Acta Zoologica (Stockh.), Vol. 66, No. 2, pp. 97-110, 198.5 0001-7272B.5 $3.00+ .OO Printed in Great Britain Pergamon Press Ltd. 0 1985 The Royal Swedish Academy of Sciences Balancer Fine Structure of the Pleurodeles Larva Harold Fox Department of Zoology, University College, London, U.K. (Received 22 November 1984) Abstract Fox, H. 1985. Balancer fine structure of the Pleurodeles larva. (Department of Zoology, University College, London, U.K.-Acta zool. (Stockh.) 66, 97-110. The paired balancers of larval Pleurodeles waltl, a urodelan species of the Amphibia, were investigated throughout their life span until final degeneration, using electron microscopy. The evidence from cellular ultrastructure illustrates the mucus-secreting function of these organs, and the outer epithelial cells actively synthesize muco-proteinaceous substance. An extensive granular endoplasmic reticulum and well developed Golgi complexes participate in the mucus manufacture. The balancer epidermis is also extensively innervated throughout by non-myelinated neurites, most of them without Schwann cells, a feature which argues strongly in favour of the organ also having a sensory function, whose nature has yet to be determined. Harold Fox, Department of Zoology, University College London, Cower Street, London WCI E 6BT, U. K. Introduction Balancers are a pair of slender rod-like struc- tures, each one extending outwards from the side of the head just behind the eye, of some species of amphibian larvae. They are illustrated among others by Fox (1955) in young larvae of Triturus cristatus carnifex and T. c. karelini, by Hambur- ger (1950) in Ambystoma maculatum from Harrison stages 34 to 45 (they have regressed in H stage 46), and by Gallien and Durocher (1957) in Pleurodeles waltl, as tiny buds at GD stage 28 which elongate and ultimately regress from GD stage 39 to 42, to disappear finally by G D stage 45. Balancers are restricted to some families of urodeles, the Ambystomidae, Salamandridae and Hynobiidae (Harrison 1925), but even here their presence is variable, for they are lacking in some species of Ambystoma (A. rnexicanum), are found only occasionally in A . tigrinum (Nicholas 1924) and are rudimentary in Salamandra. Balancers were probably first described in urodele larvae by Spallanzani (1784-1789) and later named as such by Clarke (1880), from their resemblance to balancers of dipterous insects (see Latta 1919). Clarke first suggested that they prop up the head until the future forelimbs serve this function. Balancers can lay claim to a number of names, for example: crochets de Kusconi (van Bambeke 1880), stalked suckers (Balfour 1881), Kieferfort- satz (Maurer 1888), Stutzorgan (Egert 1913), crochets d’adherence (Rusconi 1821-1823), Rus- conische Hakchen, Uncini di Rusconi, Balancier- Strang (see Harrison 1925), or balancing sticks, HeikBshijijB in the Japanese equivalent (Oyama 1930). Among the earlier workers who investigated balancers by light microscopy (see also Nicholas 1924, Harrison 1925, Murayama 1928), they were considered to be the homologues of external gills by Spallanzani (1784-1789), of the mandibular arch (Maurer 1888, Orr 1888) or of the hyoid arch (Kingsley 1899). Indeed, Spallanzani erroneously illustrated balancers having the appearance of tufted gills. Others homologized balancers with anuran embryonic adhesive organs (Rusconi 1821, van Bambeke 1880), as well as with the stalked suctorial disc of Triturus (Balfour 1881, Latta 1919). Furthermore, homol- ogy with the tentacular apparatus of adult caecil- ians and the tentacle of the larval Xenopus laevis was advocated by Cope (1889), Boulenger (foot- note in Leslie 1890), Bles (1905), Gadow (1909) and Cunningham (1912). By an ingeneous argu- ment Bles (1905) concluded that if balancers are modified mandibular external gills and they are 97
Transcript
Acta Zoologica (Stockh.), Vol. 66, No. 2, pp. 97-110, 198.5 0001-7272B.5 $3.00+ .OO Printed in Great Britain Pergamon Press Ltd.
0 1985 The Royal Swedish Academy of Sciences
Balancer Fine Structure of the Pleurodeles Larva
Harold Fox
Department of Zoology, University College, London, U.K. (Received 22 November 1984)
Abstract Fox, H. 1985. Balancer fine structure of the Pleurodeles larva. (Department of Zoology, University College, London, U.K.-Acta zool. (Stockh.) 66, 97-110.
The paired balancers of larval Pleurodeles waltl, a urodelan species of the Amphibia, were investigated throughout their life span until final degeneration, using electron microscopy. The evidence from cellular ultrastructure illustrates the mucus-secreting function of these organs, and the outer epithelial cells actively synthesize muco-proteinaceous substance. An extensive granular endoplasmic reticulum and well developed Golgi complexes participate in the mucus manufacture. The balancer epidermis is also extensively innervated throughout by non-myelinated neurites, most of them without Schwann cells, a feature which argues strongly in favour of the organ also having a sensory function, whose nature has yet to be determined.
Harold Fox, Department of Zoology, University College London, Cower Street, London WCI E 6BT, U. K .
Introduction
Balancers are a pair of slender rod-like struc- tures, each one extending outwards from the side of the head just behind the eye, of some species of amphibian larvae. They are illustrated among others by Fox (1955) in young larvae of Triturus cristatus carnifex and T. c. karelini, by Hambur- ger (1950) in Ambystoma maculatum from Harrison stages 34 to 45 (they have regressed in H stage 46), and by Gallien and Durocher (1957) in Pleurodeles waltl, as tiny buds at GD stage 28 which elongate and ultimately regress from GD stage 39 to 42, to disappear finally by GD stage 45.
Balancers are restricted to some families of urodeles, the Ambystomidae, Salamandridae and Hynobiidae (Harrison 1925), but even here their presence is variable, for they are lacking in some species of Ambystoma (A. rnexicanum), are found only occasionally in A . tigrinum (Nicholas 1924) and are rudimentary in Salamandra.
Balancers were probably first described in urodele larvae by Spallanzani (1784-1789) and later named as such by Clarke (1880), from their resemblance to balancers of dipterous insects (see Latta 1919). Clarke first suggested that they prop up the head until the future forelimbs serve this function.
Balancers can lay claim to a number of names, for example: crochets de Kusconi (van Bambeke 1880), stalked suckers (Balfour 1881), Kieferfort- satz (Maurer 1888), Stutzorgan (Egert 1913), crochets d’adherence (Rusconi 1821-1823), Rus- conische Hakchen, Uncini di Rusconi, Balancier- Strang (see Harrison 1925), or balancing sticks, HeikBshijijB in the Japanese equivalent (Oyama 1930).
Among the earlier workers who investigated balancers by light microscopy (see also Nicholas 1924, Harrison 1925, Murayama 1928), they were considered to be the homologues of external gills by Spallanzani (1784-1789), of the mandibular arch (Maurer 1888, Orr 1888) or of the hyoid arch (Kingsley 1899). Indeed, Spallanzani erroneously illustrated balancers having the appearance of tufted gills. Others homologized balancers with anuran embryonic adhesive organs (Rusconi 1821, van Bambeke 1880), as well as with the stalked suctorial disc of Triturus (Balfour 1881, Latta 1919). Furthermore, homol- ogy with the tentacular apparatus of adult caecil- ians and the tentacle of the larval Xenopus laevis was advocated by Cope (1889), Boulenger (foot- note in Leslie 1890), Bles (1905), Gadow (1909) and Cunningham (1912). By an ingeneous argu- ment Bles (1905) concluded that if balancers are modified mandibular external gills and they are
97
98 Harold Fox
also the homologues of Xenopus laevis larval tentacles (neither relationship substantiated, nevertheless), therefore Xenopus tentacles are derivations of external gills.
Balancers occasionally were considered to have a sensory function (Gadow 1909, Cunning- ham 1912), but no real evidence was offered: indeed, this function was discounted by Kingsley (1902), Egert (1913), Latta (1919) and Harrison (1925), though it was well known that the organ was innervated. Others believed the balancer to be respiratory (Spallanzani 17841789), or an adhesive organ (Rusconi 1821, van Bambeke 1880, Balfour 1881, Baird-in Cope 1889, Latta 1919), or to function mechanically as a prop (the most commonly held view) to balance the larva and support the body (Bell 1907), keep its head off the mud and elevate the pericardial region in the pond-lake habitat (Clarke 1880, Gadow 1909, Cunningham 1912, Egert 1913, Latta 1919, Harrison 1925, Oyama 1930). Cunningham (1912) also believed that the balancer was active- ly mobile, probably because of Orr's (1888) erroneous description of muscles from the ptery- goid musculature supposedly extending into it. A moveable balancer was denied by Harrison (1925), who also discredited the presence of a hollow core of bone beneath the balancer epidermis reaching the tip, previously claimed to be present by Latta (1919). The basement mem- brane (basement lamella) of the balancer of Diemyctylus viridescens was considered by Bell (1907) to be of collagen, and though its true nature was not understood by Harrison (1925), he correctly related it to a type of connective tissue.
Limited and inconclusive experimental work described transplantation and regeneration of the balancers (Bell 1907, Nicholas 1924, Kollros 1940). Essentially it appears that there is little if any regeneration and the life span of balancers is determined by intrinsic factors (Nakamura 1930).
It is likely that until now the only published description of the urodelan balancer by electron microscopy is that by Anderson and Kollros (1962), in Ambystoma opacum and A . jefferson- ianum. However, they did not deal with its innervation. The balancer was recently reviewed by Fox (1984, 1985a).
The following account of the ultrastructure of the larval balancer of Pleurodeles waltl, together with changes occurring during its subsequent development, differentiation and regression, provides further information and enlarges our understanding of this somewhat enigmatic organ. Among other things the results confirm its func- tion as a mucus-manufacturing and secreting structure, and in addition demonstrate its exten- sive epidermal innervation, predominantly by
naked neurites, an arrangement which supports a heretofore barely considered view of a sensory function, as yet to be specified.
Material and Methods
Pleurodeles waltl larvae at different stages of the Gal- lien and Durocher (1957) series, maintained in tap water in aquaria at 18-2loC, were anaesthetised in 1:10OOO MS 222 and the paired balancers removed and fixed ice-cold in a combined glutaraldehyde-osmic acid fixative (Hirsch and Fedorko 1968). For dehydration, Araldite embedding, sectioning (by use of a diamond knife) and staining sections, standard methods were used (see Fox 1974, 1983).
The balancers examined by electron microscopy were of GD stages 32(1) (larva 7 mm long with 3 pairs of external gills), 33b(l) (larva 8 mm long and the fore-limb bud visible), 34(1) (larva 9 mm long; the first hatching specimens), 36(3) (larva 11 mm long and the fore-limb buds cylindrical in shape), 37(1) (larva 11.8 mm long and the fore-limb palette-shaped), 38(1) (larva 12 mm long; first feeding stage), 39(3) (larva with 2 digits to the fore-limb), 40(2) (larva with elongate and branching gill filaments), 41(2) (larva with 3 digits to the fore-limbs). The number of specimens used is in parentheses. In this particular population of Pleurodeles larvae, balancers in G D stage 41 were either a stump or, if of normal length, appeared degenerate. Balancers were absent in GD stage 42 specimens.
Skin of Pleurodeles larvae from the head at the base of the balancer (stage 37), fore-limb bud (stage 40) and the base of the tail (stages 3 5 4 ) was also examined for comparison with that of the balancer.
Silver-grey transverse sections of balancers of the above stages, including a longitudinal series at GD stage 39 and transverse sections of the skin of the other regions of the body, were examined under a JEOL JEM 100 CX 11 electron microscope. A young larva of Pleurodeles waltl 10 mm long and probably in the mid 30s stage, used for light microscopy, was sectioned transversely at 10 pm and stained by Ehrlich's haemat- oxylin and aqueous eosin. Sections showing the balancer were examined and photographed with a Zeiss photo- microscope for gross structure and topography.
Results
At stage 32 the balancer of Pleurodeles is about 0.5 mm long and typically the epidermis is 2-3 cell layers thick. Epithelial cells are up to 21 pm long and 15 pm deep and cuboidal in shape. Adjacent cells join by tight junctions and by desmosomes that likewise join other epithelial cells throughout the epidermis. Outer cells have a large irregularly-shaped nucleus with a promin- ent nucleolus. The cytoplasm is richly supplied with polysomes and an extensive granular endo-
Balancer of Pleurodeles 99
plasmic reticulum (RER), and there are tonofila- ments and mitochondria (Fig. 1). Well developed Golgi complexes are present alongside the nuc- leus, a feature of outer cells throughout the series. Sometimes the Golgi complex has a semi- circular profile of 4-5 stacks of somewhat dense cisternae, an arrangement seen in other balan- cers. At the terminals of the cisternae there are numerous dense vesicles, some clearly similar in size and appearance to the large number of secretory granules in the rest of the cell (see Figs. 5 , 8).
The free external surface of the skin of the balancer is ridged or, in some instances, apparently microvillous, and lined by a mucus layer up to 5 p,m thick: the layer is equally prominent in other stages (Fig. 2). Mucus ves- icles in various stages of discharge line the exter- nal surface and supply the mucus layer. Mucus originates from secretory granules, probably of glycoprotein, widely distributed in cells of the outer layer (Fig. 1) but absea from those of the basal layer. Granules average about 200-250 nm in diameter, though larger less-dense granules about 550 nm in diameter occur. Large yolk bodies and liposomes are present.
Ciliary cells are occasionally seen at the skin surface; ciliary roots are seen located near a few mucus vesicles of the cell. Ciliary cells appear to be mitochondria-rich, a property in addition to their cilia that distinguishes them from the other epithelial cells.
Basal epithelial cells are generally similar in appearance throughout most of balancer life (Fig. 3). There is a high content of polysomes, an RER, albeit far less substantial in amount than in outer cells, but there are more mitochondria. Tonofilaments occur and their amount increases in older balancers. There are no Figures of Eberth or hemidesmosomes in basal cells at any stage of balancer life, though sometimes sugges- tions of incipient hemidesmosomes are recogniz- able. In most basal cells the Golgi complexes are not as conspicuous as in outer cells. Basal cells are bounded by an adepidermal membrane (and space) and below randomly oriented collagen fibrils make up an incipient basement lamella.
Between the epithelial cells of the epidermis there are neurites containing small mitochondria, neurotubules, vesicles and glycogen, features recognized in neurites of other stages. Frequently nerves are enveloped by adjacent epithelial cells.
At stage 33 the balancer outer epithelial cells have an extensive RER which, as in other stages, often appears whorled around the nucleus to fill much of the cell (see Fig. 4). Prominent Golgi complexes are active in granule formation. Secre- tory granules of varied size and electron-density appear more plentiful than in the previous stage,
the less-dense granules situated nearer the outer surface. Pigment granules and larger dense secre- tory granules of similar size are often difficult to categorize. Below the outer cell surface extensive roundish muco-granular areas, 5 p,m in diameter or even larger and often containing pigment, are occasionally recognizable in this and other stages.
Vestigial cilia occur at the surface; they are never found in balancers of older stages. Large lipid droplets are present in cells of different layers; indeed they are still found in stages 36 and 37.
Well developed unmyelinated neurites (some- times found in groups of 3-5 in the basal layer) are present, frequently partly or wholly sur- rounded by neighbouring epithelial cells.
The basement lamella is less extensive than in the previous stage, demonstrating its variability in size and degree of collagenolytic differenti- ation in different larval stages and in different regions of the balancer (vide infru). Below the collagen layer there are fibroblasts with a high content of RER.
At stage 34 the balancerlis about 1 mm long and about 0.1 mm thick: the epidermis is up to 15 pm deep. Secretory granules of outer cells appear less electron-dense and often clearly membrane-bound. Multivesicular bodies and other rounded lucent vesicles are present and mitochondria have well developed cristae. Near the surface the content of tonofilaments has increased compared with previous stages, and pigment granules can occur in groups of 20 or more in this region. Basal cells have increased their content of RER, there is some development of an SER and multivesicular bodies, pigment and an occasional centriole are seen. Autophag- ous vacuoles uncommonly occur in cells of different layers. Neurites are widely distributed in the epidermis.
At stage 36, possibly around the peak period of mucus secretion by the balancer, the RER and Golgi complexes of outer cells are prominent (Fig. 4). Granule origin from the Golgi complex was particularly noticeable in an example at this stage (Fig. 5). Tonofilaments, especially those near the surface, frequently form a network between secretory granules. Mucus vesicles can occur to a depth of 2-3 p,m below the surface, which implies a high level of mucus secretory activity. Adjacent dense secretory material in the surface mucus vesicles join one another by fine filamentous strands. Within the cell, mucus ves- icles are of varied appearance and their dense vacuolated substance frequently merges into ambient lucent areas. Other secretory granules appear to be in the process of dissolving in the cytoplasm. Presumably the product diffuses to
100 Harold Fox
Balancer of Pleurodeles 101
Fig. 4. Outer epithelial cell of a balancer near the tip, of a stage 36larva, showing the whorled arrangement of the RER and the packed secretory granules that supply the mucus of the vesicles at the surface. Golgi complexes are located at the periphery of the RER. Fig. 5. Go@ complex in an outer balancer cell at stage 36, showing the likely derivation of the secretory granules from the Golgi cisternae.
the surface to join other mucus ready for dis- charge.
At the tip of the balancer the basement lamella collagen is only poorly developed: in contrast in
Fig. 1. Outer region of a surface epithelial cell of a balancer at stage 32. There is a high content of RER and secretory granules, and mucus granules at the surface. Golgi complexes are located on either side of the irregularly-shaped nucleus. Fig. 2. An example of the thick mucus layer in a stage 32 larva, found at the surface of the balancer. Fig. 3. Basal epithelial cells of the balancer of a stage 36 larva. There is a substantial content of RER and polysomes and numerous mitochondria. The matrix includes tonofilaments. The basement lamella is well developed, though variable in thickness in different regions of the balancer, and the collagen fibrils are oriented at random. Note the neurites between the cells.
another example at stage 36, further proximally the balancer showed more substantial develop- ment.
At the centre of the balancer a bundle of unmyelinated nerves (dimensions in one example at this stage 5.5 pm long by 2.7 pm wide in section) enveloped by Schwann cells doubtless supplies neurites distributed in the epidermis, often near the surface and extending to the tip. The central nerve may contain up to 25 separate nerve fibres, the largest about 1.2 pm in diam- eter. A similar nerve bundle occurs in balan- cers of other stages. A well developed capillary lies adjacent to the central nerve.
Balancer epithelial cells of a stage 37 larva are generally little changed from those of. the pre- vious stage. However, occasionally large inner epithelial cells are seen to contain numerous secretory granules. These cells are always bound- ed basally by extensions of basal cellular tissue
102 Harold Fox
and doubtless they are outer cells, part of whose external surface is overlain by an adjacent outer epithelial cell. As a general rule only outer epithelial cells manufacture secretory granules for discharge at the surface.
In the well innervated epidermis, in one inst- ance in the basal layer, a neurite, about 0.6 pm in diameter, was found enveloped by 1% turns of a Schwann cell sheath, about 0.06 pm thick (Fig.
Compared with previous stages, outer balancer cells of a stage 38 specimen have far fewer dense secretory granules, though a. RER and Golgi complex are still well developed, and cells of both layers of the epidermis have a higher con- tent of tonofilaments (Figs. 7, 8). There is clearly a reduction in balancer mucus-secretory activity. Small cytolysomes occur in outer cells, indeed they are sometimes found in younger balancers. Inner cells, however, have extensive areas of degeneration, that may include mucoid granules and their degraded products, large and small cytolysomes, pigment, lipid and vacuoles (Fig. 9). Nuclei are not pycnotic.
In the basal region a lengthwise section of a neurite revealed elongate neurotubules, and other neurites sectioned transversely, in groups of up to 12, in addition contained vesicles, mitochondria and glycogen granules (Fig. 10).
A balancer at stage 39 sectioned horizontally had the RER and Golgi complexes of outer epithelial cells reduced in amount compared with the previous stage; likewise the number of secre-
6).
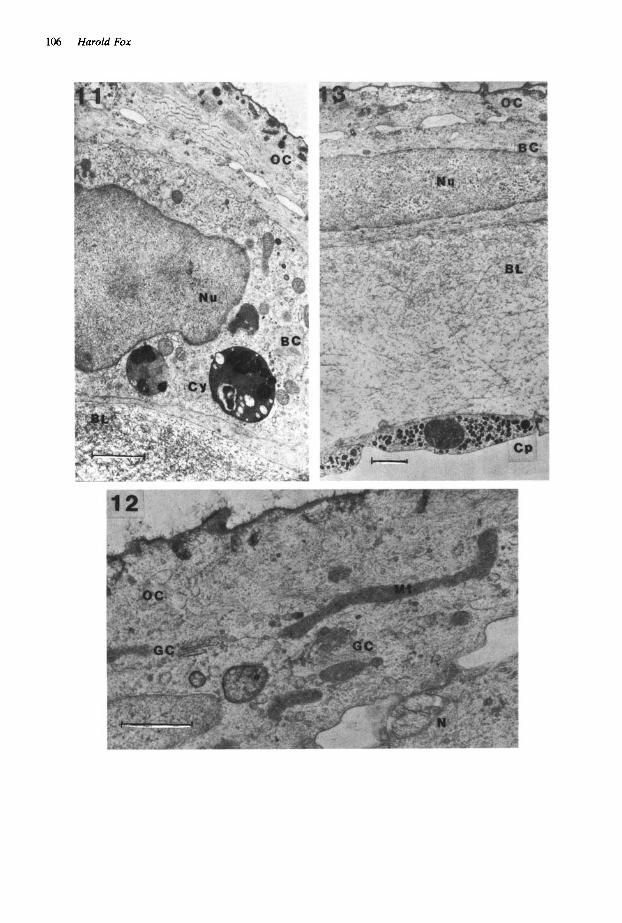
tory granules and mitochondria was less. Few mucus vesicles line the outer surface. Cells of both layers contain cytolysomes 0.9-3.6 pm in diameter, of rounded membrane-bound lucent bodies, probably of h .ghly degenerate substance. Other cellular organelle material shows a greater or lesser degree of degeneration. Cytolysomes in the inner cells are *more plentiful, larger and substantial. Balancer cells of the transversely sectioned series at stage 39 were more flattened and fibrous than those of the horizontal senes (Fig. 11). Hemispherical shaped Golgi complexes and associated small dense granules still occur in outer cells. Large cytolysomes in the basal cells are often electron-dense and include pigment (Fig. 11). Balancers of both stage 39 examples are well innervated.
The more prematurely degenerate balancer of two stage 40 specimens of Pleurodeles investi- gated may have belonged to an unhealthy larva, though the donor seemed normal in outward appearance. Flattened fibrous epithelial cells of the lesser degenerate balancer contained few mucus vesicles at the free surface of the outer cells, though a layer of mucus is still present. Cellular organelles are further reduced in num- ber compared with those in the previous stage. Nevertheless, small Golgi complexes are still associated with a lesser number of dense gran- ules. Large and small liposomes, pigment gran- ules and degenerate organelle tissue occur in cytolysomes, seen especially within inner cells. Some dense granules in the outer cells presum-
Fig. 6. Neurite enveloped by a Schwann cell sheath, in the basal region of the epidermis of a balancer at stage 37. the 1% turns of the wrapping Schwann cell sheath are clearly recognizable.
Balancer of Pleurodeles 103
ably are glycoproteinaceous; smaller granules could well be primary lysosomes, enzymic in nature and involved in tissue autolysis. More highly degenerate stage 40 balancer cells are flattened, fibrous and ragged in appearance, and some outer cells seem to have sloughed. Nuclei are pycnotic, in contrast to those of the other stage 40 balancers that are normal in appearance. Most of the cell organelles are highly degenerate, the RER lumina are swollen and the Golgi complexes reduced in size.
Some larval Pleurodeles at stage 41 had balan- cers of practically normal size, though showing features of degeneration, others retained a balan- cer merely as a stump on one side of the head, and the rest had lost their balancers. Balancers of nearly normal length had highly fibrous flattened epithelial cells, roughly intermediate in appear- ance between those of the balancers of the two preceding stage 40 specimens. Nevertheless, elongate non-degenerate mitochondria in outer cells demonstrate the uneven pattern of organelle degeneration in balancer cells (Fig. 12). Small neurites occur between the cell layers as in the case of the preceding stage. There is an extensive unchanged basement lamella and a central capil- lary has endothelial walls containing numerous dense granules (Fig. 13).
Where the balancer stump merges into the head, its degenerate fibrous epithelial cells are generally similar in appearance to those of the other stage 41 specimen. A large non-degenerate nerve was seen in the sub-epidermal region of the stump, and there were occasional neurites in the epidermis.
Fig. 7. Outer epithelial cell of a balancer at stage 38. The whorled RER and the Golgi complex are well represented, though at this time there are fewer secre- tory granules than in balancers of previous stages. Nevertheless, mucus vesicles still line the free surface and there is a substantial mucus layer. Note the neurite below the outer cell partially enveloped by it.
Fig. 8. Well developed Golgi complex in an outer epithelial cell of a balancer at stage 38, showing associated granules forming. It is possible that there is a switch- over from secretory granule formation to that of primary lysosomes around this stage, when cellular autolysis commences.
104 Harold Fox
Fig. 9. Balancer epidermis in a stage 38 larva (see also Fig. 7), showing a large area of degeneration in the basal epithelial cell. A clear difference in RER content between outer and basal cells is apparent, and likewise there are secretory granules in outer cells in contrast to their absence in basal cells.
Skin from Other Regions of the Larva of Pleurodeles
In general the ultrastructure of the skin of differ- ent regions of a Pleurodeles larva, such as that of the head near the balancer, the fore-limbs and the base of the tail, for example, is similar. Roberts and Hayes (1983) likewise described head skin of the Xenoplrs larva to be similar to that of other regions of the body. However,-in Pleurodeles clear differences are apparent be- tween skin of the larval balancer and that of the adjacent head, as well as with skin from other
regions of the body. Outer epithelial cells for much of balancer life (until about stage 38) have an extremely high content of RER and secrete a copious mucus (see Figs. 2, 7, 9). Figures of Eberth and hemidesmosomes do not occur in the basal cells. The epidermis is extensively inner- vated (see Fig. 10). There are no lateral line organs, nor is the basement lamella collagen arranged orthogonally. In contrast, outer epidermal cells of the head skin possess only a modest RER and light content of polysomes, with far less mucus secretion than in balancer cells (Fig. 14), and the basal cells include well
Balancer of Pleurodeles 105
. Fig. 10. Basal region of a balancer epidermis, at stage 38, showing 12 neurites and/or their branches; an example of the extensive innervation in the balancer epidermis.
developed Figures of Eberth and hemidesmo- somes. The basement lamella shows a pro- nounced orthogonality (Fig. 15). Furthermore, lateral line organs occur and the epidermis appears to be less well innervated than in the balancer. The presence of striated muscles below the skin of the head is a useful criterion for distinguishing balancer and head skin, for the balancer does not contain them.
Discussion
The larval urodelan balancers have long been known to secrete mucus, especially at their ter- minal region. Viewed by electron microscopy, surface balancer cells of Pleurodeles and also those of Ambystoma (Anderson and Kollros 1962) include an RER that seems to be more extensive than that found in the body epidermal cells of these species, and of other larval urodeles (Kelly 1966, Warburg and Lewinson 1977, Greven 1980), anurans (Fox 1972, 1974, 1977) and caecilians (Welsch and Storch 1973, Fox un- pub].). Careful examination of the epidermis of Pleurodeles larvae of the head near the balancer, the base of the tail and of the fore-limb confirms this view. The arrangement of the RER in the outer balancer cells is similar to that in develop- ing mucocytes of mucus glands in the Salamandra terdigitata larva (Delfino et al. 1982). The extent and arrangement of the RER and the enormous number of secretory granules in the balancer cells
reflects a highly active protein synthesis. Presum- ably the resulting protein is transported to the well developed Golgi saccules (see Perry and Waddington 1966). where glycosilation takes place (Alberts et al. 1983). Dense granules orig- inating at the terminals of the Golgi cisternae probably contain glycoprotein, the source of mucus in the surface mucus vesicles. Basal cells have few if any secretory granules and presum- ably they synthesize little glycoprotein, a conclu- sion reinforced by their lesser content of RER and reduced Golgi complexes compared with these components in outer cells.
The basement lamella (basement membrane of Anderson and Kollros 1962) of the balancer reaches a maximum width of 4-6 pm at stage 37. Collagen fibrils orientate randomly and there is no orthogonal configuration (see Weiss and Fer- ris 1954), though in transverse section of older stages there is a slight preferential orientation of some collagen fibrils that extend parallel to the adepidermal membrane.
Degeneration of the balancer is by autolysis, and during later stages of balancer life at first small and then gradually larger areas of cytoplas- mic degeneration-the cytolysomes-appear, paripassu with a high content of tonofilaments in cells of both layers. At this period the number of secretory granules and level of mucus secretion are reduced. Yet small dense granules still origin- ate at the terminals of the Golgi cisternae. In prehatching stages of Xenopus laevis, the Golgi complexes of hatching gland cells give origin to granules containing polysaccharides. At hatching
106 Harold Fox
Balancer of Pleurodeles 107
Fig. 14. Outer epithelial cells of head skin near the base of the balancer, in a stage 37 larva. These cells should be compared with similar outer cells of a balancer at stages 32 (Fig. l ) , 36 (Fig. 4) and 38 (Figs. 7, 9), which emphasise the mucus- secreting activity of the balancer. Fig. 15. Basal cell of head skin near the base of the balancer, in a stage 37 larva, showing the presence of Figures of Eberth and hemidesmosomes, and below a basement lamella of orthogonally arranged collagen fibrils. The fine structure should be compared with the basal cells of the balancers at stage 34 (Fig. 3) and stage 41 (Fig. 13), where the head cell features are absent.
Fig. 11. Flattened balancer cells at stage 39. A modest amount of RER, and a Golgi complex and surface mucus vesicles are recognized in the outer cell. Large dense lysosomes occur within the inner cell. The basement lamella is extensive, but there is no orthogonal arrangement of the collagen fibrils. Fig. 12. A flattened fibrous outer epithelial cell of a balancer at stage 41. A reduced Golgi complex may originate primary lysosomes, for there are few secretory granules or surface mucus vesicles. Presumably there is little active mucus secretion. Mitochondria can be well formed and elongate. Neurites are present between the cell layers. Fig. 13. Similar flattened fibrous epithelial cells of a balancer, at stage 41, as in Fig. 12. There is an extensive basement lamella, about 5 pm thick, with no orthogonality. The endothelial cells of the underlying capillary include numerous small dense granules.
108 Harold Fox
they produce acid phosphatase-rich granules, probably primary lysosomes, involved in auto- lytic degeneration (Yoshizaki and Katagiri 1975), for it is well known that primary lysosomes arise from Golgi cisternae (Fox 1974). It seems likely, therefore, that similar events occur in balancer cells; the Golgi complex first budding off secre- tory granules during the balancer mucus- secretory phase, and thereafter primary lyso- somes that elicit autolysis during the degener- ation phase.
Autotomy of the balancer could be the result of a high degree of autolysis in a specific region (perhaps proximal) of the balancer, the so de- graded organ is then either forceably or passively discarded. Shrinkage appears to be an overall response to widespread autolytic phenomena. Cessation of the blood supply or curtailment of balancer innervation do not appear to be of signif- icance in influencing its degeneration.
Basement lamella collagen of the larval anuran tail is phagocytosed by mesenchymal mac- rophages utilising lysosomal enzymes (Kemp 1963, Gona 1969, Fox 1972, 1981). However, there is no macrophagic invasion of balancer col- lagen during its degeneration, and the basement lamella remains unchanged and well developed, at least until the terminal stages of balancer life. Presumably the integrity of the collagen is vit- iated owing to the overall cellular degeneration of the balancer, and collagen breakdown is an in- direct result of these events.
From the evidence of ultrastructure it is clear that the balancer is a mucus-secreting organ, similar t o the cement glands of Xenopus embryos, whose cells likewise have an extensive RER (Perry and Waddington 1966) and secrete a glycoproteinaceous mucus (Ling and Lyerla 1976). The balancer secretion may serve for: adherence of the larva to flora or other material in the aquatic habitat, as originally described by Rusconi (1821-1823); or assist its balance when the balancers (perhaps) prop up the head; or in antibiosis. The balancer is not actively mobile for there is no musculature along its length, though this does not preclude some indirect degree of mobility if it were activated near its base by later- al head muscles.
The balancer epidermis of Pleurodeles is exten- sively innervated by branches from a centrally located unmyelinated nerve (enveloped by Schwann cells), a branch of the trigeminal nerve of the gasserian ganglion (Harrison 1925). Neurites without Schwann cell sheaths are distrib- uted in the basal layer in groups of up to 12 branches, or singly between cells of the same or different layers. In one rare instance a nerve with a Schwann cell sheath was found in the basal layer, probably the first report of such a feature
in a larval amphibian epidermis. Sheathed nerves occur in the basal layer of the epidermis of adult Rana temporaria, and in the nuptial pads of the male breeding Rana, where nerves meet the Mer- kel cells (Whitear 1974, 1983).
Skin of the body and tail of embryonic and young larval Xenopus is innervated by naked sen- sory terminals of the Rohon-Beard cells. The nerves detect transient touch stimuli (Roberts and Hayes 1977, Clarke et al. 1984). Head skin is innervated by branches of the maxillary and ophthalmic profundus nerves of the trigeminal ganglion. Skin of this region is similar in structure to that from other regions of the body, and so is the fine structure of their neurones. Head skin neurites include: Type 1 tonically firing move- ment detectors with homolateral innervation; Type 2 quickly adapting rapid transient detectors that cross the mid-line to innervate both sides of the head. Most neurites occur singly in the skin, and the approximately 20% remainder occur in groups of up to three, usually in the basal epidermal layer (Hayes and Roberts 1983).
The nature of the innervation of the balancer of Pleurodeles (or for that matter of any urodele larva) is not understood. Certainly, deformation of the neurite membrane could provide a stimu- lus for nerve activity (Whitear 1983) and a mechano-sensory function would seem likely, though a chemo-sensory function cannot be ruled out. Balancers of Pleurodeles and the tentacles of the adult caecilian Zchthyophis kohtaoensis are extensively innervated by naked neurites (Fox 1985b). Tentacles of the larval Xenopus laevis have epidermal naked neurites, but in addition Merkel cells in synaptic association with them (Fox and Whitear 1978, Ovalle 1979, Tachibana et al. 1980). Tentacles of Xenopus are probably mechano-sensory in function (Ovalle 1979), for their nerves respond to mechansal pressure. There is also a twitch withdrawal response, elic- ited by the tentacular musculature at its base, after crushing the tentacle tip (Cannone and Kel- ly 1977).
In newly hatched Xenopus laevis larvae, Roberts and Blight (1975) showed that stimul- ation of the cement gland by touch inhibits swim- ming, and the sensory unmyelinated neurites (1 pm or less in diameter) in the skin of the gland are responsible for this effect. The result explains why tadpoles cease to move when the cement gland makes contact with a firm object. Again, when a tadpole hangs by a mucus thread the cement gland receptors probably fire impulses at a low rate, which through central connections inhibit the degree of activity of the tadpole.
A high level of mucus secretion and an exten- sive innervation of the balancer of Pleurodeles are similar to the morpho-physiological arrange-
Balancer of Pleurodeles 109
ment of the adhesive cement gland of Xenopus. Whether the balancer functional activity of urodele larvae is similar poses problems for future experimentation.
Acknowledgements
Thanks are due to Jonathan Slack for generously supplying the Pleurodeles larvae. Technical assistance was provided by Brian Pirie and Roy Mahoney and by Mary Whitear, who also advised me during the preparation of this work.
Abbreviations in the Figures
AM BC B L CP CY FE GC HD IJ LP MV Mu M I N Nu oc OG
PG RER scs SG T
adepidermal membrane basal cell of the epidermis basement lamella cap i 11 a r y cytolysome Figure of Eberth Golgi complex hemidesmosomes intercellular junction lipid droplet mucus vesicle mucus mitochondrion neurite nucleus outer cell of the epidermis orthogonally arranged collagen fibrils of the
Scale marks on all the figures represent 1 pm, except for those on Figs. 5 and 6, which are 0.5 pm, and on Fig. 9, which is 2 pm.
References
Alberts, k., Bray, D., Lewis, J., RafJ M., Roberts, K., Watson, J . D. 1982. Molecular Biology of the Cell, Garland, New York.
Anderson, E., Kollros, J . J . 1962. The ultrastructure and development of balancers in Ambystoma embryos with special reference to the basement membrane. J. Ultrastruct. Res. 6, 35-56.
Baird, S. F. 1851. Quoted in Cope, E . D. (1889) pp. 59-60.
Balfour, F. M. 1881. A Treatise on Comparative Embryology, Vol. 2, London.
van Bambeke, Ch. 1880. Nouvelles recherches sur I’embryologie des batraciens. Archs Biol., Paris 1,
Bell, E. T. 1907. On regeneration and transplantation of the balancers of embryos of Diemyctylus (with a note on the external gills). Anat. Anz . 31,283-291.
Bles, E. J . 1905. The life history of Xenopus laevis. Trans. R. SOC. Edinb. 41, 78%821.
Boulenger, G. A. 1890. Footnote in Leslie, J. M. (1890). Cannone, A. J . , Kelly, P. J . 1977. The tentacles of
Xenopus laevis-Evidence for a mechanoreceptive role. S. Afr. med. J . 52, 407.
Clarke, S. F. 1880. The development of Amblystoma punctatum. Part 1. External. Stud. biol. Lab. Johns Hopkins Univ. 2, 105-125.
Clarke, J . D. W., Hayes, B. P., Hunt, J . P., Roberts, A. 1984. Sensory physiology, anatomy and immuno- histochemistry of Rohon-Beard neurones in embryos of Xenopus laevis. J. Physiol., Lond. 348,
Cope, E. D. 1889. The Batrachia of North America. Bull. U.S. natn. Hist. Mus. 34, 1-515.
Cunningham, J. T. 1912. Reptiles, Amphibians, Fishes and Lower Chordates. Animal Life and Evolution- ary Natural History. London.
Delfino, G., Brizzi, R., Calloni, C. 1982. Development of cutaneous glands in Salamandra terdigitata (LacCptde, 1788) (Amphibia; Urodela); findings by light and electron microscopy. Z. mikrosk. -anat. Forsch. 96, 948-971.
Egert, F. 1913. Die Kopfanhange der Amphibien. Zool. Anz . 42, 280-288.
Fox, H. 1955. Early development of two species of the salamander Triturus cristatus. Copeia 1930, (No. 2),
Fox, H. 1972. Tissue degeneration: an electron micros- copic study of the tail skin of Rana temporaria during metamorphosis. Archs Biol., Liege 83,373- 394.
Fox, H. 1974. The epidermis and its degeneration in the larval tail and adult body of Rana temporaria and Xenopus laevis (Amphibia:Anura). J . Zool. Lond.
Fox, H. 1977. The anuran tadpole skin: changes occur- ring in it during metamorphosis and some compari- sons with that of the adult. In Comparative Biology of Skin (Ed. R. I . C. Spearman), Symp. zool. SOC. Lond. 39, 269-289.
Fox, H. 1981. Cytological and morphological changes during amphibian metamorphosis. In Metamorph- osis A Problem in Developmental Biology (Eds. L. I . Gilbert and E. Frieden), pp. 327-362, Plenum, New York.
Fox, H. 1983. The skin of Ichthyophis (Amphibia: Caecilia): an ultrastructural study. J . Zool. Lond. 199, 223-248.
Fox, H. 1984. Amphibian Morphogenesis, Humana Press, New Jersey.
Fox, H. 1985a. The skin of the amphibia, Springer, in press.
Fox, H. 1985b. The tentacles of khthyophis (Amphibia: Caecilia) with special reference to the skin. J . Zool. Lond. 205, 223-234.
Fox, H., Whitear, M. 1978. Observations on Merkel cells in amphibians. Biologie cellulaire 32,223-232.
305-380.
5 1 1-525.
131-133.
174, 217-235.
110 Harold Fox
Gadow, H. 1909. Amphibia and Reptiles (Eds. S . F . Harmer, A. E. Shipley), Vol. 8, Macmillan, London.
Gallien, L., Durocher, M. 1957. Table chronologique du developpement chez Pleurodeles waltlii Michah. Bull. biol. Fr. Belg. 91, 97-114.
Gona, A. G. 1969. Light and electron microscopic study on thyroxine-induced in vitro resorption of the tadpole tail fin. Z . Zellforsch. 95, 483-494.
Greven, H. 1980. Ultrastructural investigations of the epidermis and the gill epithelium in the intrauterine larvae of Salamandra salamandra (L.) (Amphi- bia:Urodela). Z . mikrosk.-anat. Forsch. 94, 196- 208.
Hamburger, V. 1950. A Manual of Experimental Embryology, University of Chicago Press, Illinois.
Harrison, R. G. 1925. The development of the balancer in Amblystoma, studied by the method of trans- plantation and in relation to the connective-tissue problem. J . exp. Zool. 41, 349-427.
Hayes, B. P., Roberts, W. 1983. The anatomy of two functional types of mechanoreceptive free nerve endings in the head skin of Xenopus embryos. Proc.
Hirsch, J., Fedorko, M. E. 1968. Glutaraldehyde and osmium tetroxide combined fixation method. J . Cell Biol. 38, 615-627.
Kelly, D. E. 1966. Fine structure of desmosomes, hemidesmosomes and an adepidermal globular layer in developing newt epidermis. J . Cell Biol. 28,
Kemp, N. E. 1963. Metamorphic changes of dermis in skin of frog larvae exposed to thyroxine. Devl Biol. 7, 244-254.
Kingsley, J . S. 1899. Text Book of Vertebrate Zoology, George Bell, London.
Kollros, J . J . 1940. The disappearance of the balancer in Amblystoma larvae. J . exp. Zool. 85, 33-52.
Lana, J . S. 1919. The morphology of the so-called balancers in certain species of Amblystoma. Anat. Rec. 17, 63-71.
Leslie, J . M. 1890. Notes on the habits and oviposition of Xenopus laevis. Proc. zool. SOC. Lond. 1890,69-71.
Ling, B. Y., Lyerla, T. A. 1976. Acid phosphatase activity in the development of the cement gland in Xenopus laevis. J . exp. Zool. 195, 191-197.
Maurer, F. 1888. Die Kiemen und ihre Gefasse bei anuren und urodelen Amphibien, und die Umbil- dung der beiden ersten Arterienbogen bei Tele- ostiern. Morph. Jb,. 14, 17S222.
Murayama, T. 1928. Uber die Balanzierstrange bei Hynobiuslarven. Folia anat. jap. 6, 37.5-387.
Nakamura, 0. 1938. Studies on amphibian metamorph- osis. V. Experiments on the disappearance of the balancer in urodele, Triturus pyrrhogaster (Boie).
R. SOC. B 218, 61-76.
51-72.
Bot. ZOO^. 6, 1051-1055. Nicholas, J . S. 1924. The development of the balancer in
Amblystoma tigrinum. Anat. Rec. 28, 317-329. Orr, H. 1888. Note on the development of amphibians,
chiefly concerning the central nervous system; with additional observations on the hypophysis, mouth and the appendages and skeleton of the head. Q. J . microsc. Sci. 29, 295-324.
Ovalle, W . K. 1979. Neurite complexes with Merkel cells in larval tentacles of Xenopus laevis. Cell Tissue Res. 204, 233-241.
Oyama, J . 1930. Balancer in Diemictylus pyrrhogaster and in Hynobius nebulosus. Copeia 1930, (No. 173) 103-106.
Perry, M. M., Waddington, C. H. 1966. The ultrastruc- ture of the cement gland in Xenopus laevis. J. Cell Sci. 1, 19S200.
Roberts, A., Blight, A. 1975. Anatomy, physiology and behavioural role of sensory nerve endings in the cement gland of embryonic Xenopus. Proc. R. SOC.
Roberts, A, , Hayes, B. P. 1977. The anatomy and function of ‘free’ nerve endings in an amphibian skin sensory system. Proc. R. SOC. B 196,415-429.
Rusconi, M. 1821-1823. Observations on the natural history and structure of the aquatic salamander, and on the development of the larva of these animals from the egg, up to the perfect animal (translation by D. E.). In Edinb. Phil. J . 9, 107-127 (1823).
Spallanzani, A. L. 1784-1789. Dissertation relative to the natural history of animals and vegetables (trans- lation from the Italian), 2, London.
Tachibana, T., Sakakura, y., Nawa, T. 1980. Merkel cell differentiation in the developing tentacles of Xenopus laevis. Acta anat. nippon. 55, 588-599.
Warburg, M . R., Lewinson, D. 1977. Ultrastructure of epidermis of Salamandra salamandra followed throughout ontogenesis. Cell Tissue Res. 181,36!+ 393.
Weiss, P., Ferris, W. 1954. Electronmicroscopic study of the texture of the basement membrane of larval amphibian skin. Proc. natn. Acad. Sci. U.S.A. 40, 528-540.
Welsch, U . , Storch, V. 1973. Die Feinstruktur verhorn- ter und nichtverhornter ektodermaler Epithelien und der Hautdrusen embyonaler und adulter Gym- nophionen. Zool. J . Anat. 90, 323-342.
Whitear, M. 1974. The nerves in frog skin. J . Zool. Lond. 172,503-529.
Whitear, M. 1983. The question of free nerve endings in the epidermis of lower vertebrates. Acta biol. hung. 34, 303-319.
Yoshizaki, N., Katagiri, C. 1975. Cellular basis for the production and secretion of the hatching gland enzyme by frog embryo. J . exp. Zool. 192,203-212.