Bacteriocin PJ4 Active Against Enteric Pathogen Produced by Lactobacillus helveticus PJ4 Isolated from Gut Microflora of Wistar Rat (Rattus norvegicus): Partial Purification and Characterization of Bacteriocin Prasant Kumar Jena & Disha Trivedi & Harshita Chaudhary & Tapasa Kumar Sahoo & Sriram Seshadri Received: 8 October 2012 / Accepted: 17 December 2012 # Springer Science+Business Media New York 2013 Abstract The increase of multidrug-resistant pathogens and the restriction on the use antibiotics due to its side effects have drawn attention to the search for possible alternatives. Bacteriocins are small antimicrobial peptides produced by numerous bac- teria. Much interest has been focused on bacteriocins because they exhibit inhibitory activity against pathogens. Lactic acid bacteria possess the ability to synthesize anti- microbial compounds (like bacteriocin) during their growth. In this study, an antibac- terial substance (bacteriocin PJ4) produced by Lactobacillus helveticus PJ4, isolated from rat gut microflora, was identified as bacteriocin. It was effective against wide assay of both Gram-positive and Gram-negative bacteria involved in various diseases, including Escherichia coli, Bacillus subtilis, Pseudomonas aeruginosa, Enterococcus faecalis, and Staphylococcus aureus. The antimicrobial peptide was relatively heat- resistant and also active over a wide pH range of 2–10. It has been partially purified to homogeneity using ammonium sulfate precipitation and size exclusion chromatogra- phy and checked on reverse-phase high-performance liquid chromatography. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis of bacteriocin PJ4 purified through size exclusion chromatography resolved ~6.5 kDa protein with bacteriocin activity. The peptide is inactivated by proteolytic enzymes, trypsin, and lipase but not when treated with catalase, α-amylase, and pepsin. It showed a bactericidal mode of action against the indicator strains E. coli MTCC443, Lactobacillus casei MTCC1423, and E. faecalis DT48. Such characteristics indicate that this bacteriocin may be a potential candidate for alternative agents to control important pathogens. Appl Biochem Biotechnol DOI 10.1007/s12010-012-0044-7 P. K. Jena : D. Trivedi : H. Chaudhary : S. Seshadri (*) Institute of Science, Nirma University, Sarkhej-Gandhinagar Highway, Chharodi, Ahmedabad 382481 Gujarat, India e-mail: [email protected]T. K. Sahoo School of Life Sciences, Sambalpur University, Jyotivihar, Burla, 768019 Sambalpur, Odisha, India
Transcript
Bacteriocin PJ4 Active Against Enteric PathogenProduced by Lactobacillus helveticus PJ4 Isolatedfrom Gut Microflora of Wistar Rat (Rattus norvegicus):Partial Purification and Characterization of Bacteriocin
Received: 8 October 2012 /Accepted: 17 December 2012# Springer Science+Business Media New York 2013
Abstract The increase of multidrug-resistant pathogens and the restriction on the useantibiotics due to its side effects have drawn attention to the search for possiblealternatives. Bacteriocins are small antimicrobial peptides produced by numerous bac-teria. Much interest has been focused on bacteriocins because they exhibit inhibitoryactivity against pathogens. Lactic acid bacteria possess the ability to synthesize anti-microbial compounds (like bacteriocin) during their growth. In this study, an antibac-terial substance (bacteriocin PJ4) produced by Lactobacillus helveticus PJ4, isolatedfrom rat gut microflora, was identified as bacteriocin. It was effective against wideassay of both Gram-positive and Gram-negative bacteria involved in various diseases,including Escherichia coli, Bacillus subtilis, Pseudomonas aeruginosa, Enterococcusfaecalis, and Staphylococcus aureus. The antimicrobial peptide was relatively heat-resistant and also active over a wide pH range of 2–10. It has been partially purifiedto homogeneity using ammonium sulfate precipitation and size exclusion chromatogra-phy and checked on reverse-phase high-performance liquid chromatography. Sodiumdodecyl sulfate–polyacrylamide gel electrophoresis of bacteriocin PJ4 purified throughsize exclusion chromatography resolved ~6.5 kDa protein with bacteriocin activity. Thepeptide is inactivated by proteolytic enzymes, trypsin, and lipase but not when treatedwith catalase, α-amylase, and pepsin. It showed a bactericidal mode of action againstthe indicator strains E. coli MTCC443, Lactobacillus casei MTCC1423, and E. faecalisDT48. Such characteristics indicate that this bacteriocin may be a potential candidatefor alternative agents to control important pathogens.
P. K. Jena :D. Trivedi : H. Chaudhary : S. Seshadri (*)Institute of Science, Nirma University, Sarkhej-Gandhinagar Highway, Chharodi,Ahmedabad 382481 Gujarat, Indiae-mail: [email protected]
T. K. SahooSchool of Life Sciences, Sambalpur University, Jyotivihar, Burla, 768019 Sambalpur, Odisha, India
Keywords Lactobacillus helveticus . Bacteriocin . Purification . Characterization .
Mode of action . Pathogen
Introduction
Infections caused by enteric pathogens are an important cause of morbidity and mortalityworldwide and have a major impact on public health [1]. Enteric bacteria comprised ofSalmonella species, Shigella species, Proteus species, Klebsiella species, Escherichia coli,Pseudomonas species, Vibrio cholerae, and Staphylococcus aureus which are major etio-logic agents of enteric infection [2]. Enteric pathogens are often transmitted by means offood or water (foodborne diseases) and are responsible for acute gastroenteritis; some causesystemic disease that may have chronic complications. The rise in antibiotic-resistantbacteria has generated interest in the scientific community to the prophylactic and therapeu-tic uses of probiotics and to reconsider them as alternatives to antibiotics [3, 4].
Microbial therapeutics is expanding and commensal beneficial bacteria are being imple-mented as treatment and prevention strategies for immune disorders and infectious diseases[5]. A group of antimicrobials peptides, called bacteriocins, have been studied because theyhold a great potential in controlling antibiotic-resistant pathogens. They often act towardspecies related to the producer with a very high potency and specificity. The commonmechanisms of killing by bacteriocins are destruction of target cells by pore formationand/or inhibition of cell wall synthesis [6]. The bacteriocins are active against numerousfoodborne and human pathogens, are produced by “generally regarded as safe” micro-organisms—Lactobacilli, and are readily degraded by proteolytic host systems, which makethem attractive candidates for biotechnological applications [5].
The intestinal microbiota has been identified as a rich source of potential probioticbacteria which produce novel antimicrobial and, more specifically, antipathogenic bacter-iocins having exceptional potential with respect to beneficially modulating the gastrointes-tinal microbiota and specifically inhibiting specific gastrointestinal pathogens [7]. Differentbacteria produce different types of bacteriocins that potentially reach high concentrations incertain local regions of the gut; these compounds act in a nontargeted manner and theircontribution to probiotic functionality has not been investigated as extensively. Lactobacil-lus spp. produce antimicrobial factors and bacteriocins, including lantibiotics, small heat-stable, nonlanthionine containing membrane-active peptides, larger heat-labile proteins, andcomplex bacteriocins containing one or more chemical moieties [8, 9]. Probiotics areproducing diverse antimicrobial agents and may be beneficial for the treatment and preven-tion of a variety of infectious diseases caused by oral, enteric pathogens and urogenitalinfections [3, 9]. These strains may show therapeutic alternatives in the multidrug-resistantpathogens [10, 11].
Some bacteriocins are active against certain Gram-negative bacteria, such as E. coli andSalmonella Typhimurium [12]. Bacteriocins are ribosomally synthesized and extracellularlyreleased bioactive peptides or proteins displaying antimicrobial activity against other bacte-ria. They are generally low-molecular-weight proteins that enter into the target cells bybinding to cell surface receptors [13].
Therefore, there is an enormous need to explore and isolate more and more bacteria fromnew sources capable of producing novel bacteriocins and to characterize them for furtherapplications. Very few bacteriocins has been identified and characterized from the Lactoba-cillus helveticus species. The bacteriocins lactocin LP27 and helveticin J were produced byL. helveticus LP27 and L. helveticus 481, respectively [14,15].
Appl Biochem Biotechnol
In the present study, we report the purification and characterization of a bacteriocinproduced by L. helveticus PJ4, showing bactericidal activity against the indicator organism,enteric E. coli and other selected bacteria.
To the best of our knowledge, this is the first report describing the characterizationand partial purification of the bacteriocin component of L. helveticus PJ4 isolatedfrom the gut microflora of male Wistar rat (Rattus norvegicus), an experimentalanimal.
Materials and Methods
Bacterial Strains, Growth Conditions, and Media
L. helveticus PJ4 (NCBI Accession No. JQ068823) isolated from feces of male Wistar ratswas routinely propagated in de Man, Rogosa, and Sharpe (MRS) medium (HiMedia,Mumbai, India). For bacteriocin production, the strain PJ4 was grown in MRS medium.The indicator organism used in bacteriocin assay, E. coli MTCC433, was propagated inbrain–heart infusion (BHI) broth. In addition, several Gram-positive and Gram-negativestrains (Table 1), from various sources, were used in the determination of spectrum ofactivity. All cultures were raised at 30–37 °C in MRS or BHI broth as shown in Table 1.All chemicals were obtained from Sigma-Aldrich (USA) and all media components werepurchased from HiMedia (India). Antimicrobial activity was determined by the agar spot testmethod. Activity was expressed as arbitrary units (AU) per milliliter, with 1 AU defined asthe reciprocal of the highest dilution showing a clear zone of inhibition [12].
In Vivo Characterization of L. helveticus PJ4 Strain for Probiotics Characteristics
Eight- to 10-week-old male Wistar rats weighing 200–250 g were procured from theLaboratory Animals Centre of Zydus Research Center (ZRC), Ahmedabad. Approval to
Table 1 Antagonism of bacteriocin PJ4 produced by L. helveticus PJ4 against various target strains
Target strains Collection Growth medium Growth temperature (°C) Inhibition (mm)
Escherichia coli MTCC443 BHI 37 27±0.45
Lactobacillus plantarum fecal isolates MRS 37 27±0.19
Values are presented as the mean ± standard error of the mean (SEM) (n=3)
BHI brain–heart infusion agar, MRS de Man, Rogosa, and Sharpe agar (HiMedia)
Appl Biochem Biotechnol
work with Wistar rats was obtained from the Ethics Committee of the Nirma Univer-sity (ethics reference number: IS/BT/PHD10-11/001) from the Ministry of Environ-ment and Forests, Government of India and Committee for the Purpose of Control andSupervision of Experiments on Animals. The animals were acclimatized at a temper-ature of 25±2 °C and relative humidity of 50–60 % under 12/12 h light/darkconditions for 1 week before experiments. These rats were divided into two groups(six rats each). One group was fed orally with 1 ml of 0.8 % saline containingapproximately 107–108 cells of L. helveticus PJ4 daily for a month. Another group fedwith 1 ml of saline alone served as a control. Weights of animals were checkedweekly. Fecal samples of treated and untreated rats were enumerated for microbiolog-ical population. One gram of feces sample was diluted in 0.8 % saline. Diluents(0.1 ml) were spread plated on MRS agar, yeast and mold agar, and violet red bileagar. Plates were incubated for 48 h at 37 °C, and resultant colonies were counted.
Mode of Bacteriocin Action
Growth of the Test Microorganisms in Presence of Bacteriocin
BHI broth was inoculated with 1 % (v/v) E. coliMTCC443; MRS broth was inoculated with1 % (v/v) Enterococcus faecalis DT48 and Lactobacillus casei MTCC1423 at the earlyexponential phase and then incubated at 37 °C. The 10-ml filter-sterilized cell-free super-natant was added to the cultures (90 ml), and changes in optical density (at 600 nm) wererecorded every hour for 16 h (Fig. 2). Control cells were treated with the inactive bacteriocin(treated for 20 min at 121 °C).
Determination of the Reduction of Viable Cells of Target Microorganisms in the Presenceof Bacteriocin
Early stationary phase cultures of E. coli MTCC443 (16 h old), E. faecalis DT48 (16 h old),and L. caseiMTCC1423 (18 h old) were harvested (10,000 rpm, 5 min, 4 °C), washed twicewith sterile saline, and resuspended in 10 ml saline. Equal volumes of the cell suspensionsand filter-sterilized (0.20-μm; Axiva) bacteriocin PJ4 were mixed. Viable cell numbers weredetermined before and after incubation for 1 h at 37 °C by plating onto suitable agarmedium. Cell suspensions of E. coli MTCC443, E. faecalis DT48, and L. casei MTCC1423and with no added bacteriocin served as controls.
Molecular Mass Determination of Bacteriocin PJ4
A 24-h-old culture of L. helveticus PJ4 obtained in MRS broth at 37 °C was centri-fuged (15 min, 10,000 rpm) and the pH adjusted to 6.0 with 1 N NaOH. To avoidproteolytic degradation of the bacteriocin, cell-free supernatants were treated for 10 minat 80 °C. Ammonium sulfate was slowly added to the cell-free supernatants to 60 %saturation and stirred for 4 h at 4 °C and centrifuged (10,000 rpm, 30 min, 4 °C). Theprecipitate was resuspended in 10 ml of 25 mM ammonium acetate buffer (pH 6.5) anddesalted by dialysis using a 1,000-Da cutoff dialysis membrane (Sigma) against thesame buffer. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE)was used for further separation, as described by Schägger and Von Jagow [16]. Low-molecular-weight markers with sizes ranging from 2.5 to 45.0 kDa (Amersham Bio-sciences, Germany) were used.
Appl Biochem Biotechnol
Bacteriocin Purification
Bacteriocin was purified from a 1,000-ml culture of L. helveticus PJ4, grown in MRS brothas described above. Cells were removed by centrifugation at 14,000 rpm for 10 min at roomtemperature. Purification of bacteriocin was achieved by using a multistep protocol.
Ammonium Sulfate Precipitation
Culture supernatant was brought to 90 % saturation with solid ammonium sulfate, and afterstirring overnight at 4 °C, the precipitate was collected by centrifugation (10,000 rpm,10 min, 4 °C). The precipitate was dissolved in 60 ml sodium phosphate buffer (20 mM,pH 6.0), and the bacteriocin suspension was desalted by dialyzing through a 2-kDa cutoffdialysis membrane (Sigma) against the same buffer for 24 h. The dialyzed suspension wascentrifuged at 14,000 rpm for 15 min at 4 °C. The supernatant was filtered through a 0.2-μmmembrane and checked for antimicrobial activity by agar well diffusion assay by using E.coli MTCC443 as the indicator strain and the same was labeled as fraction I.
Size Exclusion Chromatography
Fraction I was then loaded on a Superdex 75 (10/300 GL) prepacked Tricon column (GEHealthcare, USA) linked to a high-performance liquid chromatography (HPLC; Agilent, 1100)system equilibrated with sodium phosphate buffer (pH 6.7) with 200 mM NaCl at a flow rate of0.5 ml/min [17]. The eluted peaks were fractionated at a volume 0.5 ml and were checked forantimicrobial activity described earlier. The active fractions were pooled and named as fraction II.
To check the homogeneity of purified active fraction II eluted from the size exclusion chromatog-raphy loaded in ACE-5, C-18-300 reverse-phase (RP) column (250×4.6 mm; ACE CapillaryColumn, Advance Chromatography Technology, Scotland) using anHPLC system (Agilent, USA).The column was equilibrated with solvent A (HPLC-grade water containing 0.1 % trifluoroaceticacid [TFA]). The elution was performed using linear gradient from solvent A to 100 % acetonitrilein 0.1 % TFA (solvent B) for 60 min. The flow rate (0.3 ml/min) and temperature (60 °C) wasmaintained and eluted analytes were monitored by an ultraviolet detector at 210 nm.
Effect of pH, Temperature, and Enzymes on the Activity of Bacteriocin
L. helveticus PJ4 was grown in MRS medium for 20 h at 37 °C. The cells were harvested(10,000 rpm, 15 min, 4 °C) and the cell-free supernatant was incubated for 2 h in thepresence of trypsin (Sigma), proteinase K (Sigma), pepsin (Sigma), lipase (Sigma), and α-amylase (Sigma) at a final concentration of 1.0 mg/ml. Enzyme activity was terminated byboiling for 5 min, and the residual activity was determined by using the well diffusionmethod. An untreated sample was used as a control (100 %).
The effect of pH on the activity of bacteriocin PJ4 was tested by adjusting cell-freesupernatants from pH 2.0 to 10.0 (at increments of two pH units) with sterile 1 M NaOH or1 M HCl for 1 h at 37 °C. After incubation, pH was neutralized to pH 6.0. Antimicrobialactivity was tested by well diffusion method.
To determine the effects of temperature, purified antimicrobial samples were incubated inde-pendently at 30, 45, 60, 75, and 100 ° C for 1 h. A nonheated sample was used as a control (100%).
Appl Biochem Biotechnol
Results and Discussion
The aims of this study was to screen the antimicrobial protein producing L. helveticus PJ4from fecal microflora of male Wistar rats and to screen their bacteriocins as potential naturalantibacterial agents for use against few selected pathogens.
Spectrum of Activity
Bacteriocin PJ4 presented a broad spectrum of activity, being inhibitory against many entericbacteria and foodborne pathogens. The antibacterial activity of bacteriocin PJ4 was not onlyevident against Gram-positive bacteria but also against Gram-negative bacteria (Table 1). Similarresults were recorded for the cell-free supernatant and for the semipurified bacteriocin (Fig. 3).The bacteriocin PJ4 showed high activity againstE. coli, S. aureus,E. faecalis, andEnterococcusfaecium but showed low activity against Pseudomonas aeruginosa, Shigella flexneri, Klebsiellapneumoniae, and Salmonella Typhimurium. Most of the bacteriocins described for L. helveticusare active against a much broader range of genera and species [15]. It is important to outline thatthe bioactivity against E. coli, Salmonella, and Shigella enteric pathogens is of increasingimportance. Activity against Gram-negative bacteria is also a relevant characteristic, detectedby several authors in other Lactobacilli. Bacteriocin R1333 produced by Lactobacillus sakeiR1333 isolated from smoked salmon and enterocin LR/6 produced by E. faeciumLR/6were alsoactive against Gram-negative bacteria [18, 19]. The activity of bacteriocin PJ4 observed againstGram-negative bacteria (Escherichia, Pseudomonas, and Salmonella) is an unpredicted result.Earlier studies [20] reported that bacteriocins of lactic acid bacteria are inefficient to inhibitGram-negative bacteria because the outer membrane hinders the site for bacteriocin action.However, a few bacteriocins produced by Lactobacillus plantarum have been reported to beactive against Gram-negative bacteria. BacteriocinsST26MS (2.8 kDa) and ST28MS (5.5 kDa)produced by L. plantarum ST26MS and ST28MS, respectively, can inhibit Acinetobacter,Escherichia, and Pseudomonas, and antimicrobial activity against Gram-negative bacteria isalso a relevant characteristic that was previously detected by several authors in other micro-organisms including bacteriocins produced by other Lactobacilli [21].
In Vivo Probiotic Characterization
When male Wistar rats were fed with L. helveticus PJ4 (approximately 108 cells/ml for amonth), there was no sign of any illness and they were similar as per control rats in terms ofweight gain per week. There was a marked decrease in the count of yeast and mold (3.297±0.23 to 2.889±0.22) and coliform bacteria (6.365±0.15 to 2.795±0.18) in L. helveticus PJ4-treated samples compared to untreated rats. A significantly increased count of Lactobacilliwas found in treated rat as compared to untreated rats (Table 2). These results indicate the
Table 2 Comparison of microbial flora in treated and untreated group with L. helveticus PJ4
Strain Culture media L. helveticus PJ4treated (Log CFU/g)
L. helveticus PJ4untreated (Log CFU/g)
Lactobacilli MRS agar 10.084±0.24 8.352±0.27
Yeast and mold Yeast and mold agar 2.889±0.22 3.297±0.23
Coliforms Violet red bile agar 2.795±0.18 6.365±0.15
Values are presented as the mean ± SEM (n=3)
Appl Biochem Biotechnol
antimicrobial nature of L. helveticus PJ4 inside the gut of the rat. Hence, it could beadministered orally as food supplement.
Mode of Action
E. coli MTCC443 treated with bacteriocin PJ4 (200 AU/ml) increased from OD600 0.145 to0.459 over 7 h (Fig. 3). The control E. coli MTCC443 (not treated with bacteriocin PJ4)increased from OD600 0.145 to 1.096 over the same period (Fig. 1a). E. faecalis DT48treated with bacteriocin PJ4 (350 AU/ml) increased from OD600 0.138 to 0.351 over 7 h(Fig. 1b). The control E. faecalis DT48 (not treated with bacteriocin PJ4) increased fromOD600 0.137 to 0.725 over the same period. L. casei MTCC1423 treated with bacteriocinPJ4 (600 AU/ml) increased from OD600 0.068 to 0.227over 7 h (Fig. 3). The control L. casei
Fig. 1 The effect of bacteriocin PJ4 on the growth of a E. coli MTCC443, b E. faecalis DT48, and c L. caseiMTCC1423. Growth of test microorganisms in presence of bacteriocin PJ4 (triangles) and in the absence ofbacteriocin PJ4 (squares). The arrow indicates the time point at which the bacteriocin was added
Appl Biochem Biotechnol
MTCC1423 (not treated with bacteriocin PJ4) increased from OD600 0.067 to 0.356 over thesame period (Fig. 1c). When stationary-phase cells of E. faecalis DT48, L. caseiMTCC1423, and E. coli MTCC443 (107–108 CFU/ml) were treated with the bacteriocinproduced by L. helveticus PJ4, complete death of L. casei MTCC1423 and E. coli resultedafter 1 h contact time. No viable cells of L. casei MTCC1423 and E. coli MTCC443 weredetected, while low levels (102–103 CFU/ml) of viable cells of E. faecalis DT48 weredetected. No significant changes in cell numbers of E. faecalis DT48, L. casei MTCC1423,and E. coli were recorded in the untreated (control) sample. Previously, similar results wereobtained by bacteriocins HA-6111-2 and HA-5692-3 produced by Pediococcus acidilactici[22], pediocin-like bacteriocin ST5Ha from E. faecium [23], and bacteriocin from L. acid-ophilus La-14 [24].
Molecular Mass Determination
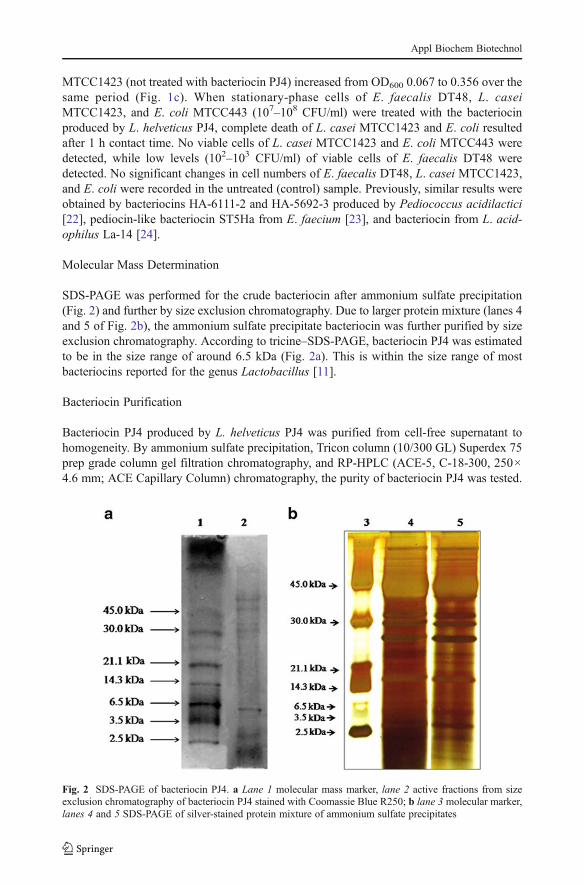
SDS-PAGE was performed for the crude bacteriocin after ammonium sulfate precipitation(Fig. 2) and further by size exclusion chromatography. Due to larger protein mixture (lanes 4and 5 of Fig. 2b), the ammonium sulfate precipitate bacteriocin was further purified by sizeexclusion chromatography. According to tricine–SDS-PAGE, bacteriocin PJ4 was estimatedto be in the size range of around 6.5 kDa (Fig. 2a). This is within the size range of mostbacteriocins reported for the genus Lactobacillus [11].
Bacteriocin Purification
Bacteriocin PJ4 produced by L. helveticus PJ4 was purified from cell-free supernatant tohomogeneity. By ammonium sulfate precipitation, Tricon column (10/300 GL) Superdex 75prep grade column gel filtration chromatography, and RP-HPLC (ACE-5, C-18-300, 250×4.6 mm; ACE Capillary Column) chromatography, the purity of bacteriocin PJ4 was tested.
Fig. 2 SDS-PAGE of bacteriocin PJ4. a Lane 1 molecular mass marker, lane 2 active fractions from sizeexclusion chromatography of bacteriocin PJ4 stained with Coomassie Blue R250; b lane 3 molecular marker,lanes 4 and 5 SDS-PAGE of silver-stained protein mixture of ammonium sulfate precipitates
Appl Biochem Biotechnol
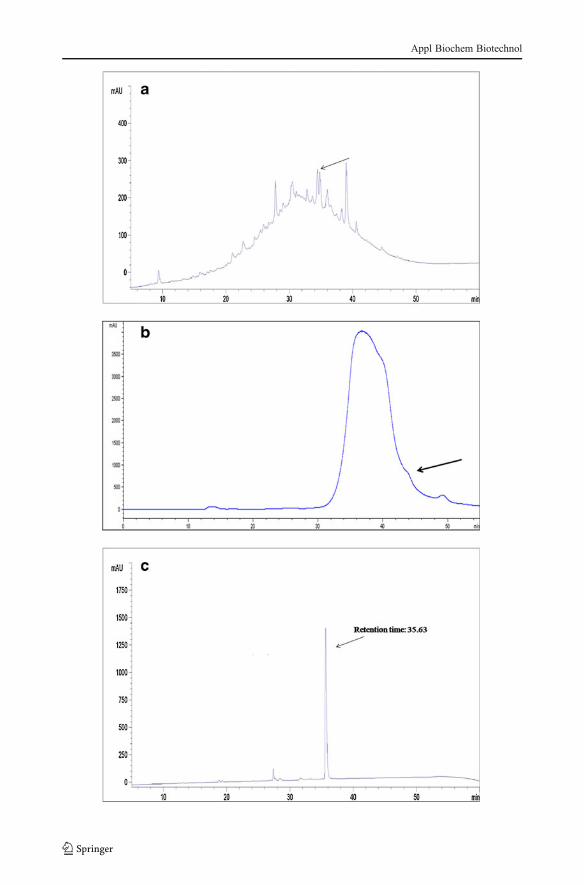
Cell-free supernatants from 24-h cultures in MRS broth at 37 °C were used for bacteriocinpurification. The activity against E. coli MTCC443 presented by the proteins precipitatedwith ammonium sulfate and reconstituted in ammonium acetate buffer was similar to thatpresented by the fractions after size exclusion chromatography on Superdex 75 column. Theantimicrobial activity of fraction I and fraction II is shown in Fig. 3. The HPLC analysis ofcrude bacteriocin after ammonium sulfate precipitation is shown in Fig. 4b. After sizeexclusion chromatography analysis (Fig. 4b), the active fraction II was reinjected in RP-HPLC. A partial purified active fraction was obtained at a retention time of 35 min (Fig. 4c).Similar purification protocol was used by other researchers for purification of bacteriocins[12, 19, 23, 25].
Effect of pH, Temperature, and Enzymes on the Activity of Bacteriocin
The antimicrobial protein purified from L. helveticus PJ4 (bacteriocin PJ4) was tested for itssensitivity to pH, temperature, and enzymes; the results are summarized in Table 3. Bacte-riocin PJ4 remained stable after incubation for 1 h at pH values from 2.0 to 6.0, but itsactivity was reduced during pH 8.0–10.0 and highest activity shows at pH 6.0. These resultsdemonstrated that the bacteriocin was resistant to acidic conditions. Bacteriocins weresimilar with regard to their sensitivity to inactivation by temperature. Like most of theknown bacteriocins, they were mainly heat-tolerant at pH 6.5 [26]. The bacteriocin PJ4showed greater thermal stability. It retained 95.84 % of its initial activity after 1 h incubationat 60 °C, whereas it retained only 69.85 % activity at 100 °C. Only bacteriocins ET30 andET31 were moderately heat-stable at 60 °C and 100 °C, thus resembling nisin produced byL. lactis WNC20, which was inactivated after 15 min at 121 °C at pH 7.0 [27], or pediocinPA-1, showing about 40 % activity lost after 15 min of heating at 121 °C in the pH range of2.5–9.0 [28].
Fig. 3 Antimicrobial activity of protein during purification steps: A activity of fraction II of size exclusionchromatography, B activity of fraction1of ammonium sulfate precipitation, C bacteriocin-like peptide withproteinase K, D bacteriocin-like peptide with catalase and proteinase K
Appl Biochem Biotechnol
Appl Biochem Biotechnol
The antibacterial activity of bacteriocin PJ4 was completely abolished after treatmentwith proteolytic enzymes (proteinase K), whereas α-amylase and pepsin had no effect.Treatment of partial purified bacteriocin with catalase did not result in any changes ofantibacterial activity (Table 3), indicating that hydrogen peroxide was not responsible forinhibition. These results indicate that bacteriocin PJ4 is a kind of peptide that does notcontain lipid or carbohydrate groups. Again, it exhibited high thermal and pH stability. Inthis case, the secreted organic acids can be ruled out since the pH of the growth medium wasalways in the neutral range (6.5–7.0). These properties are similar to that of a bacteriocinproduced by Lactobacillus sake C2 [29].
Conclusion
The antimicrobial activity of lactic acid bacteria may be due to a number of factors,including decreased pH levels, competition for substrates, and production of substanceswith a bactericidal or bacteriostatic action, including bacteriocins. The antimicrobial com-ponents secreted by probiotic strains may help to avoid pathogen colonization of mammalianintestine. Again, these antimicrobial components may find applications as food preservativesand in clinical studies.
�Fig. 4 HPLC chromatogram of primary separation after ammonium sulfate precipitation (a), reinjection insize exclusion chromatography (b), and the active fraction again injected in RP-HPLC for protein purificationpeak (c) (retention time, 35.63)
Table 3 Effects of pH, tempera-ture, and enzymes on the activityof bacteriocin PJ4
Values are presented as the mean± SEM (n=3)
Treatment Residual antimicrobial activity (%)
E. coli MTCC443
pH
2 71.23±0.16
4 83.25±0.43
6 91.76±0.23
8 54.17±0.42
10 32.04±0.24
Temperature
30 100
45 100
60 95.84±0.38
75 83.58±0.29
100 69.85±0.37
Enzyme
α-Amylase 100
Pepsin 83.64±0.43
Protease K 0
Trypsin 0
Lipase 0
Catalase 95±0.38
Appl Biochem Biotechnol
This is the first report characterizing the bacteriocin component of L. helveticus PJ4isolated from the gut microflora of male Wistar rat (R. norvegicus), an experimental animal.The bacteriocin was most effective in inhibiting the growth of various pathogenic strains likeE. coli, E. faecalis, S. aureus, E. faecium, K. pneumoniae, and P. aeruginosa. The partiallypurified bacteriocin was not only biodegradable, but also stable in a wide range of pH values(2.0–10.0), heat-resistant (30–100 °C), and has a size about 6.5 kDa. The bacteriocin PJ4may have potential application in the prevention and treatment of a few enteric diseases ingastrointestinal tract, which is a major problem in gastrointestinal disorders. However, thisneeds to be confirmed in future in vivo experiments. In this study, we report the partialcharacterizations of the antimicrobial compound bacteriocin PJ4; however, the identificationand chemical characterizations of these compounds must be carried out to elucidate theircomplete structure.
Acknowledgments The authors are thankful to the Nirma Education and Research Foundation (NERF),Ahmedabad for providing the infrastructure and financial support.
References
1. Culligan, E. P., Hill, C., & Sleator, R. D. (2009). Probiotics and gastrointestinal disease: successes,problems and future prospects. Gut pathogens, 1(19), 1–12.
2. Ballal, M., & Shivananda, P. G. (2002). Rotavirus and enteric pathogens in infantile diarrhoea in Manipal,South India. Indian Journal of Pediatrics, 69(5), 393–396.
3. Fuller, R. (1991). Probiotics in human medicine. Gut, 32(4), 439–442.4. Ahmed, F. E. (2003). Genetically modified probiotics in foods. Trends in Biotechnology, 21, 491–497.5. Beshkova, D., & Frengova, G. (2012). Bacteriocins from lactic acid bacteria: microorganisms of
potential biotechnological importance for the dairy industry. Engineering in Life Sciences, 12(4),1–14.
6. Hassan, M., Kjos, M., Nes, I. F., Diep, D. B., & Lotfipour, F. (2012). Natural antimicrobial peptides frombacteria: characteristics and potential applications to fight against antibiotic resistance. Journal of AppliedMicrobiology, 113(4), 723–736.
7. O'Shea, E. F., Cotter, P. D., Stanton, C., Ross, R. P., & Hill, C. (2012). Production of bioactive substancesby intestinal bacteria as a basis for explaining probiotic mechanisms: bacteriocins and conjugated linoleicacid. International Journal of Food Microbiology, 152(3), 189–205.
8. Klaenhammer, T. R. (1993). Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol-ogy Review, 12, 39–86.
9. Reid, G., & Burton, J. (2002). Use of Lactobacillus to prevent infection by pathogenic bacteria.Microbesand Infection, 4, 319–324.
10. Servin, A. L. (2004). Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens.FEMS Microbiology Reviews, 28(4), 405–440.
11. Spinler, J. K., Taweechotipatr, M., Rognerud, C. L., Ou, C. N., Tumwasorn, S., & Versalovic, J. (2008).Human-derived probiotic Lactobacillus reuteri demonstrate antimicrobial activities targeting diverseenteric bacterial pathogens. Anaerobe, 14(3), 166–171.
12. De Vuyst, L., & Vandamme, E. J. (1994). Bacteriocins of lactic acid bacteria: Microbiology, geneticsand applications. In L. de Vuyst & E. J. Vandamme (Eds.), Lactic acid bacteria and bacteriocins:their practical importance (pp. 1–11). London, United Kingdom London: Blackie Academic andProfessional.
13. Gong, H. S., Meng, X. C., & Wang, H. (2010). Plantaricin MG active against Gram-negative bacteriaproduced by Lactobacillus plantarum KLDS1.0391 isolated from “Jiaoke”, a traditional fermented creamfrom China. Food Control, 21, 89–96.
14. Heu, S., Oh, J. H., Kang, Y. S., Ryu, S., Cho, S. K., Cho, Y. S., et al. (2001). Cloning, expression, andpurification of glycinecin A, a bacteriocin produced by Xanthomonas campestris pv. glycines 8ra. Appliedand Environmental Microbiology, 67, 4105–4110.
15. Upreti, G. C., & Hinsdill, R. D. (1975). Production and Mode of Action of Lactocin 27: bacteriocin from aHomofermentative Lactobacillus. Antimicrobial Agents and Chemotherapy, 7(2), 139–145.
Appl Biochem Biotechnol
16. Joerger, M. C., & Klaenhammer, T. R. (1986). Characterization and purification of helveticin J andevidence for a chromosomally determined bacteriocin produced by Lactobacillus helveticus. Journal ofBacteriology, 167(2), 439–446.
17. Schägger, H., & Von Jagow, G. (1987). Tricine-Sodium Dodecyl Sulfate-Polyacrylamide Gel Electro-phoresis for the Separation of Proteins in the Range from 1 to 100 kDa. Analytical Biochemistry, 166,368–379.
18. Georgalaki, M. D., Berghe, E. V., Kritikos, D., Devreese, B., Beeumen, J. V., Kalantzopoulos, G., et al.(2002). Macedocin, a Food-Grade Lantibiotic Produced by Streptococcus macedonicus ACA-DC 198.Applied and Environmental Microbiology, 68(12), 5891–5903.
19. Todorov, S. D., Prevost, H., Lebosis, M., Dousset, X., LeBlanc, J. G., de Melo, G., et al. (2011).Bacteriocinogenic Lactobacillus plantarum ST16Pa isolated from papaya (Carica papaya) — Fromisolation to application: characterization of a bacteriocin. Food Research International, 44, 1351–1363.
20. Kumar, M., Tiwari, S. K., & Srivastava, S. (2010). Purification and characterization of enterocin LR/6, anew bacteriocin from Enterococcus faecium LR/6. Applied Biochemistry and Biotechnology, 160, 40–49.
21. Todorov, S. D., & Dicks, L. M. T. (2005). Characterization of bacteriocins produced by lactic acid bacteriaisolated from spoiled black olives. Journal of Basic Microbiology, 45, 312–322.
22. Albano, H., Todorov, S. D., Van Reenen, C. A., Hogg, T., Dicks, L. M. T., & Teixeira, P. (2007).Characterization of a bacteriocin produced by Pediococcus acidilactici isolated from “Alheira”, afermented sausage traditionally produced in Portugal. International Journal of Food Microbiology, 116,239–247.
23. Todorov, S. D., Wachsman, M., Tomé, E., Dousset, X., Destro, M. T., Dicks, L. M. T., et al. (2010).Characterisation of an antiviral pediocin-like bacteriocin produced by Enterococcus faecium. FoodMicrobiology, 27, 869–879.
24. Todorov, S. D., Furtado, D. N., Saad, S. M. I., de Melo, G., & Franco, B. D. (2011). Bacteriocinproduction and resistance to drugs are advantageous features for Lactobacillus acidophilus La-14, apotential probiotic strain. The New Microbiologica, 34, 357–370.
25. Giri, S. S., Sukumaran, V., Sen, S. S., Vinumonia, J., Nazeema, B. B., & Jena, P. K. (2011). AntagonisticActivity of Cellular Components of Potential Probiotic Bacteria, Isolated from the Gut of Labeo rohita,Against Aeromonas hydrophila. Probiotics and Antimicrobial Proteins, 3, 214–222.
26. Todorov, S., Onno, B., Sorokin, O., Chobert, J. M., Ivanova, I., & Dousset, X. (1999). Detection andcharacterization of a novel antibacterial substance produced by Lactobacillus plantarum ST31 isolatedfrom sourdough. International Journal of Food Microbiology, 48, 167–177.
27. Ray, B. (1994). Pediocins of Pediococcus species. In L. De Vuyst & E. J. Vandanme (Eds.), Bacteriocinsof Lactic Acid Bacteria: Microbiology, Genetics and Applications (pp. 465–495). London: Chapman andHall.
28. Noonpakdee, W., Santivarangkna, C., Jumriangrit, P., Sonomoto, K., & Panyim, S. (2003). Isolation ofnisin-producing Lactococcus lactis WNC20 strain from nham, a traditional Thai fermented sausage.International Journal of Microbiology, 81, 137–145.
29. Goa, Y., Jia, S., Goa, Q., & Tan, Z. (2010). A novel bacteriocin with a broad inhibitory spectrum producedby Lactobacillus sake C2, isolated from traditional Chinese fermented cabbage. Food Control, 1, 76–81.