B 1 and B 2 kinin receptor participation in hyperproliferative and inflammatory skin processes in mice Evelise Fernandes Pietrovski a , Ka ´ tia Sabrina Paludo b , Daniel Augusto Gasparin Bueno Mendes a , Fernando de Souza Fonseca Guimara ˜es b , Silvio Sanchez Veiga b , Dorly de Freitas Buchi b , Raphael Gomes Fonseca c , Aleksander Roberto Zampronio a , Michael Bader d , Joa ˜o Bosco Pesquero c , Juliano Ferreira e , Michel Fleith Otuki f , Daniela Almeida Cabrini a, * a Department of Pharmacology, Universidade Federal do Parana ´, Curitiba, PR, Brazil b Department of Cellular Biology, Universidade Federal do Parana ´, Curitiba, PR, Brazil c Department of Biophysics, Universidade Federal de Sa ˜o Paulo, Sa˜o Paulo, SP, Brazil d Max-Delbru ¨ck-Centre for Molecular Medicine (MDC), Wiltbergstr, Berlin-Buch, Germany e Department of Chemistry, Universidade Federal de Santa Maria, Santa Maria, RS, Brazil f Department of Pharmaceutical Sciences, Universidade Estadual de Ponta Grossa, Ponta Grossa, PR, Brazil 1. Introduction Hyperproliferative epidermal diseases range from benign hyperplasias such as psoriasis, to cell carcinomas [1]. Cell carcinomas (squamous and basal) and actinic keratoses represent common hyperproliferative disorders associated with aberrant keratinocyte differentiation [2]. Psoriasis is probably the most prevalent immune-mediated hyperproliferative skin disorder associated with abnormal epidermal differentiation accompanied by an inflammatory infiltrate of T lymphocytes, macrophages, and dendritic cells, thereby manifesting it from head to toe in genetically susceptible individuals as red thick scaly plaques [3– 6]. Traditional treatments for the disease have several short- comings, including inconvenience (as is the case with topical therapy and phototherapy) and toxicity (as is the case with systemic treatment) [7]. A better understanding of the inflamma- tory nature of psoriasis and a further characterisation of epidermal homeostasis should provide clues to the prevention and treatment of these cutaneous diseases [6]. Journal of Dermatological Science 64 (2011) 23–30 A R T I C L E I N F O Article history: Received 19 April 2011 Received in revised form 22 June 2011 Accepted 28 June 2011 Keywords: Kinin receptor Kinin receptor knockout mice SSR 240612 FR 173657 Epidermis hyperproliferation A B S T R A C T Background: Kinins are released during dermal injury and inflammation and seem to contribute to the pathogenesis of cutaneous diseases. Objective: Participation of kinins in skin inflammatory process was evaluated using knockout mice and non-peptide kinin receptor antagonists. Methods: Chronic skin inflammation was induced by multiple applications of TPA in mice ear. Results: The B 2 knockout mice (B 2 À/À ) showed a significant increase of ear weight (23 10%) and epidermal cellular hyperproliferation and acanthosis formation upon histological analysis when compared with wildtype mice. Also, evaluation of PCNA levels by Western blot and immunohistochemistry confirmed the increase in the epidermis hyperproliferation in the ear skin of B 2 À/À mice. In contrast, no modification in these parameters was detected in B 1 knockout mice (B 1 À/À ). However, mice lacking both kinin receptors (B 1 B 2 À/À ) presented a considerable reduction of epidermis thickness and in PCNA levels. Following the establishment of skin inflammation (5th day of TPA application) treatment with the non-peptide antagonists SSR 240612 (B 1 receptor antagonist), FR 173657 (B 2 receptor antagonist), or SSR 240612 plus FR 173657 topically applied, caused a significant inhibition of ear weight (20 5%, 34 4% and 32 6%, respectively). In the histological analysis, the antagonists produced a reduction in epidermal hyperplasia and acanthosis formation; but the treatment with a combination of the two antagonists did not increase efficacy. Conclusion: Kinin receptors seem to be involved in the control of the keratinocyte hyperproliferative process, and non-peptide kinin receptor antagonists may be useful tools in the treatment of hyperproliferative skin disorders. ß 2011 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights reserved. * Corresponding author at: Departamento de Farmacologia, Setor de Cie ˆncias Biolo ´ gicas, Universidade Federal do Parana ´, Centro Polite ´ cnico, Jardim das Ame ´ ricas, P.O. Box 19031, Curitiba 81530-900, Parana ´, Brazil. Tel.: +55 41 3361 1539; fax: +55 41 3266 2042. E-mail address: [email protected](D.A. Cabrini). Contents lists available at ScienceDirect Journal of Dermatological Science jou r nal h o mep ag e: w ww .elsevier .co m /jds 0923-1811/$36.00 ß 2011 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.jdermsci.2011.06.016
Transcript
Journal of Dermatological Science 64 (2011) 23–30
B1 and B2 kinin receptor participation in hyperproliferative and inflammatory skinprocesses in mice
Evelise Fernandes Pietrovski a, Katia Sabrina Paludo b, Daniel Augusto Gasparin Bueno Mendes a,Fernando de Souza Fonseca Guimaraes b, Silvio Sanchez Veiga b, Dorly de Freitas Buchi b,Raphael Gomes Fonseca c, Aleksander Roberto Zampronio a, Michael Bader d,Joao Bosco Pesquero c, Juliano Ferreira e, Michel Fleith Otuki f, Daniela Almeida Cabrini a,*a Department of Pharmacology, Universidade Federal do Parana, Curitiba, PR, Brazilb Department of Cellular Biology, Universidade Federal do Parana, Curitiba, PR, Brazilc Department of Biophysics, Universidade Federal de Sao Paulo, Sao Paulo, SP, Brazild Max-Delbruck-Centre for Molecular Medicine (MDC), Wiltbergstr, Berlin-Buch, Germanye Department of Chemistry, Universidade Federal de Santa Maria, Santa Maria, RS, Brazilf Department of Pharmaceutical Sciences, Universidade Estadual de Ponta Grossa, Ponta Grossa, PR, Brazil
A R T I C L E I N F O
Article history:
Received 19 April 2011
Received in revised form 22 June 2011
Accepted 28 June 2011
Keywords:
Kinin receptor
Kinin receptor knockout mice
SSR 240612
FR 173657
Epidermis hyperproliferation
A B S T R A C T
Background: Kinins are released during dermal injury and inflammation and seem to contribute to the
pathogenesis of cutaneous diseases.
Objective: Participation of kinins in skin inflammatory process was evaluated using knockout mice and
non-peptide kinin receptor antagonists.
Methods: Chronic skin inflammation was induced by multiple applications of TPA in mice ear.
Results: The B2 knockout mice (B2�/�) showed a significant increase of ear weight (23 � 10%) and
epidermal cellular hyperproliferation and acanthosis formation upon histological analysis when compared
with wildtype mice. Also, evaluation of PCNA levels by Western blot and immunohistochemistry confirmed
the increase in the epidermis hyperproliferation in the ear skin of B2�/� mice. In contrast, no modification in
these parameters was detected in B1 knockout mice (B1�/�). However, mice lacking both kinin receptors
(B1B2�/�) presented a considerable reduction of epidermis thickness and in PCNA levels. Following the
establishment of skin inflammation (5th day of TPA application) treatment with the non-peptide antagonists
SSR 240612 (B1 receptor antagonist), FR 173657 (B2 receptor antagonist), or SSR 240612 plus FR 173657
topically applied, caused a significant inhibition of ear weight (20 � 5%, 34 � 4% and 32 � 6%, respectively). In
the histological analysis, the antagonists produced a reduction in epidermal hyperplasia and acanthosis
formation; but the treatment with a combination of the two antagonists did not increase efficacy.
Conclusion: Kinin receptors seem to be involved in the control of the keratinocyte hyperproliferative
process, and non-peptide kinin receptor antagonists may be useful tools in the treatment of
hyperproliferative skin disorders.
� 2011 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rightsreserved.
Contents lists available at ScienceDirect
Journal of Dermatological Science
jou r nal h o mep ag e: w ww .e lsev ier . co m / jds
1. Introduction
Hyperproliferative epidermal diseases range from benignhyperplasias such as psoriasis, to cell carcinomas [1]. Cellcarcinomas (squamous and basal) and actinic keratoses representcommon hyperproliferative disorders associated with aberrant
* Corresponding author at: Departamento de Farmacologia, Setor de Ciencias
Biologicas, Universidade Federal do Parana, Centro Politecnico, Jardim das
0923-1811/$36.00 � 2011 Japanese Society for Investigative Dermatology. Published b
doi:10.1016/j.jdermsci.2011.06.016
keratinocyte differentiation [2]. Psoriasis is probably the mostprevalent immune-mediated hyperproliferative skin disorderassociated with abnormal epidermal differentiation accompaniedby an inflammatory infiltrate of T lymphocytes, macrophages, anddendritic cells, thereby manifesting it from head to toe ingenetically susceptible individuals as red thick scaly plaques [3–6]. Traditional treatments for the disease have several short-comings, including inconvenience (as is the case with topicaltherapy and phototherapy) and toxicity (as is the case withsystemic treatment) [7]. A better understanding of the inflamma-tory nature of psoriasis and a further characterisation of epidermalhomeostasis should provide clues to the prevention and treatmentof these cutaneous diseases [6].
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–3024
Schremmer-Danninger et al. [8] have demonstrated that kinincomponents (tissue kallikrein, B1 and B2 receptors) are constitu-tively present in normal human skin. Moreover, low molecularweight kininogen mRNA increases and both kinin receptors areupregulated in psoriatic skin. Later, the same authors were able tolocalise kinin components in normal, traumatised (surgery),basalioma and lichenificated skin [9]. Also, we have previouslyshowed that both kinin receptors seemed to be important inneurogenic cutaneous inflammatory responses [10]. These findingssupport the possibility that the kinin system serves an importantrole in skin homeostasis and in pathological states. However, therole of kinins in the inflammatory process is poorly understood, butit is probable that kinins contribute to the pathogenesis ofcutaneous diseases such as psoriasis [11].
Kinins are peptide mediators that are involved in a series ofpathophysiological processes. They exert most of their effects byinteracting with two classes of G-protein-coupled receptors calledB1 and B2 [12]. The B2 receptor mediates the action of bradykinin(BK) and kallidin or lysyl-bradykinin (Lys-BK), which are the firstset bioactive kinins formed in response to injury from kininogenprecursors through the action of plasma and tissue kallikreins.Whereas the B1 receptor mediates the effects of des-Arg9-BK andLys-des-Arg9-BK, the second set bioactive kinins which are formedthrough the actions of carboxypeptidases on BK and Lys-BK,respectively [13]. It has been demonstrated that B2 receptors arewidely expressed in a constitutive manner throughout mostmammalian tissues. By contrast, B1 are not constitutivelyexpressed to a great extent under normal conditions, but areupregulated following inflammatory, infectious or traumaticstimuli [14]. However, as already mentioned, the B1 receptor isconstitutively expressed in normal skin [8,15]. Furthermore, BKand Lys-BK are released during dermal injury and inflammation asa result of activation of kallikreins. In the skin, kinins play a role inthe mediation of cutaneous responses such as reactive cellproliferation, pain and inflammation [8].
Therefore, we aimed to evaluate the role of the kinins inhyperproliferative and chronic inflammatory processes in skinusing non-peptide kinin receptor antagonists and kinin receptorknockout mice for a better understanding of the mechanismsunderlying epidermal proliferation that might contribute to thedesign of new optimal treatments for psoriasis.
2. Materials and methods
2.1. Reagents
The following reagents were used: 12-O-tetradecanoylphorbol-acetate (TPA), dexamethasone acetate, phenylmethylsulphonylfluoride, dithiothreitol, leupeptin, pepstatin A, aprotinin, soybeantrypsin inhibitor, sodium orthovanadate, phosphate-bufferedsaline (PBS), 4-(2-hydroxyethyl)-1-piperazineethanesulphonic ac-id (HEPES), Tween-20, glycerol, b-mercaptoethanol, dimethylsulphoxide (DMSO), bromophenol blue, bovine serum albumin(BSA) (Sigma Chemical Co., St. Louis, USA); NaCl, sodium fluoride,ethylenediamine tetraacetic acid (EDTA), acetone, formaldehyde,acetic acid, Tris–HCl, MgCl2, methanol (Merck and Co., Inc.,Whitehouse Station, USA); hydrogen peroxide (H2O2), eosin,hematoxylin, absolute ethanol, floxine B, xylol, propylene glycol(Vetec, Rio de Janeiro, Brazil); anti-PCNA polyclonal antibodies,horseradish peroxidase (HRP)-conjugated secondary antibodies(Santa Cruz Biotech. Inc., Santa Cruz, USA); DAB substrate kit,mouse IgG1 immunoglobulin isotype control (BD Pharmingen, SanJose, CA, USA). SSR 240612 and FR 173657 were kindly provided bySanofi-Aventis (Germany) and Astellas Pharma Inc. (Japan),respectively. Moreover, these non-peptide antagonists were pre-
solubilised in DMSO (2%), propylene glycol (15%) and ethanol(33%), after being diluted in acetone (50% of final solution).
2.2. Animals
Experiments were conducted using male and female Swiss miceor wild type mice (C57BL/6), B1 receptor knockout mice (B1
�/�), B2
receptor knockout mice (B2�/�) or double kinin receptor knockout
mice (B1B2�/�) (20–30 g) housed at 22 � 2 8C under a 12-h light/12-
h dark cycle and with access to food and water ad libitum. Theseanimals were allowed to adapt to the laboratory for at least 2 h beforetesting. Swiss mice were obtained at the Sector of Biological Sciences,Universidade Federal do Parana (UFPR, Curitiba, Brazil). WildtypeC57BL/6, B1
�/�, B2�/� or B1B2
�/�were obtained from the Departmentof Biophysics (UNIFESP, Sao Paulo, Brazil). Deletion of the entirecoding sequence for B1, B2 and both kinin receptors was achievedaccording to the methodologies described previously by Pesquero etal. [16], Rupniak et al. [17] and Cayla et al. [18], respectively.Experiments were performed after approval of the protocol by theInstitutional Ethics Committee and were carried out in accordancewith the current guidelines for the care and ethical treatment oflaboratory animals. Treatments were performed at 12 � 2 h. Datafrom 4 to 8 animals per group were analysed.
2.3. Mouse model of chronic skin inflammation
The chronic skin inflammation was induced by application of2.0 mg/ear of 12-O-tetradecanoylphorbol-13-acetate (TPA) dis-solved in 20 ml of acetone to the right ear. TPA was applied onalternating days over the course of 9 days in B1
�/�, B2�/�, B1B2
�/�,wildtype animals or in Swiss mice [19]. The non-peptideantagonists SSR 240612 (non-peptide B1 receptor antagonist),FR 173657 (non-peptide B2 receptor antagonist), and thecombination of SSR 240612 plus FR 173657 were applied topically(0.1 mg/ear) in Swiss mice, twice daily, during the last 4 days ofthe experiment. Dexamethasone was used as the reference drug(0.1 mg/ear) and was applied in the same manner as the non-peptide antagonists. At the end of the chronic experiment, animalswere sacrificed and samples (6 mm punch) were removed fromthe right ear and the biopsies were individually weighed.Subsequently, the samples were stored at �80 8C until furtheranalysis.
2.4. Preparation of nuclear fractions
The tissues were prepared as previously described for Medeiroset al. [20] with some modifications. In brief, tissues werehomogenised in ice-cold buffer A (10 mM HEPES, pH 7.4;containing 1.5 mM MgCl2, 10 mM KCl, 0.5 mM phenylmethylsul-phonyl fluoride, 5 mg/ml leupeptin, 1.5 mg/ml trypsin inhibitor,7 mg/ml pepstatin A, 1 mg/ml leupeptin, 10 mg/ml aprotinin, 2 mMsodium orthovanadate, 50 mM sodium fluoride and 0.5 mMdithiothreitol). The homogenates were chilled on ice undervigorous shaking for 15 min. The nuclear fraction was precipitatedby centrifugation at 10,000 � g for 30 min. The supernatantcontaining the cytosolic fraction was stored at �80 8C until use.The membrane pellet was resuspended in buffer B (20 mM HEPES,pH 7.4; containing 1.5 mM MgCl2, 420 mM NaCl, 0.2 mM EDTA,25% (v/v) glycerol, 0.5 mM phenylmethylsulphonyl fluoride,0.5 mM dithiothreitol, 5 mg/ml trypsin inhibitor, 7 mg/ml pepsta-tin A, 5 mg/ml pepstatin A, and 10 mg/ml aprotinin) and incubatedwith continuous shaking at 4 8C for 30 min. The nuclear fractionwas then centrifuged for 30 min at 10,000 � g and the supernatantwas aliquoted and stored at �80 8C. The protein concentration wasdetermined using a Bradford Protein Assay (Bio-Rad Laboratories,USA).
Fig. 1. The effect of the absence of kinin receptors on the oedema caused by TPA
multiple applications in the ear skin. Ear weight after multiple topical applications
of TPA in wildtype (+/+), kinin B1 receptor knockout (B1�/�), kinin B2 receptor
knockout (B2�/�) and double knockout (B1B2
�/�mice). Bars show the mean � S.E.M.
for 6–8 animals. Significantly different from wildtype, **P < 0.01, ***P < 0.001.
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–30 25
2.5. Evaluation of proliferating cell nuclear antigen (PCNA) levels by
Western blot
Cell lysates of nuclear fraction (30 mg protein) were boiled insodium dodecyl sulphate (SDS) sample buffer for 5 min beforeelectrophoresis on 12.5% SDS–polyacrylamide gel. After transfer-ring to a polyvinylidene fluoride (PVDF) membrane (GE Health-care), the blots were blocked with a 5% fat-free dry milk-TBSTbuffer (Tris-buffered saline containing 0.5% Tween-20) for 1 h atroom temperature and then washed with TBST buffer. Themembranes were incubated for 2 h at room temperature with1:200 dilutions of primary antibodies for PCNA (Santa CruzBiotech. Inc., USA). Blots were washed three times with TBST at5 min intervals followed by incubation with a 1:500 dilution of theappropriate horseradish peroxidase conjugated secondary anti-bodies for 1 h and again washed in TBST three times. Thetransferred proteins were visualised with an Enhanced Chemilu-minescence (ECL) Detection Kit according to the manufacturer’sinstructions (Amersham Pharmacia Biotech, England).
2.6. Evaluation of PCNA levels by immunohistochemistry
Ear samples were fixed in ALFAC (formaldehyde, ethanol 80%,glacial acetic acid) solution. After washing with PBS, ears weredehydrated using increasing ethanol concentrations, xylene andembedded in paraffin. The samples were sectioned onto silanecoated slides and the embedded tissue sections were deparaffi-nised in xylene and rehydrated. The endogenous peroxidaseactivity in skin tissue was quenched in 3% H2O2/methanol for10 min and rinsed twice for 5 min each in PBS. The nonspecific siteswere blocked in 1% bovine serum albumin (BSA) in PBS for 30 minfollowed by incubation with primary antibody for 2 h (1:50dilution of anti-PCNA goat polyclonal antibody, Santa Cruz Biotech.Inc., USA). The sections were then washed and incubated withhorseradish peroxidase (HRP)-conjugated secondary antibodiesfor 1 h. The colour reaction was observed using a DAB substrate kit(BD Pharmingen, San Jose, CA, USA). The sections were counter-stained with Harris hematoxylin, washed in tap water, dehydratedthrough alcohol and xylene, and mounted using permount.Individual sections were examined by light microscopy (40�) toquantitate the immunopositive reaction. The relative numbers ofimmunohistochemically stained PCNA protein were determined inpixels using the Image J1 software (NIH, USA).
2.7. Histology
Ear samples were fixed in ALFAC solution. Each sample was cutlongitudinally into equal halves. Half of each was embedded inparaffin, cut into 5 mm sections and stained with hematoxylin–eosin. Sections were scanned under the microscope and arepresentative area was selected for qualitative light microscopicanalysis of inflammatory and hyperproliferative cellular responsewith 20� and 40� objectives.
Epidermal thickness (considered as the distance from the basallayer to the stratum corneum, and also all formed acanthosis inslice), and the skin thickness (considered as the distance betweenone stratum corneum up to the other) were randomly measured on6 different visual areas on each slice [3–5] using Adobe Photo-shop1 software. The mean value was then calculated for eachsample.
2.8. Statistical analysis
The results are presented as mean � S.E.M. The percentagesvalues for each individual group are calculated as mean � S.E.M.compared with the percentage values for wildtype mice group or
Swiss mice control group after chronic treatment with TPA. Data weresubjected to analysis of variance (ANOVA) complemented by New-man–Keul’s post hoc test. P < 0.05 was considered as indicative ofsignificance.
3. Results
3.1. TPA-induced mouse ear oedema in knockout mice
Phorbol esters, like TPA, have been demonstrated to induce skininflammation and hyperproliferative responses in animals [21].Accordingly, repeated topical applications of TPA induced an equalincrease in ear weight in wildtype mice, B1
�/� or B1B2�/� mice
(Fig. 1). Also, no difference in ear weight was detected on basalsamples from wildtype and knockout mice. In contrast, the B2
�/�
mice exhibited an additional increase in ear weight whencompared with the wildtype mice (23 � 10%) after chronictreatment with TPA. Following the establishment of the inflammatoryprocess, chronic treatment with dexamethasone (0.1 mg/ear) pro-moted a complete inhibition of the oedema (ear weight) on day 9 ofTPA treatment. Fig. S1 exemplifies representative pictures of the earsafter repeated TPA applications in wildtype, B1
�/�, B2�/�, and B1B2
�/�
mice.The histological analysis showed that repeated treatment with
vehicle caused no alteration in cutaneous morphology (Fig. 2A–D).Multiple applications of TPA induced an intense oedema formationand epidermal hyperproliferation accompanied by projection ofthe epidermis into the dermis, which is a process known asacanthosis, well-observed in the photo from wildtype mice(Fig. 2E), as well as in samples from all transgenic animals(Fig. 2F–H). Actually, no difference of total skin thickness wasdetected between groups, except for dexamethasone treatment(reduction of 45 � 3%) (Fig. 2I). The increase in epidermal thicknesswas 44 � 12% greater in B2
�/� mice when compared to the wildtypegroup (Fig. 2J). Moreover, the lack of both kinin receptors (B1B2
�/�
mice) presented a significant reduction of epidermal width (21 � 5%)(Fig. 2J), and minor acanthosis formation (Fig. 2H), in response torepeated applications of TPA when compared to wildtype mice.Dexamethasone treatment shows a decrease of 56 � 2% in epidermalthickness (Fig. 2J).
3.2. Evaluation of PCNA levels by Western blot and
immunohistochemistry
PCNA protein quantification showed that wildtype, B1�/� and
B2�/� mice exhibited an increase in protein expression on the 9th
day after chronic TPA treatment (Fig. 3). However, the PCNAexpression was significantly enhanced in B2
�/� mice whencompared with wildtype mice (58 � 6%). In contrast, B1B2
�/� mice
Fig. 2. Histological analysis after chronic TPA-induced inflammation in kinin receptor knockout mice. (A–H) Representative pictures of histological sections from the ears of
mice (20�, HE-stained), (I) measurement of skin thickness and (J) measurement of epidermis thickness. Ears of wildtype (+/+) (A), kinin B1 receptor knockout (B1�/�) (B), kinin
B2 receptor knockout (B2�/�) (C) and double knockout (B1B2
�/�) (D) mice treated with vehicle. Ears of wildtype (+/+) (E), kinin B1 receptor knockout (B1�/�) (F), kinin B2
receptor knockout (B2�/�) (G) and double knockout (B1B2
�/�) (H) mice treated with multiple TPA applications. Bars show the mean � S.E.M. for 3–4 sections. Significantly
different from wildtype, *P < 0.05, ***P < 0.001.
Fig. 3. The effect of the absence of kinin receptors on the epidermis cell proliferation
after TPA multiple applications in the ear skin. Evaluation of PCNA expression in
�) and double knockout (B1B2�/�) mice. (A) Western blot analysis of PCNA
expression was performed after chronic TPA treatment. (B) Quantification of PCNA
protein expression. Each bar represents the mean � S.E.M. for 2 independent
experiments. Significantly different from wildtype, *P < 0.05.
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–3026
exhibited a significant decrease in PCNA protein (76 � 7%) at the endof the chronic TPA treatment, indicating a minor change in cellularproliferation in mice deficient of both kinin receptors (Fig. 3).
The changes in PCNA expression caused by repeated TPAapplications in the skin were also evaluated with immunohisto-chemical technique (Fig. 4). In the ear of wildtype mice, apronounced staining for PCNA was observed, thereby indicating anincrease of cellular proliferation in the epidermis of these animals.Similar result was observed in the ear skin from mice without B1
receptors (B1�/�) after repeated applications of TPA. On the other
hand, in the skin ear of B2�/� mice, the PCNA staining was
demonstrated to be significantly enlarged (36 � 16%), while theB1B2
�/� mice exhibited reduced staining for PCNA (43 � 12%). Thesefindings confirm the previous results demonstrating a decrease incellular proliferation in tissue deficient of both kinin receptors. Also,in all knockout and wildtype mice, the PCNA staining was highlyspecific in the basal layer of the epidermis with no detectabledifferences among them (Fig. 4).
3.3. Effect of non-peptide kinin antagonists on multiple TPA skin
applications in the mouse ear
After the establishment of the chronic inflammatory process inthe skin of the ear, the chronic topical application of the non-peptide kinin antagonists SSR 240612 (B1 receptor antagonist) orFR 173657 (B2 receptor antagonist) (0.1 mg/ear) caused asignificant inhibition of ear weight gain (Fig. 5). The calculated
Fig. 4. Representative pictures of PCNA staining in the skin of knockout mice after multiple applications of TPA in the ear. Representative pictures of PCNA
immunohistochemical of transverse slices of hematoxylin-stained ears (increase of 400�, scale of 250 mm) from knockout mice at basal or collected on day 9 after the
multiple applications of TPA (2.0 mg/ear). Bars show the mean � S.E.M. for the quantification of 5 different areas (40�). Significantly different from wildtype, **P < 0.01,
***P < 0.001.
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–30 27
inhibition value for SSR 240612 was 20 � 5%, while the calculatedinhibition for FR 173657 was 34 � 4%. The combination treatment ofFR 173657 and SSR 240612 (0.1 mg/ear) did not cause an increase inthe inhibitory effect (32 � 6%) on the TPA-induced ear oedema whencompared to the results of a single drug treatment. Moreover, topicalapplication of vehicle did not alter the ear weight when compared tobasal group that received no treatment (Fig. 5). Fig. S2 exemplifiesrepresentative pictures of the ears of mice after repeated TPAapplication that directly illustrated the effect of non-peptide kininantagonists on reducing ear oedema.
Next, we investigated H&E-stained ear sections from Swissmice treated with non-peptide kinin antagonists (SSR 240612, FR
Fig. 5. Effect of topical treatment with kinin receptor antagonists during multiple
applications of TPA in the ear. Topical effect of non-peptide kinin receptor
antagonists SSR 240612 (B1 receptor antagonist), FR 173657 (B2 receptor
antagonist), SRR 240612 plus FR 173657, and vehicle (V) on ear weight after
multiple topical applications of TPA. Bars show the mean � S.E.M. for 4–6 animals.
Significantly different from control (C), *P < 0.05, ***P < 0.001.
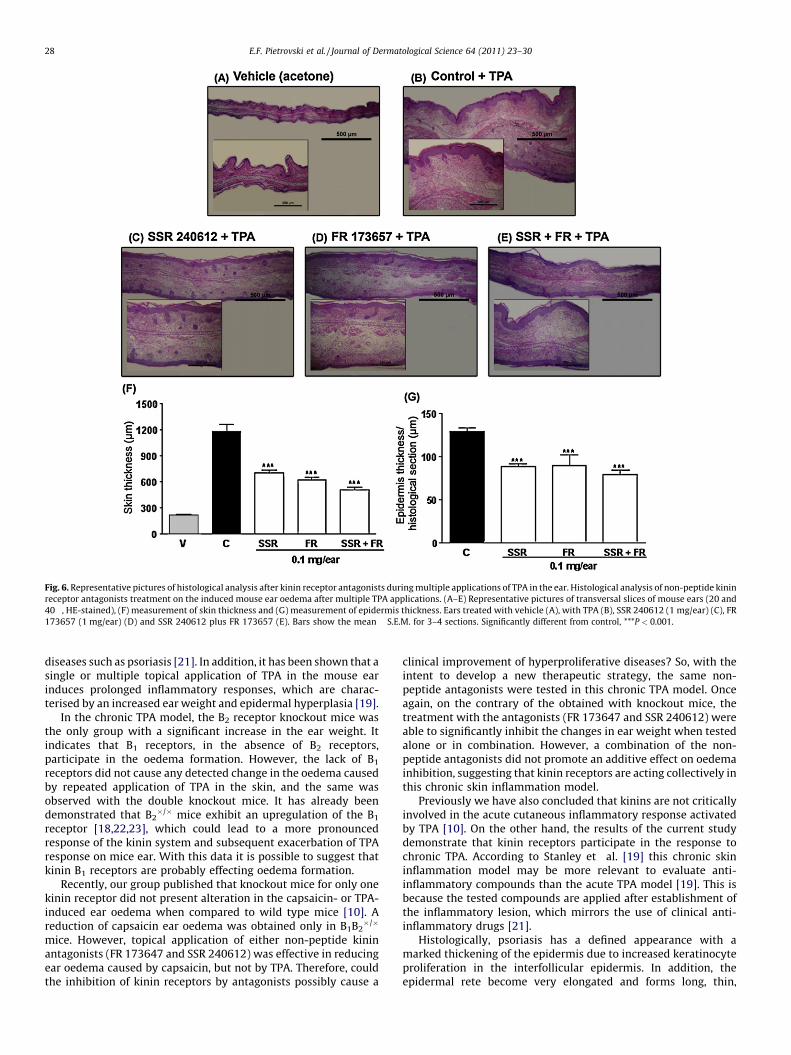
173657 and SSR 240612 plus FR 173657). Chronic application ofTPA resulted in a marked increase in ear thickness with clearevidence of oedema and epidermal hyperplasia when compared tovehicle (Fig. 6A, B and F). Treatments with either non-peptideantagonist were able to reduce ear thickness, epidermal hyperpla-sia and acanthosis (Fig. 6C–E). Mice treated with SSR 240612, FR173657, or SSR 240612 plus FR 173657 presented a significantreduction of total skin thickness measure with inhibition values of49 � 3%, 57 � 2% and 69 � 2%, respectively (Fig. 6F). Similarly,treatments with the antagonists were also able to inhibit theepidermal thickness presenting inhibition values of 32 � 2%, 30 � 9%and 38 � 4%, respectively for SSR 240612, FR 173657 and SSR 240612plus FR 173657 (Fig. 6G).
Besides, topical treatments with non-peptide antagonists werealso able to restrain cellular proliferation detected by PCNA-immunostaining (Fig. 7). When compared to control group, SSR240612, FR 173657 and SSR 240612 plus FR 173657 applicationsinhibited the increase in TPA-induced PCNA staining in 38 � 3%,26 � 7% and 31 � 3%, respectively (Fig. 7G). Repeated applications ofvehicle (acetone) did not interfere with basal PCNA parameters(Fig. 7A, B and G).
4. Discussion
This study provides evidence that kinin receptors clearlyparticipate in chronic skin inflammatory and hyperproliferativeprocesses in the epidermis after repeated TPA treatment. Phorbolesters, such as TPA, the phlogistic agent used with multipleapplications in this study, induce skin inflammation and hyper-proliferative responses in animals that resemble the signs of skin
Fig. 6. Representative pictures of histological analysis after kinin receptor antagonists during multiple applications of TPA in the ear. Histological analysis of non-peptide kinin
receptor antagonists treatment on the induced mouse ear oedema after multiple TPA applications. (A–E) Representative pictures of transversal slices of mouse ears (20 and
40�, HE-stained), (F) measurement of skin thickness and (G) measurement of epidermis thickness. Ears treated with vehicle (A), with TPA (B), SSR 240612 (1 mg/ear) (C), FR
173657 (1 mg/ear) (D) and SSR 240612 plus FR 173657 (E). Bars show the mean � S.E.M. for 3–4 sections. Significantly different from control, ***P < 0.001.
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–3028
diseases such as psoriasis [21]. In addition, it has been shown that asingle or multiple topical application of TPA in the mouse earinduces prolonged inflammatory responses, which are charac-terised by an increased ear weight and epidermal hyperplasia [19].
In the chronic TPA model, the B2 receptor knockout mice wasthe only group with a significant increase in the ear weight. Itindicates that B1 receptors, in the absence of B2 receptors,participate in the oedema formation. However, the lack of B1
receptors did not cause any detected change in the oedema causedby repeated application of TPA in the skin, and the same wasobserved with the double knockout mice. It has already beendemonstrated that B2
�/� mice exhibit an upregulation of the B1
receptor [18,22,23], which could lead to a more pronouncedresponse of the kinin system and subsequent exacerbation of TPAresponse on mice ear. With this data it is possible to suggest thatkinin B1 receptors are probably effecting oedema formation.
Recently, our group published that knockout mice for only onekinin receptor did not present alteration in the capsaicin- or TPA-induced ear oedema when compared to wild type mice [10]. Areduction of capsaicin ear oedema was obtained only in B1B2
�/�
mice. However, topical application of either non-peptide kininantagonists (FR 173647 and SSR 240612) was effective in reducingear oedema caused by capsaicin, but not by TPA. Therefore, couldthe inhibition of kinin receptors by antagonists possibly cause a
clinical improvement of hyperproliferative diseases? So, with theintent to develop a new therapeutic strategy, the same non-peptide antagonists were tested in this chronic TPA model. Onceagain, on the contrary of the obtained with knockout mice, thetreatment with the antagonists (FR 173647 and SSR 240612) wereable to significantly inhibit the changes in ear weight when testedalone or in combination. However, a combination of the non-peptide antagonists did not promote an additive effect on oedemainhibition, suggesting that kinin receptors are acting collectively inthis chronic skin inflammation model.
Previously we have also concluded that kinins are not criticallyinvolved in the acute cutaneous inflammatory response activatedby TPA [10]. On the other hand, the results of the current studydemonstrate that kinin receptors participate in the response tochronic TPA. According to Stanley et al. [19] this chronic skininflammation model may be more relevant to evaluate anti-inflammatory compounds than the acute TPA model [19]. This isbecause the tested compounds are applied after establishment ofthe inflammatory lesion, which mirrors the use of clinical anti-inflammatory drugs [21].
Histologically, psoriasis has a defined appearance with amarked thickening of the epidermis due to increased keratinocyteproliferation in the interfollicular epidermis. In addition, theepidermal rete become very elongated and forms long, thin,
Fig. 7. The effect of the treatment with kinin receptor antagonists on the epidermis cell proliferation after TPA multiple applications in the ear skin. Representative pictures of
PCNA immunohistochemical of transverse slices from Swiss mice ears stained with hematoxylin (increase of 400�, scale of 250 mm) collected on day 9 after the multiple
applications of TPA (2.0 mg/ear). Groups: naive (A), vehicle (B), control (C), SSR 240612 (D), FR 173657 (E) and SSR 240612 plus FR 173657 (F). Treatments were started on the
5th day of the experiment. Bars show the mean � S.E.M. for 16–31 sections marked with anti-PCNA antibody and stained with hematoxylin as described previously. Significantly
different from control, ***P < 0.001.
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–30 29
downward projections into the dermis [4]. Our results showed thatepidermal hyperplasia and acanthosis were pronounced in wildtype, B1
�/� and B2�/� mice, which indicated marked keratinocyte
proliferation in these animals. Curiously, the epidermal hyperpla-sia was reduced in B1B2
�/�mice, suggesting that a deletion of bothkinin receptors causes reduction in keratinocyte hyperprolifera-tion, clearly confirmed in representative ear pictures of B1B2
�/�
mice. Similarly, the treatment with non-peptide antagonistscaused a decline of epidermal thickness. Therefore, non-peptideantagonists could be topically used to reduce the histologicalalterations of the skin present in psoriasis.
In order to confirm kinin participation in the regulation ofcellular proliferation, PCNA protein expression levels wereevaluated. PCNA is a nuclear protein that is necessary for DNAsynthesis in eukaryotes. Its synthesis and expression are related to
the proliferating state of cells and its level of expression reflects theproliferating activity of cells in tumours and precanceroses [24].Using the Western blot technique, our results showed that PCNAexpression was increased in B2
�/� mice, while a significantreduction was exhibited by B1B2
�/� mice. This result confirmsthat only the absence of both kinin receptors is necessary to reducehyperproliferative responses in the keratinocytes of the epidermis.The confirmation of this result was obtained with the quantifica-tion of PCNA by immunohistochemistry, which showed areduction of PCNA staining in double knockout mice. In addition,the PCNA staining was more specific in the basal layer of theepidermis. Schremmer-Danninger et al. demonstrated a specificimmunolabelling for kinin B2 receptors in basal keratinocytes inthe epidermis of normal skin [9]. The fact that these sites arespecifically confined in skin tissue and is associated with a high
E.F. Pietrovski et al. / Journal of Dermatological Science 64 (2011) 23–3030
mitotic activity suggests that BK may play an important role inepidermal cell proliferation [25]. However, the PCNA staining wasmore intense in B2
�/� knockout mice, and again, it appeared thatthe presence of one type of kinin receptor was enough to maintainkinin system functioning. On the other hand, when the lack ofactivity of kinin receptors was caused by a selective antagonist(SSR 240612 and FR 173657), and not by knocking out the gene, thereduction in PCNA staining happened with either antagonist. Thus,with the establishment of the hyperproliferative process, thetreatment with a B1 or with a B2 antagonist caused a reduction inthe cell proliferation induced by TPA, effect that was not intensifiedby the association of both antagonists. Therefore, this datademonstrate that non-peptide kinin antagonists topically appliedin an animal model of skin inflammation are effective interferingwith cell hyperproliferation, and in this situation the inactivity ofonly one kinin receptor is sufficient to reach the purpose.
This study indicated that B1 and B2 kinin receptors could becontributing to the induction of cell proliferation in skin diseases.Accordingly, previous studies have reported kinin involvement inthe control of cell proliferation. Schremmer-Danninger et al.described that BK induces phosphoinositol turnover in keratino-cytes, as well as the formation of 1,2-diglyceride, tyrosinephosphorylation and cell growth [25]. Poblete et al. reportedthat in psoriatic human skin, there are many neutrophilsimmunoreactive for tissue kallikrein in the epidermis and psoriaticscales [11]. Several studies have reported that kinins may increaseDNA synthesis and cell division in several cellular systems [26].Nevertheless, some data are contradictory and state that kinins donot stimulate keratinocyte proliferation [27–29] or induce a weakresponse when compared with that produced by epidermal growthfactor (EGF) [30]. These previously mentioned works wereperformed in vitro and some used epidermal scales of psoriaticpatients. Thus, our work complements these previous studies andemploys an in vivo model to clarify kinin actions in the skin.
The obtained results showed that kinins most likely contributeto the pathogenesis of psoriasis by regulating cell hyperprolifera-tion. Furthermore, we have demonstrated that the non-peptideantagonists FR 173657 and SSR 240612 were potent inhibitors in
vivo when topically applied, thereby highlighting a possible newalternative for the local treatment of hyperproliferative skindisorders.
Acknowledgments
This study was supported by grant from Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico (CNPq). E.F. Pietrovski isa PhD student of pharmacology and thanks CAPES for fellowshipsupport. The authors thank Sanofi-Aventis (Germany) and AstellasPharma Inc. (Japan) for the generous supply of SSR 240612 and FR173657, respectively.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, in
the online version, at doi:10.1016/j.jdermsci.2011.06.016.
References
[1] Fang M, Wee SA, Ronski K, Fan H, Tao S, Lin Q. Evidence of EGR1 as adifferentially expressed gene among proliferative skin diseases. GenomicMed 2007;1:75–85.
[2] Ayli EE, Li W, Brown TT, Witkiewicz A, Elenitsas R, Seykora JT. Activation of Src-family tyrosine kinases in hyperproliferative epidermal disorders. J CutanPathol 2008;35:273–7.
[3] Jean J, Lapointe M, Soucy J, Pouliot R. Development of an in vitro psoriatic skinmodel by tissue engineering. J Dermatol Sci 2009;53:19–25.
[4] Lowes MA, Bowcock AM, Krueger JG. Pathogenesis and therapy of psoriasis.Nature 2007;445:866–73.
[5] Schon MP, Boehncke WH. Psoriasis. N Engl J Med 2005;352:1899–912.[6] Nickoloff BJ, Bonish BK, Marble DJ, Schriedel KA, Dipietro LA, Gordon KB, et al.
Lessons learned from psoriatic plaques concerning mechanisms of tissuerepair, remodeling, and inflammation. J Investig Dermatol Symp Proc2006;11:16–29.
[7] Gottlieb AB. Psoriasis: emerging therapeutic strategies. Nat Rev Drug Discov2005;4:19–34.
[8] Schremmer-Danninger E, Hermann A, Fink E, Fritz H, Roscher AA. Identifica-tion and occurrence of mRNAs for components of the kallikrein–kinin systemin human skin and in skin diseases. Immunopharmacology 1999;43:287–91.
[9] Schremmer-Danninger E, Naidoo S, Neuhof C, Valeske K, Snyman C, Sander C,et al. Visualisation of tissue kallikrein, kininogen and kinin receptors in humanskin following trauma and in dermal diseases. Biol Chem 2004;385:1069–76.
[10] Pietrovski EF, Otuki MF, Regoli D, Bader M, Pesquero JB, Cabrini DA, et al. Thenon-peptide kinin receptor antagonists FR 173657 and SSR 240612: preclini-cal evidence for the treatment of skin inflammation. Regul Pept 2009;152:67–72.
[11] Poblete MT, Reynolds NJ, Figueroa CD, Burton JL, Muller-Esterl W, Bhoola KD.Tissue kallikrein and kininogen in human sweat glands and psoriatic skin. Br JDermatol 1991;124:236–41.
[12] Campos MM, Leal PC, Yunes RA, Calixto JB. Non-peptide antagonists for kininB1 receptors: new insights into their therapeutic potential for the manage-ment of inflammation and pain. Trends Pharmacol Sci 2006;27:646–51.
[13] Leeb-Lundberg LM, Marceau F, Muller-Esterl W, Pettibone DJ, Zuraw BL.International union of pharmacology. XLV. Classification of the kinin receptorfamily: from molecular mechanisms to pathophysiological consequences.Pharmacol Rev 2005;57:7–77.
[14] Calixto JB, Medeiros R, Fernandes ES, Ferreira J, Cabrini DA, Campos MM. KininB1 receptors: key G-protein-coupled receptors and their role in inflammatoryand painful processes. Br J Pharmacol 2004;143:803–18.
[15] Chai KX, Ni A, Wang D, Ward DC, Chao J, Chao L. Genomic DNA sequence,expression, and chromosomal localization of the human B1 bradykinin recep-tor gene BDKRB1. Genomics 1996;31:51–7.
[16] Pesquero JB, Araujo RC, Heppenstall PA, Stucky CL, Silva Jr JA, Walther T, et al.Hypoalgesia and altered inflammatory responses in mice lacking kinin B1receptors. Proc Natl Acad Sci U S A 2000;97:8140–5.
[17] Rupniak NM, Boyce S, Webb JK, Williams AR, Carlson EJ, Hill RG, et al. Effects ofthe bradykinin B1 receptor antagonist des-Arg9[Leu8]bradykinin and geneticdisruption of the B2 receptor on nociception in rats and mice. Pain1997;71:89–97.
[18] Cayla C, Todiras M, Iliescu R, Saul VV, Gross V, Pilz B, et al. Mice deficient forboth kinin receptors are normotensive and protected from endotoxin-inducedhypotension. FASEB J 2007;21:1689–98.
[19] Stanley PL, Steiner S, Havens M, Tramposch KM. Mouse skin inflammationinduced by multiple topical applications of 12-O-tetradecanoylphorbol-13-acetate. Skin Pharmacol 1991;4:262–71.
[20] Medeiros R, Otuki MF, Avellar MC, Calixto JB. Mechanisms underlying theinhibitory actions of the pentacyclic triterpene alpha-amyrin in the mouseskin inflammation induced by phorbol ester 12-O-tetradecanoylphorbol-13-acetate. Eur J Pharmacol 2007;559:227–35.
[21] Gabor M. Mouse ear inflammation models. In: Gabor M, editor. Mouse earinflammation models and their pharmacological applications. Budapeste:Akademiai Kiado; 2000. p. 15–33.
[22] Madeddu P, Varoni MV, Palomba D, Emanueli C, Demontis MP, Glorioso N,et al. Cardiovascular phenotype of a mouse strain with disruption of bradyki-nin B2-receptor gene. Circulation 1997;96:3570–8.
[23] Duka I, Kintsurashvili E, Gavras I, Johns C, Bresnahan M, Gavras H. Vasoactivepotential of the B(1) bradykinin receptor in normotension and hypertension.Circ Res 2001;88:275–81.
[24] Gambichler T, Bischoff S, Bechara FG, Altmeyer P, Kreuter A. Expression ofproliferation markers and cell cycle regulators in T cell lymphoproliferativeskin disorders. J Dermatol Sci 2008;49:125–32.
[25] Schremmer-Danninger E, Heinz-Erian P, Topfer-Petersen E, Roscher AA. Au-toradiographic localization and characterization of bradykinin receptors inhuman skin. Eur J Pharmacol 1995;283:207–16.
[26] Bhoola KD, Figueroa CD, Whorthy K. Bioregulation of kinins: kallikreins,kininogens, and kininases. Pharmacol Rev 1992;44:1–80.
[27] Johnson RM, King KL, Morhenn VB. Comparison of second messengerformation in human keratinocytes following stimulation with epidermalgrowth factor and bradykinin. Second Messengers Phosphoproteins1992;14:21–37.
[28] Jung EM, Betancourt-Calle S, Mann-Blakeney R, Griner RD, Bollinger Bollag W.Sustained phospholipase D activation is associated with keratinocyte differ-entiation. Carcinogenesis 1999;20:569–76.
[29] Vidal MA, Astroza A, Matus CE, Ehrenfeld P, Pavicic F, Sanchez T, et al. Kinin B2receptor-coupled signal transduction in human cultured keratinocytes. JInvest Dermatol 2005;124:178–86.
[30] Coutant KD, Ryder NS. Bradykinin upregulates immediate-early gene mRNA inhuman keratinocytes. Arch Dermatol Res 1996;288:2–6.