Binding of the Bacteriophage T4 Transcriptional Activator, MotA, to T4 Middle Promoter DNA: Evidence for both Major and Minor Groove Contacts Mridula Sharma, Philip Marshall and Deborah M. Hinton* Laboratory of Molecular and Cellular Biology, National Institute of Diabetes and Digestive and Kidney Diseases National Institutes of Health Bethesda, MD 20892, USA During infection, the bacteriophage T4 transcriptional activator MotA, the co-activator AsiA, and host RNA polymerase are needed to transcribe from T4 middle promoters. Middle promoters contain a 10 region recognized by the s 70 subunit of RNA polymerase and a MotA box cen- tered at 30 that is bound by MotA. We have investigated how the loss or modification of base determinants within the MotA box sequence 5 0 TTTGCTTTA3 0 (positions 34 to 26 of a middle promoter) affects MotA function. Gel retardation assays with mutant MotA boxes are con- sistent with the idea that MotA uses minor groove contacts upstream and major groove contacts downstream of the center GC, and does not require any specific base feature at the C G base-pair at position 30. In particular, the 5-methyl residue on the thymine residue at position 29, a major groove contact, contributes to MotA binding, while converting the T A at 32 to a C I base-pair, a change that affects the major but nor the minor groove, yields a MotA box that is similar to wild-type. However, methylation interference analyses indicate that neither the binding of MotA nor the binding of polymerase/MotA/AsiA to the middle pro- moter P uvsX is inhibited by premethylation of guanine and adenine residues, suggesting that binding does not require minor groove contact with any specific T A base-pair. Using gel retardation analyses, we calcu- late an apparent dissociation constant of 130 nM for MotA binding to the wild-type MotA box. Previous work has shown that the N-terminal region of MotA is needed for an interaction between MotA and s 70 . We suggest that this MotA-s 70 interaction helps to stabilize the relatively weak interaction of MotA with the 30 region of middle promoter DNA. # 1999 Academic Press Keywords: activation; AsiA; bacteriophage; MotA; transcription *Corresponding author Introduction During infection, transcription from bacterio- phage T4 middle promoters requires the host RNA polymerase, the T4 transcriptional activator MotA, and the T4 co-activator AsiA (Mattson et al., 1974; Hinton, 1991; Ouhammouch et al., 1995; reviewed by Stitt & Hinton, 1994). MotA is a DNA-binding protein that recognizes a MotA box sequence ((t/a)(t/a)TGCTT(t/c)A) located in the 30 region of middle promoter DNA (Brody et al., 1983; Guild et al., 1988; Hinton, 1991). In addition, both MotA and AsiA interact with the sigma-70 (s 70 ) subunit of RNA polymerase (Stevens & Rhoton, 1975; Stevens, 1976; Gerber & Hinton, 1996). The MotA box sequence is highly conserved among T4 middle promoters (Stitt & Hinton, 1994). However, single base-pair changes throughout this sequence are tolerated in both MotA binding and transcription assays, and some natural middle promoters deviate from the consensus sequence at the most highly conserved base-pairs (Marshall et al., 1999). In addition, wild-type T4 DNA con- tains glucosylated, 5-hydroxymethyl cytosine resi- dues (reviewed by Carlson et al., 1994). This Present address: M. Sharma, AgResearch, Ruakura, East Street, Hamilton, New Zealand; P. Marshall, Ottawa General Hospital Research Institute, 501 Smythe Road, Ottawa, Ontario, Canada K1H 8L6. Abbreviations used: s 70 , sigma-70; DMS, dimethyl sulfate; TBP, TATA-binding protein. E-mail address of the corresponding author: [email protected]Article No. jmbi.1999.2928 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 290, 905–915 0022-2836/99/300905–11 $30.00/0 # 1999 Academic Press
Transcript
Article No. jmbi.1999.2928 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 290, 905±915
Binding of the Bacteriophage T4 TranscriptionalActivator, MotA, to T4 Middle Promoter DNA: Evidencefor both Major and Minor Groove Contacts
Mridula Sharma, Philip Marshall and Deborah M. Hinton*
Laboratory of Molecular andCellular Biology, NationalInstitute of Diabetes andDigestive and Kidney DiseasesNational Institutes of HealthBethesda, MD 20892, USA
Present address: M. Sharma, AgREast Street, Hamilton, New ZealanOttawa General Hospital ResearchRoad, Ottawa, Ontario, Canada K1
Abbreviations used: s70, sigma-7sulfate; TBP, TATA-binding protein
During infection, the bacteriophage T4 transcriptional activator MotA,the co-activator AsiA, and host RNA polymerase are needed to transcribefrom T4 middle promoters. Middle promoters contain a ÿ10 regionrecognized by the s70 subunit of RNA polymerase and a MotA box cen-tered at ÿ30 that is bound by MotA. We have investigated how the lossor modi®cation of base determinants within the MotA box sequence50TTTGCTTTA30 (positions ÿ34 to ÿ26 of a middle promoter) affectsMotA function. Gel retardation assays with mutant MotA boxes are con-sistent with the idea that MotA uses minor groove contacts upstreamand major groove contacts downstream of the center GC, and does notrequire any speci®c base feature at the C �G base-pair at position ÿ30. Inparticular, the 5-methyl residue on the thymine residue at position ÿ29, amajor groove contact, contributes to MotA binding, while converting theT �A at ÿ32 to a C �I base-pair, a change that affects the major but nor theminor groove, yields a MotA box that is similar to wild-type. However,methylation interference analyses indicate that neither the binding ofMotA nor the binding of polymerase/MotA/AsiA to the middle pro-moter PuvsX is inhibited by premethylation of guanine and adenineresidues, suggesting that binding does not require minor groove contactwith any speci®c T �A base-pair. Using gel retardation analyses, we calcu-late an apparent dissociation constant of 130 nM for MotA binding to thewild-type MotA box. Previous work has shown that the N-terminalregion of MotA is needed for an interaction between MotA and s70. Wesuggest that this MotA-s70 interaction helps to stabilize the relativelyweak interaction of MotA with the ÿ30 region of middle promoter DNA.
During infection, transcription from bacterio-phage T4 middle promoters requires the host RNApolymerase, the T4 transcriptional activator MotA,and the T4 co-activator AsiA (Mattson et al., 1974;Hinton, 1991; Ouhammouch et al., 1995; reviewedby Stitt & Hinton, 1994). MotA is a DNA-binding
esearch, Ruakura,d; P. Marshall,Institute, 501 SmytheH 8L6.0; DMS, dimethyl.ing author:
protein that recognizes a MotA box sequence((t/a)(t/a)TGCTT(t/c)A) located in the ÿ30 regionof middle promoter DNA (Brody et al., 1983; Guildet al., 1988; Hinton, 1991). In addition, both MotAand AsiA interact with the sigma-70 (s70) subunitof RNA polymerase (Stevens & Rhoton, 1975;Stevens, 1976; Gerber & Hinton, 1996).
The MotA box sequence is highly conservedamong T4 middle promoters (Stitt & Hinton, 1994).However, single base-pair changes throughout thissequence are tolerated in both MotA binding andtranscription assays, and some natural middlepromoters deviate from the consensus sequence atthe most highly conserved base-pairs (Marshallet al., 1999). In addition, wild-type T4 DNA con-tains glucosylated, 5-hydroxymethyl cytosine resi-dues (reviewed by Carlson et al., 1994). This
906 Binding of Bacteriophage T4 MotA Protein to DNA
modi®cation, which protects the phage from hostrestriction systems as well as phage-encoded endo-nucleases that are speci®c for unmodi®ed DNA,puts a large residue in the major groove of T4DNA. Although the presence of the modi®cation isnot required for MotA-dependent transcriptionfrom many T4 middle promoters (Hinton, 1991;Schmidt & Kreuzer, 1992; Ouhammouch et al.,1995; Adelman et al., 1997), its presence does affectMotA activation from promoters that have particu-lar MotA box sequences (Marshall et al., 1999). Toinvestigate how speci®c base determinants con-tribute to middle promoter function, we haveassayed how various base modi®cations affect thebinding of MotA to the MotA box sequence andthe formation of transcription complexes withpolymerase alone or with polymerase/MotA/AsiA. We have also determined an apparent dis-sociation constant (Kd(app)) using gel retardationassays. We suggest that in the transcription com-plex, the interaction of MotA with s70 helps tostrengthen the binding of MotA to the promoterDNA.
Results
Determination of an apparent dissociationconstant Kd(app) for MotA binding to DNA
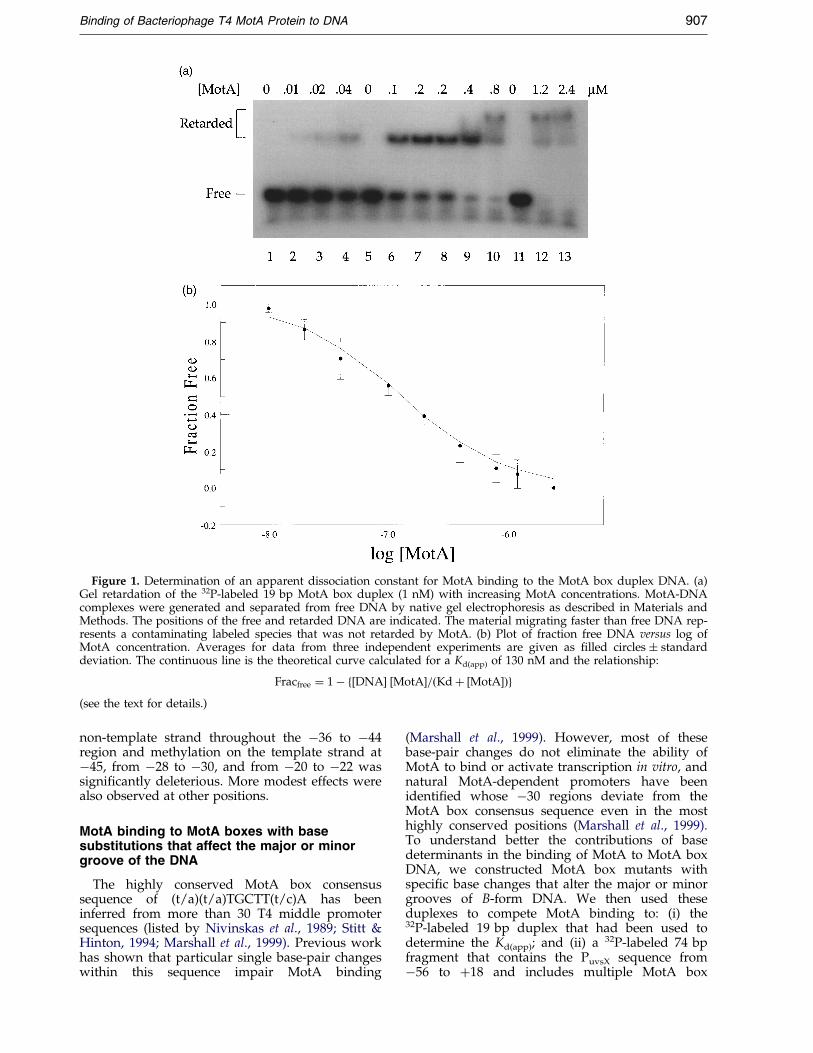
The procedure described by Carey (1988, 1991)was used to determine a Kd(app) for MotA bindingto DNA. In the absence of competitor, a MotA con-centration of 130 nM was needed to retard 50 % ofa 19 bp duplex that contains a single match to theMotA box sequence, 50TTTGCTTTA (Figure 1(a)and (b). This gives a Kd(app) of 130 nM if MotAbinds as a monomer. This Kd(app) agrees with thework by Cicero et al. (1998), who recently deter-mined a Kd of 220 nM, based on steady-state ¯uor-escence anisotropy. A plot of Fracfree versus log[MotA] (Figure 1(b)) demonstrates that our exper-imental data ®t the theoretical curve that is gener-ated using the Kd(app) of 130 nM and assuming thatthe binding of MotA can be represented by thesimple equilibrium:
MotA�DNA !MotA �DNA
(see Materials and Methods for details). However,in the gel retardation assays, a single retardedband was seen at the lower MotA concentrations(Figure 1(a), lanes 2-4, 6-8), but slower migratingcomplexes appeared as the MotA concentrationwas increased (lanes 9, 10, 12, 13). Using a differentMotA box DNA substrate, Cicero et al. (1998) haveshown that the fastest moving species consists of asingle MotA monomer bound to a single MotAbox substrate, but the slower moving complexescontain additional MotA proteins. Thus, eventhough our data ®t the theoretical curve for simplebinding, it is likely that the Kd(app) contains contri-butions from other types of binding.
Methylation of guanine and adenine residuesdoes not interfere with the binding of MotA orpolymerase/AsiA/MotA to the T4 middlepromoter PuvsX
Treatment of DNA with dimethyl sulfate (DMS)methylates the N7 position of guanine and the N3position of adenine residues, positions that lie inthe major and minor grooves, respectively, ofB-form DNA (Siebenlist & Gilbert, 1980;Hendrickson & Schleif, 1985; Wang et al., 1998). Toanalyze complexes between MotA and the T4middle promoter PuvsX we used a 200 bp fragment,which included the T4 sequences from 94 bpupstream to 83 bp downstream of the start of tran-scription. This DNA contains three MotA bindingsites, centered at ÿ51, ÿ30, and �49 (March-Amegadzie & Hinton, 1995), but only the site at -30 is required for transcription (Marshall et al.,1999). In methylation interference assays, no sig-ni®cant differences were observed in the cleavagepatterns of DNA that was bound with MotA versusthe DNA that was free (Figures 2(a) and (b), upperpanels, red and green scans, respectively). Theseresults are consistent with the idea that MotA doesnot require major groove contact with any speci®cguanine residue or minor groove contact with anyspeci®c adenine residue for binding. However, wecannot eliminate the possibility that our failure toobserve interference was due to the presence ofmultiple MotA binding sites in the PuvsX DNA.
PuvsX contains a perfect match to the s70 ÿ 10consensus sequence (Figure 2(c)) and in vitro, it isbound and transcribed ef®ciently by either poly-merase alone or by polymerase/AsiA/MotA (acti-vated transcription) (Hinton et al., 1996a).However, transcription by polymerase alone doesnot occur during T4 infection (Guild et al., 1988;reviewed by Stitt & Hinton, 1994). We performedmethylation interference assays using PuvsX DNAand polymerase alone or polymerase/AsiA/MotA.Within the ÿ30 MotA box region, the cleavage pat-terns obtained with polymerase/AsiA/MotA weresimilar to those observed with MotA alone(Figure 2(a) and (b), upper panels, black scans). Inaddition, only a few differences between the clea-vage patterns obtained with MotA or polymerase/AsiA/MotA were seen in other regions of the pro-moter. There was some enhancement by methyl-ation of the guanines at positions ÿ14, ÿ15 and ofthe guanine and adenine residues at positions ÿ11to �3 on the non-template strand, and some inter-ference from guanine methylation on the templatestrand at �10. In contrast, many differences wereobserved between the cleavage patterns of boundand free DNA obtained after formation of the com-plex with polymerase alone (Figure 2(a) and (b),lower panels). Of particular interest is that N7guanine methylation at positions ÿ14 and ÿ15on the non-template strand interfered with theformation of the complex with polymerase alonewhile it somewhat enhanced the formation of theactivated complex. In addition, methylation on the
Figure 1. Determination of an apparent dissociation constant for MotA binding to the MotA box duplex DNA. (a)Gel retardation of the 32P-labeled 19 bp MotA box duplex (1 nM) with increasing MotA concentrations. MotA-DNAcomplexes were generated and separated from free DNA by native gel electrophoresis as described in Materials andMethods. The positions of the free and retarded DNA are indicated. The material migrating faster than free DNA rep-resents a contaminating labeled species that was not retarded by MotA. (b) Plot of fraction free DNA versus log ofMotA concentration. Averages for data from three independent experiments are given as ®lled circles � standarddeviation. The continuous line is the theoretical curve calculated for a Kd(app) of 130 nM and the relationship:
Fracfree � 1ÿ f�DNA� �MotA�=�Kd� �MotA��g(see the text for details.)
Binding of Bacteriophage T4 MotA Protein to DNA 907
non-template strand throughout the ÿ36 to ÿ44region and methylation on the template strand atÿ45, from ÿ28 to ÿ30, and from ÿ20 to ÿ22 wassigni®cantly deleterious. More modest effects werealso observed at other positions.
MotA binding to MotA boxes with basesubstitutions that affect the major or minorgroove of the DNA
The highly conserved MotA box consensussequence of (t/a)(t/a)TGCTT(t/c)A has beeninferred from more than 30 T4 middle promotersequences (listed by Nivinskas et al., 1989; Stitt &Hinton, 1994; Marshall et al., 1999). Previous workhas shown that particular single base-pair changeswithin this sequence impair MotA binding
(Marshall et al., 1999). However, most of thesebase-pair changes do not eliminate the ability ofMotA to bind or activate transcription in vitro, andnatural MotA-dependent promoters have beenidenti®ed whose ÿ30 regions deviate from theMotA box consensus sequence even in the mosthighly conserved positions (Marshall et al., 1999).To understand better the contributions of basedeterminants in the binding of MotA to MotA boxDNA, we constructed MotA box mutants withspeci®c base changes that alter the major or minorgrooves of B-form DNA. We then used theseduplexes to compete MotA binding to: (i) the32P-labeled 19 bp duplex that had been used todetermine the Kd(app); and (ii) a 32P-labeled 74 bpfragment that contains the PuvsX sequence fromÿ56 to �18 and includes multiple MotA box
Figure 2 (Legend shown opposite)
908 Binding of Bacteriophage T4 MotA Protein to DNA
Figure 2. Methylation interference analyses. Densitometry scans of peak intensity (relative OD units) versus baseposition of the methylation interference footprints obtained with the PuvsX fragment labeled on either (a) the templateor (b) the non-template strand. In each case, the upper panel shows scans of DNA that was bound by MotA (red),DNA that was not bound by MotA (green) and DNA that was bound by polymerase/MotA/AsiA (black); the lowerpanel shows DNA that was bound by polymerase alone (red) and DNA that was not bound by polymerase alone(green). (See Materials and Methods for details.) For the scans shown in (a), the peak heights (in OD units) at baseposition �10 were 0.33 (DNA not bound by MotA), 0.24 (DNA bound by MotA), 0.57 (DNA bound by polymerase/Mot/AsiA), 0.26 (DNA not bound by polymerase), and 0.16 (DNA bound by polymerase). For the scans shown in(b), the peak heights (in OD units) at base position ÿ51 were 0.48 (DNA not bound by MotA), 0.32 (DNA bound byMotA), 0.55 (DNA bound by polymerase/MotA/AsiA), 0.34 (DNA not bound by polymerase), and 0.32 (DNAbound by polymerase). (c) Sequence of PuvsX from positions ÿ56 to �18. The ÿ10 region, which contains the con-sensus sequence for s70, and the MotA box sequence centered at ÿ30 are shown in bold. Other matches to the MotAbox consensus sequence are indicated by the arrows.
Binding of Bacteriophage T4 MotA Protein to DNA 909
sequences (see Figure 2(c)). Unlike the 19 bpduplex, the larger DNA formed several retardedcomplexes in the gel retardation assays (March-Amegadzie & Hinton, 1995; Figure 3, this study),presumably because of the multiple MotA boxesand because of the non-speci®c binding that wasalso seen with the 19 bp DNA (Figure 1(a)). How-ever, even though quantitative differences wereobserved when comparing a competitor with eachof these DNAs, the overall results were similar(Table 1).
A T �A to C �I conversion alters the major grooveof B-form DNA with little effect on the minorgroove (Starr & Hawley, 1991). Thus, changing allthe T �A base-pairs to C � I base-pairs in the bindingsite of the TATA-binding protein (TBP), whichbinds in the minor groove of its target DNA (J. L.Kim et al., 1993; Y. Kim et al., 1993), had no effecton TBP binding (Starr & Hawley, 1991). The wild-type MotA box sequence used for our assays has
the sequence 50TTTGCTTTA30. As seen in Table 1and Figure 3 (lane 5), converting all the T �A base-pairs to C �I base-pairs resulted in a poor competi-tor. This result is inconsistent with binding thatonly involves minor groove contacts. However, notall of the T �A base-pairs were equivalent. Chan-ging just the upstream T �A base-pairs (positionsÿ34 to ÿ32) produced a competitor that was muchless impaired (Table 1 and Figure 3, lane 6), and aT �A to C � I conversion at position ÿ32 produced acompetitor that was similar to wild-type (Table 1and Figure 3, lane 7). The importance of theÿ32 T �A base-pair for MotA function has pre-viously been shown by the fact that a ÿ32 T �A toA �T change impairs MotA binding (Marshall et al.,1999). That result was con®rmed here in a compe-tition with the 74 bp fragment (Table 1). Thus, asimple explanation for these results is that there isa minor groove contact between MotA and theT �A base-pair at position ÿ32. However, the meth-
Figure 3. Competition gel retar-dation assay. Assays without (lane1) or with (lanes 2-12) MotA con-tained the 32P-labeled 74 bp PuvsX
fragment and, where indicated, a1000-fold excess of the indicatednon-radioactive competitor. SeeTable 1 for sequences of the compe-titor DNAs and Materials andMethods for reaction conditions.The positions of free and retardedDNA are indicated.
Table 1. Gel retardation assays with wild-type and mutant MotA box sequences
How effective is the competitorb versus the MotA box duplexes of
MotA Box presentas competitora 19 bpc (%) 74 bpc (%)
ÿ34 ÿ 26Wild-type TTTGCTTTA 100 100
AAACGAAAT
multiple mutant TTATACTTA 11 3AATATGAAT
all C �I CCCGCCCCI 31 8IIICGIIIC
ÿ34 to ÿ32 C �I CCCGCTTTA Not tested 72IIICGAAAT
ÿ32 C �I TTCGCTTTA 103 89AAICGAAAT
ÿ32 A �T TTAGCTTTA Not tested 10AATCGAAAT
ÿ31, ÿ30 I �C, C �I TTTICTTTA 65 67AAACIAAAT
ÿ31 I �C TTTICTTTA 88 94AAACGAAAT
ÿ30 C �I TTTGCTTTA 76 88AAACIAAAT
ÿ30 A �T TTTGATTTA 110 99AAACTAAAT
ÿ30 A �G TTTGATTTA 0 0AAACGAAAT
ÿ29 C �I TTTGCCTTA 60 44AAACGIAAT
ÿ29 Cme �I TTTGCCmeTTA 106 106AAACGI AAT
ÿ29 C �G TTTGCCTTA 42 26AAACGGAAT
ÿ29 Cme �G TTTGCCmeTTA 83 86AAACGG AAT
ÿ29 dU �A TTTGCUTTA 74 69AAACGAAAT
ÿ28 C �I TTTGCTCTA 52 32AAACGAIAT
ÿ28 Cme �I TTTGCTCmeTA 56 37AAACGAI AT
ÿ28 dU �A TTTGCTUTA 92 91AAACGAAAT
a The MotA box sequence (shown underlined below) was present within the double stranded oligomer:
50 AATTATTTGCTTTAGATTA 30
30 TTAATAAACGAAATCTAAT 50
b Calculated using the formula given in Materials and Methods. For experiments with the labeled 19 bp duplex, the fraction boundwithout competitor was 59(�4.7)% and the fraction bound with wild-type competitor was 23(�2.0)%. For experiments with thelabeled 74 bp duplex, the fraction bound without competitor was 95(�1.2)% and the fraction bound with the wild-type competitorwas 28(�3.3)%. Data represent averages of two to ten experiments except for the competition of the 19 bp duplex with the ÿ30 A �Tand ÿ30 A �G mutants, which was done once.
c The 19 and 74 bp MotA duplexes are described in Materials and Methods.
910 Binding of Bacteriophage T4 MotA Protein to DNA
ylation interference results using PuvsX indicatedthat the N3 methylation of the adenine residue atÿ32 is not inhibitory, suggesting that either thiscontact is not affected by the N3 modi®cation orthat it can be lost without seriously affecting bind-ing. In addition, the ÿ32 T �A to A �T base-pairchange is found in some natural T4 middle promo-
ters, supporting the idea that a minor groove con-tact here is not a requirement for MotA function.
The T �A base-pairs at positions ÿ29 and ÿ28 arehighly conserved among T4 middle promoters.Changing either the ÿ29 T �A to C �G or the ÿ28T �A to G �C reduced MotA binding (Marshall et al.,1999), suggesting that MotA might recognize base
Binding of Bacteriophage T4 MotA Protein to DNA 911
features at these base positions. Here we con®rmedthat the ÿ29 T �A to C �G change is deleterious(Table 1; Figure 3, lane 9). In addition, we foundthat conversion of the T �A base-pair to a C �I base-pair at either position ÿ29 or ÿ28 resulted in animpaired competitor for MotA binding, a resultthat is compatible with MotA making a major, butnot a minor, groove contact at these positions. The5-methyl residue of a thymine base is a majorgroove determinant for some DNA binding pro-teins (Dubendorff et al., 1987; Fliess et al., 1988;Maslak et al., 1993). The importance of this methylgroup can be assayed by the T �A to C �I change,which removes the 5-methyl residue but alsomakes other changes to the major groove, or by aT �A to dU �A base-pair conversion, which simplyremoves the 5-methyl residue (Delort et al., 1985).For the T �A at position ÿ28, the presence of the5-methyl group was inconsequential (Table 1).Conversion of the T �A base-pair to a dU �A base-pair had practically no effect, and converting theT �A base-pair to Cme � I was as deleterious as con-verting it to C �I. In contrast, the presence of the 5-methyl group at position ÿ29 either in a Cme �I or aCme �G base-pair signi®cantly improved the abilityof the oligomer to compete for MotA binding(Table 1 and Figure 3, lanes 8, 9, 11 and 12). More-over, loss of the methyl group (the T �A to dU �Aconversion) had a more serious effect here than atposition ÿ28 (Table 1 and Figure 3, lane 10). Theeffect of some of these changes at position ÿ29 wasalso assessed in gel retardation experiments usinglabeled oligomers without competitor (Table 2).These results indicated that the ÿ29 Cme �I and ÿ29dU �A mutants require twofold and fourfold moreMotA, respectively, than does the wild-type inorder to bind half of the DNA. This argues that the5-methyl group at position ÿ29 contributes toMotA binding and its presence decreases theKd(app) by a factor of around four.
The methylation interference analyses indicatedthat MotA does not require a major groove contactwith the guanines at positions ÿ31 or ÿ30. A G �Cto I �C conversion removes the 2-amino residuefrom the guanine residue, a minor groove determi-nant for some proteins (Schick & Martin, 1993;Wang et al., 1998). A mutant MotA box in whichboth of the G �C base-pairs (positions ÿ31 and ÿ30)had been changed to I �C bps gave some reductionin binding, but changing either one of these gua-
Table 2. Binding of MotA to wild-type, ÿ29 dU �A, and ÿ29
a Fraction of the (50-32P)-labeled 19 bp fragment containing the wretarded during gel electrophoresis. (See Table 1 for the sequencesaveraged from two to four experiments except for the the ÿ29Cme � I
nine residues separately had practically no effect atposition ÿ31 and only a modest effect at positionÿ30 (Table 1). In addition, we have found that themore drastic change of a C �G to A �T change atposition ÿ30 produces a competitor that is indistin-guishable from wild-type (Marshall et al., 1999;Table 1), and changing position ÿ30 to either anA �T, G �C, or T �A still results in a MotA box thatcan be activated in vitro (Marshall et al., 1999).Taken together, our results suggest that MotA doesnot need any speci®c base feature at position ÿ30and that the guanine residue at position ÿ31 is notcontacted in the major groove or through its 2-amino group in the minor groove. However, wefound that changing the C �G base-pair at positionÿ30 to an A �G base-pair eliminated binding(Table 1), suggesting that a disruption of base pair-ing at this position cannot be tolerated.
Discussion
Bacteriophage T4 middle promoters (listed byNivinskas et al., 1989; Stitt & Hinton, 1994;Marshall et al., 1999) have two regions of con-served sequence, centered at ÿ10 and ÿ30. In theÿ10 region, all identi®ed middle promoters containa strong match to the s70 ÿ 10 consensus sequence(TAnnnT). This match is critical for MotA-depen-dent transcription (Marshall et al., 1999). Its pre-sence also explains the ability of polymerase aloneto use middle promoters when they are present incloned DNA (reviewed by Stitt & Hinton, 1994).However, during infection this transcription bypolymerase alone is not observed, and middle pro-moter activity requires MotA and AsiA. The ÿ30region of a middle promoter contains the bindingsite for MotA (Brody et al., 1983; Guild et al., 1988;Hinton, 1991). Although a highly conserved MotAbox consensus sequence of 50(t/a)(t/a)TGCTT(t/c)A can be deduced for this region, it has beendif®cult to determine which base determinants areimportant for MotA action. MotA tolerates changeswithin the most conserved portion of thissequence. Furthermore, in the natural T4 template,each cytosine residue is modi®ed by the additionof a glucosylated, hydroxymethyl moiety at baseposition 5. Although this addition modi®es thecenter of the MotA box sequence in the majorgroove, it is not required for MotA-dependent tran-scription from many T4 middle promoters which
ild-type or the indicated mutant MotA box sequences that wasof the DNAs and Materials and Methods for details.) Values aremutant with 400 nM MotA which is from a single experiment.
Figure 4. Model for RNA polymerase, MotA andAsiA at a T4 middle promoter. The subunits of RNApolymerase (a2bb
0s70) and the T4 proteins MotA andAsiA are depicted on middle promoter DNA with thes70 ÿ 10 region and MotA box region centered at ÿ30shown in bold. The NTD (N-terminal domain) and CTD(C-terminal domain) are indicated for the a subunits ofRNA polymerase and for MotA, and domains 2.4 and4.2 are indicated for s70. Placement of the a2, b, b0 sub-units about the promoter DNA and of domain 2.4 s70
on the ÿ10 region is based on analyses with E. coli pro-moters (for reviews, see Busby & Ebright, 1994;Hochschild & Dove, 1998). The a CTDs are shown withjagged lines to depict ADP-ribosylation of Arg265 in theCTD of the a subunits by T4-encoded proteins (for areview, see Stitt & Hinton, 1994). Placement of MotAand AsiA proteins is discussed in the text.
912 Binding of Bacteriophage T4 MotA Protein to DNA
have a center motif of GCTT (Hinton, 1991;Schmidt & Kreuzer, 1992; Ouhammouch et al.,1995; Adelman et al., 1997). However, the modi®-cation does affect promoters with certain otherMotA box sequences, suggesting that it may con-tribute to MotA function depending on thesequence context (Marshall et al., 1999).
We have asked how base modi®cations affectthe binding of MotA to the MotA box and the for-mation of transcription complexes by polymerasealone or with polymerase/MotA/AsiA. For thecomplex with PuvsX and polymerase alone, wefound protein-DNA contacts that are typical forpolymerase and host promoters (Hinton et al.,1996a; this study). Although cleavage patterns inmethylation interference assays are dependent onthe exact promoter sequence, previous studies withEscherichia coli promoters have found that methyl-ation on the non-template strand just upstream ofthe ÿ10 or ÿ35 regions and guanine methylationon the template strand around position ÿ30 isinhibitory (Siebenlist et al., 1980; Simpson, 1982;Minchin & Busby, 1993). Likewise, for PuvsX, gua-nine methylation at positions ÿ14 and ÿ15 of thenon-template strand and ÿ30 of the templatestrand particularly interfered with the formation ofthe complex of PuvsX and polymerase alone. Inthe T4 template, the cytosine residue modi®cationprovides a major groove modi®cation at thesesites. Thus, this natural modi®cation may contrib-ute to the suppression of transcription by polymer-ase alone.
The protein-DNA contacts made in the activatedcomplex differed signi®cantly from those made bypolymerase alone. Major groove modi®cation ofguanine residues did not interfere with the bindingof MotA or polymerase/MotA/AsiA to PuvsX, con-sistent with the fact that the natural major groovemodi®cation by T4 at its C �G base-pairs is notinhibitory. Within the MotA box sequence of50TTTGCTTTA30 (corresponding to positions ÿ34to ÿ26 of a middle promoter), our results suggestthat MotA uses minor groove contacts upstreamand major groove contacts downstream of the cen-ter G �C and does not require any speci®c base fea-ture at the C �G base-pair at position ÿ30. Thisconclusion con¯icts with the previous suggestionthat the DNA-binding domain of MotA is similarto that of the TATA-binding protein (Finnin et al.,1994). Crystal structures have shown that TBPbends the DNA sharply as it binds in the minorgroove of its target sequence (J. L. Kim et al., 1993;Y. Kim et al., 1993). As expected from such astructure, converting all of the T �A base-pairs ofthe TATA box to C � I base-pairs results in a DNAthat is still fully competent to bind TBP (Starr &Hawley, 1991) and N3 methylation of adenine resi-dues within the binding sequence is quite deleter-ious (Starr & Hawley, 1991; Lee et al., 1991). Thus,we conclude that MotA binding and activationfrom unmodi®ed DNA is not analogous to that ofTBP and its TATA box. However, we cannot elim-inate the possibility that MotA contacts modi®ed
T4 DNA in a way that differs from what we havedetermined with unmodi®ed templates.
The Kd measurements, again using unmodi®edDNA (Cicero et al., 1998; this study), are consistentwith the idea that MotA is not a tight DNA bind-ing protein. Despite this fact, stoichiometric ratiosof MotA relative to DNA are suf®cient for MotAactivation of transcription or to generate a stablepre-transcription complex of polymerase/MotA/AsiA and PuvsX (March-Amegadzie & Hinton,1995; Hinton et al., 1996a). This suggests that someaspect of the transcription complex stabilizes theinteraction of MotA with the DNA. Based on workwith E. coli promoters, a model of how the sub-units of RNA polymerase are positioned about ahost promoter has emerged (for reviews, see Busby& Ebright, 1994; Hochschild & Dove, 1998). In thismodel, a central domain of s70 (domain 2.4) andthe far C-terminal domain of s70 (domain 4.2)directly contact the ÿ10 and ÿ35 regions, respect-ively, of the promoter DNA. For a T4 middle pro-moter, it appears that a strong contact betweendomain 2.4 and the ÿ10 region is retained, but con-tact between domain 4.2 and the ÿ35 region of thepromoter has been disrupted. Instead, the C termi-nus of s70 interacts with AsiA (Severinova et al.,1996; Adelman et al., 1998; Severianov & Muir,1998). MotA also interacts with s70, and this inter-action requires the N-terminal domain of MotA(Gerber & Hinton, 1996). Other work hassuggested that the C-terminal half of MotA con-
Binding of Bacteriophage T4 MotA Protein to DNA 913
tains the DNA-binding domain (D.M.H., unpub-lished results). We propose that the s70-MotAinteraction anchors the C-terminal half of MotA tothe MotA box sequence (Figure 4). Given thatMotA is bound to the ÿ30 region of the promoter,it seems likely that the C-terminal region of s70 isalso involved in the s70-MotA interaction. Thus,together MotA and AsiA create new protein-protein and protein-DNA contacts, which thengenerate a polymerase with an altered sequencespeci®city for the ÿ30 region, but not the ÿ10region of promoter DNA.
Materials and Methods
Proteins and reaction buffers
Wild-type MotA and AsiA were obtained as described(Hinton et al., 1996b). E. coli RNA polymerase was pur-chased from USB of Amersham Life Science.
The 1 � Kglu transcription buffer (Zou & Richardson,1991) contains 40 mM Tris-acetate (pH 7.9), 150 mM pot-assium glutamate, 4 mM magnesium acetate, 0.1 mMEDTA, 0.1 mM DTT, and 100 mg/ml bovine serumalbumen.
DNA
The wild-type MotA box 19 bp duplex DNA containspositions ÿ37 to ÿ19 of the T4 middle promoter P57a
(Guild et al., 1988) and has a single match to the MotAbox consensus sequence. (The promoter positionsassigned to this MotA box (shown in Table 1) match thepositions of the MotA box in the T4 middle promoterPuvsX.) The (50-32P)-labeled and unlabeled 19 bp duplexwas obtained as described (Marshall et al., 1999). The(50-32P)-labeled 200 bp PuvsX DNA, containing PuvsX
sequences from positions ÿ94 to �83, was generated andpuri®ed as described (March-Amegadzie & Hinton,1995).
Densitometry of autoradiograms
After autoradiography, ®lms were scanned using aScanmaster III Plus from Howtek, Inc. Quanti®cationwas performed using Diversity One software from Pro-tein-Databases, Inc.
Dissociation constant determination
An apparent dissociation constant Kd(app) for MotAbinding to DNA was determined from gel retardationassays according to the procedure described by Carey(1988, 1991). The labeled wild-type 19 bp duplex (1 nM )was incubated at 4 �C for 30 minutes in a reaction (10 ml)containing from 0 to 2400 nM MotA and 1 � Kglu trans-cription buffer. Reactions were loaded on native 12 %(w/v) polyacrylamide (37.5 acrylamide:1 bis-acrylamide)gels, and protein-DNA complexes were resolved fromfree DNA by electrophoresis at 10 V/cm for three hoursat 4 �C. The fraction of free DNA (Fracfree) remaining ata particular MotA concentration ([MotA]X) was deter-mined as follows:
Fracfree � �amount of free DNA at �MotA�X-background��amount of free DNA �MotA�0- background�
The background arose from a radioactive species thatmigrated slightly faster than the labeled duplex (seeFigure 1(a)). The background was determined at a MotAconcentration of 2400 nM, which was suf®cient to retardall of the legitimate duplex. The fraction bound was cal-culated as 1-Fracfree and plotted versus the MotA concen-tration. Three data sets were averaged for Figure 1(b).Two sets were obtained with the duplex labeled on thetop strand; one set with the duplex labeled on the bot-tom strand. The Kd(app) value is the MotA concentrationneeded to bind 50 % of the DNA. Because the concen-tration of MotA in the gel retardation assays was inexcess over that of the DNA, our data should ®t theequation:
1-Fracfree � Fracbound � �DNA� �MotA�=�Kd�app� � �MotA��The curve in Figure 1(b) was generated from thisequation using the Kd(app) value of 130 nM.
Gel retardation assays
Gel retardation assays using the (50-32P)-labeled wild-type MotA box duplex, the ÿ29 dU �A mutant, or theÿ29 Cme � I mutant (Table 2) were performed as describedabove for the Kd(app) determination, except that the con-centration of the DNA was 4 nM.
For the competition gel retardation assays, reactions(10 ml) containing labeled 74 bp duplex (0.01 pmol) orlabeled 19 bp duplex (0.04 pmol), 1 � Kglu transcriptionbuffer, 2 pmol MotA (for the 74 bp substrate) or 4 pmolMotA (for the 19 bp substrate), and 0 or 10 pmol of thevarious unlabeled 19 bp duplexes were incubated at 4 �Cfor 30 minutes. Protein-DNA complexes were separatedfrom free DNA by electrophoresis on native 8 % (for the74 bp substrate) or 12 % (for the 19 bp substrate) poly-acrylamide (37.5 acrylamide:1 bis-acrylamide) gels run in1 � TBE buffer at 10 V/cm for three hours at 4 �C. Thecompetitive effectiveness of a particular competitor Xrelative to the wild-type 19 bp duplex was calculated as:
��Fraction bound without competitor-fraction
bound with competitor X�=�fraction bound
without competitor-fraction bound with wild-type
as competitor�� X 100
Methylation interference assays
The (50-32P)-labeled PuvsX 200 bp fragment (0.7 pmol in205 ml of a solution containing 50 mM Tris-HCl (pH 8.0),10 mM MgCl2, 0.02 mM EDTA) was treated with 1 mlDMS for three minutes at room temperature. DMS stopsolution (51 ml) was added and the DNA precipitatedwith ethanol twice. The pellet was then washed twicewith ethanol, dried, and dissolved in TE.
Methylated DNA (0.1 pmol) was incubated for ®veminutes at 37 �C in a reaction (10 ml) containing 1 � Kglutranscription buffer and as indicated, no protein; 5 pmolMotA; 1 pmol RNA polymerase; or 5 pmol MotA � 5 p-mol AsiA � 1 pmol RNA polymerase. Protein-DNAcomplexes were challenged by the addition of (polydI:dC) (1 ml of solution at 360 ng/ml) for 1.5 min at 37 �Cand then separated from free DNA by electrophoresis asdescribed (Hinton et al., 1996a). For the incubations ofDNA with 5 pmol of MotA alone or with polymerasealone, both the DNA present in the protein-DNA com-
914 Binding of Bacteriophage T4 MotA Protein to DNA
plex (bound DNA) and the DNA migrating as free DNAwere excised from the gel. (In each case, about 50 % ofthe DNA was bound.) For the incubationsof DNA with polymerase �MotA � AsiA, almost all ofthe DNA was in the complex with polymerase. Theremaining DNA was in a complex with the excess MotApresent in the reaction. Only the species in the complexwith polymerase �MotA � AsiA was excised. DNA wasisolated after electroelution onto NA45 membranes asdescribed (Hinton et al., 1996a). The DNA was thencleaved at sites of guanine and adenine methylationusing the procedure by Wang et al. (1998). DNA frag-ments were separated on denaturing 8 % polyacrylamide(19 acrylamide: 1 bis-acrylamide), 7 M urea gels run in1/2 � TBE.
Acknowledgements
We are grateful to Nancy Nossal and Stoney Simonsfor helpful discussions.
References
Adelman, K., Orsini, G., Kolb, A., Graziani, L. & Brody,E. N. (1997). The interaction between the AsiA pro-tein of bacteriophage T4 and the s70 subunit ofEscherichia coli RNA polymerase. J. Biol. Chem. 272,27435-27443.
Adelman, K., Brody, E. N. & Buckle, M. (1998). Stimu-lation of bacteriophage T4 middle transcription bythe T4 proteins MotA and AsiA occurs at two dis-tinct steps in the transcription cycle. Proc. Natl Acad.Sci. USA, 95, 15247-15252.
Brody, E., Rabussay, D. & Hall, D. H. (1983). Regulationof transcription of prereplicative genes. In Bacterio-phage T4 (Mathews, C. K., Kutter, E., Mosig, G. &Berget, P. B., eds), pp. 174-183, American Societyfor Microbiology, Washington, DC.
Busby, S. & Ebright, R. (1994). Promoter structure, pro-moter recognition, and transcription activation inprokaryotes. Cell, 79, 743-746.
Carey, J. (1988). Gel retardation at low pH resolves trprepressor-DNA complexes for quantitative study.Proc. Natl Acad. Sci. USA, 85, 975-979.
Carey, J. (1991). Gel retardation. Methods Enzymol. 208,103-117.
Carlson, K., Raleigh, E. A. & Hattman, S. (1994). Restric-tion and modi®cation. In Molecular Biology of Bac-teriophage T4 (Karam, J. D., Drake, J. W., Mosig, G.,Hall, D. H., Eiserling, F. A., Black, L. W., Spicer,E. K., Kutter, E., Carlson, K. & Miller, E. S., eds),pp. 369-381, American Society for Microbiology,Washington, DC.
Cicero, M. P., Alexander, K. A. & Kreuzer, K. N. (1998).The MotA transcriptional activator of bacteriophageT4 binds to its speci®c DNA site as a monomer. Bio-chemistry, 37, 4977-4984.
Delort, A.-M., Neumann, J. M., Molko, D., HerveÂ, M.,TeÂoule, R. & Dinh, S. (1985). In¯uence of uracildefect on DNA structure: 1H NMR investigation at500 MHz. Nucl. Acids Res. 13, 3343-3355.
Dubendorff, J. W., deHaseth, P. L., Rosendahl, M. S. &Caruthers, M. H. (1987). DNA functional groupsrequired for formation of open complexes betweenEscherichia coli RNA polymerase and the l PR pro-moter. J. Biol. Chem. 262, 892-898.
Finnin, M. S., Hoffman, D. W. & White, S. W. (1994).The DNA-binding domain of the MotA transcrip-tion factor from bacteriophage T4 shows structuralsimilarity to the TATA-binding protein. Proc. NatlAcad. Sci. USA, 91, 10972-10976.
Fliess, A., Wolfes, H., Seela, F. & Pingoud, A. (1988).Analysis of the recognition mechanism involved inthe EcoRV catalyzed cleavage of DNA using modi-®ed oligodeoxynucleotides. Nucl. Acids Res. 16,11781-11793.
Gerber, J. S. & Hinton, D. M. (1996). An N-terminalmutation in the bacteriophage T4 motA gene yieldsa protein that binds DNA but is defective for acti-vation of transcription. J. Bacteriol. 178, 6133-6139.
Guild, N., Gayle, M., Sweeney, T., Hollingsworth, T.,Modeer, T. & Gold, L. (1988). Transcriptional acti-vation of bacteriophage T4 middle promoters bythe motA protein. J. Mol. Biol. 199, 241-258.
Hendrickson, W. & Schleif, R. (1985). A dimer of AraCprotein contacts three adjacent major groove regionsat the Ara I DNA site. Proc. Natl Acad. Sci. USA, 82,3129-3133.
Hinton, D. M. (1991). Transcription from a bacterio-phage T4 middle promoter using T4 motA proteinand phage-modi®ed RNA polymerase. J. Biol. Chem.266, 18034-18044.
Hinton, D. M., March-Amegadzie, R., Gerber, J. S. &Sharma, M. (1996a). Characterization of pre-tran-scription complexes made at a bacteriophage T4middle promoter: involvement of the T4 MotA acti-vator and the T4 AsiA protein, a s70 binding pro-tein, in the formation of the open complex. J. Mol.Biol. 256, 235-248.
Hinton, D. M., March-Amegadzie, R., Gerber, J. &Sharma, M. (1996b). The bacteriophage T4 middletranscription system: T4-modi®ed RNA polymerase,AsiA (Sigma-70 binding protein), and the transcrip-tional activator MotA. Methods Enzymol. 274, 43-57.
Hochschild, A. & Dove, S. L. (1998). Protein-protein con-tacts that activate and repress transcription. Cell, 92,597-600.
Kim, J. L., Nikolov, D. B. & Burley, S. K. (1993). Co-crystal structure of TBP recognizing the minorgroove of a TATA element. Nature, 365, 520-527.
Kim, Y., Geiger, J. H., Hahn, S. & Sigler, P. B. (1993).Crystal structure of a yeast TBP/TATA-box com-plex. Nature, 365, 512-520.
Lee, D. K., Horikoshi, M. & Roeder, R. G. (1991). Inter-action of TFIID in the minor groove of the TATAelement. Cell, 67, 1241-1250.
March-Amegadzie, R. & Hinton, D. M. (1995). The bac-teriophage T4 middle promoter PuvsX: analysis ofregions important for binding of the T4 transcrip-tional activator MotA and for activation of tran-scription. Mol. Microbiol. 15, 649-660.
Marshall, P., Sharma, M. & Hinton, D. M. (1999). Thebacteriophage T4 transcriptional activator MotAaccepts various base pair changes within its bindingsequence. J. Mol. Biol. 285, 931-944.
Maslak, M., Jaworski, M. D. & Martin, C. T. (1993).Tests of a model for promoter recognition by T7RNA polymerase: thymine methyl group contacts.Biochemistry, 32, 4270-4274.
Mattson, T., Richardson, J. & Goodin, D. (1974). Mutantof bacteriophage T4D affecting expression of manyearly genes. Nature, 250, 48-50.
Minchin, S. & Busby, S. (1993). Location of close con-tacts between Escherichia coli RNA polymerase and
Binding of Bacteriophage T4 MotA Protein to DNA 915
guanine residues either with or without ÿ35 regionsequences. Biochem J. 289, 771-775.
Nivinskas, R. G., Raudonikiene, A. & Guild, N. (1989).A new early gene upstream of the middle gene 31in bacteriophage T4: cloning and expression. Mol.Biol. (Moscow), 23, 739-749.
Ouhammouch, M., Adelman, K., Harvey, S. R., Orsini,G. & Brody, E. N. (1995). Bacteriophage T4 MotAand AsiA proteins suf®ce to direct Escherichia coliRNA polymerase to initiate transcription at T4middle promoters. Proc. Natl Acad. Sci. USA, 92,1415-1419.
Schick, C. & Martin, C. T. (1993). Identi®cation ofspeci®c contacts in T3 RNA polymerase-promoterinteractions: kinetic analysis using small syntheticpromoters. Biochem. 32, 4275-4280.
Schmidt, R. P. & Kreuzer, K. N. (1992). Puri®ed motAprotein binds the ÿ30 region of a bacteriophage T4middle-mode promoter and activates transcriptionin vitro. J. Biol. Chem. 267, 11399-11407.
Severinov, K. & Muir, T. M. (1998). Expressed proteinligation, a novel method for studying protein-pro-tein interactions in transcription. J. Biol. Chem. 273,16205-16209.
Severinova, E., Severinov, K., FenyoÈ , D., Marr, M.,Brody, E. N., Roberts, J. W., Chait, B. T. & Darst, S.(1996). Domain organization of the Escherichia coliRNA polymerase s70 subunit. J. Mol. Biol. 263,637-647.
Siebenlist, U. & Gilbert, W. (1980). Contacts betweenEscherichia coli RNA polymerase and an early pro-moter of phage T7. Proc. Natl Acad. Sci. USA, 77,122-126.
Siebenlist, U., Simpson, R. B. & Gilbert, W. (1980). E. coliRNA polymerase interacts homologously with twodifferent promoters. Cell, 20, 269-281.
Simpson, R. B. (1982). Evidence for close contactsbetween RNA polymerase and promoter DNA. InPromoters: Structure and Function (Rodriguez, R. &Chamberlin, M., eds), pp. 164-180, PraegerScienti®c, Praeger Publishers, New York.
Starr, D. B. & Hawley, D. K. (1991). TFIID binds in theminor groove of the TATA box. Cell, 67, 1231-1240.
Stevens, A. (1976). A salt-promoted inhibitor of RNApolymerase isolated form T4 phage-infected E. coli.In RNA Polymerase (Losick, Chamberlin, ed.), pp.617-627, Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY.
Stevens, A. & Rhoton, J. C. (1975). Characterization ofan inhibitor causing potassium chloride sensitivityof an RNA polymerase from T4 phage-infectedEscherichia coli. Biochemistry, 14, 5074-5079.
Stitt, B. & Hinton, D. (1994). Regulation of middle-modetranscription. In Molecular Biology of Bacteriophage T4(Karam, J. D., Drake, J. W., Mosig, G., Hall, D. H.,Eiserling, F. A., Black, L. W., Spicer, E. K., Kutter,E., Carlson, K. & Miller, E. S., eds), pp. 142-160,American Society for Microbiology, Washington,DC.
Wang, H., Glansdorff, N. & Charlier, D. (1998). Thearginine repressor of Escherichia coli K-12 makesdirect contacts to minor and major groove determi-nants of the operators. J. Mol. Biol. 277, 805-824.
Zou, L. & Richardson, J. P. (1991). Enhancement of tran-scription termination factor rho activity with potass-ium glutamate. J. Biol. Chem. 266, 10201-10209.
Edited by M. Yaniv
(Received 8 March 1999; received in revised form 28 May 1999; accepted 7 June 1999)














![Bacteriophage [Compatibility Mode] (2)](https://static.documents.pub/doc/80x56/577cd7461a28ab9e789e8922/bacteriophage-compatibility-mode-2.jpg)












