF expression in primary cultures of human muscle precursor cells, the
myoblasts. Quantitative RT-PCR revealed robust transcript levels for the neurotrophin in
these undifferentiated cells under normal growth conditions (Figure 1A, first column).
Double immunofluorescence performed with a specific antibody showed BDNF protein in all
CD56/NCAM positive myoblasts (Figure 1B). Staining with another anti-BDNF antibody
confirmed this observation (not shown). Then, we induced myoblast differentiation and
fusion into multinucleated myotubes by the addition of specific culture medium, and
monitored BDNF mRNA levels over two weeks. Neurotrophin expression remained high
during the first 10 days and slightly increased afterwards (Figure 1C). Double
immunofluorescence confirmed the presence of BDNF in cultured myotubes (Figure 1D). As
BDNF action may be mediated by two distinct receptors, we investigated the expression of
TrkB and p75NTR at mRNA levels in parallel to BDNF. As already described [10], p75NTR
mRNA was expressed in undifferentiated myoblasts (Figure 1A, second column) and
increased during myogenesis (Figure 1C). In contrast, TrkB transcript levels were barely
Page 9
This article is protected by copyright. All rights reserved.
detectable both in myoblasts and in differentiated cultures (Figure 1A third column and
Figure 1C). Moreover, cytofluorimetric and double immunofluorescence experiments
confirmed the lack of TrkB at protein level in cultured myoblasts (Figure 1E-F). Therefore
human muscle cells produce BDNF in vitro and may respond to it through p75NTR.
BDNF is displayed by satellite cells and mature myofibers in vivo
As p75NTR is expressed in satellite cells in vivo [10] and myoblasts strongly produced
BDNF in vitro, we checked expression of the neurotrophin in satellite cells in situ. Double
immunofluorescence on non-myopathic tissues showed that satellite cells stained positive for
BDNF and p75NTR (Figure 1G-H). Accurate quantification revealed that more than 80% of
CD56/NCAM-positive satellite cells expressed the neurotrophin (Figure 1I). Furthermore, in
accordance with in vitro observations on myotubes, in situ hybridization and
immunohistochemistry experiments on skeletal muscle tissues showed that all mature
myofibers were positive for BDNF (Figure 1J-K). In contrast, the receptor TrkB was not
displayed by myofibers but its expression was restricted to some vessels (Figure 1L, black
arrow). These in vitro and in vivo observations suggest that BDNF may play a physiological
role in human skeletal muscle and that p75NTR is the only available receptor for BDNF
signalling in human muscle precursor cells.
BDNF regulates human myogenesis in vitro
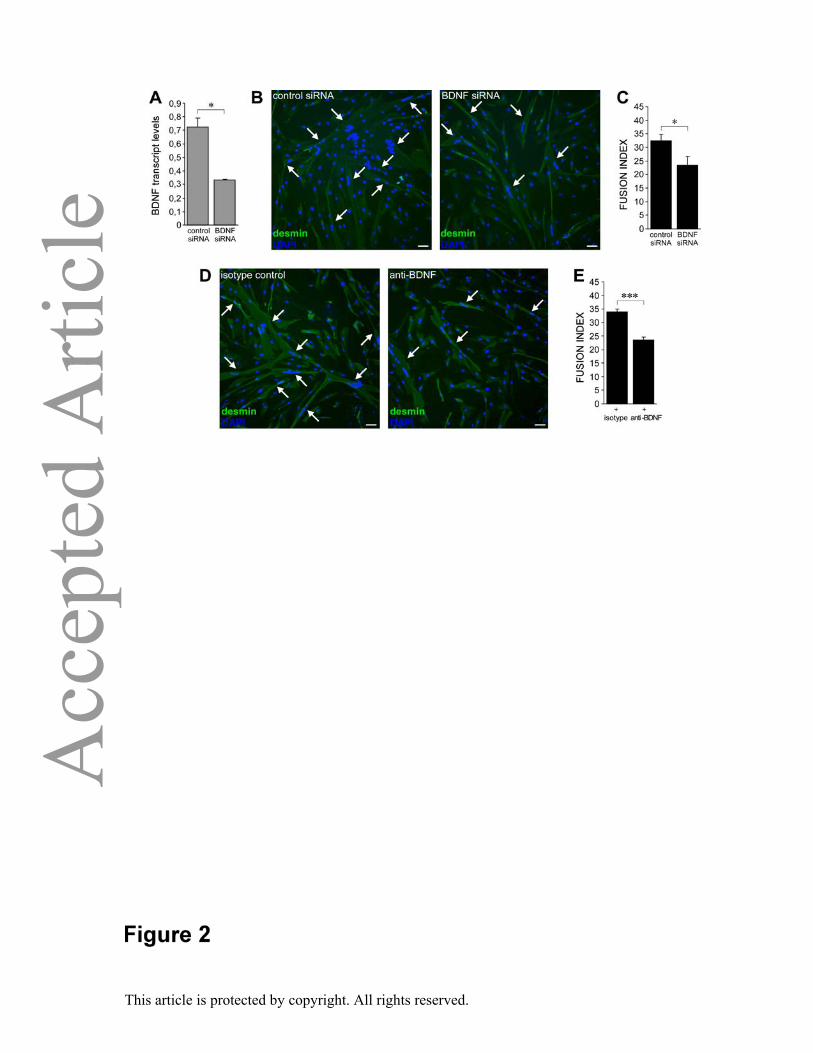
We performed functional in vitro studies to evaluate the involvement of BDNF in human
myogenesis. For this purpose we induced myoblast differentiation while hampering BDNF
expression or activity. First, we reduced BDNF mRNA availability by specific RNA
Page 10
This article is protected by copyright. All rights reserved.
interference in undifferentiated myoblasts (0.33±0.01 vs. 0.72±0.07 in BDNF-silenced and
control cultures respectively ; p value = 0.011; degrees of freedom = 2; t Stat = 9.48; t
Critical = 4.3; Figure 2A), and then induced in vitro differentiation for 6 days. Importantly,
myoblasts transfected with specific BDNF siRNA showed lower fusion index than control
cultures (23.3±3.3 vs. 32.4±2.4 in BDNF-silenced and control cultures respectively ; p value
= 0.02; degrees of freedom = 4; t Stat = 3.79; t Critical = 2.78; Figure 2B-C). Second, we
blocked BDNF protein activity in differentiating cultures by adding a specific neutralizing
antibody [34]. Parallel cultures where myoblasts were induced to differentiate in the presence
of an isotype antibody were used as controls. At day 6 myogenesis was significantly reduced
in cultures exposed to the anti-BDNF antibody (23.6±0.9 vs. 34±1 in anti-BDNF antibody-
treated and control cultures respectively; p value = 0.0002; degrees of freedom = 4; t Stat =
12.98; t Critical = 2.78; Figure 2D-E). In conclusion, the functional assays showed that
blockade of BDNF either at transcript or at protein level impairs human myogenesis.
Immune cells in inflammatory myopathies are preferentially located near regenerating
fibers and produce BDNF
We extended analysis for BDNF to muscle biopsies from patients affected by idiopathic
inflammatory myopathies (IIM). First of all, we checked BDNF transcript levels in healthy
and diseased tissues by quantitative PCR experiments and found detectable and comparable
expression of BDNF in the two groups (0.97±0.21 in control samples vs. 0.78±0.16 in IIM; p
value = 0.82; Figure 3A). Then, immunohistochemical experiments in pathological tissues
confirmed that mature (CD56 negative, asterisks in Figure 3B-E) myofibers were positive for
BDNF under inflamed settings. Moreover a number of CD56 positive regenerating fibers
(triangles, Figure 3B-E) displayed enhanced BDNF immunoreactivity. Importantly, the same
fibers expressed p75NTR (triangles, Figure 3D), suggesting that autocrine usage of BDNF
Page 11
This article is protected by copyright. All rights reserved.
supports in vivo regeneration. Further, as expected, inflamed tissues were characterized by
massive immune cell infiltration. We noticed that immune cells were not homogenously
distributed within the tissue, but were mostly organized into clusters. Such clusters were
topologically juxtaposed near CD56 positive regenerating fibers (Figure 3F-G). To quantify
this observation we counted the number of immune cells in the vicinity of regenerating fibers
or of mature myofibers in 8 biopsies from IIM patients. Such analysis confirmed that immune
cells were preferentially located near newly forming fibers (5.7±4.7 immune
cells/regenerating fiber vs. 1±1.7 immune cells/mature fiber; p value = 0.027; degrees of
freedom = 9; t Stat = 2.64; t Critical = 2.26; Figure 3H). IIM are considered autoimmune
disorders of skeletal muscle [36]. It was in fact shown that some immune cells, mainly
located within myofibers, are activated and secrete cytolytic factors [37]. To further
characterize the inflammatory infiltrates in inflamed muscle we checked the expression of the
proliferation marker Ki-67 and of the cytolytic factor perforin. We found that perforin
releasing T cells were extremely rare, located near mature myofibers and not in the vicinity of
CD56 positive regenerating fibers (Supplementary Figure 1A-B). Similar distributions were
observed for Ki-67-positive macrophages (Supplementary Figure 1C-D) or T cells (not
shown). In contrast, immunohistochemistry for BDNF revealed several immune cells strongly
positive for the neurotrophin (white arrows, Figure 3E) and frequently localized near
p75NTR-expressing regenerating myofibers (triangles, Figure 3C-D). Accurate quantification
of this observation confirmed that, while immune cells located near mature myofibers rarely
showed staining for the neurotrophin, those cells in the vicinity of regenerating fibers often
expressed BDNF (1.37±1.39 BDNF positive immune cells/regenerating fiber vs. 0.021±0.018
BDNF positive immune cells/mature fiber; p value = 0.029; degrees of freedom = 7; t Stat =
2.74; t Critical = 2.36; Figure 3I). Fine histological characterization by double
immunofluorescence and confocal microscopy showed that BDNF was produced by T
Page 12
This article is protected by copyright. All rights reserved.
lymphocytes (both CD4+ and CD8+ cells Figure 3J-K) and macrophages (Figure 3L) but not
by B cells (not shown). In summary, in inflamed tissue both myocytes and immune cells
contribute to BDNF production. The preferential distribution of BDNF producing immune
cells around p75NTR positive regenerating fibers strongly suggests that such immune cells
are not pathogenic but rather sustain skeletal muscle repair.
Discussion
In this study we identified the neurotrophin BDNF as a factor endogenously produced by
both skeletal muscle precursor cells and mature myofibers that may positively regulate
human myogenesis. In IIM several immune cells infiltrating skeletal muscle may contribute
to BDNF production and preferentially accumulate near p75NTR positive regenerating fibers.
Furthermore, BDNF positively regulates human myogenesis, identifying this factor as a
candidate ligand triggering p75NTR action in human skeletal muscle.
Human skeletal muscle regeneration requires the activation of satellite cells which is
regulated by stimuli derived from the microenvironment, such as growth factors. Recent
reports on rodent muscle highlight the role of the neurotrophin BDNF in this process [20,27].
In fact, BDNF is expressed in rodent satellite cells [20] and regulates myogenesis in vivo
[27]. In our study the fine characterization of the sources of BDNF in human skeletal muscle
revealed that this neurotrophin is expressed by both precursor and differentiated cells.
Quantification of satellite cells expressing the neurotrophin showed that the great majority of
precursors is BDNF positive. This observation is in harmony with the in vivo expression of
p75NTR. Consistent with in vivo data, high levels of BDNF were found in primary myoblasts
and in multinucleated myotubes in vitro. Regarding the receptor TrkB, it has been detected at
mRNA or protein level in total extracts from animal muscle [12,14,24], however its presence
in muscle cells has not been described. In contrast, p75NTR is expressed by skeletal muscle
Page 13
This article is protected by copyright. All rights reserved.
and regulates myogenesis via modulation of RhoA-GTP signaling in differentiating cells
[10,11,31]. In fact, we described recently that p75NTR positive precursor cells display a
repertoire of gene products that accounts for their enhanced differentiation properties and that
p75NTR blockade either at the transcript or protein level impairs myogenesis [10]. However,
the precise downstream signaling activated by NT binding to p75NTR in myocytes remains
largely unknown, and future studies are needed to resolve this issue.
Based on our data shown here and in previous publications [10,31,32], the receptor TrkB is
absent in human myocytes in vitro and in vivo, while p75NTR appears on BDNF-expressing
satellite cells and regenerating fibers as well as in cultured cells. These evidences indicate
that the neurotrophin BDNF is commonly produced by human muscle cells and that, due to
the expression of p75NTR, muscle cells may be not only the source but also the target of
BDNF. As the neurotrophin receptor p75NTR is a key regulator of human myogenesis [10],
we hypothesized that myoblast responses to BDNF could modulate skeletal muscle cell
differentiation. Functional experiments in the in vitro model of myocyte differentiation
showed that the reduction in BDNF transcript levels or in protein availability hampered cell
fusion. This is in accordance with the in vivo data obtained in a transgenic mouse model
where BDNF depletion was achieved in muscle precursor cells [27].
Overall, these observations demonstrate that BDNF is essential for skeletal muscle
differentiation and that its action is presumably mediated by p75NTR. A limitation of the
study is that the molecular mechanisms triggered by BDNF during myogenesis have not been
characterized and require further investigations.
The characterization of muscle tissues from patients affected by IIM for BDNF protein
expression led to novel additional observations. First of all, mature myofibers consistently
expressed BDNF also in IIM. Interestingly, we described recently upregulation of p75NTR in
Page 14
This article is protected by copyright. All rights reserved.
mature muscle fibers in IIM [31]. In vitro blocking experiments of p75NTR expression by
RNA interference in myotubes showed that p75NTR-silenced myotubes were more
susceptible to apoptosis after exposure to IL-1 than control myotubes [31]. These evidences
suggest that BDNF-p75NTR axis activates a tissue protective response. Moreover, we
showed here that in IIM immune cells are not evenly distributed within the tissue, but that
they accumulate in regenerating areas. This might indicate an exacerbated immune response
against newly-forming fibers. However, these immune cells were not in a proliferative state
or did not produce cytolytic factors, dampening the hypothesis of a pathogenic immune
response in such regions.
Opposed to the evidence about the involvement of immunity in the etiopathogenesis of
inflammatory myopathies, some authors demonstrated a crosstalk between immune and
muscle cells that positively regulates muscle homeostasis and repair. For example,
macrophages seem to play a primary role in muscle regeneration as they may regulate re-
activation of muscle satellite cells. An in vitro study showed that human myoblasts attract
monocytes producing specific chemoattractants such as monocyte chemoattractant protein-1,
macrophages-derived chemokine and VEGF, and that co-culture with macrophages increases
myoblast proliferation [38]. Further, after cardiotoxin-induced damage, skeletal muscle
regeneration is severely impaired if mice are treated with a specific antibody blocking
macrophage infiltration into the damaged tissues. In particular, muscles show enhanced
fibrosis, low numbers of satellite cells and of newly forming myotubes [39], indicating a clear
role for macrophages in sustaining muscle regeneration after injury. Different activation
states for macrophages have been described in vitro [40]. Classical activation (M1) is
characterized by production of proinflammatory cytokines and represents the primary
response of macrophages to tissue injury. Interestingly, macrophages with an inflammatory
phenotype are found in human dystrophic muscle [41], where they are prominently located in
Page 15
This article is protected by copyright. All rights reserved.
necrotic foci [42]. Conversely, the anti-inflammatory profile (M2) has been associated with
muscle repair [41,43].
Our data are consistent with a model where BDNF acts as a crosstalk factor between immune
and muscle cells. We demonstrated the preferential accumulation of BDNF-releasing immune
cells near regenerating fibers. Immunofluorescence experiments combined with confocal
microscopy revealed that both (CD4+ and CD8+) T lymphocytes and macrophages
contributed to BDNF production. These findings are not surprising as BDNF production by
human thymocytes and circulating immune cells has been previously described [35,44,45]. It
becomes important however to consider that the localization of BDNF producing immune
cells was preferentially within regeneration areas and that such cells were not activated and
did not produce cytolytic factors. These observations support the hypothesis that protective
immune responses may also take place in inflamed skeletal muscle and that BDNF may be a
mediator for regeneration.
Acknowledgements
We acknowledge the EurobioBank and the Italian Telethon Network of Genetic Biobanks
(GTB07001F) for providing myoblast cell lines, and Dr. M. Mora from Neurological Institute
Carlo Besta for anti-dystrophin antibody. This work was supported by the Association
Française contre les Myopathies (AFM).
Author contributions
EC carried out experiments, analysed and interpreted the data, wrote the manuscript. FB
carried out ISH experiments. IL and NL contributed reagents/material/analysis tool. SP
provided tissue specimens. CF designed the study, conceived the experiments, analysed and
interpreted the data, wrote the manuscript. All authors approved the submitted manuscript.
Page 16
This article is protected by copyright. All rights reserved.
Online Supporting Information
Figure S1. Perforin expressing T cells or Ki67 positive macrophages are localized near
mature fibers.
References
1. Collins CA. Satellite cell self-renewal. Curr.Opin.Pharmacol. 2006; 6: 301-6.
2. Boonen KJ,Post MJ. The muscle stem cell niche: regulation of satellite cells during
regeneration. Tissue Eng.Part B.Rev. 2008; 14: 419-31.
3. Ten Broek RW, Grefte S,Von den Hoff JW. Regulatory factors and cell populations
involved in skeletal muscle regeneration. J.Cell.Physiol. 2010; 224: 7-16.
4. Bibel M,Barde YA. Neurotrophins: key regulators of cell fate and cell shape in the
vertebrate nervous system. Genes Dev. 2000; 14: 2919-37.
5. Wang XH,Poo MM. Potentiation of developing synapses by postsynaptic release of
neurotrophin-4. Neuron 1997; 19: 825-35.
6. Xie K, Wang T, Olafsson P, et al. Activity-dependent expression of NT-3 in muscle cells
in culture: implications in the development of neuromuscular junctions. J.Neurosci. 1997;
17: 2947-58.
7. Gonzalez M, Ruggiero FP, Chang Q, et al. Disruption of Trkb-mediated signaling induces
disassembly of postsynaptic receptor clusters at neuromuscular junctions. Neuron 1999;
24: 567-83.
8. Henderson CE, Yamamoto Y, Livet J, et al. Role of neurotrophic factors in motoneuron
development. J.Physiol.Paris 1998; 92: 279-81.
Page 17
This article is protected by copyright. All rights reserved.
9. Davis-Lopez de Carrizosa MA, Morado-Diaz CJ, Morcuende S, et al. Nerve growth factor
regulates the firing patterns and synaptic composition of motoneurons. J.Neurosci. 2010;
30: 8308-19.
10.Colombo E, Romaggi S, Medico E, et al. Human Neurotrophin Receptor p75NTR Defines
Differentiation-Oriented Skeletal Muscle Precursor Cells: Implications for Muscle
Regeneration. J.Neuropathol.Exp.Neurol. 2011; 70: 133-42.
11.Deponti D, Buono R, Catanzaro G, et al. The low-affinity receptor for neurotrophins
p75NTR plays a key role for satellite cell function in muscle repair acting via RhoA.
Mol.Biol.Cell 2009; 20: 3620-7.
12.Funakoshi H, Frisen J, Barbany G, et al. Differential expression of mRNAs for
neurotrophins and their receptors after axotomy of the sciatic nerve. J.Cell Biol. 1993;
123: 455-65.
13.Yamamoto M, Sobue G, Yamamoto K, et al. Expression of mRNAs for neurotrophic
factors (NGF, BDNF, NT-3, and GDNF) and their receptors (p75NGFR, trkA, trkB, and
trkC) in the adult human peripheral nervous system and nonneural tissues.
Neurochem.Res. 1996; 21: 929-38.
14.Ip FC, Cheung J,Ip NY. The expression profiles of neurotrophins and their receptors in rat
and chicken tissues during development. Neurosci.Lett. 2001; 301: 107-10.
15.Sheard PW, Musaad K,Duxson MJ. Distribution of neurotrophin receptors in the mouse
neuromuscular system. Int.J.Dev.Biol. 2002; 46: 569-75.
16.Maisonpierre PC, Belluscio L, Friedman B, et al. NT-3, BDNF, and NGF in the
developing rat nervous system: parallel as well as reciprocal patterns of expression.
Neuron 1990; 5: 501-9.
17.Timmusk T, Palm K, Metsis M, et al. Multiple promoters direct tissue-specific expression
of the rat BDNF gene. Neuron 1993; 10: 475-89.
Page 18
This article is protected by copyright. All rights reserved.
18.Griesbeck O, Parsadanian AS, Sendtner M, et al. Expression of neurotrophins in skeletal
muscle: quantitative comparison and significance for motoneuron survival and
maintenance of function. J.Neurosci.Res. 1995; 42: 21-33.
19.Koliatsos VE, Clatterbuck RE, Winslow JW, et al. Evidence that brain-derived
neurotrophic factor is a trophic factor for motor neurons in vivo. Neuron 1993; 10: 359-67.
20.Mousavi K,Jasmin BJ. BDNF is expressed in skeletal muscle satellite cells and inhibits
myogenic differentiation. J.Neurosci. 2006; 26: 5739-49.
21.Liem RS, Brouwer N,Copray JC. Ultrastructural localisation of intramuscular expression
of BDNF mRNA by silver-gold intensified non-radioactive in situ hybridisation.
Histochem.Cell Biol. 2001; 116: 545-51.
22.Copray S, Liem R, Brouwer N, et al. Contraction-induced muscle fiber damage is
increased in soleus muscle of streptozotocin-diabetic rats and is associated with elevated
expression of brain-derived neurotrophic factor mRNA in muscle fibers and activated
satellite cells. Exp.Neurol. 2000; 161: 597-608.
23.Garcia N, Tomas M, Santafe MM, et al. Localization of brain-derived neurotrophic factor,
neurotrophin-4, tropomyosin-related kinase b receptor, and p75 NTR receptor by high-
resolution immunohistochemistry on the adult mouse neuromuscular junction.
J.Peripher.Nerv.Syst. 2010; 15: 40-9.
24.Gomez-Pinilla F, Ying Z, Roy RR, et al. Voluntary exercise induces a BDNF-mediated
mechanism that promotes neuroplasticity. J.Neurophysiol. 2002; 88: 2187-95.
25.Ogborn DI,Gardiner PF. Effects of exercise and muscle type on BDNF, NT-4/5, and TrKB
expression in skeletal muscle. Muscle Nerve 2010; 41: 385-91.
26.Omura T, Sano M, Omura K, et al. Different expressions of BDNF, NT3, and NT4 in
muscle and nerve after various types of peripheral nerve injuries. J.Peripher.Nerv.Syst.
2005; 10: 293-300.
Page 19
This article is protected by copyright. All rights reserved.
27.Clow C,Jasmin BJ. Brain-derived neurotrophic factor regulates satellite cell differentiation
and skeltal muscle regeneration. Mol.Biol.Cell 2010; 21: 2182-90.
28.Kust BM, Copray JC, Brouwer N, et al. Elevated levels of neurotrophins in human biceps
brachii tissue of amyotrophic lateral sclerosis. Exp.Neurol. 2002; 177: 419-27.
29.Dalakas MC. Polymyositis, dermatomyositis and inclusion-body myositis. N.Engl.J.Med.
1991; 325: 1487-98.
30.Engel A,Franzini-Armstrong C. Myology. McGraw-Hill, Newyork 1994;.
31.Colombo E, Romaggi S, Blasevich F, et al. The neurotrophin receptor p75NTR is induced
on mature myofibres in inflammatory myopathies and promotes myotube survival to
inflammatory stress. Neuropathol.Appl.Neurobiol. 2012; 38: 367-78.
32.Colombo E, Romaggi S, Mora M, et al. A role for inflammatory mediators in the
modulation of the neurotrophin receptor p75NTR on human muscle precursor cells.
J.Neuroimmunol. 2012; 243: 100-2.
33.Molteni R, Calabrese F, Bedogni F, et al. Chronic treatment with fluoxetine up-regulates
cellular BDNF mRNA expression in rat dopaminergic regions.
Int.J.Neuropsychopharmacol. 2006; 9: 307-17.
34.Colombo E, Cordiglieri C, Melli G, et al. Stimulation of the neurotrophin receptor TrkB
on astrocytes drives nitric oxide production and neurodegeneration. J.Exp.Med. 2012; 209:
521-35.
35.Berzi A, Ayata CK, Cavalcante P, et al. BDNF and its receptors in human myasthenic
thymus: implications for cell fate in thymic pathology. J.Neuroimmunol. 2008; 197: 128-
39.
36.Dalakas MC. Pathophysiology of inflammatory and autoimmune myopathies. Presse Med.
2011; 40: e237-47.
Page 20
This article is protected by copyright. All rights reserved.
37.Orimo S, Koga R, Goto K, et al. Immunohistochemical analysis of perforin and granzyme
A in inflammatory myopathies. Neuromuscul.Disord. 1994; 4: 219-26.
38.Chazaud B, Sonnet C, Lafuste P, et al. Satellite cells attract monocytes and use
macrophages as a support to escape apoptosis and enhance muscle growth. J.Cell Biol.
2003; 163: 1133-43.
39.Segawa M, Fukada S, Yamamoto Y, et al. Suppression of macrophage functions impairs
skeletal muscle regeneration with severe fibrosis. Exp.Cell Res. 2008; 314: 3232-44.
40.Chazaud B, Brigitte M, Yacoub-Youssef H, et al. Dual and beneficial roles of
macrophages during skeletal muscle regeneration. Exerc.Sport Sci.Rev. 2009; 37: 18-22.
41.Arnold L, Henry A, Poron F, et al. Inflammatory monocytes recruited after skeletal
muscle injury switch into antiinflammatory macrophages to support myogenesis.
J.Exp.Med. 2007; 204: 1057-69.
42.Desguerre I, Mayer M, Leturcq F, et al. Endomysial fibrosis in Duchenne muscular
dystrophy: a marker of poor outcome associated with macrophage alternative activation.
J.Neuropathol.Exp.Neurol. 2009; 68: 762-73.
43.Tidball JG. Inflammatory processes in muscle injury and repair.
Am.J.Physiol.Regul.Integr.Comp.Physiol. 2005; 288: R345-53.
44.Kerschensteiner M, Gallmeier E, Behrens L, et al. Activated human T cells, B cells, and
monocytes produce brain-derived neurotrophic factor in vitro and in inflammatory brain
lesions: a neuroprotective role of inflammation? J.Exp.Med. 1999; 189: 865-70.
45.Besser M,Wank R. Cutting edge: clonally restricted production of the neurotrophins brain-
derived neurotrophic factor and neurotrophin-3 mRNA by human immune cells and
Th1/Th2-polarized expression of their receptors. J.Immunol. 1999; 162: 6303-6.
Page 21
This article is protected by copyright. All rights reserved.
Figure legends
Figure 1. BDNF is expressed in human muscle precursor cells and in mature myofibers. (A)
BDNF, p75NTR and TrkB mRNA levels in cultured myoblasts. (B) Double
immunofluorescence for BDNF and CD56/NCAM in cultured myoblasts. (C) Regulation of
BDNF, p75NTR and TrkB mRNA levels upon differentiation. Quantitative real-time PCR
was performed at distinct time points. In (A) and (C) mRNA levels of target genes were
graphically reported as percentage of the housekeeping gene PPIA. (D) Double
immunofluorescence for BDNF and dystrophin in cultured myotubes. (E) Cytofluorimetric
analysis of cultured myoblasts for TrkB surface expression. (F) Double immunofluorescence
for TrkB and CD56/NCAM in cultured myoblasts. (G-H) BDNF immunoreactivity in a
CD56/NCAM (G) or p75NTR (H) positive satellite cell of adult skeletal muscle. (I)
Percentage of CD56/NCAM positive satellite cells expressing BDNF in human adult skeletal
muscle. Circles represent distinct tissue samples, red bar represents mean. At least 40 CD56
positive satellite cells in at least 2 sections per sample were counted. (J) In situ hybridization
for BDNF in human adult skeletal muscle. (K-L) Immunohistochemistry for BDNF (K) and
TrkB (L) in human adult skeletal muscle. Black arrow indicates TrkB positive vessel. In (A)
and (C) error bars represent standard deviations. Scale bar 30 μm in (B), (D), (F) and (L), 3,5
μm in (G) and (H), 50 μm in (J), 100 μm in (K).
Page 22
This article is protected by copyright. All rights reserved.
Page 23
This article is protected by copyright. All rights reserved.
Figure 2. BDNF regulates myogenesis in vitro. (A-C) BDNF silencing in myoblasts and
effects on myogenesis. (A) BDNF silencing efficiency monitored 3 days after differentiation
induction by quantitative RT-PCR. BDNF mRNA levels were graphically reported as
percentage of the housekeeping gene PPIA. (B-C) Immunofluorescence for desmin and DAPI
(B) and fusion index evaluation at day 6 after differentiation induction (C) in cultures treated
with BDNF or control siRNA. (D-E) Immunofluorescence for desmin and DAPI (D) and
fusion index quantification (E) in a blocking experiment with anti-BDNF antibody or isotype
control administered during cell differentiation. Arrows indicate myotubes (cells with more
than 2 nuclei). Error bars represent standard deviations. Experiments were performed in
triplicates and at least 1000 nuclei per replicate were counted. Similar observations were
obtained in at least three independent experiments in two primary cell lines. Scale bar 50 μm.
* P< 0.05, *** P<0.001
Page 24
This article is protected by copyright. All rights reserved.
Page 25
This article is protected by copyright. All rights reserved.
Figure 3. BDNF is produced by immune cells located near regenerating fibers in
inflammatory myopathies. (A) Quantitative PCR for BDNF mRNA tissue levels in control
biopsies (n=7) and in idiopathic inflammatory myopathies specimens (n=14). BDNF mRNA
levels were graphically reported as percentage of the housekeeping gene PPIA. Error bars
represent SEM. (B-E) Newly forming fibers are basophilic at H&E (B), and stain positively
for CD56 (C), p75NTR (D) and BDNF (E). Several immune cells localized near p75NTR and
CD56/NCAM positive regenerating fibers, and were positive for BDNF. Stainings were
performed on serial sections. Triangles, asterisks and white arrows indicate regenerating
myofibers, mature myofibers and groups of BDNF-positive immune cells respectively. (F-G)
haematoxylin-eosin and CD56/NCAM stained serial sections of IIM muscle. Right panels
show enlarged areas with mature (asterisks, upper) or regenerating (triangles, lower) fibers.
Asterisks and triangles point out fibers included in the quantification. (H) Numbers of
immune cells localized in the proximity of regenerating or mature myofibers. (I) Numbers of
BDNF positive immune cells localized in the proximity of regenerating or mature myofibers.
Dots represent distinct tissue samples, red bars represent means. In (H) and (I) all fields of at
least 2 sections per sample were analyzed; mature fibers near regenerating fibers were not
included in the quantification. Double immunofluorescence for BDNF and CD4 (J), CD8 (K)
or CD68 (L) in IIM muscle. Left panels represent single stainings. Arrows indicate BDNF
positive cells. Scale bar 30 μm in (B-E), 100 μm in (F-G), 20 μm in (J-L). * P < 0.05.
Page 26
This article is protected by copyright. All rights reserved.
Page 27
This article is protected by copyright. All rights reserved.
Supplementary Figure. 1 Perforin expressing T cells or Ki67 positive macrophages are
localized near mature fibers. Left panels show immunohistochemistry for CD56. Right panels
show triple immunofluorescence for CD3 and perforin (A and B) or CD68 and Ki67 (C and
D) in combination with desmin. Middle panels represent single stainings. Arrows indicate
perforin or Ki67 positively stained cells. Asterisks indicate the same fibers in left and right
panels. Scale bar 50 μm.