The only way a bird can obtain energy is by expending metabolic energy while foraging for food items that contain appropriate amounts and forms of energy. Birds are often specialized to use only certain kinds of food. The implication is that natural selection has shaped the morphological and physiological traits that determine the ability of a bird to obtain and process food. These general considerations, however, have not yet led to a useful predictive theory of avian foraging.
There are two distinct approaches to the relationship between energetics and foraging. The first is empirical and emphasizes documenting the energy budget of foraging organisms (e.g., Bryant and Westerterp 1980; Walsberg 1983; Stalmaster and Gessaman 1984; Goldstein 1990; Kooyman et al. 1992). The second is theoretical and is based on the assumption that birds forage in such a way to optimize the amount of energy that they consume (Stephens and Krebs 1986). The goal of this paper is to examine the energetics of foraging from a broad perspective that incorporates both of these approaches. I examine the energetics of foraging from an evolutionary perspective, with the intention of establishing how considerations of energetics should influence how we view the evolutionary significance of avian foraging structures and behaviors.
A number of reviews have examined the question of whether foraging behavior of birds and other animals can be considered as optimal solutions to the problem of obtaining energy (Schoener 1971; Pyke et al. 1977; Krebs et al. 1983; Pyke 1984; Stephens and Krebs 1986). These reviews have suggested that, in general, optimal foraging models can predict some, but not all of the aspects of foraging behavior exhibited by organisms. Critics of optimal foraging theories point out
that these failures indicate that the theories are only crude approximations of the way that birds forage, at best, and hence are of limited usefulness in explaining why and how birds forage (Myers 1983; Gray 1987). The approach taken in this review is to enumerate the energetic costs and benefits of foraging and how they affect the specific strategy used in obtaining resources. This information will then be used to assess the importance of optimization theories.
A GENERAL MODEL OF FORAGING ENERGETICS
The scale of a foraging model in space and time may have profound consequences for its implications, particularly when the model is an optimization model (Stephens and Krebs 1986; Gass and Roberts 1992). Initially, my approach is descriptive rather than predictive. The approach taken is to view the costs and benefits of foraging on the rate of energy intake during a typical day. Let E be the amount of energy obtained by a bird during the time T it spends foraging during the day. Then the net rate of energy intake over that time period is
E ET -EF -Ep -E/
T Ts +TH +Tp +T/
where
E T = total assimilable energy obtained during foraging
E F = energy spent searching, pursuing, and handling food
E p = energy spent in physiological processing of food
E[ = energy spent in interactions with other organisms during foraging
Ts = time spent searching for and pursuing food
T H = time spend handling food
Tp = time spent during physiological processing of food
T[ = time spent interacting with other organisms during foraging
(8.1)
Each of the terms in Equation 8.1 may have significant impacts on the way a bird forages and eventually may influence the kinds of adaptations that a species develops for energy acquisition over evolutionary time. Before considering the traits of a bird that affect the energy costs and benefits implied in Equation 8.1 and how these may ultimately affect the evolution of foraging adaptations, consider each of the terms in Equation 8.1 in more detail.
The term E T is the total amount of energy that can be assimilated from the food actually ingested by the foraging organism. Diets of different birds vary consider-
252 Energetics of Avian Foraging
ably in the amount of energy that can be derived from the food ingested (Robbins 1983). Seeds and leaves have relatively low amounts of energy that can be derived from ingested material, particularly if they have secondary compounds that reduce their digestibility (see, e.g., Grazal et al. 1989). Animal tissue generally contains more assimilable energy per unit of food. The energy content of foods may also vary seasonally, particularly in plant materials. Behavioral adjustments can be made so that the bird eats only parts of the food (Kaspari 1990), in which case, the actual energy derived can be much higher (Robbins 1983).
While obtaining food, birds must expend a certain amount of metabolic energy in activities that allow the discovery, capture, and handling of food items prior to consumption, hence, it is necessary to include E F' Ts and THin Equation 8.1. For some types of food, this energy expenditure may be relatively insignificant. For example, many plant food items are widely available and easily obtained so that the bird foraging on them need only locate appropriate items and consume them. At the other extreme, some animal food items are relatively rare and require substantial energy for pursuit, capture, killing, and consumption.
Once food enters the digestive tract, energy must be spent on physiological processes that break down the food into energy and nutrients used for maintenance and production (see Chapter 3). The terms Tp and E p in Equation 8.1 represent the time and energy, respectively, spent in these physiological processes. The time and energy spent on physiological processing are related to food quality. For foods that are high in energy, such as many animal tissues and nectar, time and energy expended in physiological processing is relatively small. For plant and animal foods that have low concentrations of energy and other nutrients, physiological processing costs are much higher. It should be noted here that a bird can begin physiological processing while engaging in other foraging activities. So Tp should strictly be interpreted as the amount of time that the bird spends when its only foraging activity is digestion. This time would begin after the individual becomes satiated and ceases other foraging activities. Thus, it is possible that this time could be zero even ifthe energy expended }n digestion was large. In some birds, however, satiation occurs relatively rapidly, so that Tp should also include time necessary for the gut to empty.
As birds move about their environments obtaining food, they encounter other individuals of the same and different species that are attempting to use the same resources. In addition, they are exposed to predators (e.g., Grubb and Greenwald 1982; Lima 1985,1987; Valone and Lima 1987). Therefore, it seems appropriate to include in the energetic equation terms that identify the amount of time and energy spent resolving these interactions during active foraging (E[ ~d T[). Although birds interact with other organisms in contexts other than foraging, there is a proportion of those interactions that would not be experienced ifthe bird were not actively foraging. This proportion is added to the cost offoraging in Equation 8.1.
Traits That Affect the Energetic Cost of Foraging 253
TRAITS THAT AFFECT THE ENERGETIC COST OF FORAGING
In most theoretical and empirical analyses of foraging behavior, emphasis is placed on the behavioral and morphological traits that affect the ability of the individual to obtain food. Rarely, physiological traits are examined (Speakman 1987). These traits generally are thought to have evolved by natural selection. Although some traits may arise by other processes and eventually become incorporated into a species' foraging repertoire, it is thought that natural selection must eventually affect the evolution of such traits. This section discusses the kinds of traits that affect the energetic costs of foraging in detail.
Foraging Energy Balance and Natural Selection
Lotka (1925) was one of the first biological theoreticians to examine the relationship between energy and natural selection. He pointed out that the direction of evolution should be profoundly affected by the ability of organisms to process energy (Lotka 1922a, 1922b). Those that do so more effectively are able to obtain a greater proportion of resources than those with lesser abilities. Hence, evolution leads, according to Lotka, to an increase in the ability of organisms to use energy efficiently and rapidly. Natural selection can be interpreted as an energetic principle that Lotka (1922b) called "the persistence of stable forms." Of interest here is Lotka's explicit connection between the ability of organisms to obtain and process energy and natural selection.
Equation 8.1 is intended to include the costs and benefits of foraging in terms of energy per unit time. The foraging behavior that a bird of a given species uses in obtaining food will have consequences for the overall energy balance that the bird maintains over time. Clearly, the foraging behavior employed by an individual will affect the overall energy balance that it maintains. The rate of daily energy acquisition (R), where R = EfT, determines that amount of energy that an individual has to perform necessary metabolic and behavioral work needed to sustain it. Thus, R must be >0 over a sufficient number of days to provide for an adequate amount of energy available for maintenance. Although R might be < 0 on occasion, if it is negative for a sufficient number of days, then the individual will die. The ability of an individual to maintain a positive daily energy intake will be referred to as its foraging energy balance.
An organism might maintain a positive foraging energy balance yet not have sufficient energy left over after maintenance to reproduce. If there is a threshold rate of daily energy intake that is necessary for maintenance, say Rm, then R must exceed Rm by a sufficient magnitude so that the organism can use that energy to reproduce. Natural selection then operates when some organisms exceed their
254 Energetics of Avian Foraging
maintenance requirements and others do not. Consistent patterns of death of individual organisms with particular traits that affect their foraging or a consistent inability of those organisms to obtain sufficient energy to reproduce may cause a shift in the distribution of foraging traits in a population, and therefore, evolution of more efficient foraging traits.
An important part of an organism's foraging repertoire is its ability to shift behavioral modes when it encounters resources of differing quality (Helfinan 1990). In general, birds possess more than one specific kind of foraging mode, and using different traits when presented with different spatial and temporal patterns of prey dispersion has measurable energetic consequences. For example, American kestrels (Falco sparverius) expend more energy when hunting for prey by hovering in the air but also increase their intake of prey when compared to hunting by searching from a perch (Rudolph 1982). Hovering increases net energetic benefit when wind speed is relatively high. Natural selection should operate both on the nature of the individual foraging traits of an organism and on its ability to use different modes appropriately. Thus, behavioral plasticity should itself be subjected to natural selection as a foraging trait (Morse 1971).
The end result ofthis relationship between energy and foraging behavior is that we should expect to observe traits that affect foraging to be "designed" for efficiency of energy processing. This expectation has led to a research program in which mathematical criteria are used to establish optimality rules for behavioral traits (and sometimes other traits) of birds based on the kind of cost-benefit approach implicit in Equation 8.1. The quantity being optimized is most often rate of energy acquisition or rate of delivery of energy to a central place such as a nest or storage area (Stephens and Krebs 1986). Such approaches, although they have been moderately successful, do not account for the fact, first observed by Lotka (1925:346-354), that natural selection does not operate directly on the rate of energy acquisition, but on the life history traits that are affected by the foraging energy balance maintained by individuals (Watt 1985; see also Chapter 1 this volume, Steams 1992). Hence, such optimality models can never, in principle, completely describe the behavioral attributes that result from the operation of natural selection.
The Foraging Apparatus
Given that natural selection has shaped many, if not all, of the traits used by birds to forage, it is possible to examine that set of traits that influence the ability of an individual to maintain a positive foraging energy balance. This set of traits can be pictured as the "foraging apparatus": a single functional unit in the organism that is used to search for, capture, consume, and process food. The foraging apparatus of a bird is composed of three kinds of traits: physiological, morphological, and behavioral.
Traits That Affect the Energetic Cost of Foraging 255
Physiological Traits The most important physiological trait of an organism that affects its foraging must be its digestive efficiency (Karasov 1990; Chapter 3). Digestion can be broken into two parts. First the bird must mechanically break down food and then it must absorb appropriate nutrients into its bloodstream (assimilation). Digestive efficiency is determined by the morphological and chemical characteristics of the digestive tract as they interact with the biochemical and mechanical properties of food. The digestive efficiency determines the amount of energy that the bird must expend on digestion and the amount oftime that must be expended on digestion after the bird has stopped actively foraging. Karasov (1990) examined many of the energetic and physiological constraints on digestion. He argued that these constraints lead to a number of trade-offs that seem to be involved in digestion. For example, Karasov (1990) compared gut retention times in a nectarivore and small frugivore. When corrected for body mass, the frugivore retained the food mass in the gut for a much shorter time and was less efficient at extracting glucose than the nectarivore. Presumably the foraging energy balance in the frugivore was maintained by the larger absolute mass of food that could be ingested or by other behavioral adjustments.
In the present context, digestion interacts with many other aspects of the foraging apparatus. For example, birds that eat animals tend to have less complex digestive systems, but must expend a great deal of time and energy pursuing prey items. Herbivorous birds, on the other hand, have relatively complex digestive systems and must expend considerable energy breaking down food particles in the crop before digestion, but their food can be gathered with relatively little expenditure of energy.
Morphological Traits A number of external morphological traits are associated with the pursuit, capture, and consumption of prey items. In general, the size and shape of different appendages should influence the ability of a bird to capture prey items, and consequently affect the daily rate of energy intake. Furthermore, because ecological conditions vary seasonally, morphological traits involved in foraging may often reflect periods of time when food is especially limiting (Dunning 1986). I consider several examples below.
Perhaps the most obvious trait influencing foraging of a bird is the bill. Birds of distantly related taxa that use similar prey items often have similar bill shapes (see, e.g., illustrations in Faaborg 1988). Such variation in bill shape among taxa clearly indicates an effect of natural selection. But more detailed functional analyses of bill size and shape within and among closely related species are often more enlightening. For example, consider the variability in bill morphology and the accompanying muscle structure among several species of Galapagos finches studied by Bowman (1961). Although large species have larger bills and associated muscles than small birds, relative sizes of muscles working the jaw (proportional to the fraction of total body mass that the muscle represents) is greater in larger
256 Energetics of Avian Foraging
species (Fig. 8.1). Thus, large species are able to process much harder seeds than small species. Within a single species (GeospizaJortis), Boag and Grant (1981) showed that individuals oflarger size had a higher survival rate than smaller individuals during a period of extreme food limitation, when availability of small seeds decreased drastically due to drought conditions.
Figure 8.1 Variation among species of Galapagos finches in relative muscle masses for two different muscles involved in feeding. Note that the relative muscle mass increases linearly with body mass. Thus, larger species devote a larger proportion of their body mass to feeding muscles. Lines represent the least squares regression for each data set. Data from Bowman (1961, Tables 33 and 34).
Traits That Affect the Energetic Cost of Foraging 257
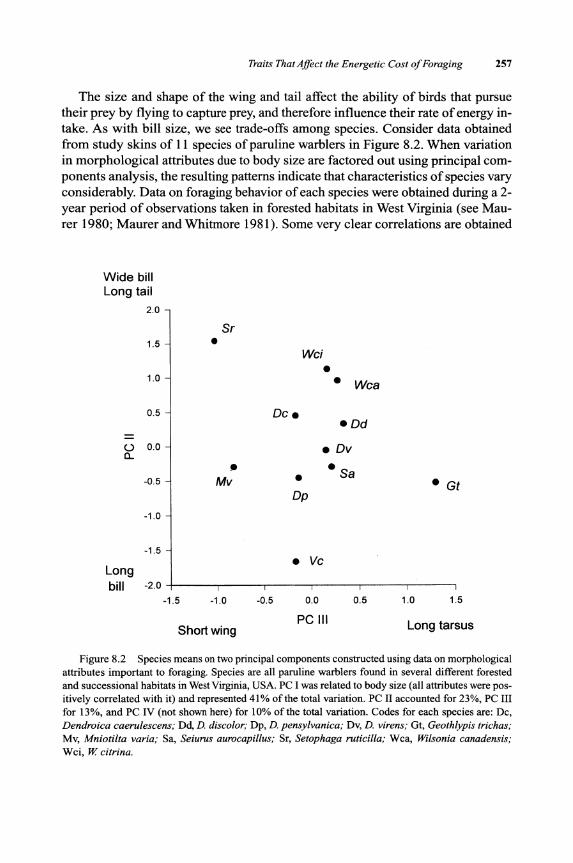
The size and shape of the wing and tail affect the ability of birds that pursue their prey by flying to capture prey, and therefore influence their rate of energy intake. As with bill size, we see trade-offs among species. Consider data obtained from study skins of 11 species of paruline warblers in Figure 8.2. When variation in morphological attributes due to body size are factored out using principal components analysis, the resulting patterns indicate that characteristics of species vary considerably. Data on foraging behavior of each species were obtained during a 2-year period of observations taken in forested habitats in West Virginia (see Maurer 1980; Maurer and Whitmore 1981). Some very clear correlations are obtained
Wide bill Long tail
2.0
Sr 1.5 •
Wei
• 1.0 • Wea
0.5 De. .Dd
0 0.0 • Dv a... It
• Sa -0.5 Mv • • Gt Dp
-1.0
-1.5
• Ve Long bill -2.0
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
PC III Long tarsus Short wing
Figure 8.2 Species means on two principal components constructed using data on morphological attributes important to foraging. Species are all paruline warblers found in several different forested and successional habitats in West Virginia, USA. PC I was related to body size (all attributes were positively correlated with it) and represented 41 % of the total variation. PC II accounted for 23%, PC III for 13%, and PC IV (not shown here) for 10% of the total variation. Codes for each species are: Dc, Dendroica caerulescens; Dd, D. discolor; Dp, D. pensylvanica; Dv, D. virens; Gt, Geothlypis trichas; Mv, Mniotilta varia; Sa, Seiurus aurocapillus; Sr, Setophaga ruticilla; Wca, Wilsonia canadensis; Wci, W citrina.
258 Energetics of Avian Foraging
between morphological variation and particular foraging behaviors. For example, species that obtain prey by aerial sallies have longer tails than those that glean insects from vegetation (Table 8.1). Richman and Price (1992) found similar relationships between foraging behavior and morphological variation among species of Phylloscopus warblers in the Himalayas. Such correlations between the size and shape of appendages and the kinds of foraging behavior that a species exhibits implies that there are inherent energetic trade-offs that occur when natural selection favors a certain kind of foraging behavior. Of course, these correlations do not imply that morphological traits used in foraging are exclusively determined by natural selection for maximal foraging efficiency, since other factors, such as escape from predation and constraints on flight during migration may also contribute to their evolution.
The length of the legs is also often related to the location where a species characteristically forages. Warblers that spend a great deal of time foraging on the ground have relatively longer tarsi than those that forage in trees (Table 8.1; see also Osterhaus 1962). The length of legs may be particularly important for many shorebirds, because their foraging may be limited to certain water depths by leg length. Species that have long legs may be able to pursue prey in relatively deep water, but may be less successful in habitats closer to the shore.
TABLE 8.1 Correlations among Foraging Behaviors and Morphological Traits of Several Species of Wood Warblers (Parulinae) Found Together in Forested Habitats in West Virginia, USA
bPrincipal component I is a measure of body size, and was not included here. Components II-IV are indicators of shape.
The Allometry of Foraging Behavior 259
Behavioral Traits Birds must actively pursue, capture, and consume the food that they obtain their energy from. These activities often require an elaborate series of behaviors that must be performed with great accuracy in order to be successful. Birds must sense their food, which can be cryptically colored, and react to these sensory stimuli appropriately. Furthermore, in order to maintain a positive energy balance and hence have sufficient energy available for reproduction, birds must choose among food items that vary in their energy and nutrient content.
Species show profound variation in the degree to which they specialize on resources (Morse 1971; Sherry 1990). How do such differences between species arise? Greenberg (1990 and references therein) has proposed that specialization evolves from the variation among individuals within species in their aversion to approaching novel resources. Greenberg has shown clear differences among species in their hesitancy to approaching new resources. He points out that further research is needed to determine the genetic component of this type of behavior. His hypothesis, as yet only tested qualitatively, merits further attention. Optimal foraging theories make quantitative predictions regarding behavioral specialization in relationship to resource abundance (see Optimal Foraging Theory), and an interesting direction for future research would be to develop models that relate these predictions to Greenberg's neophobia hypothesis.
THE ALLOMETRY OF FORAGING BEHAVIOR
Body mass is closely tied to the metabolic machinery (Peters 1983; Robbins 1983; Calder 1984) and behaviors (Goudie and Piatt 1991) used by an organism to process food and extract usable energy from it. In general, large animals require more gross energy but have lower energetic expenditures per gram body mass. In mammals, large herbivores can use poorer quality food than small herbivores (Dement and Van Soest 1985), but similar relationships have not been examined for birds. Smaller birds generally tend to require food items with higher concentrations of energy than larger birds, and the smallest birds are nectarivores. Body mass also influences the costs of flight (Pennycuick 1975), which should have an impact on energetic costs for foraging.
Bryant and Westerterp (1980) obtained estimates of foraging rates in birds (kJ/h) for 67 species of birds. A linear regression on a double log plot of foraging rate on body mass showed a good fit to the data (Fig. 8.3). Bryant and Westerterp (1980) fit a linear regression to the logarithmic data on foraging rates and obtained the following relationship (excluding seabirds, which are outliers in Fig. 8.3)
W l- •• « • • 0::: 100 C) • Z (!) « 10 0:: • 0 lJ..
1 1 10 100 1000 10000
BODY MASS (9)
Figure 8.3 Allometric variation of daily energy expenditures (field metabolic rate) and foraging rates among birds. Lines represent linear regressions. On the average, the scaling of foraging rates and energy expenditures indicates that a bird must forage for 5 hours a day to meet its expenditure of en-ergy. See text for further discussion.
Nagy and Obst (1991) used data in Nagy (1987) and added data from 15 ·additional species to obtain an estimate of the scaling of daily energy expenditure (kJ/d) to body mass as
(8.3)
The daily energy expenditure (kJ/d) divided by the foraging rate (kJ/h) gives the number of hours per day that a bird must forage to meet its energy expenditures.
Optimal Foraging Theory 261
Given these estimates of Fr and Ed' on average a bird must forage about 5 h per day to meet its own energy requirements. Very clearly this time will vary seasonally (e.g., Pulliam 1980), but the important point to note is that the amount of time spent foraging is independent of body size, since E/Fr ex: M-O.Ol. Large and small birds differ significantly in the ways that they obtain food. Larger organisms generally make fewer attempts at securing prey since they are capable of taking larger prey items (Peters 1983) but may spend longer consuming them. These considerations imply that there are trade-offs in the costs and benefits of foraging between large and small birds.
Since the costs and benefits of foraging must be measured in units of energy, trade-offs involved in the evolution of the relationship between foraging and body mass must be energetic. Recall that larger Galapagos finches tended to have proportionately larger mass for muscles controlling bill movements (Fig. 8.2). This suggests such an energetic trade-off. Larger birds require a larger absolute amount of food than smaller birds but have a lower per unit mass metabolic rate. The energetic benefit of being large allows a bird to build more bill muscle mass. This, in turn, allows larger birds to process harder seeds (regardless of seed size). The cost is that in being larger, the bird must increase its total consumption. In terms of Equation 8.1, a large bird must increase total energy gain (Er) and decrease time of handling its food items (TH ) to increase its rate of energy consumption if it is to support its larger mass. Notice that the reverse is also true, that is, a small bird can decrease its rate of energy consumption because it has less total energetic costs. Thus, depending on the distribution of food items, this energetic trade-off could provide the basis for selection that would cull individuals that were too large or too small. Further research is needed to demonstrate how natural selection on the allometry of the components of foraging behavior leads to such tradeoffs.
OPTIMAL FORAGING THEORY
Given that natural selection has shaped the adaptations of species for foraging, a large body of literature has been developed to explain how natural selection can lead to behaviors that maximize the rate of energy intake within constraints imposed by the environment (see review by Stephens and Krebs 1986). A number of assumptions are required in order for these models to be valid, some of which are examined in this section in detail. Of particular importance are assumptions that relate to how evolution occurs. There seems to have been a tendency of many foraging theorists to gloss over some of the details of the evolutionary mechanisms and the assumptions they entail. My intent is to outline some of these issues by casting the energetics of foraging within a quantitative genetics framework. This will be helpful in pointing out some of the specific limitations of many current optimal foraging approaches and in suggesting future directions for research.
262 Energetics of Avian Foraging
After outlining the evolutionary assumptions of optimal foraging, I review some of the major results of optimal foraging theory, describe some recent theoretical advances, and update empirical studies of optimal foraging using data on birds conducted in the past several years (since about 1985). Summaries and discussion of earlier studies can be found in Pyke et al. (1977), Krebs et al. (1983), Pyke (1984), and Stephens and Krebs (1986). Whatever their limitations are, the advantage of optimal foraging theories is that they produce testable predictions. The difficulties with them have been mentioned previously. Despite these difficulties, continued research into these models will provide some fundamental insights into the relationship between energetics and the evolution of foraging behavior.
General Premises of Optimal Foraging
The proposition that animals forage to optimize their rate of energy intake is based on a number of premises that should be considered before examining specific models (Pyke 1984). Note that "optimize" does not necessarily mean "maximize." Some models include constraints that result in an optimal rate of energy intake that is less than maximum. In the sense used in most optimal foraging models, optimal means the highest rate of energy intake that allows the individual to meet the constraints imposed by the environment. An example of this is models in which the organism must meet certain nutritional constraints. These models predict that the organism may choose food items that are rich in nutrients but may lower its rate of energy intake.
The first general assumption made by all optimal foraging models is that the optimal design in terms of energy and nutrient intake (or delivery of energy to a central place) corresponds to the phenotype that has the highest fitness. As the discussion in previous sections indicates, it is reasonable to expect that natural selection has given rise to the attributes of a bird included in its foraging apparatus, but it is not clear that this necessarily points to optimization. Since the relationships of most of these foraging attributes to the life history components that determine fitness are virtually unknown in most studies of optimization (but see Lemon 1991), this assumption is rarely tested.
The second general assumption is that the physiological, morphological, and behavioral components that determine the optimal phenotype are heritable. There is relatively little evidence on heritabilities of traits that determine the foraging apparatus. The best evidence is for morphology (see, e.g., Grant 1986; Boag and van Noordwijk 1987). Heritability of physiological traits such as digestive efficiency and behavioral traits such as neophobia are virtually unknown. The inclusion in the phenotype of significant nonheritable components (e.g., learned behaviors, environmental effects) that affect individual productivity may lead to a population that is at evolutionary equilibrium but still experiences a significant fitness gradi-
Optimal Foraging Theory 263
ent. This possibility has been discussed by Price et al. (1988) for breeding dates in birds, but they point out that any quantitative genetic characteristic may show this kind of evolutionary equilibrium. I consider this possibility for foraging models below. A clearer understanding of the genetic mechanisms underlying variation in the foraging apparatus is needed in order to assess the degree to which these traits are heritable.
A third important assumption is that the current environment in which an organism is living is responsible for the evolution of its adaptations. This is a universal assumption of the so-called neodarwinian paradigm of evolution and an assumption that has been widely criticized (see, e.g., Wiens 1976; Lewontin 1979; Gould and Lewontin 1979). With the advent of techniques for comparative studies (Brooks and McLennan 1991; Harvey and Pagel 1991), testing this assumption for any given study is now a real possibility. Unfortunately, such techniques require that estimates of the phylogeny of the taxa being compared are available. Phylogenies are rarely available, but work by systematists is proceeding at an encouraging pace. Future studies incorporating comparative data on foraging energetics for groups of species with reliable estimates of their phylogenies will provide fundamental advances in our understanding of the energetics of avian foraging (see Phylogenetic Patterns of Foraging Adaptations).
The effects of departure of real biological systems from these assumptions on the usefulness and validity of optimization studies could be serious, but are unknown. Defenders of the optimization paradigm argue that departures from these assumptions cannot be excessive because there has been good success in predicting patterns from optimization models (Maynard Smith 1978, 1982; Parker and Maynard Smith 1990; Mitchell and Valone 1990). Yet as has been shown in the past, predictions from optimization models are still open to quantitative and qualitative failures of their predictions (see Optimal Foraging in Birds). In addition, there is the distinct possibility that optimal foraging models get the right answer for the wrong reasons. That is, qualitative predictions from optimal foraging models may not be unique, so that other, more realistic models that do not have their basic premises in question may make the same predictions. I believe that it would be healthy for optimal foraging to be challenged by theoretical alternatives. Many viable theoretical alternatives have yet to attain the same degree of sophistication (Ward 1992, 1993; Nonacs and Dill 1993). The limitations of optimal foraging models are not sufficient to lead to their abandonment (Stephens and Krebs 1986; Stephens 1990).
The three assumptions just discussed are rarely dealt with in a manner that allows their precise definition. In the next section, I outline a model based on the assumption that the characteristics that define the foraging apparatus that will be optimized by evolution are determined by quantitative genetic variation (e.g., Falconer 1989). This assumption is based on the premise that the genetic loci that determine these traits are no different in kind than loci that determine qualitative
264 Energetics of Avian Foraging
(Mendelian) traits, but are simply more in number and cumulative in effect (Crow 1986). The intent of this model is to outline more precisely what the assumptions underlying optimal foraging imply about the process of evolution that has given rise to the characteristics that define the foraging apparatus of a species.
The Quantitative Genetics of Optimal Foraging
In examining the effects of natural selection on the energetics of foraging, it is useful to examine explicitly how natural selection might cause changes in the foraging apparatus. This will be particularly useful later, when examining optimal foraging models. These models are only infrequently framed explicitly in terms of the genetic mechanisms underlying them (e.g., Schoener 1971; McNamara and Houston 1986; Abrams 1991). As just discussed, assumptions are made regarding these mechanisms, but those assumptions have rarely been examined theoretically or empirically. Here I sketch one way that these assumptions might be examined theoretically.
Assume that the foraging apparatus of a species can be described by a numher of quantitative genetic characters referring to the morphological, physiological, and behavioral attributes involved in foraging. Let the vector X represent a set of random variables describing these traits such that X; is the ith trait. The genetic variances of traits and their covariances are given by the matrix G, where
G=[aij]i,j= 1,2, ... ,p (8.4)
aij is the genetic covariance between traits i andj, and a ii = a; is the genetic variance of trait i. The genetic variances describe how much quantitative genetic vari
. ation exists for each trait. This variation is generally thought to be additive (e.g., independently assorting loci), although allelic interactions may also be included. The covariances describe how traits are related to one another, for example, how the genetic component of bill length varies with the genetic component of bill width (presumably because some of the same genes affect both traits).
Now assume, as we have above, that the foraging apparatus, as a unit, is responsible for the rate of energy intake, R, given by Equation 8.1. The rate of energy intake can then be considered as a function of the traits, that is
(8.5)
The rate of energy intake can itself be considered a quantitative characteristic. However, it is under genetic control only so far as the individual traits that make it up are. Thus, it is useful to make a distinction between the traits that comprise the foraging apparatus and the rate of energy intake derived from them. The traits that comprise the foraging apparatus I will refer to as primary phenotypic traits and the rate of energy intake will be called a secondary phenotypic trait, since its
Optimal Foraging Theory 265
properties are derived from the primary traits. If there is genetic variation within and among the primary phenotypic traits, then there will be a distribution of genetically determined rates of energy intake among individuals in the population.
The genetic variance of R can be obtained approximately from a Taylor series expansion as
(S.6a)
where
(S.6b)
I-Li is the population mean of trait i and IT ij is the appropriate genetic variance or covariance of a primary phenotypic trait. The important point to get from Equation 8.6 is that the relationship between the secondary trait (R) and the primary traits as defined in Equation 8.5 constrains the patterns of genetic variation of the secondary trait. Thus, the rate of energy intake varies among different combinations of primary traits such as bill width, tail length, physiological rates of food processing, etc. An important assumption behind much of foraging theory is that certain combinations of primary foraging traits will maximize the rate of energy intake. Let the vector X* represent such a combination, thenf(X*) = Rmax' There may be several alternative combinations X~, XZ, etc., such thatf(X~) = f(XZ) =
Rmax' Combinations of primary traits are called strategies, and those that maximize R are rate maximizing strategies.
Equations 8.5 and 8.6 define the relationship between the energetics of foraging and the foraging apparatus. Optimal foraging theories also require that natural selection affect the foraging apparatus via the rate of energy intake. Therefore, they make assumptions regarding the relationship between energy intake and fitness. These relationships, orfitnessfunctions, define the optimalforaging strategy as that strategy (or strategies) for which the fitness function is maximum. Let W be the fitness of a phenotype, then
W=g(R) (S.7)
is the fitness function for strategies defined by Equation 8.5. The variance in fitness among individuals (or strategies) is approximately
(S.Sa)
where
(S.Sb)
266 Energetics oj Avian Foraging
Thus, the fitness function determines the shape of the distribution of fitnesses among strategies, and ultimately the optimal strategy, or if several rates of energy intake have equal fitnesses, the optimal strategies.
Fitness functions can be one of two kinds. When the rate maximizing strategy has the highest fitness, then the fitness function is a rate-maximizing fitness function. Since natural selection leads to the strategy with the highest fitness, if the fitness function is rate maximizing, then evolution will lead to organisms that maximize their rate of energy intake (Fig. 8.4A,B). It is possible that a strategy will exist in the population that has higher fitness than the rate maximizing strategy. In this case, we have a nonrate-maximizing fitness function. Such a fitness function will cause evolution to produce organisms that do not maximize their rate of energy intake. Examples of optimal foraging models that produce nonrate maximizing strategies are discussed in the next section (General Classes of Optimal Foraging Models).
Recall that the second assumption underlying optimal foraging is that the characteristics defining the foraging apparatus are heritable so that the strategies they produce are heritable. Although there is evidence that individual morphological traits have a heritable component (Boag and van Noordwijk 1987), the heritability of foraging strategies has not been examined in any study of which I am aware. The degree of heritability may have profound implications for the evolution of foraging behavior. Consider the situation in which the rate of energy intake has a significant nonheritable component, such as a learned behavior or an environmental effect (e.g., a nutritional component of food that varies among items but cannot be detected). Price et al. (1988) examined optimal evolution of breeding date in birds under a similar set of assumptions. Following their approach, the rate of energy intake can be modeled as
R=R' +n+e (8.9)
where
R' is the heritable component of the foraging rate given by Equations 8.5 and 8.6
n is a nonheritable component due to learning and environment that has variance (J' n 2
e is a random, nonheritable component with zero mean and variance (J'2
The fitness as a function of R can be obtained by assuming that the heritable and nonheritable effects of foraging on fitness are independent so that
W(R,n) = W(R) Wen) (8.10)
That is, since Rand n are independent, the overall fitness function is simply the product of the fitness functions for each effect. Following Price et al. (1988), let the nonheritable effect offoraging on fitness cause an exponential increase in fitness, then the overall fitness is
C/) C/) w Z tLL
C/) C/) W Z tLL
C/) C/) W Z tLL
Optimal Foraging Theory 267
A
B
c R*
RATE OF ENERGY INTAKE (R)
Figure 8.4 Some hypothetical functions relating rate of energy intake to fitness. A and B represent rate-maximizing fitness functions, and C is a nonrate-maximizing function. Rmax is the maximum rate of energy intake possible given a particular set of foraging adaptations (including behavioral, physiological, and morphological traits), and R* is the rate of energy intake that maximizes fitness.
W(R,n) rx exp (an) g(R) (8.11)
where g(R) is the fitness function for the heritable effect and <X is a constant related to the intensity of the nonheritable effect, that is, when <X is small, there is little effect of nonheritable factors on fitness.
The optimal strategy can be found by substituting the expression in Equation 8.9 for R in Equation 8.11 and integrating over nand e to obtain fitness as a func-
268 Energetics of Avian Foraging
tion of R'. I examined numerical solutions for several different fitness functions (g). For rate-maximizing fitness functions, the relationship in Equation 8.9 does not affect the optimal strategy. The nonheritable effect may increase or decrease the intensity of selection leading to the optimum, but not its correspondence with the maximal rate of energy intake. But for nonrate-maximizing fitness functions, when the population reaches the optimal strategy defined by the environment (R* in Fig. 8.4), there will continue to be individuals with nonoptimal strategies that persist. This possibility suggests that as long as there are significant nonheritable effects of foraging on fitness, tests of optimal foraging models may not be capable of quantitative validation. Even if quantitative agreement is obtained in a particular study, it does not necessarily follow that the model is validated until the heritable component of the behavior being studied is documented. Clearly, much more research needs to be done before we have a clear understanding of the relationships between foraging and fitness.
General Classes of Optimal Foraging Models
Optimal foraging models can be divided into several general categories. Here I outline these classes and describe some of major predictions that arise from them.
Perhaps the simplest models of optimal foraging are those that predict the optimal diet of an organism. These models generally assume that the forager encounters prey items one at a time, and bases its decision to attack the prey on the ratio of energy gained and expenditure of time. In relation to Equation 8.1 this assumes that energetic costs of obtaining prey items and physiologically processing them are effectively zero, that there is no interference interactions with other organisms (Maurer 1990 extended this model to include interference interactions), and that time spent in physiological processing of food is nil. The basic result, obtained by a number of workers (e.g., Pulliam 1974; Charnov 1976a), is that the optimally foraging bird will always attack a prey item or never attack it, depending on its profitability. Individuals should never attack an item part of the time: Diet selection is an all-or-none phenomena. This result is based on the assumption that organisms can rank prey items based on their energetic content relative to the time required to obtain them. Pulliam (1975) demonstrated that if a nutrient was in short supply in the diet, the optimal foraging strategy would not be rate maximizing because of the need to preferentially forage on items that had a high nutrient content.
The second class of optimal foraging models are those that predict how an optimally foraging animal will use patches of resources. The model assumes that the forager divides up its time between foraging optimally within a patch and moving between patches. In terms of Equation 8.1, there would be such an equation governing foraging in each patch and the optimal rate of foraging would be determined primarily by the bird's choices about when to leave patches. The basic result, often called the marginal value theorem, states that organisms will forage in
Optimal Foraging Theory 269
a patch until the rate of prey acquisition of that patch falls to the average rate for all patches (Charnov 1976b), i.e., that all patches have prey reduced to the same level by an optimally foraging organism. Dunning (1990) discussed some of the assumptions of patch models and examined tests of these assumptions.
An extension of the patch model that is particularly important to birds are models dealing with central place foraging. In these models, the forager attempt~ to optimize its rate of delivery of energy to a central place, like a nest or storage area (e.g., Orians and Pearson 1979). Central place foragers can be single-prey loaders, that is, they bring only a single prey item back to the central place, such as most raptors, or they can be multiple-prey loaders and bring back several items at the same time. The general prediction that emerges from the single-prey loader models is that the average energetic content of prey items delivered to the nest should increase with distance from the nest that they are captured. Thus, a raptor foraging near its nest could catch smaller prey for its nestlings than a raptor foraging farther from its nest. This is due primarily to the constraint that a bird foraging far away from its nest must be selective and choose only the highest quality prey, while birds foraging nearer the nest need not be as selective to maximize their rate of energy intake. Likewise, multiple prey loaders bringing food to a central place should increase the size of the load (e.g., the number of prey items carried in the bill) when foraging at greater distances from the central place.
Recent Theoretical Results
One of the most important advances in optimal foraging theory are results that extend these "classical" models of optimal foraging in situations where some of their assumptions are not met. For example, foraging birds are assumed by the basic model of diet selection to encounter prey items sequentially (one after the other), so the choice of diet is made by independent decisions to consume single prey items based on their profitability and the rate of intake. If birds encounter more than one prey item at a time, then they must make a decision among several items. The rate maximizing choice in such a situation may be very different from that made for sequential prey encounters (Stephens and Krebs 1986).
Another important idea that has been examined recently is the fact that most of the time, prey items are not identical, so foraging birds must deal with a certain amount of uncertainty regarding the profitability of prey items. If this is the case, the optimally foraging bird must deal with the variability of prey items as well as their average value. Several workers have shown that in this case, an optimal foraging bird will accept a higher degree of variability in prey quality when the average value of the prey item is great than if its average value is small (Caraco 1980; Caraco and Lima 1985; Stephens and Charnov 1982). Selection of the optimal diet or patch choice may not follow the simple rules of basic models under conditions of uncertainty.
270 Energetics of Avian Foraging
Some researchers have pointed out that optimization of the rate of energy intake may be constrained by many factors (e.g., Mangel and Clark 1986; Ludwig and Rowe 1990). For example, while foraging, many species may increase their risk of predation. If this is the case, then the optimal solution to foraging may be to decrease foraging effort when resources (and predators) are abundant (e.g., Abrams 1991). Balancing conflicting effects of foraging and other behaviors on fitness may playa major role in the evolution of attributes that constitute the foraging apparatus of a bird. Most fitness functions relating the rate of energy intake to fitness probably will be nonrate-maximizing for this reason.
Optimal Foraging in Birds
To examine the question of whether foraging behavior in birds tends to be rate maximizing, I reviewed 63 papers containing tests of the three general classes of optimal foraging models discussed above using data on birds. Krebs et al. (1983) and Stephens and Krebs (1986) reviewed 41 studies using birds published before 1985. I obtained 24 additional papers of optimal foraging studies published since their reviews (references available upon request). Some interesting patterns emerge when the cumulative results of these studies are combined.
Of the three types of optimal foraging models, the diet model has been tested more than the other two (Table 8.2). Roughly equal proportions oflaboratory and field studies have been performed. However, patch models have been tested in the laboratory more often than in the field, and central place foraging models have been tested most often in the field. For diet and patch models, experiments are done more often than observational studies, but the opposite is true for central place foraging models.
Studies were classified by their degree of agreement with the models. I recognized three categories: (1) agreement, where the results of the study generally
TABLE 8.2 Results of Empirical Tests of Optimal Foraging Models Using Data on Birds
Methods Level of Support
Agree- Incon-Model Type No. Studies Lab Field Expa Obsa ment Partial sistent
Diet 30 15 15 17 13 7 13 10 Patch 17 14 3 15 2 5 8 4 Central place
lends quantitative support to the model being tested; (2) partial agreement, where the study results were qualitatively or partially consistent with model predictions or assumptions; and (3) inconsistent, where the study results qualitatively falsified the model being tested. Only 14 out of 63 studies (22%) gave quantitative support for an optimal foraging model. The majority of studies were in partial agreement with a model. Surprisingly, 30% of the studies falsified the model being tested. These results were consistent across model types (Table 8.2).
The high rate of falsification of optimal foraging models by studies done using birds might suggest to some that these models tell us little about the energetic basis of avian foraging. However, this conclusion is unwarranted. When falsified, the reasons for model failures are often illuminating in themselves and suggest many times that the fitness functions that govern optimal decisions are often constrained by other aspects of the foraging apparatus to be nonrate-maximizing. This often suggests profitable avenues for further investigation that lead to further insights into the morphological, physiological, and behavioral factors that affect the rate of energy acquisition in birds. The ability to falsify the predictions and assumptions of optimal foraging models is clearly a strength of the general approach because it allows theory and observation to be focused on the same problem. If other questions in ecology could be approached in the same way, we would likely have a much clearer understanding of how ecosystems and communities work.
FUTURE DIRECTIONS FOR RESEARCH
Much more research needs to be done before we come to a clear understanding of the energetics of foraging behavior. The general lines of research I have outlined previously need further work. But just as importantly, it is necessary to begin to integrate different perspectives on the energetics of avian foraging that encompass the wide variety of processes incorporated into the general model in Equation 8.1. This integration will provide answers to some persistent problems in our understanding of the evolution of foraging behavior. Here I outline several of many directions along which this integrative research might proceed.
Physiological Constraints on Foraging
The physiological basis of foraging is currently poorly understood. Emerging understanding of the physiology of avian digestion (e.g., Bell 1990; Karasov 1990) is contributing new information that needs to be integrated into the existing theoretical framework of the energetics offoraging. Speakman (1987) examined the apparent digestive efficiency of two species of shorebirds for several dietary items. Speakman found that prey varied significantly in their content of digestible material. These findings were used to examine the assumptions of the optimal diet model.
272 Energetics of Avian Foraging
The general point made was that if prey items varied in their digestibility, then the predictions made by the diet model could be reversed. That is, if a prey item containing a relatively high amount of energy relative to its ease of capture also had a high amount ofundigestible material, prey items oflower quality might be included in the optimal diet if they were more digestible. Thus, deviations from the predictions of the diet model may sometimes be caused by variations in the actual energy that can be absorbed from items rather than their apparent energy content.
Physiological processes are involved in many aspects of foraging in addition to digestion. Costs of movement and prey handling require the expenditure of energy, and hence the metabolism of energy reserves. Since metabolic rate is so closely tied to body mass, much of interspecific variation in foraging behavior may be directly related to body mass variation among species (see THE ALLOMETRY OF FORAGING BEHAVIOR). However, the effects of variation in body mass among individuals of the same species have received less attention. Some of this intraspecific variation may be directly related to the health or vigor of the individual or its ability to accumulate energy stores, and hence, there may be a sizeable component of such variation that is related to the fitness of an individual and may provide ways to develop new models of foraging energetics (see, e.g., Dunbrack and Giguere 1987).
Fitness, Heritability, and Foraging Behavior
Relatively little attention has been paid to the assumption underlying most foraging models that requires the most fit phenotypes to correspond to rate maximizing strategies (see General Premises of Optimal Foraging). This assumption has received little empirical study, despite its importance. Lemon (1991) experimentally varied the rate of energy intake of four populations of Zebra Finches (Taeniopygia guttata). In populations with higher rates of energy consumption, both survival and fecundity were increased over that of populations fed at lower rates. This finding lends support to the assumed link between fitness and energy intake, and suggests that the fitness function for this species is rate maximizing. Lemon's study has two drawbacks. First, in controlling other factors such as time or energy necessary to avoid predators and variation in the nutrient content of the food, etc., Lemon's experiment does not exclude the possibility that in many natural situations, fitness depends not only on the rate of energy intake but on other factors. Hence, fitness may not be maximized by maximizing the rate of energy intake. Second, there is no assessment of the heritability of energy intake rates. In fact, the rates were imposed on the population by the experimenter, so there was very likely a strong nonheritable component to energy intake rates in the experiment. In another examination of the relationship between energy intake and reproductive success, Pierotti and Annett (1991) found that reproductive success
Future Directions for Research 273
varied significantly among Herring Gulls (Larus argentatus) using different types of diets but that net energy intake on these diets was roughly equivalent.
Lack of information on the heritabilities of foraging traits will continue to limit empirical tests of foraging theories (Raphael and Maurer 1990). Certainly the experimental protocols necessary to begin the assessment of this critical assumption are available and it is surprising that virtually no data exist to test this point. Perhaps the problem is that questions regarding the genetic basis for optimal behaviors have received little attention in discussions of the assumptions of theoretical models of optimization (Maynard Smith 1982; Pyke 1984; Schoener 1987; Parker and Maynard Smith 1990; Mitchell and Valone 1990). Most of these treatments acknowledge that there must be some genetic basis for the traits being studied, but rarely are the problems directly addressed. Schoener (1987) suggested that perhaps foraging behaviors are flexible enough that genetics would have little influence on variability. If so, then the problems with nonheritability discussed above may limit the validity of empirical tests. Mitchell and Valone (1990) argued that the genetic mechanisms that might cause the phenotype to deviate from the optimal solution to a problem may themselves be the objects of natural selection, but there are few good data to evaluate this argument (Orr and Coyne 1992).
Some criticisms of optimal foraging theory arise due to a misunderstanding about the relationship between fitness, heritability, and the phenotype. For example, Gray (1987: 72) criticizes the optimization approach by claiming that the "fittest organisms (best designed) cannot be identified a priori, and hence 'fitness' is tautologically equated with reproductive success." But this misses the point of most optimal foraging models: that design criteria are not based on fitness, but maximization of energetic efficiency. As suggested by analysis of the quantitative genetics of foraging (Equations 8.5-8.10), the functional relationship between the foraging apparatus and rate of energy intake determines the design criteria. There is nothing about fitness contained in these design criteria. A functional relationship between fitness and rate of energy intake defines the relationship between the design criterion and natural selection. Furthermore, this functional relationship does not depend on the particular definition one uses for fitness (e.g., individual reproductive success, intrinsic rate of increase, etc.). This is not circular reasoning, but deductive argumentation at its best. Gray (1987) further criticizes optimization theories because, in his view, they assume that the population is in evolutionary equilibrium. This assumption is also mistaken because optimization models are used to predict the direction of selection and not the final end state of a population (Parker and Maynard Smith 1990). If the environment changes, clearly the fitness functions defining the direction of selection will also change. The population is assumed only to continue to evolve, not to reach an evolutionary equilibrium. Optimal foraging theory is on sound footing logically. Nevertheless, it is apparent that we need both
274 Energetics of Avian Foraging
tighter theoretical formulations and better empirical data on the genetical basis of foraging behavior.
Phylogenetic Patterns of Foraging Adaptations
An approach to studying the process of adaptation that is an alternative to the optimization approach discussed in detail in this paper is the comparative method (Brooks and McLennan 1991, Harvey and Pagel 1991). The general technique is to obtain data on the relatedness among species and then use this to test hypotheses about the sequence of events that led to the current distribution of traits among species. In their study of Phyl/oscopus warblers, Richman and Price (1992) estimated the phylogeny of the genus using molecular data. Using the estimated phylogeny, they used the comparative approach suggested by Felsenstein (1985) to document the relationship between foraging behavior, morphology, and habitat selection. They found that after accounting for similarities due to shared ancestry, there were strong correlations between the morphology, ecology, and foraging behavior of these species. For example, species that had relatively short, wide bills tended to forage less while standing on the foraging substrate.
The analyses reported in this paper (Figs. 8.1-8.3) are comparative in nature. The trends seen in them must be considered provisional until it is clear what contribution phylogeny makes to the patterns observed. For example, the correlation between relative muscle size and body size in the Galapagos finches may simply be fortuitous because the species being studied are all related in some unknown manner. Techniques for comparative analyses exist that can shed light on the evolutionary scenarios that might be responsible for the current relationships between these traits. Some data exist that might be useful for reconstructing the phylogeny of the finches (e.g., Yang and Patterson 1981), but comparative analyses must await further advances in their systematics.
CONCLUSIONS
Foraging plays a key part in the energetic balance of the individual organism. Each organism must maintain a rate of energy intake that allows it to fulfill its maintenance and growth needs. If the rate of energy intake exceeds a certain level, there will be sufficient energy left for reproduction. Hence, natural selection should be a primary mechanism for the evolution offoraging traits. Considerations of the kinds of traits involved in foraging suggest that it is an integrated activity that involves the use of many morphological, physiological, and behavioral characteristics. This integrated set of traits, termed here the foraging apparatus, determines the ability of an individual organism to secure energy. Many times, trade-offs among individual traits that make up the foraging apparatus are apparent from comparative analyses.
References 275
Optimal foraging theory has produced a number of interesting, and more importantly, testable hypotheses that can be applied to the energetics of foraging. However, from a consideration of the quantitative genetics of optimal foraging, it is clear that there are some limitations to the use of these models that researchers should be aware of when designing and interpreting studies intended to test these models. In particular, if there is a significant nonheritable component to the rate of energy intake that affects fitness, the equilibrium foraging strategy could be quite different from the optimal one. This result might be particularly important in field studies that cannot control nonheritable effects. Despite these limitations, optimal foraging should continue to receive theoretical and empirical attention.
Studies testing optimal foraging theories using data on birds indicate that strong agreement with model assumptions and predictions rarely occurs (less than one fourth of all studies obtained strong support for the model being tested). Most often, data were in partial agreement with expectations. This suggests that optimal foraging models provide only approximate solutions to the behavioral problems encountered by foraging birds. Although departures from model predictions and assumptions are often specific to a particular study, it is likely that some of the problems pointed out by the quantitative genetics approach may lead to the kinds of departures from optimal foraging expectations seen in many studies.
The ultimate goal of studies of foraging energetics should be to integrate perspectives from ecology, physiology, behavior, and evolution into a more coherent, predictive theory of foraging energetics of which optimal foraging would be a subset. Some lines along which this integration may occur are studies of the physiological basis of foraging, the heritability of foraging characteristics, and comparative analyses of foraging traits among species. The last line of investigation must wait until better phylogenies are obtained from avian systematists. Nevertheless, the prospects of a more unified approach to avian energetics should encourage ecologists, physiologists, behaviorists, and systematists to pursue integrative studies of the energetics of avian foraging.
REFERENCES
Abrams, P. A. 1991. Life history and the relationship between food availability and foraging effort. Ecology 72:1242-1252.
Bell, G. P. 1990. Birds and mammals on an insect diet: a primer on diet composition analysis in relation to ecological energetics. Stud. Avian Bio!. 13 :416-422.
Boag, P. T. and P. R. Grant. 1981. Intense natural selection in a population of Darwin's Finches (Geospizinae) in the Galapagos. Science 214:82-85.
Boag, P. T., and A. J. van Noorwijk. 1987. Quantitative genetics. In Avian Genetics, eds. F. Cooke and P. A. Buckley, pp. 45-78. Academic Press, London.
276 Energetics of Avian Foraging
Bowman, R. I. 1961. Morphological differentiation and adaptation in the Galapagos finches. Univ. California Publ. Zool. 58: 1-302.
Brooks, D. R. and D. A. McLennan. 1991. Phylogeny, Ecology, and Behavior. The University of Chicago Press, Chicago.
Bryant, D. M. and K. R. Westerterp. 1980. Energetics of foraging and free existence in birds. In Acta XVII Congressus Internationalis Ornithologici, ed. R. von N6hring pp. 292-299. Verlag der Deutschen Ornithologen-Gesellschaft, Berlin.
Calder, W A., III. 1984. Size, Function, and Life History. Harvard University Press, Cambridge, Massachusetts.
Caraco, T. 1980. On foraging time allocation in a stochastic environment. Ecology 61: 119-128.
Caraco, T. and S. L. Lima. 1985. Foraging juncos: interaction of reward mean and variability. Anim. Behav. 33:216-224.
Chamov, E. L. 1976a. Optimal foraging: the attack strategy ofa mantid. Am. Nat. 110:141-151.
Charnov, E. L. 1976b. Optimal foraging: the marginal value theorem. Theor. Pop. BioI. 9:129-136.
Crow, 1. F. 1986. Basic Concepts in Population, Quantitative, and Evolutionary Genetics. W H. Freeman, New York.
Dement, M. Wand P. 1. Van Soest. 1985. A nutritional explanation for body size patterns of ruminant and non-ruminant herbivores. Am. Nat. 125:641-672.
Dunbrack, R. L. and L. A. Giguere. 1987. Adaptive responses to accelerating costs of movement: a bioenergetic basis for the type-III functional response. Am. Nat. 130: 147-160.
Dunning, 1. B., Jr. 1986. Shrub-steppe bird assemblages revisited: implications for community theory. Am. Nat. 128:82-98.
Dunning, 1. B. 1990. Meeting the assumptions of foraging models: an example using tests of avian patch choice. Stud. Avian BioI. 13:462-470.
Faaborg, 1. 1988. Ornithology: an Ecological Approach. Prentice-Hall, Englewood Cliffs, New Jersey.
Falconer, D. S. 1989. Introduction to Quantitative Genetics, 2nd ed, Longman, New York.
Felsenstein,1. 1985. Phylogenies and the comparative method. Am. Nat. 125: 1-15.
Gass, C. L. and W M. Roberts. 1992. The problem of temporal scale in optimization: three contrasting views of hummingbird visits to flowers. Am. Nat. 140:829-853.
Goldstein, D. L. 1990. Energetics of activity and free living in birds. Stud. Avian Bioi. 13:423-426.
Goudie, R. I. and 1. F. Piatt. 1991. Body size and foraging behavior in birds. In Acta XX Congressus Internationalis Ornithologici, ed. B. B. Bell et aI., pp. 811-816. New Zealand Ornithological Congress Trust Board, Wellington, N.Z.
Gould, S. 1. and R. C. Lewontin. 1979. The spandrels of San Marco and the Panglossian paradigm: a critique ofthe adaptationist program. Proc. Royal Soc. London (B) 205:581-598.
Grant, P. R. 1986. Ecology and Evolution of Darwin s Finches. The University of Chicago Press, Chicago.
Gray, R. D. 1987. Faith and foraging: a critique of the "paradigm argument from design." In Foraging Behavior, eds. A. C. Kamil, 1. R. Krebs and H. R. Pulliam, pp. 69-140. Plenum Press. New York.
References 277
Grazal, A, S. D. Strahl, R. Parra, M. G. Dominguez, and A Neher. 1989. Foregut fermentation in the Hoatzin, a neotropicalleaf-eating bird. Science 245:1236-1238.
Greenberg, R. 1990. Ecological plasticity, neophobia, and resource use in birds. Stud. Avian Bioi. 13:431-437.
Grubb, T. c., and L. Greenwald. 1982. Sparrows and a brushpile: foraging responses to different combinations of predation risk and energy cost. Anim. Behav. 30:637-640.
Harvey, P. H. and M. D. Pagel. 1991. The Comparative Method in Evolutionary Biology. Oxford University Press, Oxford.
Helfman, G. S. 1990. Mode selection and mode switching in foraging animals. Adv. Stud. Behav. 19:249-298.
Karasov, W. H. 1990. Digestion in birds: chemical and physiological determinants and ecological implications. Stud. Avian Bioi. 13:391-415.
Kaspari, M. 1990. Prey preparation and the determinants of handling time. Anim. Behav. 40:118-126.
Kooyman, G. L., Y. Cherel, Y. Le Maho, J. P. Croxall, P. H. Thorson, V. Ridoux, and C. A Kooyman. 1992. Diving behavior and energetics during foraging cycles in king penguins. Ecol. Monogr. 62:143-163.
Krebs, J. R., D. W. Stephens, and W. J. Sutherland. 1983. Perspectives in optimal foraging. In Perspectives in Ornithology, eds. A. H. Brush and G. A. Clark, Jr., pp. 165-216. Cambridge University Press, New York.
Lemon, W. C. 1991. Fitness consequences of foraging behaviour in the zebra finch. Nature 352: 153-155.
Lewontin, R. C. 1979. Fitness, survival, and optimality. In Analysis of Ecological Systems. eds. D. J. Hom, R. D. Mitchell, and G. R. Stairs, pp. 3-21. Ohio State University Press, Columbus, Ohio.
Lima, S. L. 1985. Maximizing feeding efficiency and minimizing time exposed to predators: a trade-off in the black-capped chickadee. Oecologia 66:60-67.
Lima, S. L. 1987. Initiation and termination of daily feeding in dark-eyed juncos: influences of predation risk and energy reserves. Oikos 53:3-11.
Lotka, A J. 1922a. A contribution to the energetics of evolution. Proc. Nat. Acad. Sci. (USA) 8:147-150.
Lotka, A J. 1922b. Natural selection as a physical principle. Proc. Nat. Acad. Sci. (USA) 8:151-153.
Lotka, A. J. 1925. Principles of Physical Biology. Williams and Wilkins, Baltimore.
Ludwig, D. and L. Rowe. 1990. Life-history strategies for energy gain and predator avoidance under time constraints. Am. Nat. 135 :686-707.
Mangel, M. and C. W. Clark. 1986. Towards a unified foraging theory. Ecology 67: 1127-1138.
Maurer, B. A 1980. Avian Foraging and Habitat Structure in an Eastern Deciduous Forest in West Virginia. MS thesis, West Virginia University, Morgantown.
Maurer, B. A 1990. Extensions of optimal foraging theory for insectivorous birds: implications for community structure. Stud. Avian Bioi. 13:455-461.
Maurer, B. A and J. H. Brown. 1988. Distribution of biomass and energy use among species of North American terrestrial birds. Ecology 69: 1923-1932.
278 Energetics oj Avian Foraging
Maurer, B. A. and R. C. Whitmore. 1981. Foraging of five bird species in two forests with different vegetation structure. Wilson Bull. 93:478-490.
Maynard Smith, 1. 1978. Optimization theory in evolution. Ann. Rev. Eco!. Syst. 9:31-56.
Maynard Smith, 1. 1982. Evolution and the Theory of Games. Cambridge University Press, Cambridge.
McNamara, 1. M. and A. I. Houston. 1986. The common currency for behavioral decisions. Am. Nat. 127:358-378.
Mitchell, W. A. and T. 1. Valone. 1990. The optimization research program: studying adaptations by their function. Q. Rev. Bioi. 65:43-52.
Morse, D. H. 1971. The insectivorous bird as an adaptive strategy. Ann. Rev. Eco!. Syst. 2:177-200.
Myers, 1. P. 1983. Commentary. In Perspectives in Ornithology, eds. A. H. Brush and G. A. Clark, Jr., pp. 216-22l. Cambridge University Press, New York.
Nagy, K. A. 1987. Field metabolic rate and food requirement scaling in mammals and birds. Eco!. Monogr. 57: 111-128.
Nagy, K. A. and B. S. Obst. 1991. Body size effects on field energy requirements of birds: what determines their field metabolic rates? In Acta XX Congressus Internationalis Ornithologici, ed. B. B. Bell et aI., pp. 793-799. New Zealand Ornithological Congress Trust Board, Wellington, N.Z.
Nonacs, P. and L. M. Dill. 1993. Is satisficing an alternative to optimal foraging theory? Oikos 67:371-375.
Orians, G. H. and N. E. Pearson. 1979. On the theory of central place foraging. In Analysis of Ecological Systems, eds. D. 1. Hom, R. D. Mitchell, and G. R. Stairs, pp. 155-177. Ohio State University Press, Columbus.
Orr, H. A. and 1. A. Coyne. 1992. The genetics of adaptation: a reassessment. Am. Nat. 140:725-742.
Osterhaus, O. B. 1962. Adaptive modifications in the leg structure of some North American warblers. Am. Mid!. Nat. 68:474-486.
Parker, G. A. and 1. Maynard Smith. 1990. Optimality theory in evolutionary biology. Nature 348:27-33.
Pennycuick, C. 1. 1975. Mechanics of flight. In Avian Biology, vol. 5, eds. D. S. Farner and 1. R. King, pp. 5-17. Academic Press, New York.
Peters, R. H. 1983. The Ecological Implications of Body Size. Cambridge University Press, Cambridge.
Pierotti, R., and C. A. Annett. 1991. Diet choice in the herring gull: constraints imposed by reproductive and ecological factors. Ecology 72:319-328.
Price, T., M. Kirkpatrick, and S. 1. Arnold. 1988. Directional selection and the evolution of breeding date in birds. Science 240:798-799.
Pulliam, H. R. 1974. On the theory of optimal diets. Am. Nat. lO8:59-75.
Pulliam, H. R. 1975. Diet optimization with nutrient constraints. Am. Nat. lO9:765-768.
Pulliam, H. R. 1980. Do chipping sparrows forage optimally? Ardea 68:75-82.
Pyke, G. H. 1984. Optimal foraging theory: a critical review. Ann. Rev. Eco!. Syst. 15:523-575.
References 279
Pyke, G. H., H. R. Pulliam, and E. L. Charnov. 1977. Optimal foraging: a selective review of theory and tests. Q. Rev. Bioi. 52:137-154.
Raphael, M. G. and B. A. Maurer. 1990. Biological considerations for study design. Stud. Avian Bioi. 13:123-125.
Remsen, J. v., Jr. and S. K. Robinson. 1990. A classification scheme for foraging behavior of birds in terrestrial habitats. Stud. Avian Bioi. 13: 144-160.
Richman, A. D. and T. Price. 1992. Evolution of ecological differences in the Old World leaf warblers. Nature 355:817-821.
Robbins, C. T. 1983. Wildlife Feeding and Nutrition. Academic Press, New York.
Rudolph, S. G. 1982. Foraging strategies of American kestrels during the breeding season. Ecology 63: 1268-1276.
Schoener, T. W 1971. Theory offeeding strategies. Ann. Rev. Ecol. Syst. 2:369-404.
Schoener, T. W 1987. A brief history of optimal foraging theory. In Foraging Behavior eds. A. C. Kamil, J. R. Krebs, and H. R. Pulliam, pp. 5-67. Plenum Press, New York.
Sherry, T. W 1990. When are birds dietarily specialized? Distinguishing ecological from evolutionary approaches. Stud. Avian BioI. 13:337-353.
Speakman, J. R. 1987. Apparent absorption efficiencies for redshank (Tringa totanus L.) and oystercatcher (Haematopus ostralegus L.): implications for the predictions of optimal foraging models. Am. Nat. 130:677-691.
Stalmaster, M. V. and J. A. Gessaman. 1984. Ecological energetics and foraging behavior of overwintering bald eagles. Ecol. Monogr. 54:407-428.
Steams, S. C. 1992. The Evolution of Life Histories. Oxford University Press, Oxford.
Stephens, D. W 1990. Foraging theory: up, down, and sideways. Stud. Avian Bioi. 13 :444-454.
Stephens, D. Wand E. L. Charnov. 1982. Optimal foraging: some simple stochastic mod-els. Behav. Ecol. Sociobiology 10:251-263.
Stephens, D. W and J. R. Krebs. 1986. Foraging Theory. Princeton University Press, Princeton, New Jersey.
Walsberg, G. E. 1983. Avian ecological energetics. In Avian Biology, vol. 7, eds. D. S. Farner, J. R. King, and K. C. Parkes, pp. 161-220. Academic Press, New York.
Ward, D. 1992. The role of satisficing in foraging theory. Oikos 49: 188-125.
Ward, D. 1993. Foraging theory, like all other fields of science, needs multiple working hypotheses. Oikos 67:376-378.
Watt, W. B. 1985. Bioenergetics and evolutionary genetics: opportunities for a new synthesis.Am. Nat. 125:118-143.
Wiens, J. A. 1976. Population responses to patchy environments. Ann. Rev. Ecol. Syst. 7 :81-120.
Yang, S. Y. and J. L. Patterson, 1981. Genic variability and differentiation in the Galapagos finches. Auk 98:230-242.
Valone, T. J., and S. L. Lima. 1987. Carrying food items to cover for consumption: the behavior of ten bird species under the risk of predation. Oecologia 71 :286-294.