HELICOBACTER PYLORI, PART I 08894553/00 $15.00 + .OO BACTERIOLOGY AND TAXONOMY OF HELICOBACTEX PYLORI Helen M. Windsor, PhD, and Jani ORourke, BSc Helicobacter pylori is a gram-negative, spiral-shaped organism associ- ated with gastrointestinal disease in humans. It has a worldwide preva- lence, with approximately 50% of the world's population infected. Before the first isolation and documentation of this organism from the human stomach in 1982, it was assumed that the human stomach was a sterile environment because of the high levels of acid, which would exclude it as an ecologic niche for any organism. This bacterium is the human- adapted Helicobacter primarily found in the gastric mucosa and areas of gastric metaplasia in the duodenum and occasionally in Meckel's diverticulum and the 76 It has been cultured rarely from fecesI1O2 and saliva.53 It can be detected by polymerase chain reaction (PCR) in dental plaque and 78 In the latter instances, the viability of the bacteria is in question. H. pylori also has been found in nonhuman primates and cats.12,29,44 H. pylori detection in animals is not common and could be due to human contact with animals. To date, no environmental reservoir has been shown. TAXONOMY The organism now known as H. pylori first was introduced to the scientific community in 1982. MarshalP2 and Warredo9 described a From the Helicobacter pylori Research Laboratory, Department of Microbiology, University of Western Australia, Perth (HW); and the School of Microbiology and Immunology, University of New South Wales, Sydney go), Australia GASTROENTEROLOGY CLINICS OF NORTH AMERICA VOLUME 29 * NUMBER 3 SEMEMBER 2000 633
Transcript
HELICOBACTER PYLORI, PART I 08894553/00 $15.00 + .OO
BACTERIOLOGY AND TAXONOMY OF HELICOBACTEX
PYLORI
Helen M. Windsor, PhD, and Jani ORourke, BSc
Helicobacter pylori is a gram-negative, spiral-shaped organism associ- ated with gastrointestinal disease in humans. It has a worldwide preva- lence, with approximately 50% of the world's population infected. Before the first isolation and documentation of this organism from the human stomach in 1982, it was assumed that the human stomach was a sterile environment because of the high levels of acid, which would exclude it as an ecologic niche for any organism. This bacterium is the human- adapted Helicobacter primarily found in the gastric mucosa and areas of gastric metaplasia in the duodenum and occasionally in Meckel's diverticulum and the 76 It has been cultured rarely from fecesI1O2
and saliva.53 It can be detected by polymerase chain reaction (PCR) in dental plaque and 78 In the latter instances, the viability of the bacteria is in question. H. pylori also has been found in nonhuman primates and cats.12,29,44 H. pylori detection in animals is not common and could be due to human contact with animals. To date, no environmental reservoir has been shown.
TAXONOMY
The organism now known as H. pylori first was introduced to the scientific community in 1982. MarshalP2 and Warredo9 described a
From the Helicobacter pylori Research Laboratory, Department of Microbiology, University of Western Australia, Perth ( H W ) ; and the School of Microbiology and Immunology, University of New South Wales, Sydney go), Australia
GASTROENTEROLOGY CLINICS OF NORTH AMERICA
VOLUME 29 * NUMBER 3 SEMEMBER 2000 633
634 WINDSOR & OROURKE
campylobacter-like bacterium that was seen in large numbers in the gastric mucus of patients with chronic gastritis and duodenal ulcers. S k i r r o ~ , ~ ~ a microbiologist in the Campylobacter field, suggested that if the organisms were proved to be a member of the genus Campylobacter, the name Campylobacter pyloridis (derived from the Greek pylorus for gatekeeper) would be apt because of their specific location and associa- tion. Successful culture of the bacterium resulted in the acceptance of this name,65 which was later corrected to Campylobacter pylori." Ongoing study of this organism, particularly at the genetic level, resulted in the formation of a new genus, Helicobacter (helico = curved; bacter = staff), with H. pylori as the type species.43 The analysis of the sequence of the 16s rRNA gene led to the differentiation of H. pylori from Campylobacter species, and this molecular tool also has been instrumental in the classi- fication of the other members of this genus. Other important features that differentiated Helicobacter species from Campylobacter species in- cluded the possession of sheathed flagella, unique fatty acid profile, lack of respiratory quinones, active urease enzyme, and a distinct protein pro- file.
MORPHOLOGY
H. pylori is a spiral to curved, rod-shaped bacterium approximately 0.5 km in diameter and 3 to 5 km long. This organism possesses the characteristic ultrastructure of a gram-negative bacterium. In tissue sections and Gram-stained smears from biopsy specimens, the bacteria usually appear smaller and more curved than cultured organisms, which are longer and less spiral.@ In older cultures, cells are seen to ball up, form U-shaped structures, and lose their cytoplasmic cylinders and membrane integrity, resulting in the formation of coccoid cells.17 It has been proposed that this coccoid form is a viable but nonculturable form of the organism, which allows it to survive in hostile environments outside the gastric mucus.18 Studies provide evidence for the concept that these coccoid forms are degenerative and nonculturable, however, with a significant decrease in the amount and integrity of RNA and DNA and a loss of membrane potential.55 This concept has been supported by molecular data, but it remains a controversial issue.1o5
H. pylori have 4 to 7 polar sheathed flagella, which enable the bacterium to move freely in viscous environments such as gastric mucus (Fig. 1).& Several studies have shown that this motility is essential for the bacterial colonization of its host.28 The flagella sheath is a membrane containing proteins and lipopolysaccharides, which probably protects the flagella filaments from the gastric The flagella filament contains two different flagellin proteins, FlaA and FlaB, both of which have been shown to be necessary for the motility of the organism. The genes for these proteins are not lmked on the chromosome and are regulated by different promoters (az8 and a"). It has been proposed that
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLON 635
Figure 1. Electron micrograph of Helicobacter pylori showing the polar sheathed flagella. Bar = 1.2 prn.
these latter features allow the bacterium to adapt to different environ- mental condition^.^^
PHYSIOLOGIC AND BIOCHEMICAL CHARACTERISTICS
H. pylovi is nutritionally fastidious and can be cultivated on defined media, such as blood agar, under microaerobic conditions or in the presence of air enriched with 10% carbon dioxide. Optimal growth is obtained at 37°C after 4 to 5 days for primary culture or 2 days for subsequent subculture. It is urease, catalase, and oxidase positive. Con- firmation of these phenotypic traits in collaboration with the characteris- tic spiral morphology and growth of small translucent colonies on selec- tive agar usually is adequate for the identification of H. pylori in the patient. The urease activity of this bacterium is striking, and the amounts produced have allowed accurate diagnosis in patients by direct detection of the enzyme in gastric biopsy specimens and by breath tests using carbon isotopes labeled with urea. Many roles have been proposed for
636 WINDSOR & OROURKE
this enzyme. It is known to be important for colonization and survival of the bacterium in the gastric environment.26 The hydrolysis of urea to ammonia by urease could have a buffering effect, protecting the bacterium from In vitro studies have shown that H. pylori cannot survive in acidic conditions without the presence of urea, and urea inhibits its growth in alkaline conditions.2o, 70, 87 This enzyme as well as amino acids, especially alanine, serine, and glutamine, and ammonia have been shown to be used as nitrogen sources for protein synthesis. Urease also has been proposed as an important virulence factor. Shedding of this enzyme from the cell surface may allow the bacterium to avoid host defense mechanisms, and the enzyme itself could have a direct effect on the host by inducing tissue damage or altering the immune response.67 The presence of catalase and superoxi- dase dismutase enzymes and the recA gene product, which is able to repair DNA damage resulting from oxidative attack, are thought to be important defense mechanisms for this bacterium against the oxidative damage to which it would be susceptible in areas of inflamed gastric
Additional biochemical characteristics of H . pylori include its inabil- ity to reduce nitrates or hydrolyze hippurate and indoxyl acetate. It exhibits leucine arylamidase, alkaline phosphatase, and gamma-gluta- myltranspeptidase activities. It does not grow in the presence of 1% glycine or 1.5% sodium chloride. Its major fatty acids are tetradecanoic acid (14:O) and 19-carbon cyclopropane fatty acid (19:OqC), and its major isoprenoid is MK-6.43 Initially, because of the use of standard methods to detect carbohydrate metabolism, H. pylori was thought to be asacchar- olytic. Studies have shown, however, that it can take up and use glucose by the pentose phosphate and Entner-Doudoroff pathways and glycoly- sis. Fermentation of glucose may lead to production of mixed acid products or the provision of metabolites for the Krebs cycle. Features of anaerobic and aerobic respiration have been reported, and there is evi- dence that H. pylori can degrade lipids as sources of carbon, phosphate, and energy.47
H. pylori is resistant to nalidixic acid, trimethoprim, sulfonamides, and vancomycin and is sensitive to penicillin, ampicillin, cephalothin, kanamycin, gentamicin, rifampin, and tetracycline. Variable resistance to metronidazole and clarithromycin is known. Antimicrobial sensitivity patterns are useful for the identification of H. pylori and have been employed in the design of selective supplements for the culture of the organism. It is known, however, that they do not mimic the sensitivities of the bacterium in its host. This situation has necessitated the use of multiple antimicrobial agents in combination with bismuth compounds or acid-suppressive agents or both for the successful eradication of the organism from infected persons.
H. pylori has been shown to have other characteristics, many of which are implicated as virulence factors and are discussed in detail elsewhere. Briefly, these characteristics include the following. Of strains, 50% produce a vacuolating cytotoxin, coded for by the gene ~ U C A . ~ ~ There are at least five different alleles of vacA with certain alleles commonly
mucosa.79, 85, 89.94, 103
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLORI 637
associated with more serious forms of gastric di~ease.~ The more patho- genic strains of H. pylori also possess a pathogenicity island (PAI), of which the cagA gene is a marker.19 The cag PA1 consists of 31 genes that encode for a type IV secretion system, which allows transfer of macromolecules outside the cell and between cells. The lipopolysaccha- ride of H. pylori has a low reactivity and is composed of mainly Lewis antigens.74 This latter feature allows H. pylori to mimic human cell surface glycolipids or glycoproteins. Two possible consequences of this molecular mimicry could be the ability of the bacterium to evade host defense mechanisms and the induction of cross-reactive autoantibodies, leading to mucosal damage.6 Several adhesins and other outer mem- brane proteins have been found in this organism, and they have been implicated in its virulence and development of antigenic ~a r i a t ion .~~
GENETICS
In 1997, the complete genomic sequence of H. pylori strain 26695 was published.'05 This bacterium has a single circular chromosome of 1,667,867 base pairs and 1590 predicted coding sequences (i.e., about 91% of the total genome), of which 1091 matched database sequences of genes are known from other organisms. The small size of the H. pylori genome (1.7 mebagase [Mb]), as compared with that of other bacteria that live in a wide range of habitats, such as Escherichia coli (4.6 Mb) and Pseudornonas aeruginosa (5.8 Mb), shows that H. pylori has fewer regula- tory genes that help other bacteria adapt to new environments. These findings support epidemiologic evidence that H. pylori lives only in the human stomach and that it does not possess enzymatic pathways to survive in other environments. It was also determined from the pre- dicted coding sequences that the amino acids, arginine and lysine, occur twice as frequently in H. pylori proteins as in those of Haemophilus influenzae and E. coli, perhaps reflecting an adaptation of H. pylori to gastric acidity.
Study of the genome sequence has shown that there are more than 20 previously unknown H. pylori genes that are likely to be needed for the assembly or activity of flagella, which are essential for mobility and colonization by the bacteri~m.~ There are also 12 possible genes coding for the acquisition of iron, which is an essential micronutrient for the survival of H. pylori. This knowledge has not helped biochemists in their understanding of iron acquisition, however, because some genes have been found in multiple copies, and others that were expected to be found (after in vitro experiments of the uptake of iron in various forms) were not identified. This is clearly a field where the genome sequence has provided more questions than answers.
Other genes that have been identified from the complete genome sequence include genes coding for outer membrane proteins and lipo- polysaccharide molecules, the known cagA gene and its associated PAI, and the area coding for the cytotoxic protein, VacA. Before the publica- tion of the complete genomic sequence, physical and genetic maps of
638 WINDSOR & OROURKE
the genomes of at least five H. pylori strains had been c~nstructed,'~, 99
and important genes coding for urease activity:2, 56 Cytotoxin2, 23, lo7 and adhesin production,31 and flagellin formation98 had been cloned and characterized. Many different techniques have been used to attempt to find similarities between isolates of H. pylori (see subsequently); for example, in a study using random amplified polymorphic DNA primers in a new PCR fingerprinting technique, 60 independent clinical isolates could be distinguished from one another.' One constant fact emerged from all these studies of the genetics of H. pylori, and that was the genomic variability between strains. Alm et a14 compared the genomic sequences of two unrelated H. pylori isolates and found that the overall genomic organization and predicted groups of proteins were quite simi- lar. About 7% of the genes were specific to each individual strain, however, and most of these were clustered in a single hypervariable region. Previously, comparison of the genetic maps of five H. pylori strains showed that there was no gene clustering or characteristic ar- rangement of 17 known genes on the chromosomes of these strains.49 There is variation in the DNA base composition of different strains. The DNA from 32 H. pylori strains had an average G + C content of 35.2 mol% with a range of 34.1 to 37.5 mol%, but strain 26695 used by Tomb et aP5 had an average G + C content of 39 mol%, with one region having a 43% ratio.8
This research shows that H. pylori has extensive diversity within certain regions of the circular chromosome (macrodiversity) as well as DNA sequence diversity within the genes themselves (microdiversity). These differences may be due to frequent mutations within a particular gene, possibly base substitutions that conserve important amino acids or allow substitutions that conserve functional a~tivity?~ Another mecha- nism that may contribute to this diversity is the evidence of a conjuga- tion-like mechanism of DNA transfer in H. p y h i X The ability of the organisms to take up DNA from lysed bacteria in the gut by transforma- tion or conjugation shows that there is a horizontal DNA exchange of genetic material between strains in the same environment.
Studies of H. pylori strains worldwide show that approximately 50% of H. pylori strains carry plasmids, with sizes ranging from 1.5 to 40 kb.72, Io4 These plasmids have highly variable restriction patterns and are all cryptic (they do not code for any known phenotype). Other gram- negative bacteria often carry plasmids that confer some form of antibiotic resistance to these strains, but in H. pylori no plasmids of this type have been isolated, and antibiotic resistance genes are located on the chromosome.
MOLECULAR TYPING
The DNA from most bacterial species gives a characteristic finger- print or pattern when analyzed, which can be used by epidemiologists to find similarities between strains within the human population and by
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLON 639
clinicians to determine if continued bacterial infection is due to treatment failure or reinfection. In the case of H. pylori, molecular typing has not proved to be an easy task because of the high degree of genetic variabil- ity among strains. Many different methods have been tried by different researchers, and a review of the various methods follows.
Early studies using conventional gel electrophoresis and restriction enzyme digestion using enzymes such as Hind I11 and Hue I11 showed that each set of clinical isolates of H. pylovi had a unique digestion pattern.60. 82 Similarly, ribotyping, using enzyme-digested ribosomal RNA and hybridizing with a labeled probe, highlighted some similarities between strains but is not an ideal method because of strain diversity.lm Pulsed-field gel electrophoresis with enzymes such as Not I and Nru I gave discrete, well-separated DNA fragments in some strains, but in other strains no consistent results could be obtained, and almost all unrelated strains gave different 99 This method was one of those used to construct the first genetic map of H. p y l ~ r i . ~ ~
A technique that does not rely on restriction sites or on individual genes but uses arbitrary oligonucleotides as primers in PCR amplifica- tion of random DNA fragments is random amplified polymorphic DNA (RAPD).', 66 This method has been used in a study of 64 H. pylori isolates, 60 of which were from patients in the same hospital. Each of the strains had a distinct fingerprint, but duplicate isolates from some of the patients 2 to 4 months after treatment failure were identical to the original amplification profiles, showing the reproducibility of this tech- nique as well as the failure of the treatment to eradicate the original organism (rather than reinfection with a new strain). RAPD now is widely used as a sensitive fingerprinting method, but it has limitations because whole chromosomal DNA is required so that a pure culture of the isolate is needed, and this method cannot be used on archival material.
Another method that uses PCR but does not have the limitations of RAPD is restriction fragment-length polymorphism (PCR-RFLP).2 This method has been used to differentiate between H. pylori strains using amplified sections of a specific gene, which are examined after restriction enzyme digestion to find allelic differences. Different H. pylori genes that have been compared using this method are the urease genes (ureA, ureB, ureC, and ~ r e D ) , ~ ~ , 73, 83 flagellin genes VaA and ~ I u B ) , ~ ~ and the adhesin gene (hpaA).3O This method can be used on archival material, but prob- lems can arise if more than one strain is present in the specimen. Careful addition of the size of the resulting fragments after RFLP can be used to prove the existence of a mixed population of H. pylori, however.
One of the newest methodologies using PCR is the amplification of DNA between repetitive DNA sequences (REP-PCR).Q 71 In one study, H. pylori isolates were obtained from 70 individuals using REP-PCR." Each H. pylori strain except for two had a distinct DNA fingerprint. Single-colony DNA fingerprints of H. pylori from the same patient were identical, suggesting that each patient harbored a single strain. Com- puter-assisted cluster analysis of the REP-PCR DNA fingerprints showed
640 WINDSOR & OROURKE
two large clusters of isolates, one associated with simple gastritis and the other with duodenal ulcer disease, suggesting that duodenal ulcer isolates, as a group, are more similar to one another and different from gastritis isolates. These results suggest that disease-specific strains of H. pylori may exist.
OTHER HELICOBACTERS
The genus Helicobacter at present consists of 18 official genera with another 10 potentially novel species.80 Of the official genera, 7 are of gastric origin, and the remaining 11 are found in the intestinal tract of a wide variety of animal species (Table 1). Several of these latter organisms also have been implicated in human disease.
Gastric Helicobacters
Helicobacter-like bacteria have been found in an extensive range of animal species as well as humans. H. pylori is the dominant human- adapted organism. The other gastric helicobacters include H. nemestrinae in pig-tailed macaques, H. acinonyckis in cheetahs, H. mustelae in ferrets, H . felis in cats and dogs, and H. bizzozeronii and H. salomonis in dogs.'l, 27, 43, 45, 48, The most prevalent animal helicobacter is Gastrospirillum kominis or Helicobacter keilmannii. These bacteria have been observed in a variety of nonhuman primates, in native and domestic cats and dogs,
Table 1. NATURAL HOSTS AND USUAL SITE OF ISOLATION OF HELICOBACTER SPECIES
Helicobacter Species Main Host Origin
H. pylori H. mustelae H. nemestrinae H. felis H. acinonychis H. bizzozeronii H. salomonis H. heilmannii H. cinaedi H. fennelliae H. muridarum H. canis H. pullorum H. pametensis H. hepaticus H. bilis H. cholecystus H. trogontum H. rodentiurn
Human Ferret Macaque monkey Cat, dog Cheetah Dog Dog Human, cat, dog, pig Human, hamster Human, hamster Rat, mouse Dog Poultry Tern Mouse Mouse Hamster Rat Mouse
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLORI 641
-
in pigs, and in a small percentage of humans.58 Morphologically, they are a large helical organism, 0.4 to 0.9 pm in diameter and 4 to 10 pm long with bipolar bundles of sheathed flagella, but as yet they have not been cultured successfully in vitro. In their natural environment of gastric mucus, they are motile, exhibiting a corkscrew pattern of move- ment, and generally biopsy specimens from infected animals or humans are positive for urease activity.
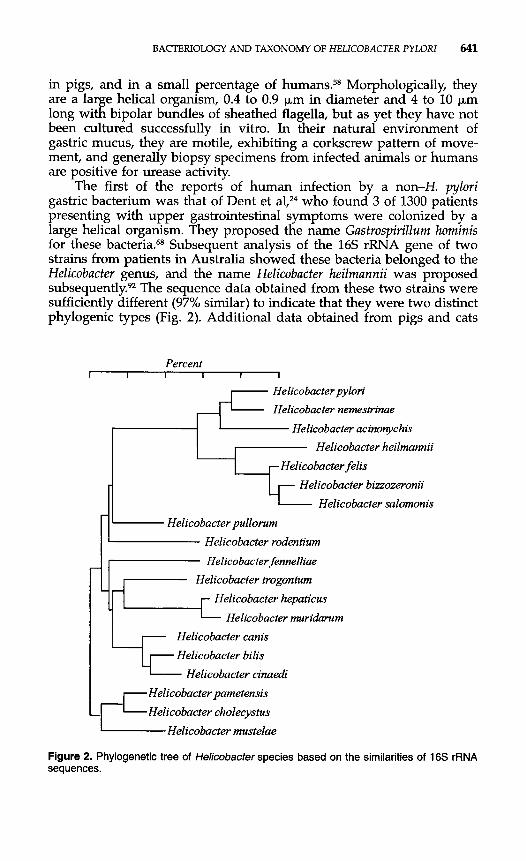
The first of the reports of human infection by a non-H. pylori gastric bacterium was that of Dent et al,24 who found 3 of 1300 patients presenting with upper gastrointestinal symptoms were colonized by a large helical organism. They proposed the name Gastrospirillurn horninis for these bacteria.68 Subsequent analysis of the 16s rRNA gene of two strains from patients in Australia showed these bacteria belonged to the Helicobacter genus, and the name Helicobacter heilrnannii was proposed subsequently?2 The sequence data obtained from these two strains were sufficiently different (97% similar) to indicate that they were two distinct phylogenic types (Fig. 2). Additional data obtained from pigs and cats
- Helicobacter pullorurn
Helicobacter rodentium
Helicobacter fennelliae Helicobacter trogontum -
Percent
Figure 2. Phylogenetic tree of Helicobacter species based on the similarities of 16s rRNA sequences.
642 WINDSOR & OROURKE
have shown almost complete homology of the 16s rRNA gene from the gastric bacteria in these animals with one or other of the two human
69 To date, only one other helicobacter has been successfully cultured from a dyspeptic patient with ga~tritis.~ Studies have shown that although this organism is similar to the H. heilmannii-like bacteria, it is closer to H . bizzozeronii (K Jalava: personal communication). There also have been two cases of patients diagnosed with severe acute gastri- tis in which bacteria ultrastructurally resembling H. felis were seen.57, Overall, most patients in whom the H. heilmannii-like bacteria are seen are diagnosed with mild chronic gastritis. Increasing numbers of reports of infections by these bacteria show they can be associated with more serious forms of gastric disease, however, including peptic ulceration,1° acute infection^,^ and gastric Of most concern is the high incidence of gastric mucosa-associated lymphoid tissue lymphoma asso- ciated with H. heilmannii when compared with similar populations in- fected with H. py10ri.~~
Nongastric Helicobacters
Lower bowel helicobacters now are being found commonly in the gastrointestinal mucus in a variety of animals, with suggestions these bacteria could be associated with human disease and as such are exam- ples of a zoonosis. They can be isolated from rodents (Helicobacter bilis, Helicobacter hepaticus, Helicobacter muridarum, Helicobacter trogontum, Heli- cobacter rodentium), hamsters (Helicobacter fennelliae, Helicobacter cinaedi, Helicobacter cholecystus), dogs (Helicobacter canis), sheep and pigs (Flex- ispira rappini), and chickens and birds (Helicobacter pullorum, Helicobacter pametensis).80 (Although F. rappini is not an official member of the genus, phylogenetic studies have shown it is a Helicobacter species). In their natural hosts, several of these species have been found in association with varying disease states, including diarrhea (H. canis),96 hepatitis (H. canis, H. hepaticus, H . bilis, H. pullor~m),~~, 36, 38, 95 hepatic adenoma and hepatocellular carcinoma (H. hepati~us),~~, *08 abortion ( E rappini),13 and cholangiofibrosis and pancreatitis (H. cholecyst~s).~~ Experimental infec- tions of immunocompromised mice with H. hepaticus or H. bilis also have shown lesions resembling those seen in inflammatory bowel disease.l6. 90
Of more relevance to clinicians and gastroenterologists is the in- creasing number of reports of finding these helicobacters in humans. Helicobacters have been found in blood cultures (H. cinaedi, H. fennelliae, E ~appini)~~, 52* 93, lol; associated with diarrhea (H. pullorurn, H. fennelliae, H. cinaedi, H. cunis)l5, 95, 96; and in cases of proctitis and protocolitis, with or without the presence of human immunodeficiency virus, in homosexual men (H. fennelliae, H. cinaedi).lo6 There have been two indi- vidual reports of the presence of Helicobacter species (H. bilis, H. pullorurn, and F. rappini) in bile and gallbladder samples from patients with chronic cholecystitis, with the suggestion they could be involved in gallbladder cancer.34, 86 Although it seems that the lower bowel helicobacters are
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLON 643
members of the normal microbiota of the gastrointestinal tract, there is increasing evidence that under certain conditions these bacteria can translocate to other organs, such as the liver and gallbladder, where they could induce more serious diseases.
SUMMARY
As the scientific community approaches the twentieth anniversary of the first isolation of H. pylori, it appears that despite the wealth of articles published in journals throughout the world every month, there are still many unanswered questions about the microbiology of this bacterium and others in the genus HeZicobacter.
References
1. Akopyantz N, Bukanov NO, Westblom TU, et al: DNA diversity among clinical isolates of Helicobacter pylori detected by PCR-based RAPD fingerprinting. Nucleic Acids Res 205137-5142, 1992
2. Akopyanz N, Bukanov NO, Westblom TU, et al: PCR-based RFLP analysis of DNA sequence diversity in the gastric pathogen Helicobacter pylori. Nucleic Acids Res 2062214225, 1992
3. Al-Himyary AJS, Zabaneh lU, Zabaneh SS, et al: Gustrospirillum hominis in acute gastric erosion. South Med J 871147-1150, 1994
4. Alm RA, Ling LSL, Moir DT, et al: Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 397176-180, 1999
5. Andersen LP, Norgaard A, Holck S, et al: Isolation of a Helicobacter heilmanii-like organism from the human stomach. Eur J Clin Microbiol Infect Dis 15:95-96, 1996
6. Appelmelk BJ, Simoonssmit I, Negrini R, et al: Potential role of molecular mimicry between Helicobacter pylori lipopolysaccharide and host Lewis blood group antigens in autoimmunity. Infect Immun M2031-2040, 1996
7. Atherton JC, Cao P, Peek RM, et a1 Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori-association of specific vaca types with cytotoxin production and peptic ulceration. J Biol Chem 270:17771-17777, 1995
8. Beji A, Megraud F, Vincent P, et al: GC content of DNA of Campylobacter pylori and other species belonging or related to the genus Campylobacter. Ann Inst Pasteur Microbiol 139:527-534, 1988
9. Berg DE, Hoffman PS, Appelmalk BJ, et al: The Helicobacter pylori genome sequence: Genetic factors for long life in the gastric mucosa. Trends Microbiol 5:468-474, 1997
10. Borody TJ, George LL, Brand1 S, et al: Helicobacter pylori-negative duodenal ulcer. Am J GastroenterolS6:1154-1157, 1991
11. Bronsdon MA, Goodwin CS, Sly LI, et al: Helicobacter nemestrinae sp. nov., a spiral bacterium found in the stomach of a pigtailed macaque (Macaca nemestrina). Int J Syst Bacteriol 41:148-153, 1991
12. Bronsdon MA, Schoenknecht F D Campylobacter pylori isolated from the stomach of the monkey, Macaca nernestrina. J Clin Microbiol26:17251728, 1988
13. Bryner JH, Ritchie AE, Pollet L, et al: Experimental infection and abortion of pregnant guinea pigs with a unique spirillum-like bacterium isolated from aborted ovine fetuses. Am J Vet Res 48:91-95, 1987
14. Bukanov NO, Berg DE: Ordered cosmid library and high-resolution physical-genetic map of Helicobacter pylori strain NTCT11638. Mol Microbiol 11:509-523, 1994
15. Bumens AP, Stanley J, Schaad UB, et al: Novel Campylobacter-like organism resembling
644 WINDSOR & OROURKE
Helicobacter fennelliae isolated from a boy with gastroenteritis and from dogs. J Clin Microbiol 31:191&1917, 1993
16. Cahill RJ, Foltz CJ, Fox JG, et al: Inflammatory bowel disease-an immunity-mediated condition triggered by bacterial infection with Helicobacter hepaticus. Infect Immun 65:31263131, 1997
17. Catrenich CE, Makin KM: Characterization of the morphologic conversion of Helico- bacter pylori from bacillary to coccoid forms. Scand J Gastroenterol 26:58-64, 1991
18. Cellini L, Allocati N, Dicampli E, et al: Helicobacter pylori-a fickle germ. Microbiol Immunol38:25-30, 1994
19. Censini S, Lange C, Xiang ZY, et al: Cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc Natl Acad Sci
20. Clyne M, Labigne A, Drumm 8: Helicobacter pylori requires an acidic environment to survive in the presence of urea. Infect Immun 63:1669-1673, 1995
21. Cothi de G, Newbold K, OConnor H: Carnpylobacter-like organisms and heterotopic gastric mucosa in Meckel's diverticula. J Clin Pathol42:132-134, 1989
22. Covacci A, Censini S, Bugnoli M, et al: Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duo- denal ulcer. Proc Natl Acad Sci U S A 905791-5795, 1993
23. Cover TL, Blaser MJ: Purification and characterization of the vacuolating toxin from Helicobacter aulori. 1 Biol Chem 26710570-10575, 1992
U S A 93:14648-14653, 1996
24. Dent JC, M<y'CAM, Uff JC, et al: Spiral organisms in the gastric antrum. Lancet 296, 1987
25. Dieterich C, Wiesel P, Neiger R, et al: Presence of multiple Helicobacter heilrnannii strains in an individual suffering from ulcers and in his two cats. J Clin Microbiol
26. Eaton KA, Brooks CL, Morgan DR, et al: Essential role of urease in pathogenesis of gastritis induced by Helicobacter pylori in gnotobiotic piglets. Infect Immun 59:2470- 2475, 1991
27. Eaton KA, Dewhirst FE, Radin MJ, et al: Helicobacter acinonyx sp. nov., isolated from cheetahs with gastritis. Int J Syst Bacteriol 43:99-106, 1993
28. Eaton KA, Morgan DR, Krakowka S: Motility as a factor in the colonisation of gnotobiotic piglets by Helicobacter pylori. J Med Microbiol 37123-127, 1992
29. Euler AR, Zurenko GE, Moe JB, et al: Evaluation of two monkey species (Macaca rnulatta and Macaca fascicularis) as possible models for human Helicobacter pylori disease. J Clin Microbiol28:2285-2290, 1990
30. Evans DG, Evans DJ, Lampert HC, et al: Restriction fragment length polymorphism in the adhesin gene hpaA of Helicobacter pylori. Am J Gastroenterol90:1282-1288, 1995
31. Evans DG, Karjalainen TK, Evans DJ, et al: Cloning, nucleotide sequence, and expres- sion of a gene encoding an adhesin subunit protein of Helicobacter pylori. J Bacteriol 175:67&683, 1993
32. Ferrero RL, Cussac V, Courcoux P, et al: Construction of isogenic urease-negative mutants of Helicobacter pylori by allelic exchange. J Bacteriol 17442124217, 1992
33. Forbes KJ, Fang Z, Pennington TH: Allelic variation in the Helicobacter pylori flagellin genes JaA and PUB: Its consequences for strain typing schemes and population structure. Epidemiol Infect 114257-266, 1995
34. Fox JG, Dewhirts FE, Shen ZL, et al: Hepatic Helicobacter species identified in bile and gallbladder tissue from Chileans with chronic cholecystitis. Gastroenterology
35. Fox JG, Dewhirst FE, Tully JG, et al: Helicobacter hepaticus sp nov, a microaerophilic bacterium isolated from livers and intestinal mucosal scrapings from mice. J Clin Microbiol 32:1238-1245, 1994
36. Fox JG, Drolet R, Higgins R, et al: Helicobacter canis isolated from a dog liver with multifocal necrotizing hepatitis. J Clin Microbiol342479-2482, 1996
37. Fox JG, Li X, Yan L, et al: Chronic proliferative hepatitis in A/JCR mice associated with persistent Helicobacter hepaticus infection-a model of Helicobacter-induced carci- nogenesis. Infect Immun 64:1548-1558, 1996
38. Fox JG, Yan LL, Dewhirst FE, et al: Helicobacter bilis sp nov, a novel Helicobacter
36~1366-1370, 1998
114~755-763, 1998
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLORI 645
species isolated from bile, livers, and intestines of aged, inbred mice. J Clin Microbiol 33445-454, 1995
39. Foxall PA, Hu L, Mobley HL: Use of polymerase chain reaction-amplified Helicobacter pylori urease structural genes for differentiation of isolates. J Clin Microbiol 30:739- 741, 1992
40. Franklin CL, Beckwith CS, Livingston RS, et al: Isolation of a novel helicobacter species, Helicobacter cholecystus sp nov, from the gallbladders of syrian hamsters with cholangiofibrosis and centrilobular pancreatitis. J Clin Microbiol 342952-2958, 1996
41. Geis G, Suerbaum S, Forsthoff B, et al: Ultrastructure and biochemical studies of the flagellar sheath of Helicobacter pylori. J Med Microbiol 38:371-377, 1993
42. Go MF, Chan KY, Versalovic J, et al: Cluster analysis of Helicobacter pylori genomic DNA fingerprints suggests gastroduodenal disease-specific associations. Scand J Gas- troenterol 30640-646, 1995
43. Goodwin CS, Armstrong JA, Chilvers T, et al: Transfer of Campylobacter pylori and Campylobacter mustelae to Helicobacter pylori gen. nov. and Helicobacter mustelae comb. nov. respectively. Int J Syst Bacteriol 39:397405, 1989
44. Handt LK, Fox JG, Dewhirst FE, et al: Helicobacter pylori isolated from the domestic cat: Public health implications. Infect Immun 62:2367-2374, 1994
45. Hanninen ML, Happonen I, Saari S, et al: Culture and characteristics of Helicobacter bizzozeronii, a new canine gastric helicobacter sp. Int J Syst Bacteriol 46160-166, 1996
46. Hazell SL, Lee A, Brady L, et al: Campylobacter pyloridis and gastritis: Association with intercellular spaces and adaptation to an environment of mucus as important factors in colonization of the gastric epithelium. J Infect Dis 153:658-663, 1986
47. Hazell SL, Mendz GL: How Helicobacter pylori works-an overview of the metabolism of Helicobacter pylori. Helicobacter 23-12, 1997
48. Jalava K, Kaartinen M, Utriainen M, et al: Helicobacter salomonis sp. nov., a canine gastric Helicobacter sp. related to Helicobacter felis and Helicobacter bizzozeronii. Int J Syst Bacteriol 47975-982, 1997
49. Jiang Q, Hiratsuka K, Taylor DE: Variability of gene order in different Helicobacter pylori strains contributes to genome diversity. Mol Microbiol 20:833-842, 1996
50. Josenhans C, Suerbaum S Flagella and motility of Helicobacter pylori. In Moran AP (ed): Pathogenesis and Host Response in Helicobacter pylori. Bad Homburg, Normed Verlag, 1997, pp 6-15
51. Kemper CA, Mickelsen P, Morton A, et al: Helicobacter-(Campylobacter) fennelliae-like organisms as an important but occult cause of bacteraemia in a patient with AIDS. J Infect 26:97-101, 1993
52. Kiehlbauch JA, Tauxe RV, Baker CN, et al: Helicobacter cinaedi-associated bacteremia and cellulitis in immunocompromised patients. Ann Intern Med 121:90-93, 1994
53. Krajden S, Fuksa M, Anderson J, et al: Examination of human stomach biopsies, saliva, and dental plaque for Carnpylobacter pylori. J Clin Microbiol 273397-1398, 1989
54. Kuipers EJ, Israel DA, Kusters JG, et al: Evidence for a conjugation-like mechanism of DNA transfer in Helicobacter pylori. J Bacteriol 180:2901-2905, 1998
55. Kusters JG, Gerrits MM, Vanstrijp J, et al: Coccoid forms of Helicobacter pylori are the morphologic manifestation of cell death. Infect Immun 65:3672-3679, 1997
56. Labigne A, Cussac V, Courcoux P: Shuttle cloning and nucleotide sequences of Helicobacter pylori genes responsible for urease activity. J Bacteriol1731920-1931,1991
57. Lavelle JP, Landas S, Mitros FA, et al: Acute gastritis associated with spiral organisms from cats. Dig Dis Sci 393744-750, 1994
58. Lee A, ORourke J: Gastric bacteria other than Helicobacter pylori. Gastroenterol Clin North Am 2221-42, 1993
59. Leunk RD, Johnson PT, David BC, et al: Cytotoxic activity in broth-culture filtrates of Campylobacter pylori. J Med Microbiol 26:9>99, 1988
60. Majewski SI, Goodwin CS: Restriction endonuclease analysis of the genome of Campy- lobacter pylori with a rapid extraction method: Evidence for considerable genomic variation. J Infect Dis 157465471, 1988
61. Mapstone NP, Lewis FA, Tompkins DS, et a1 PCR identification of Helicobacter pylori in faeces from gastritis patients. Lancet 341:447, 1993
646 WINDSOR & OROURKE
62. Marshall BJ: Unidentified curved bacillus on gastric epithelium in active chronic gastritis. Lancet 1:1273-1275, 1983
63. Marshall BJ, Barrett LJ, Prakash C, et al: Urea protects Helicobacter (Campylobacter) pylori from the bactericidal effect of acid. Gastroenterology 99:697-702, 1990
64. Marshall BJ, Goodwin CS: Revised nomenclature of Campylobacter pyloridis. Int J Syst Bacteriol 3768, 1987
65. Marshall BJ, Royce H, Annear DI, et al: Original isolation of Cumpylobacter pyloridis from human gastric mucosa. Microbiol Lett 25:8S8, 1984
66. Marshall DG, Chua A, Keeling PWN, et al: Molecular analysis of Helicobacter pylori populations in antral biopsies from individual patients using randomly amplified polymorphic DNA (RAPD) fingerprinting. FEMS Immunol Med Microbiol 10:317- 323, 1995
67. McGee DJ, Mobley HTL: Mechanisms of Helicobacter pylori infection: Bacterial factors. In Westblom TU, Czinn SJ, Nedrud JG (eds): Gastroduodenal Disease and Helicobacter pylori: Pathophysiology, Diagnosis and Treatment. Berlin, Springer, 1998, pp 15.5180
68. McNulty CAM, Dent JC, Curry A, et al: New spiral bacterium in gastric mucosa. J Clin Pathol42:585-591, 1989
69. Mendes EN, Queiroz DMM, Coimbra RS, et al: Experimental infection of wistar rats with Gastrospirillum suis. J Med Microbiol 44:10%109, 1996
70. Meyer Rosberg K, Scott DR, Rex D, et al: The effect of environmental pH on the proton motive force of Helicobacter pylori. Gastroenterology 111:88&900, 1996
71. Miehlke S, Genta RM, Graham DY, et al: Molecular relationships of Helicobacter pylori strains in a family with gastroduodenal disease. Am J Gastroenterol 943366368, 1999
72. Minnis JA, Taylor TE, Knesek JE, et al: Characterization of a 3.5-kbp plasmid from Helicobacter pylori. Plasmid 34:22-36, 1995
73. Moore RA, Kureishi A, Wong S, et al: Categorization of clinical isolates of Helicobacter pylori on the basis of restriction digest analyses of polymerase chain reaction-amplified ureC genes. J Clin Microbiol31:1334-1335, 1993
74. Moran AP: The role of lipopolysaccharide in Helicobacter pylori pathogenesis. Aliment Pharmacol Therapeut lO(supp1 1):39-50, 1996
75. Morgner A, Bayerdorffer E, Meining A, et al: Helicobacter heilmannii and gastric cancer. Lancet 346:511-512, 1995
76. Morris A, Nicholson G, Zwi J, et al: Campylobacter pylori infection in Meckel's diverti- cula containing gastric mucosa. Gut 30:1233-1235, 1989
78. Nguyen AMH, Engstrand L, Genta RM, et al: Detection of Helicobacter pylori in dental plaque by reverse transcription polymerase chain reaction. J Clin Microbiol 31:783-787, 1993
79. Odenbreit S, Wieland B, Haas R Cloning and genetic characterization of Helicobacter pylori catalase and construction of a catalase-deficient mutant strain. J Bacteriol 178:6960-6967, 1996
80. On SL, Lee A, ORourke JL, et al: Genus Helicobacter. In Goodwin, Armstrong, Chilvers, et a1 (eds): Bergey's Manual of Systematic Bacteriology, vol 2. 2000
81. Orlicek SL, Welch DF, Kuhls TL: Septicemia and meningitis caused by Helicobacter cinaedi in a neonate. J Clin Microbiol 31:569-571, 1993
82. Owen RJ, Fraser J, Costas M, et al: Signature patterns of DNA restriction fragments of Helicobacter pylori before and after treatment. J Clin Pathol 43:646449, 1990
83. Owen RJ, Slater ER, Xerry J, et al: Development of a scheme for genotyping Helico- bacter pylori based on allelic variation in urease subunit genes. J Clin Microbiol 363710-3712, 1998
84. Paster BJ, Lee A, Fox JG, et al: Phylogeny of Helicobacterfelis sp. nov., Helicobacter mustelae, and related bacteria. Int J Syst Bacteriol 41:31-38, 1991
85. Pesci EC, Pickett CL: Genetic organization and enzymatic activity of a superoxide dismutase from the microaerophilic human pathogen, Helicobacter pylori. Gene 143:lll-116, 1994
86. Roe IH, Chin YJ, Kim JT, et al: Identification of Helicobacter in diseased human bile. Gastroenterology 116(supp1):~0890, 1999
BACTERIOLOGY AND TAXONOMY OF HELICOBACTER PYLOlU 647
87. Sachs G, Meyer-Rosberg K, Scott DR, et al: Acid, protons and Helicobacter pylori. Yale J Biol Med 69:301-316, 1996
88. Salama SM, Jiang Q, Chang N, et al: Characterization of chromosomal DNA profiles from Helicobacter pylori strains isolated from sequential gastric biopsy specimens. J Clin Microbiol 33:249&2497, 1995
89. Schmitt W, Odenbreit S, Heuermann D, et al: Genetic characterisation of the Helico- bacter pylori recA gene. Gut 37(suppl):A21, 1995
90. Shomer NH, Dangler CA, Schrenzel MD, et al: Helicobacter bilis-induced inflammatory bowel disease in scid mice with defined flora. Infect Immun 65:4858-4864, 1997
91. Skirrow MB: Taxonomy and biotyping: Report on the session. In Pearson DA, Shirow MB, Rowe B, et a1 (eds): Campylobacter II. London, Public Health Laboratory Service, 1983, pp 3S38
92. Solnick JV, ORourke J, Lee A, et al: An uncultured gastric spiral organism is a newly identified helicobacter in humans. J Infect Dis 168:379-385,1993
93. Sorlin P, Vandamme P, Nortier J, et al: Recurrent "Flexispira rappini" bacteremia in an adult patient undergoing hemodialysis: Case report. J Clin Microbiol 371319-1323, 1999
94. Spiegelhalder C, Gerstenecker 8, Kersten A, et al: Purification of Helicobacter pylori superoxide dismutase and cloning and sequencing of the gene. Infect Immun 61:5315 5325, 1993
95. Stanley J, Linton D, Bumens AP, et al: Helicobacter pullorum sp nov-genotype and phenotype of a new species isolated from poultry and from human patients with gastroenteritis. Microbiology 140:3441-3449, 1994
96. Stanley J, Linton D, Bumens AP, et al: Helicobacter canis sp. nov., a new species from dogs-an integrated study of phenotype and genotype. J Gen Microbiol 139:2495- 2504, 1993
97. Stolte M, Kroher G, Meining A, et al: Comparison of Helicobacter pylori and H. heilmannii gastritis-matched control study involving 404 patients. Scand J Gastroen- terol 32:28-33, 1997
98. Suerbaum S, Josenhans C, Labigne A Cloning and genetic characterization of the Helicobacter pylori and Helicobacter mustelae flaB flagellin genes and construction of H. pylori flaA-negative and PUB-negative mutants by electroporation-mediated allelic exchange. J Bacteriol 1753278-3288, 1993
99. Taylor DE, Eaton M, Chang N, et a1 Construction of a Helicobacter pylori genome map and demonstration of diversity at the genome level. J Bacteriol 174:680&6806, 1992
100. Tee W, Lambert J, Smallwood R, et al: Ribotyping of Helicobacter pylori from clinical specimens. J Clin Microbiol30:1562-1567, 1992
101. Tee W, Street AC, Spelman D, et al: Helicobacter cinaedi bacteraemia-varied clinical manifestations in three homosexual males. Scand J Infect Dis 28:199-203, 1996
102. Thomas JE, Gibson GR, Darboe MK, et al: Isolation of Helicobacter pylori from human faeces. Lancet 340:1194-1195, 1992
103. Thompson SA, Blaser MJ: Isolation of the Helicobacter pylori recA gene and involve- ment of the recA region in resistance to low pH. Infect Immun 63:21852193, 1995
104. Tjia TN, Harper WES, Goodwin CS, et al: Plasmids in Campylobacter pyloridis. Microb- ios 36:7-11, 1987
105. Tomb JF, White 0, Kerlavage AR, et al: The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 388:539-547, 1997
106. Totten PA, Fennel1 CL, Tenover FC, et al: Carnpylobacter cinaedi (sp. nov.) and Campylo- bacter fennelliae (sp. nov.): Two new Campylobacter species associated with enteric disease in homosexual men. J Infect Dis 151:131-139, 1985
107. Tummuru MKR, Cover TL, Blaser MJ: Cloning and expression of a high molecular mass major antigen of Helicobacter pylori-evidence of linkage to cytotoxin production. Infect Immun 61:1799-1809, 1993
108. Ward JM, Fox JG, Anver MR, et al: Chronic active hepatitis and associated liver tumors in mice caused by a persistent bacterial infection with a novel Helicobacter species. J Natl Cancer Inst 861222-1227, 1994
109. Warren JR: Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1:1273, 1983
648 WINDSOR & OROURKE
110. Wegmann W, Aschwanden M, Schaub N, et al: Gastritis associated with Gastrospirillum
111. Yang HT, Li XT, Xu ZM, et al: "Helicobacter heilmannii" infection in a patient with hominis-a zoonosis? Schweiz Med Wochenschr 121245-254, 1991
gastric cancer. Dig Dis Sci 40:1013-1014, 1995
Address reprint requests to
Helen M. Windsor Helicobacter pylori Research Laboratory
Department of Microbiology University of Western Australia