40
Bio 402/502 Section II, Lecture 5 mRNA export, RNA surveillance, and Transport of proteins in and out of the nucleus Dr. Michael C. Yu
| Date post: | 19-Dec-2015 |
| Category: |
Documents |
| View: | 213 times |
| Download: | 0 times |

Bio 402/502Section II, Lecture 5
mRNA export, RNA surveillance, and
Transport of proteins in and out of the nucleus
Dr. Michael C. Yu

mRNA export - formation of an export competent mRNP
Sees formation of mRNP as transcription commences
Balbiani Rings (Chironomus tentans)
• Why export as a protein/DNA complex? RNAs are too big and lack the signals to interact w/ nuclear export receptors
• Specific “adaptor” proteins must first bind to the RNA and chaperone this molecule to the export receptor, which, in turn, guides the RNA across the NPC

There are other types of RNA transport in a cell

(Stutz & Izaurralde,2003)
Factors involved in mRNA export are co-transcriptionally recruited
• THO complex: major role in transcriptional elongation and recruitment of mRNA export factors
Model from yeast:
• Mex67 - promotes translocation across NPC
• Yra1 - mRNA export factor, interacts with Mex67

(Cullen, 2003)
(Sub2p)
(Yra1p) (Mtr2p)
(Mex67p)
(yeast homolog is indicated in parentheses)
Proteins involved in the nuclear export of mRNAs

(Linder & Stutz, 2001)
• Sub2, Yra1p and hnRNP proteins such as Npl3p associate co-transcriptionally with the mRNA in yeast.
• In mammalian cells, Aly/REF(Yra1) and UAP56(Sub2) are part of the exon-junction complex (EJC) on the spliced mRNA (not shown). UAP56 is replaced by the TAP-p15 (Mex67-Mtr2 in yeast) heterodimers
• The Mex67-Mtr2 heterodimers mediate the interaction of the mRNP with components of the nuclear pore complex (NPC).
• The DEAD box protein Dbp5p is required for release of mRNP on the cytoplasmic side of the NPC.
• DEAD box-mediated ATPase activities important for mRNA export are indicated by stars.
Path of transporting mRNA to the nuclear pore complex

Genetic approach to identify genes involved in mRNA export process
(Lei et al, 2003)
Mutagenized cells or collection of non-essential gene KOs
Non-essential genes
essential genes
Growth at permissive temperature
Shift to non-permissive temperature
RNA FISH w/ oligo dT
RNA FISH w/ oligo dT

(Stutz & Izaurralde, 2003)
Nuclear mRNA accumulation is observed after shifting mex67 TS mutant to the restrictive temperature (37°C)
Visualization of poly(A) mRNA is accomplished by in situ using fluorescently-labeled oligo-dT probe
Mex67(yeast) and NXF1(Drosophila) are essential genes involved in mRNA export

• Yra1p and Nab2p are essential for mRNP docking to the Mlp export gate at the nuclear periphery.
• mRNP complexes produced in the GFP-yra1-8 mutant strain are retained by the Mlp selective filter.
• mRNP stalling negatively feeds back on mRNA synthesis.
• Loss of Mlp1p or Mlp2p alleviates the negative effect on mRNA synthesis and allows a fraction of transcripts to reach the cytoplasm.
(Vinciguerra et al., 2005)
Linking mRNA biogenesis with mRNA export: Mlp proteins
Mlp proteins: filamentous proteins on the nuclear side of NPC v

(Vinciguerra & Stutz, 2004)
• The perinuclear Mlp1p protein contributes to mRNP surveillance by retaining unspliced transcripts within the nucleus
• This is achieved possibly via recognition of a component associated with the 5´ splice site.
Mlp proteins act as selective filters at NPC entrance

• Nab2p, a shuttling mRNA binding protein involved in polyA tail length regulation, directly interacts with Mlp proteins.
• Possible mechanism: by signaling proper 3´ end formation.
Nab2 is responsible for the docking of mRNPs to Mlp
(Vinciguerra & Stutz, 2004)

•An intact 5’ splice site and branchpoint are required for nuclear retention of pre-mRNAs
•Numerous splicing factors, including U1 snRNA and branchpoint binding protein (BBP/SF1), have been found to affect nuclear retention of pre-mRNAs
•In yeast, perinuclearly located Mlp1 physically retains improperly spliced pre-mRNAs but does not affect the splicing process itself
Thus, it appears that Mlp1 retains pre-mRNAsthat assemble into a spliceosome but fail toproceed through splicing before reaching thenuclear pore complex
(Galy, V. et al, 2004)
Mlp1 is involved in the nuclear retention of unspliced mRNA

What happens to produced messages? pathways for mRNA degradation
mRNA DEGRADATION
mRNA DECAY NMD
‘turnover’ ‘surveillance’
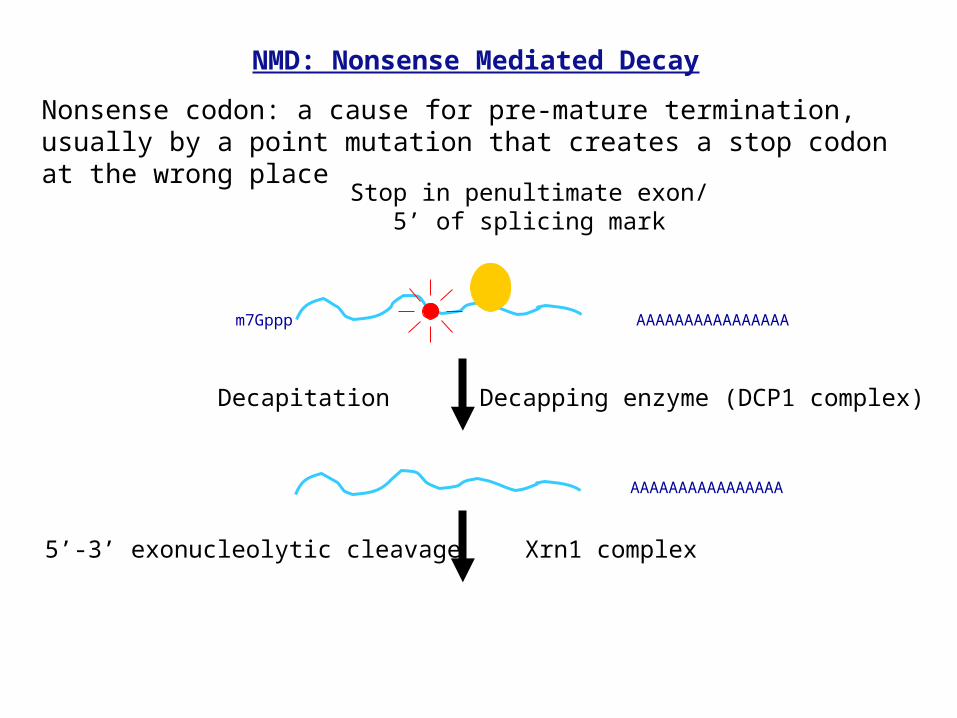
m7Gppp
Stop in penultimate exon/5’ of splicing mark
AAAAAAAAAAAAAAAA
Decapitation Decapping enzyme (DCP1 complex)
5’-3’ exonucleolytic cleavage Xrn1 complex
AAAAAAAAAAAAAAAA
NMD: Nonsense Mediated Decay
Nonsense codon: a cause for pre-mature termination, usually by a point mutation that creates a stop codon at the wrong place

(Wagner, E. and Lykke-Anderson, J., 2002)
mRNAs are checked for possible defects during its synthesis

(Vasudevan and Peltz, 2003)
Exosome
Exosome
Nuclear mRNA surveillance checks many different kinds of wrong mRNAs

The TRAMP complex is responsible for RNA degradation
(Anderson, 2005)
TRAMP complex:
- Trf4p: poly(A) polymerase- Air2p: RNA binding protein- Mtr4p: ATP-dependant RNA helicase
- Trf5p is highly related to Trf4p- Air1p is highly related to Air2p

- The TRAMP complex interacts with RNAs or RNP complexes, making them targets for degradation.
-The zinc finger domains of Air2p might be involved in substrate binding. The RNA is then polyadenylated by Trf4p.
- Exosome recruitment and activation requires the intact TRAMP complex. The activated exosome then rapidly deadenylates the RNA and can penetrate into regions of stable structure.
- Helicase activity of Mtr4p is important for dissociation or remodeling of stable RNP structures to allow passage of the exosome.
(LaCava et al., 2005))
Molecular mechanism by which TRAMP complex degrades mRNA

What happens to messages made? pathways for mRNA degradation
mRNA DEGRADATION
mRNA DECAY NMD
‘turnover’ ‘surveillance’

AAAm7Gppp
poly A shortening Deadenylase complex
AAAAAAAAAAAAAAAAm7Gppp
Decapitation Decapping enzyme (DCP1 complex)
AAA
5’-3’ exonucleolytic cleavage
Decay of mRNA
Normal decay of mRNAs
Avg. mRNA half-lives:
E. coli: 4 min (2-10 min)
Yeast: 22 min (4-40 min)
Humans: 10 hours (0.5-24
hours)
Xrn1 complex

Mature mRNAs for translation
mRNAs that are inactive for translation
Possible fate for these mRNAs: storage, decay,
transport
Regulation of mRNA activity by multiple trans-factors

(Sheth et al, 2003)
Factors involved in mRNA decay are localized to P-bodies in the cytoplasm

(Wagner, E. and Lykke-Anderson, J. 2002)
Summary: molecular mechanisms for mRNA turnover
• regulated and non-regulated turn-over but apparently coordinated
• ordered pathways (e.g. deadenylation, decapping, exonucleolytic degradation)
• cross-talk between translation and turnover
• important regulation via non-coding RNA
• turnover occurs in specific cytoplasmic compartments
• NMD: recognition of premature stop codons

CytoplasmicCytoplasmicfilamentfilament
CytoplasmCytoplasm
NucleusNucleus
CytoplasmicCytoplasmicringring
Inner ringInner ring
BasketBasket
Distal ringDistal ring
The Nuclear Pore ComplexThe Nuclear Pore Complex
RibosomeRibosome
~150Å
~2000Å
NPC: all traffic between nucleus & cytoplasm in higher eukaryotes occurs via NPC (exception: mitosis)

More on the nuclear pore complex
NPC size: diameter = 120nm, 8-fold rotational symmetry, MW = 125 million daltons, composed of 50-100 different proteins called nucleoporins (Nups)
NPC extension is thought to be the initial cargo docking site

The nuclear pores on the membrane
Nuclear basket

Small molecules can diffuse freely through the nuclear pore
However, large molecular requires active transport

Soluable nuclear transport receptors facilitates the transport
(J. Lingappa, 2003)
• Karyopherins: nuclear transport receptors that bind to Nups.
• Different factors are utilized for export (exportin) and import (importin)
• Nuclear transport factors bind to cargo (or adaptors that bind cargo) that contains either nuclear localization sequence (NLS) or nuclear export signal (NES)
• Types of cargo: proteins and RNAs

Summary of factors involved in nuclear protein import cycle
(Stewart, 2007)

Nuclear localization sequence is required for protein nuclear import
Functional NLS is required for nuclear localization (mutation in NLS results in cytosolic localization)
NLS: consists of either one or two stretches of basic amino acids

The nuclear import/export cycle
(Lei and Silver, 2004)
1. In the cytoplasm, NLS-containing cargo is bound by the heterodimeric import receptor (importin /

The nuclear import/export cycle
(Lei and Silver, 2004)
2. Ran-GTP binds to importin, causing conformational change in the importin, which releases the cargo

The nuclear import/export cycle
(Lei and Silver, 2004)
3. If cargo is phosphorylated, it can interact with exportins and Ran-GTP, allowing the complex to exit the nucleus

The nuclear import/export cycle
(Lei and Silver, 2004)
3. In the cytoplasm, hydrolysis of Ran-GTP to Ran-GDP causes dissociation of the export complex

The molecular machineries behind nuclear protein transport
Nucleoporins (known as Nups):
• Make up the cytoplasmic filaments, nuclear basket, and line the pore
• Contain different types of FG (Phe/Gly) repeats
• Nuclear transport factors make multiple sequential contacts with distinct
Nups, resulting in docking and translocation of nuclear transport complexes
• Exact mechanism by which Nups mediate translocation of complexes remains
unclear. but may involve sequential association with of receptor plus cargo with FG repeats lining pore
(Bayliss, R. et al. 2000)

RanGTP/GDP exchange: driving force behind cargo exchange
Ran-GTP: • A small GTPase that gives directionality to nuclear transport, see below• Members of the Ras oncogene family (like Rab, which regulates vesicle fusion)• Binds to nuclear transport receptors
(Matsurra and Stewart, 2004)
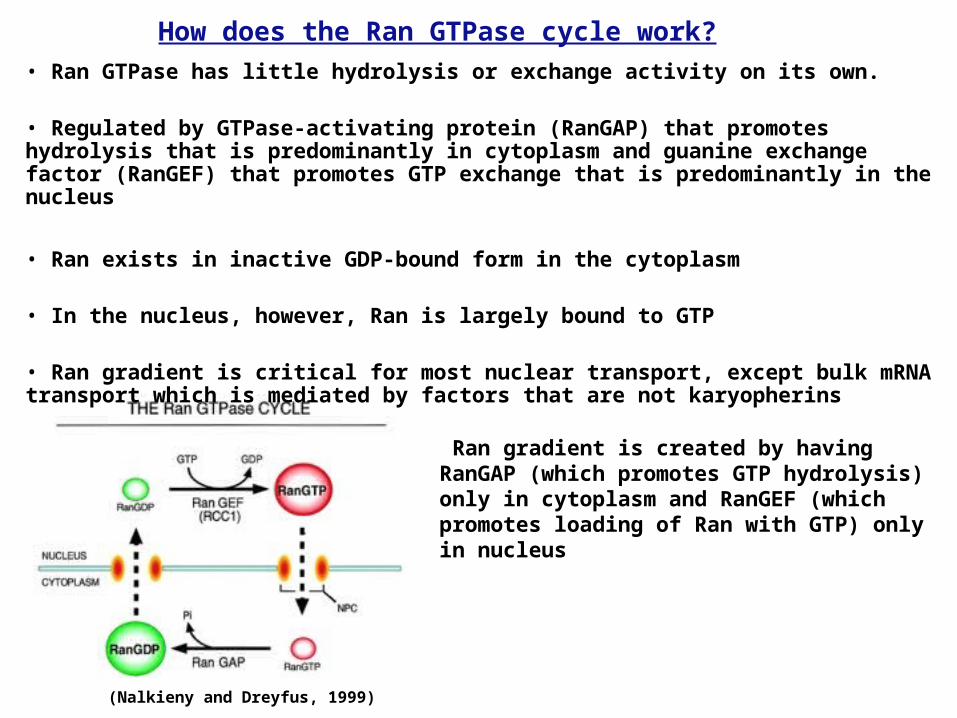
• Ran GTPase has little hydrolysis or exchange activity on its own.
• Regulated by GTPase-activating protein (RanGAP) that promotes hydrolysis that is predominantly in cytoplasm and guanine exchange factor (RanGEF) that promotes GTP exchange that is predominantly in the nucleus
• Ran exists in inactive GDP-bound form in the cytoplasm
• In the nucleus, however, Ran is largely bound to GTP
• Ran gradient is critical for most nuclear transport, except bulk mRNA transport which is mediated by factors that are not karyopherins
(Nalkieny and Dreyfus, 1999)
How does the Ran GTPase cycle work?
Ran gradient is created by having RanGAP (which promotes GTP hydrolysis) only in cytoplasm and RanGEF (which promotes loading of Ran with GTP) only in nucleus

Mechanism of proteins that shuttles between nucleus and cytoplasm
1. Proteins contain both nuclear localization and export signals (NLS + NES).
2. These proteins shuttle back and forth between nucleus and cytosol.
3. Rate of export and import determines in which compartment it resides.
4. Export/ import of shuttling proteins can be regulated, i.e. by phosphorylation-dephosphorylation of residues adjacent to NLS or NES signals resulting in blockade/exposure of signals.

Experimental approaches studying nuclear trafficking
Heterokaryon assay - determines if a nuclear protein shuttles in/out of nucleus
1. Nuclei transfected/injected to express tagged protein (+ marker protein that stays in "donor" nucleus)
2. Fuse cells to another set of cells using PEG.
3. Examine whether tagged protein shows up in cytoplasm alone (export), or in recipient nucleus as well (export followed by import, implying shuttling) using IF
Heterokaryon: solid arrowHeLa cell = arroweadXenopus cell: dotted arrow
Green: nuclear restricedRed: shuttling

Experimental approaches studying nuclear trafficking
Immunofluorescent tags
• Transfect cells with proteins tagged with GFP, RFP, YFP, etc. Assess nuclear vs. cytoplasmic location by IF (immunofluorescence)
• Or, you can transfect cells which are epitope tagged and use antibodies conjugated with fluorescently-tag to perform IF.
SRPK1: cytoplasm SC35: nuclear Combined image
(Ding et al, 2006)
Confocal microscopy

















